










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.42 n.1-2 Córdoba ene./jul. 2007
Panorama de la familia Asteraceae (= Compositae) en la Republica Argentina *
Liliana Katinas1, Diego G. Gutiérrez1, Mariana A. Grossi1 y Jorge V. Crisci1
*Trabajo publicado en homenaje a la Dra Irma J. Gamundi en conmemoración de su 80º aniversario.
1División Plantas Vasculares, Museo de La Plata , Paseo del Bosque s/n, B1900FWA La Plata.
2Laboratorio de Sistemática y Biología Evolutiva (LASBE), Museo de La Plata , Paseo del Bosque s/n, B1900FWA La Plata.
Resumen: Se presenta un panorama de la familia Asteraceae en la República Argentina , que incluye una breve historia taxonómica, datos numéricos de tribus, géneros y especies, ejemplos de endemismos y daptaciones ecológicas y una reseña sobre la hipótesis del origen de la familia en Patagonia. En la Argentina, las asteráceas representan la familia más numerosa con 227 géneros (cinco son endémicos) y ca. 1400 especies (92 son adventicias y 382 son endémicas). En términos de taxones nativos, se hallan representadas en nuestro país todas las tribus de Asteraceae excepto Inuleae (sensu Bremer). Las tribus Arctoteae y Calenduleae están representadas por taxones adventicios exclusivamente y Cardueae posee sólo una especie nativa. Barnadesieae y Mutisieae son las únicas tribus con géneros endémicos de Argentina y también presentan el porcentaje más alto de géneros y especies en el país en comparación con el resto de las tribus. El mayor número de especies nativas, endémicas y no endémicas, se da en las provincias fitogeográficas Altoandina y Patagónica. La tribu más representada en número de géneros es Heliantheae, y en número de especies nativas es Senecioneae.
Palabras clave: Asteraceae; Panorama; República Argentina.
Summary: An overview of the plant family Asteraceae ( = Compositae) in Argentina. An overview of the family Asteraceae in Argentina that includes a brief taxonomic history, numerical data of tribes, genera and species, examples of endemisms and ecological adaptations, and a discussion on the hypothesis on the origin of the family in Patagonia is given. Asteraceae is the most numerous family in Argentina , with 227 genera (five are endemic) and ca. 1400 species (92 are adventitious and 382 are endemic). In relation to the native taxa, all tribes of Asteraceae occur in Argentina excepting Inuleae (sensu Bremer). Tribes Arctoteae and Calenduleae are represented exclusively by adventitious taxa, and Cardueae has only one native species. Barnadesieae and Mutisieae are the only tribes with endemic genera in Argentina and they also present the highest percentage of genera and species occurring in the country when compared with the rest of the tribes. The highest number of native species occurs in the Altoandean and Patagonian phytogeographic provinces. The most represented tribe in number of genera is Heliantheae, and in number of species is the tribe Senecioneae.
Key words: Asteraceae; Overview; Argentina.
INTRODUCCIÓN
Los crisantemos, margaritas, dalias, girasoles, cardos, la achicoria y la lechuga son algunos de los representantes de la familia Asteraceae, Compositae, o asteráceas o compuestas en un lenguaje coloquial. Las asteráceas comprenden más de 1700 géneros y unas 24.000-30.000 especies distribuidas por todo e mundo, excepto en la Antártida , que incluyen desde pequeñas hierbas de 1 cm de altura hasta árboles de más de 30 m . Las asteráceas se reconocen por su estructura reproductiva, el capítulo donde las flores se disponen en forma sésil sobre un receptáculo ensanchado.
Asteraceae es la familia más numerosa en nuestro país (Cabrera, 1961; Zuloaga et al., 1999) y ello sumado a su amplia distribución en la Argentina y a las hipótesis que consideran a la Patagonia como el probable lugar de origen de las asteráceas (Stuessy et al., 1996), justifica la presentación de un compendio de la familia en la República Argentina. Para ello se actualizó toda la información concerniente a Asteraceae desde la publicación del catálogo de las plantas vasculares de la República Argentina (Zuloaga & Morrone, 1999) hasta el presente. La información se extrajo de material de herbario, de viajes de campo y de la literatura base (Bremer, 1994; Zuloaga & Morrone, 1999; Zuloaga et al., 1999) y se actualizó mediante la consulta a las principales revistas de sistemática botánica que generalmente tratan taxones de Argentina: Annals of the Missouri Botanical Garden, Compositae Newsletter, Darwiniana, Hickenia, Kew Bulletin, Kurtziana, Novon, Systematic Botany, Taxon. También se consultó el tratamiento de la familia Asteraceae en Argentina en floras, catálogos y sinopsis: Cabrera (1971a, 1974, 1978), Ariza Espinar (1994, 1997, 2000, 2005), Freire (1995, 1996), Katinas (1995, 1996), Ezcurra (1996), Sancho (1996), Urtubey (1996), Cabrera & Freire (1997), Cerana (1997), Delucchi & Ariza Espinar (1997), Petenatti & Ariza Espinar (1997), Ariza Espinar & Delucchi (1998a, b), Ariza Espinar & Urtubey (1998), Bortiri (1999), Cabrera et al. (1999, 2000), Troiani & Steibel (1999), Giuliano (2000), Bártoli & Tortosa (2003), Novara (2003), Sancho & Ariza Espinar (2003), Ariza Espinar & Novara (2005), Freire et al. (2005). Para la clasificación en subfamilias y tribus se sigue a Bremer (1994) y para la clasificación de las provincias fitogeográficas a Cabrera (1971b). El objetivo de este trabajo es presentar un panorama de las asteráceas que habitan la República Argentina que incluye datos numéricos actualizados respecto a las tribus, géneros y especies, y otros datos de interés.
Breve historia taxonómica de la familia
Si bien las asteráceas ya fueron reconocidas por Teofrasto hace unos 300 años A. C., la familia fue fundada con el nombre de Compositae ( nom. cons. et nom. alt .) por Giseke en 1792. El código internacional de nomenclatura botánica (Greuter et al., 2000) permite también el uso del nombre Asteraceae Dumortier ( nom. cons .), nombre asignado a la familia en 1822. Las contribuciones más importantes al conocimiento y la sistemática de la familia comienzan con el francés Henri Cassini a través de numerosas publicaciones durante el período 1812-1831 que incluyen descripciones muy detalladas de la morfología de las asteráceas. Cassini además estableció 19 tribus en la familia (Cassini, 1816). Lessing (1832) redujo a ocho el número de tribus y de Candolle (1838) a tres. Otro gran contribuyente al conocimiento de la familia fue George Bentham (1873), que trató a las asteráceas en su monumental obra Genera Plantarum (1862-1883) junto con Joseph Dalton Hooker. Fue Bentham quien estableció 13 tribus, que son las tradicionalmente usadas hoy.
Luego de Bentham hubo muchos botánicos en todo el mundo que realizaron importantes aportes al conocimiento de las asteráceas, pero si nos referimos a la Argentina , el gran botánico español que marcó los estudios en esta familia de plantas fue Angel Lulio Cabrera (1908-1999). Cabrera fue un generador y catalizador del estudio de Asteraceae en la Argentina y sus trabajos sobre la sistemática de asteráceas son considerados clásicos y de consulta obligada en todo el mundo. Parte de su legado incluye cerca de un centenar de trabajos en distintos taxones de asteráceas, el tratamiento de la familia en floras de nuestro país, y su valiosísimo herbario depositado principalmente en los herbarios del Museo de La Plata (LP) y del Instituto de Botánica Darwinion en San Isidro (SI), provincia de Buenos Aires. Cabrera y autores subsiguientes se basaron en la clasificación de Bentham (1873) en el tratamiento de Asteraceae en las floras locales. En su clave para la determinación de géneros de Asteraceae de Argentina, Cabrera (1961) considera la subfamilia Tubuliflorae con las tribus Anthemideae, Arctotideae, Astereae, Calenduleae, Cynareae, Eupatorieae, Helenieae, Heliantheae, Inuleae, Mutisieae, Senecioneae y Vernonieae, y la subfamilia Liguliflorae con la tribu Cichorieae. Un hito importante en la historia taxonómica de las asteráceas y que trajo gran controversia fue la denominada New Synantherlogy («Nueva Sinanterología», pues los especialistas en asteráceas se llaman sinanterólogos por la morfología de las anteras), establecida en numerosos trabajos de los botánicos estadounidenses Harlod Robinson y Robert M. King (King & Robinson, 1970). Esta tendencia consistía en un mayor énfasis en el uso de los microcaracteres (caracteres anatómicos, citológicos y químicos) en lugar de los macrocaracteres para tomar decisiones taxonómicas. A modo de ejemplo, entre 1967 y 1976 King y Robinson publicaron no menos de 167 trabajos científicos sobre Eupatorieae, en los que dividieron el género Eupatorium , que tenía en ese momento cerca de 600 especies, en numerosos géneros menores (McVaugh, 1982). Si se siguiera el criterio de King y Robinson en las floras locales, el género Eupatorium ya no existiría en la Argentina dado que de sus 82 especies argentinas, 20 se transfirieron a Chromolaena y el resto a 33 géneros diferentes. En las floras regionales de Argentina se ha seguido en general el uso taxonómico tradicional de tribus y géneros mencionado previamente, y sólo en unos pocos casos, como por ejemplo en la tribu Liabeae (Cabrera et al., 1999), se siguió el criterio de King y Robinson.
Uno de los aportes más significativos a la clasificación y filogenia de las asteráceas llegó con los estudios moleculares de ADN de Jansen & Palmer (1987). Su descubrimiento inicial fue que los géneros muestreados de asteráceas poseían una inversión de 22 kilobases en el genoma cloroplástico, excepto en los tres géneros Barnadesia , Chuquiraga y Dasyphyllum de la subtribu Barnadesiinae (perteneciente en ese momento a la tribu Mutisieae) y algunas familias afines a Asteraceae. Ello hizo que unos años después esta subtribu, muy representada en la Argentina , pasara al rango de subfamilia Barnadesioideae y que sea considerado el taxón basal en el árbol evolutivo de la familia.
También en el año 1987 el botánico sueco Kåre Bremer produce el primer cladograma de las asteráceas esencialmente sobre la base de caracteres morfológicos y unos años después produce un tratado sobre la familia (Bremer, 1994). La clasificación de Bremer (1994) toma en cuenta los avances en el conocimiento de la filogenia de la familia y comprende tres subfamilias: la subfamilia Barnadesioideae con la tribu Barnadesieae (e.g., Chuquiraga avellanedae ), la subfamilia Cichorioideae con las tribus Arctoteae (= Arctotideae; e.g., Gazania tenuifolia , gazanias), Cardueae (= Cynareae; e.g., Carduus thoermeri , cardos), Lactuceae (= Cichorieae; e.g., Lactuca sativa , lechuga), Liabeae (e.g., Liabum acuminatum ), Mutisieae (e.g., Mutisia decurrens , mutisias) y Vernonieae (e.g., Vernonia mollisima ), y la subfamilia Asteroideae con las tribus Anthemideae (e.g., Argyranthemum frutescens , margaritas), Astereae (e.g., Baccharis trimera , carquejas), Calenduleae (e.g., Calendula officinalis , caléndulas), Eupatorieae (e.g., Eupatorium bupleurifolium , Ageratum houstonianum ), Gnaphalieae (e.g., Helichrysum bracteatum , siemprevivas), Helenieae (e.g., Tagetes minuta , chinchilla), Heliantheae (e.g., Helianthus annuus , girasol), Inuleae (e.g., Inula helenium ), Plucheeae (e.g., Tessaria dodoneifolia, pájaro bobo) y Senecioneae (e.g., Senecio vulgaris ). Las principales diferencias entre la clasificación de Bremer (1994) y las clasificaciones previas es: (1) la consideración de Barnadesioideae como subfamilia independiente (antes la subtribu Barnadesiinae de la tribu Mutisieae); (2) la división de la tribu Inuleae en las tres tribus Inuleae sensu stricto, Gnaphalieae y Plucheeae; y (3) la consideración de Liabeae como tribu independiente (antes la subtribu Liabinae de la tribu Senecioneae, e.g., Cabrera, 1961).
Estos estudios van en paralelo con el avance de los análisis moleculares que llevaron en años recientes a la disgregación en numerosas tribus o subfamilias de la mayoría de las tribus tradicionales (Panero & Funk, 2002; Funk et al., 2005). A modo de ejemplo la tribu Mutisieae que en su sentido amplio tiene 74 géneros y unas 865 especies (Cabrera, 1977; Katinas et al., en prensa) queda dividida, de acuerdo a los resultados moleculares, en al menos tres subfamilias, una tribu y dos clados sin asignación taxonómica (Funk et al., 2005).
Número de tribus, géneros y especies
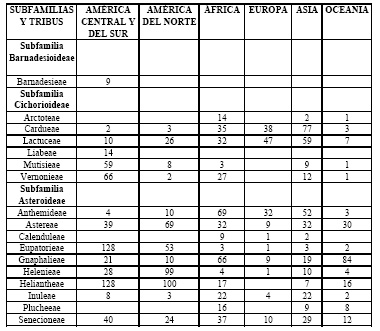
Las asteráceas se encuentran en todos los continentes excepto la Antártida dado que sus miembros han sido extraordinariamente exitosos en la adaptación a los más diversos hábitats. Su distribución en los continentes se muestra en la Tabla 1: (1) en las filas puede verse que hay tribus ampliamente distribuidas (e.g., Eupatorieae, Helenieae), mientras que otras son endémicas de un continente o parte de un continente (e.g., Barnadesieae, Liabeae) o tienen distribuciones relativamente restringidas (e.g., Arctoteae, Plucheeae); (2) las columnas muestran que hay tribus que son típicamente del Nuevo Mundo, algunas de ellas concentradas en América del Norte (Astereae, Helenieae) y otras en América Central y América del Sur (e.g., Eupatorieae, Heliantheae, Vernonieae). También hay tribus claramente concentradas en el Viejo Mundo (e.g., Anthemideae, Arctoteae, Calenduleae). En su conjunto, existe una mayor representación de las tribus en el hemisferio sur.
Tabla 1. Número de géneros nativos de Asteraceae por tribu en los distintos continentes o partes de continentes.
Considerando taxones nativos y adventicios, las asteráceas representan la familia vegetal más numerosa en la República Argentina , con 227 géneros, cinco son endémicos y cerca de 1400 especies. De la totalidad de especies 92 son exóticas o adventicias y unas 382 son endémicas; de las especies endémicas un poco menos de la mitad pertenecen al género Senecio . Es interesante comparar estos números con otros países como por ejemplo el Perú y Ecuador, que están entre los países sudamericanos más ricos en biodiversidad (Tabla 2). Los valores de la Tabla 2 muestran que Argentina no está muy alejada en el número de taxones nativos y endémicos con Perú y/ o Ecuador lo cual indica la riqueza de asteráceas en nuestro país si tenemos en cuenta que Perú tiene 17.560 plantas vasculares (Brako & Zarucchi 1993; Vásquez et al., 2002) y Ecuador unas 15.900 (Jørgensen & León-Yañez, 1999; Valencia et al., 2000), en comparación con la Argentina que posee 9.690 (Zuloaga & Morrone, 1999) a pesar de tener una superficie mayor que los otros dos países.
Tabla 2. Comparación del número de géneros y especies de Asteraceae en la Argentina con países muy ricos en biodiversidad como Perú y Ecuador.

En la República Argentina , se hallan representadas todas las tribus de asteráceas, excepto Inuleae (sensu Bremer, 1994) (Tabla 3). Las tribus Arctoteae y Calenduleae están representadas por taxones adventicios exclusivamente y Cardueae posee sólo una especie nativa ( Centaurea tweediei ). La Tabla 3 muestra que Heliantheae es la tribu con mayor número de géneros (42) en Argentina seguida de Mutisieae (con 34 géneros). Pero si se tiene en cuenta qué porcentaje de géneros de estas tribus existe en Argentina respecto al total mundial, Heliantheae tiene representados un 22 % del total de sus géneros, mientras que Mutisieae tiene el 46 %. Otro tanto ocurre con Barnadesieae que sólo tiene nueve géneros, pero siete de ellos (el 78 %) habita nuestro país; Liabeae tiene el 26,6 % y en el resto de las tribus los valores son menores al 15 %. Respecto al número de especies nativas en Argentina (Tabla 3), Senecioneae, Asteraeae y Eupatorieae tienen los valores más altos, 289, 246 y 150 especies respectivamente y en estos casos se debe a que estas tribus tienen en cada una de ellas un género con numerosas especies. El género Senecio contribuye con 270 (Cabrera et al., 1999) a las 289 especies de Senecioneae en Argentina, Baccharis con 96 (Giuliano, 2000) a las 246 especies de Astereae, y Eupatorium con 82 (Cabrera & Freire, 1997) a las 150 especies de Eupatorieae. Por otro lado, otras tribus como por ejemplo Mutisieae y Heliantheae tienen también un alto número de especies pero éstas forman parte de diversos géneros. Si se tiene en cuenta qué grado de representación de especies tiene cada tribu en la Argentina , las tribus Barnadesieae (con un 28,3 % de sus especies que habitan nuestro país) y Mutisieae (con el 16,6 %) tienen los valores más altos, mientras que en el resto de las tribus los valores son menores al 9 %.
Tabla 3. Tribus de la familia Asteraceae presentes en la Argentina , con el número y porcentaje (del total de taxones en Argentina) de géneros y especies nativas (incluye taxones endémicos y no endémicos), de géneros y especies adventicias y de especies endémicas.

A nivel de las provincias políticas de la Argentina, la Figura 1 muestra el porcentaje de tribus, a nivel específico, en cada provincia. La tribu Astereae, por ejemplo, tiene una muy buena representación en todas las provincias. Por el contrario la tribu Heliantheae, que posee el mayor número de géneros en nuestro país, tiene muy buena representación en algunas provincias (Formosa con 22 %, Santiago del Estero con 21%) pero es muy escasa en otras (Santa Cruz con 1 %). La tribu Eupatorieae está muy bien representada en provincias con clima cálido como Misiones (24 %) y Chaco (20 %), Senecioneae es abundante en Neuquén (31 %) y Mendoza (25 %), y Mutisieae en Santa Cruz (29 %) y San Juan (26 %). Las provincias de Salta y Jujuy poseen el mayor número de especies nativas de asteráceas, y Tierra del Fuego es la que posee el menor número (Fig. 2A). Salta, Catamarca y Jujuy comparten un alto número de especies endémicas, mientras que Formosa, Santa Fe y Misiones son las que poseen el menor número de endemismos (Fig. 2B). Los endemismos de las provincias políticas fueron considerados como especies endémicas de la Argentina presentes en una provincia determinada, es decir que muchas veces los endemismos mencionados no son exclusivos de esa provincia. Respecto a las especies exóticas o adventicias, es la provincia de Buenos Aires la que posee el número más alto en tanto que Formosa es la provincia con el menor número (Fig. 2C).
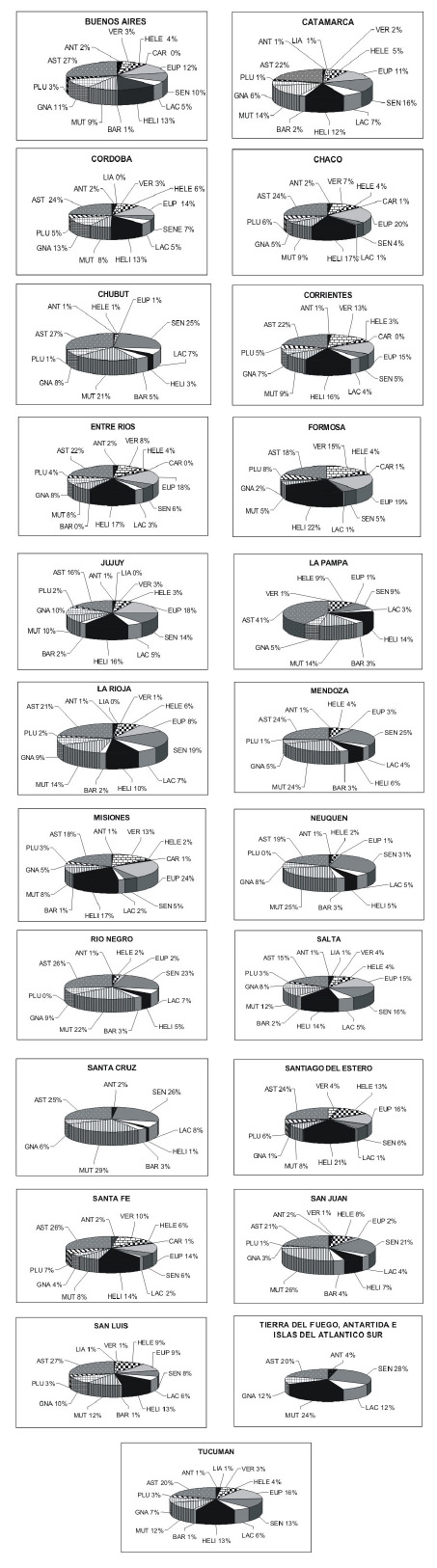
Fig. 1 . Representación en porcentaje de las tribus de Asteraceae en las provincias políticas de Argentina. ANT = Anthemideae; ARC = Arctoteae; AST = Astereae; BAR = Barnadesieae; CAL = Calenduleae; CAR = Cardueae; EUP = Eupatorieae; GNA = Gnaphalieae; HELE = Helenieae; HELI = Heliantheae; LAC = Lactuceae; LIA = Liabeae; MUT = Mutisieae; PLU = Plucheeae; SEN = Senecioneae; VER = Vernonieae.

Fig. 2 . Número de especies nativas y no nativas de Asteraceae en las provincias políticas de Argentina. A . Número de especies nativas (endémicas y no endémicas). B . Número de especies endémicas de Argentina, presentes en cada provincia pero no exclusivas de ésta. C . Número de especies adventicias. BAI = Buenos Aires; CAT = Catamarca; CBA = Córdoba; CHA = Chaco; CHU = Chubut; CRR = Corrientes; ERI = Entre Ríos; FOR = Formosa; JUJ = Jujuy; LPA = La Pampa ; LRI = La Rioja ; MEN = Mendoza; MIS = Misiones; NEU = Neuquén; RNE = Río Negro; SAL = Salta; SCR = Santa Cruz; SDE = Santiago del Estero; SFE = Santa Fe; SJU = San Juan; SLU = San Luis; TDF = Tierra del Fuego e Islas del Atlántico Sur; TUC = Tucumán.
Es interesante analizar esta información también a nivel de las provincias fitogeográficas de Argentina (Cabrera, 1971b). En general, todas las tribus están representadas en todas las provincias fitogeográficas (Fig. 3); las Senecioneae por ejemplo, son más abundantes en la provincia Altoandina, y las Eupatorieae en la Paranense y de las Yungas. La provincia Subantártica es la que posee la menor representación de tribus. La figura 4 muestra los datos numéricos de géneros, especies nativas (incluyendo las endémicas), endémicas exclusivamente y adventicias en cada provincia fitogeográfica en un mapa de la Argentina. En cuanto al número de especies nativas (no endémicas), las provincias Altoandina y Chaqueña son las que poseen los valores más altos, y la Subantártica y del Monte los valores más bajos (Fig. 5A). La provincia Altoandina se destaca también por su alto número de endemismos, seguida por la Patagónica, el menor número se registra en las provincias del Espinal y Paranense (Fig. 5B). La provincia Pampeana posee el mayor número de taxones adventicios (aunque también tiene un alto número de especies nativas), seguida de la Chaqueña y del Espinal, mientras que la Altoandina posee el número más bajo (Fig. 5C).

Fig. 3 . Representación en porcentaje de las tribus de Asteraceae en las provincias fitogeográficas de Argentina. ANT = Anthemideae; ARC = Arctoteae; AST = Astereae; BAR = Barnadesieae; CAL = Calenduleae; CAR = Cardueae; EUP = Eupatorieae; GNA = Gnaphalieae; HELE = Helenieae; HELI = Heliantheae; LAC = Lactuceae; LIA = Liabeae; MUT = Mutisieae; PLU = Plucheeae; SEN = Senecioneae; VER = Vernonieae.
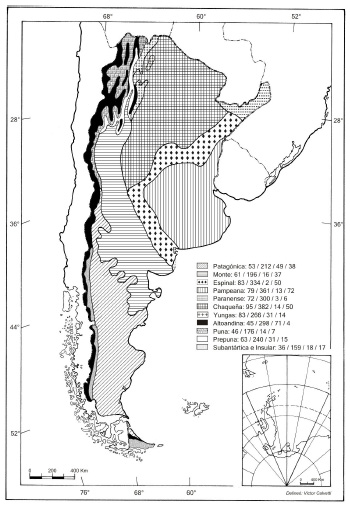
Fig. 4 . Mapa de las regiones fitogeográficas de la Argentina de acuerdo con Cabrera (1971b) indicando el número de géneros y especies de Asteraceae por provincia. La primera cifra indica el número de géneros; la segunda cifra indica el número de especies nativas (incluye endémicas y no endémicas); la tercera cifra indica el número de especies endémicas; y la cuarta cifra indica el número de especies adventicias.

Fig. 5 . Número de especies nativas y no nativas de Asteraceae en las provincias fitogeográficas de Argentina. A . Número de especies nativas (excluyendo las endémicas). B . Número de especies endémicas exclusivas de cada provincia fitogeográfica. C . Número de especies adventicias. ALT = Provincia Altoandina; CHA= Provincia Chaqueña; ESP = Provincia del Espinal; MON = Provincia del Monte; PAM = Provincia Pampeana; PAR = Provincia Paranense; PAT = Provincia Patagónica; PPU = Provincia Prepuneña; PUN = Provincia Puneña; SAI = Provincia Subantártica e Insular; YUN = Provincia de las Yungas.
La comparación entre los datos de las provincias políticas y las fitogeográficas arroja algunos resultados interesantes. El alto número de especies nativas, endémicas y no endémicas, en las provincias de Jujuy y Salta ocurre en la provincia fitogeográfica Altoandina. En las provincias patagónicas, Neuquén, Río Negro, Chubut, Santa Cruz y Tierra del Fuego, el número alto de endemismos se da en la provincia fitogeográfica Patagónica y no en la Subantártica. Es evidente que para las asteráceas las provincias importantes en número de taxones nativos son la Altoandina y la Patagónica. Esta información puede resultar importante a la hora de elegir áreas de conservación de la biodiversidad, dado que si bien las áreas de conservación se seleccionan dentro de las provincias políticas, debería considerarse en que área fitogeográfica de la provincia se halla la mayor riqueza de especies.
Endemismos
Argentina posee cinco géneros endémicos de Asteraceae, Ameghinoa , Burkartia , Dolichlasium (Mutisieae), Duseniella y Huarpea (Barnadesieae) y la familia ocupa el segundo lugar entre las plantas vasculares en cuanto al número de géneros endémicos luego de Brassicaceae (con nueve géneros endémicos) (Zuloaga et al., 1999). Los géneros endémicos de asteráceas habitan exclusivamente las provincias fitogeográficas Patagónica y Altoandina, y es importante remarcar que están restringidos a sólo dos tribus, filogenéticamente relacionadas.
Con respecto a los endemismos a nivel específico, hay taxones endémicos de provincias fitogeográficas, como por ejemplo Senecio jujuyensis y S. punae de la Puna; Aster patagonicus , Brachyclados caespitosus y Chuquiraga avellanedae de la estepa patagónica; Stevia sanguinea de las Yungas; Eupatorium militare , Grindelia chacoensis y G. ragonesei de la región Chaqueña. Algunas especies son endémicas de áreas muy restringidas, por ejemplo Baccharis phyteuma es endémica del Delta del Paraná; Senecio melanopotamicus se conoce únicamente para las barrancas del Río Negro, en los alrededores de Carmen de Patagones; Nardophyllum deserticola crece en Plaza Huincul, en el centro de la provincia de Neuquén. Algunos endemismos de cerros son Stevia centinelae hallada únicamente en la cima del cerro Centinela, Jujuy, a 2650 m altura; Conyza magnimontana colectada en la sierra Grande, en el este de la provincia de Río Negro; Senecio repollensis del cerro Repollo, provincia de Neuquén; Baccharis tandilensis , Hieracium tandilense , Senecio bravensis de las Sierras de Tandil, provincia de Buenos Aires; Hieracium chacoense y Senecio leucopeplus de Sierra de la Ventana , provincia de Buenos Aires. Las Islas Malvinas también incluyen asteráceas endémicas como Chevreulia lycopodioides, Erigeron incertus , Gamochaeta antarctica , Leucheria suaveolens , Nassauvia gaudichaudii , N. serpens , Senecio littoralis y S. vaginatus.
Ecología
La República Argentina tiene importantes características topográficas y posee una amplísima gama de suelos y climas que reúnen condiciones únicas para la distribución de las asteráceas. La topografía está afectada por la latitud y altitud y es muy variable de este a oeste y de norte a sur. Uno de los accidentes topográficos más notables es la cordillera de los Andes al oeste del país (que incluye el Aconcagua de 6.959 m , la montaña más alta de América) y que ha tenido y tiene una gran influencia ecológica e histórica en la distribución de las asteráceas.
Existen asteráceas en la mayoría de los biomas, que se adaptan a diferentes suelos, climas y relieves. Hay especies típicas de suelos salinos (e.g., Baccharis spartioides , Cyclolepis genistoides , Lepidophyllum cupressiforme ), de suelos húmedos a inundables (e.g., Enhydra anagallis , Gymnocoronis spilanthoides , Senecio bonariensis ), de suelos rocosos (e.g., Eupatorium commersonii , Haplopappus rigidus ), algunas son dunícolas (e.g., Grindelia aegialitis , Noticastrum acuminatum , Senecio crassiflorus ), otras habitan las selvas (e.g., Adenostemma brasilianum , Mikania micrantha , Stevia jujuyensis ), las vegas altoandinas (e.g., muchas especies de Werneria , Senecio hieracium ), o las orillas de ríos (e.g., Gnaphalium gaudichaudianum , Tessaria dodoneifolia ).
Muchas asteráceas tienen adaptaciones particulares a estos distintos ambientes. En la provincia fitogeográfica de la Puna las asteráceas poseen adaptaciones al déficit de agua, baja humedad atmosférica, gran radiación solar y temperaturas nocturnas inferiores a cero durante todo el año, entre otros factores. Algunas adaptaciones morfológicas a la Puna son: tallos espiniformes ( Nardophyllum armatum , Proustia cuneifolia ), formación de braquiblastos ( Nassauvia axillaris ), plantas acaules ( Perezia ciliosa , Senecio breviscapus , Trichocline auriculata ), hojas pequeñas ( Baccharis incarum ), hojas carnosas (Xenophyllum papposum ), hojas enroscadas ( Baccharis grisebachii , Chiliotrichiopsis keidelii ), hojas espiniformes (Chuquiraga acanthophylla , Nassauvia axillaris ), hojas escamiformes (especies de Parastrephia ). También hay adaptaciones anatómicas, como por ejemplo estomas en criptas ( Chiliotrichiopsis keidelii , Baccharis polifolia ) y gran desarrrollo del esclerénquima ( Senecio dryophyllus ) (Cabrera, 1968). Algunas adaptaciones foliares en asteráceas de Patagonia son por ejemplo hojas angostas, y fuertemente recurvadas en los márgenes o conduplicadas ( Brachyclados caespitosus , Burkartia lanigera ) o con el mesófilo isolateral (Baccharis magellanica , Grindelia chiloensis , Haplopappus prunelloides ) (Pyykkö, 1966). La variación en la forma del limbo foliar en especies del género Chuquiraga (Barnadesieae) es concordante con diferencias en la altitud, de modo que las especies que poseen limbo foliar ancho habitan en las zonas altas de las montañas, las de hojas lanceoladas en la Puna , y las de hojas aciculares en las zonas desérticas de menor altitud (Ezcurra et al., 1997). También hay géneros de asteráceas con anatomía Kranz, como Chrysanthellum , Flaveria , Glossocardia , Isostigma , Parthenium y Pectis que puede variar con el ambiente. En el género Isostigma (Heliantheae) el tipo de anatomía Kranz en las distintas especies difiere de acuerdo al grado de humedad de las áreas que éstas ocupan (Peter & Katinas, 2003). El género Chaetanthera (Mutisieae) posee adaptaciones en sus granos de polen a los climas secos y fríos de las provincias fitogeográficas Altoandina y Puneña y al clima árido de tipo mediterráneo de Chile central (Tellería & Katinas, 2004). El polen de Chaetanthera muestra por ejemplo: 1) una exina gruesa con el tectum interno muy desarrollado en algunas especies, que aumentaría el área de resistencia del polen reduciendo la tendencia a encorvarse producida por las columelas, y esto sería crucial para la regulación del volumen del grano de polen; 2) las endoaperturas se unen en una banda equatorial o endocíngulo que incrementaría la plasticidad de las paredes para una rápida hidratación; y 3) algunas especies poseen parasincolpo que puede considerarse también como una estrategia de hidratación del polen porque las grietas de la exina que delimitan el campo apocolpial incrementarían la flexibilidad del grano de polen. De este modo Chaetanthera combina características contra la desecación con mecanismos que aseguran una rápida hidratación en ambientes muy secos.
Las asteráceas no son sólo diversas en número de especies y ambientes en los que habitan, sino que también desempeñan múltiples funciones en los ecosistemas. Como integrantes dominantes de los ecosistemas pueden mencionarse las asociaciones de Baccharis notosergila , con B. articulata y Eupatorium buniifolium en comunidades clímax en la provincia de Buenos Aires (Cabrera, 1949). Elementos característicos que forman poblaciones abundantes en la estepa patagónica son por ejemplo Nassauvia fuegiana y N. glomerulosa ; en el monte Chuquiraga erinacea ; en la Puna Parastrephia lepidophylla y especies de Nardophyllum ; y en la Prepuna Proustia cuneifolia.
Muchas especies son pioneras en sucesiones primarias de distintos ambientes, facilitando el establecimiento de otras especies al favorecer la maduración de los suelos. Algunos ejemplos son Elephantopus mollis, Eupatorium inulifolium, E. laevigatum, Ptercaulon lorentzii (Salimon & Negrelle, 2001), Sonchus oleraceus (Walker et al., 2006). Existen también entre las asteráceas especies con un alto grado de adaptación a diversos hábitats y que constituyen malezas muy invasoras y resistentes, por ejemplo en las tribus Cardueae ( Carduus acanthoides , C. pycnocephalus ; Cabrera, 1963) y Eupatorieae ( Ageratum conyzoides , Chromolaena odorata ; Retief, 2002).
Argentina: ¿Lugar de origen de las Asteráceas?
Los registros confiables más antiguos de polen fósil de asteráceas en Argentina son de Patagonia y corresponden a géneros afines a la tribu Mutisieae del período Oligoceno tardío-Mioceno (Barreda, 1993). Ello sumado a que la subfamilia Barnadesioideae, con gran representación en los Andes y en Patagonia, es considerada la más basal en el árbol evolutivo de las asteráceas, llevó a los investigadores a postular la hipótesis que el origen de la familia se halla en el sur de América del Sur. Bremer (1992) aplicó el método de biogeografía histórica de áreas ancestrales (Crisci et al., 2000, 2003) a un cladograma de Asteraceae basado en datos morfológicos para determinar el probable lugar de origen de la familia. Sus resultados indican a Brasil, al sur del Amazonas como parte del área ancestral más probable y excluyen de esta área a África y Eurasia. Stuessy et al. (1996) presentaron un escenario muy detallado del origen de la familia también sobre la base de un cladograma obtenido con datos morfológicos. Ellos postularon que las dos familias hermanas Asteraceae y Calyceraceae habrían divergido como familias separadas en el sur de América del Sur (Argentina y Chile) en un área de transición de bosques tropicales y templados en el Oligoceno temprano (38 millones de años atrás), en lo que es actualmente la estepa patagónica. Durante el Mioceno (25- 5 m .a.) comenzó la diferenciación de las Barnadesioideae en los géneros actuales. En este período el levantamiento andino creó condiciones ecológicas especiales para la creación de nuevos hábitats y oportunidades evolutivas. De acuerdo con Stuessy et al. (1996) el mejor candidato para un linaje temprano se hallaría en la tribu Mutisieae dada su similitud con Barnadesioideae. Los resultados de Gustafsson et al. (2001) sobre la base de datos moleculares coincide con Stuessy et al. (1996). Recientemente se ha reportado el descubrimiento de tipos polínicos relacionados a Mutisieae del Paleoceno Superior-Eoceno en Sudáfrica (Zavada & de Villiers, 2000; Scott et al., 2006). Estos autores postulan un posible origen africano de las asteráceas, pero estos registros necesitan una confirmación estratigráfica (Viviana Barreda, com. pers.). Funk et al. (2005) proponen que el origen de los miembros actuales de Asteraceae fue en el sur de América del Sur con una posterior radiación al África, dando así origen a la mayoría de las tribus que conocemos hoy.
AGRADECIMIENTOS
A los editores de este volumen al permitirnos homenajear con nuestro modesto aporte a Irma Josefa Gamundi. Se agradece a Hugo Calvetti por el delineado de la figura 4. Este trabajo fue realizado con los subsidios otorgados por The John Simon Guggenheim Foundation (LK), National Geographic Society (grant # 7646-04, JVC, LK), y el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
BIBLIOGRAFÍA
ARIZA ESPINAR, L. 1994. Familia Asteraceae: Tribu Vernonieae, Tribu Eupatorieae. Prodromo de Fl. Fanerog. de Argentina Central 1: 1-65. [ Links ]
ARIZA ESPINAR, L. 1997. Asteraceae, parte 7. Tribu VII. Anthemideae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 46: 3-35. [ Links ]
ARIZA ESPINAR, L. 2000. Familia Asteraceae: Tribu Heliantheae. Prodromo de Fl. Fanerog. de Argentina Central . 2: 1-111. [ Links ]
ARIZA ESPINAR, L. 2005. Familia Asteraceae: Tribu Astereae. Prodromo de Fl. Fanerog. de Argentina Central . 3: 1-139. [ Links ]
ARIZA ESPINAR, L. & G. DELUCCHI. 1998a. Asteraceae, parte 10. Tribu X. Arctoteae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 59: 1-6. [ Links ]
ARIZA ESPINAR, L. & G. DELUCCHI. 1998b. Asteraceae, parte 11. Tribu XI. Cardueae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 60: 1-26. [ Links ]
ARIZA ESPINAR, L. & J. L. NOVARA. 2005. Flora del Valle de Lerma. Asteraceae Dumort. Tribu. III. Astereae. Aportes Bot. Salta 7: 1-120. [ Links ]
ARIZA ESPINAR, L. & E. URTUBEY. 1998. Asteraceae, parte 12. Tribu XIII. Lactuceae (excepto Hypochoeris y Hieracium ). In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 61: 1-30. [ Links ]
BARREDA, V. D. 1993. Late Oligocene?- Miocene pollen of the families Compositae, Malvaceae, and Polygonaceae from the Chenque Formation, Golfo San Jorge Basin , southeastern Argentina . Palynology 17: 169-186. [ Links ]
BÁRTOLI, A. & R. TORTOSA. 2003. Asteraceae, parte 16. Tribu III. Astereae, Subtribus Bellidinae, Asterinae: Grindelia. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 81: 42-54. [ Links ]
BENTHAM, G. 1873. Compositae. In : BENTHAM, G. & HOOKER, J. D. (eds.), Genera Plantarum , vol. 2. Lovell Reeve, London . [ Links ]
BORTIRI, E. 1999. Asteraceae, parte 14. Tribu XIII. Lactuceae: Hypochoeris . In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 63: 3-25. [ Links ]
BRAKO, L. & J. ZARUCCHI. 1993. Catalogue of the flowering plants and Gymnosperms of Peru . Missouri Bot. Garden Press. St. Louis , Missouri . [ Links ]
BREMER, K. 1987. Tribal interrelationships of the Asteraceae. Cladistics 3: 210-253. [ Links ]
BREMER, K. 1992. Ancestral areas: A cladistic reinterpretation of the center of origin concept. Syst. Biol. 41: 436-445. [ Links ]
BREMER, K. 1994. Asteraceae: Cladistics and classification . Timber Press, Portland , Oregon . [ Links ]
CABRERA, A. L. 1949. Las comunidades vegetales de los alrededores de La Plata (Prov. de Buenos Aires, Rep. Argentina). Lilloa 20: 269-376. [ Links ]
CABRERA, A. L. 1961. Compuestas Argentinas: Clave para la determinación de los géneros. Revista Mus. Argent. Ci. Nat., Bernardino Rivadavia Inst. Nac. Invest. Ci. Nat., Bot . 2: 291-362. [ Links ]
CABRERA, A. L. 1963. Compositae. In : CABRERA, A. L. (ed.), Fl. Prov. Buenos Aires , Colecc. Ci. Inst. Nac. Tecnol. Agropecu. 4(6a): 1-443. [ Links ]
CABRERA, A. L. 1968. Geo-ecología de las regiones montañosas de las Américas tropicales. Colloq. Geogr. 9: 91-116. [ Links ]
CABRERA, A. L. 1971a. Compositae. In : CORREA, M. N. (ed.), Fl. Patagónica , Colecc. Ci. Inst. Nac. Tecnol. Agropecu. 8(7): 1-451. [ Links ]
CABRERA, A. L. 1971b. Fitogeografía de la República Argentina. Bol. Soc. Argent. Bot . 14: 1-42. [ Links ]
CABRERA, A. L. 1974. Compositae. In : BURKART, A. (ed.), Fl. Il. Entre Ríos , Colecc. Ci. Inst. Nac. Tecnol. Agropecu. 6(6a): 106-554. [ Links ]
CABRERA, A. L. 1977. Mutisieae- Systematic review. In: HEYWOOD, V. H., J. B. HARBORNE & B. L. TURNER (eds.), The Biology and Chemistry of the Compositae , vol. 2, pp. 1039-1066. Academic Press, London. [ Links ]
CABRERA, A. L. 1978. Compositae. In : CABRERA, A. L. (ed.), Fl. Prov. Jujuy , Colecc. Ci. Inst. Nac. Tecnol. Agropecu. 13(10): 1-726. [ Links ]
CABRERA, A. L. & S. E. FREIRE. 1997. Asteraceae, parte 8. Tribu II. Eupatorieae (excepto Mikania ). In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 47: 3-54, 76-104. [ Links ]
CABRERA, A. L., S. E. FREIRE & L. ARIZA ESPINAR. 1999. Asteraceae, parte 13. Tribu VIII. Senecioneae. Tribu VIII bis. Liabeae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 62: 3-180. [ Links ]
CABRERA, A. L., J. V. CRISCI, G. DELUCCHI, S. E. FREIRE, D. A. GIULIANO, L. IHARLEGUI, L. KATINAS, A. A. SÁENZ, G. SANCHO & E. URTUBEY. 2000. Catálogo ilustrado de las Compuestas (= Asteraceae) de la Provincia de Buenos Aires, Argentina: Sistemática, Ecología y Usos . COBIOBO (Comisión de Biodiversidad Bonaerense), Nº 2, PROBIOTA (Programa para el Estudio y Uso Sustentable de la Biota Austral ) Nº 1, Convenio Secretaría de Política Ambiental, U.N.L.P, Provincia de Buenos Aires, Argentina. [ Links ]
CANDOLLE, A. P. DE. 1838. Prodromus Systematis Naturalis Regni Vegetabilis , vol. 7. Paris. [ Links ]
CASSINI, H. 1816. Quatriéme memoire sur le famille des synantherées, contenant l'analyse de l'ovaire et de ses accesoires. J. Phys. Chim. Hist. Nat. Arts 85: 5-21. [ Links ]
CERANA, M. M. 1997. Asteraceae, parte 8. Tribu II. Eupatorieae: Mikania . In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 47: 54-76. [ Links ]
CRISCI, J. V., L. KATINAS & P. POSADAS. 2000. Introducción a la teoría y práctica de la biogeografía histórica . Sociedad Argentina de Botánica, Buenos Aires, Argentina. [ Links ]
CRISCI, J. V., L. KATINAS & P. POSADAS. 2003. Historical biogeography: An Introduction. Harvard University Press, Boston, Massachusetts. [ Links ]
DELUCCHI, G. & L. ARIZA ESPINAR. 1997. Asteraceae, parte 9. Tribu IX. Calenduleae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 48: 2-4. [ Links ]
DUMORTIER, B. C. J. 1822. Observations botaniques, dédiées à la Societé d'Horticulture de Tournay . Tournay, France. [ Links ]
EZCURRA, C. 1996. Asteraceae, parte 5. Tribu XII. Mutisieae. Subtribu 1. Barnadesiinae: Chuquiraga . In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 31: 5-13. [ Links ]
EZCURRA, C., A. RUGGIERO & J. V. CRISCI. 1997. Phylogeny of Chuquiraga sect. Acanthophyllae (Asteraceae-Barnadesioideae), and the evolution of its leaf morphology in relation to climate. Syst. Bot . 22: 151-163. [ Links ]
FREIRE, S. E. 1995. Asteraceae, parte 2. Tribu IV. Inuleae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 14: 3-60. [ Links ]
FREIRE, S. E. 1996. Asteraceae, parte 3. Tribu XII. Mutisieae, Subtribu 2. Gochnatiinae (excepto Gochnatia ). In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 28: 3-8, 12-17. [ Links ]
FREIRE, S. E., G. SANCHO, E. URTUBEY, N. BAYÓN, L. KATINAS, D. GIULIANO, D. GUTIÉRREZ, A. A. SÁENZ, L. IHARLEGUI, C. MONTI & G. DELUCCHI. 2005. Catalogue of Asteraceae of Chacoan plain, Argentina. Compositae Newslett. 43: 1-126. [ Links ]
FUNK, V. A., R. J. BAYER, S. KEELEY, R. CHAN, L. WATSON, B. GEMEINHOLZER, E. SCHILLING, J. L. PANERO, B. G. BALDWIN, N. GARCÍA-JACAS, A. SUSANNA & R. K. JANSEN. 2005. Everywhere but Antarctica: Using a supertree to understand the diversity and distribution of the Compositae. Biol. Skr . 55: 343- 374. [ Links ]
GISEKE, P. D. 1792. Praelectiones in ordines naturales plantarum . Hamburg, Germany. [ Links ]
GIULIANO, D. A. 2000. Asteraceae, parte 15. Tribu III. Astereae, parte A. Subtribu c. Baccharinae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 66: 3-73. [ Links ]
GREUTER, W., J. MCNEILL, F. R. BARRIE, H. M. BURDET, V. DEMOULIN, T. S. FILGUEIRAS, D. H. NICOLSON, P. C. SILVA, J. E. SKOG, P. TREHANE, N. J. TURLAND & D. L. HAWKSWORTH. 2000. International Code of Botanical Nomenclature (St. Louis Code) , adopted by the Sixteenth International Botanical Congress, St. Louis, Missouri, July-August 1999. Regnum Vegetabile 138. Koeltz Scientific Books, Königstein. [ Links ]
GUSTAFSSON, M. H. G., A. S. R. PEPPER, V. A. ALBERT & M. KÄLLERSJÖ. 2001. Molecular phylogeny of the Barnadesioideae (Asteraceae). Nord. J. Bot . 21: 149-160. [ Links ]
JANSEN, R. K. & J. D. PALMER. 1987. A chloroplast DNA inversion marks an ancient evolutionary split in the sunflower family (Asteraceae). Proc. Natl. Acad. Sci. U.S.A . 84: 5818-5822. [ Links ]
JØRGENSEN, P. M. & S. LEÓN-YÁÑEZ (eds.). 1999. Catalogue of the vascular plants of Ecuador . Monogr. in Syst. Bot. Missouri Bot. Garden 75: 1-1181. [ Links ]
KATINAS, L. 1995. Asteraceae, parte 1. Tribu XII. Mutisieae. Subtribu 4. Nassauviinae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 13: 5-54. [ Links ]
KATINAS, L. 1996. Asteraceae, parte 4. Tribu XII. Mutisieae. Subtribu 3. Mutisiinae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 29: 3-40. [ Links ]
KATINAS, L., J. PRUSKI, G. SANCHO, S. E. FREIRE & M. C. TELLERÍA. The subfamily Mutisioideae (Asteraceae). Monogr. in Syst. Bot. Missouri Bot. Garden 107 (en prensa). [ Links ]
KING, R. M. & H. ROBINSON. 1970. The new synantherology. Taxon 19: 6-11. [ Links ]
LESSING, C. F. 1832. Synopsis Generum Compositarum . Duncker & Humblot, Berlin. [ Links ]
MC VAUGH, R. 1982. The new synantherlogy vs. Eupatorium in Nueva Galicia. Contr. Univ. Mich. Herb . 15: 181-190. [ Links ]
NOVARA, L. J. 2003. Catálogo de la flora de la Puna en el noroeste argentino. Aportes Bot. Salta, Ser. Misceláneas 2: 1-56. [ Links ]
PANERO, J. L. & V. A. FUNK. 2002. Toward a phylogenetic subfamilial classification for the Compositae (Asteraceae). Proc. Biol. Soc. Wash . 115: 909-922. [ Links ]
PETENATTI, E. M. & L. ARIZA ESPINAR. 1997. Asteraceae, parte 6. Tribu VI. Helenieae. In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 45: 3-35. [ Links ]
PETER, G. & L. KATINAS. 2003. A new type of Kranz anatomy in Asteraceae. Austral. J. Bot. 51: 217-226. [ Links ]
PYYKKÖ, M. 1966. The leaf anatomy of East Patagonian xeromorphic plants. Ann. Bot. Fenn. 3: 453-622. [ Links ]
RETIEF, E. 2002. The tribe Eupatorieae (Asteraceae) in Southern Africa. In : ZACHARIADES, C., R. MUNIAPPAN & L.W. STRATHIE (eds.), Proccedings of the fifth international workshop on biological control and management of Chromolaena odorata , pp. 81-89. Durban, South Africa. [ Links ]
SALIMON, C. I. & R. R. B. NEGRELLE. 2001. Natural regeneration in a Quaternary coastal plain in southern Brazilian Atlantic rain forest. Brazilian Archives of biology and technology 44: 155-163. [ Links ]
SANCHO, G. 1996. Asteraceae, parte 4. Tribu XII. Mutisieae, Subtribu 2. Gochnatiinae: Gochnatia . In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 28: 9-11. [ Links ]
SANCHO, G. & L. ARIZA ESPINAR. 2003. Asteraceae. Tribu III. Astereae, parte B. Subtribus Bellidinae, Asterinae (excepto Grindelia y Haplopappus ). In : HUNZIKER, A. T., Flora Fanerogámica Argentina 81: 3-42, 54-58, 64- 102. [ Links ]
SCOTT, L., A. CADMAN & I. MCMILLAN. 2006. Early history of Cainozoic Asteraceae along the Southern African west coast. Rev. Palaeobot. Palynol . 142: 47-52. [ Links ]
STUESSY, T. F., T. SANG & M. L. DEVORE. 1996. Phylogeny and biogeography of the subfamily Barnadesioideae with implications for early evolution of the Compositae. In : HIND, D. J. N. & H. J. BEENTJE (eds.), Compositae: Systematics vol. 1, pp. 463-490. Proceedings of the International Compositae Conference, Kew, 1994, Royal Botanic Gardens, Kew. [ Links ]
TELLERÍA, M. C. & L. KATINAS. 2004. A comparative palynologic study of Chaetanthera (Asteraceae, Mutisieae) and allied genera. Syst. Bot. 29: 752-773. [ Links ]
TROIANI, H. O. & P. H. STEIBEL. 1999. Sinopsis de las Compuestas (Compositae Giseke) de la provincia de La Pampa, Rep. Argentina. Rev. Fac. Agronomía 10, Supl. 1: 1-90. [ Links ]
URTUBEY, E. 1996. Asteraceae, parte 5. Tribu XII. Mutisieae. Subtribu 1. Barnadesiinae (excepto género Chuquiraga ). In : HUNZIKER, A. T. (ed.), Flora Fanerogámica Argentina 31: 3-4, 14-24. [ Links ]
VALENCIA, R., N. PITMAN, S. LEÓN-YÁÑEZ & P. M. JØRGENSEN (eds.). 2000. Libro rojo de las plantas endémicas del Ecuador 2000 . Herbario QCA, Pontificia Universidad Católica del Ecuador, Quito. [ Links ]
VÁSQUEZ, R., R. ROJAS & E. RODRÍGUEZ. 2002. Adiciones a la flora peruana: Especies nuevas, nuevos registros y estados taxonómicos de las Angiospermas para el Perú. Arnaldoa 9: 43-110. [ Links ]
WALKER, L. R., P. J. BELLINGHAM & D. A. PELTZER. 2006. Plant characteristics are poor predictors of microsite colonization during the first two years ofprimary succession. J. Veg. Sci. 17: 397-406. [ Links ]
ZAVADA, M. S. & S. E. DE VILLIERS. 2000. Pollen of the Asteraceae from the Paleocene-Eocene of South Africa. Grana Palynol . 39: 39-45. [ Links ]
ZULOAGA, F. O. & O. MORRONE (eds.). 1999. Catálogo de las plantas vasculares de la República Argentina. II. Dicotyledoneae. Monogr. Syst. Bot. Missouri Bot. Garden . 74: 1-1269. [ Links ]
ZULOAGA, F. O., O. MORRONE & D. RODRÍGUEZ. 1999. Análisis de la biodiversidad en plantas vasculares de la Argentina. Kurtziana 27: 17-167. [ Links ]
Recibido el 04 de Diciembre de 2006,
aceptado el 27 de Marzo de 2007.

