











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkThe bee flies (Bombyliidae) constitute one of the largest families of Diptera, and most of its species are parasitoids or hyperparasitoids of other insects, primarily the immature stages of Coleoptera, Hymenoptera, Lepidoptera, and Diptera (Yeates & Greathead, 1997). In Argentina, there are 75 species in 25 genera and 7 subfamilies (Lamas & Evenhuis, 2014; Torretta et al., 2021); however, this figure may be higher because this family has not been extensively studied in this country (Lamas & Evenhuis, 2014).

Figs 1-2: Lateral habitus of species of Toxophora (Bombyliidae, Toxophorinae) reared from nests of wasps obtained in trap-nests in forests in Reserva El Bagual, Formosa province, northern Argentina. 1. Toxophora aurea Macquart. 2. Toxophora leucon Séguy. Black arrow indicates macrochetae in pronotum. Scale bars = 2 mm.
The subfamily Toxophorinae comprises three tribes: Gerontini, Systropodini and Toxophorini (Li et al., 2020). The two latter are recorded in Argentina (Lamas & Evenhuis, 2014). The tribe Toxophorini includes only the genus Toxophora Meigen, with 51 species, of which 11 are present in the Neotropical Region (Evenhuis & Greathead, 2015) and only two in Argentina: T. amicula Seguy and T. leucon Séguy (Lamas & Evenhuis, 2014). A new record, representing the third one in Argentina, for T. aurea Macquart is presented herein. The studied individuals are deposited in the following institutions: Cátedra de Botánica General (FAUBA), Facultad de Agronomía, Universidad de Buenos Aires, Argentina, Museo Argentino de Ciencias Naturales Bernardino Rivadavia (MACN), and Museo de La Plata (MLP).
The species of Toxophora are medium-sized flies, with a stout body with humped thorax, enlarged pronotum and macrochetae and with abdomen strongly convex (Figs. 1-2), wider than thorax at base and covered dorsally with yellow or white scales forming different patterns of spots and marks (Cunha et al., 2011). These species are ectoparasitoids of solitary wasps and bees (Hymenoptera) (Yeates & Greathead, 1997; Lamas et al., 2003), although the natural history is better known for the Nearctic species [summarized inHull (1973)], there were few records for the Neotropical species: Assis & Camillo (1997) mentioned Toxophora sp. in one nest of Pachodynerus brevithorax (Saussure)
(Vespidae), Auko et al. (2014) reared one individual of T. leucon from one nest of Cyphomenes anisitsii (Bréthes) (Vespidae), Rocha-Filho et al. (2019) obtained one individual of T. leucon from a nest of Trypoxylon nitidum F. Smith (Crabronidae), and Rocha-Filho et al. (2020) reared one individual of T. leucon from one nest of Ancistroceroides cf. atripes (Fox) (Vespidae), one individual of an undetermined species of Toxophora from one nest of Hypancistrocerus advena Sausure (Vespidae) and several individuals of Toxophora amphitea Walker from nests of A. cf. atripes, Minixi brasilianum (Saussure) and Pachodynerus guadulpensis (Saussure) (all Vespidae). In a study about the pupae of two Toxophora species (Lamas et al., 2003), Pachodynerus praecox Saussure and Centris analis (Fabricius) (Apidae) were mentioned as hosts of T. leucon and one unidentified species of Parancistrocerus Bequaert (Vespidae: Eumeninae) as host of Toxophora zikani (d’Andretta & Carrera).
Here, we reported data about the biology of T. aurea (Fig. 1) and T. leucon (Fig. 2) in the Reserva El Bagual (26°18’ S, 58°49’ W), Formosa province, in northern Argentina, from individuals reared from wasp nests. The data were obtained from trap-nests placed in two heights (understory and canopy) in two forests within this Biological Reserve (Torretta & Marrero, 2019) and from one nest built in a stem of Urolepis hecatantha (DC.) R.M. King & H. Rob. (Asteraceae).
In total, 1,080 trap-nests, arranged in 72 bundles comprised by 15 canes each were placed in the field from October 2012 to April 2013 and inspected every season (spring: November 11-15; early summer: January 7-11, late summer: February 24-28 and autumn: March 29-April 2). Each trap-nest consisted of one hollow bamboo cane, which was cut so that a nodal septum closed one end of the cane (Aguiar & Garófalo, 2004) . In each forest [see Torretta & Marrero (2019) for a detailed description of the study area], 36 bundles of trap-nests were placed in three transects separated by more than 1,000 m. In each transect the bundles were placed at intervals of 100-200 m and paired at two different heights (one bundle 1-2 m above the ground and another one 8-9 m within -or in the base of- the crown of tallest trees). At each visit, the traps with nests were removed and taken to the laboratory. There, the cells were separated in plastic vials with cotton plugs and numbered from 1 to n (starting from the innermost) and were kept in the laboratory at room temperature (ca. 15-25 °C) until adult emergence. Since trap-nests were collected at intervals 30-45 days, development time can only be estimated with an error of ± 20 days (Thiele, 2005) . Of a total of 660 brood cells in 204 built nests (Torretta & Marrero, 2019), only four brood cells from four nests were attacked by Toxophora spp. The data obtained for Toxophora spp. from trap-nesting programme are presented in Table I.
Moreover, one nest of C. anisitsii was collected in January 2013, on a stem of U hecatantha to 1.2 m above ground. The nest was spheroidal and built with mud containing four brood cells. On 24 March 2013 one male and one female of wasps emerged, and on 4 May 2013 one female of T. leucon.
Both species of Toxophora attacked wasp nests of three different families: Crabronidae, Sphecidae and Vespidae. The information for T. aurea represents the first report about its reproductive ecology. This species attacked Isodontia costipennis (Spinola), a species that uses pappi of Asteraceae (modified calyxes of flowers of this plant family) as cell partitions and captured Orthoptera as prey for their offspring. In the studied forests, I. costipennis built nine nests [one in canopy (two brood cells) and eight in understory (22 brood cells)]; of them T aurea only attacked one brood cell from one nest in the understory (Table I). From other brood cell of same nest reared several individuals of eulophid wasp Melittobia sp. (Table I). Nests from other populations of I. costipennis in Brazil are parasitized by flies of Sarcophagidae (Buschini & Woiski, 2006) and by ichneumonid wasp Messatoporus sp. (Soares et al., 2001) and in Argentina there are reports of Messatoporus transversostriatus (Spinola) (Martinez & Torretta, 2015). Our record represents the first association of a fly of the family Bombyliidae with this species of sphecid wasps.
On the other hand, in the studied trap-nests, females of T. leucon attacked nests of Pachodynerus guadulpensis (Saussure) and Zethus dicomboda (Spinola), two eumenid wasp species that use mud as cell partitions and captured caterpillars as prey. The first species built 27 nests [nine in canopy (71 brood cells) and 18 in understory (102 brood cells)]; of them, only two brood cells from two nests (one nest from understory and one from canopy) were parasitized by T leucon. One of these nests, was also attacked by Melittobia sp.(Table I). Other nests of P guadulpensis were attacked by Anthrax sp. (Bombyllidae, one brood cell), Amobia sp. (Sarcophagidae, several individuals from two brood cells form one nests) and one species of Ichneumonidae (two individuals from one brood cell). Nests from other populations of P guadulpensis were attacked in Argentina by two species of Messatoporus (Martinez & Torretta, 2015) and, in Brazil, they were parasitized by Sarcophagidae and Bombyliidae flies, and Chrysididae and Ichneumonidae wasps (Buschini & Buss, 2010), and T. amphitea (Rocha-Filho et al., 2020)
The other eumenid wasp Z dicomdoba only built four nests (19 brood cells), all in the understory (Torretta & Marrero, 2019). From one of these brood cells emerged one female of T. leucon. For this wasp species, information about organism predators is scarce, Torretta (2015) reported attacks by cuckoo wasp Chrysis boutheryi (Bréthes) (Chrysididae) in Argentina, and in our study site two nests were parasitized by other chrysidid wasp (Torretta, in prep.).
Our data for T. leucon agree with the limited data available for the species. The females attacked different species of wasps, which build mud nests as Vespidae: Eumeninae (Auko et al., 2014) and Crabronidae (Rocha-Filho et al., 2019, 2020). The emergence patterns of two parasitic Toxophora and its hosts were synchronized for all studied nests.
Parasitism of these species of Toxophora in our study site seems to be low. One possible explanation is that they have small populations, as can be seen by the small number of individuals in the collections and/or its scarce (or null) biological information. However, Torretta & Marrero (2019) reported that the mortality rate for assemblages of cavity-nesting bee and wasp species was higher than the parasitism rate in these forests, suggesting that mortality could be conditioned by the environmental conditions.
Toxophora aurea Macquart
Material examined. Argentina. Chaco: Resistencia, 1 male, 10-XII-1935, J.B. Daguerre (MACN). Formosa: San Francisco de Laishi, Reserva El Bagual, 1 female, 23-I-2013 (emergence date), J.P. Torretta (FAUBA). Misiones: San Ignacio, 1 female, 20-XII-2013, J.P. Torretta (FAUBA); 1 male, ex Hyptis sp., 14-I-2020, J.P. Torretta, A. Avalos, S. Reposi & L.J. Álvarez (MACN); 1 female, 15-I-2020, J.P. Torretta, A. Avalos, S. Reposi & L.J. Álvarez (MACN). Santa Fe: Piquete, 2 females, 5-I-1928, S.J. Bridarolli (MACN). Santiago del Estero: 1 female, without date, Wagner (MLP).
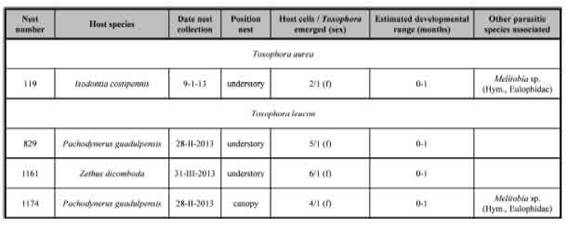
Table I: Species of Toxophora (Bombyliidae, Toxophorinae) rearing from nests of wasps obtained in trap-nests in forests in Reserva El Bagual, Formosa province, northern Argentina. f: female. Hym.: Hymenoptera.
Distribution: Argentina, Bolivia, Brazil and Paraguay.
In summary, in this paper we increase to 76 the number of species of Bombyliidae recorded from Argentina, present the first report about the reproductive ecology of T aurea and add information on the biology of this genus of bee flies.
ACKNOWLEDGMENTS
To the family Gotz and Alparamis S.A. for their permission to conduct this study in Reserva El Bagual, and to A. Di Giacomo for logistical support. To A. Roig-Alsina for his collaboration in the determination of C. anisitsii. To P. Mulieri (MACN) and L. Álvarez (MLP) that have loaned material for this study. To J.M. Castagnino for the photographs of Toxophora spp. CJEL thanks the financial support received from the “Conselho Nacional de Desenvolvimiento Científico e Tecnológico” - CNPq (proc. n. 302751/2019-0).

