










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.3 Buenos Aires jun./sept. 2005
Nuevos restos fósiles de Mendozasaurus neguyelap (Sauropoda, Titanosauria) del Cretácico Tardío de Mendoza, Argentina
Bernardo J. González Riga 1
1 Unidad de Paleovertebrados, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Centro Regional de Investigaciones Científicas y Tecnológicas, Avda. Ruiz Leal s/n, Parque Gral. San Martín, 5500 Mendoza, Argentina. bgonriga@lab.cricyt.edu.ar
Abstract. N ew fossil remains of M endozasaurus neguyelap (Sauropoda, Titanosauria ) from the Late Cretaceous of Mendoza, Argentina . In South America, most titanosaur species are represented by incomplete skeletal elements lacking well-preserved cervical vertebrae. In this context, the discovery of cervical remains assigned to Mendozasaurus neguyelap González Riga is relevant from a systematic viewpoint. The fossils were found in the paleontological site and assemblage of the holotype, Río Neuquén Subgroup, late Turonian – late Coniacian from Mendoza Province, Argentina. The cervical vertebrae of Mendozasaurus exhibit differences with those of most titanosaurs; however, they share with Isisaurus colberti (Jain and Bandyopadhyay) from Maastrichthian of India the presence of: a) short vertebral centra (ratio: total length/height of cotyle less than 2.5), b) large and deep supradiapophyseal fossa, and c) relatively tall neural spines (ratio: vertebral height / centrum length more than 1.5). The fossils recovered show an autapomorphic character that enlarges the diagnosis of Mendozasaurus : tall, laminar and transversally expanded mid-posterior cervical neural spines that are wider than vertebral centra and fan-like' or subrhomboid' in shape due to lateral expansions and a subrounded dorsal border. The fossil record of titanosaurs shows a notable morphologic diversity in the cervical series. In particular, Mendozasaurus neguyelap and Isisaurus colberti possess tall neural spines associated with the proportionally shortest cervical centra of any titanosaur. This unusual morphology suggests the development of relatively wide, robust and short necks in Late Cretaceous sauropods from Argentina and India.
Resumen. En América del Sur la mayoría de las especies de titanosaurios están representadas por restos esqueletales incompletos que no tienen preservadas sus vértebras cervicales. En este contexto, el hallazgo de restos cervicales de Mendozasaurus neguyelap González Riga es relevante desde el punto de vista sistemático. Los restos provienen del mismo sitio paleontológico y asociación fósil del holotipo, Subgrupo Río Neuquén, Turoniano tardío - Coniaciano tardío de la provincia de Mendoza, Argentina. Las vértebras cervicales de Mendozasaurus exhiben caracteres novedosos con respecto a las de otros titanosaurios, si bien comparten con Isisaurus colberti (Jain y Bandyopadhyay) del Maastrichthiano de la India la presencia de: 1) centros vertebrales relativamente cortos (relación longitud del centro/altura del cótilo: inferior a 2,5), 2) profunda y amplia fosa supradiapofisial y 3) altas espinas neurales (relación altura vertebral/longitud del centro: mayor a 1,5). Los restos hallados muestran un carácter que amplía la diagnosis de Mendozasaurus : vértebras cervicales medias/posteriores con espinas neurales altas, laminares y transversalmente expandidas (más anchas que los centros) de contorno anterior en abanico o sub-romboidal por el desarrollo de expansiones laterales y un extremo dorsal sub-redondeado. El registro fósil de titanosaurios exhibe una notable diversidad morfológica en la serie cervical. En particular Mendozasaurus neguyelap e Isisaurus colberti se encuentran entre los titanosaurios que poseen los centros cervicales proporcionalmente más cortos asociados con altas espinas neurales. Esta inusual morfología evidencia el desarrollo de cuellos relativamente gruesos, robustos y cortos en saurópodos del Cretácico Tardío de Argentina y la India.
Key words. Dinosauria. Titanosauria. Cervical vertebrae. Late Cretaceous. Mendoza. Argentina.
Palabras clave. Dinosauria. Titanosauria. Vértebras cervicales. Cretácico Tardío. Mendoza. Argentina.
Introducción
En América del Sur el registro de dinosaurios saurópodos titanosaurios es abundante y diverso. Si bien se han descrito más de una decena de especies, la mayoría de ellas están representadas por esqueletos incompletos integrados principalmente por vértebras caudales y huesos apendiculares desarticulados. En este contexto, los restos cervicales son escasos. Por ejemplo, en los titanosarios Andesaurus delgadoi (Calvo y Bonaparte, 1991), Aeolosaurus rionegrinus (Powell, 1986, 2003; Salgado y Coria, 1993; Salgado et al ., 1997b), Pellegrinisaurus powelli (Salgado, 1996), Argyrosaurus superbus (Lydekker, 1893; Powell, 1986) y Argentinosaurus huinculensis (Bonaparte y Coria, 1993) no se conocen restos cervicales. Así mismo, los restos cervicales disponibles de Rocasaurus muniozi (Salgado y Azpilicueta, 2000), Laplatosaurus araukanicus (Huene, 1929; Powell, 1986; Salgado, 2003), Gondwanatitan faustoi (Kellner y Acevedo, 1999) y Titanosaurus ? nanus (Lydekker, 1893; Powell, 1986) son incompletos y relativamente fragmentarios. En contraste, sólo Saltasaurus loricatus (Bonaparte y Powell, 1980; Powell, 2003), Neuquensaurus australis (Salgado et al ., en prensa), Titanosauria indet. DGM Serie A y Serie B de Brasil (Powell, 1987; 2003), Rinconsaurus caudamirus (González Riga y Calvo, 2001; Calvo y González Riga, 2003) y nuevos ejemplares recientemente hallados (Calvo et al ., 1997) muestran vértebras cervicales bien preservadas. Evidentemente, este registro incompleto de la secuencia cervical limita el estudio del clado. Por ello, el hallazgo de nuevos restos cervicales es relevante desde el punto de vista sistemático y merece posteriores análisis filogenéticos y morfo-funcionales.
En esta contribución se describen y analizan sistemáticamente vértebras cervicales asignadas a Mendozasaurus neguyelap. La serie tipo de esta especie está integrada por vértebras dorsales y caudales, y la mayor parte de las extremidades anteriores y posteriores (González Riga, 2002; 2003a).
Los fósiles descritos fueron hallados y preparados por el autor y asistentes entre los años 2001-2003. Provienen de la localidad Arroyo Seco, sur del cerro Guillermo, provincia de Mendoza, del mismo sitio paleontológico y asociación fósil del holotipo de Mendozasaurus . En este sitio han sido hallados huesos de titanosaurios de diverso tamaño y desarrollo ontogenético junto con restos fragmentarios de dinosaurios Theropoda. Los fósiles provienen de facies de desbordamento ( crevasse splay ) relacionadas con sistemas fluviales de moderada - alta sinuosidad (meandriformes) desarrollados sobre extensas llanuras de inundación (figura 1). El sitio paleontológico corresponde a un modo tafonómico ( sensu Behrensmeyer y Hook, 1992) denominado acumulaciones óseas de desbordamiento, caracterizado por acumulaciones autóctonas y parautóctonas de huesos articulados y desarticulados depositados en facies fluviales de desbordamiento (González Riga et al ., 2003; 2004). En su historia tafonómica se reconoce una primera etapa bioestratinómica que comprendió procesos de desarticulación y orientación de huesos por corrientes de desbordamiento. Los restos, cubiertos por sedimentos antes de la completa descomposición de los tejidos blandos, provienen de vaques feldespáticos micáceos que indican intensos procesos de iluviación y bioturbación. La segunda etapa tafonómica (fosildiagénesis) incluyó procesos de permineralización y compresión litostática (González Riga, 2002). Desde el punto de vista paleoambiental y paleoecológico, la abundancia de grandes vertebrados herbívoros, troncos de coníferas, trazas y bioturbación indican, para esta región de la cuenca Neuquina en el intervalo Turoniano tardío-Coniaciano tardío, una intensa actividad biológica y desarrollo de vegetación en planicies fluviales relativamente bien drenadas sujetas a inundaciones periódicas (González Riga, 2003b).

Figura 1. Columna estratigráfica del Subgrupo Río Neuquén en Arroyo Seco (cerro Guillermo, provincia de Mendoza, Argentina), mostrando niveles fosilíferos y asociaciones de facies/ Stratigraphic column of the Río Neuquén Subgroup at Arroyo Seco site (cerro Guillermo, Mendoza Province, Argentina) showing the fossiliferous levels and facies associations. Referencias/ References : 1, conglomerados/ conglomerates ; 2, areniscas/ sandstones ; 3, pelitas/ pelites ; 4, estratificación entrecruzada en artesa/ trough-cross bedding ; 5, estratificación entrecruzada planar/ planar cross bedding ; 6, laminación horizontal/ horizontal bedding ; 7, ondulitas/ riples ; 8, dirección de paleocorriente/ paleocurrent data ; 9, fósiles de dinosaurios/ dinosaur fossils ; 10, trazas y bioturbación/ traces and bioturbation .
Abreviaturas institucionales. DGM, Museo de la Divisao Geologia y Mineralogia, Rio de Janeiro, Brasil; IANIGLA-PV, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Colección Paleovertebrados, Mendoza, Argentina; MCF-PVPH, Museo Carmen Funes, Paleontología de Vertebrados, Plaza Huincul, Neuquén, Argentina; PVL, Colección de Paleontología de Vertebrados de la Fundación Instituto Miguel Lillo, Tucumán, Argentina.
Paleontología sistemática
S aurischia Seeley, 1888
S auropoda Marsh, 1878
T itanosauria Bonaparte y Coria, 1993
Mendozasaurus neguyelap González Riga, 2003a
Figuras 2-8
Diagnosis modificada . Titanosaurio de gran tamaño (18-25 m de largo) caracterizado por la siguiente asociación de caracteres únicos: 1, vértebras cervicales medias-posteriores con espinas neurales altas, laminares y transversalmente expandidas (más anchas que los centros) de contorno anterior en abanico o sub-romboidal por el desarrollo de expansiones laterales y un extremo dorsal sub-redondeado; 2, cavidad interzigapofisial de vértebras caudales anteriores extendida dorsoventralmente y limitada dorsalmente por las láminas espinopostzigapofisial y espinoprezigapofisial; 3, centros caudales medios suavemente procélicos con cóndilos posteriores muy reducidos y desplazados dorsalmente; 4, espinas neurales laminares en vértebras caudales medias-posteriores, con un borde dorsal horizontal y un ángulo ánterodorsal recto; 5, grandes placas óseas subcónicas-subesféricas sin cingulum . Además la presencia de las siguientes sinopomorfías caracteriza al nuevo taxón: centros cervicales medios relativamente cortos (longitud total/altura del cótilo < 2,5), profunda y amplia fosa supradiapofisial en vértebras cervicales; relación: altura de la vértebra cervical/longitud del centro > 1,5; procesos transversos cervicales con forma de ala en vistas anterior y ventral debido a la presencia de amplias láminas prezigodiapofisiales de borde anterior convexo; lámina prespinal extendida hasta la base de la espina neural en vértebras dorsales anteriores; pleurocelos acuminados en vértebras dorsales anteriores; ausencia de articulación hipósfeno-hipantro en vértebras dorsales anteriores; centros caudales anteriores fuertemente procélicos con cóndilos posteriores prominentes; arcos neurales de las vértebras caudales medias y posteriores ubicados en la mitad anterior de los centros; ángulo ánterodorsal de la espina neural de las caudales medias situado anteriormente con respecto al margen anterior de las postzigapófisis; espinas neurales axialmente elongadas y laminares en vértebras caudales medias; prezigapófisis relativamente largas en vértebras caudales medias; hemapófisis con articulaciones proximales abiertas; placa esternal de contorno semilunar con un borde posterior relativamente recto; borde anterior de la hoja escapular cóncavo en su porción proximal y recto en su porción distal; húmero con un borde proximal relativamente recto que se curva en su extremo medial; metacarpos sin facetas articulares distales; fémur con una protuberancia lateral debajo del trocánter mayor.
Observaciones. Con respecto a la diagnosis original (González Riga, 2003a) se agrega aquí un carácter cervical (carácter 1) y se excluyen dos caracteres de las vértebras dorsales anteriores propuestos anteriormente como probables autapomorfías: la presencia de dos fosas infrapostzigapofisiales subtriangulares y el desarrollo de láminas postzigapostespinales paralelas a los planos de las facetas postzigapofisiales. Estos caracteres se encuentran presentes, con algunas variantes morfológicas, en algunos titanosaurios recientemente descubiertos en el norte de Patagonia; en particular en un nuevo taxón procedente de Rincón de los Sauces (Calvo y González Riga, en preparación) y en Bonitasaura salgadoi (Apesteguía, 2004 y com. pers.).
Por otro lado, la reciente publicación de un ejemplar bien preservado de Epachthosaurus sciutoi (Martínez et al ., 2004) del Cenomaniano tardío-Turoniano temprano de Patagonia, permite reconocer similitudes con Mendozasaurus neguyelap . En primer lugar, Epachtosaurus muestra una progresiva reducción de los cóndilos articulares posteriores en sus caudales medias y posteriores (Martínez et al ., 2004). No obstante, no muestran la morfología presente en Mendozasaurus (carácter 3), donde la cara articular posterior de los centros vertebrales es plana en su mitad inferior y muestra un cóndilo muy reducido o casi ausente en su mitad superior (González Riga, 2003a). En segundo lugar, si bien las espinas neurales de las vértebras caudales mediasposteriores son semejantes en ambos taxones, se reconocen diferencias. En contraste con Mendozasaurus (véase carácter 4), Epachthosaurus posee espinas neurales con un ángulo ántero-dorsal obtuso y un borde dorsal que desciende suavemente hacia posterior. Finalmente, las espinas neurales de las vértebras caudales medias-posteriores de Andesaurus delgadoi (Calvo y Bonaparte, 1991), si bien también algo similares a las descritas, se diferencian de Mendozasaurus por poseer un borde dorsal ligeramente curvado que se elevada hacia posterior y un ángulo anterodorsal más redondeado.
Material asignado . IANIGLA-PV 076/1-4: cuatro vértebras cervicales asociadas, una de ellas casi completa, pertenecientes a un ejemplar adulto. IANIGLA-PV 084: una cervical posterior atribuida a un espécimen adulto de gran tamaño.
Procedencia estratigráfica y geográfica. Niveles asignados al Subgrupo Río Neuquén (Turoniano tardío - Coniaciano tardío según Leanza y Hugo, 2001), Arroyo Seco, región sur del cerro Guillermo, departamento de Malargüe, provincia de Mendoza, cuenca Neuquina.
Descripción
Desde el punto de vista metodológico se adopta la nomenclatura vertebral propuesta por Janensch (1929; 1950) y Wilson (1999).
Vértebras cervicales medias posteriores (figuras 2-3). Se ha recuperado una vértebra cervical casi completa que yacía in situ acostada sobre su cara anterior. Esta vértebra estaba articulada con otra cervical, la cual sólo ha preservado parte de su espina neural y postzigapófisis. La vértebra más completa (figura 2, IANIGLA-PV 076/1) exhibe un centro vertebral opistocélico, dorsoventralmente deprimido. Es relativamente corto, con un índice de elongación (IE = longitud del centro /altura del cótilo sensu Wedel et al ., 2000) inferior a 2,5 (tabla 1). No se observan evidencias de aplastamiento sobre el arco neural, lo que indica que la inusual longitud del centro constituye un carácter morfológico, si bien la convexidad del cóndilo pudo haber sido levemente modificada por presión litostática. El cóndilo muestra un contorno elipsoidal más reducido que el cótilo. Las parapófisis son extensas y se orientan lateroventralmente. Se localizan en la mitad anterior del centro y se unen al mismo mediante una amplia lámina centroparapofisial ( sensu Wedel et al ., 2000) que se extiende hasta el borde posterior del cuerpo vertebral.

Tabla 1. Vértebras cervicales de Mendozasaurus neguyelap (medidas en mm)/ Cervical vertebrae of Mendozasaurus neguyelap (measurements in mm). (*medida estimada/ estimated measurement )

Figura 2. Vértebra cervical media-posterior de Mendozasaurus neguyelap en vista anterior (A), lateral (B) y ventral (C) / mid-posterior cervical vertebra of Mendozasaurus neguyelap in anterior (A) , lateral (B) and ventral (C) views , IANIGLA-PV 076/1. Referencias/ References : cm: cresta medial/ medial crest ; dp: diapófisis/ diapophysis ; lcda / acdl: lámina centrodiapofisial anterior/ anterior centrodiapophyseal lamina ; lcdp / cdl: lámina centrodiapofisial posterior/ posterior centrodiapophyseal lamina ; lepo / spol: lámina espinopostzigapofisial/ spinopostzygapophyseal lamina ; lepr / sprl: lámina espinoprezigapofisial/ spinoprezygapophyseal lamina ; el: expasión lateral de la espina neural/ lateral expansion of the neural spine ; lprd / prdl: lámina prezigadiapofisial/ prezygadiapophyseal lamina ; pp: parapófisis/ parapophysis ; poz: postzigapófisis/ postzygapophysis ; prz: prezigapófisis/ prezygapophysis . Escala gráfica: 10 cm/ Scale bar: 10 cm .

Figura 3. Vértebra cervical media-posterior de Mendozasaurus neguyelap en vista posterior/ mid-posterior cervical vertebra of Mendozasaurus neguyelap in posterior view , IANIGLA-PV 076/1. Referencias / References : dp: diapófisis/ diapophysis ; lepo / spol: lámina espinopostzigapofisial/ pinopostzygapophyseal lamina ; el: expansión lateral de la espina neural/ lateral expansion of the neural spine ; pp: parapófisis/ parapophysis ; poz: postzigapófisis/ postzygapophysis ; Escala gráfica: 10 cm/ Scale bar: 10 cm.
La cara ventral del centro (figura 2.C) es fuertemente cóncava en sentido láteromedial y muestra una suave quilla o cresta medial en la porción anterior. En contraste, la porción posterior es convexa en sentido anteroposterior y lateromedial.
La cara lateral del centro (figura 2.B) exhibe una amplia y profunda cavidad limitada ventralmente por la lámina centroparapofisial posterior y dorsalmente por una lámina horizontal accesoria. Sobre ésta se desarrollan dos cavidades infradiapofisiales de contorno subtriangular separadas por la lámina centrodiapofisial anterior. Esta lámina, parcialmente preservada, es una estructura muy delgada (~1-2 mm). Las láminas centrodiapofisiales anteriores y posteriores se unen cerca de la faceta articular de la diapófisis.
La diapófisis se encuentra sobre la parapófisis yen una posición relativamente baja, a nivel de la mitad del centro vertebral. Los procesos transversos están muy extendidos lateralmente. Se encuentran soportados por amplias láminas prezigodiapofisiales y centrodiapofisiales posteriores, las que forman una extensa superficie posteroventralmente inclinada. El borde anterior convexo de las láminas prezigodiapofisiales otorga a los procesos transversos forma de ala en vista ventral y anterior (figuras 2.A, 2.C). Esta morfología constituye un carácter novedoso entre los titanosaurios. Si bien en algunos otros saurópodos aparecen láminas prezigodiapofisiales relativamente extensas, no muestran procesos transversos con forma de ala. Por ejemplo, en Jobaria tiguidensis , un Eusauropoda del Cretácico Inferior de Nigeria, Sereno et al . (1999) describen un proceso anterior accesorio cerca de la prezigapófisis. Por otro lado, en la última cervical y primera dorsal de Haplocanthosaurus priscus , Hatcher (1903) describe expansiones de los procesos transversos que probablemente se relacionan con la inserción de los músculos de la escápula.
El arco y espina neural están desplazados hacia anterior (figura 2.B), de tal forma que la prezigapófisis sobrepasa el borde anterior del cóndilo. Lateralmente, el arco neural exhibe una amplia y profunda cavidad supradiapofisial que llega casi hasta el plano medial de la vértebra. Esta cavidad está limitada dorsalmente por las láminas espinopostzigapofisial y espinoprezigapofisial. Tanto las prezigapófisis como las postzigapófisis están proyectadas lateralmente (figura 2.A) de manera que alcanzan una posición más lateral que las parapófisis. Este carácter también está presente en otros titanosaurios como Saltasaurus y en el Titanosauria indet. DGM Serie A de Brasil (Powell, 1987; Bonaparte, 1999). Las prezigapófisis muestran amplias facetas articulares de contorno subtriangular. Están unidas entre sí mediante una robusta lámina intraprezigapofisial, la cual describe un contorno cóncavo hacia dorsal. Las láminas espinoprezigapofisiales constituyen pilares que llegan hasta la mitad de la altura de la espina neural.
En vista posterior (figura 3), el arco neural muestra dos gruesas láminas espinopostzigapofisiales que se extienden dorsalmente, pero no convergen. Entre ellas se desarrolla una amplia cavidad, ya que la lámina postespinal está ausente.
La espina neural es indivisa, alta, laminar y lateralmente muy expandida. En vistas anterior y posterior exhibe un peculiar contorno en abanico o subromboidal, con un borde dorsal redondeado y dos expansiones laterales que describen un ángulo obtuso. El tamaño relativo de la espina en relación al centro vertebral, como así también su morfología caracterizada por dos expansiones laterales, constituye una probable autapomorfía de Mendozasaurus . El borde dorsal de la espina neural es relativamente grueso (~30-35 mm) y ligeramente cóncavo en su cara posterior. No se observa una lámina prespinal, aunque un reborde óseo parcialmente preservado sugiere la presencia de una incipiente y corta lámina prespinal situada en la base de la espina neural.
Existen materiales fósiles en preparación (IANIGLA-PV 84) procedentes del mismo sitio que corresponden a un ejemplar adulto de talla gigantesca. Entre los fósiles recuperados se destaca una vértebra cervical que tiene unos 100 cm de ancho y 90 cm de altura. Observaciones preliminares permiten reconocer en esta cervical una morfología similar a la descripta, con algunas variantes en el contorno de la espina neural, que es relativamente más alta y de contorno anterior más cuadrangular. No obstante, la falta de material más completo y articulado impide, por el momento, describir con precisión los cambios morfológicos de la espina neural a lo largo de la secuencia cervical y dorsal.
Vértebra cervical posterior (figura 4). Se ha hallado una vértebra cervical posterior incompleta (IANIGLA-PV 076/3) que ha preservado su centro vertebral, y su prezigapófisis y diapófisis derecha. Ha sufrido un aplastamiento litostático en sentido dorsoventral y su arco neural ha sido desplazado hacia su flanco derecho (figura 4.C). El centro vertebral es opistocélico y más largo que el centro de la cervical media-posterior. La distorsión del centro no permite conocer con precisión la altura del cótilo. No obstante, si la misma puede estimarse en 180 mm, el índice de elongación del centro vertebral sería inferior a 2,5 (tabla 1). A diferencia de la vértebra cervical mediaposterior, la cara ventral del centro (figura 4.B) es anteroposteriormente convexa en su extremo anterior y cóncava en su porción posterior. Carece de cresta medial y muestra parapófisis con facetas articulares extensas, ubicadas en la mitad del centro.
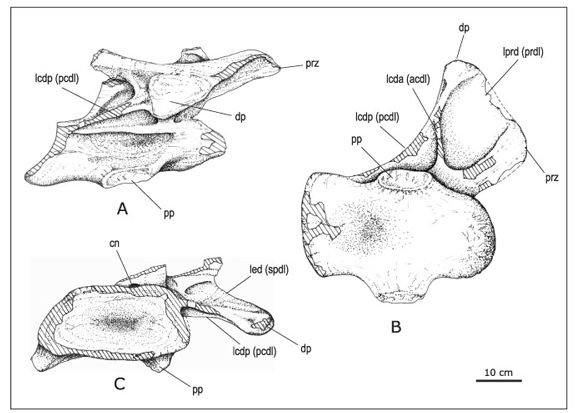
Figura 4. Vértebra cervical posterior de Mendozasaurus neguyelap en vista lateral (A), ventral (B) y posterior (C)/ posterior cervical vertebra of Mendozasaurus neguyelap in lateral (A), ventral (B), and posterior (C) views , IANIGLA-PV 076/3; Referencias/ References : cn: canal neural/ neural canal ; dp: diapófisis/ diapophysis ; lcda / acdl: lámina centrodiapofisial anterior/ anterior centrodiapophyseal lamina ; lcdp / pcdl: lámina centrodiapofisial posterior/ posterior centrodiapophyseal lamina ; led / spdl: lámina espinodiapofisial/ spinodiapophyseal lamina ; lprd / prdl: lámina prezigadiapofisial/ prezygadiapophyseal lamina ; pp: parapófisis/ parapophysis ; prz: prezigapófisis / prezygapophysis . Escala gráfica: 10 cm/ Scale bar: 10 cm.
La cara lateral del centro no presenta variaciones con respecto al centro cervical medio-posterior. Se observa una profunda y elongada cavidad lateral, sin septos (figura 4.A). Dorsalmente, una lámina horizontal accesoria separa esta cavidad de una fosa lateral subtriangular limitada por las láminas centrodiapofisiales anterior y posterior. El borde posteroventral del centro vertebral se extiende posteriormente con respecto a su borde posterodorsal. Este carácter, presente en otros titanosaurios, está relacionado con la curvatura natural del cuello en posición de vida (Lehman y Coulson, 2002).
Comparte con la cervical media-posterior (IANIGLA-PV 076/1) el gran desarrollo lateral de los procesos transversos, con extensas láminas prezigodiapofisial y centrodiapofisial posterior (hay 400 mm entre el centro del canal neural y la diapófisis). En contraste con la cervical media-posterior, la diapófisis muestra en esta cervical una faceta articular más robusta y de contorno subtriangular (figura 3.A). La diapófisis está soportada por las láminas centrodiapofisial posterior, centrodiapofisial anterior, prezigadiapofisial y espinodiapofisial. Esta última lámina, característica de las vértebras dorsales de los saurópodos (Wilson, 1999), comienza a aparecer en Mendozasarus en este sector posterior de la serie cervical.
Las láminas centrodiapofisial anterior y posterior son muy delgadas y se unen antes de alcanzar la diapófisis, a diferencia de lo que acontece en la cervical media-posterior. La lámina centrodiapofisial posterior, cerca de la faceta articular de la diapófisis, circunscribe una pequeña depresión posterior (figura 4.B). La cara dorsal de este arco neural no está bien preservada y la prezigapófisis preservada ha sido fuertemente aplastada. No obstante, se reconoce la presencia de láminas espinoprezigapofisiales y espinodiapiofisiales que soportan dorsalmente al proceso transverso.
Vértebra cervico-dorsal (figuras 5.A-B). Se ha preservado un arco neural incompleto que corresponde a una vértebra cervico-dorsal o primera dorsal (IANIGLA-PV 076/4). A pesar de su carácter fragmentario, es significativo porque sirve de nexo entre las cervicales descriptas y la dorsal anterior (IANIGLA-PV066; González Riga, 2003a), la cual probablemente corresponde a una tercera dorsal (figura 5.C).
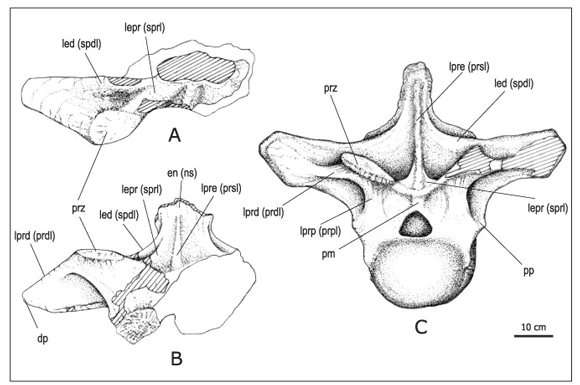
Figura 5. Mendozasaurus neguyelap . A-B, vértebra cervico-dorsal en vista dorsal (A) y anterior (B)/ cervico-dorsal vertebra in dorsal (A) and anterior (B) views , IANIGLA-PV 076/4; C, vértebra dorsal anterior en vista anterior/ anterior dorsal vertebra in anterior view , IANIGLA-PV 066. Referencias/ References : dp: diapófisis/ diapophysis ; en / ns: espina neural/ neural spine ; led / spdl: lámina espinodiapofisial/ spinodiapophyseal lamina ; lepr / sprl: lámina espinoprezigapofisial/ spinoprezygapophyseal lamina ; lprd / prdl: lámina prezigadiapofisial/ prezygadiapophyseal lamina ; lpre / prsl: lámina prespinal/ prespinal lamina ; lprp / prpl: lámina prezigaparapofisial / prezygaparapophyseal lamina ; pp: parapófisis/ parapophysis ; prz: prezigapófisis/ prezygapophysis . Escala gráfica: 10 cm/ Scale bar: 10 cm.
Por un lado, este arco cervico-dorsal (figuras 5.AB) comparte con las vértebras cervicales la forma de los procesos transversos, con amplias láminas prezigodiapofisial y centrodiapofisial posterior, y la posición relativamente baja de la diapófisis, ubicadas a nivel de la base del arco neural. Además, su lámina espinoprezigapofisial constituye una estructura robusta, no tan extendida dorsalmente como en las cervicales, pero más larga y desarrollada que en la tercera dorsal anterior (figura 5.C).
Por otro lado, este arco neural comparte con las vértebras dorsales la presencia de una lámina espinodiapofisial, característica en las vértebras dorsales de los saurópodos (Wilson, 1999). No obstante, en este arco cervico-dorsal la lámina espinodiapofisial es más delgada que la lámina espinoprezigapofisial, al contrario de lo que acontece en la tercera vértebra dorsal (figura 5.C). En esta última, la lámina espinodiapofisial constituye una larga estructura que forma el borde dorsal del proceso transverso. Además, el arco cervico-dorsal, al igual que las vértebras dorsales, presenta una lámina prespinal bien definida.
Finalmente, un carácter que no puede describirse con precisión en los restos preservados, es la forma en que las anchas espinas neurales de las vértebras cervicales reducen paulatinamente su desarrollo transversal hacia la serie dorsal. No obstante, las evidencias disponibles permiten suponer que las anchas espinas neurales de la serie cervical de Mendozasaurus alcanzan su máximo desarrollo en la sección media-posterior de la serie cervical (figura 2), desde donde reducen sensiblemente su diámetro transversal hacia la sección cervico-dorsal (figuras 5.A-B). Finalmente, la tercera dorsal (figura 5.C) muestra una espina neural relativamente angosta, aunque debe tenerse en cuenta el deficiente estado de preservación de sus bordes laterales, que sugiere un mayor desarrollo transversal que el preservado. Es importante resaltar que en los saurópodos es frecuente observar un cambio morfológico notable en la transición cervico-dorsal que involucra la forma de la espina neural, de las costillas, del arco y de los centros vertebrales (McIntosh, 1990; Bonaparte, 1999; Wedel et al ., 2000).
Comparaciones
Los restos cervicales de los titanosaurios Malawisaurus dixeyi (Jacobs et al ., 1993) del Cretácico Inferior de África, Isisaurus colberti (Jain y Bandyopadhyay, 1997; Wilson y Upchurch, 2003) del Maastrichtiano de la India, Rapetosaurus krausei (Curry Rogers y Forster, 2001; 2004) del Maastrichtiano de Madagascar, Alamosaurus sanjuanensis (Lehman y Coulson, 2002) del Maastrichtiano de América del Norte, Titanosauridae indet. DGM Serie A del Cretácico Tardío de Brasil (Powell, 1987; 2003), Neuquensaurus australis (Salgado et al ., en prensa) del Campaniano temprano de Argentina, Saltasaurus loricatus (Bonaparte y Powell, 1980; Powell 1992) del Campaniano-Maastrictiano de Argentina, y Rinconsaurus caudamirus (Calvo y González Riga, 2003) y Mendozasaurus neguyelap (González Riga, 2003a), ambos del Turoniano tardío-Coniaciano tardío de Argentina, muestran una notable diversidad morfológica.
Un primer análisis comparativo (figura 6; tabla 2) permite reconocer grandes diferencias de tamaño entre las vértebras cervicales de los titanosaurios, siendo las de Isisaurus colberti y Mendozasaurus neguyelap las de mayor tamaño. Así mismo, se observa una notable variación en la forma y altura de las espinas neurales, en la posición y forma de las zigapófisis, en el desarrollo de láminas y cavidades suprazigapofisiales y en las proporciones de los centros vertebrales.
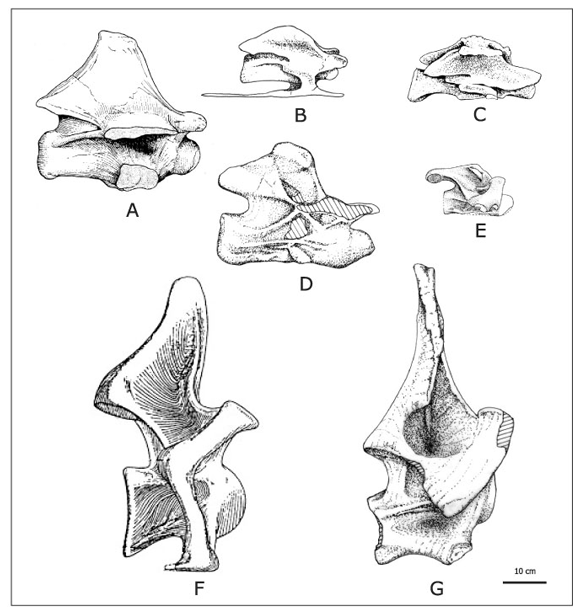
Figura 6. Vértebras cervicales medias y posteriores de titanosaurios en vista lateral/ middle and posterior cervical vertebrae of titanosaurs in lateral views . A, Malawisaurus dixeyi , cervical media-posterior/ mid-posterior cervical (según Jacobs et al ., 1993); B, Titanosauria indet. DGM Serie A de Brasil, cervical posterior / posterior cervical (redibujado de Powell, 1987), C, Alamosaurus sanjuanensis , cervical media/ middle cervical (según Lehman y Coulson, 2002); D, Rinconsaurus caudamirus , cervical posterior/ posterior cervical (redibujado de Calvo y González Riga, 2003), E, Saltasaurus loricatus , cervical media/ middle cervical (según Powell, 1992); F, Isisaurus colberti , cervical media-posterior/ mid-posterior cervical (según Jain y Bandyopadhyay, 1997); G, Mendozasaurus neguyelap , cervical media-posterior/ mid-posterior cervical. Escala gráfica: 10 cm/ Scale bar: 10 cm.
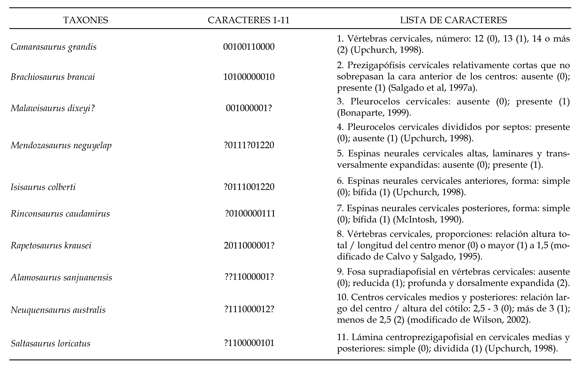
Tabla 2. Caracteres cervicales de Camarasaurus grandis (Cope, 1877), Brachiosaurus brancai (Janensch, 1950) y titanosaurios mencionados en el texto/ Cervical characters of Camarasaurus grandis (Cope, 1877), Brachiosaurus brancai (Janensch, 1950) and titanosaurs mentioned in the text.
Las vértebras cervicales de Mendozasaurus son notablemente diferentes a la del titanosaurio basal Malawisaurus dixeyi (Jacobs et al ., 1993). Esta especie africana muestra centros relativamente largos, con índices de elongación (EI) > 3, y ausencia de pleurocelos o cavidades laterales (figura 6.A). Su espina neural, proporcionalmente más baja que la de Mendozasaurus , exhibe un borde dorsal axialmente elongado. En adición, su arco neural muestra un escaso desarrollo de láminas diapofisiales (Wilson y Sereno, 1998), y carece de lámina prezigodiapofisial y de cavidad supradiapofisial. Evidentemente, Malawisaurus exhibe una morfología distinta a la de Mendozasaurus y otros titanosaurios más derivados (Calvo, 1999; Bonaparte et al ., 2000; González Riga, 2002).
Por otro lado, las cervicales de Mendozasaurus son claramente distintas a las de Rapetosaurus krausei (Curry Rogers y Forster, 2001), a la del Titanosauria indet. Serie A de Brasil (Powell, 1987) y a la de Alamosaurus sanjuanensis (Lehman y Coulson, 2002) (figura 6, B-C). En contraste con Mendozasaurus , estos titanosaurios exhiben centros cervicales elongados (IE > 3), arcos neurales relativamente bajos, ausencia de amplias cavidades supradiapofisiales y espinas neurales dorsalmente reducidas.
Rinconsaurus caudamirus (Calvo y González Riga, 2003) también muestra cervicales muy diferentes a las de Mendozasaurus , con centros relativamente largos (IE > 3), espinas neurales bajas y fosas supradiapiofisiales profundas pero reducidas (figura 6.D). En particular Rinconsaurus se destaca por la presencia de una lámina centroprezigapofisial dividida que delimita una elongada depresión debajo de la prezigapófisis. Un carácter parcialmente similar ha sido descrito en dorsales anteriores de Saltasaurus loricatus (Powell, 1992; 2003) y en Diplodocidae (Wilson, 2002).
Las cervicales de Saltasaurus loricatus (figura 6.E) (Bonaparte y Powell, 1980; Powell 1992) son también notablemente distintas a las de Mendozasaurus . Por un lado, los centros vertebrales, si bien no tan elongados como en otros titanosaurios (IE = 2,5-3), son proporcionalmente más largos que los de Mendozasaurus . Además los centros de Saltasaurus muestran pleurocelos pequeños divididos por láminas, lo que es considerado una reversión autapomórfica por Wilson (2002). Por otro lado, las espinas neurales de Saltasaurus están tan reducidas dorsalmente que apenas sobrepasan la altura de las postzigapófisis (Powell, 1992; Bonaparte, 1999). Además, son transversalmente anchas y muestran un perfil lateral asimétrico, con un borde anterior fuertemente inclinado hacia las prezigapófisis. En adición, la espina neural se conecta con robustas láminas espinopostzigapofisiales que se inclinan suavemente hacia posterior (González Riga, 2002). En contraste con Saltasaurus , el Saltasaurinae Neuquensaurus australis (Huene, 1929; Powell, 1986) muestra espinas neurales relativamente más altas. Si bien los restos cervicales de Neuquensaurus son fragmentarios, nuevos materiales procedentes de Cinco Saltos (Formación Anacleto, provincia de Río Negro) (Salgado et al ., en prensa), amplían el conocimiento sistemático de este género. Tal como fue propuesto por Salgado et al ., (1997a), Neuquensaurus y Saltasaurus comparten la presencia de prezigagapófisis relativamente cortas que no sobrepasan el borde anterior de los centros (y su correlato, postzigapófisis proyectadas posteriormente), carácter sinapomórfico de Saltasaurinae. Las cervicales posteriores de Neuquensaurus exhiben una espina neural más elevada que la de Saltasaurus y que termina dorsalmente en una protuberencia hemiesférica, carácter también presente en el titanosaurio de Periópolis DGM Serie A de Brasil (Salgado et al ., en prensa).
En contraste con los titanosaurios analizados, Mendozasaurus neguyelap comparte con Isisaurus colberti (Jain y Bandyopadhyay, 1997) los siguientes caracteres:
1) Centros vertebrales cervicales relativamente cortos, con IE < 2,5. La longitud de las vértebras cervicales varía en los distintos géneros y especies, y también en una misma especie durante su desarrollo ontogenético (Carpenter y McIntosh, 1994). El índice de elongación permite reconocer titanosaurios con: a) centros muy cortos (EI < 2,5) como los de Mendozasaurus neguyelap e Isisaurus colberti , b) centros cortos (EI = 2,5-3) como los de Saltasaurus , c) centros largos (EI > 3) como los de Alamosaurus , y d) centros muy largos (EI > 4) como los de Rapetosaurus .
En los saurópodos la longitud de los centros cervicales está relacionada con la longitud del cuello y su posición de vida (Wedel et al ., 2000). La variación ontogenética puede observarse en diferentes materiales asignados a Saltasaurus . Por ejemplo, en especimenes adultos los centros cervicales medios y posteriores (Powell, 2003, láminas 48 y 49; PVL 4017-30 y 4017-139) son relativamente largos (EI > 2,5); mientras que ejemplares juveniles (PVL 4017-4) exhiben centros proporcionalmente cortos (EI < 2). Es oportuno mencionar que algunos restos de Neuquensaurus australis , como los descriptos por Salgado et al ., (en prensa), comprenden centros cervicales posteriores relativamente cortos (EI < 2,5). Evidentemente, el acortamiento de los centros cervicales se registra tanto en titanosaurios de gran porte con altas espinas neurales y amplias fosas supradiapofisiales ( Mendozasaurus , Isisaurus ) como en titanosaurios derivados de tamaño medio ( Saltasaurus , Neuquensaurus ) agrupados en el clado Saltasaurinae.
En la mayoría de los titanosaurios se desconoce el número de vértebras que integran la serie cervical. No obstante, si se asume que Mendozausaus tenía 16 vértebras cervicales, tal como posee Rapetosaurus krausei (Curry Rogers y Foster, 2001), es de suponer que exhibía un cuello relativamente corto que no habría superado los 6 m de longitud (figura 7). En contraste el cuello de Mendozasaurus habría sido relativamente grueso, con un diámetro dorsoventral en su sección media de unos 120 cm, a juzgar por el tamaño de sus vértebras cervicales que tienen 71 cm de altura y 80 cm de ancho.

Figura 7. Mendozasaurus neguyelap . A, reconstrucción esqueletal con indicación de los huesos preservados/ skeletal reconstruction showing preserved bones ; B, reconstrucción en vida realizada por el autor. Observaciones: La reconstrucción de la cabeza y el número de cervicales se basa en restos bien preservados de Rapetosaurus krausei (Curry Rogers y Foster, 2001; 2004). La reconstrucción en vida de Mendozasaurus se fundamenta en ejemplares adultos de tamaño medio (largo del fémur: 150 cm). Sin embargo es importante destacar que fueron hallados huesos que pertenecen a un espécimen adulto un 30 % más grande que el ejemplar dibujado (largo estimado del fémur: 190 cm)/ Life restoration carried out by the author. Observations: the restoration of the head and the number of cervicals are based on well-preserved remains of Rapetosaurus krausei (Curry Rogers and Foster, 2001; 2004). The life restoration of Mendozasaurus is supported on mid-sized adults (femur length: 150 cm); however, it is important to emphasize that an adult 30 percent larger than the specimen illustrated has been found (estimated femur length: 190 cm). Escala gráfica: 1 m/ Scale bar: 1 m.
2) Fosa supradiapofisial dorsoventralmente extendida, limitada por las láminas espinoprezigapofisial, espinopostzigapofisial, prezigadiapofisial y postzigadiapofisial. En Euhelopus zdanskyi (Wiman, 1929) y la mayoría de los titanosaurios se observa una importante reducción de las láminas del arco cervical (Wilson y Sereno, 1998; Wilson, 2002). Esta simplificación de la estructura laminar puede describirse con mayor precisión si se tiene en cuenta la presencia o ausencia de algunos caracteres del arco cervical, tal como la fosa supradiapofisial (tabla 2), presente en Apatosaurus louisae y otros saurópodos (Gilmore, 1936; Wilson, 1999). En la mayoría de los titanosaurios esta fosa está ausente ( Malawisaurus , Alamosaurus ), es reducida ( Rinconsaurus ) o aparece como una cavidad lateral sin bordes definidos ( Saltasaurus ). En contraste, Mendozasaurus e Isisaurus muestran una fosa supradiapofisial con amplio desarrollo dorsoventral, limitada por las láminas espinoprezigapofisial, espinopostzigapofisial, prezigadiapofisial y postzigadiapofisial. En particular, Mendozasaurus muestra una cavidad supradiapofisial tan profunda que casi llega hasta el plano axial. A diferencia de lo que acontece en Isisaurus colberti , esta fosa está parcialmente limitada hacia dorsal por la expansión lateral de la espina neural (figura 6.G).
3) ) Vertebras cervicales con altas espinas neurales. En la mayoría de los Titanosauriformes Salgado et al ., (1997a) las vértebras cervicales exhiben espinas neurales relativamente bajas y centros axialmente elongados. En contraste, Mendozasaurus neguyelap e Isisaurus colberti muestran espinas neurales relativamente altas, donde la relación: altura total de la vértebra/longitud del centro es mayor a 1,5.
Discusión y conclusiones
Si bien las vértebras cervicales de Mendozasaurus neguyelap exhiben similitudes con las de Isisaurus colberti , muestran algunas diferencias significativas relacionadas con la forma de los procesos transversos y la espina neural. En particular se destaca en Mendozasaurus un carácter novedoso en las cervicales medias-posteriores que constituye una probable autapomorfía: la presencia de espinas neurales laminares y expandidas transversalmente, más anchas que los centros y de contorno anterior en abanico o subromboidal por el desarrollo de expansiones laterales y un extremo dorsal sub-redondeado. Estas expansiones de la espina neural se proyectan lateralmente con respecto a la lámina espinopostzigapofisial y se bifurcan ventralmente en dos láminas, una que termina dentro de la cavidad supradiapofisial y otra que delimita el borde posterior de la cavidad supradiapofisial (figura 2.B). En contraste, Isisaurus exhibe espinas neurales de contorno posterior subtriangular o subtrapezoidal (Jain y Bandyopadhyay, 1997, figs. 3-5). Además, como carece de expansiones laterales, el borde lateral de la espina neural está definido por las láminas espinopostzigapofisiales (figura 6.F).
Las espinas cervicales de Mendozasaurus también son diferentes a las de un nuevo titanosaurio basal (MCF-PVPH-233) procedente de la Formación Lohan Cura, Aptiano-Albiano de la provincia de Neuquén (Bonaparte, González Riga y Apesteguía, en estudio). En este taxón las vértebras cervicales posteriores poseen espinas neurales levemente más anchas que los centros y de contorno romboidal. No obstante, en este titanosaurio de Neuquén la espina neural es relativamente más baja y muestra un borde dorsal con un ángulo más definido. Además, se observan otras diferencias: en Mendozasaurus el contorno subromboidal está dado por proyecciones laterales de la espina neural. En cambio, en este nuevo taxón, el borde lateral de la espina neural está formado por proyecciones de las láminas espinoprezigapofisiales laterales, láminas que están ausentes en Mendozasaurus .
El registro fósil de vértebras cervicales de titanosaurios, tanto en América del Sur como en otras partes del mundo, muestra una notable diversidad morfológica que involucra la forma y altura de las espinas neurales, la posición de las zigapófisis, el desarrollo de láminas y cavidades diapofisiales y las proporciones de los centros vertebrales. En particular, Mendozasaurus neguyelap se encuentra entre los titanosaurios que poseen los centros cervicales proporcionalmente más cortos asociados con altas espinas neurales, lo que evidencia la presencia de saurópodos con cuellos relativamente anchos, robustos y cortos en el Cretácico Tardío de Argentina. Es probable que la diversa morfología cervical de los titanosaurios esté relacionada con distintas adaptaciones ecológicas. No obstante se requieren detallados estudios paleobiomecánicos y paleoecológicos para establecer las relaciones existentes entre la estructura ósea de estos grandes vertebrados y sus particulares adaptaciones morfofuncionales.
Agradecimientos
Deseo expresar mi agradecimiento a J. Bonaparte y W. Volkheimer por sus orientaciones como directores de beca postdoctoral del Consejo Nacional de Investigaciones Científicas y Técnicas; a J. Calvo por su permanente apoyo y asesoramiento en este trabajo; a L. Salgado por la revisión del manuscrito y sus oportunas observaciones; a K. Curry Rogers (Science Museum of Minnesota, EE.UU), por la revisión del manuscrito y por brindar generosamente información inédita sobre titanosaurios; a S. Apesteguía por sus comentarios; a S. Londero, por su inestimable apoyo en el montaje del laboratorio; a M. Bourguet, E. Previtera, C. Sancho y C. Pirrone por su importante colaboración en los trabajos de campo y laboratorio durante los años 2001-2003. Este estudio ha sido apoyado por el IANIGLA-CRICYT y financiado por la Agencia Nacional de Promoción Científica y Tecnológica (PICT 07-08277; director J. Calvo).
Bibliografía
Apesteguía, S. 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. Naturwissenschaften 91: 493-497. [ Links ]
Behrensmeyer, A.K. y Hook, R.W. 1992. Paleoenvironmental contexts and taphonomic modes. En: A.K. Behrensmeyer, J.D. Damuth, W.A DiMichele, R. Potts, H.D. Sues y S.L. Wing (eds.), Terrestrial ecosystems through time: evolutionary paleoecology of terrestrial plants and animals , University of Chicago Press, Chicago, pp. 15-136. [ Links ]
Bonaparte, J.F. 1999. Evolución de las vértebras presacras en Sauropodomorpha. Ameghiniana 36: 115-187. [ Links ]
Bonaparte, J.F. y Coria, R.A. 1993. Un nuevo y gigantesco saurópodo Titanosaurio de la Formación Río Limay (Albiano-Cenomaniano) de la provincia del Neuquén, Argentina. Ameghiniana 30: 271-282. [ Links ]
Bonaparte, J.F. y Powell, J.E. 1980. A continental assemblage of tetrapods from the Upper Cretaceous beds of El Brete, northwestern Argentina (Sauropoda, Coelurosauria, Carnosauria, Aves). Mémoires de la Société Geólogique de France 139: 19-28. [ Links ]
Bonaparte, J.F., Heinrich, W.D. y Wild, R. 2000. Review of Janenschia Wild, with the description of a new sauropod from the Tendaguru beds of Tanzania and a discussion on the systematic value of procoelous caudal vertebrae in the Sauropoda. Palaeontographica 256: 25-76. [ Links ]
Calvo, J.O. 1999. Dinosaurs and other vertebrates of the Lake Ezequiel Ramos Mexía area, Nequén-Patagonia, Argentina. En: Y. Tomida, T.H. Rich, y P. Vickers Rich (eds.), Proceedings of the Second Gondwanan Dinosaur Symposium, National Science Museum Monographs 15: 13-45. [ Links ]
Calvo, J.O. y Bonaparte, J.F. 1991. Andesaurus delgadoi n.g.n. sp. (Saurischia, Sauropoda) dinosaurio Titanosauridae de la Formación Río Limay (Albiano - Cenomaniano), Neuquén, Argentina. Ameghiniana 28: 303-310. [ Links ]
Calvo, J.O. y González Riga, B.J. 2003. Rinconsaurus caudamirus gen. et sp. nov., a new titanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia, Argentina. Revista Geológica de Chile 30: 333-353. [ Links ]
Calvo, J.O. y Salgado, L. 1995. Rebbachisaurus tessonei sp. nov., a new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. GAIA 11: 13-33. [ Links ]
Calvo, J.O., Coria, R.A. y Salgado, L. 1997. Uno de los más completos titanosáuridos (Dinosauria-Sauropoda) registrados en el mundo. Ameghiniana 34: 534. [ Links ]
Carpenter, K y McIntosh, J. 1994. Upper Jurassic sauropod babies from the Morrisson Formation. En: K. Carpenter, K. Hirsch y J. Horner (eds.), Dinosaur Eggs and Babies , Cambridge University Press, Cambridge, pp. 165-278. [ Links ]
Cope, E.D. 1877. On a gigantic saurian from the Dakota epoch of Colorado. Paleontological Bulletin (Philadelphia) 25: 5-10. [ Links ]
Curry Rogers, K. y Foster, C.A. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature 412: 530-534. [ Links ]
Curry Rogers, K. y Foster, C.A. 2004. The skull of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 24: 121-144. [ Links ]
Gilmore, C.W. 1936. Osteology of Apatosaurus with special reference to specimens in the Carnegie Museum. Memoirs of the Carnegie Museum 11: 175-300. [ Links ]
González Riga, B.J. 2002. [ Estratigrafía y dinosaurios del Cretácico Tardío en el extremo sur de la provincia de Mendoza, R. Argentina. Tesis Doctoral, Universidad Nacional de Córdoba, Argentina, 280 pp., Inédita]. [ Links ]
González Riga, B.J. 2003a. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Amehginiana 40: 155-172. [ Links ]
González Riga, B.J. 2003b. Bioestratinomía y fosildiagénesis de un sitio paleontológico con saurópodos Titanosauridae del Cretácico Tardío de Mendoza, Argentina. Ameghinana 40, Suplemento Resúmenes : 58R. [ Links ]
González Riga, B.J. y Calvo, J.O. 2001. A new genus and species of Titanosaurid Sauropod from the Upper Cretaceous of Rincón de los Sauces, Neuquén, Argentina. Journal of Vertebrate Paleontology, Abstracts 21: 55A. [ Links ]
González Riga, B.J., Calvo, J.O., Astini, R.A., y Parras, A.M. 2003. Preservación tafonómica de titanosaurios (Dinosauria, Sauropoda) en facies fluviales del sector septentrional de la Cuenca Neuquina. Ameghiniana 40, Suplemento Resúmenes : 102R. [ Links ]
González Riga, B.J. Calvo, J.O., Londero, S., Piccolella, F., Bourguet, M., Previtera, M.E., Pirrone, C. E. y Sancho, C. 2004. Hallazgo de dinosaurios saurópodos en la región de Cañadón Amarillo (Mendoza, Argentina) y su relación con los ambientes fluviales. Ameghiniana 41 , Suplemento Resúmenes : 49R. [ Links ]
Hatcher, J.B. 1903. Osteology of Haplocanthosaurus , with description of a new species. Memoirs of the Carnegie Museum 2: 1-72. [ Links ]
Huene, F. 1929. Los Saurisquios y Ornitisquios del Cretácico Argentino. Anales del Museo de La Plata (Serie 2) 3: 1-196. [ Links ]
Jacobs, L.L, Winkler, D.A., Downs W.R.y Gomani, E.M. 1993. New material of an early Cretaceous titanosaurid sauropod dinosaur from Malawi. Palaeontology 36: 523-534. [ Links ]
Jain, S.L. y Bandyopadhyay, S. 1997. New titanosaurid (Dinosauria: Sauropoda) from the Late Cretaceous of Central India. Journal of Vertebrate Paleontology 17: 114-136. [ Links ]
Janensch, W. 1929. Material und Formengehalt der Sauropoden in der Ausbeute der Tendaguru- expedition. Palaeontographica , Suppl . 7: 1-34. [ Links ]
Janensch, W. 1950. Die Wirbelsäule von Brachiosaurus brancai . Palaeontographica 7: 27-93. [ Links ]
Kellner, A.W.A. y Acevedo, S.A. K. 1999. A new sauropod dinosaur (Titanosauria) from the Late Cretaceous of Brazil. En: Y. Tomida, T.H. Rich, y P. Vickers Rich (eds.), Proceedings of the Second Gondwanan Dinosaur Symposium, National Science Museum Monographs 15: 111-142. [ Links ]
Leanza, H.A. y Hugo, C.A. 2001. Cretaceous red beds from southern Neuquén Basin (Argentina): age, distribution and stratigraphic discontinuities. Asociación Paleontológica Argentina, Publicación Especial 7: 117-122. [ Links ]
Lehman, T.M. y Coulson, A.B. 2002. A juvenile specimen of the sauropod dinosaur Alamosaurus sanjuanensis from the Upper Cretaceous of Big Bend National Park, Texas. Journal of Paleontology 76: 156-172. [ Links ]
Lydekker, R. 1893. Contributions to the study of the fossil vertebrates of Argentina. I, The dinosaurs of Patagonia. Anales del Museo de La Plata, Paleontología 2: 1-14. [ Links ]
Marsh, O.C. 1878. Principal characters of American Jurassic dinosaurs. American Journal of Science , Serie 3, 16: 411-416. [ Links ]
Martínez, R.D., Jiménez, O., Rodríguez, J. Luna, M. y Lamanna, M.C. 2004. An articulated specimen of the basal Titanosaurian (Dinosauria: Sauropoda) Epachthosaurus sciuttoi from the Early Late Cretaceous Bajo Barreal Formation of Chubut Province, Argentina. Journal of Vertebrate Paleontology 24: 107-120. [ Links ]
McIntosh, J.S. 1990. Sauropoda. En: D. Weishampel, P. Dobson and H. Osmolska (eds.), The Dinosauria , University of California Press, Berkeley, pp. 345-401. [ Links ]
Powell, J.E. 1986. [ Revisión de los Titanosáuridos de América del Sur. Tesis Doctoral, Universidad Nacional de Tucumán, Argentina, 340 pp. Inédita]. [ Links ]
Powell, J.E. 1987. Morfología del esqueleto axial de los dinosaurios titanosáuridos (Saurischia, Sauropoda) del estado de Minas Gerais, Brasil. Anais do 10º Congresso Brasileiro de Paleontología (Río de Janeiro): 151-171. [ Links ]
Powell, J.E. 1992. Osteología de Saltasaurus loricatus (Sauropoda-Titanosauridae) del Cretácico Superior del noroeste argentino. En: J.L. Sanz y A.D. Buscalioni (eds.), Los Dinosaurios y su entorno biótico , Instituto Juan de Valdés, Cuenca, pp. 165-230. [ Links ]
Powell, J.E. 2003. Revision of South American Titanosaurid dinosaurs: palaeobiological, palaeobiogeographical and phylogenetic aspects. Records of the Queen Victoria Museum, Launceston , 173 pp. [ Links ]
Salgado, L. 1996. Pellegrinisaurus powelli nov. gen. et sp. (Sauropoda, Titanosauridae) from the Upper Cretaceous of Lago Pellegrini, northwestern Patagonia, Argentina. Ameghiniana 33: 355-365. [ Links ]
Salgado, L. 2003. Should we abandon the name Titanosauridae? Some comments on the taxonomy of titanosaurian sauropod (Dinosauria). Revista Española de Paleontología 18: 15-21. [ Links ]
Salgado, L. y Azpilicueta, C. 2000. Un nuevo saltasaurino (Sauropoda, Titanosauridae) de la provincia de Río Negro (Formación Allen, Cretácico Superior), Patagonia, Argentina. Ameghiniana 37: 259-264. [ Links ]
Salgado, L. y Coria, R.A. 1993. El género Aeolosaurus (Sauropoda-Titanosauridae) en la Formación Allen (Campaniano-Maastrichtiano) de la Provincia de Río Negro, Argentina. Ameghiniana 30: 119-128. [ Links ]
Salgado, L., Coria, R.A. y Calvo, J.O. 1997a. Evolution of Titanosaurid Sauropods. I: Phylogenetic analysis based on the postcraneal evidence. Ameghiniana 34: 3-32. [ Links ]
Salgado, L., Coria, R.A. y Calvo, J.O. 1997b. Presencia del género Aelosaurus (Sauropoda, Titanosauridae) en la Formación Los Alamitos, Cretácico Superior de la Provincia de Río Negro, Argentina. Geociencias 2: 44-49. [ Links ]
Salgado, L., Apesteguía, S. y Heredia, S. (en prensa). A new specimen of Neuquensaurus australis , a Late Cretaceous saltasaurine titanosaur from north Patagonia. Journal of Vertebrate Paleontology . [ Links ]
Seeley, H.G. 1887. On the classification of the fossil animals commonly called Dinosauria. Royal Society of London 43: 165-171. [ Links ]
Sereno, P.C., Beck, A.L., Dutheil, D.B., Larsson, H., Lyon, G.H., Moussa, B., Sadleir, R.W., Sidor, C.A., Varricchio, D.J., Wilson. G.P. y Wilson. J.A. 1999. Cretaceous Sauropods from the Sahara ad the Uneven Rate of Skeletal Evolution Among Dinosaurs. Science 286: 1342-1347. [ Links ]
Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43-103. [ Links ]
Wedel, M.J., Cifelli, R.L. y Kent Sanders, R. 2000. Osteology, paleobiology and relationships of the sauropod dinosaur Sauroposeidon. Acta Palaeontologica Polonica 45: 343-388. [ Links ]
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639-653. [ Links ]
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 217-276. [ Links ]
Wilson, J.A. y Sereno, P. 1998. Early Evolution and Higher-level Phylogeny of Sauropod Dinosaurs. Journal of Vertebrate Paleontology 18, Supplement 2: 1-68. [ Links ]
Wilson, J.A. y Upchurch, P. 2003. A revision of Titanosaurus Lydekker (Dinosauria-Sauropoda), the first dinosaur genus with a Gondwanan' distribution. Journal of Systematic Palaeontology 1: 125-160. [ Links ]
Wiman, C. 1929. Die Kreide-Dinosaurier aus Shantung. Palaeontologia Sinica (Series C) 6: 1-67. [ Links ]
Recibido: 10 de febrero de 2004.
Aceptado: 11 de noviembre de 2004.

