










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.11 n.2 Mendoza jul./dic. 2004
Adaptaciones morfológicas de la lengua de Desmodus Rotundus Rotundus (Chiroptera: Phyllostomidae) en función de la alimentación
Marcos I. Mollerach1 y Susana Mangione2
1Facultad de Ciencias Naturales e Instituto Miguel Lillo, y PIDBA (Programa de Investigaciones de Biodiversidad Argentina), Universidad Nacional de Tucumán, Argentina. 2Fundación Miguel Lillo, Miguel Lillo 255, 4000, Tucumán, Argentina.
ASBTRACT: Morphological adaptations of the tongue of Desmodus rotundus (Chiroptera: Phyllostomidae) related with their feeding habits. An analysis of the external morphology and histomorphology of the tongue, as well as an histochemical analysis of the glands of Desmodus rotundus, are here presented with the purpose of establishing a correlation between these structures and the feeding habits of this species. In the lingual surface they have the typical filiform papillae modified to different degrees of keratinization, and there are fungiform-like papillae in less amount. The intrinsic musculature and the presence of lynphocitary infiltration are described; the presence of important nervous packages was observed.
Key words. Desmodus rotundus. Feeding habits. Histology. Tongue.
INTRODUCCIÓN
Desmodus rotundus es una especie que se alimenta principalmente de sangre (Greenhall et al., 1983). En estudios de contenido estomacal se ha observado que un 73.4% se compone de sangre de mamíferos, 10.1% de sangre de aves, y un 16.5% de leche (Villa-R. et al., 1969). Sus hábitos son gregarios y forman colonias de hasta 5000 individuos (Crespo et al., 1961). Habitan en cavernas o cuevas y utilizan otros refugios temporarios durante la digestión (Greenhall et al., 1983). Esta especie es de importancia económico-sanitaria ya que puede transmitir enfermedades como rabia paralítica y trypanosomiasis (Barquez et al., 1999). Su principal fuente de alimento es la sangre de animales domésticos, grandes mamíferos nativos y, eventualmente, humanos (Arata et al., 1967). Su distribución se extiende desde el norte de México, hasta el sur de Argentina, y se conocen pocos registros en la región Amazónica (Barquez et al., 1999). Como la gran mayoría de los murciélagos, son de hábitos nocturnos o vespertinos y de reposo diurno (Crespo et al., 1961).
La musculatura lingual intrínseca en mamíferos está muy desarrollada. Estos músculos son el genio-glossus y el hyo-glossus, derivados durante la ontogenia a partir del genio-hyoideus (Edgeworth, 1935). Esta musculatura adquiere características propias de acuerdo con los hábitos de cada grupo en particular. En la mecánica de la alimentación la lengua tiene gran importancia, y éstas, en vertebrados, presentan numerosas diferencias de forma y complejidad que abarcan variaciones de los componentes como epitelios con corpúsculos sensitivos y glándulas epiteliales, musculatura, glándulas del cuerpo lingual, inervación y vasos sanguíneos.
DiSanto (1960) analizó correlativamente, desde la estructura, la función de las glándulas salivales mayores y menores de la cavidad bucal de Desmodus rotundus murinus. Wetterer et al. (2000) realizaron un análisis de la filogenia de Chiroptera considerando, entre otros, los caracteres linguales macroscópicos de tipos y distribución de papilas, musculatura y morfología externa.
En el presente estudio se describe la anatomía macroscópica y microscópica de la lengua de Desmodus rotundus, con el objeto de establecer correspondencias entre estructura y función, y un aporte al conocimiento de la anatomía general del grupo y variaciones observadas.
MATERIALES Y MÉTODOS
Se analizaron ejemplares depositados en la Colección Mamíferos Lillo (CML), Universidad Nacional de Tucumán y Fundación Miguel Lillo (CML, 4816, 3210, 6168, 4268). Para los estudios histológicos las lenguas extraídas fueron fijadas en solución de formaldehído 4% en agua destilada e incluidas en Paraplast in toto. Para aspectos macroscópicos se observaron lenguas de ejemplares intactos bajo lupa binocular. Los cortes se realizaron en forma semi-seriada con micrótomo de deslizamiento a 6 µm de espesor. Las muestras se colorearon con Hematoxilina de Ehrlich-Eosina (McManus y Mowry, 1968) y Tricrómico de Gallego (Vívoli, 1969) para observación de estructuras; Azul de Alcian 8GS, pH 2,5 y 0,5 combinadas con Ácido periódico-reactivo de Schiff, para análisis histoquímicos de detección de muco proteínas y muco polisacáridos ácidos y neutros, respectivamente (Pearse, 1960). Como complemento de las técnicas anteriores se realizó coloración con Azul de Toluidina a pH 5,6 para identificar mucoproteínas y mucopolisacáridos fuertemente ácidos.
Las observaciones con lupa binocular de los detalles de morfología externa, se realizaron mediante pincelado con Azul de Metileno para contraste de relieves en la superficie de la lengua in toto. Las abreviaturas empleadas en el texto son las siguientes: H-E (Hematoxilina de Ehrlich-Eosina), Gallego (Tricrómico de Gallego), AB (Azul de Alcian), PAS (Ácido periódico-reactivo de Schiff ) y TB (Azul de Toluidina).
RESULTADOS
Observaciones macroscópicas
La lengua de D. rotundus es fusiforme, estrecha y alargada. Dorsalmente posee un surco medio que se extiende desde la región media anterior hasta la posterior. Latero-ventralmente hay dos grandes pliegues longitudinales que forman surcos profundos. En observaciones con lupa, las papilas de la zona apical en sus dos tercios anteriores son pequeñas con relación a las del tercio posterior. Estas papilas del ápice o punta lingual se extienden hasta el inicio del tercio posterior. Son de forma cónica y, como el ancho supera ampliamente al espesor, resultan aplanadas antero-posteriormente. En la cara aboral tienen una concavidad. Sus ápices rematan abruptamente en una punta que se dirige hacia atrás. Las papilas del tercio posterior aumentan de tamaño desde el surco medio hacia la región lateral del dorso lingual. Al igual que las anteriores, son aplanadas anteroposteriormente y superan el ancho aproximadamente en tres veces al espesor; son de forma cónica con un ápice abrupto. Su orientación no está generalizada en un orden ya que se muestran desestructuradas y, en general, desordenadas. Se observan, además, algunas pequeñas papilas circulares que tienen una distribución aparentemente azarosa sobre la superficie dorso-anterior de la lengua.
Microanatomía
En corte sagital se observan la mucosa, la submucosa y, por debajo, la musculatura. La mucosa está constituida por el epitelio de revestimiento plano estratificado con distintos grados de queratinización en la mitad anterior y no queratinizado en la mitad posterior del cuerpo y raíz lingual. Completa la mucosa lingual una túnica propia, formada por tejido conectivo laxo sin límites precisos con la submucosa que es muy delgada; carece de muscularis mucosae.
Las papilas que se observan en la región dorsal de la lengua son de dos tipos, unas filiformes y otras similares a las clásicas fungiformes. la morfología de las papilas filiformes, que a su vez pueden ser caracterizadas en tres tipos, varía en la punta propiamente dicha, en los dos tercios anteriores y en el tercio posterior de la lengua. las papilas de la punta lingual son muy semejantes a las papilas filiformes queratinizadas, características de roedores. Su aspecto es cónico, con el eje papilar perpendicular a la superficie lingual, con queratina que cubre todo el ápice. A las siguientes, que se extienden por detrás de las primeras hasta los dos tercios de la longitud total de órgano, las hemos denominado "papilas filiformes raedoras", tienen el ápice dirigido hacia la región posterior, formando una pequeña concavidad por detrás del cuerpo papilar, y están fuertemente queratinizadas (Figs. 1, 2). en cortes sagitales el proceso de queratinización aparece como una formación asimétrica en sentido oral-aboral donde la cara córnea del epitelio (convexo) se dirige hacia la abertura bucal con obvia función lacerante a modo de pala. Evidentemente el desarrollo del proceso de queratinización en este "órgano raedor" implica diferencias en los índices de producción de queratina en dos sectores diferentes de la papila. El último tipo observado de papila filiforme se ubica en el tercio posterior; éstas son altas, gruesas y de ápices redondeados (Figs. 3, 4); el desarrollo de unas con respecto a otras es bastante variable. El eje de tejido conectivo de estas papilas es más abundante y laxo que en las papilas descriptas anteriormente. Las papilas filiformes de la región dorsal se extienden hacia los laterales hasta llegar a una forma casi inconspicua y desaparecer por completo en la región ventral. Las papilas similares a fungiformes son aplanadas y en la superficie tienen estratificación de queratina (Fig. 5).

Fig. 1. Vista dorsal anterior de la lengua: m) músculo estriado; pfr) papila filiforme raedora; pfsr) papila filiforme similar a las de roedores; sm) submucosa. Coloración: Ticrómico de Gallego; aumento 20X; barra = 90µm.
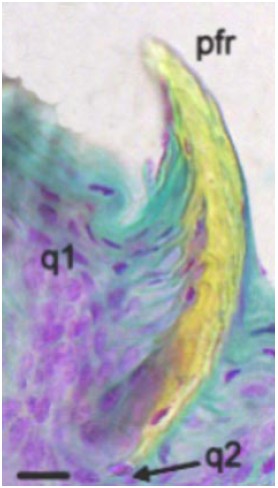
Fig. 2.Papila raedora: q1) proceso de queratinización 1; q2) proceso de queratinización 2; pfr) papila filiforme raedora. Coloración: Tricrómico de Gallego, contraste de fases (Ph 1); aumento 63X; barra = 12µm.

Fig. 3.Detalle de papilas filiformes: dg) ducto glandular; e) epitelio estratificado no queratinizado; m) músculo; pf) papila filiforme. Coloración: tricrómico de Gallego; aumento 20X; barra = 30µm.

Fig. 4.Vista dorsal de la región posterior: e) epitelio; gl) glándulas salivales linguales; i) infiltrado linfocitario; m) músculo; sm) submucosa. Coloración: Tricrómico de Gallego. Aumento: 20X. Barra = 42µm.

Fig. 5.Vista de superficie dorsal media: pf) papila filiforme; pfu) papila fungiforme.Coloración: Tricrómico de Gallego; aumento 20X; barra = 42µm.
Se realizó una búsqueda cuidadosa de corpúsculos gustativos sobre toda el área epitelial lingual, pero no se encontraron. Sin embargo cabe destacar la presencia de nervios que subyacen muy cercanos al epitelio del dorso de la lengua. En la región posterior, o raíz lingual, se observan paquetes nerviosos entre el tejido conectivo interfascicular muscular.
El cuerpo lingual está formado por musculatura estriada esquelética dispuesta en numerosos fascículos con dirección antero-posterior, inmediatamente por debajo de la lámina propia del epitelio dorso lingual; por debajo se encuentran Haces dorsoventrales a modo de columnas que alternan con aces de recorrido transversal, que en corte sagital resultan en paquetes apilados unos sobre otros, ordenados también en columnas.
Entre las bases papilares del dorso lingual desembocan ductos de glándulas salivales (Figs. 4, 6) de estructura tubular contorneada, cuyas unidades secretoras responden a la morfología de glándulas serosas (Figs. 3, 6, 7, 8). Éstas son adenómeros glandulares agrupados y, a diferencia de las glándulas salivales mayores, no constituyen lóbulos individualizables ni poseen cápsula de tejido conectivo, sino que se encuentran dispuestas entre las fibras musculares linguales. Las unidades secretoras de las glándulas salivales linguales están conformadas por dos tipos de células piramidales que se encuentran irregularmente alternadas, tanto en el adenómero como en el conducto de éstas. Los conductos excretores conservan su capacidad secretora casi hasta la zona misma de su desembocadura (Fig. 1). Las células que constituyen dichas glándulas que se observan en mayor número las hemos denominado de "tipo a", y las que se encuentran en menor número, de "tipo b" (Figs. 6, 7, 8).El citoplasma de ambos tipos celulares tiene pequeñas granulaciones basófilas, y los núcleos voluminosos y eucromáticos están generalmente ubicados en su base, tienen finos gránulos de heterocromatina y nucleolos pequeños. Basándose en su naturaleza histoquímica se pueden también detectar e identificar los dos tipos de células glandulares que ya se describieron en base a las coloraciones estructurales.

Fig. 6.Desembocadura glandular: a) célula "tipo a", ortocromática; ag) acino glandular; b) célula "tipo b", metacromática; dg) ducto glandular con desembocadura de glándula; pf) papila filiforme; sm) submucosa. Coloración: Azul de Toluidina; aumento: 20X; barra = 19µm.

Fig. 7.Glándula y ducto: a) células de "tipo a", con contenidos de alcianofilia moderada; b) células de "tipo b", con contenidos fuertemente alcianófilos; ds) ducto secretor; m) músculo. Técnica: Azul de Alcián, pH 0.5; aumento 40X; barra = 26µm.
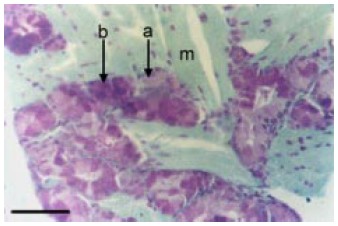
Fig. 8.Glándula salival lingual. Acinos mixtos con células de "tipo a y b"; m) fibras musculares estriadas. Coloración: Tricrómico de Gallego; aumento 63X; barra = 54µm.
Las células del tipo "a" de alcianofilia moderada con AB pH 0.5 y 2.5, dan reacción ortocromática con TB pH 5.6 y las de tipo "b", son fuertemente alcianófilas con AB pH 0.5 y 2.5, y meta cromáticas con TB pH 5.6. Las de tipo "a" no se colorean con refuerzo de coloración obtenida con PAS, lo que sí se observa en las de tipo "b".
Puede observarse un infiltrado linfocitario difuso que no reviste semejanzas micro anatómicas con nódulos linfáticos en la zona postero-dorsal en el tejido conectivo interfascicular e interglandular (Fig. 4).
DISCUSIÓN Y CONCLUSIONES
Según lo observado, y de acuerdo con la información ecológica y de comportamiento disponibles, las estructuras de la lengua de Desmodus rotundus están funcionalmente adaptadas para el tipo de alimentación característica de esta especie. A escala microscópica se observa que las papilas de la región anterior están dispuestas de manera concordante con su función lacerante, con un alto grado de queratinización en la cara anterior y con sus bordes dirigidos hacia atrás, de tal manera que rompen los tejidos de la presa haciendo que el sangrado sea permanente; además la concavidad observada en esas papilas, funciona a modo de pala para arrastrar el tejido desprendido y su posterior deglución. Las papilas del tercio posterior, sin grado alguno de queratinización, aparentemente participan en la sujeción y conducción de sangre y tejidos para la posterior ingesta, formando un canal sobre la superficie lingual.
Nuestras observaciones respecto a la distribución de las papilas en D. rotundus no coinciden con lo descrito por Wetterer et al. (2000), ya que las denominadas filiformes raedoras por nosotros se disponen en diseño diferente al señalado por estos autores para las mismas papilas que denominan "horny papilla"; con respecto a las papilas similares a fungiformes ("basketlike papilla") ellos sólo registran 3 en una hilera central anteroposterior pero en nuestros ejemplares se observaron dispersas sobre el dorso lingual y en mayor número.
La presencia de nervios que subyacen al epitelio, y la aparente ausencia de estructuras sensoriales de tipo epitelial como indicadores de quimiopercepción, sugieren la posible existencia de mecano receptores de ubicación profunda en el epitelio, o en relación con su lámina basal, que no son detectables con estudios de microscopía óptica. Esta conclusión se basa en el conocimiento de que, en estrecha relación con las láminas basales de epitelios linguales, están descriptas ultraestructuralmente células en sinapsis con terminaciones nerviosas, de ubicación similar en vertebrados (Toyoshima, 1989).
La musculatura, de prominente desarrollo en todas direcciones, denota gran movilidad del órgano lingual, con participación activa en la ingesta. Es probable que conjuntamente con la función lacerante de las papilas, el animal realice presión con la lengua in toto sobre la herida, manteniendo de forma constante el flujo de sangre y evitando mecánicamente la coagulación de la misma, a lo que se suma la acción de un factor proteolítico que hidroliza fibrina, secretado por las glándulas submaxilares demostrado experimentalmente en Desmodus rotundus murinus (DiSanto, 1960).
Sería prematuro, con el estudio realizado, establecer una homología entre el infiltrado linfocitario observado en la lengua de esta especie y las tonsilas linguales descriptas para otros vertebrados, cuya ubicación es aproximadamente la misma, y afirmar o rechazar su relación de homología requeriría estudios de tipo ontogénicos.
La presencia de glándulas en la lengua indica algún tipo de participación en la digestión temprana de los componentes sanguíneos que se suma a las secreciones de las glándulas salivales en conjunto. Respecto a los resultados de coloraciones histoquímicas se infiere, como observó DiSanto (1960) para Desmodus rotundus murinus, que las glándulas linguales poseen características morfológicas, como así también en la calidad de sus secreciones, que no se constituyen en francamente mucosas o serosas, según la clasificación clásica de glándulas salivales de vertebrados, sino que sus células presentan aspectos comunes tanto a células de secreción mucosa como también serosa, aspecto que tambien se observa para Desmosdus rotundus. Los tipos celulares "a" y "b" denotan, por su fuerte basofilia citoplasmática, que poseen gran cantidad de ribosomas indicando capacidad secretora de proteínas, sin embargo, también reaccionan positivamente a coloraciones para mucinas y muco proteínas. Las características de las células glandulares linguales determinan a estas glándulas como formadoras de una secreción mucosa constituidas por un complejo de glicoconjugados: mucoproteínas, mucopolisacáridos ácidos carboxilados y fosfatados principalmente, originados por las células de tipo "a". Se suma a ellas un componente ácido (sulfatado) en menor proporción originado por las células de tipo "b". DiSanto (1960) denominó a estos tipos celulares como pseudoserosas y pseudomucosas, a nuestro parecer con criterio acertado.
Los surcos de la región ventral-lateral también fueron observados por Wetterer et al. (2000), quienes los indicaron como carácter del género.
La lengua es un órgano clave entonces, no sólo para analizar por sus implicancias fisiológicas, sino también en el momento de considerar la variedad de caracteres en ella observados y utilizados por Wetterer et al. (2000) con fines taxonómicos. De esto resulta la necesidad de intensificar estudios del tipo aquí aportado para enriquecer la información relacionada con este órgano en los distintos grupos de Chiroptera.
AGRADECIMIENTOS
A la Facultad de Ciencias Naturales de la Universidad Nacional de Tucumán y a la Fundación Miguel Lillo por brindar lugar de trabajo; al Dr. Rubén Barquez y al PIDBA por facilitar los ejemplares y bibliografía pertinentes para el desarrollo del mismo. Al Dr. David Flores por la lectura crítica del manuscrito y sus acertadas observaciones.
LITERATURA CITADA
ARATA AA, JB VAUGHAN y ME THOMAS. 1967. Food habits of certain Colombian bats. Journal of Mammalogy 48(4):653-655. [ Links ]
BARQUEZ RM., MA MARES y JK BRAUN. 1999. The Bats of Argentina. Special Publications, Museum of TexasTechUniversity 42:275 pp. [ Links ]
CRESPO JA, JM VANELLA, BD BLOOD y JM DE CARLO. 1961. Observaciones ecológicas del vampiro Desmodus r. rotundus (Geoffroy) en el norte de Córdoba. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" e Instituto nacional de Investigaciones de las Ciencias Naturales, Ciencias Zoológicas 6:131-351. [ Links ]
DISANTO PE. 1960. Anatomy and Histochemistry of the Salivary Glands of the Vampire Bat, Desmodus rotundus murinus. Journal of Morphology 301-335. [ Links ]
EDGEWORTH FH. 1935. The cranial muscles of vertebrales. Cambridge at the University Press 493 pp. [ Links ]
GREENHALL AM, G JOERMANN y U SCHMIDT. 1983. Desmodus rotundus. Mammalian species 202:1-6. [ Links ]
McMANUS J FA y RW MOWRY. 1968. Técnica Histológica. Editorial Atika, España 612 pp. [ Links ]
PEARSE E. 1960. Histoquímica Teórica y Aplicada, Editorial Aguilar, Madrid 597 pp. [ Links ]
TOYOSHIMA K. 1989. Chemoreceptive and mechanoreceptive paraneurons in the tongue. Archives of Histology and Cytolology 52:383-388. [ Links ]
VILLA-R B y M VILLA-C. 1969. Algunos murciélagos del norte de Argentina. Miscellaneous Publications of the Museum of Natural History, University of Kansas 51:407-428. [ Links ]
VÍVOLI FA. 1969. Técnica Micrográfica. Editorial Rosalo, Buenos Aires 544 pp. [ Links ]
WETTERER AL, MV ROCKMAN y NB SIMMONS. 2000. Phylogeny of Phyllostomid Bats ( Mammalia: Chiroptera): Data from diverse morphological system, sex chromosomes, and restriction sites. Bulletin of the American Museum of Natural History 248:200 pp. [ Links ]
Recibido 31 marzo 2004.
Aceptación final 27 setiembre 2004.

