











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. INTRODUCCIÓN
La descomposición es un proceso biológico por el cual se transforman las sustancias orgánicas en inorgánicas, siendo el aporte de hojarasca y su posterior mineralización un proceso de retroalimentación positiva para la productividad del bosque (Gallardo y Merino, 2007). Como proceso biológico se encuentra regulada por factores del ambiente físico tales como la temperatura, la humedad, la radiación ultravioleta, las características fisicoquímicas del suelo, el ambiente biótico tales como la cantidad y calidad de la hojarasca, las características de las comunidades de invertebrados y microorganismos presentes en el suelo y la cantidad y frecuencia del aporte de hojarasca al suelo entre otras Mungai y Motavalli, 2006; Carrera et al., 2009).
Para algunos autores el patrón general en la pérdida de peso de la hojarasca durante la descomposición comprende dos fases, una inicial en la cual se produce un rápido lavado de compuestos solubles y la descomposición de sustancias lábiles como azúcares, algunos fenoles, almidones y proteínas; y una segunda fase más lenta, en la cual comienza la descomposición de sustancias recalcitrantes tales como celulosa, hemicelulosa, taninos y lignina (Aceñolaza y Gallardo Lancho, 1994; Arellano et al., 2004; Goma-Tchimbakala y Bernhard-Reversat, 2006). Asimismo la tasa de descomposición, visualizada como pérdida de peso seco de hojarasca, está afectada por una serie de variables ambientales relacionadas a la edad del bosque (Aceñolaza y Gallardo, 1994).
Como antecedente de estudios de descomposición de hojas en el área de estudio, se destacan los efectuados para especies nativas (xerófilas) del Espinal Mesopotámico (Mendoza, 2015); sin embargo existen pocos antecedentes específicos para Carya illinoinensis (Wang.) K. Koch (Mungai y Motavalli, 2006) y ninguno para la región.
El cultivo de C. illinoinensis se introduce alrededor de la década del cincuenta en el Delta del Paraná (Madero et al., 2007), y posteriormente se comenzó a expandir en diversas regiones del país. Esta especie, originaria de Norteamérica, es cultivada para la obtención de un fruto seco de alto valor comercial, y con un creciente interés regional (Castillo et al., 2013).
El presente trabajo tiene como objetivo analizar la descomposición de hojas de C. illinoinensis en bosques cultivados de diferentes edades, bajo la hipótesis que, el incremento en la cobertura de la canopia forestal induce cambios a nivel de sotobosque que influyen en la tasa de descomposición. Una mayor cobertura vegetal generaría un microambiente más húmedo a nivel de suelo y con menor radiación solar.
2. MATERIALES Y MÉTODOS
Área de estudio. El estudio se realizó en parcelas comerciales de C. illinoinensis situados en el departamento de Villaguay, provincia de Entre Ríos, Argentina.
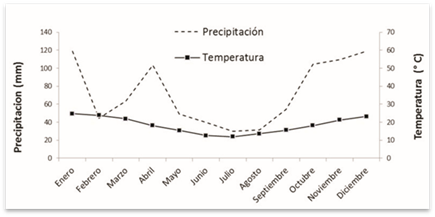
El clima es templado húmedo; las precipitaciones medias anuales son de 1000 mm año-1 (INTA, 2000); las lluvias se distribuyen uniformemente a lo largo del año, pero con incrementos en otoño y primavera. La temperatura media anual es de 16 ºC; la media del mes más frío 11 °C (Julio) y del mes más cálido 25 °C (Febrero), (Figura 1).
Los suelos son de textura arcillosa, profundos, con escaso drenaje, con concreciones calcáreas a partir de los 60 cm de profundidad y deficitarios en P asimilable. Taxonómicamente los suelos del área de estudio corresponden al orden Vertisol, con pH levemente ácido (6,3 ± 0,7) y contenido de materia orgánica de 5,4 ± 3,0 %, dependiendo del uso del suelo (INTA, 2000).
La vegetación natural del área de estudio corresponde a la Provincia Fitogeográfica del Espinal, Distrito del Ñandubay (Cabrera, 1994) y los bosques nativos dominados por especies arbóreas tales como Prosopis nigra, Prosopis affinis y Acacia caven tienen un uso silvopastoril (Mendoza et al., 2012; 2014a; 2014b). La implantación de bosques cultivados de C. illinoinensis en la región, tiene por finalidad la producción de frutos para su comercialización (nuez pecan).
Caracterización de los bosques de C. illinoinensis. Los bosques estudiados poseen un marco de plantación de 10 x 10 m. Las diferentes edades de los bosques están delimitadas espacialmente y fueron identificados como Bosque 1 (7 años de edad, B1 ), Bosque 2 (12 años de edad, B2 ) y Bosque 3 (16 años de edad, B3 ); las características dasonómicas y sus aportes de hojarasca se detallan en el Tabla 1.
Tabla 1. Características dasométricas de los bosques de C. illinoinensis. D (densidad), AB (área basal), DAP (diámetro a la altura del pecho) h (altura), Cob (cobertura), Ah (aporte de hojarasca).
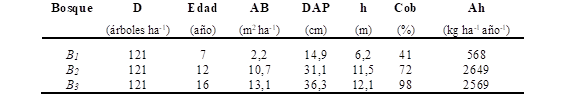
Metodología de estudio
Se empleó la técnica estandarizada de bolsas de descomposición o “litter bags” (Aceñolaza y Gallardo, 1994; Carranza et al., 2012) que consiste en la colocación de hojarasca de peso conocido en bolsas de descomposición que se retiran a lo largo del tiempo por dos años. A tal fin se dispusieron un total 84 muestras (28 por cada bosque) en dimensiones de 20 x 20 cm2 con malla plástica de 1,5 mm de luz con 3,00 g (peso seco) de hojas recolectadas en otoño desde la copa de árboles y sin signos de enfermedades. Posteriormente se dispusieron sobre el suelo bajo la copa de árboles en cada bosque de C. illinoensis para simular condiciones naturales de descomposición. Las mismas se fueron retirando progresivamente y al azar (4 bolsas por bosque) con una frecuencia de 6, 27, 67, 137, 308, 373, 468 días desde el inicio del ensayo. En cada fecha de muestreo, se determinó peso seco remanente de las bolsas retiradas de la parcela (Santa Regina et al., 1997 y Palma et al., 2002) limpiando los residuos de hojas con aire, secándolos a una temperatura de 80 °C durante 3 h y pesando la materia seca con balanza electrónica de acuerdo al procedimiento propuesto por Chapman y Pratt (1979).
Ajuste de las curvas de descomposición
Se determinó el índice de descomposición k de acuerdo al modelo modificado de Olson (1963), que se observa en la Ecuación 1:
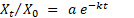 Ecuación 1
Ecuación 1
Donde: Xt : peso seco del material remanente en un tiempo t (g), Xo : peso seco inicial (g), a: parámetro de la fracción lábil, k: constante de descomposición (año-1), t: tiempo transcurrido (año).
A partir de la k, se calculó la vida media o tiempo necesario en años para alcanzar una descomposición del 50 % y del 99 % del material de partida (t50% = 0,6931/k y t99% = 6,9077/k) y la velocidad inicial de pérdida de peso (ak) (Arunachalam y Singh, 2002).
Una vez ajustadas las curvas de descomposición se determinaron los coeficientes “r2” para estudiar la bondad del ajuste.
Análisis de los resultados
Para comparar los valores obtenidos de cada una de estos cuatro parámetros (k, t50%, t99% y ak) entre los tres bosques, se utilizó un análisis de la varianza de un factor con la formulación habitual 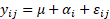 , siendo
, siendo 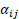 el efecto del bosque i y
el efecto del bosque i y 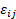 el error aleatorio con
el error aleatorio con 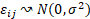 . Para la comparación de las medias estimadas en cada bosque se utilizó el test HSD de Tukey con un nivel de significación de 0.05.
. Para la comparación de las medias estimadas en cada bosque se utilizó el test HSD de Tukey con un nivel de significación de 0.05.
Además se ha utilizado un modelo lineal mixto de análisis de la varianza con un factor inter-sujetos (bosque) con cuatro réplicas y un factor intra-sujetos de medidas repetidas (tiempo), con el fin de conocer el efecto de la edad de plantación (tipo de bosque) y el del tiempo transcurrido en la descomposición de las hojas. La formulación matemática del modelo fue la siguiente:
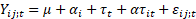
con i = 1, 2, 3 para los tres bosques; j = 1, 2, 3, 4 para las cuatro muestras dentro de cada bosque, t =1, 2, 3, 4, 5, 6 y 7 para los tiempos de medición, y siendo:
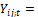 peso de la muestra j en el bosque i en el tiempo de medición t.
peso de la muestra j en el bosque i en el tiempo de medición t.
µ = efecto de media general.
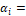 efecto principal del bosque i.
efecto principal del bosque i.
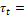 efecto principal del tiempo t.
efecto principal del tiempo t.
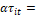 efecto de interacción del bosque i con el tiempo de medición t.
efecto de interacción del bosque i con el tiempo de medición t.
 error aleatorio para el peso de la muestra j en el bosque i en el tiempo de medición t.
error aleatorio para el peso de la muestra j en el bosque i en el tiempo de medición t.
Las hipótesis del modelo fueron:
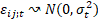
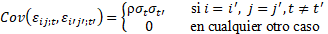
La estructura de varianzas del modelo lineal mixto contiene por tanto 6 parámetros de varianza que fueron estimados por el método de máxima verosimilitud restringida (REML). Las comparaciones entre las medias estimadas por el modelo se hicieron utilizando test t individuales con nivel de significación 0,05. La normalidad de los residuales studentizados del modelo se analizó utilizando el test de Kolmogorov-Smirnov, que proporcionó un p-valor mayor que 0,15.
3. RESULTADOS
Pérdidas de peso en bosques de diferentes edades
Comparando las pérdidas de peso de las hojas colocadas en bolsas de descomposición desde el inicio hasta los 67 días, los porcentajes de peso seco residuales oscilaron en torno al 80 %. Los porcentajes de peso remanente en las bolsas de descomposición estuvieron en torno al 50 % a los 137 días del inicio del ensayo y hacia la finalización (468 días) los valores oscilaron entre el 5 y el 13 % (Tabla 2).
Tabla 2. Porcentaje de peso remanente de las hojas en bosques de C. illinoinensis (B1 , B2 y B3 ) Media ± (desviación estándar) con n = 4.
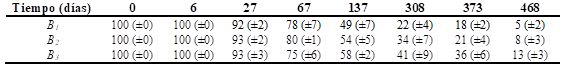
Las tasas de descomposición de las hojas evaluadas por el índice k, registraron valores de 1,85; 1,55 y 1,21 año-1 en los B1, B2 y B3 (con diferencias significativas entre sí, Tabla 3), indicando una máxima velocidad de descomposición en el B1 (bosque más joven) y mínima descomposición en el B3 (bosque de más edad), con valores intermedios en el B2 (bosque intermedio). El parámetro a alcanzó un valor en torno a 1,0 indicando que el 100 % de la fracción de hojas interviene en la descomposición. Además, sólo mostró diferencias significativas entre B1 y B3 , con valores intermedios en B2 (Tabla 3).
El tiempo que tardaría en descomponerse la mitad de las hojas de C. illinoinensis expresado como t50% fue de 0,37; 0,45 y 0,58 años para B1 , B2 y B3 ; respectivamente (con diferencias significativas entre sí) mientras que el tiempo estimado para la descomposición del 99 % sería de 3,7; 4,4 y 5,7 años para B1 , B2 y B3 ; respectivamente (con diferencias significativas entre sí; Tabla 3). Coherentemente con los parámetros t50% yt99%, las velocidades iniciales en las pérdidas de peso expresadas como ak fueron máximas en B1 y mínimas en B3 , con diferencias significativas entre bosques (Tabla 3).
El modelo de regresión ajustado para la estimación de la descomposición de las hojas tuvo coeficientes de determinación r2 altos que oscilaron entre 0.96 y 0.98, lo cual denotó que el modelo fue apropiado para el análisis de dichos datos (Tabla 3).
Tabla 3. Parámetros de la descomposición en hojas de C. illinoinensis obtenidos de ajustar los datos a la Ecuación 1.
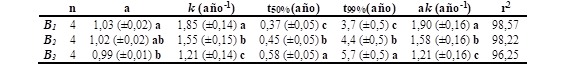
Siendo a parámetro adimensional; t50% y t99% (tiempo en años para alcanzar una descomposición del 50% y del 99% respectivamente) y ak (velocidad inicial de pérdida de peso). Letras minúsculas diferentes indican diferencias significativas (p <0,05) entre bosques.
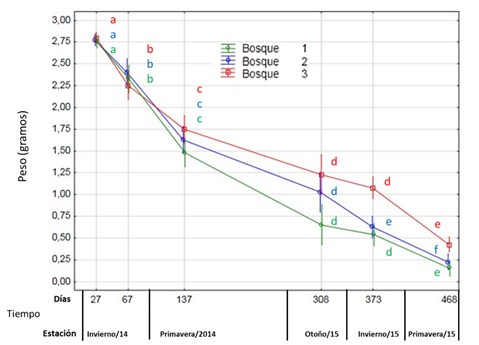
Patrones de pérdida de peso entre bosques. Analizando el patrón temporal en la pérdida de peso por descomposición de hojas de C. illinoinensis (Fig. 2), se observa que la misma registró disminuciones a tasas similares en la mayoría de los puntos analizados, a excepción de los B1 y B3 que, durante el último tramo del proceso de descomposición, a partir del invierno del año 2015.
4. DISCUSION
Tasas de descomposición en bosques de diferentes edades. El valor medio de la k (1,5 año-1) hallado para la secuencia de bosques de C. illinoinensis en Entre Ríos (Argentina) fue menor respecto al 3,2 año-1 hallados por Mungai y Motavalli (2006) para la fracción recalcitrante de la misma especie cultivada en el centro norte de Missouri (EEUU). Estas diferencias observadas en los valores de k para una misma especie cultivada en diferentes regiones geográficas, podrían estar influenciadas por la variable clima ya que, si bien ambos sitios poseen características similares en precipitaciones, la temperatura media anual de Missouri es menor por encontrarse en una situación de mayor continentalidad y con 6 grados de latitud más que el sitio del presente estudio.
Los valores de k hallados para estos bosques de C. illinoinensis cultivados en un clima templado (Tabla 3) fueron superiores a los valores de referencia que oscilaron entre 0,08 y 0,47 año-1, para especies de regiones templadas propuestos por Melillo et al. (1982) y Aerts (1997).
Comparando las tasas de descomposición de C. illinoinensis, respecto a la descomposición de hojas de especies nativas tales como Acacia caven (Molina) Molina, Prosopis affinis Spreng., Prosopis nigra (Griseb.) Hieron.; típicas de la vegetación del Espinal Mesopotámico (Mendoza, 2015), se observó una mayor descomposición en C. illinoinensis (k = 1,5 año-1) del orden del doble respecto al valor medio observado en nativas (k = 0,8 año-1). Esto mismo, fue probado para otras especies leñosas en bosques de Yungas (Tucumán, Argentina) cuando se comparó la descomposición entre exóticas y nativas (Fernández y Aragón, 2014). Si bien los estudios que compararon la descomposición de hojas de especies exóticas versus nativas parecerían indicar que en general las exóticas se descompondrían más rápido; en ecosistemas mediterráneos de España (Godoy et al., 2010) se concluyó que los factores de mayor incidencia en las tasas de descomposición fueron la composición química referida al contenido de lignina y la relación carbono/nitrógeno. Además, los valores del parámetro “a” obtenido en el ajuste al modelo de Olsen de la descomposición de las hojas de C. illinoinensis corrobora que el 100 % del material utilizado en el ensayo interviene en el proceso de descomposición, al presentar este parámetro un valor en torno a 1,0.
Las diferencias halladas entre los valores de k en bosques de C. illinoinensis de diferentes edades (Tabla 3), se atribuyeron a los porcentajes de cobertura de copa observada entre bosques (Tabla 1), siendo el parámetro dasonómico más influyente en las condiciones microclimáticas al interior de cada bosque, modificando la radiación indecente sobre el suelo y en consecuencia la temperatura del suelo bajo copa. Todos estos factores fueron descriptos por Mungai y Motavalli (2006) como variables a considerar en el proceso de descomposición. Consecuentemente, la mayor k de descomposición en B1 se asoció a una menor cobertura de copa asociada a una mayor radicación solar y una mayor temperatura, posibilitando más actividad biológica de los descomponedores; en contraposición a lo sucedido en el B3 donde las condiciones de mayor cobertura se relacionaron a una menor actividad biológica.
Los resultados observados sugieren que la cobertura variable entre bosques, con una radiación incidente variable, determinarían distintas temperaturas a escala de micrositio, siendo el factor más influyente en comparación con la humedad edáfica que permanecería más o menos constante debido a que las precipitaciones se distribuyen uniformemente a lo largo del año (Fig. 1). Sin embargo, en contraposición a lo observado para nuestro estudio, (Mungai y Motavalli, 2006) no encontraron diferencias en las pérdidas de peso cuando compararon la descomposición de hojas de C. illinoinensis bajo la conopia y fuera de la canopia. Aceñolaza y Gallardo (1994) describieron diferencias altamente significativas en los índices de descomposición del bosque maduro de Alnus acuminata H.B.K. respecto al bosque joven y senil; siendo mayor la descomposición en hojas de etapas iniciales y finales donde su cobertura fue menor y por consiguiente un mayor ingreso de luz al nivel de suelo; los autores asociaron esta situación a mejores condiciones de temperatura para los descomponedores.
Las diferencias en las velocidades de descomposición asociadas a las diferentes coberturas arbóreas descriptas en C. illinoinensis, también fueron halladas para otras especies exóticas por Kourtev et al. (2002), quienes concluyeron que solo algunas especies registraron distintas velocidades en la descomposición cuando la cobertura es variable. Estas variaciones microclimaticas al interior de los bosques, que inciden en los procesos de descomposición de hojas, fueron expuestas por Bahamonde et al. (2009).
Patrones en la descomposición de hojas entre bosques. En los tres bosques se observaron máximas velocidades de descomposición hasta los 137 días (Fig. 2); lo cual fue coherente con los análisis de Coleman et al. (2004) quienes plantearon que durante los estados iniciales (primeras semanas) la velocidad de descomposición se acelerada debido a la lixiviación de compuestos solubles y al desarrollo de poblaciones microbianas que empiezan a consumir los compuestos que son fácilmente degradables. Esto también se atribuye a que el material utilizado fue solamente hojas, no considerándose las demás fracciones de la hojarasca que contienen más lignina (como la fracción tallos) que suele tener mayor resistencia a la descomposición, como fue estudiado por Ibarra et al. (2011).
El patrón de las pérdidas de peso entre bosques no tuvieron diferencias significativas entre los 27 y 67 días de estudio; con porcentajes de hojas remanentes (a los 67 días) que oscilaron entre un 22 y 25 % (diferenciándose significativamente entre B1 y B3 ; Fig. 2); lo cual tuvo similitud con estudios realizados en otras especies exóticas como Ligustrum lucidum y Morus alba pertenecientes a un bosque serrano de Córdoba, Argentina, que tuvieron pérdidas de peso después de 43 días de entre un 15 % y 30 % (Furey et al., 2014). Se registraron diferencias significativas entre bosques a partir de los 137 días, acentuándose hacia la finalización del estudio las diferencias B3 (con una menor tasa de descomposición) respecto a los demás (Fig. 2). Las mayores tasas de pérdida de peso seco del bosque joven B1 implican rápida mineralización y por lo tanto mayores tasas de retorno de nutrientes al suelo en etapas iniciales, que se ralentizan con la edad del monte productivo.
5. CONCLUSIONES
Se concluye que, las diferencias significativas en las tasas de descomposición de hojas en bosques de diferentes edades de C. illinoinensis, se asocian a las diferentes coberturas arbóreas; conjeturando que las mejores condiciones para la descomposición de hojas en B1 , se relacionaron con mayores temperaturas edáficas superficiales vinculadas a una mayor radiación solar, posibilitando una mayor actividad biológica para los descomponedores.

