










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680
Rev. Soc. Entomol. Argent. vol.69 no.3-4 Mendoza jul./dic. 2010
TRABAJOS CIENTÍFICOS
Estudio taxonómico del género monotípico Aphractus (Orthoptera: Tettigoniidae, Pseudophyllinae)
Taxonomic study of the monotypic genus Aphractus (Orthoptera: Tettigoniidae, Pseudophyllinae)
Vera, Alejandro
Departamento de Biología. Facultad de Ciencias Básicas. Universidad Metropolitana de Ciencias de la Educación. José Pedro Alessandri 774. Santiago, Chile; e-mail: alveras2@gmail.com
RESUMEN. Se redescribe la especie Aphractus acuminatus Brunner von Wattenwyl, 1895. Se describen por primera vez el macho, el huevo, la espermateca, el espermatóforo y la estrategia de defensa. Se presenta la distribución geográfica y se discute la morfología genital.
PALABRAS CLAVE. Pseudophyllinae; Aphractini; Morfología genital; Chile.
ABSTRACT. The species Aphractus acuminatus Brunner von Wattenwyl, 1895 is redescribed. The male, eggs, spermateca, spermatophore, and defense strategy are described for the first time. Geographical distribution is presented and genital morphology is discussed.
KEY WORDS. Pseudophyllinae; Aphractini; Genital morphology; Chile.Katydids.
Recibido: 26-VIII-2010;
Aceptado: 18-X-2010
INTRODUCCIÓN
La tribu Aphractini se compone en Chile de cuatro géneros y siete especies, los géneros Aphractus Brunner von Wattenwyl, Paraphractus Beier y Polycleptidella Beier son monoespecíficos y se les conoce casi únicamente por su descripción original. A. acuminatus Brunner von Wattenwyl, 1895 fue descrita a partir de un único espécimen hembra cuya localidad es "Chile", este mismo espécimen fue redescrito por Beier (1962). Posteriormente sólo se le ha citado en varios catálogos, pero sin nuevos aportes al conocimiento de la especie (Elgueta et al. 1999).
Desde el trabajo de Brunner von Wattenwyl (1895), los rasgos morfológicos informativos para la sistemática en el estudio de los Orthoptera han sufrido importantes modificaciones, lo que hace necesario examinar rasgos omitidos inicialmente, más aún cuando se desconoce la morfología del macho. La genitalia de ambos sexos presenta, tanto en sus partes esclerosadas como en sus partes blandas, importantes rasgos para sugerir especies y relaciones sistemáticas. Habitualmente, se describen rasgos observables externamente como los cercos, la placa subgenital y el ovipositor (Buzzetti, 2008; Naskrecki & Lopes-Andrade, 2005); sin embargo, se ha observado una variación intraespecífica sobre estos rasgos cuando se comparan poblaciones fragmentadas históricamente (North & Shaw, 1979), lo que sugiere explorar una mayor diversidad de estructuras genitales. El falo membranoso de los Tettigoniidae puede presentar estructuras esclerosadas (titiladores sensu Ander, 1956) que han sido utilizadas en sistemática, aun cuando no hay acuerdo sobre su homología dentro del orden (Desutter-Grandcolas, 2003). La mayor parte de los Ensifera transfieren sus espermios mediante espermatóforos posicionados externamente en la hembra, estos se acompañan de un tubo que conduce los espermios a la espermateca. Generalmente, este mecanismo se relaciona con órganos fálicos que no operan como verdaderos órganos intromitentes (Alexander & Otte, 1967), y en muchos Tettigoniidae su complejidad morfológica responde a tareas como limpieza del tracto genital femenino, asiento del espermatóforo y ducto respectivo y la adición de un espermatofilax (Gwynne, 2001). Atribuir las funciones a los componentes del falo requiere de un acabado conocimiento de la interacción genital durante la cópula, lo que habitualmente no se ha logrado.
Actualmente la colección de Orthoptera del Museo Nacional de Historia Natural, Santiago-Chile, reúne varios especímenes de Aphractus acuminatus para ambos sexos y varias localidades. Adicionalmente, se han observado especímenes vivos in situ y en condiciones de laboratorio, lo que ha permitido reunir los datos que hacen posible incrementar el conocimiento de esta especie en cuanto a su distribución, estrategias defensivas, morfología del espermatofilax, describir por primera vez el macho y la morfología genital de sus partes blandas, redescribir la hembra y describir su espermateca y el huevo.
MATERIAL Y MÉTODOS
Los datos de los especímenes estudiados se detallan junto a la descripción y han sido depositados en la Colección Entomológica del Museo Nacional de Historia Natural, Santiago-Chile (MNHN). Los especímenes utilizados para el estudio genital han sido conservados en etanol del 70% y posteriormente tratados con KOH 10% por 12 hrs., sin calentar. La descripción sigue la propuesta de Montealegre-Z, 2003, Montealegre-Z et al. 2004 y a Naskrecki, 2008; la morfología genital sigue la nomenclatura de Desutter-Grandcolas, 2003; la nomenclatura de la venación alar sigue a Béthoux & Nel, 2001, 2002; la descripción del huevo y la descripción de las estrategias de defensa están de acuerdo con Hartley, 1974 y Gwynne, 2001, respectivamente.
RESULTADOS
Aphractus Brunner von Wattenwyl, 1895
Aphractus Brunner von Wattenwyl, 1895: 100. Especie tipo: Aphractus acuminatus Brunner von Wattenwyl, 1895, por monotipia.
Diagnosis. Cápsula cefálica de superficie lisa; pronoto liso, con suturas tenues, dorsalmente plano; tegmen lanceolado, en las hembras cubre el ovipositor; segundo par de alas sub-circulares cerca de la mitad del largo de los tégmenes, de color blanco y negro; ovipositor recto, gradualmente aguzado dorsalmente; cercos largos, delgados, dirigidos posteriormente, rectos con una ligera curvatura dorsal, en machos el extremo apical con una espina dirigida medialmente.
Aphractus acuminatus Brunner von Wattenwyl, 1895 (Figs. 1- 17)
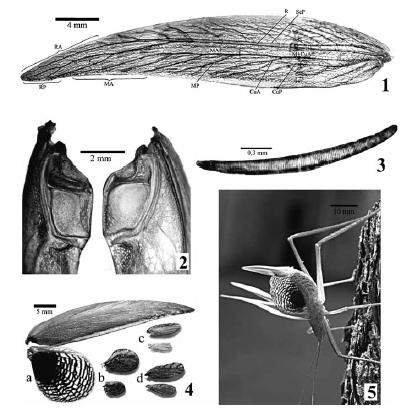
Figs. 1-5. 1, Aphractus acuminatus ♀, ala anterior con la venación indicada; 2, A. acuminatus ♂, porción anterior del tegmen en vista dorsal; 3, A. acuminatus ♂, fila en vista ventral; 4, ♀♀, par de alas derechas en vista dorsal de: a) Aphractus, b) Polycleptis, c) Paraphractus, d) Polycleptidella; 5, A. acuminatus ♀, in vivo con alas extendidas exhibiendo conducta de defensa secundaria.

Figs. 6-8. Aphractus acuminatus ♂. 6, Extremo distal del abdomen en reposo, a) ventral, b) dorsal, c) lateral; 7, Extremo abdominal con lóbulos del ectofalo extendidos, a) lateral, b) posterior; 8, Extremo abdominal con epifalo extendido, a) lateral, b) posterior.

Figs. 9-17. Aphractus acuminatus. 9, ♂, ápice de los bastones del titilador; 10, ♀, ovipositor en vista lateral; 11, Espermatóforo y expermatofilax, a) lateral, b) ventral; 12, ♀, placa sugenital, en vista ventral; 13, ♀, disección con ovariolas y espermateca, el cuadro señala la figura 14; 14, ♀, espermateca; 15, Huevos, a) lateral, b) ventral; 16, Detalle de las micropilas, vistas por transparencia en un huevo aclarado; 17, Detalle del corion perforado del capuchón.
Aphractus acuminatus Brunner von Wattenwyl, 1895: 100; Kirby, W.F. 1906: 310; Pirán, 1942: 246; Beier, 1962: 417; 1963:122; Otte, D., 1997: 200; Elgueta et al., 1999: 15.
Descripción de imagos
Medidas corporales (Promedio ± Desviación Estándar en mm):
♀n=10: largo corporal 33,4 ± 4,7; antena 58,7 ± 5,4; fémures pro/meso/meta 13,7 ± 0,5 / 12,4 ± 0,5 / 22,8 ± 1,3; tegmen 36,8 ± 3,1; ala posterior 15,2 ± 0,8; ovipositor 15 ± 1; cerco 4,1 ± 0,8.
♂n=10: largo corporal 31,1 ± 2,6; antena 49,9 ± 4; fémures pro/meso/meta 11,8 ± 0,4 / 10,3 ± 0,5 / 20,4 ± 1,6; tegmen 28,9 ± 1,1; ala posterior 12,5 ± 0,9; cerco 3,3 ± 0,5.
Imago macho: Cabeza: de superficie lisa, fastigio corto y puntiagudo. Pronoto, margen anterior convexo, margen posterior prominente, convergente y de ápice redondeado; pro, meso y metazona de superficie plana, con punteado espaciado y poco profundo, suturas transversales tenues, lóbulo lateral de margen recto y de borde realzado, seno humeral ausente; procesos proesternales agudos y cortos. Patas delgadas, las medias de menor longitud que las anteriores; espina procoxal desarrollada, delgada y aguda; tibias anteriores sin espinas dorsales, tibias medias con o sin un par y tibias posteriores con 11 o más pares; fémures posteriores delgados sin impresiones musculares. Alas (Figs. 1-5): tegmen lanceolado, con su porción distal curvada posteriormente, en reposo sobrepasa el abdomen; alas posteriores semicirculares (Fig. 4), casi un tercio de la longitud del tegmen; Órgano estridulador ocupa 1/5 de la longitud del tegmen, fila (Fig. 3) alargada de 2mm, con alrededor de 124 dientes lameliformes, ambas alas presentan espejo (Fig. 2).
Abdomen (Figs. 6-9): Tergos poco esclerosados, décimo tergo estrecho (Figs.: 6a-c), con epiprocto (ep) alongado, casi tan largo como el décimo tergo, prominente y con forma de triángulo isósceles (Fig. 6b), en vista lateral es aplanado (Fig. 6c); paraproctos (pa) cortos, robustos y con un tubérculo ventral esclerosado (Ptv) (Fig. 7a-b); cercos (ce) hirsutos, largos y delgados, casi dos veces la longitud del décimo tergo, con su mitad distal ligeramente curvada dorsalmente, con ápice redondeado, con una espina oscura dirigida medialmente (Figs. 6a-c). Noveno esterno con placa subgenital (Psg) trapezoidal, dividida medialmente, con el margen posterior estrecho y hendido; estilos (es) hirsutos, cilíndricos y rectos, 0,3 veces la longitud del cerco (Fig. 6a).
Genitalia (Figs. 7-9): Ectofalo (Ecf) (Fig.8a-b) compuesto de un lóbulo dorsal (Ld) y un lóbulo ventral (Lv) bien diferenciados. El lóbulo ventral (Fig. 7a-b) está subdividido en dos lóbulos laterales (Lvl) y un elemento central (Lvc); cada lóbulo lateral es aguzado con forma de hojuela, mientras que el elemento central está compuesto de dos lóbulos delgados, paralelos, ligeramente aplanados distalmente, con ápices redondeados y divergentes, cada uno presenta un pliegue de división transversal (Lvcpl) que define una mitad distal y otra basal (Fig. 7b). El lóbulo dorsal (Fig.7a y 8b.) está subdividido en dos lóbulos laterales (Ldl) y un elemento central (Ldc); cada lóbulo lateral es grande grueso, aguzado apicalmente y con forma general de hojuela, el elemento central está compuesto de un gran lóbulo plano, ancho y hendido medialmente, provoca un margen distal bilobulado (Fig. 8b).
Epifalo (Epf) (Fig. 8a) en forma subcónica, diferenciado en un titilador evertible, se compone de un esclerito basal (Epeb) y dos bastones articulados (Epba); el esclerito presenta tres ramas, dos de ellas ubicadas en la base y opuestas, con forma de lámina transversal con extremos redondeados, esta base articula la eversión del titilador; la rama media se proyecta en una quilla con dos aristas, en su cima articulan los bastones (Fig. 8b); cada bastón es delgado, recto y muy esclerosado, tan largo como los cercos, con ápice curvado lateralmente y opuesto en cada bastón, los ápices son agudos y están armados de múltiples espinas dirigidas posteriomente (Fig. 9).
Imago hembra: Alas (Figs. 1 y 4) semejantes a las del macho, tegmen sin órgano estridulador, en reposo cubren el ovipositor, RA, RP y MA presentan al menos una horquilla más que en los machos. Cercos (Figs. 10-11) largos (casi un tercio del largo del ovipositor), delgados con el extremo distal curvado dorsalmente. Epiprocto y paraprocto semejantes al macho, pero estos últimos sin el tubérculo ventral. Placa subgenital (Fig. 12) transversal, con el margen posterior ligeramente convexo y un par de procesos submediales dirigidos anteriormente y convergentes por sobre la placa subgenital, cada uno con forma de láminas de ápice redondeado. Ovipositor (Ov) (Fig. 10) de largo similar al fémur posterior, casi recto, aguzado gradualmente, con una ligera inflexión dorsal en la mitad de su largo, de superficie lisa, valva dorsal dentada en su tercio medio, los dientes son poco realzados y decrecen hasta desaparecer. Espermateca (es) (Figs. 13-14) de cápsula esférica, tubo ondulado, tan largo como dos veces el diámetro de la cápsula, con glándula accesoria (gl) simple, como un único tubo cilíndrico plegado, oviductos (ovd) de largo similar al ovipositor.
Espermatóforo (espf) (Figs. 11a y b) de forma esférica, externo, espermatofilax (espfx) transversal, ovoide, de 6,5x5mm.
Coloración: en ambos sexos es semejante. En general, pueden presentarse en tonos amarillo-pardusco o en reemplazo, por gris. Cabeza: antenas grises; clípeo, labro, palpos, frente y genas amarillo pálido, vértex con banda postocular amarilla, con una banda longitudinal medial gris, línea media amarilla. Pronoto, el diseño cefálico se continúa con la banda gris y la línea media amarilla, el resto de la superficie es amarilla a anaranjada, lóbulos laterales amarillo pálido. Meso y meta noto con banda medial gris. Patas pardas, pueden o no presentar bandas longitudinales amarillas. Alas: tegmen amarillo a pardo con pequeñas y dispersas manchas grises, algunos con las venas grises; margen posterior con una banda amarilla. Ala posterior, región costal anaranjada, lámina blanca y venas negras, región basal negra. Abdomen: amarillo a gris, tergos con una banda oscurecida en la línea media, ovipositor rojo con una mancha gris en la base de la valva dorsal.
Huevos: (Figs. 15-17)
Longitud 7±1mm; ancho 1,4±0,5mm. De forma subcilíndrica, comprimido lateralmente, extremo posterior redondeado, extremo anterior con un capuchón, de la mitad del largo de la cápsula. Cápsula de color pardo, con un reborde dorso-ventral crenulado, corion sin ornamentos (Fig. 15), con 7 a 11 micropilas alineadas sobre el borde crenulado, en la mitad ventroposterior (Fig. 16). Capuchón de color pardo, más oscuro que la cápsula, con quilla ventral y ápice agudo dirigido dorsalmente (Fig.15b), superficie estructurada como una red de poros ovoides, de márgenes realzados (Fig.17).
Material examinado. Colección MNHN: Chile Concepción: (1M) fundo Curaco alto 8-12-2003 leg. G. Arraigada; (1N) Escuadrón 10-04-1996, (1N) Periquillo 24-1-17, (2N) Periquillo 1-02-1997 col. Cekalovic; (3M) Hualqui 235m. 2-1-2004 leg. S. Rothmann. Malleco Angol 27-1-1993 col. M. Cerda. (1N) Linares La vega (5km de Parral) 5-11-1991 leg. M. Elgueta. (1N) Cauquenes: Canelillos (El Chovellen) 5-11-1991 leg. M. Elgueta, (1H, 1M) Los Ruiles 23-25-1-2004 leg. A. Vera. Material en cautiverio para observación in vivo de la cópula, en colección del autor: (13M, 9H) Chile VII Región, Cauquenes, sector Los Ruiles, IX-XII-2004, col. A. Vera.
Distribución geográfica y estacional. En Chile entre los 35º58'S hasta los 37º48'S; en la franja costera hasta los 71º50'W. Frecuentemente asociada a la Cordillera de la Costa. Las ninfas abundan durante los meses de junio a diciembre; los adultos a partir de septiembre y hasta febrero.
Hábitat. En el sotobosque, refugiado durante el día en Chusquea sp., Blechnum chilensis y Teline monspessulana (especie exótica); durante la noche son activos caminadores, se les ha observado alimentarse de Nothofagus glauca a gran altura. Se les encuentran en ambientes fragmentados por plantaciones de Pinus sp., (por ejemplo: Reserva Nacional Los Queules y Reserva Nacional Los Ruiles).
Estrategias de defensa. La defensa primaria es el camuflaje, donde su coloración y forma semeja hojas y tallos de Chusquea sp. Las defensas secundarias consisten en dejarse caer o saltar hacia atrás. La insistencia del depredador (estímulo) provoca una respuesta exagerada (Fig. 5): levanta su cuerpo tres veces más alto que el estado inicial, simultáneamente abre sus alas exponiendo el segundo par, curva el abdomen dorsalmente casi asomándose entre las alas (en las hembras se expone el intenso color rojo del ovipositor), secreta abundante líquido rojo y fétido desde la boca y exhibe un rápido movimiento de balanceo circular mientras huelle.
DISCUSIÓN
En cuanto a las genitalias, éstas muestran una gran riqueza de caracteres. La genitalia masculina es muy compleja y los bastones del titilador son de longitud semejante al tubo de la espermateca, con lo que podrían cumplir una función de limpieza previa a la cópula, observaciones inéditas en otros Aphractini muestran que también presentan estos bastones articulados, pero de diferente morfología.
Aphractus acuminatus es el único Aphractini con tégmenes desarrollados y alas posteriores con coloración distintiva, como se muestra en la figura 4 las restantes especies presentan alas muy reducidas. Esta diferencia podría relacionarse con su conducta defensiva exagerada. Otros Aphractini exhiben semejantes estrategias primarias de defensa, pero no manifiestan esta conducta exagerada, sólo Polycleptis presenta alas posteriores de color negro o azul metálico y las exhibe al verse agredido.
En este estudio, se muestra una gran variedad de información que es inexistente en otros miembros de Aphractini, lo que hace imposible su comparación. Esto sugiere la necesidad de un análisis similar de las restantes especies.
AGRADECIMIENTOS
Al Museo Nacional de Historia Natural Santiago-Chile Entomología, por facilitar laboratorios y equipos; a los evaluadores del manuscrito; a quienes me facilitaron bibliografía: Mario Elgueta, Holger Braun, André Nel y Piotr Naskrecki; y a todos los que me entregaron algún dato o un momento de conversación sobre el tema, todos miembros de la Sociedad Chilena de Entomología.
BIBLIOGRAFÍA CITADA
1. ALEXANDER, R. D. & D. OTTE. 1967. The evolution of genitalia and mating behavior in crickets (Gryllidae) and other Orthoptera. Miscellaneous Publications Museum of Zoology, University of Michigan 133: 1-62. [ Links ]
2. ANDER, K. 1956. Orthoptera Saltatoria. In: Tuxen (eds.) Taxonomist´s Glossary of genitalia in Insects, pp. 53-63. Copenhagen. [ Links ]
3. BEIER, M. 1962. Orthoptera: Tettigoniidae (Pseudophyllinae I). Das Tierreich 73: 1-468. [ Links ]
4. BEIER, M. 1963. Tettigoniidae: subfam. Pseudophyllinae. Orthopterorum Catalogus 5: 1-246. [ Links ]
5. BÉTHOUX, O. & A. NEL. 2001. Venation pattern of Orthoptera. Journal of Orthoptera Research 10 (2): 195-198 [ Links ]
6. BÉTHOUX, O. & A. NEL. 2002. Venation pattern and revision of Orthoptera sensu nov. and sister groups. Phylogeny of palaeozoic and mesozoic Orthoptera sensu nov. Zootaxa 96: 1-88. [ Links ]
7. BRUNNER VON WATTENWYL, C. 1895. Monographie der Pseudophylliden. Verhandlungen der Kaiserlich-Königlichen Zoologisch-botanischen Gesellschaft in Wien 45: 1-282. [ Links ]
8. BUZZETTI, F. M. 2008. A new species of Incanotus Beier from Ecuador (Orthoptera: Tettigoniidae: Pseudophyllinae). Entomotropica 23 (1): 37-41. [ Links ]
9. DESUTTER-GRANDCOLAS, L. 2003. Phylogeny and the evolution of acoustic communication in extant Ensifera (Insecta, Orthoptera). Zoologica Scripta 32 (6): 525-561. [ Links ]
10. ELGUETA, M., A. CAMOUSSEIGHT & C. S. CARBONELL. 1999. Catálogo de Orthoptera (Insecta) de Chile. Publicación Ocasional, Museo Nacional de Historia Natural, Chile, 54: 60pp. [ Links ]
11. GWYNNE, D. T. 2001. Katydids and Bush-Crickets Reproductive Behavior and Evolution of the Tettigoniidae. Cornell University Press, Ithaca. [ Links ]
12. HARTLEY, J. C. 1964. The structure of the eggs of the British Tettigoniidae (Orthoptera). Proceedings of the Royal Entomological Society of London 39 (7-9): 111-117. [ Links ]
13. KIRBY, W. F. 1906. A synonymic catalogue of Orthoptera. Vol. II. Orthoptera Saltatoria. Part I. (Achetidae et Phasgonuridae.) Catalogue of Orthoptera. British Museum, Londres. [ Links ]
14. MONTEALEGRE-Z, F. & G. K. MORRIS. 2004. The spiny devil katydids, Panacanthus Walker (Orthoptera: Tettigoniidae): an evolutionary study of acoustic behaviour and morphological traits. Systematic Entomology 29: 21-57. [ Links ]
15. MONTEALEGRE-Z, F., P. A. GUERRA & G. K. MORRIS. 2003. Panoploscelis specularis (Orthoptera: Tettigoniidae: Pseudophyllinae): extraordinary female sound generator, male description, male protest and calling signals. Journal of Orthoptera Research 12 (2): 173-181 [ Links ]
16. NASKRECKI, P. & C. LOPES-ANDRADE. 2005. A new genus and species of katydids of the tribe Polyancistrini (Orthoptera: Tettigoniidae: Pseudophyllinae) from Brazil, an apparent pest of Eucalyptus plantations. Zootaxa 952: 1-8 [ Links ]
17. NASKRECKI, P. 2008. Sylvan katydids (Orthoptera: Tettigoniidae: Pseudophyllinae) of the Guinean Forests of West Africa hotspot: an overview and descriptions of new species. Zootaxa 1712: 1-41. [ Links ]
18. NORTH, R. C. & K. C. SHAW. 1979. Variation in distribution, morphology and calling song of two populations of Pterophylla camellifolia (Orthoptera: Tettigoniidae). Psyche 86: 363-374. [ Links ]
19. OTTE, D. 1997. Tettigonioidea. Orthoptera Species File 7. The Orthopterists' Society and The Academy of Natural Sciences of Philadelphia, Philadelphia. [ Links ]
20. PIRÁN, A. 1942. Catálogo sistemático y zoogeográfico de Tettigonioideos argentinos. Revista de la Sociedad Entomológica Argentina 11 (3): 240-287. [ Links ]

