










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.45 n.3 Buenos Aires jul./sep. 2008
Nuevos aportes para la sistemática de los Glyptodontia Ameghino 1889 (Mammalia, Xenarthra, Cingulata)
Juan Carlos Fernicola1
1Sección Paleontología de Vertebrados, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". Av. Angel Gallardo 470, 1405 Buenos Aires, Argentina. CONICET jctano@macn.gov.ar
Abstract. New contributions to the systematic of Glyptodontia Ameghino 1889 (Mammalia, Xenarthra, Cingulata). An exhaustive revision of the systematic of the glyptodonts, including different classifications and phylogenetic hypothesis proprosed since the XIX century is presented herein. According to the current classification, proposed as preliminary, a cladistic analysis based on 84 craniodental characters was carried out for the following taxa: Glyptodontinae [Glyptodon], Doedicurinae [Doedicurus] Propalaehoplophorinae [Propalaehoplophorus and Eucinepeltus] and Hoplophorinae (=Sclerocalyptinae) Hoplophorini (=Sclerocalyptini) [Neosclerocalyptus (=Sclerocalyptus), Eosclerocalyptus and Hoplophractus] Plohophorini [Stromaphorus, Pseudoplohophorus and Plohophorus] Panochthini [Panochthus] and Lomaphorini [Urotherium]. The results of this analysis support the monophyly of the glyptodonts as well as a basal dichotomy represented by the Propalaehoplophorinae as the sister group of the remaining glyptodonts. The relationships between the taxa of this group do not support the previous monophyletic hypotheses of the Hoplophorinae (= Sclerocalyptinae) neither of their tribes Hoplophorini (= Sclerocalyptini) and Plohophorini. On the contrary, different groups not formally proposed until today are clearly supported as the derived clades (Glyptodon + Doedicurus) and (Panochthus + Neosclerocalyptus). These results, together with the preliminary character of the current classification, allow to propose a new classification for the glyptodonts. On the other hand, the combination of the phylogenetic and the stratigraphic information allows locating the first dichotomy of the glyptodonts in Colhuehuapian times (c.a. 20 Ma.). The second dichotomy, assignable to Chasicoan/Huayquerian times (c.a. 10 Ma.), allows to recognize a gap in the paleontological record of at least 10 Ma. Lastly, the inclusion of the genus Hoplophractus within Eosclerocalyptus is questioned.
Resumen. En este trabajo se presenta una exhaustiva revisión de la sistemática de los gliptodontes, incluyendo las distintas clasificaciones e hipótesis filogenéticas propuestas desde el siglo XIX. De acuerdo a la clasificación vigente, formulada con carácter preliminar, se realizó un análisis cladístico, basado en 84 caracteres craneomandibulares, de los siguientes taxones: Glyptodontinae [Glyptodon], Doedicurinae [Doedicurus], Propalaehoplophorinae [Propalaehoplophorus y Eucinepeltus] y Hoplophorinae (=Sclerocalyptinae), Hoplophorini (=Sclerocalyptini) [Neosclerocalyptus (=Sclerocalyptus), Eosclerocalyptus y Hoplophractus] Plohophorini [Stromaphorus, Pseudoplohophorus y Plohophorus] Panochthini [Panochthus] y Lomaphorini [Urotherium]. Los resultados obtenidos sustentan la monofilia de los gliptodontes y una dicotomia basal representada por los Propalaehoplophorinae como el grupo hermano de los restantes gliptodontes. Las relaciones entre los taxones de este último grupo no avalan las hipótesis monofiléticas previas de los Hoplophorinae (=Sclerocalyptinae) ni de sus tribus Hoplophorini (=Sclerocalyptini) y Plohophorini. Al contrario, distintos agrupamientos hasta hoy no postulados se encuentran claramente soportados como los clados más derivado (Glyptodon + Doedicurus) y (Panochthus + Neosclerocalyptus). Estos resultados, juntamente con el carácter preliminar de la clasificación vigente permitieron proponer una nueva clasificación para los gliptodontes. Por otra parte, la mixtura de la información filogenética con el registro estratigráfico permitió ubicar la primera dicotomía del grupo en tiempos colhuehuapenses (c.a. 20 Ma.), mientras que la segunda dicotomía asignable a tiempos Chasiquense/Huayqueriense (c.a. 10 Ma.) permitió reconocer una prolongada ausencia en el registro paleontológico de al menos 10 Ma. Por último, se cuestiona la inclusión del género Hoplophractus dentro de Eosclerocalyptus.
Key words. Glyptodontia; Systematic; Phylogeny; Classification.
Palabras clave. Glyptodontia; Sistemática; Filogenia; Clasificación.
Introducción
Los gliptodontes, ampliamente registrados en yacimientos cenozoicos americanos, fueron históricamente agrupados en los Cingulata, conjuntamente con pampaterios y armadillos, de los que se los ha diferenciado por presentar un caparazón dorsal esencialmente inmóvil, diferente número de dientes trilobulados, miembros elefantinos y una aparato masticatorio telescopado por debajo del cráneo (Hoffstetter, 1958; Fariña y Vizcaíno, 2001). El taxón más antiguo conocido del grupo, Glyptatelus Ameghino, proviene del Eoceno medio de Patagonia, Argentina (Scillato-Yané, 1976) mientras que el último registro se ubica en el entorno de los últimos 10.000 años. Desde sus inicios, la clasificación y filogenia de los gliptodontes ha generado distintas controversias. Desde una perspectiva histórica, fue Ameghino (1889) quien, basado en dos hipótesis morfoevolutivas que daban cuenta de las características óseas de la cola y la coraza dorsal, propuso el primer esquema filogenético de los gliptodontes. Sobre las mismas características, aunque valorizadas en forma desigual, más algunas características del endoesqueleto, Castellanos (1931, 1932, 1959) y Hoffstetter (1958) modificaron de diferentes formas la clasificación de los gliptodontes y plantearon distintas hipótesis genealógicas entre los mismos. Sin embargo, fue la propuesta sistemática preliminar de Hoffstetter (1958) la que tuvo un amplio consenso en la comunidad científica y como tal es la actualmente vigente. En la misma, Hoffstetter (1958) dividió a los gliptodontes en las subfamilias Glyptatelinae, Propalaehoplophorinae, Doedicurinae, Glyptodontinae y Hoplophorinae y esta última en siete tribus. Desde una perspectiva cladística, los gliptodontes sólo fueron incluidos como un grupo terminal en el estudio de las relaciones de los cingulados (Engelmann, 1985; Gaudin y Wible, 2006). En este trabajo se presenta un análisis filogenético de los gliptodontes basado en 84 caracteres craneomandibulares con el fin de poner a prueba la monofilia del grupo así como la de distintos agrupamientos supragenéricos propuestos por Hoffstetter (1958). Al mismo tiempo, se presenta una exhaustiva síntesis de las propuestas filogenéticas y clasificatorias de los autores antes mencionados con el fin de sustentar las modificaciones propuestas en la clasificación de los gliptodontes.
Relaciones entre los gliptodontes y los restantes cingulados
Hasta fines del siglo XX, los gliptodontes fueron considerados ancestros de los armadillos actuales (v. gr. Flower, 1882), relación que, como sustuvo Ameghino (1884a, 1889), se habría originado en la mayor antigüedad que presentaban, hasta ese entonces, los gliptodontes con respecto a los armadillos. Fue Ameghino (1884a, 1889) quien planteó una relación genealógica inversa, basándose en la idea de que lo estructuralmente simple antecede a lo complejo y que la fusión de los elementos óseos es una característica derivada. Así, los gliptodontes fueron considerados derivados por presentar una coraza sin bandas móviles completas, columna vertebral con distintos grado de fusión y molariformes mayoritariamente trilobulados. Contrariamente, los armadillos fueron considerados ancestrales por presentar un número variable de bandas móviles en la coraza, columna vertebral no fusionada y una morfología dentaria en la que predomina la forma elíptica. Esta hipótesis filogenética fue posteriormente avalada por varios autores que abordaron distintos aspectos filogenéticos de los cingulados (Castellanos, 1932, 1959; Hoffstetter, 1958; Engelmann, 1985; Gaudin y Wible, 2006). Tres taxones fueron estrechamente relacionados a los gliptodontes. Ameghino (1889, 1898), Castellanos (1937) y Patterson y Pascual (1972) reconocieron a los pampaterios como los armadillos ancestrales de los gliptodontes, mientras que Engelmann (1985) los consideró como el grupo hermano del clado conformado por gliptodontes y armadillos eutatinos (Eutatus y Proeutatus). Esta última hipótesis no encuentra sustento en el estudio filogenético de Gaudin y Wible (2006), quienes encontraron un fuerte apoyo al clado conformado por pampaterios y gliptodontes.
El tercer grupo de cingulados reconocidos como intermediarios entre armadillos y gliptodontes fueron los Palaeopeltidae, grupo cuya historia taxonómica es profusa y compleja (véase Patterson y Pascual, 1972 y Fernicola et al., 2008). Ameghino (1895) los consideró como gliptodontes basales, mientras que Castellanos (1959) los posicionó como el grupo hermano de sus Doedicurinae (véase adelante). Patterson y Pascual (1972) lo reconocieron como un grupo independiente e intermedio entre armadillos y gliptodontes, aunque Hoffstetter (1958), basado en la atelodoncia del grupo, reconoció que podrían representar un grupo divergente de peltefilinos.
Marco histórico sistemático
En 1889, Florentino Ameghino presentó la primera síntesis sistemática de su orden Glyptodontia, conformado por tres familias: Glyptodontidae [Glyptodon Owen, Neothoracophorus Ameghino y Cochlops Ameghino], Hoplophoridae [Propalaehoplophorus Ameghi-no, Asterostemma Ameghino, Palaehoplophorus Ame-ghino, Zaphilus Ameghino, Hoplophorus Lund [= Scle-rocalyptus sensu Ameghino, 1891a] Lomaphorus Ameghino, Plohophorus Ameghino, Nopachthus Ameghino, Panochthus Burmeister, Protoglyptodon Ameghino y Eleutherocercus Koken] y Doedicuridae [Neuryurus Ameghino, Comaphorus Ameghino, Doedicurus Burmeister, Plaxaplous Ameghino y Pseudoeuryurus Ameghino]. Basándose en la morfología exoesquelética, Ameghino (1889) estableció una dicotomía basal entre los gliptodontes sustentada en los morfotipos caudales largo y corto (figura 1). Esta última era: "... corta, cónica, gruesa en la base, y de punta roma, formada por una sucesión de anillos movibles en toda su longitud, es la forma más evolucionada y de consiguiente la de aparición más reciente..." (Ameghino, 1889: 881). En este tipo, Ameghino (1889) reconoció al género Cochlops como el taxón más antiguo y primitivo, del que se habrían originado dicotómicamente los géneros Glyptodon y Neothoracophorus. Esta hipótesis fue propuesta con reservas por Ameghino (1889) ya que reconoció que en Cochlops, "...la cola, aunque anillada y con tubérculos cónicos-puntiagudos, era probablemente más larga, y de consiguiente más cercana á las del otro tipo...". Los restantes géneros de gliptodontes fueron incluidos por Ameghino (1889) en el segundo tipo de cola que caracterizó como "... larga, formada por anillos movibles planos en la base, es el tipo más primitivo y más antiguo, si bien cada uno de ellos ha sufrido en su evolución modificaciones especiales" (Ameghino, 1889: 881). La forma primitiva de este tipo debía "... presentarse en sus más antiguos antecesores con el tubo igualmente anillado, ó dividido por secciones transversales que se unían de adelante hacia atrás para formar el mencionado tubo; en esta forma debía presentarse la cola del Propalaehoplophorus, que no debía ser diferente de la de los Dasypidae." (Ameghino, 1889: 881). Esta forma habría evolucionado hacia la unión de los anillos más caudales, dando origen a un tubo caudal continuo presente en los restantes miembros del grupo. Así conformado el primer anidamiento, Ameghino (1889) postuló otras dos líneas evolutivas basadas en la morfología externa de las placas del caparazón. La primera daba cuenta de un aumento progresivo del número de hileras y figuras periféricas, por lo que derivó directamente de Propalaehoplophorus al género Palaehoplophorus, taxón en el que comenzaría a esbozarse dicho patrón estructural. De este último, derivó dicotómicamente a Hoplophorus [= Sclerocalyptus sensu Ameghino (1891a)], aunque ésta no posee un mayor número de figuras periféricas que Propalaehoplophorus, y a Plohophorus, género que presenta dos hileras de figuras periféricas. De este último se originarían Nopachthus y Panochthus, taxones que muestra una superficie externa completamente cubierta por figuras pequeñas.

Figura 1. Relaciones filogenéticas de los gliptodontes. [Modificado de Ameghino, 1889] / phylogenetic relationships of glyptodonts. [Modified from Ameghino, 1889].
La segunda tendencia evolutiva se iniciaría en una pérdida gradual de la morfología externas de las placas óseas que culminaría en la ausencia total de figuras externas. Así, Ameghino (1889) relacionó estrecha y secuencialmente los géneros Neuryurus, Plaxaplous y Doedicurus, pudiéndose intercalar entre los dos primeros a Pseudoeuryurus, ya que este autor (Ameghino, 1889: 851) lo describió como una forma intermedia entre ambos. El origen de esta secuencia evolutiva lo colocó tentativamente en el género Plohophorus, incertidumbre despejada por Ameghino (1895) al considerar a Neuryurus antiquus (= Urotherium antiquus) como un taxón muy próximo a Plohophorus.
Posteriormente y sin modificar sus propuesta genealógica, Ameghino (1891a) agrupó a Propalaehoplophorus, Cochlops y Asterostemma en una nueva familia, Propalaehoplophoridae, incluyendo posteriormente en ésta a los géneros Eucinepeltus Ameghino (1891b), Glyptatelus Ameghino (1897), Metopotoxus Ameghino (1898) y Lomaphorelus Ameghino (1902).
Scott (1903-1904) agrupó en la familia Glyptodontidae los géneros: Asterostemma, Cochlops, Eucinepeltus, y Propalaehoplophorus, y con dudas en Doedicuridae a Metopotoxus. A diferencia de Ameghino (1889) sostuvo que Cochlops y Eucinepeltus no revestían importancia filogenética, Asterostemma y Propalaehoplophorus estarían relacionados con los ancestros de los gliptodontes más modernos (v. gr. Glyptodon) y Metopotoxus, podría relacionarse con Panochthus.Castellanos (1931, 1932, 1959) abordó el estudio sistemático de los gliptodontes, basándose en las propuestas morfoevolutivas ameghinianas antes mecionadas, aunque con diferentes resultados (figura 2). En su primera propuesta, Castellanos (1931, 1932) reconoció las familias Palaeopeltidae, Glyptodontidae, Propalaehoplophoridae (Propalaehoplorinae y Glyptatelinae), Doedicuridae (Doedicurinae y Panochthinae) y Hoplophoridae (=Sclerocalyptidae) (Hoplophoridae (=Sclerocalyptinae) y Plohophorinae [= Palaehoplophorinae Castellanos, 1931) (figura 2.1). Filogenéticamente, ubicó a los Palaeopeltidae como una rama basal divergente y a los Propalaehoplophoridae como el grupo ancestral de los restantes gliptodontes. Dentro de esta última familia, dividió a los Glyptatelinae en dos grupos innominados. Un grupo se iniciaría con Glyptatelus, del que surgiría un "taxón desconocido" que luego habría originado divergentemente a Eucinepeltus, Asterostemma y Metopotoxus. De estos últimos se separarían sendas líneas evolutivas que conducen a cuatro géneros de la subfamilia Hoplophorinae (=Sclerocalyptinae). Metopotoxus iniciaría la secuencia que termina en Hoplophorus (= Sclerocalyptus sensu Ameghino, 1891a), previa aparición de Eosclerocalyptus C. Ameghino, mientras que Asterostemma daría origen linealmente a Parahoplophorus Castellanos y posiblemente a Zaphilus. El segundo grupo de la subfamilia Glyptatelinae incluyó a Lomaphorelus como el ancestro de la secuencia evolutiva de los Hoplophorinae (=Sclerocalyptinae) Lomaphorops Castellanos, Trachycalyptus Ameghino y Lomaphorus. Estas hipótesis parafiléticas contrastan con las propuestas previas, ya que el origen no está en Propalaeohoplophorus sino en Glyptatelus.

Figura 2. 1, Relaciones filogenéticas de los gliptodontes basadas en Castellanos (1931, 1932) y 2, relaciones filogenéticas de los gliptodontes basadas en Castellanos (1959) [Modificado de Castellanos, 1959] / 1, phylogenetic relationships of glyptodonts based on Castellanos (1931, 1932) and 2, phylogenetic relationships of glyptodonts based on Castellanos (1959). [Modified from Castellanos, 1959].
Los Propalaehoplophorinae, compuestos por Cochlops, Glyptotherium Osborn y Propalaehoplophorus, se habrían bifurcado en dos líneas evolutivas, luego de haber pasado por un "taxón desconocido". La primera se iniciaría en Cochlops, dando un "ancestro desconocido" del que divergentemente, ya como Glyptodontidae, surgiría una rama que desembocaría en el género Neothoracophorus y en otra que, previo paso por Paraglyptodon Castellanos, conduciría a Glyptodon. Esta propuesta recupera la relación genealógica entre Cochlops y Glyptodon de Ameghino (1889) que fuera descartada por Scott (1903-1904). La posición intermedia de Paraglyptodon coincide con las ideas ameghinianas, dado que este último género fue erigido por Castellanos (1932) sobre la especie Glyptodon chapadmalensis Ameghino (1908).
La segunda línea genealógica de Castellanos (1931, 1932) tuvo como origen, directa o indirectamente, el género Propalaehoplophorus, del que surgirían cuatro linajes evolutivos. El primero, representado por un "taxón desconocido" que en América del Norte originaría a Glyptotherium. El segundo, caracterizado por presentar más de una hilera de figuras periféricas, se iniciaría en Stromaphorus del que divergentemente surgirían Pseudoplohophorus y Plohophorus, y de este último Plohophoroides. El tercer grupo comenzaría con Palaehoplophorus, del que surgiría divergentemente Protoglyptodon. Las últimas dos hipótesis se diferencian de las de Ameghino (1889), ya que para éste Palaehoplophorus y Plohophorus eran dos taxones ancestrales del grupo que presentaba tubo caudal, mientras que para Castellanos (1931, 1932) estos taxones sólo representan ramas terminales.La última línea evolutiva comprende a los Doedicuridae, cuyo carácter distintivo sería la presencia de figuras cóncavas rugosas en el tubo caudal, que surgiría de un "ancestro desconocido" incluido en el "tronco común de los Propalaehoplophorus". De éste y luego de otro "ancestro desconocido", surgirían los Panochthinae Nopachthus-Propanochthus-Panochthus y los Doedicurinae. Estos últimos se dividen en dos ramas evolutivas, una se iniciaría con el género Urotherium, y divergentemente de éste Pseudoeuryrus y Neuryurus, y la otra luego de pasar por un "taxón desconocido", conduciría a Eleutherocercus-Comaphorus y Palaeodoedicurus Castellanos. De este último surgiría el género Xiphuroides Castellanos, que originaría por un lado a Prodaedicurus Castellanos y Doedicurus, y por otro a Plaxaplous. Taxonómicamente, la reunión de panoctinos y doedicurinos difiere de la propuesta de Ameghino (1889), quien los reconcocía en dos familias distintas, mientras que las relaciones filogenéticas son parcialmente coincidentes entre ambos autores. Cabe destacar que dicha coincidencia genealógica no tiene la misma base morfológica, dado que Ameghino (1889) los relacionó mediante las placas óseas mientras que Castellanos (1931, 1932) mediante el tubo caudal. En 1940, Castellanos modificó su clasificación, traspasando a Urotherium, Pseudoeuryurus y Neuryurus desde Doedicurinae a Plohophorinae. Posteriormente, Castellanos (1946) reconoció a Palaehoplophorus como el ancestro del género Protoglyptodon, y las líneas evolutivas que comenzarían con Lomaphorops, Urotherium y Nopachthus. En 1959, Castellanos presentó su última síntesis sistemática de los gliptodontes, en la que se destacan varias hipótesis genealógicas nuevas (figura 2.2) y cambios taxonómicos.
En los Glyptodontoidea Castellanos (1959) posicionó a las familias Palaeopeltidae, con el género Palaeopeltis, y Glyptodontidae subdividida en Propalaehoplophorinae, Glyptodontinae, Doedicurinae y Hoplophorinae (=Sclerocalyptinae). En la primera subfamilia incluyó todos los géneros que Castellanos (1931, 1932, 1940) había asignado a la subfamilia Glyptatelinae y posicionó a Cochlops en la subfamilia Glyptodontinae. Glyptatelus fue considerado por Castellanos (1959) como el ancestro de tres líneas evolutivas: Metopotoxus-Eucinepeltus, Propalaehoplophorus-Asterostemma y Cohlops-Glyptodon. Esta última corresponde a la subfamilia Glyptodontinae. El restante Propalaehoplophorinae, Lomaphorelus, fue considerado por Castellanos (1959) como posible ancestro de las tribus Trachycalyptini, Hoplophorini y Plohophorini de su Hoplophorinae (=Sclerocalyptinae). En la primera tribu ubicó algunos taxones que Castellanos (1940) había incluido en las subfamilias Plohophorinae y Hoplophorinae (=Sclerocalyptinae) modificando sus relaciones gnealógicas. En concreto, Trachycalyptus sería el ancestro de la secuencia Lomaphorops-Lomaphorus, mientras que la relación Urotherium-Pseudoeuryurus fue modificada al ser considerados como géneros divergentes de un ancestro común. Con respecto a los Plohophorini, Castellanos (1959) retomó parcialmente su esquema sistemático de los años ‘30 y desechó sus propuestas de 1946. Al respecto, un "ancestro desconocido" habría originado a Palaehoplophorus y de éste Parahoplophorus (ex Hoplophorinae). De dicho "ancestro" se habrían originados linealmente dos "ancestros desconocidos". Del primero surgirían linealmente Protoglyptodon y Plohophorops, mientras que divergentemente del segundo lo harían Pseudoplohophorus y la secuencia Plohophorus-Plohophoroides. Cabe mencionar que la posición del último "ancestro" mencionado era ocupada por el género Stromaphorus (Castellanos, 1931), mientras que en este esquema dicho género fue considerado como un taxón terminal de los Hoplophorini. En esta última tribu Castellanos (1959) sólo mantuvo la supuesta relación ancestro-descendiente entre los géneros Eosclerocalytpus y Neosclerocalyptus, mientras que la posición de los restantes taxones de dicha tribu fueron modificadas. Al respecto, Zaphilus, previamente reconocido como un descendiente de Parahoplophorus, ahora lo sería de Sclerocalytpus.
Por último, Castellanos (1959) incluyó en los Doedicurinae los restante taxones, aunque con rango de tribu: Doedicurini y Panochthini. Esta última tribu se habría originado directamente del Doedicurini Castellanosia Kraglievich, taxón sólo conocido por elementos de esqueleto caudal. Cabe destacar que Ameghino (1889) había relacionado los panoctinos con los plohoforinos dada la similitud presente entre los osteodermos de ambos taxones. Por otra parte, el género Palaeopeltis, considerado como una rama basal y lateral de los gliptodontes (Castellanos, 1931, 1932), fue posicionado por Castellanos (1959) como el grupo hermano de los Doedicurinae, y ambos como una línea divergente de los restantes gliptodontes.
Hoffstetter (1958) presentó una detallada síntesis crítica de la sistemática de los gliptodontes, reconociendo que "Le sectionnement en sous-familles et en tribus est plus délicat. Beaucoup de formes sont très imparfaitement connues, certaines par une seule plaque. Dans ces conditions, il est illusoire d'essayer d'établir des relations phylétiques et des divisions naturelles. On s'est résolu à accepter, au moins pour les formes anciennes, un sectionnement horizontal, qui n'a évidemment qu´un caractère provisoire". Así, dividió la familia Glyptodontidae en: Glyptatelinae [Glyptatelus y Lomaphorelus]; Propalaehoplophorinae [Asterostemma, Cochlops, Eucinepeltus, Metopotoxus y Propalaehoplophorus]; Doedicurinae [Castellanosia, Comaphorus, Doedicuroides, Doedicurus, Eleutherocercus, Palaeodaedicurus, Plaxaplous, Prodaedicurus y Xiphuroides]; Glyptodontinae [Glyptodon, Glyptodontidium, Glyptotherium y Paraglyptodon] y Hoplophorinae (=Sclerocalyptinae sensu Hoffstetter 1958). Este último incluía siete tribus: Hoplophorini (=Sclerocalyptini sensu Hoffstetter 1958) [Eosclerocalyptus, Eosclerophorus, Hoplophractus, Hoplophorus, Isolinia, Neosclerocalyptus y Trabalia]; Palaehoplophorini [Aspidocalyptus, Chlamyphractus, Palaehoplophorus, Protoglyptodon y Pseudoeuryurus y]; Plohophorini [Berthawyleria, Coscinocercus, Parahoplophorus, Phlyctaenopiga, Plohophoroides, Plohophorops, Plohophorus, Pseudoplohophorus, Stromaphoropsis, Stromaphorus, Teisseiria y Zaphilus]; Lomaphorini [Lomaphorops, Lomaphorus, Peiranoa, Trachycalyptus y Urotherium]; Neothoracophorini [Neothoracophorus]; Neuryurini [Neuryurus] y Panochthini [Nopachthus, Panochthus y Propanochthus]. Las diferencias más notables con los esquemas propuestos por Castellanos (1931, 1932, 1940, 1959) se presentan en la exclusión de los Panochthini y Neothoracophorini de las subfamilias Doedicurinae y Glyptodontinae, respectivamente.
Hoffstetter (1958) propuso relaciones a nivel tribal y subfamiliar, basado en la similitud morfológica del caparazón y el tubo caudal, entre algunos géneros incluidos en esos niveles taxonómicos (figura 3). Así, consideró a los Gliptatelinae como el grupo ancestral de los Propalaehoplophorinae y en éstos reconoció una clara similitud entre Metopotoxus y Panochthus y consideró que Asterostemma era un género muy próximo de Propalaehoplophorus y posible ancestro de numerosos gliptodontes recientes.

Figura 3. Relaciones filogenéticas de los gliptodontes basadas en Hoffstetter (1958). [Tomado de Hoffstetter, 1958] / phylogenetic relationships of gliptodonts based on Hoffstetter (1958).
Hoffstetter (1958) dividió a los Hoplophorinae (=Sclerocalyptinae) en siete tribus. Los Palaehoplophorini fueron caracterizados por la presencia de grandes perforaciones presentes en las placas óseas y en el tubo caudal. En su descripción del género Palaehoplophorus reconoció que la ausencia de placas terminales en el tubo caudal indicaría un estado muy primitivo, que habría inducido a varios autores a reconocerlo como el ancestro de varios grupos más recientes. Sin embargo, la presencia de más de una hilera de figuras periféricas mal definidas en las placas del caparazón estaría indicando para Hoffstetter (1958) que no podía tratarse de una forma ancestral generalizada. En este contexto, relacionó indirectamente esta tribu con los Plohophorini a través de Pseudoplohophorus, aunque reconoció que este último bien podría representar un Palaehoplophorini.
Con respecto a la tribu Plohophorini, Hoffstetter (1958) reconoció que el aumento de figuras periféricas registrado en este grupo, estaría indicando una estrecha relación filogenérica con la tribu Panochthini. Por otra parte, Hoffstetter (1958) sostuvo que la morfología de las placas óseas del caparazón relacionaría a los Lomaphorini Urotherium y Trachycalyptus con la tribu Neothoracophorini y el Neuryurini Neuryurus, respectivamente. Con respecto a este último género, también reconoció que la presencia de ampollas cónicas en el tubo caudal lo asemejan a los géneros Hoplophorus (sensu Paula Couto, 1957) y Panochthus, mientras que el escudo cefálico de este último es muy similar al de Neuryurus. La última tribu reconocida por Hoffstetter (1958), Hoplophorini (=Sclerocalyptini), no fue relacionada filogenéticamente con las restantes tribus y sólo reconoció a Eosclerocalyptus como el ancestro directo de Neosclerocalyptus.
Por último, Hoffstetter (1958) reconoció que las relaciones genealógicas de la subfamilia Glyptodontinae eran un enigma y sólo reconoció a Glyptodontidium como la forma ancestral del grupo que presentaba una escultura externa de las placas óseas similar a la de Propalaehoplophorus. Con respecto a la subfamilia Doedicurinae, Hoffstetter (1958) no postuló ninguna hipótesis filogenética.
Materiales y métodos
Los ejemplares estudiados en este trabajo se listan en el apéndice I.
Morfología craneana. La particular morfología craneana de los gliptodontes hace imprescindible estandarizar la forma de obtener las medidas o relaciones entre los diferentes accidentes morfológicos evaluados en este trabajo. En este sentido, los cráneos se posicionaron con los últimos molariformes paralelos al plano horizontal (figura 4), de forma tal que las observaciones puedan ser repetidas por otros investigadores.

Figura 4. Cráneo de Glyptodon en vista lateral. Posición en la que fueron evaluados los caracteres craneanos / skull of Glyptodon in lateral view. Position in which the cranial characters were evaluated.
Análisis filogenético. Un conjunto de 12 géneros de gliptodontes y 4 taxones externos fueron evaluados para 84 caracteres craneomandibulares (apéndice II y III). La matriz resultante se analizó bajo pesos iguales y en forma exhautiva con el programa NONA 2.0 (Goloboff, 1993). Los índices de soporte fueron obtenidos mediante Bremmer Support (Bremmer, 1994) [BS] y Parsimony Jacknife (Farris et al., 1996) [PJ].
Grupo interno. Atento a al clasificación de Hoffstetter (1958) se estudiaron un conjunto de ejemplares que representan los siguientes grupos: Glyptodontinae (Glyptodon), Doedicurinae (Doedicurus) Propalaehoplophorinae (Propalaehoplophorus y Eucinepeltus) y Hoplophorinae (=Sclerocalyptinae) Hoplophorini (=Sclerocalyptini) (Neosclerocalyptus, Eosclerocalyptus y Hoplophractus) Plohophorini (Stromaphorus, Pseudoplohophorus y Plohophorus) Panochthini (Panochthus) y Lomaphorini (Urotherium).
Comentarios sobre Hoplophractus. Perea (2005) lo sinonimizó el género Hoplophractus a Eosclerocalyptus destacando que las diferencias craneanas (altura de las coanas) y de la coraza se condicen con un diferenciación específica. No obstante, la altura de las coanas, así como otro conjunto de caracteres craneanos (v. gr. número de molariformes trilobulados) se presentan constante a nivel génerico en los restantes taxones de gliptodontes (Fernicola, 2005). Así, es posible considerar, al menos, a las especies aquí incluidas Hoplophractus tapinocephalus y Eosclerocalyptus planus como géneros distintos (Fernicola, 2005).
Comentarios sobre Sclerocalyptus. Lund (1839) describió un nuevo género y especie que denominó Hoplophorus euphractus, sobre distintos elementos óseos hallado en Minas Gerais, Brasil. Dicho nombre genérico fue reemplazado por Sclerocalyptus, por Ameghino (1891a) al considerarlo preocupado. Dicha sinonimia, aceptada por la comunidad paleontológica, fue cuestionada por Simpson (1945), quien consideró a Sclerocalyptus como un sinónimo objetivo de Hoplophorus. Paula Couto (1957) describió un ejemplar de Hoplophorus euphractus, que por sus características no podía ser considerado cogenérico de las restantes especie pampeanas de "Sclerocalyptus". En tal contexto, revalidó la sinonimia objetiva de Sclerocalyptus con Hoplophorus, propuesta por Simpson (1945), e incluyó en un nuevo género, Neosclerocalyptus a las restantes especies incluidas hasta ese entonces en Sclerocalyptus, designando como especie tipo a Glyptodon ornatus. Cabe destacar que esta última había sido erróneamente reconocida como tipo de Hoplophorus por Ameghino (1891a), ya que Hoplophorus euphractus debió ser considerada la especie tipo por monotipia (Paula Couto, 1947). La propuesta de Paula Couto (1957) no sólo refleja la objetividad necesaria en estos casos, sino que de haber preservado el nombre genérico Sclerocalyptus bajo el concepto de nomen protectum (ICZN, 2000) se habría originado una gran confusión taxonómica, dado que el mismo habría quedado restringido a las formas de Brasil. Por lo expuesto en este trabajo se sigue la propuesta de Paula Couto (1957).
Grupos externos. De acuerdo a la reseña histórica presentada, los grupos externos utilizados en este trabajo corresponden a los géneros Eutatus, Pampatherium, Dasypus y Euphractus.
Distribución cronológica. La presencia de gliptodontes en el continente americano desde el Eoceno hasta el Holoceno nos indujo a mixturar los esquemas "cronológicos" propuestos bajo los conceptos de Edad Mamífero (Pascual et al. 1965, 1966; Marshall et al., 1983; Flynn y Swisher, 1995) y Piso/Edad (Cione y Tonni, 1995a y 1995b; Cione et al., 2000). En tal sentido, el esquema utilizado abarca el Piso/Edad Laventense hasta la actualidad, de Cione et al. (2000) y Madden et al. (1997), y el esquema de "Edades Mamífero" prelaventense de Flynn y Swisher (1995). La primera aparición de los géneros y taxones supragenéricos utilizados en este trabajo fue tomada de Marshall y Patterson (1981), Marshall et al. (1983), Pascual et al. (1965), Pascual et al. (1996), Cabrera (1944), Cione et al. (1999, 2000) Scillato-Yané et al. (1995) y Perea (2005).
Resultados
Tres árboles más parsimoniosos de 181 pasos fueron obtenidos (CI 0,70; CIinf 0,69; RI 0,79). El conflicto entre esas hipótesis se produce por la posición fluctuante del género Urotherium (figura 5.1). La exclusión de este último, permite obtener un solo árbol de 177 pasos (CI 0,72; CIinf 0,71; RI 0,80) (figura 5.2). NODO A [Glyptodontia Ameghino 1889] (BS: 17 y PJ: 100). El conjunto de sinapomorfías no ambiguas de máximo ajuste [ci: 100; ri: 100] que sustentan este clado son: cóndilo articular de la mandíbula posterior a la hilera dentaria por menos de dos molariformes (3.2); ángulo entre la rama horizontal y la rama ascendente mayor a 90° (5.1); distancia entre el borde posterior de la mandíbula con respecto al cóndilo occipital mayor o igual a la longitud lateromedial del cóndilo (8.1); forma de la porción posterodorsal de la porción angular (9.1); sínfisis mandibular proyectada medialmente en toda su extensión (14.2); superficie oclusal de los molariformes superiores plana (23.2); pared medial del foramen suborbitario claramente lateral con respecto al borde labial de la hilera dentaria (39.2); paladar sigmoideo (47.1); plano anterior de los nasales en línea con el plano anteroventral de los premaxilares (54.1); plano posterior de los nasales posterior al plano de los forámenes lagrimales (56.1); relación entre la longitud del rostro y la longitud entre los forámenes lagrimales menor a 0.9 (58.2); cavidad glenoidea posterior al último molariforme por menos de una longitud anteroposterior de este último (64.2); sutura yugal-escamoso en forma de S (66.2); eje principal de la sutura yugalescamosa dirigido posteroventralmente (67.1); lámina frontal descendente desarrollada ventralmente (71.2) (figuras 6 y 7).

Figura 5. Hipótesis más parsimoniosa obtenida en el análisis filogenético de 12 géneros de gliptodontes y 4 grupos externos de armadillos utilizando Nona (Goloboff, 1993). 1, excluyendo al género Urotherium (largo: 180; CI: 0,71; RI: 0,79). 2, incluyendo al género Urotherium (largo: 184; CI 0,70; RI: 0,78). / more parsimonious hypothesis obtained in the phylogenetic analysis of 12 glyptodonts and 4 outgroup of armadillos using NONA (Goloboff, 1993). 1, excluding to the genus Urotherium (Length: 180; CI: 0,71; RI: 0,79). 2, including to the genus Urotherium (Length: 184; CI 0,70; RI: 0,78). Sinapomorfías no ambiguas (en negritas sinapomorfías no homoplásticas, en letra común sinapomorfías homoplásticas) presentes en los nodos internos son discutidas en el texto / unambiguous synapomorphies (in bold type non-homoplastic synapomorphies, in plane type homoplastic synapomorphy) present in the internal nodes are discussed in the text. Nodo A / Node A (Glyptodontia): 3 (2), 5 (3), 6 (0), 8 (1), 9 (1), 14 (2), 15 (1), 18 (0), 23 (2), 25 (1), 30 (2), 39 (2), 41 (1), 45 (1), 47 (1), 54 (1), 56 (1), 58 (2), 60 (1), 62 (1), 64 (2), 66 (2), 67 (2), 69 (1), 71 (2), 75 (2), 78 (1), 81 (1), 83 (1); Nodo B / Node B (Propalaehoplophoroinei): 29 (2), 49 (3); Nodo C / Node C (Glyptodontoinei): 20 (2), 35 (1), 36(1), 64 (3), 80 (1), 81 (2); Nodo D / Node D: 2 (3); Nodo E / Node E: 41 (0), 43 (1); Nodo F / Node F: 45 (2), 46 (2), 81 (3); Nodo G / Node G (Glyptodontoidea): 15 (0), 25 (0); Nodo H / Node H (Glyptodontidae): 74 (2); Nodo I / Node I (Glyptodontinae): 41 (1), 77 (1), 79 (2), 80 (0); Nodo J / Node J (Panochthidae): 1 (3), 28 (2), 30 (3), 44 (1), 52 (1), 53 (1), 54 (2), 57 (1), 70 (0), 72 (1), 76 (1), 82 (1). Números debajo de las ramas indican los valores de Bremer Support (izquierda de la diagnal) y Parsimony Jackknife (derecha de la diagonal) de los nodos ubicados a su derecha. Sólo se muestran los valores de jackknife superiores a 50% / numbers below branches indicate Bremer Support (left of the diagonal) and Parsimony Jackknife (right to the diagonal) values of the nodes at their right. Only jackknife values above 50% are shown.

Figura 6. Mandíbulas de Glyptodon en vistas lateral (1) y medial (3) y de Euphractus en vista lateral (2). Abreviaturas. CA, cóndilo articular; PA, proceso angular; PC, proceso coronoideo; SM, sínfisis mandibular. Los caracteres y estados ilustrados representan las sinapomorfías del Nodo A (Glyptodontia) discutidos en el texto. / mandibles of Glyptodon in lateral (1) and medial (3) views and the Euphractus in lateral view (2). Abbrevations. CA, articular condyle; PA, angular process; PC, condyloid process; SM, mandibular symphisys. The drawn characters and states represent synapomorphies of Node A (Glyptodontia) are discussed in the text.

Figura 7. Cráneo de Eucinepeltus en vista lateral (1) y vista anterior (2). Abreviaturas. ES, escamoso; JU, yugal. Los caracteres y estados ilustrados corresponden a sinapomorfías del Nodo A (Glyptodontia) discutidos en el texto / skull of Eucinepeltus in lateral view (1) and anterior view (2). Abbrevations. ES, squamosal; JU, jugal. Drawn characters and states correspond to synapomorphies of Node A (Glyptodontia) are discussed in the text.
Otras sinapomorfías no ambiguas que avalan el NODO A presentan bajo ajuste como consecuencia de la homoplasia existente con los géneros externos Dasypus: ocho dientes mandibulares (18.0) [ci: 67; ri:50]; cavidad glenoidea en línea con el cóndilo occipital (62.1) [ci: 67; ri:50] y ausencia de cresta lateral en el proceso coronoides (6.0) [ci: 50; ri:50] y Eutatus: foramen lagrimal próximo a la tabla craneana (75.2) [ci: 67; ri:50]. Al mismo tiempo, la porción rugosa del pterigoide ubicada a nivel del último molariforme (60.1) y yugal superpuesto al escamoso en vista lateral (69.1), se presentan como sinapomorfías no ambiguas pero de menor ajuste [ci: 50; ri: 75] dadas sendas reversiones (1 -> 0) presentes, respectivamente, en Doedicurus (60.0) y Glyptodon (69.0), con respecto a los grupos externos.
Otra sinapomorfía no ambigua es la relación alto/ancho de la apertura nasal posterior mayor a 0,75 y menor a 1,1 (30.1) [ci: 66: ri:80]. Este último carácter también se presenta como una homoplasia (30.2) entre el clado (Panochthus + Neosclerocalyptus) y el género Eosclerocalyptus. El fuerte desarrollo del proceso cigomático del maxilar con respecto a la longitud total de la hilera dentaria (45.1) [ci: 75; ri: 88] es una sinapomorfía no ambigua, cuyo desajuste obedece a la homoplasia (45.3) presente entre el clado (Panochthus + Neosclerocalyptus) y el género Doedicurus.
Otro conjunto de sinapomorfías no ambiguas sustentan la monofilia de los gliptodontes y como reversiones apoyan clados más derivados. En concreto, el foramen lagrimal en línea o medial a la pared externa del foramen suborbitario (41.1) [ci: 33; ri: 60] presenta dos reversiones: [1->0] en el NODO E con respecto a los grupos externos y [0->1] en el clado (Glyptodon + Doedicurus) con respecto al nodo anterior. Asimismo, la porción angular de la mandíbula proyectada ventromedialmente (15.1) [ci: 50; ri: 75] y primer molariforme aproximadamente isodiamétrico (25.1) [ci: 50; ri: 75], revierten [1->0] en el NODO G con respecto a los grupos externos. Por otra parte, el plano occipital inclinado a menos de 80° (81.1) [ci: 75; ri: 91] se resuelve como una sinapomorfía de los gliptodontes, mientras que menores inclinaciones sustentan agrupamientos más derivados (NODO C).
Por último, el buen desarrollo del proceso yugal del frontal (78.1) [ci: árbol 1: 50; árboles 0 y 2: 40] y la cresta occipital fuertemente desarrollada (83.1) [árbol 1: ci: 20 ri: 33; árboles 0 y 2 ci: 25 ri: 50] son sinapomorfías basales, pero presentan un muy bajo ajuste dada la conflictividad presente en la porción más derivada de los cladogramas.
NODO B [Infraorden Propalaehoplophoroinei Ameghino 1891a] (Propalaehoplophorus + Eucinepeltus) (BS: 1 y PJ: 61). Dos sinapomorfías no ambiguas definen este grupo: apertura nasal posterior en forma de cerradura (29.2) [ci: 100; ri: 100] y borde anterior del primer alvéolo maxilar anterior al borde posterior de los forámenes incisivos (49.3) [ci: 75: ri: 80], estado también presente en el género Doedicurus. NODO C [Infraorden Glyptodontoinei Gray 1869] (Stromaphorus + NODO D) (BS: 5 y PJ: 84). Las sinapomorfías de máximo ajuste que definen este agrupamiento son: últimos molariformes de la mandíbula trilobulados con el primer lóbulo triangular (20.2); foramen oval posterior con respecto a la cavidad glenoidea (35.1); ausencia de un foramen vascular anterior al foramen oval (36.1) y cavidad glenoidea posterior a la hilera dentaria por menos de un molariforme (64.3). Otras dos sinapomorfías no ambiguas, aunque de menor ajuste, avalan este clado: cóndilo occipital más desarrollado dorsoventralmente (80.1) [ci. 50; ri. 75], característica presente como una reversión [1-> 0] en el clado (Glyptodon + Doedicurus) y en el género polimórfico [01] Neosclerocalyptus y el plano occipital dirigido entre 45° y 65° (81.2) [ci. 60; ri. 91], que revierte [3-> 2] en el género Neosclerocalyptus (figura 8).

Figura 8. Cráneo de Eosclerocalyptus en vista lateral (1) y mandíbula de Panochthus en vista dorsal (2). Abreviaturas. CG, cavidad glenoidea; FO, foramen oval. Los caracteres y estados ilustrados corresponden a sinapomorfías de los Nodo C (Glyptodontoinei) y Nodo G (Glyptodontoidea) discutidas en el texto / skull of Eosclerocalyptus in lateral view (A) and mandible of Panochthus in dorsal view (B). Abbrevations. CG, glenoid fossa; FO, foramen ovale. Drawn characters and states correspond to synapomorphies of Node C (Glyptodontoinei) and Node G (Glyptodontoidea) are discussed in the text.
NODO D (Hoplophractus + NODO E) (BS: 1). Una sóla sinapomorfía no ambigua soporta este agrupamiento: tres molariformes posteriores ocultos por la rama ascendente de la mandíbula (2.3) [ci: 60; ri. 60]. Esta característica se presenta como una reversión [3 > 2] en el género Doedicurus con respecto a los géneros Eutatus, Pampatherium, Propalaehoplophorus, Eucinepeltus y Stromaphorus.
NODO E (Eosclerocalyptus + (NODO F)) (BS: 1). Este clado presenta una sinapomorfía no ambigua de máximo ajuste: posición anterior del foramen suborbitario con respecto a la hilera dentaria (43.1) [ci: 100; ri: 100] y como fuera mencionado una reversión representada por el foramen lagrimal lateral al foramen suborbitario (41.0) [ci: 33; ri: 60]. NODO F (Pseudoplohophorus + (NODO G)) (BS: 1). Tres sinapomorfías no ambiguas sustentan este agrupamientos: mayor desarrollo de la superficie externa del proceso descendente con respecto a la superficie interna (46.2) [ci: 100; ri:100]; proceso descendente del arco cigomático representa más del 60% y menos del 80% de la hilera dentaria del maxilar (45.2) [ci: 75; ri:88], cuyo desajuste se debe a un posible paralelismo [2 -> 3] entre el clado (Panochthus + Neosclerocalyptus) y el género Doedicurus. Por último, el ángulo del plano occipital menor a 45° (81.3) [ci:60; ri:83], se presenta como una sinaporfía en la hipótesis que presenta a Urotherium y Plohophorus como un grupo monofilético, hipótesis que contempla una reversión [3 -> 2] entre Urotherium y Neosclerocalyptus, mientras que en las otras dos hipótesis sólo adquiere un carácter potencial.
NODO G [Superfamilia Glyptodontoidea Gray, 1869] (Plohophorus Urotherium (Glyptodon + Doedicurus) (Panochthus + Neosclerocalyptus)) (BS: 2 y PJ: 53). Dos sinapomorfías no ambiguas sustentan este clado: proyección ventromedial de la región angular ausente (15.0) [ci:50; ri: 75] y primer molariforme del maxilar alargado anteroposteriormente (25.0) [ci:50; ri: 75]. Características que, como fuera mencionado, se presentan como sendas reversiones [1 -> 0] con respecto a los grupos externos (figura 8). Las tres resoluciones internas del NODO G obedecen a la posición fluctuante del género Urotherium (figura 5.1). Las hipótesis 1 (Urotherium (Plohophorus (Glyptodon + Doedicurus))) y 2 ((Urotherium + Plohophorus) (Glyptodon + Doedicurus)) se sustentan en el foramen lagrimal ventral al borde ventral del cóndilo occipital (74.2) [ci: 66; ri: 75]. A su vez, el clado (Plohophorus (Glyptodon + Doedicurus)), presente en la primera hipótesis, se apoya en una cresta occipital ausente o poco desarrollada (83.0) [ci: 25; ri: 50], siendo la misma una homoplasia con el género Panochthus y una reversión de los grupos externos Eutatus, Euphractus y Pampatherium. Por otra parte, el clado (Urotherium + Plohophorus), presente en la segunda hipótesis, se sustenta en el rudimentario desarrollo del proceso yugal del frontal (78.0) [ci: 50; ri: 66], siendo el mismo una reversión con respecto a los grupos externos.
En la hipótesis 3 se reconocen dos grupos, ((Plohophorus (Glyptodon + Doedicurus)) + (Urotherium (Panochthus + Neosclerocalyptus))). El pimer clado se sustenta en una cresta occipital ausente o poco desarrollada (83.0) [ci: 25; ri:50], mientras que el segundo se sustenta en la ausencia de cresta sagital (70.0) [ci: 50; ri:80], estado que revierte con respecto a los géneros externos Dasypus, Eutatus y Euphractus.
En este contexto, el consenso de estas hipótesis encubre la estrecha relación existente entre Plohophorus y (Glyptodon + Doedicurus), por tal motivo, la "exclusión" del género Urotherium permite definir el NODO H [Familia Glyptodontidae Gray, 1869] ((Plohophorus + (Glyptodon + Doedicurus)) (BS: 1 PS: 54), apoyado en el foramen lagrimal ventral al borde ventral del cóndilo occipital (74.2) [ci: 66; ri: 66].Cabe destacar que el mismo estado del carácter soportaba el clado ((Urotherium + Plohophorus) (Glyptodon + Doedicurus)) y que la indefinición posicional del género Urotherium, más allá de las entradas faltantes, se debe a que comparte con el clado (Panochthus + Neosclerocalyptus) el carácter (70.0). NODO J [Familia Panochthidae Castellanos, 1927] (Panochthus + Neosclerocalyptus) (BS: 7 y PJ: 99). Este grupo se sustenta en seis sinapomorfías de máximo ajuste [ci: 100; ri: 100]: ancho de la apertura nasal anterior mayor al 95% de la longitud extracondilar (28.4), foramen suborbitario ventral (44.1), borde anterior de los premaxilares alcanza los dientes maxilares en vista lateral (53.1), lámina frontal descendente dirigida a más de 45° (72.1), foramen lagrimal intraorbital (76.1), ancho mínimo postorbital claramente mayor a la longitud extracondilar (82.1). La relación alto/ancho de la apertura nasal posterior (30.3) [ci: 66; ri: 88] se presenta como una homoplasia con el género Eosclerocalyptus (figura 9).
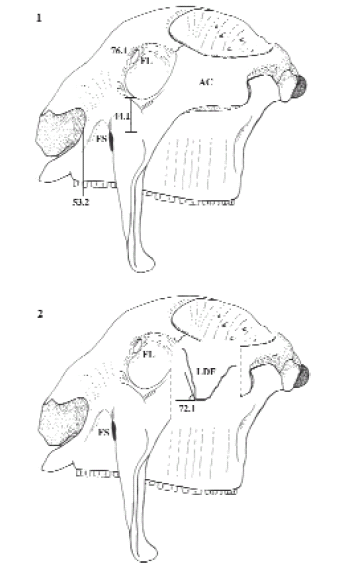
Figura 9. Cráneo de Panochthus en vista lateral. 1, con el arco cigomático y 2, sin el arco cigomático. Abreviaturas. LDF, lámina descendente del frontal; FL, foramen lagrimal; FS, foramen suborbitario. Los caracteres y estados ilustrados corresponden a sinapomorfías del Nodo J (Panochthidae) discutidos en el texto / skull of Panochthus in lateral view. 1, with the cigomatic arch and 2, with cigomatic arch cut. Abbrevations. LDF, descending sheet extensión of the frontal; FL, lacrimal foramen; FS, infraorbital foramen. Drawn characters and states correspond to synapomorphies of Node J (Panochthidae) are discussed in the text.
La posición del género Urotherium como grupo hermano del clado (Panochthus + Neosclerocalyptus) modifica la asignación de estados internos de los siguientes caracteres: borde posterior de la mandíbula ubicado a una distancia menor a 2.5 longitudes anteroposterior del último molariforme (1.3); borde anterior de los nasales ubicados posteriormente al borde anteroventral de los premaxilares (54.2); tabique nasal anterior al borde anterior de los nasales (57.1) y marcado desarrollo dorsoventral del premaxilar (52.1). Estos estados se transforman en sinapomorfías potenciales del clado (Panochthus + Neosclerocalyptus), por lo que nuevos materiales podrán dilucidar esta problemática. NODO I [Subfamilia Glyptodontinae Gray, 1869] (Glyptodon + Doedicurus) (BS: 3 y PJ: 70). Este grupo monofilético esta sustentado por: foramen lagrimal en línea con, o medial a, la pared lateral del foramen suborbitario (41.1) [ci: 33; ri: 60], reversión con respecto a los gliptodontes basales; órbita circular (79.2) [ci: 67], posible convergencia con el género externo Euphractus; cóndilo occipital con su eje lateromedial mayor o igual a su eje dorsoventral (80.0) [ci: 50; ri: 75], característica también presente en el género Neosclerocalyptus [01] y en Propalaehoplophorus, así como en todos los grupos externos, y superficie dorsal al foramen lagrimal irregular (77.1) [ci: 40; ri: 40], estado presente en los géneros Euphractus, Pampatherium, Stromaphorus, Propalaehoplophorus y Panochthus. En estos dos últimos taxones también presentan la superficie dorsal al foramen lagrimal cónica (77.2) (figura 10).
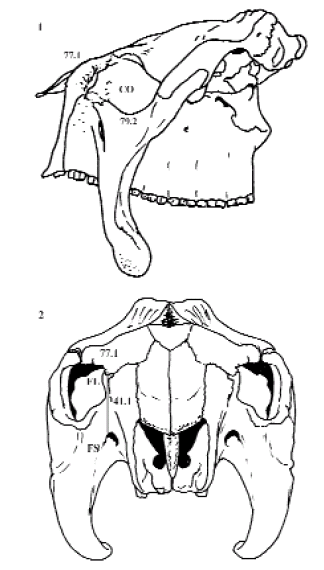
Figura 10. Cráneos de Glyptodon en vista lateral (1) y de Doedicurus en vista anterior (2). Abreviaturas. CO, cavidad orbital; FL, foramen lagrimal; FS, foramen suborbitario. Los caracteres y estados ilustrados corresponden a sinapomorfías del Nodo I (Glyptodontinae) discutidos en el texto / skulls of Glyptodon in lateral view (1) and Doedicurus in anterior view (2). Abbrevations. CO, orbital cavity;; FL, lacrimal foramen; FS, infraorbital foramen. Drawn characters and states correspond to synapomorphies of Node I (Glyptodontinae) are discussed in the text.
Discusión
Filogenia. Históricamente, la monofilia de los gliptodontes no ha sido cuestionada y la composición taxonómica del grupo sólo se ha visto afectada por la inclusión o exclusión de Palaeopeltis. De hecho, los últimos análisis filogenéticos de los cingulados han incluido a los gliptodontes como grupo terminal (v. gr. Engelmann, 1985; Gaudin y Wible, 2006). En tal sentido, este trabajo es el primero en evaluar y sustentar en un marco cladístico la monofilia de los gliptodontes, aunque no fue posible analizar la posición filogenética de los gliptatelinos y Palaeopeltis. Con respecto al primer grupo mencionado los elementos craneanos asignados sin dudas a este taxón son muy fragmentarios y no permiten evaluar los caracteres utilizados en este trabajo. La posición de Palaeopeltis es aún conflictiva y el único fragmento craneano asignado no ha podido ser estudiado. Sin embargo, como sostuviera Hoffstetter (1958), es posible que se trate de un taxón relacionado con los peltefilinos, dado que comparte con estos últimos un ordenamiento dentario atelodonte. No obstante, la posición de este género deberá ser evaluada en una marco filogenético adecuado.
Las hipótesis filogenéticas entre los distintos grupos de gliptodontes propuestas hasta 1959 se sustentaron en supuestos cambios evolutivos, originalmente planteados por Ameghino (1884a, 1889), que daban cuenta independientemente de las diferentes morfologías del tubo caudal y de la escultura externa de las placas óseas de la coraza dorsal con que fueron diagnosticados las distintos grupos reconocidos. La metodología que daba marco a estas hipótesis resolvía las contradicciones entre las mismas estableciendo un orden jerárquico de anidamiento a priori. Por lo tanto, las distintas secuencias jerárquicas pueden arrojar distintos esquemas filogenéticos. En tal sentido, Castellanos (1959), relacionó estrechamente al doedicurino Castellanosia con los panoctinos basado en la morfología caudal, ya que el primer taxón sólo era conocido a través de fragmentos caudales, mientras que para Ameghino (1889) la morfología de las placas de los panoctinos indicaría una estrecha relación con Plohophorus.
Más allá de las distintas valorizaciones aplicadas y por ende de los esquemas obtenidos, las propuestas de Ameghino (1889), Castellanos (1931, 1932, 1940, 1946, 1959) y Hoffstetter (1958) concuerdan, aunque con matices, en considerar a los propalaehoploforinos como un grupo ancestral. Sin embargo, para estos autores el grupo sería parafilético, ya que del mismo se habrían originado las distintas líneas de gliptodontes más modernos. En este estudio, los propaleohoploforinos Eucinepeltus y Propalaehoplophorus se resolvieron como un clado, hermano de los restantes gliptodontes. Sin embargo, no es posible descartar la parafilia del grupo ya que no se han incluido en este estudio los géneros Metopotoxus, Asterostemma y Cochlops.
Las relaciones obtenidas entre los restantes taxones evaluados en este trabajo son netamente contradictorias con las propuestas clásicas del grupo. Con excepción del género Urotherium, cuya posición fluctúa dentro del NODO G, la resolución de los restantes taxones (Stromaphorus (Hoplophractus (Eosclerocalyptus (Pseudoplohophorus ((Panochthus Neosclerocalyptus) + (Plohophorus (Doedicurus Glyptodon))))))), cuestiona claramente la monofilia de los Hoplophorinae (=Sclerocalyptinae sensu Hoffstetter, 1958), así como la de las tribus Plohophorini (sensu Hoffstetter, 1958) y Hoplophorini (= Sclerocalyptini sensu Hoffstetter, 1958) y permite establecer relaciones entre grupos cuya posición era incierta (v. gr. Doedicurinae y Glyptodontinae sensu Hoffstetter, 1958).
En concreto, el género Stromaphorus, taxón hermano de los restantes gliptodontes, fue considerado por Castellanos (1931, 1932, 1946) como el ancestro de Plohophorus, mientras que posteriormente el mismo autor (Castellanos, 1959) lo posicionó como una rama lateral de la línea que conduciría a Neosclerocalyptus. Por su parte, Hoffstetter (1958) consideró que los Panochthini derivarían de un taxón próximo a Stromaphorus.
El género Eosclerocalyptus fue considerado por Castellanos (1931, 1932) como el ancestro directo de Neosclerocalyptus, opinión compartida por Hoffstetter (1958), sin embargo en este estudio se ubica como el grupo hermano del clado (Pseudoplohophorus + NODO G) de forma tal que ambos géneros se posicionan claramente distantes entre sí.
La posición del género Pseudoplohophorus ha variado considerablemente en los esquemas filogenéticos clásicos. Castellanos (1931, 1932) lo reconoció como un género derivado de Stromaphorus, mientras que en 1959 lo ubicó como el grupo hermano de Plohophorus. Por otra parte, Hoffstetter (1958) consideró al género Pseudoplohophorus como un posible Plohophorini, intermediario entre esta tribu y Palaehoplophorini. Estas hipótesis no encuentran sustento en este estudio ya que los Plohophorini resultaron parafiléticos y Pseudoplohophorus no se relacionó con Plohophorus ni con Stromaphorus.
La relación entre Plohophorus / Plohophorini y Panochthini, propuesta por Ameghino (1889) y Hoffstetter (1958) respectivamente, así como la propuesta de Castellanos (1932, 1942) que deriva a los Panochthini, conjuntamente con los doedicurinos de un "tronco común de los Propalaehoplophorus" o de gliptodontes primitivos (Castellanos, 1959) no encuentran aval en este estudio. Los géneros Panochthus, único representante incluido de la tribu Panochthini, y Plohophorus se posicionaron en la porción más derivada del cladograma como el grupo hermano de Neosclerocalyptus y (Glyptodon + Doedicurus) respectivamente. Por último, la estrecha afinidad filogenética que presentan los géneros Doedicurus y Glyptodon, aunque no había sido planteada en los esquemas filogenéticos tradicionales, tampoco los contradice. En efecto, la posición filogenética propuesta para la subfamilia Glyptodontidae fue incierta ya desde Ameghino (1889), quien consideró con dudas al género Glyptodon como descendiente del género Cochlops, mientras que Scott (1904) descartó de plano esta posibilidad. Castellanos (1931, 1932, 1959) retomó la propuesta ameghiniana, aunque sin sustentarla, mientras que Hoffstetter (1958) reconoció que la posición de la subfamilia Glyptodontinae era realmente incierta.
Clasificación. Excluyendo a los palaeopéltidos, todas las clasificaciones hasta hoy propuestas, incluyen cuatro grandes grupos de gliptodontes: Hoplophorinae/dae (=Sclerocalyptinae/dae), Doedicurinae/dae y Glyptodontinae/dae y Propalaehoplophorinae/dae. Esta última subdividida por Castellanos (1931) en Glyptatelinae [Glyptatelus, Eucinepeltus, Metopotoxus, Asterostemma y Lomaphorelus] y Propalaehoplophorinae [Cochlops y Propalaehoplophorus]. Simpson (1945) y Paula Couto (1979) no reconocieron esta separación y agruparon todos esos taxones en la segunda subfamilia. Hoffstetter (1958) retomó la propuesta supragenérica de Castellanos (1931, 1932, 1940) pero sólo incluyó en los Glyptatelinae a Glyptatelus y Lomaphorelus y en los Propalaehoplophorinae a Eucinepeltus, Metopotoxus, Asterostemma, Cochlops y Propalaehoplophorus, propuesta aceptada por Scillato-Yané (1977), Carlini et al. (1997), McKenna y Bell (1997) y Vizcaíno et al., (2003). La posición de los gliptatelinos en los cingulados es conflictiva, ya que los escasos restos conocidos evidencian caracteres presentes en gliptodontes y armadillos. Con respecto a estos últimos comparten la presencia de vértebras dorsales no fusionadas (Carlini et al., 1997) y con los gliptodontes la ausencia en el caparazón de bandas móviles completas (Vizcaíno et al., 2003). Esta última característica ha sido históricamente considerada como un carácter de pertenencia a los gliptodontes, criterio que, en ausencia de estudios filogenéticos que sustenten lo contrario, es compartido en este trabajo, por lo que los gliptatelinos son incluidos en la presente clasificación.
Hoffstetter (1958) individualizó a los Propalaehoplophorinae por presentar una cola similar a la de los armadillos, primeros molariformes simples y plano occipital casi vertical, entre otros rasgos morfológicos. En cuanto a los Glyptodontinae, la presencia de una cola anillada con fuertes tubérculos cónicos, todos los dientes trilobulados y la presencia del primer dedo del pie, entre otros, caracterizaría al grupo. Por otra parte, los Doedicurinae fueron caracterizados por Hoffstetter (1958) por la ausencia de figuras en la superficie externa de las placas óseas del caparazón y, entre otros, primer molariforme simple. Como fuera mencionado, la monofilia de estos dos últimos grupos no fue evaluada, mientras que la de los propalaehoploforinos ha sido parcialmente corroborada. No obstante, el estudio filogenético preliminar de los géneros Cochlops y Metopotoxus permite reconocer que los mismos conforman un clado con Propalaehoplophorus y Eucinepeltus (Fernicola, 2006).
La subfamilia Hoplophorinae (= Sclerocalyptinae sensu Hoffstetter, 1958), grupo claramente conflictivo, fue caracterizado por Hoffstetter (1958: 577) en los siguientes términos "La sous-famille paraît surtout caractérisée par l´acquisition précoce d´un long étui caudal formant un tube cylindroconique. D´une façon générale, elle se distingue des Propaléhoplophorines par une spécialisation plus poussée des dents, du crâne et du squelette des membres. Un caractère important correspond à la disaparition du pouce en avant et en arrière, mais les documents manquent, qui permettraient d´établir la date à la-quelle cette atrophie c´est généralisée". Por otra parte, reconoció que "Tout l´ensemble des autres Glyptodontes [Glyptodontinae y Doedicurinae] connus depuis le Miocène supérieur jusqu´au Pleistocène en Amérique du Sud forme l´énorme sous-famille des Sclerocalyptinae, dans laquelle on devra distinguer au moins sept tribus provisoires.". Tribus que describió como "On peut y distinguer, surtout d´après la morphologie de la carapace et du tube caudal, un certain nombre de tribus, qui ne représentent guère que des groupements provisoires autour des quelques formes les mieux connues".
En este contexto, la tribu Hoplophorini (=Sclero-calyptini sensu Hoffstetter 1958) fue diagnosticada sobre Neosclerocalyptus como un grupo con coraza baja y alargada con alas laterales anteriores, placas delgadas con una figura central rodeada de figuras periféricas, tubo caudal cónico-cilíndrico, perfil craneano convexo por el desarrollo de los senos frontales, nasales inclinados ventralmente, apófisis postorbitaria presente y rama ascendente de la mandíbula inclinada anteriormente. Estas características, listadas por Hoffstetter (1958), fueron evaluadas y su presencia en los Hoplophorini no es constante (v. gr. el cráneo de Eosclerocalyptus no se presenta convexo) o también se observan en taxones no incluidos en la misma (v. gr. inclinación anterior de la rama ascendente de la mandíbula presente en todos los gliptodontes o placas con una figura central y una hilera de períféricas presentes en Propalaehoplophorus y Glyptodon).
En lo atinente a la tribu Plohophorini, Hoffstetter (1958: 581) reconoció que "Cette tribu, assurément synthétique, se distingue essentiellement des Sclerocalyptini [Hoplophorini] par la multiplication des figures périphériques." De los tres géneros de esta tribu, incluidos en este trabajo, sólo se conocía la coraza de Plohophorus y Stromaphorus, que presentan dichas hileras extras. En cuanto al género Pseudoplohophorus, conocido por tubos caudales, Hoffstetter (1958) abrigó la posibilidad de que este taxón y sus "vecinos" pudieran ser incluidos en la tribu Palaehoplohorini. Perea (2005) mencionó que la coraza de Pseudoplohophorus absolutus Perea carece de una doble hilera de figuras periféricas, salvo en algunas placas de la zona pleural, y remarcó la notable similitud que presenta el tubo caudal con el neotipo de Pseudoplohophorus orientalis, material sobre el que Hoffstetter (1958) hiciera sus consideraciones filogenéticas. Este hallazgo refuerza la idea de excluir este género de la tribu Plohophorini, aunque no es claro en qué grupo ubicarlo y en tal sentido, Perea (2005) lo incluyó en los Hoplophorinae (=Sclerocalyptinae), sin referirlo tribalmente.
La tribu Panochthini, caracterizada por Hoffstetter (1958) sobre el género Panochthus, incluye a los taxones Nopachthus y Propanochthus. Estos últimos, presentan en la región posterior de la coraza una amplia región de placas con una figura central elevada, en forma de ampolla, morfología presente en el Plohophorini Phlyctaenopyga Cabrera. De hecho, Cabrera (1944) planteó que Nopachthus trouessarti, sería coespecífico de Phlyctaenopyga ameghini y representaría otra especie de este último género. Sin embargo, las diferencias entre los tubos caudales de ambos taxones posiblemente contribuyó a que Phlyctaenopyga no fuera incluido en esta tribu. Por otra parte, el Lomaphorini Peiranoa Castellanos presenta las ampollas centrales mencionadas, aunque rodeadas de una zona rugosa, mientras que en el propalaehoploforino Cochlops, la figura central de la región posterior de la coraza se eleva abruptamente en forma de conos, característica que reproducen las figuras periféricas. De esta forma, sería posible establecer un conjunto de agrupamientos supragenéricos diferente si se modificara la valorización a priori de los caracteres.
Por último, la tribu Lomaphorini, caracterizada por Hoffstetter (1958) por la presencia de placas con una figura central suavemente delimitada y pérdida de figuras periféricas, ha sido modificada en algunos trabajos en cuanto a su composición genérica. Para Hoffstetter (1958) los géneros incluidos fueron Lomaphorus, Trachycalyptus, Lomaphorops, Peiranoa y Urotherium, mientras que Scillato-Yané et al. (1995) y Cione et al. (2000) ubican al último género en la tribu Neuryurini, aunque sin sustentar dicho cambio.
Como sostuviera Hoffstetter (1958), el reconocimiento de los Hoplophorinae (=Sclerocalyptinae) como grupo natural fue netamente preliminar y avanzando más, es posible sostener que dicha subfamilia actuó como reservorio de distintos taxones que de una u otra forma no podían ser incluidos en las restantes subfamilias, cuya caracterización era más sólida. En tal sentido, es posible, sin mediar análisis filogenético, postular distintos agrupamientos supragenéricos con sólo modificar las condiciones iniciales de agrupamiento. La presencia de placas óseas con una figura central bien desarrollada podría ser reconocida como un carácter de pertenencia grupal y una vez incluidos los taxones con dicha característica, asumir que las diferencias observadas entre otros elementos óseos (v. gr. tubo caudal) son producto de la diversidad interna del grupo, sin evaluar los taxones externos.
La confluencia de la problemática planteada y los resultados del análisis filogenético llevado a cabo permite proponer una nueva clasificación de los gliptodontes en la que conviven algunos de los grupos propuestos por Hoffstetter (1958) cuya monofilia no fue evaluada y varios de los clados obtenidos en este estudio. El no reconocimiento formal de los Nodos D, E y F se basó en los bajos soportes de los mismos, por lo que es posible que nueva evidencia contradiga estos agrupamientos y por lo tanto modificar la topología del árbol. Las tribus no evaluadas fueron posicionadas como incertae sedis en los Glyptodontia así como los géneros no evaluados pero cuyas tribus resultaron parafiléticas [apéndice IV]. En tal sentido, resulta importante recordar las observaciones Simpson (1945:95) "Later work on glyptodont phylogeny has been mostly by Castellanos and this has been followed in a general way, although scaled differently and expressed in slightly different terms for uniformity. The tribal subdivision of the Hoplophorinae [=Sclerocalyptinae] would be convenient and is suggested by his work, but at present it would be surely valid for only a few genera and would leave most incertae sedis or classified on ground more theoretical than subjective (sic) [objective]".
Paleobiogeografía y cronología. La confrontación de los resultados filogenéticos obtenidos con la distribución temporal de los gliptodontes estudiados permite discutir la calidad del registro fósil del grupo y proponer algunas consideraciones paleobiogeográficas generales. Por tal motivo, se incluyó la primera aparición de los grupos (v. gr. tribu o subfamilia) representados, mientras que se empleó la distribución temporal del género, cuando fue rechazada la monofilia de la tribu que lo contenía. De esta forma y teniendo en cuenta que los grupos hermanos deben aparecer simultáneamente (nodos internos), se puede inferir que la edad mínima de la primera dicotomía encontrada se remontaría a la Edad Mamífero Colhuehuapense (ca. 20 Ma, Mioceno temprano, véase Flynn y Swisher 1995), primer registro del género Propalaehoplophorus, ya que el de Eucinepeltus es más reciente. Cabe destacar que la edad mínima podría ser más antigua, dada la presencia de un registro subfamiliar indeterminado (Marshall et al., 1983) en el Deseadense de Patagonia (ca. 25 Ma, Oligoceno tardío). Con excepción de la tribu Panochthini (Piso/Edad Montehermosense) y Neosclerocalyptus (Piso/Edad Marplatense), los restantes "Hoplophorinae" Stromaphorus, Hoplophractus, Eosclerocalyptus, Plohophorus, Urotherium y Pseudoplohophorus tiene su primer registro en Piso/Edad Huayqueriense y los géneros Doedicurus y Glyptodon aparecen en el Piso/Edad Ensenadense, aunque a nivel subfamiliar los primeros registros se remontan al Piso/Edad Huayqueriense y Chapadmalalense, respectivamente. Si la posición estratigráfica que reconociera Cabrera (1944) del género Glyptodontidium fuera correcta, la subfamilia Glyptodontinae tendría su primer registro en el Piso/Edad Chasiquense y por lo tanto el origen del clado (Nodo C) debería ubicarse al menos en esos tiempos. Esta incertidumbre, sumada a la escasa y fragmentaria presencia de ejemplares fósiles asignables al lapso temporal que va desde el Colhuehuapense al Chasiquense/Huayqueriense impide el planteo de hipótesis paleobiogeográficas robustas. Dicho lapso temporal, abarca aproximadamente 10 Ma, por lo que es dable esperar la aparición ya sea de nuevas formas o de taxones conocidos pero aún no registrados en esos tiempos.
Conclusiones
Las conclusiones de este trabajo dan cuenta de distintos aspectos sobre la sistemática y evolución de los gliptodontes. Por un lado, se corroboró la monofilia del grupo, en el que se incluyeron representantes de cuatro de los cinco agrupamientos reconocidos históricamente: Propalaehoplophorinae, Hoplophorinae, Doedicurinae y Glyptodontinae. La falta de registro craneano de los Glyptatelinae imposibilitó la evaluación filogenética del mismo, aunque por las características postcraneanas hasta hoy descritas su posición en los cingulados es incierta.
En lo que respecta a las relaciones internas, los propalaehoploforinos considerados clásicamente como un grupo parafilético se resolvieron como un grupo natural, hermano de los restantes gliptodontes, mientras que los grupos previamente considerados naturales se resolvieron en forma parafilética. Entre estos últimos se incluyen las tribus Hoplophorini (=Sclerocalyptini sensu Hoffstetter, 1958) y Plohophorini, incluidas en la subfamilia Hoplophorinae (= Sclerocalyptinae sensu Hoffstetter, 1958), taxón que también se presenta parafilético. Por otra parte, se destaca la monofilia de grupos hasta hoy no propuestos como Neosclerocalyptus + Panochthus, clado que presenta el mayor soporte interno del cladograma, y Doedicurus + Glyptodon.
Por otra parte, se propone una nueva clasificación de los gliptodontes que incluye aquellos agrupamientos de buen soporte obtenidos en este trabajo y grupos cuya monofilia no ha sido evaluada en el mismo. Cabe remarcar que aun cuando no se estudiaron las mismas estructuras anatómicas y se empleó un marco metodológico distinto, es posible modificar la clasificación vigente tomada de Hoffstetter (1958) sin mediar análisis filogenético alguno con sólo valorizar diferencialmente los caracteres que sustentan varios de los agrupamientos clásicos.
Por último, la mixtura de la información filogenética con el registro estratigráfico permitió ubicar la primera dicotomía de los gliptodontes en tiempos colhuehuapenses. Los aproximadamente 10 Ma. que separan esta dicotomía de la siguiente (NODO C) permite postular la aparición de nuevas formas, posiblemente basales, en yacimientos correspondientes a ese lapso o de taxones incluidos en dicho clado (NODO C), que permitirían completar el registro fósil y así discutir distintas hipótesis paleobiogeograficas.
Apéndice I. Acrónimos y lista de ejemplares utilizados con su número de catálogo / acronyms and list of the specimens used with their catalogue numbers.
FC-DPV: Departamento de Paleontología de la Facultad de Ciencias, Universidad de la República, Uruguay. MACN-Pv: Colección Nacional de Paleovertebrados del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". MACN-A: Colección Nacional Ameghino del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". MACN-Ma: Colección Nacional de Mastozoología del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". MHD-P: Museo Histórico Departamental de Artigas. Paleontología, Uruguay. MLP: colección de la División Paleontología de Vertebrados, Facultad de Ciencias Naturales y Museo de la Plata.
Dasypus novemcintus: MACN-Ma: 49.391, 49.397, 49.350, 49.383, 50.123, 50.124, 39.461. Euphractus sexcinctus: MACN-Ma: 45.30, 50.121, 11.24, 20.3, 20.4, 14317, 34.592, 34.714, 47.204, 299. Eutatus seguini: MLP: 69-VIII-22-4, 69-VIII-1-4, 69-VIII-1-5, 69-VIII-1-8, MACN-Pv: 985, 1982, 10177. Pampatherium typum: MACN-Pv 11543/4, 11544, 11474, 11522; MLP-81-X-30-1. Pampatherium humboldtii: MHD-P-28. Propalaehoplophorus australis: MLP 16-15. Propalaehoplophorus minus: MACN A-4757. Propalaehoplophorus sp: MACN A-4754, MACN A-7655, MACN A-7656. Eucinepeltus pete-satus: MACN-A-4758, (holotipo), MACN-A-4760. Eucinepeltus sp.: MLP-84-III-1-1. Eosclerocalyptus planus: MACN-Pv 4853. Neosclerocalyptus sp.: MACN-Pv 8773, 15151, 18107, 16-28. Hoplophractus tapinocephalus: MLP-37-III-7-7 (Holotipo). Panochthus sp.: MLP P-1 (numeración antigua), MLP 16 - 38. Panochthus intermedius: MLP 16 - 37. Panochthus tuberculatus: MLP 16 - 29. Plohophorus figuratus: MLP 16-153 (holotipo). Stromaphorus compressidens: MLP-29-X-8-1. Pseudoplohophorus absolutus: FC-DPV-475 y 595 (holotipo). Urotherium antiquus: MACN A-229 - 31. Doedicurus sp.: MLP 16-24, MACN-Pv 12572, MACN-Pv 15153, MACN-Pv 2757, MACN-Pv 2762. Glyptodon reticulatus: MACN-Pv 10153, MACN-Pv 1780. Glyptodon muñizi: MACN-Pv 8706. Glyptodon sp.: MACN-Pv 2908, MACN-Pv 15152
Apéndice II. Listado de carateres y estados empleados en este estudio. Caracteres marcados con "+" son multiestado ordenado. La asignación de estado de los caracteres 28, 45, 62, 80, 82 y 83 del género Urotherium se basó en Ameghino (1889), del carácter 45 de Eosclerocalyptus en Ameghino (1919) y de los caracteres 28, 61, 82 y 83 de Hoplophractus en Cabrera (1939) / list of characters and states used in this study. Characters marked with "+" are multistate and orderer. The assignation of states of the characters 28, 45, 62, 80, 82 and 83 of Urotherium was taken of Ameghino (1889), of the character 45 of Eosclerocalyptus was taken of Ameghino (1919) and of the characters 28, 61, 82 y 83 of Hoplophractus was taken of Cabrera (1939).
Carácter 0. + Relación entre la porción desdentada anterior de la mandíbula con respecto a la longitud total de la misma: porción desdentada > al 25 % (0); < al 25% y > al 10% (1); < al 10% (2).
Carácter 1. + Distancia entre el borde posterior de la mandíbula con respecto al borde posterior del último molariforme mandibular (LPD), medida en unidades del último molariforma: > a 10 unidades (0); > a 3,5 y < 10 (1); > a 2,5 y < a 3,5 (2); < 2,5 (3).
Carácter 2. + Número de molariformes ocultos parcial o totalmente por la rama ascendente: ninguno (0); uno (1); dos (2); tres (3).
Carácter 3. + Distancia entre el borde anterior del cóndilo articular de la mandíbula con respecto a la hilera dentaria, medida en unidades del último molariforme: > a 10 unidades (0); < 10 y > a 2 (1); < a 2 (2).
Carácter 4. Posición del borde posterior de la sínfisis mandibular con respecto al primer molariforme mandibular: anterior (0); posterior (1).
Carácter 5. Angulo entre la rama horizontal y la rama ascendente: < a 90º (0); > a 90º (1).
Carácter 6. Cresta lateral del proceso coronoides: ausente (0); presente (1). Carácter 21 de Gaudin y Wible (2006).
Carácter 7. + Posición relativa entre los procesos coronoideo, condilar y angular: condilar próximo al angular (0); condilar equidistante (1); condilar próximo al coronoideo (2). Carácter 22 de Gaudin y Wible (2006).
Carácter 8. Posición del borde posterior de la mandíbula con respecto al cóndilo mandibular medida en unidades lateromediales del cóndilo articular: < a 1 (0); > a (1).
Carácter 9. Forma de la porción posterodorsal de la región angular: fuertemente cóncava, rematando en punta (0); muy suavemente cóncava o convexa (1).
Carácter 10. + Relación entre la máxima altura de la rama horizontal de la mandíbula y la longitud anteroposterior de la misma: máxima altura < al 10% de la longitud anteroposterior de la mandíbula (0); > al 10% y < 20% (1); > al 20% (2). Carácter 16 de Gaudin y Wible (2006).
Carácter 11. Posición del foramen mandibular con respecto al borde dorsal del último alvéolo mandibular: en línea o dorsal (0); ventral (1).
Carácter 12. + Distancia entre el foramen mandibular y el borde posterior del último alvéolo, medida en longitudes del último molariforme: > a 4 (0); < a 4 y > a 1 (1); < a 1 (2).
Carácter 13. Orientación del cóndilo mandibular en vista lateral: dorsal (0); anterodorsal (1).
Carácter 14. + Proyección medial de la sínfisis mandibular: ausente o rudimentaria (0); presente en su mitad ventral (1); presente en toda su extensión (2).
Carácter 15. Proyección ventromedial de la región angular: ausente (0); presente (1).
Carácter 16. Marcada crenulación en la superficie externa de la región angular: ausente (0); presente (1).
Carácter 17. + Relación entre la base y la altura del proceso coronoide: base < al 70% de la altura (0); > al 70% y < al 100% (1); > al 100% (2).
Carácter 18. + Número de dientes mandibulares: ocho (0); nueve (1); diez o más (2).
Carácter 19. Longitud que representa la hilera dentaria inferior con respecto al largo total de la mandíbula: < al 50% (0); > al 50% (1).
Carácter 20. + Forma de los últimos molariformes de la mandíbula: circulares o elípticos (0); trilobulados con el primer lóbulo circular o elíptico (1); trilobulados con el primer lóbulo triangular (2).
Carácter 21. Dientes premaxilares: ausente (0); presente (1).
Carácter 22. + Número de dientes maxilares: siete (0); ocho (1); nueve (2).
Carácter 23. + Superficie oclusal de la hilera dentaria superior: en V con el ápice dorsal, menos el último o los dos últimos molariformes (0); primeros molariformes oblicua, últimos planos (1); todas las superficies dentarias planas (2).
Carácter 24. Ramificaciones secundarias de osteodentina: ausente (0); presente (1).
Carácter 25. Forma del primer molariforme maxilar: eje antero-posterior > mayor al eje lateromedial (0); eje anteroposterior < al eje lateromedial (1).
Carácter 26. + Forma de los molariformes del maxilar: todos elípticos (0); los cuatro posteriores trilobulados (1); los cinco posteriores trilobulados (2); los seis posteriores trilobulados (3); los siete posteriores o todos trilobulados (4)
Carácter 27. + Relación entre el ancho máximo de la apertura nasal anterior y los últimos molariformes: < a la longitud interna entre los molariformes (0); > a la longitud interna y < a la longitud externa (1); > mayor a la longitud externa (2).
Carácter 28. + Relación entre el ancho máximo de la apertura nasal anterior y la longitud extracondilar: apertura nasal anterior < menor al 50% de la longitud extracondilar (0); > al 50% y < al 100% (1); > al 100% (2).
Carácter 29. Forma de la apertura nasal posterior: "U" (0); "V" (1); forma de "cerradura" (2).
Carácter 30. + Relación alto/ancho de la apertura nasal posterior: < a 0,75 [ancho representa más del 50% y menos del 75% del alto] (0); > a 0,75 y < a 1,1 [ancho mayor al 75% y menor] (1); > a 1,1 (2).
Carácter 31. Posición del borde anterior de la lámina escamosa con respecto al borde posterior de la fisura esfenorbital: posterior (0); en línea o anterior (1).
Carácter 32. Exposición del etmoides en la pared orbital: presente (0); ausente (1).
Carácter 33. Foramen maxilar en vista lateral: completamente visible (0); levemente o no visible (1).
Carácter 34. Foramen maxilar en vista ventral: visible (0); no visible (1).
Carácter 35. Posición del foramen oval con respecto a la cavidad glenoidea: en línea (0); posterior (1).
Carácter 36. Foramen transverso: presente (0); ausente (1).
Carácter 37. Posición del foramen oval con respecto al foramen maxilar: en línea (0); dorsal (1).
Carácter 38. Posición de los bordes ventrales de los forámenes maxilar y suborbitario: foramen maxilar ventral o en línea (0); foramen maxilar dorsal (1).
Carácter 39. + Posición de la pared medial del foramen suborbitario con respecto al borde labial de la hilera dentaria: pared en línea o medial a la hilera dentaria (0); pared levemente lateral (1); lateral (2).
Carácter 40. Posición relativa de los bordes externos de los forámenes suborbitario y maxilar: foramen suborbitario medial (0); foramen suborbitario en línea o lateral (1).
Carácter 41. Posición del foramen lagrimal con respecto al foramen suborbitario: foramen lagrimal lateral (0); en línea o medial a la pared externa de foramen suborbitario (1).
Carácter 42. Foramen suborbitario en vista lateral: visible (0); no visible (1).
Carácter 43. Posición del foramen suborbitario con respecto a la hilera dentaria del maxilar, medido como número de molariformes posteriores a este foramen: < a cinco (0); > a cinco (1)
Carácter 44. Posición del foramen suborbitario con respecto al borde ventral de la órbita: dorsal o levemente ventral (0); ventral por dos o más forámenes (1).
Carácter 45. + Desarrollo del proceso cigomático del maxilar con respecto a la longitud total de la hilera dentaria: proceso cigomático representa menos del 60% de la hilera dentaria (0); proceso cigomático representa menos del 60% de la hilera dentaria (1); proceso cigomático representa más del 60% y menos del 80% de la hilera dentaria (2); proceso cigomático representa más del 80% de la hilera dentaria (3).
Carácter 46. + Relación entre las caras internas de la apófisis descendente del arco cigomático: superficie externa menor (0); superficies iguales (1); superficie externa mayor (2).
Carácter 47. Forma del paladar en vista lateral: recto o cóncavo (0); sigmoideo (1).
Carácter 48. Desarrollo del paladar posterior a los forámenes incisivos: plano o suavemente elevado (0); fuertemente elevado cubriendo parcialmente los forámenes incisivos (1).
Carácter 49. + Posición del borde anterior del primer alvéolo del maxilar con respecto al plano posterior de los forámenes incisivos: posterior por una longitud mayor a una vez y media el ancho del premaxilar en el paladar (0); posterior por una longitud mayor a la mitad del ancho del premaxilar en el paladar y menor a una vez y media del mismo ancho (1); en línea o posterior por menos de la mitad del ancho del premaxilar en el paladar (2); claramente anterior (3).
Carácter 50. + Posición del plano anterior de la apertura nasal posterior con respecto a la hilera dentaria, medido sobre la máxima longitud anteroposterior del último molariforme del maxilar: posterior por más de dos longitudes (0), posterior por más de una longitud (1) anterior o posterior por menos de una longitud (2).
Carácter 51. + Posición de los forámenes vasculares anteriores del paladar con respecto al borde anterior de la hilera dentaria del maxilar: anterior al borde posterior del primer molariforme (0); posterior al borde posterior del primer molariforme (1).
Carácter 52. Fuerte desarrollo dorsoventral de la porción palatal del premaxilar: ausente (0); presente (1).
Carácter 53. Inclinación del borde anterior de los premaxilares: no alcanza la hilera dentaria del maxilar (0); alcanza la hilera dentaria del maxilar (1).
Carácter 54. + Posición del plano anterior de los nasales con respecto al plano anteroventral de los premaxilares: anterior (0); en línea (1); posterior (2).
Carácter 55. + Posición del borde anterior de los nasales con respecto al foramen lagrimal y el borde ventral de la órbita: dorsal al foramen lagrimal (0); entre el foramen lagrimal y el borde ventral de la órbita (1); ventral al borde ventral de la órbita (2).
Carácter 56. Posición del plano posterior de los nasales con respecto a los forámenes lagrimales: anterior (0); posterior (1).
Carácter 57. Posición del tabique nasal óseo con respecto al borde anterior de los nasales: a nivel o posterior (0); anterior (1).
Carácter 58. + Relación entre la longitud del rostro y el ancho entre los forámenes lagrimales: > a 1,5 (0); > a 1 y < a 1,5 (1); < a 1 (2).
Carácter 59. Posición del borde ventral del pterigoide con respecto al último molariforme: medial al borde labial del último molariforme (0); en línea o lateral al borde labial del último molariforme (1). Carácter 59 de Gaudin y Wible (2006).
Carácter 60. Posición que alcanza la porción rugosa del pterigoides o del pterigoides y aliesfenoides: no alcanza el último molariforme (0); alcanza el último molariforme (1). Carácter 61. + Distancia entre las cicatrices de la inserción muscular del rectus capitis: ampliamente separdos (0); levemente separados (1); en contacto con una cresta medial (2).
Carácter 62. + Posición de la cavidad glenoidea con respeto al cóndilo occipital: dorsal (0); en línea (1); ventral (2).
Carácter 63. Relación entre las longitudes lateromedial/antero-posterior de la cavidad glenoidea: < a 2 (0); > a 2 (1).
Carácter 64. + Posición de la cavidad glenoidea con respecto a la hilera dentaria: posterior por más de ocho molariformes (0); posterior por más de dos molariformes (1); posterior por más de un molariforme (2); posterior por menos de un molariforme (3).
Carácter 65. Posición del borde posterior del yugal con respecto a la cavidad glenoidea: a nivel (0); claramente anterior (1).
Carácter 66. Forma de la sutura yugal-escamoso en vista lateral: recta (0); forma de L (1); forma de S (2).
Carácter 67. Dirección del eje principal de la sutura yugal-escamoso: anteroventral o ventral (0); posteroventral (1).
Carácter 68. Posición del plano lateral del proceso cigomático del escamoso con respecto al plano lateral del lagrimal: en línea o medial (0); lateral (1).
Carácter 69. Superposción dorsal del yugal al escamoso en vista lateral: ausente (0); presente (1).
Carácter 70. Cresta sagital: ausente (0); presente (1).
Carácter 71. + Lámina frontal descendente: ausente (0); presente, no desarrollada ventralmente (1); presente, desarrollada ventralmente (2).
Carácter 72. Dirección de la lámina frontal descendente: < a 45° grados (0); > a 45° (1).
Carácter 73. Posición del plano más posterior del frontal con respecto a la cavidad glenoidea: anterior o en línea (0); posterior (1).
Carácter 74. + Posición del foramen lagrimal con respecto al cóndilo occipital: dorsal (0); en línea (1); ventral (2).
Carácter 75. + Posición dorsoventral del foramen lagrimal: próximo al paladar (0); equidistante (1); próximo a la tabla craneana (2).
Carácter 76. Posición del foramen lagrimal con respecto al borde anterior de la órbita: extraorbital (0); intraorbital (1).
Carácter 77. + Superficie dorsal al foramen lagrimal: lisa (0); irregular, fuertemente punteda (1); cónica de fuerte desarrollo (2).
Carácter 78. + Desarrollo del proceso yugal del frontal: ausente o rudimentario (0); bien desarrollado (1); fuertemente desarrollado (2).
Carácter 79. Forma de la órbita: rectangular (0); U (1); circular (2).
Carácter 80. Relación entre las longitudes lateromedial y dorso-ventral del cóndilo occipital: lateromedial igual o mayor (0); lateromedial menor (1).
Carácter 81. + Angulo entre el plano occipital y el plano horizontal: > a 80º (0); < a 80º y > a 65º (1); < a 65º y > a 45º (2); < a 45º (3).
Carácter 82. Relación entre el ancho mínimo postorbital con respecto al ancho extracondilar: < a 1,2 (0); > 1,2 (1).
Carácter 83. + Cresta occipital: rudimentaria o no pasa el tercio medio del occipital (0); alcanza el tercio ventral occipital (1).
Apéndice III. Matriz de datos. Los símbolos "?" y "N" representa entrada faltante y elemento no comparable respectivamente. Los polimorfismos se indican como "A" = [0,1]; "B" = [1,2]; "C" = [2,3] / data matrix. The symbols "?" and "N" represents missing data and inapplicable character respectively. The polymorphism are indicated as "A" = [0,1]; "B" = [1,2]; "C" = [2,3]

Apéndice IV. Clasificación de los gliptodontes / classification of the glyptodonts.
Magnorden Xenarthra Cope, 1889
Orden Cingulata Illiger 1811
Suborden Glyptodontia Ameghino,1889 [Nodo A]
†Asymmetrura Fariña, 1981
†Berthawyleria Castellanos, 1940
†Chacus Zurita, 2002
†Coscinocercus Cabrera, 1939
†Eonaucum Scillato-Yané y Carlini, 1998
†Eosclerophorus Castellanos, 1948
†Hoplophorus Lund, 1839
†Isolinia Castellanos, 1951b
†Parahoplophorus, Castellanos, 1931
†Phlyctaenopyga Cabrera, 1944
†Plohophoroides Castellanos, 1928
†Plohophorops Castellanos, 1935
†Stromaphoropsis Kraglievich, 1932
†Teisseiria Kraglievich, 1932
†Trabalia Kraglievich, 1932
†Trachycalyptus, Ameghino, 1908
†Uruguayurus Mones, 1987
†Zaphilus Ameghino, 1889
Tribu Neothoracophorini Castellanos, 1951
†Pseudoneothoracophorus Castellanos, 1951
†Neothoracophorus Ameghino, 1889
Tribu Palaehoplophorini Hoffstetter, 1958
†Palaehoplophoroides Scillato-Yané y Carlini, 1998
†Palaehoplophorus Ameghino, 1883
†Protoglyptodon Ameghino, 1885
†Aspidocalyptus Cabrera, 1939
†Pseudoeuryurus Ameghino, 1889
†Chlamyphractus Castellanos, 1940
Tribu Neuryurini Hoffstetter, 1958
†Neuryurus Ameghino, 1889
Familia Glyptatelidae Castellanos, 1932
†Glyptatelus Ameghino, 1897
†Clypeotherium Scillato-Yané 1977
†Neoglyptatelus Carlini, Vizcaíno y Scillato-Yané,1997
†Lomaphorelus Ameghino, 1902
†Pachyarmatherium Downing y White, 1995
Infraorden Propalaehoplophoroinei Ameghino, 1891a [Nodo B]
Superfamilia Propalaehoplophoroidea Ameghino, 1891a
Familia Propalaehoplophoridae Ameghino, 1891a
†Propalaehoplophorus Ameghino, 1887a
†Eucinepeltus Ameghino, 1891
†Asterostemma Ameghino, 1889
†Metopotoxus Ameghino, 1898
†Cochlops Ameghino, 1889
Infraorden Glyptodontoinei Gray, 1869 [Nodo C]
†Eosclerocalyptus Ameghino, 1919
†Hoplophractus Cabrera, 1939
†Stromaphorus Castellanos, 1926
†Pseudoplohophorus Castellanos, 1926
Superfamilia Glyptodontoidea Gray, 1869 [Nodo G]
Tribu Lomaphorini Hoffstetter, 1958
†Urotherium Castellanos, 1926
†Lomaphorops Castellanos, 1931
†Lomaphorus Ameghino, 1889
†Peiranoa Castellanos, 1946
†Thachycalyptoides Saint-André, 1996
Familia Panochthidae Castellanos 1927 [Nodo J]
Subfamilia Panochthinae Castellanos, 1927
Tribu Neosclerocalyptini Paula Couto,1957
†Neosclerocalyptus Paula Couto, 1957
Tribu Panochtini Castellanos, 1927
†PanochthusBurmeister, 1866
†Nopachtus Ameghino, 1888
†Propanochthus Castellanos, 1925
Familia Glyptodontidae Gray, 1869 [Nodo H]
Subfamilia Plohophorinae Castellanos, 1932
Tribu Plohophorini Castellanos, 1932
†Plohophorus Ameghino, 1887b
Subfamilia Glyptodontinae Gray, 1869 [Nodo I]
Tribu Doedicurini Ameghino, 1889
†DoedicurusBurmeister, 1874
†Comaphorus Ameghino, 1886
†Eleutherocercus Koken, 1888
†Palaeodaedicurus Castellanos, 1927
†Prodaedicurus Castellanos, 1927
†Castellanosia Kraglievich, 1932
†Xiphuroides Castellanos, 1927
†Plaxaplous Ameghino, 1884b
†Doedicuroides Castellanos, 1940
Tribu Glyptodontini Gray, 1869
†Glyptodon Owen, 1839
†Glyptodontidium Cabrera, 1944
†Stromatherium Castellanos, 1953
†Paraglyptodon, Castellanos, 1932
†Glyptostracon Castellanos, 1953
†Heteroglyptodon Roselli, 1976
†Glyptotherium Osborn, 1903
Agradecimientos
A M.S. Bargo y M. Reguero del Museo de La Plata, O. Vaccaro, A. Kramarz y J. Bonaparte del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", a M. Ubilla, D. Perea y M. Clara de la Facultad de Ciencia de Uruguay, a J. Soloví de Artigas, Uruguay, por facilitarme el acceso a las colecciones allí depositadas. A R. Fariña y S.F. Vizcaíno, quienes siempre me acompañaron en el estudio de los gliptodontes. A los uruguayos, quienes a través de la Universidad Pública y Gratuita, me dieron la posibilidad de llevar adelante mis estudios de postgrado. Este trabajo es una contribución a los proyectos UNLP N474, PIP-CONICET 5240 y PICT 26219.
Bibliografía
1. Ameghino, C. 1919. Sobre mamíferos fósiles del piso araucanense de Catamarca y Tucumán. 1º Reunión Nacional de la Sociedad Argentina de Ciencias Naturales 150-153. [ Links ]
2. Ameghino, F. 1883. Sobre una colección de mamíferos fósiles del piso mesopotamiense de la formación patagónica, recogidos por el Prof. Pedro Scalabrini. Boletín de la Academia Nacional de Ciencias (Córdoba) 5: 101-116. [ Links ]
3. Ameghino, F. 1884a. Filogenia. Principios de clasificación transformista basados sobre leyes naturales y proporciones matemáticas, pp 390. [ Links ]
4. Ameghino, F. 1884b. Excursiones geológicas y paleontológicas en la provincia de Buenos Aires. Boletín de la Academia Nacional de Ciencias (Córdoba) 6: 161-257. [ Links ]
5. Ameghino, F. 1885. Nuevos restos de mamíferos fósiles oligocenos recogidos por el Profesor Pedro Scalabrini y pertenecientes al Museo Provincial de la ciudad de Paraná. Boletín de la Academia Nacional de Ciencias (Córdoba) 8: 5-207. [ Links ]
6. Ameghino, F. 1886. Contribuciones al conocimiento de los mamíferos fósiles de los terrenos terciarios antiguos del Paraná. Boletín de la Academia Nacional de Ciencias (Córdoba) 9: 5-228. [ Links ]
7. Ameghino, F. 1887a. Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de Patagonia Austral y depositados en el Museo de La Plata. Boletín del Museo de La Plata 1: 1-26. [ Links ]
8. Ameghino, F. 1887b. Apuntes preliminares sobre algunos mamíferos extinguidos del yacimiento Monte Hermoso. Boletín del Museo de La Plata 1: 1-20. [ Links ]
9. Ameghino, F. 1888. Ameghino, F. 1888. Rápidas diagnosis de algunos mamíferos fósiles nuevos de la República Argentina. Editorial Coni, Buenos Aires pp 17. [ Links ]
10. Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias (Córdoba) 6: 1-1027. [ Links ]
11. Ameghino, F. 1891a. Mamíferos y aves fósiles argentinas. Especies nuevas adiciones y correcciones. Revista Argentina de Historia Natural 1: 240-259. [ Links ]
12. Ameghino, F. 1891b. Nuevos restos de mamíferos fósiles descubiertos por Carlos Ameghino en el Eoceno Inferior de la Patagonia Austral. Especies nuevas: adiciones y correcciones. Revista Argentina de Historia Natural 1: 289-328. [ Links ]
13. Ameghino, F. 1895. Sur les édentés fosiles de l´Argentine (examen critique, revisión et correction de l´ouvrage de M. R. Lydekker "The extinct Edentates of Argentina"). Revista del Jardín Zoológico de Buenos Aires 3: 97-192. [ Links ]
14. Ameghino, F. 1897. Mammiféres crétacés de l´Argentine. Boletín del Instituto Geográfico Argentino 18: 406-521. [ Links ]
15. Ameghino, F. 1898. Primera sinopsis geológico-paleontológica. 2º Censo de la República Argentina 1: 111-255. [ Links ]
16. Ameghino, F. 1902. Notices préliminaires sur des mammifères nouveaux des terrains crétacés de Patagonie. Boletín de la Academia Nacional de Ciencias (Córdoba) 17: 5-70. [ Links ]
17. Ameghino, F. 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadmalán. Anales del Museo Nacional de Historia Natural 10: 343-428. [ Links ]
18. Bremmer, K. 1994. Branch support and tree stability. Cladistics 10 (3): 295-304. [ Links ]
19. Burmeister, G. 1866. Lista de los mamíferos fósiles del terreno diluviano. Anales del Museo Público de Buenos Aires 1: 121-232. [ Links ]
20. Burmeister, G. 1870-1874. Monografía de los glyptodontes en el Museo Público de Buenos Aires. Anales del Museo Público de Buenos Aires 2: 1-412. [ Links ]
21. Cabrera, A. 1939. Sobre vertebrados fósiles del Plioceno de Adolfo Alsina. Revista del Museo de La Plata (nueva serie) sección Paleontología 2: 3-35. [ Links ]
22. Cabrera, A. 1944. Los gliptodontoideos del araucaniano de Catamarca. Revista del Museo de La Plata (nueva serie) sección paleontología 3: 5-76. [ Links ]
23. Carlini, A.A., Vizcaíno, S.F. y Scillato-Yané, G.J. 1997. Armored xenarthrans: a unique taxonomic and ecologic assemblage. En R. F. Kay, R. L. Cifelli, J. J. Flynn, and R. H. Madden (eds.), Vertebrate Paleontology of the Miocene Honda Group, Republic of Colombia. Smithsonian Institution Press, Washington, pp. 213-226 [ Links ]
24. Castellanos, A. 1925. Breves notas sobre la evolución de la coraza y del extremo caudal de los Loricata (Hicanodonta). Revista El Círculo número extraordinario: 93-96. [ Links ]
25. Castellanos, A. 1926. Sobre un nuevo gliptodóntido chapadmalense, Urotherium simplex n. gen. et n. sp. y las formas afines. Anales del Museo Nacional de Historia Natural de Buenos Aires 34: 263-278. [ Links ]
26. Castellanos, A. 1927. Descripción de un fragmento de tubo caudal de un doedicurino en relación con sus géneros afines. Anales del Museo de Historia Natural de Montevideo 2: 265-300. [ Links ]
27. Castellanos, A. 1928. Un nuevo representante de la subfamilia Sclerocalyptinae: Plohophoroides n. gen. y sus especies. Revista de la Universidad Nacional de Córdoba 15: 110-148. [ Links ]
28. Castellanos, A. 1931. "La librería del Glyptodon" de Ameghino. Cultura, Órgano de la Biblioteca Popular Bernardino Rivadavia 3: 4-9. [ Links ]
29. Castellanos, A. 1932. Nuevos géneros de gliptodontes en relación con su filogenia. Physis 11: 92-100. [ Links ]
30. Castellanos, A. 1935. Contribución al estudio de la paleontología del Uruguay. Restos de Pseudoplohophorus orientalis Amegh. Altura (Organo de Asociación de Farmacias) 1: 5-15. [ Links ]
31. Castellanos, A. 1937. Anotaciones sobre la línea filogenética de los clamiterios. Publicaciones del Instituto de Fisiografía y Geología de la Facultad de Ciencias Matemáticas Físico-Químicas y Naturales aplicadas a la Industria de la Universidad Nacional del Litoral. Serie técnico-científica 1: 1-35. [ Links ]
32. Castellanos, A. 1940. A propósito de los géneros Plohophorus, Nopachthus y Panochthus. Publicaciones del Instituto de Fisiografía y Geología de la Facultad de Ciencias Matemáticas Físico-Químicas y Naturales aplicadas a la Industria de la Universidad Nacional del Litoral, Serie técnico-científica 1: 1-279. [ Links ]
33. Castellanos, A. 1942. A propósito de los géneros Plohophorus, Nopachthus y Panochthus. Publicaciones del Instituto de Fisiografía y Geología de la Facultad de Ciencias Matemáticas Físico-Químicas y Naturales aplicadas a la Industria de la Universidad Nacional del Litoral, Serie técnico-científica 3: 417-592. [ Links ]
34. Castellanos, A. 1946. Un nuevo gliptodontoideo del Araucanense del valle de Yocavil (Santa María) de la Provincia de Tucumán. Publicaciones del Instituto de Fisiografía y Geología 6: 1-19. [ Links ]
35. Castellanos, A. 1948. Um novo esclerocaliptino. Divisão de Gelogia e Mineralogia, Notas preliminares e Estudos 47: 1-13. [ Links ]
36. Castellanos, A. 1951. Acotaciones al género Neothoracophorus Ameghino. Revista de la Asociación Geológica Argentina 6: 63-82. [ Links ]
37. Castellanos, A. 1951b. Un nuevo género de esclerocaliptino (Isolinia) descubierto en el Araucaniano del Valle de Los Reartes (Sierras de Córdoba). Revista de la Asociación Geológica Argentina 6: 95-100. [ Links ]
38. Castellanos, A. 1953. Anotações e retificações ao gênero Glyptodon Owen e à subfamília Glyptodontinae. Anais da Academia Brasileira de Ciências 25: 391-410. [ Links ]
39. Castellanos, A. 1959. Trascendencia de la obra de Florentino Ameghino. Revista de la Facultad de Ciencias Naturales de Salta 1: 35-56. [ Links ]
40. Cione, A.L. y Tonni, E.P. 1995a. Chronostratigraphy and "Land-Mammal Ages" in the Cenozoic of Southern South America: principles, practices, and the "Uquian" problem. Journal of Paleontology 69: 135-159. [ Links ]
41. Cione, A.L. y Tonni, E.P. 1995b. Bioestratigrafía y cronología del Cenozoico superior de la región pampeana. En M.T Alberdi, G. Leone y E.P. Tonni (eds.), Evolución Biológica y Climática de la Región Pampeana Durante los Últimos Cinco Millones de Años. Un Ensayo de Correlación con el Mediterráneo Occidental. Museo Nacional de Ciencias Naturales (Madrid). Monografías 12: 47-74. [ Links ]
42. Cione, L. A., Tonni, E.P., Bond, M., Carlini A.A., Pardiñas, U.F., Scillato-Yané, G.J., Verzi, D. y Vucetich, M.G. 1999. Ocurrence charts of Pleistocene mammals in the Pampean area, eastern Argentina. En: J. Rabassa y M. Salemme (eds.), "Quaternary Vertebrate Paleontology in South America", Quaternary of South America and Antarctic Peninsula 12: 53 - 59. [ Links ]
43. Cione, A.L., Azpelicueta, M.M. Bond, M., Carlini, A.A., Casciotta, J.R., Cozzuol, M.A., De la Fuente, M., Gasparini, Z., Goin, F.J., Noriega, J., Scillato-Yané, G.J., Soibelzon, L., Tonni, E.P., Verzi, D. y Vucetich, M.G. 2000. Miocene vertebrates from Entre Ríos province, eastern Argentina. En: F.G. Aceñolaza y R. Herbst (eds.), El Neógeno de Argentina. INSUGEO, Serie Correlación Geológica 14: 191-237. [ Links ]
44. Cope, E.D. 1889. The Edentata of North America. American Naturalist 23: 657-664. [ Links ]
45. Downing, K. F., y White, R. S. 1995. The cingulates (Xenarthra) of the Leisey Shell Pit local fauna (Irvingtonian), Hillsborough County, Florida. Bulletin of the Florida Museum of Natural History 37: 375-396. [ Links ]
46. Engelmann, G. F. 1985. The phylogeny of the Xenarthra. En G. G. Montgomery (ed.), The Evolution and Ecology of Armadillos, Sloths and Vermilinguas Smithsonian Institution Press, Washington, DC. pp. 51-64. [ Links ]
47. Fariña, R.A. 1981. Anatomía comparada de los tubos caudales de Plohophorini (Mammamlia, Glyptodontidae), del Montehermosense del Uruguay. 2° Congresso Latino-Americano de Paleontología (Porto Alegre), Anais 2: 779-793. [ Links ]
48. Fariña, R. A. y Vizcaíno, S. F. 2001. Carved teeth and strange jaws: how glyptodonts masticated. En: S.F. Vizcaíno, R.A. Fariña y C. Janis. Biomechanics and Paleobiology of Vertebrates, Acta Paleontologica Polonica, Special Issue 46: 87-102. [ Links ]
49. Farris, J.S., Albert, V.A., Källersjö, M., Lipscomb, D. y Kluge, A.G. 1996. Parsimony jackknifing outperforms neighbor-joining. Cladistics 12: 99-124. [ Links ]
50. Fernicola, J. C. 2005. [Análisis filogenético de la familia Glyptodontidae Gray 1869. Tesis Doctoral, Facultad de Ciencias, Montevideo, Uruguay, 135 pp. Inédita.]. [ Links ]
51. Fernicola, J.C. 2006. Análisis filogenético de la subfamilia Propalaehoplophorinae (Xenarthra, Glyptodontidae). 22º Jornadas Argentinas de Paleontología de Vertebrados (San Juan), pp. 19. [ Links ]
52. Fernicola, J.C., Vizcaíno, S. F. y Fariña A. R. 2008. The evolution of armored xenarthrans and a phylogeny of the glyptodonts. En: J. Loughry y S.F. Vizcaíno (eds.), The Biology of the Xenarthra. University Press of Florida, pp. 79-85. [ Links ]
53. Flower, W. H. 1882. On the mutual affinities of the animals composing the order Edentata. Proceeding of the Zoological Society of London 358-367. [ Links ]
54. Flynn, J.J., and C.C. Swisher. 1995. Cenozoic South American land mammal ages: correlation to global geochronologies. Society for Sedimentary Geology Special Publication 54: 317-333. [ Links ]
55. Gaudin, T.J. y Wible, J.R. 2006. The phylogeny of living and extinct armadillos (Mammalia, Xenarthra, Cingulata): a cranio-dental analysis. En: M.T. Carrano, T.J. Gaudin, R.W. Blob y J. R. Wible (eds.), Amniote Paleobiology: Perspectives on the Evolution of Mammals, Birds and Reptiles, The University of Chicago Press, Chicago, IL, pp. 153-198. [ Links ]
56. Goloboff, P.A. 1993. NONA versión 2.0, programa y documentación distribuidos por el autor. San Miguel de Tucumán, Argentina. [ Links ]
57. Gray, J. E. 1869. Catalogue of carnivorous, pachydermatous and edentates Mammalia in the British Museum. British Museum of Natural History, pp 398. [ Links ]
58. Hoffstetter, R. 1958. Xenarthra. En: J. Piveteau (ed.), Traité de Paléontologie Masson & Cie. Paris 6: 535-636. [ Links ]
59. Illiger, J.K.W. 1811. Prodromus systemalis mammalium et avium additis terminis zoographicis utriusque classis. Berlin, 301 pp. [ Links ]
60. International Commission on Zoological Nomenclature. 1999. International Code of Zoological Nomenclature adopted by the 20º General Assembly of the International Union of Biological Science, 1-306. [ Links ]
61. Koken, E. 1888. Eleutherocercus ein neuer Glyptodont aus Uruguay. Abhandlungen der königlichen preussischen Akademie der Wissenscaften zu Berlin 1: 1-28. [ Links ]
62. Kraglievich, L. 1932. Nuevos apuntes para la Geología et Paleontología del Uruguay. Anales del Museo de Historia Natural de Montevideo 2: 1-65. [ Links ]
63. Lund, P.W. 1839. Blik paa Brasiliens dyreverden för sidste jordomvaeltning. Anden afhandling: Pattedyrene. Det kongelige Danske Videnskabernes selskabs naturvedenskabelige og mathematiske afhandlinger 8: 61-144 [ Links ]
64. Madden, R.H., Guerrero, J., Kay, R.F., Flynn, J.J., Swisher III, C.C. y Walton, A.H. 1997. The Laventan stage and age. En: R.F. Kay, R.L. Cifelli, J.J. Flynn and R.H. Madden (eds.), Vertebrate Paleontology of the Miocene Honda Group, Republic of Colombia. Smithsonian Institution Press, Washington DC, pp. 499-519. [ Links ]
65. Marshall, L.G. y Patterson, B. 1981. Geology and geochronology of the mammal-bearing Tertiary of the Valle de Santa María and Río Corral Quemado, Catamarca province, Argentina. Fieldiana Geology 9: 1-81. [ Links ]
66. Marshall, L.G., Hoffstetter, R. y Pascual, R. 1983. Mammals and Stratigraphy: Geochronology of the Continental Mammal-bearing Tertiary of South America. Paleovertebrata, Mémoire Extraordinaire: 1-93. [ Links ]
67. McKenna, M.C., y Bell, S.K. 1997. Classification of Mammals above the Species Level. Columbia University Press, New York, 631 pp. [ Links ]
68. Mones, A. 1987. Sobre tres especies de Gliptodontes del Uruguay: Cingulata: Glyptodontidae. 4º Congreso Latinoamericano de Paleontología (Bolivia), Memoria 1: 500-515. [ Links ]
69. Osborn, H.F. 1903. Glyptotherium texanum, a new Glyptodont, from the Lower Pleistocene of Texas. Bulletin of the American Museum of Natural History 19: 491-494. [ Links ]
70. Owen, R. 1839. Description of a Tooth and part of the skeleton of the Glyptodon clavipes, a large quadruped of the edentate order, to which belongs the tesselated bony armour described and figured by Mr. Clift in his memoir on the remains of the Megatherium, brougth to England by Sir Woodbine Parish, F.G.S. Proceeding of the Geological Society of London 3: 108-113. [ Links ]
71. Pascual, R., Ortega Hinojosa, E.J., Gondar, D. y Tonni, E.P. 1965. Las edades del Cenozoico mamalífero de la Argentina, con especial atención a aquellas del territorio bonaerense. Anales de la Comisión de Investigaciones Científicas de la Provincia de Buenos Aires 6: 165-193. [ Links ]
72. Pascual, R., Ortega Hinojosa, E.J., Gondar D. y Tonni. E.P. 1966. Las Edades del Cenozoico mamalífero de la Argentina con especial atención a aquellas del territorio bonaerense. En: A.V. Borello (ed.), Paleontografía Bonaerense, 4 Vertebrata. Comisión de Investigación Científica, Buenos Aires: 3-27. [ Links ]
73. Pascual, R., Ortiz Jaureguizar, E. y Prado, J.L. 1996. Land Mammals: Paradigm for Cenozoic South American Geobiotic Evolution. Münchner Geowissenschaftliche Abhandlungen (A) 30: 265-319. [ Links ]
74. Patterson, B. y Pascual, R. 1972. The Fossil Mammal Fauna of South America. En: A. Keast, F.C. Erk, y B.Glass (eds.), Evolution, Mammals, and Southern Continents, State University of New York Press, Albany, pp. 247-309. [ Links ]
75. Paula Couto, C. de. 1947. Contribuiçao para o estudo de Hoplophorus euphractus Lund, 1839. Summa Brasiliensis Geologiae 1: 33-53. [ Links ]
76. Paula Couto, C. de. 1957. Sôbre um gliptodonte do Brasil. Boletim Divisão de Geologia e Mineralogia 165: 1-37. [ Links ]
77. Paula Couto, C. de. 1979. Tratado de Paleomastozoologia. Academia Brasileira de Ciencias, Rio de Janeiro, 590 pp. [ Links ]
78. Perea, D. 2005. Pseudoplohophorus absolutus n. sp. (Xenarthra, Glyptodontidae), variabilidad en Sclerocalyptinae y redefinición de una biozona del Mioceno Superior de Uruguay. Ameghiniana 42: 175-190. [ Links ]
79.Roselli F.L. 1976. Contribución al estudio de la geopaleontología de los departamentos de Colonia y Soriano, (República Oriental del Uruguay). Imprenta Cooperativa, Montevideo, 175 pp. [ Links ]
80. Rovereto, C. 1914. Los estratos araucanos y sus fósiles. Anales del Museo Nacional de Historia Natural 25: 1-247. [ Links ]
81. Saint-André, P-A. 1996. Deux nouveaux édentés (Mammalia, Xenarthra) Trachycalyptoides achirense nov. gen. et nov. sp. (Glyptodontidae, Sclerocalyptinae) et Xyophorus villarroeli nov. sp. (Megatheriidae, Nothrotheriinae) du Huayquérien (Miocène supérieur) de l´Altiplano bolivien. Bulletin du Muséum National d`Histoire Naturelle cuarta serie 18: 79-106. [ Links ]
82. Scillato-Yané, G.J. 1976. Sobre un Dasypodidae (Mammalia, Xenarthra) de Edad Riochiquense (Paleoceno superior) de Itaboraí (Brasil). Anais Academia Brasileira do Ciencias 48: 527-530. [ Links ]
83. Scillato-Yané, G. J. 1977. Sur quelques Glyptodontidae nouveaux (Mammalia, Edentata) du Déséadien (Oligocène inférieur) de Patagonie (Argentine). Bulletin du Muséum National d`Histoire Naturelle 64: 249-260. [ Links ]
84. Scillato-Yané, G. J. y Carlini, A. A. 1998. Nuevos Xenarhra del Friasense (Mioceno Medio) de Argentina. Stvdia Geologica Salmantisencia 34: 43-67. [ Links ]
85. Scillato-Yané, G.J., Carlini, A.A., Viscaíno, S.F. y Ortiz Jaureguizar, E.O. 1995. Los Xenarthros. En: M.T Alberdi, G. Leone y E.P. Tonni (eds.), Evolución Biológica y Climática de la Región Pampeana Durante los Últimos Cinco Millones de Años. Un Ensayo de Correlación con el Mediterráneo Occidental. Museo Nacional de Ciencias Naturales (Madrid). Monografías 12: 183-210. [ Links ]
86. Scott, W.B. 1903-1904. Mammalia of the Santa Cruz Beds. Paleontology. Part I Edentata. En: W. B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896-1899. Princeton University, New Jersey. E. Schweizerbart'sche Verlagshandlung (E. Nägele), Stuttgart 5: 1-364. [ Links ]
87. Simpson, G.G. 1945.The principles of classification and a classification of Mammals. Bulletin af the American Museum of Natural History 85: 1-254. [ Links ]
88. Vizcaíno, S.F., Rinderknecht, A. y Czerwonogora, A. 2003. An enigmatic cingulata (Mammalia: Xenarthra) from the late Miocene of Uruguay. Journal of Vertebrate Paleontology 23: 981-983. [ Links ]
89. Zurita, A.E. 2002. Nuevo gliptodonte (Mammalia: Glyptodontoidea) del Cuaternario de la provincia del Chaco, Argentina. Ameghiniana 39: 175-182. [ Links ]
Recibido: 30 de enero de 2007.
Aceptado: 10 de junio 2008.

