











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The cactus pear is an important resource for animal feeding due to its rusticity and production potential even with low water availability, especially in the semi-arid region of Brazil, as this plant presents high forage supply capacity when compared to the native vegetation of the caatinga 14, mainly during the dry season.
When dealing with historical aspects of cactus pear, the literature points to Mexico as its origin point 7. The cactus pear species most used as forage belong mainly to the genera Opuntia and Nopalea10, which present variable chemical composition according to the period of the year, plant age, cladode order, genotype, fertilization management, cultivation spacing, and other factors 6. It is of a forage that has low contents of dry matter (63.7 g kg-1), crude protein (54.2 g kg-1), neutral detergent fiber (283.0 g kg-1) and acid detergent fiber (209.3 g kg-1) 4. However, it is rich in non-fibrous carbohydrates (473.6 g kg-1) and total digestible nutrients, presenting high water content, and high animal acceptability. The cactus pear is also rich in vitamins A, complex B and C, and minerals such as calcium, magnesium, sodium, potassium, and 17 types of amino acids 13,17.
Cactus pear is also considered a strategic forage reserve and, depending on the needs of the producer and climatic conditions, the harvest frequency may vary. Depending on the crop structure, the cactus pear can be harvested manually and transported by animal traction to its place of use. In general, this operation is carried out daily, increasing production costs 23. Another factor that jeopardizes the harvest is the lack of pattern of the orchard, hindering management practices, such as fertilization. Also, direct grazing might compromise its cultivation, due to damages caused to the plants by the animals 3.
Post-harvest storage may be an alternative to lower the costs of harvesting and transportation of the material, however the storage must be done in a shady or covered place as long as it is ventilated. However, few information is available in the literature on post-harvest storage of cactus pear genotypes and its effects on the nutritional value and animal performance. It was hypothesized that the cactus pear genotypes stored for up to 60 days post-harvest would not present changes in their chemical and mineral composition enough to reduce their quality. Thus, the objective was to evaluate water loss and chemical composition of the cactus pear genotypes Doce, Baiana and Orelha de Elefante Mexicana submitted to up to 60 days of storage post-harvest.
Material and methods
Experimental site
The experiment was carried out at the Experimental Farm Alvorada of Gurguéia, which belongs to the Federal University of Piauí (UFPI), in Alvorada of Gurguéia, Piauí, Brazil. The city of Alvorada of Gurguéia is located at latitude 08°25’28” South and longitude 43°46’38” West, at an altitude of 281 m. The region has a semi-arid climate, with a dry period of approximately eight months 1.
Experimental design
The experimental design adopted was the completely randomized, in a 3 × 5 factorial arrangement, with three genotypes of cactus pear (Doce, Baiana and Orelha de Elefante Mexicana) and five storage periods (0, 15, 30, 45 and 60 days), in ten replications.
Planting and fertilization
Before planting the cactus pear genotypes, a soil sample was collected for analysis and chemical characterization of the 0-20-cm layer, which was carried out at the Soil Analysis Center of the UFPI, Bom Jesus, Piauí, Brazil. It was not necessary to perform soil correction based on soil base saturation and crop requirement. The basic fertilization consisted of the application of 50 kg ha-1 of nitrogen as urea (45% N), 50 kg ha-1 of potassium as potassium chloride (48% K2O) and 30 kg ha-1 of phosphorus as single superphosphate (18% P2O5).
The spacing used for the cultivation of the cactus pear genotypes was 1.5m × 0.1m, in a density of 66,133 plants ha-1. Three genotypes of cactus pear were planted in December 2013: Nopalea cochinillifera, genotypes Doce and Baiana, and Opuntia tuna, genotype Orelha de Elefante Mexicana (OEM). After 2 years of cultivation under rainfed conditions, the harvest was performed on December 6th, 2015. The cactus pear was harvested manually, using a 14-inches Tramontina® knife preserving a residual cladode area 23, and stored in a ventilated shed on wooden pallets, with a height of approximately 10 cm from the floor, storing 200 cladodes from each genotype, thus providing a random choice, for a period of up to 60 days of post-harvest.
Quantification of water loss
After harvesting, ten cladodes of each genotype were selected and separated for weighing, which were alwas the same cladodes to be weighed according to the storage periods. In order to measure the amount of water lost by the catus pear genotypes, it was used a digital electronic scale, with capacity of 1 g to 5 kg, model Sf-400 UNICASA®. The calculation of water loss was made through the difference between the weight of the current period and the previous period.
The cladodes of the cactus pear genotypes were subjected to natural drying due to the conditions of rainfall (mm), air relative humidity (%) and room temperature (°C). The accumulated precipitation during the entire experimental period was 387 mm. The maximum and minimum values of air relative humidity were 93.5% and 33%, respectively. The maximum temperature values ranged from 25.6°C to 38.4°C, and the minimum temperature values ranged from 19.2°C and 24.2°C.
Chemical composition
Ten cladodes of each genotype and each treatment (0, 15, 30, 45 and 60 days of storage) were randomly collected in plastic bags, duly identified and taken to the Animal Nutrition Laboratory of the UFPI, where they were chopped, weighed and taken to a forced ventilation oven at a temperature of 65°C until reaching constant weight. Later, the pre-dried samples were ground in a “Thomas Wiley” stationary mill, model SP-32 SPLAPOR® with a 1.0-mm mesh sieve, and packed in containers with covers for chemical analysis. The contents of dry matter (DM) (Method n° 934.01), mineral matter (MM) (Method n° 930.05), crude protein (CP) (Method n° 981.10) and ether extract (EE) (Method n° 920.29) were determined 2. In the analysis for the determination of neutral detergent fiber (NDF) and acid detergent fiber (ADF), it was adopted a methodology with modifications proposed by the Ankon device manual of the Ankon Technology Corporation 25. Lignin was determined on the ADF residue with 72% sulfuric acid 25. For the estimation of total carbohydrates (TCHO) the following equation was used: TCHO = 100 - (%CP + %EE + %MM) (24). The non-fiber carbohydrates (NFC) were estimated through the equation: NFC = 100 - %MM - %CP - %EE - %NDF 11.
Mineral composition
In the analysis of mineral composition, after nitric-perchloric digestion, the phosphorus (P) contents were determined by UV/VIS spectrophotometry at 660nm, by reading the blue color intensity of the phosphomolybdic complex produced by the reduction of molybdate with ascorbic acid in a spectrophotometer model IL-592 EVEN®. Whereas the contents of potassium (K), calcium (Ca), magnesium (Mg), iron (Fe), manganese (Mn), zinc (Zn), sodium (Na) and copper (Cu) were determined by atomic absorption spectrophotometry (AAS), in a spectrophotometer model AA240FS VARIAN® (20), in the Soil Analysis Center of the UFPI.
Statistical analysis
The results were submitted to analysis of variance and regression analysis (storage period), by the Scott-Knott test (cactus pear genotypes), at a level of 5% of significance, using the software SISVAR version 5.0 developed by the Federal University of Lavras 8.
Results
Quantification of water loss
Regarding the analysis of water loss of the stored cladodes, there was no effect (P=0.2589) of the interaction between the factors genotypes and storage periods (Figure 1).

Figure 1: Figura 1: Water loss of the cactus pear genotypes during post-harvest storage periods. La pérdida de agua de las variedades de pera de cactus durante los períodos de almacenamiento posteriores a la cosecha.
It was observed a significant difference between the cactus pear genotypes for water loss (P=0.0004), in which the genotype Baiana presented the highest water loss (0.218 g). There was a linear reduction (P=0.018) of water loss in all genotypes as the storage period advanced, with wate loss being observed only after 15 days os post-harvest.
Chemical composition
It was observed significant effect (P<0.05) of the interaction between cactos pear genotypes and storage periods on the variables dry matter, crude protein, ether extract, mineral matter, organic matter and non-fiber carbohydrates (Table 1).
Table 1: Tabla 1: Chemical composition of cactus pear genotypes submitted to different storage periods. Composición química de las variedades de pera de cactus forraje sometidas a diferentes períodos de almacenamiento.

1g kg-1 on DM basis; 2OEM: Orelha de Elefante Mexicana; P-value: linear regression for the storage periods, *significant at 5%; nsnon-significant at 5%; Means followed by different uppercase letters in the columns are statistically different by the Scott-Knott test (P<0.05).
1g kg-1 en base DM; 2 OEM: Oreja de elefante mexicano; Valor P: regresión lineal para los períodos de almacenamiento, * significativo al 5%; nsnon-significativo hasta el 5%; Las medias seguidas de letras mayúsculas diferentes en las columnas, difieren estadísticamente según la prueba de Scott-Knott (P <0,05).
Regarding the DM content, only the genotype Baiana had significant effect (P=0.0106) of the storage periods, presenting the highest content at 15 days (114.13 g kg-1), and decreasing over time (Table 1). Regarding the CP content, a linear increase (P=0.0006) was observed for the genotype OEM, and linear decrease for the genotypes Doce and Baiana (Table 1).The genotype OEM also presented the highest CP contents (54.2 ± 0.07 g kg-1) in all storage periods and a linear increase in the EE content (P=0.0061).
There was a linear increase in MM (P<0.0001) and decrese in OM, TCHO and NFC variables in the different storage periods (Table 1). The genotype OEM presented the lowest levels of MM except for 30 days of storage (167.60 ± 0.17 g kg-1). The increase in the MM content in this period can be explained by the wearing of OM caused by higher temperatures up to 29 days during the entire studied period.
Regarding the chemical composition of the cell wall, there was a significant difference between the cactus pear genotypes for NDF, ADF and lignin (Table 2).
Table 2: Tabla 2: Chemical composition of the cell wall of cactus pear submitted to different storage periods. Composición química de la pared celular de la pera del cactus forrajero sometida a diferentes períodos de almacenamiento.
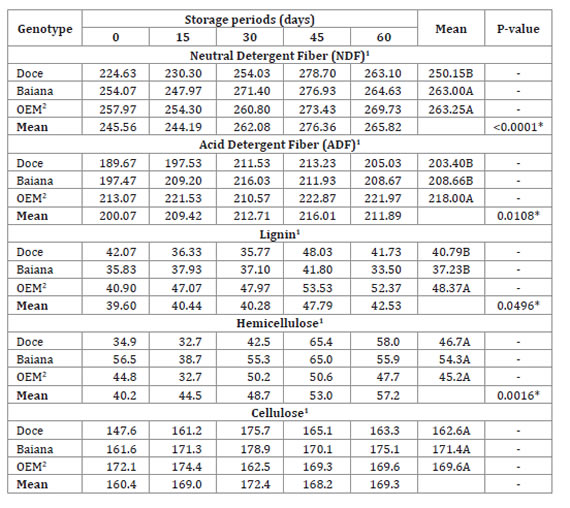
1g kg-1 on DM basis; 2OEM: Orelha de Elefante Mexicana; P-value: linear regression for the storage periods, *significant at 5%; nsnon-significant at 5%; Means followed by different uppercase letters in the columns are statistically different by the Scott-Knott test (P<0.05).
1g kg-1 en base DM; 2OEM: Oreja de elefante mexicano; Valor P: regresión lineal para los períodos de almacenamiento, * significativo al 5%; nsnon-significativo hasta el 5%; Las medias seguidas de letras mayúsculas diferentes en las columnas, difieren estadísticamente según la prueba de Scott-Knott (P <0,05).
The different storage periods caused these variabes to present a linear increase. The genotype Doce presented the lowest levels of NDF and ADF (250.15 ± 0.64 g kg-1 and 203.40 ± 0.61 g kg-1, respectively). The genotype OEM presented the highest lignin contents when compared to the others (48.37 ± 0.35 g kg-1). There was no significant difference between the cactus pear genotypes for hemicellulose and cellulose (Table 2). Nevertheless, hemicellulose presented a linear increase (P=0.0016) in the storage periods.
Mineral composition
As for the composition of macro and micronutrients of the cactus pear genotypes in the different storage periods, there was effect (P<0.05) of the the storage periods on the studied variables, exception for potassium (K). Regarding the isolated effects of the cactus pear genotypes, potassium (K), calcium (Ca), manganese (Mn) and sodium (Na) did not have significant effect (Table 3, page 267).
Table Tabla 3: 3: Composition of macronutrients and micronutrients of cactus pear genotypes submitted to different storage periods. Composición de macronutrientes y micronutrientes de las variedades de pera de cactus forraje sometidas a diferentes períodos de almacenamiento.

1g kg-1 on DM basis; 2OEM: Orelha de Elefante Mexicana; P-value: linear regression for the storage periods, *significant at 5%; nsnon-significant at 5%; Means followed by different uppercase letters in the columns are statistically different by the Scott-Knott test (P<0.05).
1g kg-1 en base DM; 2OEM: Oreja de elefante mexicano; Valor P: regresión lineal para los períodos de almacenamiento, * significativo al 5%; nsnon-significativo hasta el 5%; Las medias seguidas de letras mayúsculas diferentes en las columnas, difieren estadísticamente según la prueba de Scott-Knott (P <0,05).
The Mg content statistically differed among genotypes, where OEM presented the highest content in comparison to the genotypes Doce and Baiana (4.83 ± 0.32 g kg-1) (Table 3, page 267). In addition, it presented linear reduction (P=0.0005) in the storage periods. There was a significant difference between the genotypes for Cu and Zn. The Cu contents presented linear reduction and Zn presented linear increase, in the sorage periods. However, the genotype Doce had no effect (P=0.9531) on Zn in the post-harvest storage periods (Table 3, page 267).
For Fe, there was no effect (P=0.1208) of the storage periods. The cactus pear genotypes presented significant differences, with OEM showing the highest content (41.66 ± 4.73 mg kg-1) in all storage periods. Initially, the Mn content did not present significant difference between the genotypes, but the mean values along the storage periods showed linear increase (P<0.05), with concentration effect of this nutrient caused by the water loss.
Discussion
The loss of water (Figure 1, page 264) in the genotype Baiana is related to the respiratory activities through plant evapotranspiration. Plants with crassulacean acid metabolism (CAM) avoid water loss through evaporation by keeping the stomata closed during the day 12, thus reducing gas exchange. However, after harvesting, if opened, the stomata can no longer close. The stomatal opening throughout the entire storage period was confirmed by the emergence of new sprouts of the cactus pear genotypes, mainly in the Nopalea genus ones. The opening of the stomata may increase immediately after harvest 9. With stomata open, the vapor pressure deficit between the forage and the air is high, but thanks to their structural and functional characteristics, cacti are able to consume and lose the minimum water amount they store 18, even with the stomata opened. The increase in ambient temperature promotes an exponential increase in respiration, since the higher the temperature, the greater the metabolic activity of the tissues 15.
Research testing the effect of the post-harvest storage period on DM content and chemical composition of the cactus pear, did not find significant difference between the genotypes Gigante and Redonda up to 16 days of storage 19, but for the genotype Doce, the period of 12 days presented higher contents than day 0.
The water loss resulted in a concentration of CP in OEM (Table 1, page 265) due to the respiratory activity of the plant, which results in a decrease in the soluble carbohydrate contents, while CP concentrations are not affected by respiration, increasing in proportional terms. That cactus pear from the Opuntia genus presented higher CP content than Nopalea, ranging from 54.2 to 52.1 g kg-1 for Gigante and Redonda, respectively, and 43.1 g kg-1 for Doce 4. Differently from the present study, researchers did not find statistical difference for EE up to 21 days of storage post-harvest in OEM (23). Among the genotypes, Baiana presented the highest levels of EE up to 45 days. As lipids are the most susceptible constituents to chemical degradation and directly influence the drying and preservability of the stored product, this similarly justifies the highest water and DM losses in this genotype 16.
The decrease in carbohydrates is attributed to the shoots that appeared during the post-harvest storage period, due to the dilution effect, caused by the increase in new cladodes, which are poor in structural carbohydrates 22. As in hay making, unfavorable environmental conditions lead to the loss of soluble carbohydrates through continuous cellular respiration, and when the cladodes are stored with moisture content higher than the recommended levels (between 150 and 180 g kg-1 of DM), they easily heat up and result in increased levels of NDF, ADF and lignin 5. The increase in MM content in this period can be explained by the wearing of OM caused by higher temperatures up to 30 days during the entire study period.
When considering the plant development stage, there is an increase in the contents of structural compounds, such as hemicellulose, cellulose and lignin and consequent decrease in cellular content (Table 2, page 266). Researchers reported higher lignin contents in the genotypes Gigante and Redonda in comparison to Doce, and did not find significant differences for hemicellulose and cellulose contents, in Gigante, Redonda and Miúda in different crop densities 4. Put Silva et al. (2017) found a significant difference of crude fiber in genotype Doce between the periods 0 and 12 days of storage, with contents of 123.4 and 96.5 g kg-1, but not differing from the other periods (4, 8 and 16 days).
The water loss over time may have led to the concentration of P contents, (Table 3, page 267) since the genotypes Baiana and OEM lost more water and presented the highest concentrations of P (0.94 and 1.00 g kg-1, respectively). The reduction in the Ca content over time may be related to the emergence of shoots in the post-harvest storage. In addition, Ca influences the structure and strength of the cell wall. Reserchers found levels of Ca of 17.0 and 28.2 g kg-1 and P of 0.7 and 0.8 g kg-1 in cactus pear Gigante at 390 and 620 days after planting, respectively, in the treatment without fertilization, which are values very similar to those found in the present study 21.
Sprouts over the storage period may have influenced the decrease in Mg (Table 3, page 267) since it is rapidly transferred to young plants. In contrast, there was no difference for K and Na contents in the studied genotypes, nor for the post-harvest storage periods. Studies evaluating the mineral composition in the harvest of cactus pear cladodes under different spacing and chemical fertilization, also did not find significant difference for K and Na 21. Santos et al. (1992) noted that differences in the Fe content can be explained by the interactions of Fe with other nutrients, notably phosphorus, zinc, molybdenum and copper, which are common. There is little information available in the literature on macro and micronutrient contents in cactus pear and its effects on production, especially when it comes to storage for long post-harvest periods. In general, the reduction in some minerals during post-harvest storage may be due to the action of microorganisms, which degrade part of the constituents over time.
Conclusions
The cactus pear genotypes Doce and Orelha de Elefante Mexicana (OEM) can be stored for a period up to 60 days after harvest, with minimum losses of water content and maintainance of a stable chemical composition, mainly regaring to the variables dry matter, crude protein, ether extract, sodium and potassium.

