










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.1 Buenos Aires ene./mar. 2005
Trilobites del Cámbrico tardío-Ordovícico temprano del área de Alfarcito, Tilcara, Cordillera Oriental de Jujuy, Argentina
Fernando J. Zeballo 1 y M. Franco Tortello 2
1 Museo de Paleontología, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Av. Vélez Sarsfield 299, 5000 Córdoba, Argentina. fzeballo@hotmail.com
2 CONICET - Departamento Científico Paleontología Invertebrados, Museo de La Plata, Paseo del Bosque s/nº, 1900 La Plata, Argentina. tortello@museo.fcnym.unlp.edu.ar
Resumen. Los trilobites del área de Alfarcito (Tilcara, Provincia de Jujuy, Argentina) están representados principalmente por olénidos, kainélidos, agnóstidos, ceratopígidos, hapalopléuridos y caliménidos. Se describen por primera vez los agnóstidos de esta región y, sobre la base del buen estado de preservación del material, se discuten algunos aspectos de la morfología de los trilobites polímeros. Los registros de Parabolina frequens argentina (Kayser), Parabolinella y Onychopyge indican una edad cámbrica tardía (parte inferior de la biozona epónima) para la Formación Casa Colorada. Aunque la Formación Alfarcito resultó estéril en trilobites, las faunas de conodontes y graptolitos permitieron datar su tramo superior al Tremadociano tardío bajo. Una edad similar es asignada a los estratos suprayacentes de la Formación Rupasca, en los que se destacan los registros de Bienvillia tetragonalis (Harrington), Leptoplastides marianus (Hoek), Parabolinella argentinensis Kobayashi, Peltocare norvegicum (Moberg y Möller), Pseudokainella keideli Harrington, Pharostomina trapezoidalis (Harrington), Hapalopleura sp., Apatokephalus sp., Gymnagnostus n. sp. A y Geragnostus sp. La fauna estudiada pertenece a la Provincia Paleobiogeográfica Báltica, ya que la mayoría de los taxones representados muestran una fuerte afinidad con géneros europeos ( e.g. , Parabolina , Peltocare ), junto con algunas formas endémicas sudamericanas ( e.g. , Hapalopleura ).
Abstract. Late Cambrian - early Ordovician Trilobites from the Alfarcito area, Tilcara, Oriental Cordillera, Jujuy, Argentina. The trilobites of the Alfarcito area (Tilcara, Jujuy Province, Argentina) are mainly represented by olenids, kainellids, agnostoids, ceratopygids, hapalopleurids and calymenids. The agnostoids from this region are described for the first time. In addition, some morphological aspects of the polymerid trilobites are discussed on the basis of the good state of preservation of the material. The records of Parabolina frequens argentina (Kayser), Parabolinella and Onychopyge indicate a Late Cambrian age (lower part of the eponymous biozone) for the Casa Colorada Formation. Although the Alfarcito Formation has no trilobites, its conodont and graptolite faunas have enabled us to date the upper part of the unit as early Late Tremadoc. A similar age is assigned to the overlying strata of the Rupasca Formation, in which Bienvillia tetragonalis (Harrington), Leptoplastides marianus (Hoek), Parabolinella argentinensis Kobayashi, Peltocare norvegicum (Moberg and Möller), Pseudokainella keideli Harrington, Pharostomina trapezoidalis (Harrington), Hapalopleura sp., Apatokephalus sp., Gymnagnostus n. sp. A and Geragnostus sp. are recorded. Although the faunas studied contain some endemic forms ( e.g. , Hapalopleura ), they show strong affinities with those from Europe ( e.g. , Parabolina , Peltocare ) and are therefore assigned to the Baltic Province.
Key words. Trilobita. Latest Cambrian-Early Ordovician. Alfarcito. Jujuy. Argentina. Systematics. Biostratigraphy.
Palabras clave. Trilobita. Cámbrico tardío-Ordovícico temprano. Alfarcito. Jujuy. Argentina. Sistemática. Bioestratigrafía.
Introducción
Alfarcito es una localidad clásica del Paleozoico inferior de Jujuy, donde diversos autores realizaron estudios geológicos y paleontológicos de distinta índole durante las primeras décadas del siglo XX. Esta área comprende la localidad tipo de tres unidades litoestratigráficas (formaciones Casa Colorada, Alfarcito y Rupasca) con un contenido fosilífero muy rico y es considerada clave para profundizar el conocimiento de la bioestratigrafía de la transición Cámbrico- Ordovícico y Tremadociano en la Cordillera Oriental argentina.
En el presente trabajo se describe la estratigrafía de dos quebradas de la región de Alfarcito: Casa Colorada y San Gregorio, y se discuten las implicaciones bioestratigráficas de sus faunas de trilobites. La buena preservación del material permite ampliar el conocimiento sistemático de algunas especies, mientras que nuevos registros de graptolitos y conodon- tes permiten datar con mayor precisión los rangos de los taxones identificados (Zeballo et al ., 2005).
Antecedentes y ubicación geográfica
Daniel (1940), López y Nullo (1969), Amengual y Zanettini (1974), Moya (1988) y Aceñolaza (1996) realizaron estudios geológicos en la inmediaciones de Tilcara. En dicha región, Harrington (1938), y Harrington y Leanza (1957) describieron una profusa fauna de trilobites y definieron, además, las formaciones Casa Colorada, Alfarcito y Rupasca, las cuales fueron originalmente asignadas al Ordovícico temprano (Tremadociano).
En la presente contribución se levantaron dos perfiles correspondientes a las quebradas de Casa Colorada y San Gregorio, ambas afluentes del río Huasamayo. Éste desagua sobre la margen izquierda de la quebrada de Humahuaca a la altura de la ciudad de Tilcara, distante 85 km de la ciudad de San Salvador de Jujuy, por la Ruta Nacional Nº 9 (figura 1).
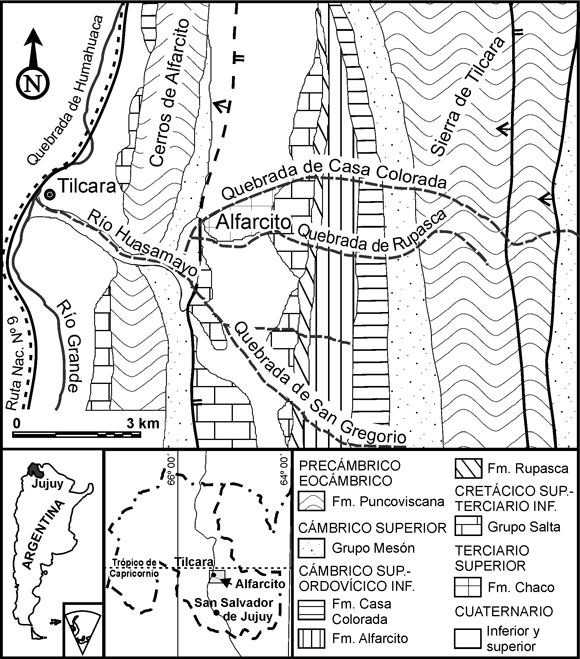
Figura 1. Mapa geológico del área de Alfarcito (modificado de López y Nullo, 1969, y Mon et al. , 1993). / Geologic map of the Alfarcito area (modified after López and Nullo, 1969, and Mon et al. , 1993).
Litoestratigrafía
La secuencia se inicia con los depósitos precámbricos- eocámbricos de la Formación Puncoviscana, compuestos por metamorfitas de bajo grado. Sobre éstos suprayacen las areniscas y ortocuarcitas de colores claros del Grupo Mesón, de edad cámbrica superior. El contacto del Grupo Mesón con la secuencia cámbrico cuspidal-ordovícica sólo se pudo observar a distancia. Esta última secuencia comprende a las formaciones Casa Colorada, Alfarcito y Rupasca. López y Nullo (1969) describen un ortoconglomerado oligomíctico de clastos angulosos, con matriz arenosa y cemento de color castaño-verdoso a violeta, de ca . 1 m de espesor en la base de la Formación Casa Colorada. El tramo superior de esta formación presenta en forma dominante lutitas de colores grises oscuros y verdes, con escasos bancos, delgados, de arenisca, aunque en la quebrada de San Gregorio expone niveles de areniscas de color gris oscuro, micáceas, con trilobites. El espesor observado osciló entre 15,50 m en la quebrada de Casa Colorada, y 48,50 m en la quebrada de San Gregorio. El contacto con la formación suprayacente es concordante, transicional. La Formación Alfarcito es una unidad heterolítica, donde alternan estratos lutíticos con potentes bancos de arenisca, de hasta 3 m de espesor. En la secuencia psamítica es frecuente encontrar icnofósiles, como Skolithos isp. y Cruziana isp. El tramo medio-superior de esta formación se caracteriza por la aparición de bancos de calcarenitas de hasta 1 m de espesor, coquinoideos, que brindaron una rica microfauna fósil. En la quebrada de San Gregorio, cerca del contacto con la unidad suprayacente, se encontraron graptolitos, asignados al género Rhabdinopora Eichwald junto con ondulitas de diversa orientación. El espesor total de la Formación Alfarcito es de 209 m en la quebrada de San Gregorio, donde aflora con pocas complicaciones tectónicas. En la quebrada de Casa Colorada se presenta con plegamiento y fallamiento en algunos sectores, lo que impide una medición correcta del espesor real. El espesor medido es de 478,50 m. El contacto con la unidad siguiente es concordante, neto. La Formación Rupasca es predominantemente lutítica, de colores gris-oscuro y verdoso, con escasas intercalaciones de bancos de arenisca y una abundante fauna de trilobites. De las lentes dispersas de coquina se recuperó una variada fauna de conodontes. El espesor medido en la quebrada de Casa Colorada es de 224,50 m y en la quebrada de San Gregorio es de 146,50 m. El contacto con las calizas oolíticas de la Formación Yacoraite, de edad cretácica superior, es discordante, de baja angularidad. Se completa la estratigrafía con las areniscas terciarias de las formaciones Chaco y Uquía, y sedimentos fluviales y aluviales cuaternarios (figura 1).
Bioestratigrafía
Los trilobites hallados permiten asignar la secuencia al Cámbrico tardío-Tremadociano (figuras 2 y 3).

Figura 2. Rangos estratigráficos de trilobites en la quebrada de Casa Colorada. / Stratigraphic ranges of trilobites in Casa Colorada creek . (P.f.a.: Zona de Parabolina frequens argentina / Parabolina frequens argentina Zone ).

Figura 3. Rangos estratigráficos de trilobites en la quebrada de San Gregorio / Stratigraphic ranges of trilobites in San Gregorio creek . (P.f.a.: Zona de Parabolina frequens argentina / Parabolina frequens argentina Zone , R.f.f.*: Intervalo con Rhabdinopora flabelliformis flabelliformis / Interval with Rhabdinopora flabelliformis flabelliformis).
La parte inferior de la sección pertenece a la Zona de Parabolina ( Neoparabolina ) frequens argentina (Harrington y Leanza, 1957; López y Nullo, 1969; Moya, 1988). Harrington y Leanza (1957) definieron esta biozona para el Tremadociano temprano del noroeste argentino y, posteriormente, Aceñolaza (1983) extendió su edad al Cámbrico tardío alto. Parabolina frequens argentina (Kayser) es un taxón de amplia representación en Argentina, Bolivia, Venezuela y Méjico. El mismo fue descripto para el Tremadociano inferior de algunas localidades ( e.g. , Angosto de Lampazar, Salta; Tortello y Rao, 2000), aunque está mejor representado en el Cámbrico Superior alto, caracterizando la mitad inferior de la biozona que lleva su nombre ( e.g. , Benedetto, 1977a; Aceñolaza, 1983; Salfity et al. , 1984; Moya et al. , 1994). En las secciones del río Santa Victoria (Harrington y Leanza, 1957), sierra de Cajas (Aceñolaza, 1968; Tortello y Esteban, 2003) y río Volcancito (Tortello y Esteban, 1999), P . frequens argentina se encuentra en los niveles más bajos de la transición Cámbrico-Ordovícico, acompañado de una diversa fauna de trilobites agnóstidos: Lotagnostus ( Lotagnostus ) sp., Lotagnostus ( Semagnostus ) zuninoi (Harrington y Leanza), Pseudorhaptagnostus ( Machairagnostus ) tmetus Harrington y Leanza, P . ( Machairagnostus ) corrugatus (Suárez-Soruco), Strictagnostus ? micropeltis (Harrington y Leanza), Micragnostus vilonii Harrington y Leanza, Gymnagnostus bolivianus (Hoek) y Leiagnostus turgidulus Harrington y Leanza, entre otros.
En nuestras colecciones, P . frequens argentina se registra en la parte superior de la Formación Casa Colorada, mientras que el tramo inferior de dicha formación es inaccesible en los dos perfiles estudiados. Por su parte, en este trabajo se identifican dos nuevos trilobites en la Formación Casa Colorada: Parabolinella argentinensis Kobayashi y Onychopyge sp. El primero posee una amplia distribución geográfica y un rango estratigráfico muy extenso. En efecto, P . argentinensis fue descripto en el Cámbrico Superior alto de México (Robison y Pantoja-Alor, 1968), el Tremadociano de Gales (Owens et al. , 1982) y el Cámbrico Superior-Tremadociano superior (biozonas de Parabolina frequens argentina , Kainella meridionalis y Bienvillia tetragonalis - Conophrys minutula ) de Bolivia y el noroeste argentino (Harrington y Leanza, 1957; Pribyl y Vanek, 1980). A su vez, Onychopyge es un elemento de mayor significación bioestratigráfica. Onychopyge riojana Harrington y Leanza es una especie característica del Cámbrico cuspidal del sistema de Famatina procedente de los niveles inferiores de la Formación Volcancito (Harrington y Leanza, 1957; Tortello y Esteban, 1999). Brandán (1974) describe ejemplares de la misma especie, posteriormente reasignados por Benedetto (1977b) a O . harringtoni , en los niveles inferiores de la Formación Santa Rosita en la quebrada de la Vizcacha, sierra de Cajas, junto con P . frequens argentina . Por su parte, Onychopyge argentina y O . longispina fueron descriptos en asociaciones similares de la sierra de Santa Victoria (Harrington y Leanza, 1957).
Si bien el registro fosilífero de la Formación Casa Colorada es escaso en número de especies, éstas sugieren una edad cámbrica tardía alta, no registrándose formas típicas de la base del Ordovícico (ej. Jujuyaspis ).
Por encima de estos niveles, existe un tramo asignable a casi la totalidad de la Formación Alfarcito, en el que no se registran trilobites. Harrington y Leanza (1957) describieron la Zona de Kainella meridionalis en esta formación, sobre la base del registro de Kainella meridionalis Kobayashi, Pseudokainella lata Kobayashi, Leptoplastides marianus (Hoek) y Angelina hyeronimi (Kayser). Este hecho no pudo ser corroborado en el presente estudio, si bien se registra una forma similar al taxón nominal ( Kainella ? sp.) en el tope de la Formación Alfarcito, en la quebrada Casa Colorada.
Finalmente, la fauna de la Formación Rupasca está compuesta por Bienvillia tetragonalis (Harrington), especie originalmente descripta sobre la base de material de la vecina localidad de Rupasca, acompañada de Leptoplastides marianus (Hoek), Parabolinella argentinensis Kobayashi, Pseudokainella keideli Harrington, Pharostomina trapezoidalis (Harrington), Peltocare norvegicum (Moberg y Möller), Hapalopleura sp., Asaphellus sp., Apatokephalus sp., Gymnagnostus n sp. A y Geragnostus sp. Si bien algunas de estas formas poseen rangos estratigráficos extensos, e.g. , P . argentinensis (véase más arriba), L. marianus (ej. Harrington y Leanza, 1957; Robison y Pantoja-Alor, 1968; Tortello y Rao, 2000), B . tetragonalis (Harrington y Leanza, 1957; Lu y Lin, 1984), la asociación caracteriza a la Zona de Bienvillia tetragonalis - Conophrys minutula (Tremadociano superior bajo) tal como fuera definida por Harrington y Leanza (1957) (véase también Henningsmoen, 1957; Branisa, 1965; Robison y Pantoja-Alor, 1968; Pribyl y Vanek, 1980; Moya et al. , 1994). No obstante, cabe destacar la significativa ausencia de Conophrys minutula Harrington en nuestras colecciones, razón por la cual la diagnosis de esta unidad bioestratigráfica puede ser revisada en el futuro (véase Waisfeld y Vaccari, 2003). En el presente trabajo se describe Hapalopleura en el área de Alfarcito, mientras que se confirma la presencia del género Peltocare en América del Sur. Gymnagnostus n. sp. A, de acuerdo a la sinonimia aquí propuesta, también fue descripto en el Ordovícico temprano (Tremadociano superior bajo) de Gales del Sur (Owens et al. , 1982), en niveles asignables a la Zona de Clonograptus tenellus . Asimismo, el registro de conodontes de la Zona de Paltodus deltifer (Subzona de Paltodus deltifer pristinus ), asociado a esta fauna de trilobites, indica una edad tremadociana tardía baja (Zeballo et al ., en prensa).
Consideraciones paleobiogeográficas
La fauna de trilobites en el área de Alfarcito muestra claras afinidades atlánticas, ya que la mayoría de los géneros presentes son comparables con formas europeas. También se hallan algunas formas endémicas (como el género Hapalopleura ) de origen sudamericano.
Shergold (1988) establece cuatro provincias biogeográficas para los trilobites durante la transición Cámbrico-Ordovícico: Báltica, Sudeste de China, Norte de China y Norteamérica, donde existiría un potencial control de temperatura y de corrientes en la distribución faunística de éstas. La fauna aquí considerada pertenece a la Provincia Báltica, que incluye a las regiones del norte de Gales, Escandinavia, Polonia, este de América del Norte, centro de México y América del Sur. Esta provincia habría sido de aguas frías debido a su posición en altas latitudes.
Paleontología sistemática
El material paleontológico estudiado está depositado en el Museo de Paleontología de la Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, bajo la sigla de repositorio CORD-PZ. Las siglas de muestreo CC y SG corresponden a muestras procedentes de las quebradas de Casa Colorada y San Gregorio, respectivamente.
Orden Agnostida Salter, 1864
Suborden Agnostina Salter, 1864
Familia Metagnostidae Jaekel, 1909
Género Geragnostus Howell, 1935
Especie tipo. Agnostus sidenbladhi Linnarsson, 1869, según designación original.
Observaciones. Durante las décadas que siguieron a la descripción original de Geragnostus Howell (1935), varios autores destacaron su fuerte similitud con otros géneros que, en conjunto, fueron considerados informalmente como " Micragnostus sensu lato " (ej., Micragnostus Howell, 1935; Oncagnostus Whitehouse, 1936; Homagnostus Howell, 1935; Trilobagnostus Harrington, 1938). En los últimos años se han realizado importantes avances para clarificar el alcance de estos taxones. De acuerdo a Fortey (1980), Geragnostus posee un surco transglabelar somero, con sus extremos extendidos hacia adelante, y un nodo axial dispuesto inmediatamente por detrás de ese surco; mientras que Micragnostus exhibe un F3 glabelar profundo, recto y transverso, y un nodo axial dispuesto en el punto medio del lóbulo posterior de la glabela o suavemente por detrás de ese punto. Siguiendo este criterio, en la actualidad Geragnostus y Micragnostus son incluidos en familias diferentes (Metagnostidae y Agnostidae, respectivamente).
Geragnostus sp.
Figuras 4.A-B
Material. Dos ejemplares completos, 24 céfalos, 20 pigidios (CORD-PZ 30549, 30551, 30619, 30648, 30657-30659, 30661, 30662, 30665, 30668, 30671, 30678, 30684, 30685, 30689, 30703, 30704, 30706, 30707, 30709, 30711, 30714, 30715, 30722, 30728, 30732, 30749, 30750, 30906, 30912, 30915, 30947-30950).

Figura 4. A-B, Geragnostus sp.; A, céfalo / cephalon , CORD-PZ 30750A, X9; B, ejemplar completo / complete specimen , CORD-PZ 30948, X8. C, Gymnagnostus n. sp. A., ejemplar completo / complete specimen , CORD-PZ 30914, X7. D-E, Parabolina ( N .) frequens argentina (Kayser); D, ejemplar completo / complete specimen , CORD-PZ 30510, X2; E, ejemplar completo / complete specimen , CORD-PZ 30502, X2. F, Gymnagnostus n. sp. A., pigidio / pygidium , CORD-PZ 30935, X13. G-J, Parabolinella argentinensis Kobayashi; G, cranidio / cranidium , CORD-PZ 30556, X2.5; H, cranidio / cranidium , CORD-PZ 30793, X1.5; I, céfalo-tórax / cephalothorax , CORD-PZ 30792, X1.5; J, cranidio / cranidium , CORD-PZ 30756, X2.5. K, Parabolina ( N .) frequens argentina (Kayser), tóraco-pigidio con hipostoma / thoracopygidium with hypostome , CORD-PZ 30516, X3.5. L, Parabolinella argentinensis Kobayashi, cranidios con campos preglabelares prominentes / cranidia with prominent preglabellar fields , CORD-PZ 30784, X2.5. M, Leptoplastides marianus (Hoek), céfalo-tórax / cephalon-thorax , CORD-PZ 30860, X4. N-O, Parabolinella argentinensis Kobayashi; N, ejemplar completo / complete specimen , CORD-PZ 30757, X1.2; O, ejemplar completo / complete specimen , CORD-PZ 30763, X1. P-R, Leptoplastides marianus (Hoek); P, céfalo-tórax / cephalon-thorax , CORD-PZ 30858A, X1.6; Q, céfalo-tórax / cephalon-thorax , CORD-PZ 30880B, X2; R, escudo axial / axial shield , CORD-PZ 30926, X6. S-T, Bienvillia tetragonalis (Harrington); S, cranidio / cranidium , CORD-PZ 30566, X4; T, cranidio / cranidium , CORD-PZ 30592, X4.
Descripción. Céfalo de contorno subcircular, convexo, de longitud y anchura equivalentes. El punto más alto localizado sobre el sector medio de la posteroglabela. Glabela bien elevada por sobre el nivel de las genas, claramente definida por surcos axiales delgados y profundos, de lados subparalelos a suavemente convergentes hacia adelante. La longitud glabelar es algo variable, oscilando entre el 62 % y el 66 % de la longitud (sag.) cefálica, mientras que la máxima anchura glabelar representa el 36-41 % de la máxima anchura cefálica. F3 muy somero, apenas esbozado en algunos ejemplares, con su tramo central de disposición transversa y sus porciones laterales dispuestas hacia adelante, ubicado en un nivel que representa el 60-64 % de la longitud glabelar (medido desde el margen posterior). Anteroglabela suavemente convexa, de contorno semioval, con su extremo anterior redondeado, abarcando en 36-40 % de la longitud (sag.) glabelar. Posteroglabela de convexidad más marcada, provista de un nodo muy delicado (sólo visible en los ejemplares mejor preservados) localizado cerca del surco F3. En vista dorsal, el extremo posterior de la posteroglabela es redondeado. Lóbulos basales subtriangulares, pequeños, de longitud y anchura equivalentes, conectados medialmente por una estrecha banda occipital, separados de la glabela y de las genas por surcos delicados. Acrolóbulo sin constricciones. Genas suavemente convexas, desprovistas de ornamentación, confluentes. Borde angosto y suavemente convexo, enangostado posterolateralmente. Surco marginal reducido. Borde y surco marginal abarcan, en conjunto, un 5,7-6,1 % del largo (sag.) cefálico. Borde posterior convexo (exsag.), puntiagudo, definido por un surco delgado, sin espinas.
Pigidio de contorno subcircular, con una anchura máxima algo mayor que la longitud (sag.). Pigaxis bien definido por surcos axiales delgados y profundos, convexo, bien elevado sobre el nivel de los campos pleurales, con sus lados subparalelos a suavemente arqueados hacia afuera y su extremo posterior subelíptico. La longitud pigaxial, excluido el semianillo articular, representa un 63-66 % de la longitud (sag.) del pigidio, mientras que la anchura máxima del pigaxis abarca un 42-48 % de la máxima anchura pigidial. Pigaxis dividido internamente en 3 lóbulos (M1-M3). M1 delimitado lateralmente por surcos axiales convergentes hacia atrás, dividido en un sector medio y 2 lóbulos laterales subtrapezoidales. F1 transverso, con su porción central levemente diferenciada. M2 más elevado y más largo que M1, definido lateralmente por surcos axiales subparalelos y provisto de un tubérculo conspicuo en forma de gota, que se prolonga sobre el área anterior de M3. Sobre el tubérculo pigidial está situado el punto más alto del pigidio. F2 delgado y somero, recto, de disposición transversa. Posteroaxis de contorno suboval, no expandido lateralmente, con una longitud (sag.) que representa un 52-56 % de la longitud (sag.) pigaxial. Acrolóbulo no constricto. Campos pleurales desprovistos de ornamentación, poco convexos, confluentes, de anchura aproximadamente uniforme. Borde de desarrollo moderado, levemente convexo, separado de los campos pleurales por un surco marginal poco profundo. Borde y surco marginal abarcan, en conjunto, un 9-12 % de la longitud pigidial. Espinas posterolaterales muy pequeñas, sólo visibles en los ejemplares mejor preservados. Borde anterior angosto (exsag.) y convexo, definido por un surco muy delgado.
Discusión. El material estudiado no presenta un estado de preservación óptimo, razón por la cual el surco transglabelar (F3) no se aprecia claramente en todos los casos. No obstante, en los ejemplares mejor conservados es posible determinar que este surco es muy somero y exhibe sus porciones laterales claramente extendidas hacia adelante. Asimismo, en algunos ejemplares se esboza un nodo glabelar localizado cerca de F3. Sobre la base de las diagnosis de Fortey (1980), Shergold et al. (1990) y Nielsen (1997), el material es asignado a Geragnostus . La presencia de tagmas de contorno subcircular, un pigaxis muy convexo con un lóbulo posterior de contorno subelíptico, un borde pigidial de anchura moderada y espinas posterolaterales muy cortas, son caracteres propios de Geragnostus lepidus Tjernvik, del Arenigiano superior de Suecia (Tjernvik, 1956: lám. 1, figs, 9-10, textfig. 27C; Ahlberg, 1992: fig. 10a-f). No obstante, el material de Alfarcito presenta un surco marginal cefálico más reducido y un posteroaxis desprovisto de nodo terminal.
Harrington y Leanza (1957) han descripto tres especies de Geragnostus en el Tremadociano superior del noroeste argentino: G . nesossii , identificado en diferentes localidades asignables al Tremadociano superior (Harrington y Leanza, 1957: figs 9.1-5, 10; Tortello, 1996a: figs 4.1-12; Tortello y Rao, 2000), se diferencia de Geragnostus sp. porque el primero presenta un surco marginal cefálico de inusual desarrollo, posteroaxis de contorno variable y espinas posterolaterales más conspicuas. Geragnostus maurii , de la Biozona de Bienvillia tetragonalis - Conophrys minutula en el tramo austral de la sierra de Mojotoro (Harrington y Leanza, 1957: figs. 8, 9.6; González, 1983), exhibe un posteroaxis truncado posteriormente y espinas posterolaterales de buen desarrollo. Por su parte, Geragnostus callaveiformis procedente de varias localidades clásicas de la Biozona de Notopeltis orthometopa (Harrington y Leanza, 1957: figs. 13.1-5, 14; Tortello, 1996b: lám. 1, figs. 6-8), se distingue principalmente porque exhibe campos pleurales más convexos, un pigaxis más largo y un posteroaxis algo expandido lateralmente en su porción media.
El material estudiado se diferencia de la especie tipo G . sidenbladhi (Linnarsson), del Tremadociano tardío (Biozona de Apatokephalus serratus ) de Suecia y Noruega ( e.g. , Ahlberg, 1989: figs. 1A-K, 2A-C, 3AG), porque este último posee el céfalo y el pigidio de contorno subcuadrado, el pigaxis menos elevado por sobre el nivel de los campos pleurales, el margen posterior del posteroaxis truncado o apenas redondeado, y espinas posterolaterales conspicuas.
Geragnostus yangtzeensis Lu, del Tremadociano de China (Lu, 1975: lám. 1, figs. 1-4), se diferencia de Geragnostus sp. porque presenta bordes y surcos marginales mucho más anchos y un posteroaxis de contorno menos semielíptico. Geragnostus crassus Tjernvik, del Tremadociano superior-Arenigiano inferior de Suecia y el sur de China (Tjernvik, 1956: lám. 1, figs. 7-8, text-fig. 27B; Ahlberg, 1992: figs. 7.a-r), posee bordes más desarrollados que Geragnostus sp. y un pigaxis más ancho.
Por su parte, G . splendens (Holub), del Arenigiano temprano de la República Checa y Suecia (Pek, 1977: lám. 1, fig. 8; Alhberg, 1992: figs. 10 k-o, 11a-i) se distingue de Geragnostus sp. porque el primero posee un posteroaxis algo expandido lateralmente en su porción media, y bordes marginales y espinas posterolaterales de mayor tamaño; mientras que G . callavei (Raw), del Tremadociano de Inglaterra (véase Rushton, 1988: lám. 66, figs. 1, 2, 4, 5, 11), presenta el extremo anterior de la glabela levemente acuminado y un anteroaxis (M1+M2) pigidial proporcionalmente pequeño en comparación con las dimensiones de M3.
Procedencia y edad. Muestras CC11-4,50 m, CC11- 0,50 m, CC11+19 m, CC11+25 m, CC11+50 m, SG9+6 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Incertae Familiae
Género Gymnagnostus Robison y Pantoja-Alor, 1968
Especie tipo. Gymnagnostus gongros Robison y Pantoja-Alor, 1968, según designación original.
Gymnagnostus n. sp. A
Figuras 4.C, F
1982. Leiagnostus aff. turgidulus Harrington y Leanza. Owens et al. : 6-7; lám. 1, figs. b-d, f.
1991. Gymnagnostus aff. turgidulus (Harrington y Leanza). Fortey y Owens: 443.
Material. Ocho ejemplares completos, 3 céfalos y 9 pigidios (CORD-PZ 30532, 30534, 30536, 30545, 30653, 30678, 30682, 30696, 30718, 30722, 30785, 30914, 30933-30935, 30939, 30941, 30944, 30951).
Descripción. Céfalo liso, de contorno subelíptico, de longitud y anchura subiguales, moderadamente convexo, con el punto más elevado localizado sobre el tercio posterior del acrolóbulo. Tramos proximales de los surcos axiales apenas esbozados, delimitando el borde externo de los lóbulos basales. Acrolóbulo no constricto. Borde muy angosto, convexo, enangostado hacia atrás, delimitado por un surco marginal muy delgado. Sagitalmente, el borde y el surco marginal ocupan, en conjunto, un 6% de la longitud cefálica.
Pigidio liso, de contorno subelíptico, aproximadamente tan largo como ancho, con una convexidad algo mayor que la del céfalo. El punto más alto del pigidio está localizado en el cuarto anterior del acrolóbulo. En dicho punto se observa un nodo axial redondeado, apenas esbozado, de dimensiones moderadas. Tramos anteriores de los surcos axiales apenas esbozados, delimitando un pigaxis (porción proximal) de lados subparalelos cuya anchura representa un 40% de la máxima anchura pigidial. Acrolóbulo con su parte posterior levemente aguzada. Semianillo articular angosto (sag.), separado del resto del pigidio por un surco delgado de disposición transversa. Borde pigidial bien definido, suavemente convexo, enangostado hacia adelante en su tercio anterior, sin espinas, separado del acrolóbulo por un surco marginal conspicuo. Surco marginal y borde representan, en conjunto, un 8-12 % de la longitud pigidial. Bordes anteriores muy delgados, separados del acrolóbulo por un surco muy angosto.
Discusión . De acuerdo a las diagnosis provistas por Robison y Pantoja-Alor (1968), Shergold et al. (1990) y Nielsen (1997), el material estudiado es referido al género Gymnagnostus . Tal asignación está fundamentada, principalmente, en el hecho de que el material presenta los surcos dorsales con un alto grado de obliteración (" effacement "), los lóbulos basales parcialmente esbozados, un borde cefálico delgado aunque bien definido, y un acrolóbulo pigidial que carece de un nodo terminal. Debido al escaso número de ejemplares examinados, el material es referido en nomenclatura abierta.
Leiagnostus aff. turgidulus fue descripto originalmente en el Tremadociano de Gales (Owens et al. , 1982: 6-7, lám. 1, figs. b-d, f) y, posteriormente, reasignado a Gymnagnostus por Fortey y Owens (1991). De acuerdo a la descripción de Owens et al. (1982), este taxón podría ser coespecífico de Gymnagnostus n. sp. A.
Gymnagnostus n. sp. A se diferencia de G . bolivianus (Hoek), del Cámbrico Superior alto de Bolivia y Argentina ( e.g. , Harrington y Leanza, 1957: fig. 21, 2a-b; Tortello y Aceñolaza, 1993: lám. 1, figs. 5-6; Tortello y Esteban, 1999: figs. 5N-O) porque este último posee los bordes cefálicos y pigidial de anchura similar.
Gymnagnostus n. sp. A se distingue de G . ellipticus (Lu y Lin), del Cámbrico Superior alto-Tremadociano inferior de China (Lu y Lin, 1984: lám. 4, figs. 6-8; Peng, 1991: lám. 1, figs. 2-5), porque este último posee un céfalo de contorno subcircular. Gymnagnostus perinflatus (Harrington y Leanza), del Cámbrico tardío del sistema de Famatina (Harrington y Leanza, 1957: fig. 21:3; Tortello y Esteban, 1999: figs. 5P-T), presenta un céfalo con sus surcos dorsales menos obliterados y un pigidio más convexo. Por su parte, la especie tipo G . gongros Robison y Pantoja-Alor, del Cámbrico cuspidal de Méjico (Robison y Pantoja- Alor, 1968: lám. 97, figs. 24-33), posee espinas posterolaterales bien definidas, un tubérculo pigaxial conspicuo y un borde pigidial anterior fuertemente geniculado.
Procedencia y edad. Muestras CC6-15 m, CC11-4,50 m, CC11-0,50 m, CC11+50 m, CC11+56 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Distribución geográfica. Gales (Owens et al. , 1982) y Argentina.
Orden Ptychopariida Swinnerton, 1915
Suborden Olenina Burmeister, 1843
Familia Olenidae Burmeister, 1843
Subfamilia Oleninae Burmeister, 1843
Género Parabolina Salter, 1849
Especie tipo. Parabolina frequens (Barrande, 1868), según designación original.
Subgénero Neoparabolina Nikolaisen y Henningsmoen, 1985
Parabolina ( Neoparabolina ) frequens argentina (Kayser, 1876)
Figuras 4.D-E, K; 6.A
1876. Olenus argentinus Kayser: 6; lám. 1, figs. 1-3.
1957. Parabolina argentina (Kayser). Harrington y Leanza: 81-85; figs. 25-26.
1957. Parabolina argentina (Kayser). Henningsmoen: 116-117 (cum syn.).
2000. Parabolina ( Neoparabolina ) frequens argentina (Kayser). Tortello y Rao: 69-71; figs. 3. D-I (cum syn.).
2003. Parabolina ( Neoparabolina ) frequens argentina (Kayser). Tortello y Esteban: 338-340; figs. 5. G-I.
Material. 24 ejemplares completos, 1 cranidio, 13 fragmentos cranidiales, 2 librígenas, 1 céfalo-tórax, 3 tóraco-pigidios, 6 pigidios (CORD-PZ 30500-30504, 30506, 30507, 30509-30512, 30515, 30516, 30789, 30908, 30917-30919, 30921-30925, 30927, 30928).
Discusión. El material coleccionado presenta un excelente estado de preservación, lo que permite observar el hipostoma de este taxón (figuras 4.K, 6.A). El largo del pigaxis, la forma de la glabela, el diseño de los surcos glabelares y de la sutura facial, y el ancho del borde anterior parecen ser altamente variables (Harrington y Leanza, 1957; Pribyl y Vanek, 1980). Siguiendo el criterio de estos autores, la presencia de 3 pares de espinas pigidiales y el largo del pigaxis de los ejemplares estudiados permiten asignar el material a Parabolina frequens argentina .
Parabolina ( Neoparabolina ) frequens argentina (Kayser) se diferencia de Parabolina ( Neoparabolina ) frequens frequens (Barrande), del Ordovícico temprano de Alemania (Sdzuy, 1955: lám. 3, figs. 58-70; text-fig. 10), por poseer 2-3 pares de espinas pigidiales en lugar de 2-4, y un borde pigidial más estrecho. A su vez, difiere de P. ( N .) frequens finmarchica Nikolaisen y Henningsmoen (1985: figs. 4, 11.C-J, 12.A-Ea), del Tremadociano de Noruega, porque tiene las espinas genales dispuestas más posteriormente.
Procedencia y edad. Muestras CC1-445 m, CC1-430 m, SG1-120 m, Formación Casa Colorada, Zona de Parabolina frequens argentina . Cámbrico tardío alto.
Distribución geográfica. Argentina (Harrington y Leanza, 1957; Brandán, 1974; Tortello y Rao, 2000; Tortello y Esteban, 2003), Bolivia (Kayser, 1876; Harrington y Leanza, 1943; Pribyl y Vanek, 1980) y Venezuela (Frederickson, 1958).
Género Parabolinella Brøgger, 1882
Especie tipo. Parabolinella limitis Brøgger, 1882 (designado por Bassler, 1915).
Parabolinella argentinensis Kobayashi, 1936
Figuras 4.G-J, L, N-O; 6.B
1936. Parabolinella argentinensis Kobayashi: 88; lám. 15, figs. 1-5.
1957. Parabolinella argentinensis Kobayashi. Harrington y Leanza: 103-105; figs. 37 (1-11), 38 (1-8).
1980. Parabolinella argentinensis Kobayashi. Pribyl y Vanek: 18-19; lám. 7, figs. 1-2; lám. 8, fig. 8 (cum syn.).
1982. Parabolinella argentinensis Kobayashi. Owens et al. : 8-9; lám. 1, figs. g-j.
Material. Cuatro ejemplares completos, 1 céfalo, 4 fragmentos cefálicos, 4 cranidios, 26 fragmentos cranidiales, 6 librígenas, 1 tóraco- pigidio, 1 fragmento torácico, 2 pigidios (CORD-PZ 30556, 30558, 30578, 30595, 30607, 30640, 30646, 30677, 30681, 30715, 30739, 30747, 30753, 30756, 30757, 30763, 30768, 30782, 30784- 30786, 30788, 30792, 30793, 30800, 30860, 30875, 30877, 30879, 30883, 30887, 30889, 30890, 30894, 30909, 30912, 30915, 30944).
Discusión. Harrington (1938) describió especímenes de las quebradas de Huasamayo y Rupasca, pero los mismos no poseen el campo preglabelar inflado, como es el caso del material coleccionado en la Formación Casa Colorada, quebrada de San Gregorio (figuras 4.H-J, L). Fortey y Hughes (1998) interpretan que los abultamientos del campo preglabelar de algunos trilobites cambro-ordovícicos podrían haber funcionado como "sacos embrionarios" ( brood pouches ), lo que indicaría la existencia de dimorfismo sexual. En nuestro caso, es interesante señalar que todos los ejemplares de P . argentinensis procedentes del tramo inferior del perfil presentan esta característica, mientras que ésta no fue documentada en el resto de la sección. Waisfeld y Vaccari (2003) señalan que los ejemplares de P. argentinensis ilustrados por Harrington y Leanza (1957) corresponderían a más de una especie, hecho que podría apoyarse con el material estudiado aquí.
Parabolinella argentinensis Kobayashi se distingue de Parabolinella coelatifrons Harrington y Leanza (1957: fig. 39.3a-h), del Cámbrico tardío del noroeste argentino, porque esta última tiene surcos axiales levemente convergentes y 15 segmentos torácicos en lugar de 19. Por su parte, Parabolinella triarthra (Callaway), del Ordovícico temprano de Inglaterra y Canadá ( e.g. , Henningsmoen, 1957: lám. 12, figs. 6-7; Owens et al. , 1982: lám. 1, fig. K) se diferencia por poseer una sutura facial menos divergente anteriormente y 21 segmentos torácicos (Robison y Pantoja- Alor, 1968).
Procedencia y edad. Muestras CC11-4,50 m, CC11-2 m, CC11-0,50 m, CC11+8 m, CC11+12,50 m, CC11+29 m, CC11+34 m, CC11+56 m, SG1-122 m, SG1-115 m, SG7 B-2 m, SG7 C-1,5 m, SG7 C, SG8-3 m, SG8+12 m, formaciones Casa Colorada y Rupasca, zonas de Parabolina frequens argentina y Bienvillia tetragonalis - Conophrys minutula . Cámbrico tardío alto-Tremadociano tardío bajo.
Distribución geográfica. Argentina (Harrington y Leanza, 1957; Malanca, 1973; Brandán, 1974), Bolivia (Pribyl y Vanek), Méjico (Robison y Pantoja-Alor, 1968) y Gales (Owens et al. , 1982).
Subfamilia Pelturinae Hawle y Corda, 1847
Género Leptoplastides Raw, 1908
Especie tipo. Conocoryphe salteri Callaway, 1877, según designación original.
Leptoplastides marianus (Hoek, en Steinman y Hoek, 1912)
Figuras 4.M, P-R; 6.C-D
1912. Parabolinopsis mariana Hoek. En: Steinmann y Hoek: 226; lám. 7, figs. 1-3. ?1957. Protopeltura mesembria Harrington y Leanza: 88; fig. 27, 2 ad.
1957. Parabolinopsis mariana Hoek. Harrington y Leanza: 89-93; figs. 29-30 ( cum. syn .).
1957. Leptoplastides marianus (Hoek). Henningsmoen: 266 (cum syn.).
2000. Leptoplastides marianus (Hoek). Tortello y Rao: 72; figs. 3. J-R ( cum. syn .).
Material. Holáspidos: 20 ejemplares completos, 6 céfalos, 6 céfalotórax, 11 tóraco-pigidios, 9 pigidios, 30 cranidios, 56 librígenas, 10 fragmentos céfalicos, 158 fragmentos cranidiales, 4 fragmentos céfalo- torácicos, 1 fragmento pigidial. Meráspidos: 24 cranidios, 6 librígenas, 4 fragmentos cranidiales (CORD-PZ 30532-30535, 30537- 30540, 30543-30545, 30553, 30557, 30560-30563, 30565, 30569, 30571, 30576, 30577, 30581, 30585, 30587-30589, 30591, 30593-30596, 30600, 30602-30605, 30608-30610, 30612, 30613, 30615, 30620, 30625, 30630, 30633, 30635, 30637-30639, 30643, 30647, 30649-30653, 30656-30658, 30660, 30661, 30663-30667, 30669, 30670, 30672, 30674-30680, 30682, 30683, 30685-30689, 30692, 30694, 30698-30702, 30704, 30705, 30707- 30714, 30717-30725, 30727-30729, 30731, 30733, 30734, 30736, 30738, 30739, 30741-30745, 30750-30752, 30754, 30755, 30758-30760, 30762, 30764, 30767-30771, 30773-30780, 30795-30798, 30856-30860, 30862, 30863, 30865, 30868, 30872, 30873, 30876-30878, 30880-30886, 30888, 30892, 30894, 30897, 30899-30901, 30903, 30904, 30906, 30907, 30911, 30915, 30919, 30926, 30936, 30939, 30943, 30945, 30947, 30948, 30950, 30951).
Discusión. Harrington (1938) describió este taxón en las quebradas de Rupasca y Huasamayo bajo el nombre de Protopeltura granulosa n. sp., siendo una especie muy abundante en toda esta región. Leptoplastides marianus presenta una alta variabilidad intraespecífica relacionada con el desarrollo ontogenético (Robison y Pantoja-Alor, 1968), que involucra el grado de expresión de los surcos glabelares y aristas oculares, el ángulo de inserción de las espinas genales, presencia de nodos o espinas torácicas, y el número de espinas pigidiales (Robison y Pantoja Alor, 1968; Rao y Tortello, 1998; Tortello y Rao, 2000). Tal variabilidad se manifiesta en el material de Alfarcito: los ejemplares meráspidos y holáspidos tempranos poseen una glabela convexa, 2 ó 3 pares de surcos glabelares, nodo occipital, aristas oculares bien marcadas, librígenas provistas de estrías que irradian desde el ojo, y un ángulo genal de 140º; mientras que los ejemplares holáspidos tardíos presentan la glabela poco convexa, surcos glabelares apenas esbozados o ausentes, un anillo occipital liso, aristas oculares poco marcadas, y un ángulo genal externo de casi 180º.
Protopeltura mesembria Harrington y Leanza es incluida tentativamente en sinonimia ya que algunas de las características descriptas por Harrington y Leanza (1957: 88, fig. 27.2a-d) para esta especie coinciden con las de los ejemplares meráspidos y holáspidos tempranos de L . marianus (ángulo genal, estrías en las librígenas, surcos glabelares y convexidad de la glabela).
Procedencia y edad. Muestras CC6-15 m, CC11-4,50 m, CC11-2 m, CC11-0,50 m, CC11, CC11+8 m, CC11+12,50 m, CC11+25 m, CC11+29 m, CC11+34 m, CC11+42 m, CC11+50 m, CC11+54 m, CC11+56 m, SG7 B-2 m, SG7 C-1,50 m, SG7 C, SG8-3 m, SG8+12 m, SG9+6 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Distribución geográfica. Argentina (Harrington, 1938; Harrington y Leanza, 1957; Malanca, 1973; Aceñolaza y González, 1977; Rao y Tortello, 1998; Tortello y Rao, 2000), Bolivia (Steinmann y Hoek, 1912) y Méjico (Robison y Pantoja-Alor, 1968).
Género Peltocare Henningsmoen, 1957
Especie tipo. Acerocare norvegicum Moberg y Möller, 1898 (según designación original).
Peltocare norvegicum (Moberg y Möller, 1898)
Figuras 5.A-D; 6.E
1898. Acerocare norvegicum Moberg y Möller: 243. 1957. Peltocare norvegicum (Moberg y Möller). Henningsmoen: 247; lám. 2, fig. 9; lám. 8; lám. 27, figs. 6, 8, 10-14 ( cum syn .).
1957. Acerocarina glaber (Harrington). Harrington y Leanza: 93; figs. 3 a-d (cum syn.).
1968. Peltocare norvegicum (Moberg y Möller). Robison y Pantoja- Alor: 793; lám. 103, figs. 14-23 ( cum syn.).
Material. Cinco ejemplares completos, 1 céfalo, 7 cranidios, 9 céfalo- tórax, 4 tóraco-pigidios, 11 pigidios, 15 librígenas, 19 fragmentos cranidiales (CORD-PZ 30580, 30586, 30591, 30595, 30597, 30599, 30602, 30603, 30606, 30608, 30616, 30617, 30624, 30628, 30642, 30644, 30657, 30671, 30673, 30674, 30682, 30695, 30722, 30728, 30730, 30737, 30739, 30746, 30748, 30856, 30857, 30859, 30861, 30865, 30866-30870, 30877, 30880-30883, 30886, 30887, 30891, 30913, 30916).
Discusión. Acerocarina glaber (Harrington) fue descripto originalmente con material de las quebradas de Rupasca y Huasamayo, en el área de Alfarcito (Harrington, 1938: 212-213, lám. 9, figs. 1, 5, 12; Harrington y Leanza, 1957: 93-95, figs. 32.3 a-d). Henningsmoen (1957) y Robison y Pantoja-Alor (1968) consideran a A . glaber como sinónimo junior de P . norvegicum , criterio que se sigue en este trabajo.
El material coleccionado posee una variabilidad muy grande. La convexidad del céfalo y del tórax es el rasgo más notorio, denotando dos tipos de preservación: ejemplares comprimidos dorso-ventralmente (figuras 5.A-B) y sin compresión (figura 5.D). Estos últimos concuerdan con el material de Oaxaca, Méjico, descripto por Robison y Pantoja-Alor (1968: lám. 100, figs. 8, 9, 11-14). Otras características notables en los ejemplares bien conservados son la presencia de omatidios en los ojos, y las líneas aterrazadas en el pigidio (figura 6.E).

Figura 5. A-D, Peltocare norvegicum (Moberg y Möller); A, ejemplar completo / complete specimen , CORD-PZ 30624, X2.5; B, toraco-pigidio / thorax - pygidium , CORD-PZ 30916, X2; C, toraco-pigidio / thorax - pygidium , CORD-PZ 30856, X1.9; D, cranidio / cranidium , CORDPZ 30869, X5.5. E, Pharostomina trapezoidalis (Harrington), cranidio / cranidium , CORD-PZ 30727, X8. F, Kainella ? sp., cranidio / cranidium , CORD-PZ 30532, X9.5. G-H, Pseudokainella keideli Harrington; G, pigidio / pygidium , CORD-PZ 30879, X1.5; H, ejemplar completo / complete specimen , CORD-PZ 30888, X5. I-K, Hapalopleura sp.; I, cranidio / cranidium , CORD-PZ 30938, X4; J, ejemplar enrollado / rolled specimen , CORD-PZ 30932, X3.5; K, ejemplar completo / complete specimen , CORD-PZ 30871, X4. L, Onychopyge sp., tórax fragmentario y pigidio / fragmentary thorax and pygidium , CORD-PZ 30794, X0.5

Figura 6. A, Parabolina ( N .) frequens argentina Kayser, hipostome / hypostoma , CORD-PZ 30514; B, Parabolinella argentinensis (Kobayashi), tórax posterior y pigidio / posterior thorax and pygidium , CORD-PZ 30646; C-D, Leptoplastides marianus (Hoek); C, librígena / librigena , CORD-PZ 30770; D, cranidio meráspido / meraspid cranidium , CORD-PZ 30768; E, Peltocare norvegicum (Moberg y Möller), pigidio / pygidium , CORD-PZ 30722; F, Onychopyge sp., cranidio / cranidium , CORD-PZ 30790; G, Apatokephalus sp., cranidio / cranidium , CORD-PZ 30750. Escala gráfica / scale bar: 1 mm.
Peltocare norvegicum (Moberg y Möller) se distingue de P . compactum Nikolaisen y Henningsmoen (1985: figs. 8, 16.A-R), del Tremadociano de Noruega, fundamentalmente porque esta última presenta regiones pleurales más estrechas (tr.).
Procedencia y edad. Muestras CC11-4,50 m, CC11-2 m, CC11-0,50 m, CC11+8 m, CC11+12,50 m, CC11+19 m, SG7 C, SG8-3 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Distribución geográfica. Argentina (Harrington y Leanza, 1957), Méjico (Robison y Pantoja-Alor, 1968), Noruega (Henningsmoen, 1957) y Suecia (Moberg y Möller, 1898).
Subfamilia Triarthrinae Ulrich, 1930
Género Bienvillia Clark, 1924
Especie tipo. Dikelocephalus ? corax Billings, 1865, según designación original.
Observaciones. En este trabajo se sigue la diagnosis propuesta por Ludvigsen y Tuffnell (1983), que caracteriza al género por poseer el surco del borde cefálico anterior dispuesto sobre el cranidio, y un campo preglabelar de buen desarrollo.
Bienvillia tetragonalis (Harrington, 1938)
Figuras 4.S-T
1938. Parabolinella tetragonalis Harrington: 196; lám. 7, figs. 3-4.
1957. Triarthrus tetragonalis (Harrington). Harrington y Leanza: 113-115; fig. 42 (1-6, ?7, ?8) (cum syn.).
1980. Triarthrus tetragonalis (Harrington). Pribyl y Vanek: 17; lám. 9, figs. 1-2 (cum syn.).
1984. Bienvillia tetragonalis jiangshanensis subsp. nov. Lu y Lin: 82- 83, lám. 6, fig. 3b; lám. 8, figs. 6-9.
Material. Dos ejemplares completos, 10 cranidios, 1 céfalo-tórax, 1 pigidio, 31 fragmentos cranidiales, 2 fragmentos torácicos (CORDPZ 30532, 30545, 30548, 30557, 30559, 30564, 30566, 30570, 30572, 30582, 30583, 30590, 30592, 30596, 30601, 30608-30611, 30623, 30631, 30635, 30636, 30638, 30663, 30732, 30735, 30738, 30742, 30753, 30755, 30759, 30766, 30772, 30863, 30864, 30866, 30880, 30896, 30898, 30910, 30933, 30946).
Discusión. Esta especie se describió originalmente con material de las quebradas de Rupasca y Huasamayo (Harrington, 1938). Los ejemplares estudiados concuerdan en general con la descripción de Harrington, aunque presentan una cierta variabilidad en el diseño de las ramas anteriores de la sutura facial.
Lu y Lin (1984: lám. 6, fig. 3b; lám. 8, figs. 6-9) describen una nueva subespecie del Tremadociano de China, Bienvillia tetragonalis jiangshanensis , que presenta las ramas anteriores de la sutura facial divergentes y las fixígenas estrechas, pero estas características parecen estar incluidas en el rango de variación morfológica de la especie. Bienvillia parchaensis (Harrington y Leanza, 1957: figs. 43.1a-h, 44.1a-e), del Ordovícico inferior del noroeste argentino, se distingue de B . tetragonalis por tener lóbulos palpebrales más grandes, los cuales están ubicados más próximos al margen posterior, un campo preglabelar más estrecho, y sólo 2 anillos en el pigaxis en lugar de 3 en B . tetragonalis .
Procedencia y edad. Muestras CC6-15 m, CC11-4,50 m, CC11-2 m, CC11-0,50 m, CC11+8 m, CC11+12,50 m, CC11+29 m, CC11+42 m, CC11+56 m, SG7 C, SG8- 3 m, SG8+12 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Distribución geográfica. Argentina (Harrington y Leanza, 1957), Bolivia (Pribyl y Vanek, 1980; Branisa, 1965), Méjico (Robison y Pantoja-Alor, 1968), Noruega (Henningsmoen, 1957) y China (Lu y Lin, 1984). Orden ASAPHIDA Salter, 1864 enmend . Fortey y Chatterton, 1988
Suborden Asaphina Salter, 1864 enmend . Fortey y Chatterton, 1988
Superfamilia Asaphoidea Burmeister, 1843
Familia Ceratopygidae Linnarsson, 1869
Género Onychopyge Harrington, 1938
Especie tipo. Onychopyge riojana Harrington, 1938, según designación original.
Onychopyge sp.
Figuras 5.L; 6.F
Material. Tres pigidios y 1 fragmento cranidial (CORD-PZ 30782, 30783, 30790, 30794).
Descripción. Cranidio de contorno subtrapezoidal. Glabela convexa, limitada por surcos axiales levemente convergentes hacia adelante, con 2 pares de surcos laterales pobremente definidos, oblicuos hacia atrás y adentro. Anillo occipital ancho, con sus extremos ligeramente dirigidos hacia adelante. Campo preglabelar estrecho, plano, con su borde anterior casi del mismo ancho (sag.). Parte posterior de las fixígenas cortas (sag.) y anchas (tr.), con sus extremos dirigidos apenas hacia atrás. Lóbulos palpebrales semielípticos, convexos, ubicados más cerca del margen posterior que del anterior. Ramas anteriores de la sutura facial fuertemente divergentes, ramas posteriores normales al axis. Las librígenas no fueron observadas.
Tórax con número indeterminado de segmentos. Anillos de bordes rectos, con pleuras ligeramente dirigidas hacia atrás, con sus extremos distales en forma de espinas, solapadas, fuertemente dirigidas hacia atrás.
Pigidio grande, con 7 anillos, segmento terminal y cresta post-axial corta. Pleura anterior muy ancha (exsag.), de forma lanceolada o subtriangular con surco interpleural profundo, que se proyecta en una espina muy larga, más allá del borde posterior del pigidio. Este último presenta líneas aterrazadas. Un par de pleuras cortas por debajo de la pleura anterior. Borde posterior plano o deprimido, ancho. Margen posterior fuertemente curvado en su parte media.
Discusión. El gran tamaño de los ojos, su ubicación (cercanos al borde posterior y adyacentes a la glabela), la presencia de la primera pleura del pigidio con una espina larga, y el borde posterior del pigidio ancho y deprimido permiten asignar el material al género Onychopyge Harrington.
Onychopyge sp. difiere de O . riojana Harrington (1938: lám. 5, fig. 20; Harrington y Leanza, 1957: fig. 95, 96.2a-c), del Cámbrico tardío de Famatina, porque la primera posee lóbulos palpebrales semielípticos en lugar de semicirculares, y la longitud de la espina de la primera pleura pigidial es mayor. Harrington y Leanza (1957: 189, figs. 96.1a-d; 96.4) describen dos especies de la Cordillera Oriental: O . argentina , y O . longispina , que se diferencian del material aquí estudiado por presentar 5 anillos axiales pigidiales en lugar de 7. Benedetto (1977b: lám. 2, figs. 14-15) describe una nueva especie, O . harringtoni , de la sierra de Cajas, provincia de Jujuy, que se distingue porque exhibe 3 pares de surcos glabelares, un área frontal más amplia y 6 anillos axiales pigidiales. Por su parte, Onychopyge sculptura Robison y Pantoja-Alor (1968: lám. 100, figs. 1-7, 10), del Cámbrico tardío de Méjico, tiene un campo preglabelar más ancho (sag.) y 5-6 anillos axiales en el pigidio.
Dado que el material es escaso, se describe en nomenclatura abierta. Ésta es la primera cita del género en el área de Alfarcito.
Procedencia y edad. Muestras SG1-122 m, SG1-120 m, Formación Casa Colorada, Zona de Parabolina frequens argentina . Cámbrico tardío alto.
Superfamilia RemopleuridioideA Hawle y Corda, 1847
Familia Kainellidae Ulrich y Resser, 1930
Género Kainella Walcott, 1925
Especie tipo. Hungaia billingsi Walcott, 1924, según designación original.
Kainella ? sp.
Figura 5.F
Material. Dos fragmentos cranidiales (CORD-PZ 30532).
Descripción. Cranidio de contorno trapezoidal, más ancho (en su parte posterior) que largo. Glabela subrectangular, de bordes axiales rectos, débilmente convergentes hacia adelante, surco preglabelar levemente curvado hacia anterior. Dos pares de surcos laterales conspicuos, cortos, oblicuos hacia adentro y atrás, que no alcanzan los surcos axiales. Campo preglabelar ancho con ornamentación de finas estrías radiales. Surco del borde profundo, con pequeños hoyuelos. Borde anterior ancho, convexo, con líneas aterrazadas. Margen anterior suavemente curvado hacia adelante. Surco occipital con extremos dirigidos hacia adelante. Anillo occipital más ancho en su parte media (sag.) y más ancho que la glabela. Ramas anteriores de la sutura facial fuertemente divergentes, ramas posteriores normales al axis recorriendo el borde posterior. Área palpebral de las fixígenas planas, lóbulos palpebrales fuertemente convexos, reniformes.
Discusión. El material coleccionado es escaso y pequeño. El mismo presenta cierta semejanza con Kainella Kobayashi, relacionada con el diseño del campo preglabelar, las ramas anteriores de la sutura facial, y el grado de desarrollo de los lóbulos palpebrales. No obstante, hasta no contar con material adicional, los ejemplares se describen en nomenclatura abierta.
Procededencia y edad. Muestra CC6-15 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophry s minutula . Tremadociano tardío bajo.
Género Pseudokainella Harrington, 1938
Especie tipo. Pseudokainella keideli Harrington, 1938, según designación original.
Pseudokainella keideli Harrington, 1938
Figuras 5.G-H
1938. Pseudokainella keideli Harrington: 174; lám. 5, figs. 11-15.
1957. Pseudokainella keideli Harrington. Harrington y Leanza: 131- 132; figs. 51, 52 (5-10) ( cum syn ).
Material. Dos ejemplares completos, 4 fragmentos cefálicos, 3 fragmentos cranidiales, 1 pigidio, 1 ejemplar meráspido (CORDPZ 30864, 30878, 30879, 30887, 30888, 30893).
Discusión. Harrington y Leanza (1957) describieron varias formas del género en el Tremadociano del noroeste argentino. Pseudokainella keideli se diferencia de P . conica (Kobayashi) (Harrington y Leanza, 1957: fig. 52.1-4) por tener un campo preglabelar más estrecho (sag.), 3 surcos laterales en la glabela y un ángulo genal interno ( sensu Henningsmoen, 1957) agudo en lugar de recto. Pseudokainella lata (Kobayashi) (Harrington y Leanza, 1957: figs. 53, 54.1-9) posee la glabela truncada, más ensanchada posteriormente, librígenas con márgenes posteriores normales al axis y un pigidio con espinas de igual tamaño. Por su parte, Pseudokainella pustulosa Harrington y Leanza (1957: figs. 55.1-8) difiere porque presenta la parte posterior de las fixígenas más anchas (tr.), pleuras con espinas más largas, un pigaxis con 3 anillos en lugar de 4, espinas pigidiales más cortas, y la superficie del exoesqueleto granulosa.
Procedencia y edad. Muestras SG7 C, SG8-3 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Distribución geográfica. Argentina (Harrington, 1938; Harrington y Leanza, 1957).
Género Apatokephalus Brøgger, 1896
Especie tipo. Trilobites serratus Boeck, 1838, subsecuente designación de Bassler, 1915.
Apatokephalus sp.
Figura 6.G
Material. 1 fragmento cranidial (CORD-PZ 30750).
Discusión. El fragmento estudiado presenta las características diagnósticas de Apatokephalus [glabela convexa, ensanchada entre los surcos laterales; campo preglabelar estrecho (sag.); borde anterior convexo y surco del borde con hoyuelos grandes], pero dada la naturaleza del mismo, se lo cita en nomenclatura abierta.
Procedencia y edad. Muestra CC11+25 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Superfamilia Trinucleoidea Swinnerton, 1915
Familia Hapalopleuridae Harrington y Leanza, 1957
Género Hapalopleura Harrington y Leanza, 1957
Especie tipo. Hapalopleura clavata Harrington y Leanza, 1957, según designación original.
Hapalopleura sp.
Figuras 5.I-K
Material. Dos ejemplares completos (uno de ellos enrollado), 2 céfalo- tórax, 1 fragmento cranidial (CORD-PZ 30545, 30871, 30930- 30932).
Descripción. Exoesqueleto dorsal ovoide, de pequeño a mediano tamaño. Cranidio subtrapezoidal. Campo preglabelar ancho (sag.), plano, 17% del largo total del cranidio, con trazas de estrías muy finas, radiales. Borde anterior levemente curvado hacia adelante, con fuerte pendiente hacia la parte ventral, más ancho en su parte media, recorrido por líneas aterrazadas. Margen anterior subacuminado. Glabela moderamente convexa, elevada por sobre las fixígenas, bien marcada por surcos axiales rectos y divergentes hacia adelante. Surco preglabelar subredondeado. Tres pares de surcos laterales: par 3 p, apenas visible, como depresiones; par 2 p, más marcado, inclinado hacia atrás y adentro, que no alcanza los surcos axiales; 1p, bien marcado, más oblicuo que los anteriores, muy próximo al surco occipital, cuyos extremos alcanzan los surcos axiales. Áreas anteriores de las fixígenas subrectangulares, áreas posteriores subtrapezoidales, con sus extremos distales dirigidos levemente hacia atrás. En los ejemplares bien conservados su superficie está recorrida por estrías conspicuas, subparalelas al borde cefálico, que se continúan de forma más débil en la glabela y las librígenas. Aristas oculares bien marcadas, elevadas por sobre las fixígenas, estrechas, largas y rectas, dirigidas levemente hacia atrás y afuera. Ojos pequeños de posición media en el largo del cranidio (en algunos ejemplares se observan omatidios hexagonales). Librígenas estrechas, triangulares, con espinas genales largas y rectas, que siguen la dirección de los márgenes laterales.
Tórax con 14 segmentos. Axis estrecho (tr.), 17% del ancho torácico máximo. Cuatro primeras pleuras con extremos terminados en espinas cortas, pleuras siguientes con extremos redondeados. Fulcras muy distales.
Pigidio pequeño, con forma de espátula. Segmentos pleurales más estrechos (sag.) que en el tórax.
Discusión. El material estudiado es muy afín a Hapalopleura clavata Harrington y Leanza (1957: figs. 109, 110.1-6), del Tremadociano inferior de la Cordillera Oriental, en particular por la forma general de la glabela y el diseño de los surcos glabelares y las aristas oculares. No obstante, el surco preglabelar y borde anterior son subredondeados y no rectos como en H . clavata , el par de surcos 1p está más próximo al surco occipital, la parte más ancha (tr.) del tórax involucra un mayor número de segmentos, y posee dos segmentos torácicos más con espinas. Ésta es la primera cita del género en el área de Alfarcito.
Hapalopleura longicornis Harrington y Leanza (1957: figs. 111, 112.1a-e) se diferencia de Hapalopleura sp. porque la primera posee una glabela lisa y más ancha, aristas oculares dirigidas en forma oblicua hacia afuera y adelante, una espina anterior larga, un tórax con un menor número de segmentos y un axis más ancho (tr.).
Procedencia y edad. Muestras CC6-15 m, SG7 C-1,5 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophrys minutula . Tremadociano tardío bajo.
Orden Phacopida Salter, 1864
Suborden Calymenina Swinnerton, 1915
Familia Calymenidae Milne-Edwards, 1840
Género Pharostomina Sdzuy, 1955
Especie tipo. Pharostomina oepiki Sdzuy, 1955, según designación original.
Pharostomina trapezoidalis (Harrington, 1938)
Figura 5.E
1938. Pilekia trapezoidalis Harrington: 191; lám. 6, fig. 2.
1957. Colpocoryphoides trapezoidalis (Harrington). Harrington y Leanza: 223-224; fig. 123, 1-6. cf.
1982. Pharostomina cf. trapezoidalis (Harrington). Owens et al. : 16; lám. 4, fig. d.
Material. Un cranidio (CORD-PZ 30727).
Discusión. Pribyl y Vanek (1980: lám. 20, figs. 4-7; text-fig. 17) describen una nueva subespecie: Pharostomina trapezoidalis quaesita , del Ordovícico temprano de Bolivia, que se distingue de P . trapezoidalis trapezoidalis por tener un cranidio con el margen anterior recto, una granulación gruesa en su superficie y el borde posterior más estrecho en sus extremos. Pharostomina mexicana Robison y Pantoja-Alor (1968: lám. 99, figs. 7-12) presenta un margen anterior más recto que el de P . trapezoidalis y un campo preglabelar mejor definido (Robison y Pantoja-Alor, 1968). Owens et al. (1982: lám. 4 fig. d) describen un céfalo del Tremadociano de Gales que, aunque incompleto, es asignado tentativamente a P . cf. trapezoidalis . Por su parte, Pharostomina tenuisulcata Benedetto (1977b: lám. 1, fig. 16), del Cámbrico Superior de la sierra de Cajas, se distingue de P . trapezoidalis por poseer sólo dos surcos glabelares, los cuales son poco definidos y más cortos, un campo preglabelar más amplio, un borde cefálico anterior curvado hacia adelante en su parte media, y la superficie del cranidio lisa.
Procedencia y edad. Muestra CC11-0,50 m, Formación Rupasca, Zona de Bienvillia tetragonalis - Conophry s minutula . Tremadociano tardío bajo.
Distribución geográfica. Argentina (Harrington y Leanza, 1957) y Bolivia (Pribyl y Vanek, 1980).
Agradecimientos
Los autores agradecen a A. García del Río por permitir el acceso a las secciones estudiadas, a J. Toconás y a N. Maine, director y maestra respectivamente, de la Escuela Provincial Nº 352 de la localidad de Alfarcito por permitir el uso de sus instalaciones, y a A. Maine por la colaboración prestada en las tareas de campaña. I. Rábano y G. Edgecombe revisaron el manuscrito y aportaron valiosas sugerencias.
Bibliografía
1. Aceñolaza, F.G. 1968. Geología estratigráfica de la región de la sierra de Cajas, Departamento Humahuaca, provincia de Jujuy. Revista de la Asociación Geológica Argentina 23: 207-222. [ Links ]
2. Aceñolaza, F.G. 1983. The tremadocian beds and the Cambrian- Ordovician boundary problems in Latin America. Papers for the Cambrian-Ordovician boundary, Academia Sinica : 88-93. [ Links ]
3. Aceñolaza, F.G. y González, O.E. 1977. El Ordovícico de la zona de Arroyo Volcán, en la sierra de Santa Bárbara, Prov. de Jujuy. Revista del Instituto Geológico Minero 2: 125-137. [ Links ]
4. Aceñolaza, G.F. 1996. [ Bioestratigrafía del límite Cámbrico-Ordovícico y Ordovícico basal en la quebrada de Humahuaca, Provincia de Jujuy, República Argentina. Tesis Doctoral, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. Inédita, 245 pp. [ Links ]].
5. Ahlberg, P. 1989. The type species of the Ordovician agnostid trilobite Geragnostus Howell, 1935. Paläontologische Zeitschrift 63: 309-317. [ Links ]
6. Ahlberg, P. 1992. Agnostid trilobites from the Lower Ordovician of southern Sweden. Transactions of the Royal Society of Edinburgh, Earth Sciences 83: 539-570. [ Links ]
7. Amengual, R. y Zanettini, J.C.M. 1974. Geología de la quebrada de Humahuaca entre Uquía y Purmamarca (Provincia de Jujuy). Revista de la Asociación Geológica Argentina 29: 30-40. [ Links ]
8. Barrande, J. 1868. Faune silurienne des environs de Hof, en Bavière. Bulletin de la Société Géologique du France 26: 31-110. [ Links ]
9. Bassler, R.S. 1915. Bibliographic index of American Ordovician and Silurian fossils. Bulletin of United States National Museum 92: 719- 1521. [ Links ]
10. Benedetto, J.L. 1977a. Algunas consideraciones acerca de la posición del límite Cambro-Ordovícico en América del Sur. Geos 23: 3-11. [ Links ]
11. Benedetto, J.L. 1977b. Una nueva fauna de trilobites tremadocianos de la Provincia de Jujuy (sierra de Cajas), Argentina. Ameghiniana 14: 186-214. [ Links ]
12. Billings, E. 1865. Palaeozoic fossils: v1. Containing descriptions and figures of new or little known species of organic remains from the Silurian rocks. Geological Survey of Canada : 169-426. [ Links ]
13. Boeck, C. 1838. Übersicht der bisher in Norwegen gefundenen Formen der Trilobiten Familie. Gaea Norvegica 1: 138-145. [ Links ]
14. Brandán, M. 1974. [ Estudio paleontológico de las unidades ordovícicas aflorantes en la región de Cajas-Espinazo del Diablo, Departamento Humahuaca, provincia de Jujuy . Seminario II, Departamento de Ciencias Naturales, Universidad Nacional de Salta, 50 pp. Inédito] [ Links ].
15. Branisa, L. 1965. Los fósiles guías de Bolivia. Index fossils of Bolivia. Boletín del Servicio Geológico de Bolivia 6: 1-82. [ Links ]
16. Brögger, W.C. 1882. Die silurischen Etagen 2 und 3 in Kristianagebiet und auf Eker . Universitäts-program für 1882, 376 pp. [ Links ]
17. Brögger, W.C. 1896. Über die Verbreitung der Euloma-Niobe Fauna (der Ceratopygenkalkfauna) in Europa. Nyt Magazin for Naturvidenskaverne 35: 16-24. [ Links ]
18. Burmeister, H. 1843. Die Organisation der Trilobiten, aus ihren lebenden Verwandten entwickelt: nebst einer systematischen Übersicht aller zeither beschribenen Arten , 147 pp. Berlín. [ Links ]
19. Callaway, C. 1877. On a new area of Upper Cambrian rocks in south Shropshire, with a description of a new fauna. Quarterly Journal of the Geological Society of London 33: 652-672. [ Links ]
20. Clark, T.H. 1924. The Paleontology of the Beekmantown Series at Levis, Quebec. Bulletin of American Paleontology 10: 1-134. [ Links ]
21. Daniel, J. 1940. Sobre la constitución, deposición transgresiva y tectónica de los estratos mesozoicos de Alfarcito, departamento de Tilcara, provincia de Jujuy . Universidad Nacional de La Plata, Tesis del Museo 3, 69 pp. [ Links ]
22. Fortey R.A. 1980. The Ordovician trilobites of Spitsbergen. III. Remaining trilobites of the Valhallfonna Formation. Norsk. Polarinstitutt Skrifter 171: 1-163. [ Links ]
23. Fortey, R.A. y Chatterton, B.D.E. 1988. Classification of the trilobite suborder Asaphina. Palaeontology 31: 165-222. [ Links ]
24. Fortey, R.A. y Hughes, N.C. 1998. Brood pouches in trilobites. Journal of Paleontology 72: 638-649. [ Links ]
25. Fortey, R.A. y Owens, R.M. 1991. A trilobite fauna from the highest Shineton Shales in Shropshire, and the correlation of the latest Tremadoc. Geological Magazine 128: 437-464. [ Links ]
26. Frederickson, E.A. 1958. Lower Tremadocian trilobite from Venezuela. Journal of Paleontology 32: 541-543. [ Links ]
27. González, C.E. 1983. [ Evaluación faunística del Tremadociano del tramo austral de la sierra de Mojotoro, provincia de Salta, República Argentina . Tesis profesional, Facultad de Ciencias Naturales, Universidad Nacional de Salta. Inédita, 35 pp. [ Links ]].
28. Harrington, H.J. 1938. Sobre las faunas del Ordoviciano Inferior del Norte argentino. Revista del Museo de La Plata, Nueva Serie (sección Paleontología) 1: 109-289. [ Links ]
29. Harrington, H.J. y Leanza, A.F. 1943. La fáunula del Tremadociano inferior de Salitre, Bolivia. Revista del Museo de La Plata, nueva serie (sección Paleontología) 2: 343-356. [ Links ]
30. Harrington, H.J. y Leanza, A.F. 1957. Ordovician trilobites of Argentina. Department of Geology, University of Kansas, Special Publication 1, 276 pp. [ Links ]
31. Hawle, I. y Corda, A.J.C. 1847. Prodrom einer Monographie der böhmischen Trilobiten. Abhandlungen der Konigliche Bohmischen Gessellschaft Wissenschaften 5: 121-292. [ Links ]
32. Henningsmoen, G. 1957. The trilobite Family Olenidae, with descriptions of Norwegian material and remarks on the Olenid and Tremadocian Series. Skrifter Utgitt av Det Norske Videnskaps- Akademi i Oslo I Matematisk-Naturvidenskapelig Klasse 1957, 303 pp. [ Links ]
33. Howell, B.F. 1935. Cambrian and Ordovician trilobites from Herault, southern France. Journal of Paleontology 9: 222-238. [ Links ]
34. Jaekel, O. 1909. Ueber die Agnostiden. Zeitschrift der Deutschen Geologischen Gesellschaft 61: 380-401. [ Links ]
35. Kayser, E. 1876. Über Primordiale und Untersilurische Fossilien aus der Argentinischen Republik. Beitrage zur Geologie und Paläontologie der Argentinischen Republik, 2. Palaeontographica Cassel, Supplementum 3: 1-33. [ Links ]
36. Kobayashi, T. 1936. On the Parabolinella Fauna from Province Jujuy, Argentina with a note on the Olenidae. Japanese Journal of Geology and Geography 13: 85-102. [ Links ]
37. Linnarsson, J.G.O. 1869. Om Vestergötlands Cambriska och Siluriska Aflagringar. Kongliga Svenska Vetenskapsakademiens, Handlingar 8: 1-89. [ Links ]
38. López, C.R. y Nullo, F.E. 1969. Geología de la margen izquierda de la quebrada de Humahuaca, de Huacalera a Maimará, Departamento Tilcara, Provincia de Jujuy, República Argentina. Revista de la Asociación Geológica Argentina 24: 173-182. [ Links ]
39. Lu, Y. 1975. Ordovician trilobite faunas of central and southwest China. Palaeontologia Sinica, New Series (B) 11: 484 pp. [ Links ]
40. Lu, Y.-H. y Lin, H.-L. 1984. Late late Cambrian and earliest Ordovician trilobites of Jianghan-Changsshan Area, Zhejiang. En: Stratigraphy and Palaeontology of systemic boundaries in China, Cambrian- Ordovician boundary . Anhui Science and Technology Publishing House, Nanjing, 1: pp. 45-143. [ Links ]
41. Ludvigsen, R. y Tuffnell, P.A. 1983. A revision of the Ordovician olenid trilobite Triarthus Green. Geological Magazine 120: 567-577. [ Links ]
42. Malanca, H.S. 1973. [ Estudio paleontológico del Ordovícico aflorante en el tramo inferior de la quebrada de Purmamarca (Dto. Tumbaya, Pcia. de Jujuy). Seminario I, Departamento de Ciencias Naturales, Universidad Nacional de Salta, 48 pp. Inédito] [ Links ].
43. Milne-Edwards, H. 1840. Histoire naturelle des Crustacés, comprenant l'anatomie, la physiologie et la classification des animaux 3, 638 pp. [ Links ]
44. Paris: Roret. Moberg, J.C. y Möller, H. 1898. Om Acerocare-zonen. Geologiska Föreningens i Stockholm Förhandlingar 20: 197-290. [ Links ]
45. Mon, R., Rahmer, S. y Mena, R. 1993. Estructuras superpuestas en la Cordillera Oriental, Provincia de Jujuy, Argentina. 12º Congreso Geológico Argentino y 2º Congreso de Exploración de Hidrocarburos , Actas 3: 48-54. [ Links ]
46. Moya, M.C. 1988. [ Estratigrafía del Tremadociano en el tramo austral de la Cordillera Oriental argentina . Tesis doctoral, Facultad de Ciencias Naturales, Universidad Nacional de Salta, 368 pp. Inédita] [ Links ].
47. Moya, M.C., Malanca, S., Monteros, J.A. y Cuerda, A. 1994. Bioestratigrafía del Ordovícico inferior en la Cordillera Oriental argentina basada en graptolitos. Revista Española de Paleontología 9: 91-104. [ Links ]
48. Nielsen, A.T. 1997. A review of Ordovician agnostid genera (Trilobita). Transactions of the Royal Society of Edinburgh, Earth Sciences 87: 463-501. [ Links ]
49. Nikolaisen, F. y Henningsmoen, G. 1985. Upper Cambrian and lower Tremadoc olenid trilobites from the Digermul Peninsula, Finnmark, northern Norway. Bulletin Norges geologiske undersøkelse 400: 1-49. [ Links ]
50. Owens, R.M., Fortey, R.A., Cope, J.C.W., Rushton, A.W.A. y Bassett, M.G. 1982. Tremadoc faunas from the Carmarthen district, South Wales. Geological Magazine 119: 1-38. [ Links ]
51. Peng, S. 1991. Tremadocian trilobites from Guotang Formation, Luxi, Western Hunan. Acta Palaeontologica Sinica 30: 141-166. [ Links ]
52. Pek, I. 1977. Agnostid trilobites of the Central Bohemian Ordovician. Sborník Geologických Ved, Paleontologie 19: 7-44. [ Links ]
53. Pribyl, A. y Vanek, J. 1980. Ordovician trilobites of Bolivia. Rozpravy Ceskoslovenské Akademie Ved 90: 1-90. [ Links ]
54. Rao, R.I. y Tortello, M.F. 1998. Tremadoc conodonts and trilobites from the Cardonal Formation, Incamayo Creek, Salta Province, northwestern Argentina. En: H. Szaniawski (ed.), Proceedings of the 6th European Conodont Symposium (ECOS VI). Palaeontologia Polonica 58: 31-45 [ Links ]
55. Raw, F. 1908. The trilobite fauna of the Shineton Shales. Reports of the British Association for the Advancement of Science 77: 511-513. [ Links ]
56. Robison, R.A. y Pantoja-Alor, J. 1968. Tremadocian trilobites from the Nochixtlán region, Oaxaca, México. Journal of Paleontology 42: 767- 800. [ Links ]
57. Rushton, A.W.A. 1988. Tremadoc trilobites from the Skiddaw Group in the English Lake District. Palaeontology 31: 677-698. [ Links ]
58. Salfity, J.A., Malanca, S., Moya, M.C., Monaldi, C.R. y Brandán, E.M. 1984. El límite Cámbrico-Ordovícico en el norte de la Argentina. 9º Congreso Geológico Argentino, Actas 1: 568-575. [ Links ]
59. Salter, J.W. 1849 . Figures and descriptions illustrative of British organic remains. Memoirs of the Geological Survey of United Kingdom, decade 2, 39 pp. [ Links ]
60. Salter, J.W. 1864. Figures and descriptions illustrative of British organic remains. Trilobites (chiefly Silurian). Memoirs of the Geological Survey of United Kingdom, decade 11, 60 pp. [ Links ]
61. Sdzuy, K. 1955. Die Fauna der Leitmitz-schiefer (Tremadoc). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 492: 1-74. [ Links ]
62. Shergold, H.J. 1988. Review of trilobite biofacies distributions at the Cambrian-Ordovician Boundary. Geological Magazine 125: 363-380. [ Links ]
63. Shergold, J.H., Laurie, J.R. y Sun, X. 1990. Classification and review of the trilobite Order Agnostida Salter, 1864: an Australian perspective. Reports of the Bureau of Mineral Resources, Geology and Geophysics 296: 1-93. [ Links ]
64. Steinmann, G. y Hoek, H. 1912. Das Silur und Cambrium des Hochlandes von Bolivia Und ihre Fauna. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 34: 176-252. [ Links ]
65. Swinnerton, H.H. 1915. Suggestions for a revised classification of trilobites. Geological Magazine, New Series 2: 538-545. [ Links ]
66. Tjernvik, T.E. 1956. On the Early Ordovician of Sweden-Stratigraphy and fauna. Bulletin of the Geological Institutions, University of Uppsala 36: 107-284. [ Links ]
67. Tortello, M.F. 1996a. Geragnostus nesosii Harrington y Leanza, 1957 (Trilobita, Agnostida) en el Tremadociano superior del noroeste argentino. 13º Congreso Geológico Argentino y 3º Congreso de Exploración de Hidrocarburos, Actas 5: 17-25. [ Links ]
68. Tortello, M.F. 1996b. Trilobites agnóstidos del Tremadociano superior alto (Ordovícico Inferior) del noroeste argentino. Memorias del 12º Congreso Geológico de Bolivia, Tomo 1: 203-209. [ Links ]
69. Tortello, M.F. y Aceñolaza, G.F. 1993. Trilobites agnóstidos del límite Cámbrico-Ordovícico de la Formación Lampazar, sierra de Cajas, Provincia de Jujuy, Argentina. Revista Española de Paleontología 8: 177-184. [ Links ]
70. Tortello, M.F. y Esteban, S.B. 1999. La transición Cámbrico-Ordovícico en la Formación Volcancito (sierra de Famatina, La Rioja, Argentina). Ameghiniana 36: 371-387. [ Links ]
71. Tortello, M.F. y Esteban, S.B. 2003. Trilobites del Cámbrico Tardío de la Formación Lampazar (sierra de Cajas, Jujuy, Argentina). Implicancias bioestratigráficas y paleoambientales. Ameghiniana 40: 323-344. [ Links ]
72. Tortello, M.F. y Rao, R.I. 2000. Trilobites y conodontes del Ordovícico temprano del Angosto de Lampazar (Provincia de Salta, Argentina). Boletín Geológico y Minero 111: 61-84. [ Links ]
73. Ulrich, E.O. 1930. Ordovician trilobites of the family Telephidae and concerned stratigraphic correlations. Proceedings of the United States National Museum 76: 1-101. [ Links ]
74. Ulrich, E.O. y Resser, C.E. 1930. Cambrian of the upper Mississipi valley. Part 1: Trilobita; Dikelocephalinae and Osceolinae. Mississippi; Bulletin of the Milwaukee Public Museum 12: 1-122. [ Links ]
75. Waisfeld, B.G. y Vaccari, N.E. 2003. Trilobites. En: J.L. Benedetto (ed.), Ordovician fossils of Argentina . Secretaría de Ciencia y Tecnología, Universidad Nacional de Córdoba, pp. 295-339. [ Links ]
76. Walcott, C.D. 1924. Cambrian geology and paleontology (V): Cambrian and Lower Ozarkian trilobites. Smithsonian Miscellaneous Collections 75: 51-60. [ Links ]
77. Walcott, C.D. 1925. Cambrian geology and paleontology (V): Cambrian and Ozarkian trilobites. Smithsonian Miscellaneous Collections 75: 61-146. [ Links ]
78. Whitehouse, F.W. 1936. The Cambrian faunas of northeastern Australia. Parts 1 and 2. Memoirs of the Queensland Museum 11: 59- 112. [ Links ]
79. Zeballo F.J., Albanesi, L.G. y Ortega, G. 2005. Conodontes y graptolitos de las formaciones Alfarcito y Rupasca (Tremadociano) en el área de Alfarcito, Tilcara, Cordillera Oriental de Jujuy, Argentina. Parte 1: Bioestratigrafía. Ameghiniana 42: 39-46. [ Links ]
Recibido: 5 de noviembre de 2003.
Aceptado: 13 de mayo de 2004.

