










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRIA. Revista de investigaciones agropecuarias
versión On-line ISSN 1669-2314
RIA. Rev. investig. agropecu. vol.44 no.1 Ciudad Autónoma de Buenos Aires abr. 2018
ARTÍCULOS
Interacción entre festuca alta infectada con endófito y Lotus tenuis bajo dos frecuencias de defoliación
Petigrosso, L.R.1; Assuero, S.G.1; Vignolio, O.R.1; Romano, Y.1; Colabelli, M.N.2; Salvat, A.2; Cristos, D.3
1 Facultad de Ciencias Agrarias, Universidad Nacional de Mar del Plata. Ruta Nacional 226 km 73,5, 7620 Balcarce, Argentina. Correo electrónico: petigrosso.lucas@inta.gob.ar; assuero.silvia@inta.gob.ar; vignolio.osvaldo@inta.gob.ar; yesiromano@hotmail.com; colabelli.mabel@inta.gob.ar
2 Instituto Nacional de Tecnología Agropecuaria (INTA), Instituto de Patobiología, Centro de Investigación en Ciencias Veterinarias y Agronómicas (CICVyA), Laboratorio de Toxicología, Castelar (Buenos Aires, Argentina). Correo electrónico: cristos.diego@inta.gob.ar
3 Instituto Nacional de Tecnología Agropuecuaria (INTA), Instituto de Tecnología de Alimentos, Centro de Investigación de Agroindustria (CIA), Laboratorio de Contaminantes Químicos, Castelar (Buenos Aires, Argentina). Correo eléctrico: salvat.adriana@inta.gob.ar
Recibido 25 de octubre de 2016
Aceptado 09 de mayo de 2017
Publicado online 27 de marzo de 2018
RESUMEN
La toxicidad de festuca alta infectada con endófito asexual Epichloë coenophiala, puede ser atenuada cuando se la siembra en mezcla con leguminosas. El objetivo de este trabajo fue simular el establecimiento de pasturas de festuca alta (Fa) infectada puras y consociadas con Lotus tenuis (Lt), y evaluar el efecto de dos frecuencias de defoliación sobre la producción de biomasa aérea, la densidad de macollos de la gramínea y de tallos de la leguminosa durante el año de implantación. El experimento se llevó a cabo al aire libre en contenedores plásticos de 0,6 m x 0,4 m x 0,2 m. El 24 de febrero de 2015, se sembraron 52 semillas de Fa infectada y 12 de Lt por contenedor en el caso de la mezcla (ej. 64 plantas en total), y el mismo número de semillas de cada especie en los respectivos monocultivos (ej. 52 y 12 plantas en los monocultivos de Fa y Lt, respectivamente). El factor experimental controlado fue la frecuencia de defoliación (a 70 mm de altura): alta y baja (cada 7-9 y 14-21 días, respectivamente, variando el intervalo con la estación del año). Se empleó un diseño completamente aleatorizado con 3 repeticiones y medidas repetidas en el tiempo (18 contenedores en total). En cada contenedor se determinó la biomasa aérea y se contó el número de macollos en Fa y el de tallos en Lt. La biomasa aérea cosechada acumulada de Fa y Lt en monocultivo y en mezcla durante el período experimental varió dependiendo de la frecuencia de defoliación y el tipo de pastura. En general, y en coincidencia con la estación de crecimiento de cada una de las especies, en los primeros cortes los monocultivos de Fa y las pasturas mezcla acumularon más biomasa aérea que los monocultivos de Lt, lo que se revirtió al final del experimento. La densidad de macollos en Fa y de tallos en Lt, tanto en monocultivos como en la mezcla, no resultó afectada por la frecuencia de defoliación, salvo al final del período experimental cuando la densidad de tallos de Lt en el monocultivo superó a la observada en la mezcla con Fa. Tanto en monocultivo como en mezcla, y contrariamente a lo observado en los cortes precedentes, la densidad de macollos en Fa y de tallos en Lt al final del período experimental tendió a ser mayor con baja frecuencia de defoliación.
Palabras clave: Schedonorus arundinaceus; Epichloë coenophiala; Leguminosa; Competencia.
ABSTRACT
The toxicity of tall fescue infected with asexual endophyte Epichloë coenophiala can be attenuated when the species is sown in mixture with legumes. The aim of this work was to simulate the establishment of pastures of infected tall fescue (Fa), both pure and consociated with Lotus tenuis (Lt), and to evaluate the effect of two defoliation frequencies on aerial biomass production and the number of tillers in Fa and stems in Lt during the year of implantation. The experiment was conducted outdoors in plastic containers (0.6 m x 0.4 m x 0.2 m). On 24 February 2015, 52 seeds of infected Fa and 12 seeds of Lt were sown in each container for the mixed pastures (i.e. 64 plants in total), and the same number of seeds of each species was used for the respective monocultures (i.e. 52 and 12 plants in Fa and Lt monocultures, respectively). The controlled experimental factor was the defoliation frequency (70 mm cutting height): high and low (every 7-9 and 14-21 days, respectively, the interval depending on the season). A completely randomized experimental design with 3 replicates and repeated measurements in time was used (18 containers in total). Accumulated aboveground harvested biomass of each container and the number of tillers of Fa and stems of Lt plants were determined during the experimental period. Accumulated biomass of Fa and Lt in monocultures and mixtures during the experimental period varied depending on the defoliation frequency and/or type of pasture. In general, and according to the growing season of the species, at the beginning of the experimental period Fa monocultures and mix pastures accumulated more biomass than Lt monocultures, while the opposite took place by the end of it. Tillers density in Fa and stems density in Lt for both monocultures and mix pastures were not affected by defoliation frequency, except by the end of the experimental period when the stems density in Lt monocultures was higher than the observed in mix pastures. Conversely to what it was observed previously, at the end of the experimental period tillers density in Fa and stems density in Lt, for both monocultures and mix pastures, tended to be higher with low than with high defoliation frequency.
Keywords: Schedonorus arundinaceus; Epichloë coenophiala; Legume; Competition.
INTRODUCCIÓN
La presencia de endófitos asexuales Epichloë spp. (Leuchtmann et al., 2014), ex Neotyphodium spp. (Glenn et al., 1996), en pasturas de festuca alta, Schedonorus arundinaceus (Schreb.) Dumont (= Festuca arundinacea Schreb.) (Hoveland et al., 1999; Clay y Schardl, 2002) otorga beneficios a las gramíneas en su crecimiento y tolerancia a estreses bióticos y abióticos (Malinowski y Belesky, 2000; White y Torres, 2009; Omacini et al., 2013). Sin embargo, debido a la producción de alcaloides tóxicos, principalmente ergovalina (Schard y Phillips, 1997; Evans et al., 2004), los vacunos, equinos y ovinos que consumen festuca alta infectada sufren una enfermedad clínica conocida como intoxicación con festuca alta o festucosis (Bacon et al., 1977; De Battista et al., 1995). Esta enfermedad les provoca menor ganancia diaria de peso, cambios hormonales, alteraciones reproductivas, síndrome gangrenoso e hipertérmico (Stuedemann y Hoveland, 1988).
La intoxicación debida al consumo de gramíneas infectadas con endófito tiene importante repercusión económica en Estados Unidos, Nueva Zelanda y Argentina, entre otros países; y se han estimado grandes pérdidas económicas (De Battista et al., 1995; Strickland et al., 2011). Sin embargo, en nuestro país no son comunes las intoxicaciones a escala de establecimientos ganaderos debido a diferentes estrategias de manejo implementadas por los productores (De Battista et al., 1997). Por una parte, por ejemplo, la toxicidad puede ser atenuada si la gramínea se siembra en mezcla con otras gramíneas libres de endófito (Evans et al., 2012) o leguminosas tales como Lotus tenuis (Waldst. et Kit. Ex Willd) y Trifolium repens L. (De Battista et al., 1997; Manzini, 1991; Tekeli y Ateş, 2005). Por otra parte, García Parisi et al. (2015) hallaron que la presencia del endófito en plantas de Lolium multiflorum L. no modificaría el crecimiento ni la fijación de nitrógeno de Trifolium repens L., y que la presencia simultánea de endófitos en gramíneas y bacterias en leguminosas resultaría positiva para la comunidad en términos de utilización de nitrógeno y productividad.
En las pasturas mezcla con leguminosas las proporciones iniciales de las especies tienden a cambiar con el tiempo, a favor de la biomasa de las gramíneas libres (León y Oesterheld 1982; Mela, 2003) o infectadas (Sutherland y Hoglund, 1989; Eerens et al., 1998; Hoveland et al., 1999; Vazquez de Aldana et al., 2013) en detrimento de las dicotiledóneas. Este cambio en las proporciones de las mezclas puede deberse a múltiples causas, como por ejemplo: especie hospedante, especies que componen la pastura, régimen de uso de la pastura, factores climáticos, disponibilidad de nutrientes, preferencia de los herbívoros y desplazamiento competitivo (León y Oesterheld, 1982, Dirihan et al., 2015). Las prácticas de manejo tendientes a mantener una cierta proporción gramínea-leguminosa bajo pastoreo deben considerar los factores antes mencionados. En tal sentido, la habilidad competitiva y la proporción de las especies en la pastura pueden ser moduladas por la frecuencia de defoliación (Manzini, 1991; Hoveland et al., 1999; Stuedemann y Seman, 2005). Así, por un lado, manejos que permitan mantener una determinada proporción de la leguminosa en la mezcla reducirían la toxicidad de la pastura por “efecto de dilución” (Salminen y Grewal, 2002; Salminen et al., 2003). Por otro lado, la toxicidad de festuca alta infectada aumenta con la edad de las plantas y con defoliaciones menos frecuentes e intensas (Stuedemann y Seman, 2005). Por lo tanto, el manejo de la defoliación es una herramienta simple y de bajo costo que permitiría reducir la habilidad competitiva de la gramínea, garantizar el crecimiento y persistencia de la leguminosa y, con ello, disminuir la toxicidad de las pasturas (Salminen y Grewal, 2002; Salminen et al., 2003). En este sentido, son escasos los trabajos publicados que investigan el efecto de la defoliación de pasturas mezcla de festuca infectada y leguminosas sobre dinámica de producción de sus componentes.
El objetivo de este trabajo fue simular el establecimiento de pasturas de festuca alta infectada, puras y consociadas con L. tenuis, y evaluar los efectos de la frecuencia de defoliación sobre la producción de biomasa aérea y la densidad de macollos/tallos de cada especie.
MATERIALES Y MÉTODOS
Sitio experimental
Se realizó un experimento al aire libre en la Unidad Integrada Balcarce (Facultad de Ciencias Agrarias UNMdP– Estación Experimental Agropecuaria INTA Balcarce) 37º45’47,94”S, 58º17’38,82”W, 130 m s.n.m. Se utilizó semilla de festuca alta infectada (Fa) proveniente de una pastura del partido de Mar Chiquita (37º 32’ S; 57º 55’ O, identificada previamente por Petigrosso et al., 2013) cosechada el 15 de diciembre de 2014 (PG 69%) y semilla escarificada de L. tenuis (Lt) cultivar Chajá (PG 90%) inoculada con Rhizobium loti (Cepa 733). Se corroboró la condición de infección endofítica de las semillas de Fa mediante análisis microscópico según el protocolo empleado por Saha et al. (1988) y Peretti (1994). Se emplearon 18 contenedores plásticos (unidades experimentales, UE) de 0,6 m x 0,4 m x 0,2 m, conteniendo suelo tamizado y homogeneizado, proveniente del horizonte A de un Argiudol típico (53 ppm de P Bray-1; 4,3% MO y 30,1 ppm N-NO3 -). El 24 de febrero de 2015 se efectuó la siembra de pasturas mezcla y monocultivos (mesocosmos) empleando una placa plástica perforada que permitió ubicar en los monocultivos de Fa y en las pasturas mezcla 13 semillas de Fa a 5 mm de profundidad en cada uno de cuatro surcos distanciados 12 cm entre sí (ej. densidad de siembra 200 plantas/m2), y en los monocultivos de Lt y en las mezclas 4 semillas de Lt en cada uno de los tres entresurcos (ej. densidad de siembra 24 plantas/ m2) (figura 1). Las fallas en la emergencia de plántulas de Fa se subsanaron dentro de los 20 días desde la siembra mediante trasplantes de plántulas extracultivadas en almácigos. La fecha de emergencia promedio fue el 10 de marzo de 2015 (Siembra-emergencia: 159 °Cd). A los 30 días de emergidas las plántulas de Fa se corroboró la presencia del endófito mediante el análisis microscópico de un macollo por planta (Latch et al., 1984). El 15 de abril de 2015 (173 ºCd desde la emergencia) se realizó un corte de “limpieza” para homogenizar la altura de las UE y a partir de esa fecha se aplicaron dos niveles de frecuencia de defoliación a 70 mm de altura desde el nivel del suelo: alta y baja (cada 7-9 días y cada 14-21 días, respectivamente, variando el intervalo con la estación del año). Los contenedores se mantuvieron libres de malezas y con adecuada disponibilidad de agua durante el período experimental. El tiempo térmico acumulado (°Cd) se calculó como la sumatoria de las diferencias entre las temperaturas medias diarias y la temperatura base (4 ºC; Colabelli et al., 1998). Para ello se utilizaron los registros de temperatura del aire en casilla meteorológica a 1,50 m de altura de la Estación de Agrometereología de la EEA Balcarce del INTA, ubicada aproximadamente a 300 m del sitio experimental. No se observaron ataques de plagas ni patógenos durante el desarrollo del experimento.
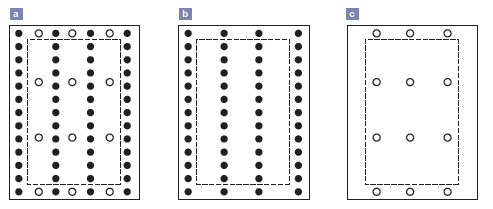
Figura 1. Arreglo espacial de las semillas en los mesocosmos (unidad experimental). a) Pastura mezcla festuca alta – lotus, b) monocultivo de festuca alta y c) monocultivo de lotus. Los círculos negros representan las semillas de festuca alta E+ y los círculos vacíos las semillas de lotus. La línea punteada encierra la zona de muestreo (en pasturas mezcla: 22 plantas de festuca alta y 6 plantas de lotus por contenedor, en monocultivo de festuca 22 plantas y en monocultivo de lotus 6 plantas).
Determinación de la biomasa aérea acumulada y del número de macollos y tallos
El primer corte experimental se realizó 21 de abril de 2015 para los tratamientos con alta frecuencia de defoliación y el 28 de abril de 2015 para los tratamientos con baja mediante análisis de varianza con medidas repetidas en el tiempo. Se realizaron comparaciones de medias (LSD p<0,05) cuando se encontraron diferencias significativas (α=0,05). Debido a que de las variables respuesta no cumplieron con los supuestos de normalidad ni homogeneidad de varianzas debieron transformarse (ej. ln de la densidad de macollos y de tallos; y ln de la biomasa aérea acumulada+ 1). Todos los análisis se realizaron empleando el software estadístico R (R Development Core Team, 2015). Los modelos se ajustaron con la función lme del paquete nlme (Pinheiro et al., 2012).
Tabla 1. Fechas de corte y tiempo térmico acumulado (TT, °Cd, temperatura base 4 °C) desde el corte limpieza el 15 de abril de 2015 (173 °Cd acumulados desde la emergencia el 10 de marzo de 2015) para ambas frecuencias de defoliación: alta (A, cada 7-9 días) y baja (B, cada 14-21 días).

RESULTADOS Y DISCUSIÓN
Biomasa aérea cosechada acumulada
En la figura 2 se observa la acumulación de la biomasa cosechada en los monocultivos y en la mezcla para ambas frecuencias de defoliación. Se detectó interacción significativa entre el tipo de pastura, frecuencia de defoliación y fecha de corte (p<0,0001). Se puede observar que hasta 593 ºCd no hubo diferencias significativas en la producción de biomasa acumulada debido a la frecuencia de defoliación, pero sí al tipo de pastura. En este sentido, se observó que las producciones de biomasa de los monocultivos de Fa y de las mezclas fueron similares y superiores a las de los monocultivos de Lt. Entre los 1034 ºCd y los 1307 ºCd se registró una interacción significativa entre el tipo de pastura y la frecuencia de defoliación. Ello se debió a que la biomasa acumulada fue mayor en los monocultivos de Fa con baja frecuencia, lo contrario ocurrió en los monocultivos de Lt, y no hubo diferencias entre frecuencias de defoliación en las mezclas. Por una parte, la producción de biomasa total acumulada al final del experimento (1604 ºCd) fue mayor en los monocultivos de Lt independientemente de la frecuencia de defoliación. Las biomasas acumuladas en los monocultivos de Fa y de las pasturas mezcla no presentaron diferencias significativas entre sí, aunque la biomasa del monocultivo de Fa con alta frecuencia de defoliación tendió a ser menor que las restantes (p=0,0901). Por otro lado, se observó que en las pasturas mezclas la contribución relativa de Lt a la biomasa acumulada total fue mayor con baja que con alta frecuencia de defoliación (35% y 24%, respectivamente).
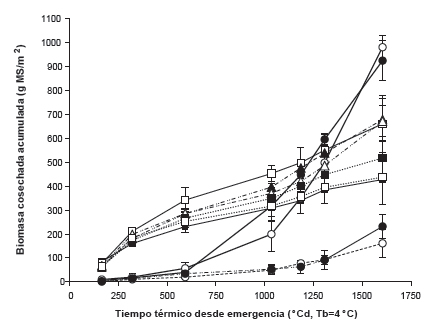
Figura 2. Acumulación de biomasa cosechada (g MS/m2, media ± EE, n=3) en pasturas en mesocosmos de festuca alta infectada (Fa) en monocultivo y en mezcla con Lotus tenuis (Lt), Lt en monocultivo y en mezcla con Fa y mezcla de ambas especies para dos frecuencias de defoliación: alta (A, 7-9 días) y baja (B, 14-21 días). Referencias: monocultivo de Fa en A __■__ y B __□__; monocultivo de Lt en A __●__ y B __○__; Fa en mezcla en A ….■…. y B ….□…., Lt en mezcla en A _ _●_ _ y B _ _○_ _; y mezcla en A _ _▲_ _ y B_ _Δ_ _.
Al comparar la producción de biomasa de Fa en monocultivo y en mezcla con Lt se observó que ninguno de los efectos principales ni sus interacciones fueron significativos (p>0,05). La falta de respuesta a la frecuencia de defoliación hallada en el presente experimento en el monocultivo de Fa durante el invierno, se contrapone con los resultados de Hart et al. (1971) quienes encontraron una mayor producción de biomasa de Fa cuando la frecuencia de defoliación era baja. Sin embargo, por un lado, las frecuencias de defoliación estudiadas por estos autores fueron más contrastantes que las estudiadas en este trabajo, la frecuencia alta fue similar (7 días), mientras que en la baja las pasturas se defoliaban una vez por mes. Por otro lado, la mayor producción del monocultivo de Fa a baja frecuencia de defoliación contradice lo reportado por Kerrisk y Thomson (1990) quienes hallaron una mayor producción de biomasa de Fa cuando la pastura fue defoliada con alta frecuencia. Sin embargo, la frecuencia alta fue similar a la baja de este experimento (15 días) y la baja frecuencia correspondió a intervalos de 30 días. En el presente trabajo se aplicaron dos frecuencias de defoliación relativamente altas ya que se ha sugerido que una alta frecuencia de defoliación podría reducir las concentraciones de ergoalcaloides en plantas de Fa infectada y, por ende, su toxicidad dado que las plantas destinarían una alta proporción de carbohidratos para el rebrote, dejando menos fotoasimilados disponibles para la síntesis de alcaloides por parte del endófito (Belesky y Hill, 1997). La biomasa acumulada de Lt no fue afectada en los tres primeros cortes ni en el monocultivo ni en la mezcla por la frecuencia de defoliación ni por el tipo de pastura (p>0,05). Entre los 1034 ºCd y 1307 ºCd se registró interacción entre el tipo de pastura y la frecuencia de defoliación (p<0,05) dado que Lt en monocultivo produjo más biomasa con alta frecuencia de defoliación que con baja frecuencia, mientras que en las mezclas no se registraron diferencias entre frecuencias. En el último corte (1604 °Cd), la acumulación de biomasa de Lt fue mayor en los monocultivos que en las mezclas (p<0,0001) y no hubo efecto significativo de la frecuencia de la defoliación (p=0,1096).
La escasa producción de biomasa aérea de Lt en los primeros cortes estuvo relacionada con las bajas tasas de crecimiento características de la especie en ese momento del año (Sevilla et al., 1996; Vignolio y Fernández, 2011). Sin embargo, en la época primavero-estival, cuando esta especie presenta las mayores tasas de crecimiento (Colabelli y Miñón, 1993; Vignolio et al., 2010), la producción de biomasa acumulada de Lt aumentó hasta superar en el último corte a las demás pasturas, independientemente de la frecuencia de defoliación. Acuña y Cuevas (1999) tampoco hallaron una respuesta significativa de la frecuencia de defoliación (6 y 8 semanas entre defoliaciones) en un monocultivo de Lt bajo riego sobre la producción de materia seca. La menor producción de biomasa de Lt en mezcla podría deberse a la mayor habilidad competitiva de la gramínea con respecto a la leguminosa (Muslera y Ratera, 1991; Hernández et al., 2005).
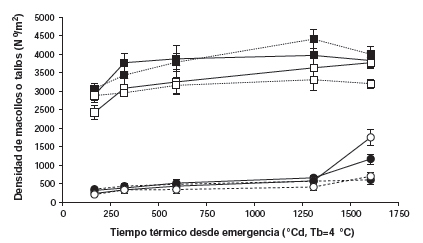
Figura 3. Dinámica de la densidad de macollos o tallos en pasturas en mesocosmos de festuca alta infectada (Fa) y Lotus tenuis (Lt) en monocultivo y en mezcla bajo dos frecuencias de defoliación: alta (A, 7-9 días) y baja (B, 14-21 días). Referencias: monocultivo de Fa en A __■__ y B __□__; monocultivo de Lt en A __●__ y B __○__; Fa en mezcla en A ….■…. y B ….□…., Lt en mezcla en A _ _●_ _ y B _ _○_ _.
Densidad de macollos de festuca alta infectada y tallos de Lotus tenuis
En la figura 3 se observa la dinámica de la densidad de macollos de Fa y de tallos de Lt en monocultivo y en mezcla para ambas frecuencias de defoliación. En el caso de Fa, ninguno de los efectos principales ni sus interacciones fue significativo (p>0,05). Sin embargo, se observó que la densidad de macollos de Fa tendió a ser mayor en alta frecuencia de defoliación tanto en monocultivo como en mezcla (p=0,0720). Estos resultados coinciden con los publicados por Parsons et al. (1983) y Lemaire y Chapman (1996) quienes hallaron que las pasturas manejadas con alta frecuencia de defoliación desarrollarían una estructura con una mayor densidad de macollos de menor tamaño que aquellas sometidas a bajas frecuencias de defoliación, lo que se explicaría por la ley de los -3/2 o de la compensación tamaño/densidad (Matthew et al., 1995).
Por un lado, la densidad de tallos de Lt solamente fue afectada por el tipo de pastura (p=0,042) y la fecha de corte (p<0,0001). Hasta los 1307 ºCd no hubo diferencias en la densidad de tallos de Lt debido al tipo de pastura ni a la frecuencia de defoliación, aunque hasta ese momento tendió a ser mayor con alta frecuencia de defoliación (p=0,0802). Al final del experimento (1604 ºCd), y en coincidencia con lo observado en la producción de biomasa aérea, la densidad de tallos de Lt fue mayor en el monocultivo que en la mezcla. La falta de respuesta del Lt a la frecuencia de defoliación también fue reportada por Acuña y Cuevas (1999) para monocultivos de Lt bajo riego sometidos a frecuencias de defoliación más bajas (6 y 8 semanas) que las de este experimento. Por otro lado, y contrariamente a lo encontrado en los cortes precedentes, a 1604 ºCd la densidad de tallos de Lt en monocultivo tendió a ser mayor a baja frecuencia de defoliación (p=0,096), probablemente porque en la frecuencia de defoliación alta el intervalo de corte fue demasiado breve, restringiendo la recuperación del área foliar y limitando la producción de fotosintatos necesarios para el rebrote de las yemas de la corona (Colabelli y Miñon, 1994; Acuña y Cuevas, 1999).
CONCLUSIONES
En función de los resultados obtenidos podemos concluir que:
1.La frecuencia de defoliación y el tipo de pastura afectaron a la biomasa aérea cosechada acumulada de Fa y Lt en monocultivo y en mezcla en forma diferencial durante el período experimental. Por un lado, la biomasa total acumulada en los monocultivos de Fa y de las pasturas mezcla no presentaron diferencias significativas entre sí, aunque la biomasa del monocultivo de Fa con alta frecuencia de defoliación tendió a ser menor que las restantes. Por otro lado, la biomasa aérea cosechada acumulada de Lt en monocultivo fue baja al principio del período experimental, pero superó a la de las pasturas mezcla y monocultivos de Fa al final del mismo debido a las mayores tasas de crecimiento de la leguminosa en la primavera.
2.La densidad de macollos de Fa y de tallos de Lt, tanto en monocultivos como en la mezcla, no resultó afectada por la frecuencia de defoliación aunque tendió a ser mayor con alta frecuencia. Solamente al final del experimento, el número de tallos en el monocultivo de Lt superó a la observada en la mezcla con Fa, lo que podría deberse a la mayor habilidad competitiva de la gramínea que afectaría negativamente el crecimiento de Lt y, a su vez, tanto en monocultivo como en mezcla y contrariamente a lo observado en cortes precedentes tendió a ser mayor con baja que con alta frecuencia de defoliación.
Los resultados de este experimento indican que las pasturas, mezcla de festuca alta y Lotus tenuis, defoliadas con una frecuencia de aproximadamente 15 días tenderían a presentar una mayor proporción de biomasa de lotus que las defoliadas más frecuentemente. Estos resultados deberían corroborarse en futuros experimentos en los que se analice, además de la dinámica de producción de biomasa de ambos componentes de la mezcla, el contenido de ergoalcaloides del forraje cosechado para verificar si disminuye la toxicidad por un efecto de dilución.
AGRADECIMIENTOS
El presente estudio se desarrolló en el marco del Proyecto 15/A473, AGR 472/15 de la Facultad de Ciencias Agrarias de la Universidad Nacional de Mar del Plata. Los autores agradecen a la empresa KWS por el suministro de la semilla de Lotus tenuis empleada.
BIBLIOGRAFÍA
1. ACUÑA, P.H.; CUEVAS, G.C. 1999. Efecto de la altura y frecuencia de la defoliación, bajo corte y pastoreo, en el crecimiento y productividad de tres especies del género Lotus en suelos arcillosos. Agricultura Técnica. 59: 296–308.
2. BACON, C.W.; PORTER, J.K.; ROBBINS, J.D.; LUTTRELL, E.S. 1977. Epichloe typhina from toxic tall fescue grasses. App. Environ. Microbiol. 34:576–581.
3. BELESKY, D.P.; HILL, S. 1997. Defoliation and Leaf Age Influence on Ergot Alkaloids in Tall Fescue. Ann. Bot. 79: 259–264
4. CLAY, K.; SCHARDL, C. 2002. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 160: 99–127.
5. COLABELLI, M.; AGNUSDEI, M.; MAZZANTI, A.; LABREVEUX, M., 1998. El proceso de crecimiento y desarrollo de gramíneas forrajeras como base para el manejo de la defoliación. INTA EEA Balcarce. Buenos Aires, Argentina. Boletín Técnico N.° 148. 1–14 pp.
6. COLABELLI, M.R.; MIÑON, D.P. 1993. Métodos de intersiembra de Lotus tenuis y Trifolium repens en pastizales de la Pampa Deprimida bonaerense. Rev. Arg. Prod. Anim. 13: 225–233.
7. COLABELLI, M.; MIÑÓN, D. 1994. Rendimiento y cambios botánicos de pasturas de Lotus tenuis puro y en mezcla bajo régimen de corte. Agricultura Técnica. 54: 39–45.
8. DE BATTISTA, J.; PERETTI, A; CARLETTI, S.; RAMIREZ, A.; COSTA, M; SCHULTZ, L. 1995. Evolución de la incidencia de la infección de Acremonium coenophialum en la oferta de semilla de festuca alta en Argentina. Período 1987-1994. Rev. Arg. Prod. Anim. 15: 300–302.
9. DIRIHAN, S.; HELANDER, M.L.; SALONIEMI, I.; GUNDEL, P.E.; SAIKKONEN, K. 2015. Effects of systemic fungal endophytes on the performance of meadow fescue and tall fescue in mixtures with red clover. Grass Forage Sci. 70: 465–473.
10. DONAGHY, D.J.; TURNER, L.R.; ADAMCZEWSKI, K.A. 2008. Effect of defoliation management on water-soluble carbohydrate energy reserves, dry matter yields, and herbage quality of tall fescue. Agr. J. 100: 122–127.
11. EERENS, J.P.; LUCAS, R.J.; EASTON, H.S.; WHITE, J.G. 1998. Influence of the ryegrass endophyte (Neotyphodium lolii) in a coolmoist environment III. Interaction with clover. NZ. J. Agr. Res. 41: 201–207.
12. EVANS, T.J.; ROTTINGHAUS, G.E.; CASTEEL, S.W. 2004. Ergot. In: Plumlee, K.H. (ed.), Clinical Veterinary Toxicology, Mosby, St. Louis, MO. 239–243 pp.
13. EVANS, T.J.; BLODGETT, D.J.; ROTTINGHAUS, G.E. 2012. Fescue toxicosis. Vet Toxicol. 87: 1166–1177.
14. GARCÍA PARISI, P.A.; LATTANZI, F.A.; GRIMOLDI, A.A.; OMACIN, M.I. 2015. Multi-symbiotic systems: functional implications of the coexistence of grass–endophyte and legume–rhizobia symbioses. Oikos. 124: 553–560.
15. GLENN, A.E.; BACON, C.W.; PRICE, R.; HANDIL, R.T. 1996. Molecular phylogeny of Acremonium and its taxonomic implications. Mycol. 88: 369–383.
16. HART, R.H.; CARLSON, G.E.; MCCLOUD, D.E. 1971. Acumulative effects of cutting management on forage yields and tiller densities of tall fescue and orchard grass. Agr. J.63: 895–898.
17. HERNÁNDEZ, S.R.; PÉREZ, J.O.; GUILLÉN, R.J.; CASTRO, E.H. 2005. Manejo de praderas asociadas de gramíneas y leguminosas para pastoreo en el trópico. Revista Electrónica de Veterinaria REDVET. (Disponible: http://www.veterinaria.org/revistas/redvet verificado: 10 de marzo de 2015). [ Links ]
18. HOVELAND, C.S.; BOUTON, J.H.; DURHAM, R.G. 1999. Fungal endophyte effects on production of legumes in association with tall fescue. Agr. J. 91: 897–903.
19. KEMP, S. 2004. Fescue to the rescue. Aust Dairy Farmer. 19:78. [ Links ]
20. KERRISK, J.J.; THOMSON, N.A. 1990. Effect of intensity and frequency of defoliation on growth of ryegrass, tall fescue and phalaris. Proc. New Zeal. Grass. Assoc. 51:135–138.
21. LATCH, G.C.M.; CHRISTENSEN, M.J.; SAMUELS, G.J. 1984. Five endophytes of Lolium and Festuca in New Zealand. Mycotaxon 20: 535–550.
22. LEMAIRE, G.; CHAPMAN, D. 1996. Tissue flows in grazed plant communities. En: HODGSON, J.; ILLIUS, A.W. (Eds.). The ecology and management of grazing system. CAB International. 3–36 pp.
23. LEÓN, R.J.C.; OESTERHELD, M. 1982. Envejecimiento de pasturas implantadas en el norte de la Depresión del Salado. Un enfoque sucesional. Rev. Fac. Agron. 3: 41–49.
24. LEUCHTMANN, A.; BACON, C.W.; SCHARDL, C.L.; WHITE,J.F.; TADYCH,M. 2014 . Nomenclatural realignment of Neotyphodium species with genus Epichloë. Mycol. 106: 202–215.
25. MALINOWSKI, D.; BELESKY, D. 2000. Adaptations of endophyte- infected cool-season grasses to environmental stresses: mechanisms of drought and mineral stress tolerance. Crop Sci. 40: 923-940. [ Links ]
26. MANZINI, N.A. 1991. Estudios sobre Acremonium sp. en festuca. INIA. Unidad de Difusión e Información Tecnológica. Montevideo- Uruguay. Serie Técnica N.º 8. 16 p. [ Links ]
27. MATTHEW, C.; LEMAIRE, G.; SACKVILLE HAMILTON, R.N. 1995. A modified self-thinning equation to describe size density relationships for defoliation swards. Ann. Bot. 76: 579-587. [ Links ]
28. MELA, T. 2003. Red clover in a mixture with grasses: yield, persistence and dynamics of quality characteristics. Agr. Food Sci. Finland. 12: 195-212. [ Links ]
29. MUSLERA PARDO, E.; RATERA GARCÍA, C. 1991. Las gramíneas. En: MUSLERA PARDO, E.; RATERA GARCÍA, C. (Eds.). Praderas y forrajes: producción y aprovechamiento. Mundi- Prensa, Madrid. 29-114 pp. [ Links ]
30. OMACINI, M.; GUNDEL, P.; SEMMARTIN M. G. 2013. Huellas de la simbiosis pasto-endófito en el agroecosistema. En: GARCÍA DE SALAMONE, I.E.; VÁZQUEZ, S.; PENNA, C.; CASSAN, F. (Eds.). Rizósfera, Biodiversidad y Agricultura Sustentable. División de Microbiología Agrícola y Ambiental, Asociación Argentina de Microbiología. Buenos Aires, Argentina. 73 p. [ Links ]
31. PARSONS, A.J.; LEAFE, E.L.; COLLET, B.; STILES, W. 1983. The physiology of grass production under grazing. I. Characteristics of lead and canopy photosynthesis of continuously-grazed swards. J. Appl. Ecol. 20: 117-126. [ Links ]
32. PERETTI, A. 1994. Manual para análisis de semillas. Hemisferio Sur, Buenos Aires. 281 p. [ Links ]
33. PINHEIRO, J.; BATES, D.; DEBROY, S.; SARKAR, D. 2012. R Development Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-103. [ Links ]
34. PETIGROSSO, L.R.; COLABELLI, M.N.; FERNÁNDEZ, O.N.; ISPIZÚA, V.; CENDOYA, M.G. 2013. Incidence of the endophyte fungus Neotyphodium coenophialum in pastures of tall fescue differing in age and soil characteristics. Afr. J. Agric. Res. 8 (22): 2655-2662. [ Links ]
35. R DEVELOPMENT CORE TEAM. 2015. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. (Disponible: http://www.R-project.org/ verificado: 15 de junio de 2015). [ Links ]
36. SAHA, C.D.; JACKSON, M.A.; JOHNSON-CICALESE, J.M. 1988. A rapid staining method for detection of endophytic fungi in turf and forage grass. Phytopathol. 78: 237-239 [ Links ]
37. SALMINEN, S.O.; GREWAL, P.S. 2002. Does decreased mowing frequency enhance alkaloid production in endophytic tall fescue and perennial ryegrass? J. Chem. Ecol. 28: 939-950. [ Links ]
38. SALMINEN, S.O.; GREWAL, P.S.; QUIGLEY, M.F. 2003. Does mowing height influence alkaloid production in endophytic tall fescue and perennial ryegrass? J. Chem. Ecol. 29: 6-10. [ Links ]
39. SCHARDL, C.L.; PHILLIPS, T.D. 1997. Protective grass endophytes. Where are they from and where are they going? Plant Dis. 81: 430-438. [ Links ]
40. SEVILLA, G.H.; FERNÁNDEZ, O.N.; MIÑON, D.P.; MONTES, L. 1996. Emergence and seedling survival of Lotus tenuis in Festuca arundinacea pastures. J. Range Manage. 49: 509-511. [ Links ]
41. STRICKLAND, J.R.; LOOPER, M.L.; MATTHEWS, J.C.; ROSENKRANS, C.F.; FLYTHE, M.D. JR.; BROWN, K.R. 2011. St. Anthony’s fire in livestock: Causes, mechanisms, and potential solutions. J. Anim. Sci. 89: 1603-1626.
42. STUEDEMANN, J.A.; HOVELAND, C.S, 1988. Fescue endophyte: History and impact on animal agriculture. J. Prod. Agr.1:39-44. [ Links ]
43. STUEDEMANN, J.A.; SEMAN, D.H. 2005. Integrating genetics, environment, and management to minimize animal toxicoses. En: ROBERTS, C.A.; WEST, C.P.; SPIERS, D.E. (Eds). Neotyphodium in Cool-Season Grasses. Blackwell, Oxford, Reino Unido. 305-324 pp. [ Links ]
44. SUTHERLAND, B.L.; HOGLUND, J.H. 1989. Effect of ryegrass containing the endophyte (Acremonium lolii) on the performance of associated white clover and subsequent crops. Proceedings of the New Zealand Grassland Association 50: 265-269. [ Links ]
45. TEKELI, A.S.; ATES, E. 2005. Yield potential and mineral composition of white clover (Trifolium repens L.) - tall fescue (Festuca arundinacea Schreb.) mixtures. J. Cent. Eur. Agr. 6: 27-34. [ Links ]
46. VIGNOLIO, O.R.; CAMBARERI, G.S.; MACEIRA, N.O. 2010. Lotus tenuis (Fabaceae). Productividad y manejo agronómico. Rev. Arg. Prod. Anim. 30: 97-116. [ Links ]
47. VIGNOLIO, O.R.; FERNÁNDEZ, O.N. 2011. Lotus tenuis seedling establishment and biomass production in Flodding Pampa grasslands (Buenos Aires, Argentina). Chil. J. Agr. Res. 71: 96-103. [ Links ]
48. VAZQUEZ DE ALDANA, B.R.; ZABALGOGEAZCOA, I.; GARCIA- CIUDAD, A.; GARCIA-CRIADO, B. 2013. An Epichloe endophyte affects the competitive ability of Festuca rubra against other grassland species. Plant Soil. 362: 201-213. [ Links ] [ Links ]

