










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana vol.47 no.2 Buenos Aires jun. 2010
ARTÍCULOS ORIGINALES
Middle-Upper Devonian palynoflora from the Tonono x-1 borehole, Salta Province, northwestern Argentina
Sol Noetinger1
1CONICET-UBA. Facultad de Ciencias Exactas y Naturales, Departamento de Geología, Universidad Nacional de Buenos Aires. Intendente Güiraldes 2160, Ciudad Universitaria, 1428 Buenos Aires, Argentina. snoetinger@gl.fcen.uba.ar.
Abstract. The surveyed microflora was recovered from six cores from the Tonono x-1 well in northwestern Argentina. Most of the studied interval corresponds to the Tonono Formation, i.e., the upper part to the Jollín Member and the basal part of the interval may be related to the Michicola Formation. The microflora totals 73 species represented by trilete spores (35 species), microplankton (30 species) including prasinophycean, acritarch and chlorophycean algae and chitinozoans (8 species). The stratigraphic distribution of these species allowed the definition of three associations. The palynoassemblage To1 (3946,5 - 3638,5 m) is characterized by species such as Grandispora douglastownense Mc Gregor, Dibolisporites eifeliensis (Lanninger) McGregor, Verrucosisporites sp. cf. V. loboziakii Marshall and Fletcher, Alpenachitina matogrossensis Burjack and Paris, Alpenachitina sp. cf. A. eisenacki Dunn and Miller and Ancyrochitina simplex Grahn, Bergamaschi and Pereira suggesting a late Eifelian to earliest Givetian age. Based on the presence of Geminospora lemurata Balme, Aneurospora greggsii (McGregor) Streel, Biharisporites parviornatus Richardson, Raistrickia aratra Allen and Leiotriletes balapucencis di Pasquo a Givetian age is proposed for palynoassemblage To2 (3367,35 - 3285 m). Palynoassemblage To3 (3073,2 - 3137,5 m) is predominantly composed of marine elements together with AOM. Few key species are recognized; however the presence of Acinosporites sp. cf. A. eumammillatus Loboziak, Streel and Burjack together with the chitinozoans Angochitina katzeri Grahn and Melo and Angochitina mourai Lange support an early Frasnian age. The distinctive increase of marine elements and AOM between To2 and To3 suggests that a maximum flooding event would have occurred there during this period.
Resumen. Palinoflora del devónico medio-superior del pozo Tonono x-1, Provincia de Salta, Noroeste de Argentina. Se presenta el estudio de seis coronas del pozo Tonono x-1 ubicado en el noroeste Argentino. La mayor parte del intervalo estudiado corresponde a la Formación Tonono; la porción superior al Miembro Jollín y la porción basal podría relacionarse con la Formación Michicola. La microflora recuperada se compone de 73 especies representadas por diversos grupos palinológicos como esporas trilete (35 especies), microplancton incluyendo prasinofitas, acritarcas, algas clorofitas (30 especies) y quitinozoos (8 especies). La distribución estratigráfica de las especies permite distinguir tres asociaciones, la asociación To1 (3946,5 - 3638,5 m), caracterizada por Grandispora douglastownense Mc Gregor, Dibolisporites eifeliensis (Lanninger) McGregor, Verrucosisporites sp. cf. V. loboziakii Marshall y Fletcher, Alpenachitina matogrossensis Burjack y Paris, Alpenachitina sp. cf. A. eisenacki Dunn y Miller y Ancyrochitina simplex Grahn, Bergamaschi y Pereira sugieren una edad eifeliana tardía-givetiana temprana. Debido a la presencia de Geminospora lemurata Balme, Aneurospora greggsii (McGregor) Streel, Biharisporites parviornatus Richardson, Raistrickia aratra Allen y Leiotriletes balapucencis di Pasquo se propone una edad givetiana para la asociación To2 (3367,35 - 3285 m). La asociación To3 (3073,2 - 3137,5 m) comprende predominantemente elementos marinos incluyendo MOA. Se reconocen las siguientes especies clave Acinosporites sp. cf. A. eumammillatus Loboziak, Streel y Burjack junto a los quitinozoos Angochitina katzeri Grahn y Melo y Angochitina mourai Lange las cuales sugieren una edad frasniana temprana. El notable incremento de material de origen marino y MOA entre To2 y To3 sugiere que un evento de máxima inundación podría haber ocurrido allí durante este período.
Key words. Palynostratigraphy; Mid-Late Devonian; Tonono Formation; Salta; Argentina.
Palabras clave. Palinoestratirgrafía; Devónico Medio-Tardío; Formación Tonono; Salta; Argentina.
Introduction
The Tarija basin comprises sedimentary rocks of Silurian to Recent age, which were deposited during several sedimentary cycles. In northwestern Argentina, the Silurian-Devonian unit occurs along an East-West axis and is represented in subsurface mostly by shale and sandstone facies that were mainly deposited in a shallow marine environment. These shales and sandstones increases to over 2000 meters in thickness in the Subandean area and the Chaco- Salteño Plain. Towards the south, these sediments become more continental in character (Disalvo, 2002). Many multidisciplinary works based on subsurface and surface information have been carried out by different oil companies, although, published accounts are less numerous (e.g., Suárez Soruco 2000; Dalenz Farjat et al., 2002; Albariño et al. 2002; Álvarez et al., 2003; Vistalli et al., 2005). The Chaco-Salteño Plain was geophysically surveyed as part of an oil exploration programme. This resulted in the discovery of small, oil/ gas hydrocarbon, fields. The Tonono Field (figure 1A) was discovered in 1969 and holds over 100,000 m3 of condensate (Disalvo, 2002).

Figure 1. A, Distribution of Palaeozoic surface deposits in the Province of Salta, northern Argentina and location of the studied Tonono x-1 Borehole. B, Comparative stratigraphy of the study area modified from Starck (1999)./ A, Distribución de los depósitos de superficie Paleozoicos en la provincia de Salta, al norte de Argentina y localización del pozo Tonono x-1. B, Estratigrafía comparada del área de estudio, modificada de Starck (1999).
This contribution presents a detailed palynological survey of a Devonian palynoflora (microspores, megaspores and microplankton) recovered from six cores of the 3945-3077 m interval in Tonono x-1 borehole. The stratigraphic distribution of the species is analysed and compared with their global ranges to assess the age and correlation of the assemblages defined in this interval. Palynofacies are also considered in order to better understand previous palaeoenvironmental interpretations.
Summary of the stratigraphy and paleontology
The stratigraphic units of the Late Silurian to Devonian rocks from northern Argentina and southern Bolivia have been assigned to supersequence hierarchies by Starck (1995, 1999). This includes a Silurian-Jurassic tectonic-stratigraphic interval that is divided into two units separated by a regional unconformity at the end of the Devonian. In the first Silurian-Devonian unit, the Cinco Picachos, Las Pavas and Aguaragüe Supersequences are characterized by stacked, kilometer scale, coarsening-upward shale and sandstone facies bounded by first order flooding surfaces. In this scheme, on the Chaco-Salteño Plain, the Tonono Formation is part of the Aguaragüe Supersequence, conformably overlying the Michicola and Rincón Formations, which in turn are part of the Las Pavas Supersequence (Starck, 1995) (figure 1B). Lithologies in the subsurface Rincón Formation include shales containing macrofossils such as Metacryphaeus sp., Calmonia subcesiva, Australocoelia tourtelotti and palaeomicroplankton suggesting a time range from the Emsian to the Givetian (Cuerda and Baldis, 1971; Russo et al., 1979, Aceñolaza et al., 2000, Grahn and Gutierrez, 2001, Grahn, 2003, Antonelli and Ottone, 2006). The Michicola Formation comprises a dozen meters of whitish-grey mostly silicified quartzites (Russo et al., 1979) and is considered, by some authors a facies variant of the Rincón Formation (see Aceñolaza et al., 2000). The Michicola Formation is correlated with the quartzites of Cerro León in Paraguay, which are supposed to be Silurian (Padula et al., 1967).
The subsurface Tonono Formation shows a sucession of dark grey to black, laminated and fissile, very micaceous, bituminous and carbonaceous shales with variable sand or silt content. Grey to whitish sandstones appear in very thin individual layers. Its thickness varies between 710 and 1960 meters; such variation is due to erosion during the Chanic orogenic phase. The uppermost part of this formation is dominated by quartz sandstones; it is known as the Jollín Sandstone Member (Padula et al., 1967) and is correlated with the lower part of the Bolivian Iquiri Formation dated as Frasnian (Dávila and Rodríguez, 1967). Several unpublished accounts assign an Emsian- Eifelian to Givetian-Frasnian age for the Tonono Formation based on its palaeontological content, which includes the remains of cf. Cyclostigma sp. and an undescribed trilobite (see Padula et al., 1967; Böttcher et al., 1984). Azcuy and Laffitte (1981) described the "association 0" from an unidentified sample from the Tonono x-1 containing several chitinozoans and other microplankton such as acritarchs, chlorophycean algae (e.g., Chomotriletes sp.), prasinophytes (e.g., Maranhites spp.) and scant scolecodonts. Within this assemblage there are spores such as Emphanisporites sp. cf. E. rotatus and Grandispora pseudoreticulata among others. This group is recognized as Upper Devonian. Barreda (1986) studied a complete Givetian-Frasnian assemblage from the Tonono Formation on the northeastern Chaco-Salteño Plain -including microplankton, spores and chitinozoans- but only published the acritarch association. Volkheimer et al. (1986) also studied the Tonono Formation from three samples from the Ramos x-1 borehole and one from the Tonono x-1, where species such as Sphaerochitina sp., Sphaerochitina sp. cf. S. schwalbi and Gotlandochitina sp. occur, assigning a Middle Devonian for the upper level of the formation. Later on, di Pasquo (2003) recognized the Macharetí Group unconformably overlying the Tonono Formation in this borehole and assigned it an early Pennsylvanian age based on palynology.
Materials and methods
The Tonono x-1 (To) well (c.a. 63º 38' W, 22º 17' S) is located in the Chaco-Salteño Plain (figure 1.A). It was drilled to a total depth of 4032 meters (13228 feet). Forty-seven core samples were collected from several intervals. One sample from the Upper Devonian section was studied by Azcuy and Laffitte (1981) and Volkheimer et al. (1986). Di Pasquo (2003) studied in detail five core samples (2984 to 2656 m) that yielded diagnostic palynomorphs of the overlying RS and BC Biozones akin to the early Pennsylvanian. An abstract was presented by Noetinger and di Pasquo (2008a) of six core samples between 3073 m and 3945 m depth (figure 2). A detailed survey of these is the subject of this work.

Figure 2. Lithostratigraphic scheme together with the wireline log of the Tonono x-1 well and the palynological sample depths / esquema litoestratigráfico junto al perfil eléctrico del pozo Tonono x-1 y las profundidades de las muestras palinológicas.
Standard palynological methods were performed to obtain organic residues from the core samples. These were crushed and then treated first with hydrochloric and then with hydrofluoric acid to remove carbonate, silica and silicates, respectively, sieved with a 25 µm mesh and finally mounted on slides with glycerine jelly.
Eight types of dispersed organic matter and palynomorph groups were identified in this study, including amorphous organic matter (AOM); structured phytodebris (SP), characterized by identifiable cuticles and wood; unstructured phytodebris (USP), known as gelified matter; black phytodebris (BP) or opaque clasts including charcoal; spores (SPO); phytoplankton (PHY) comprising acritarchs, prasinophycean and chlorophycean algae and chitinozoans (CHI). This is a simplified scheme adapted from Tyson (1995) for the types of organic components and palynomorphs found in the analysed assemblages used to calculate relative percentages allowing the definition of different palynofacies. Statistical analyses were also carried out to identify the subtle differences in the composition of the assemblages. A palynological marine index (PMI= (Marine Richness (Rm)/ Terrestrial Richness (Rt) + 1) 100, Helenes et al. 1998) was also calculated in order to support the interpretation of depositional environments. The values used are stipulated as in de Araujo Carvalho et al. (2006), where the marine and terrestrial richness were expressed as number of genera per sample. High values of PMI are interpreted as indicative of normal marine depositional conditions. As the PMI is based on the palynomorph diversity of terrestrial and marine palynomorph, it is therefore used as a substitute for terrestrial/marine ratio.
The identification of palynomorphs was undertaken using both Leitz Orthoplan and Nikon Eclipse 80i trinocular transmitted light microscopes, with x1000 maximum magnification. The photomicrographs were obtained with Motic (2.0 megapixels) and Pax-it (3.1 megapixels) videocameras and the illustrations are labelled with BAFC-Pl numbers followed by the England Finder reference. The studied samples are deposited at the Department of Geology of the Facultad de Ciencias Exactas, Físicas y Naturales (University of Buenos Aires).
Systematic paleontology
List of species
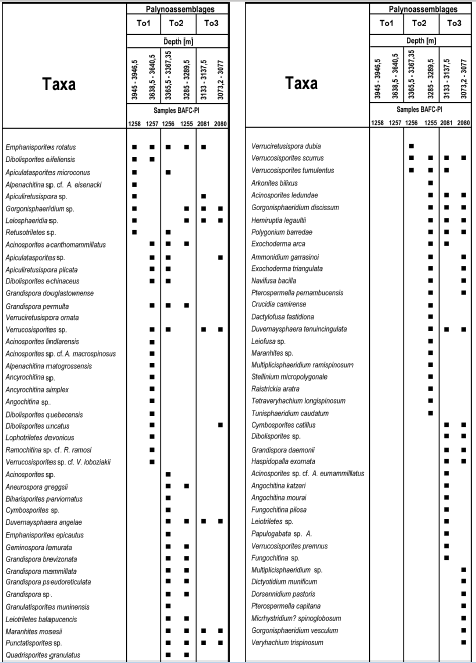
The identified palynomorph taxa are reported by major groups and in alphabetical order. Some specimens not illustrated are grouped at generic level because their poor preservation prevents a more specific assignment. Nevertheless, they are included in the stratigraphic distribution of the assemblages (figure 3), and some of them are figured together with the rest of the species as indicated in brackets (figures 4-8).
Figure 3. Stratigraphic distribution of the species recorded in the Tonono x-1 well / distribución estratigráfica de las especies en el pozo Tonono x-1.

Figure 4. 1, Acinosporites acanthomammillatus Richardson BAFC-Pl 1257 (1) L34/2. 2, Acinosporites ledundae Ottone BAFC-Pl 2081 (1) P43. 3, Acinosporites lindlarensis Riegel BAFC-Pl 1257 (1) V42/3. 4, Acinosporites sp. cf. A. eumammillatus Loboziak, Streel and Burjack BAFC-Pl 2081 (1) C38/1. 5, Acinosporites sp. cf. A. macrospinosus Richardson BAFC-Pl 1257 (1) M38/4. 6, Aneurospora greggsii (McGregor) Streel BAFC-Pl 1256 (1) Y56. 7, Apiculatasporites microconus (Richardson) McGregor and Camfield BAFC-Pl 1258 (2) S18. 8, Apiculiretusispora plicata (Allen) Streel BAFC-Pl 1257 (1) F28/3. 9, Biharisporites parviornatus Richardson BAFC-Pl 1256 (1) U19. 10, Cymbosporites catillus Allen BAFC-Pl 2080 (1) A46/2. 11, Dibolisporites echinaceus (Eisenack) Richardson BAFC-Pl 1257 (1) Y24/1. 12, Dibolisporites eifeliensis (Lanninger) McGregor BAFC-Pl 1257 (2) D49/3. 13, Dibolisporites quebecensis McGregor BAFC-Pl 1257 (1) H32/2. 14, Dibolisporites uncatus (Naumova) McGregor and Camfield BAFC-Pl 1257 (1) N45/4. 15, Emphanisporites epicautus Richardson and Lister BAFC-Pl 1256 (2) U19. 16, Emphanisporites rotatus McGregor emend. McGregor BAFC-Pl 1255 (1) C46/2. 17, Geminospora lemurata Balme BAFC-Pl 1255 (1) D57/1. Scale bar/ escala gráfica: 1, 2, 3, 5, 6, 7, 9, 11= 15 ?m. 4, 8, 10, 12, 13, 14, 15, 16, 17= 10 µm.
Trilete spores
Acinosporites acanthomammillatus Richardson 1965 (figure 4.1)
Acinosporites ledundae Ottone 1996 (figure 4.2)
Acinosporites lindlarensis Riegel 1968 (figure 4.3)
Acinosporites sp. cf. A. eumammillatus Loboziak, Streel and Burjack 1988 (figure 4.4)
Acinosporites sp. cf. A. macrospinosus Richardson 1965 (figure 4.5)
Aneurospora greggsii (McGregor) Streel in Becker, Bless, Streel and Thorez 1974 (figure 4.6)
Apiculatasporites microconus (Richardson) McGregor and Camfield 1982 (figure 4.7)
Apiculiretusispora plicata (Allen) Streel 1967 (figure 4.8)
Biharisporites parviornatus Richardson 1965 (figure 4.9)
Cymbosporites catillus Allen 1965 (figure 4.10)
Dibolisporites echinaceus (Eisenack) Richardson 1965 (figure 4.11)
Dibolisporites eifeliensis (Lanninger) McGregor 1973 (figure 4.12)
Dibolisporites quebecensis McGregor 1973 (figure 4.13)
Dibolisporites uncatus (Naumova) McGregor and Camfield 1982 (figure 4.14)
Dibolisporites spp.
Emphanisporites epicautus Richardson and Lister 1969 (figure 4.15)
Emphanisporites rotatus McGregor emend. McGregor 1973 (figure 4.16)
Geminospora lemurata Balme 1962 (figure 4.17)
Grandispora brevizonata (Menéndez and Pöthe de Baldis) di Pasquo 2007a (figure 5.1)
Grandispora daemonii Loboziak, Streel and Burjack 1988 (figure 5.2)
Grandispora douglastownense McGregor 1973 (figure 5.3)
Grandispora mammillata Owens 1971 (figure 5.4)
Grandispora permulta (Daemon) Loboziak, Streel and Melo 1999 (figure 5.5)
Grandispora pseudoreticulata (Menéndez and Pöthe de Baldis) Ottone 1996 (figure 5.6)
Grandispora spp.
Granulatisporites muninensis Allen 1965 (figure 5.7)
Leiotriletes balapucensis di Pasquo 2007a (figure 5.8)
Leiotriletes spp.
Lophotriletes devonicus (Naumova ex Chibrikova) McGregor and Camfield 1982 (figure 5.9)
Punctatisporites spp. (figure 5.10)
Raistrickia aratra Allen 1965 (figure 5.11)
Retusotriletes spp. (figure 5.12)
Verruciretusispora dubia (Eisenack) Richardson and Rasul 1978 (figure 5.13)
Verruciretusispora ornata (Menéndez and Pöthe de Baldis) Pérez Leyton ex di Pasquo 2007a (figure 5.14)
Verrucosisporites premnus (Richardson) Richardson 1965 (figure 5.15)
Verrucosisporites scurrus (Naumova) McGregor and Camfield 1982 (figure 5.16, 17)
Verrucosisporites sp. cf. V. loboziakii Marshall and Fletcher 2002 (figure 5.18)
Verrucosisporites tumulentus Clayton and Graham 1974 (figure 5.19)
Verrucosisporites spp.

Figure 5. 1, Grandispora brevizonata (Menéndez and Pöthe de Baldis) di Pasquo BAFC-Pl 1255 (2) G49/4. 2, Grandispora daemonii Loboziak, Streel and Burjack BAFC-Pl 2081 (2) J19. 3, Grandispora douglastownense McGregor BAFC-Pl 1256 (1) C21/3. 4, Grandispora mammillata Owens BAFC-Pl 1256 (1) H49/2. 5, Grandispora permulta (Daemon) Loboziak, Streel and Melo BAFC-Pl 1255 (1) F44. 6, Grandispora pseudoreticulata (Menéndez and Pöthe de Baldis) Ottone BAFC-Pl 1256 (1) R33. 7, Granulatisporites muninensis Allen BAFC-Pl 1256 (1) R31. 8, Leiotriletes balapucensis di Pasquo BAFC-Pl 1255 (1) L61. 9, Lophotriletes devonicus (Naumova ex Chibrikova) McGregor and Camfield BAFC-Pl 1257 (1) W20/2. 10, Punctatisporites sp. BAFC-Pl 2081 (2) B22/3. 11, Raistrickia aratra Allen BAFCPl 1255 (1) J59. 12, Retusotriletes sp. BAFC-Pl 1258 (1) V43/1. 13, Verruciretusispora dubia (Eisenack) Richardson and Rasul BAFC-Pl 1256 (1) Y22/4. 14, Verruciretusispora ornata (Menéndez and Pöthe de Baldis) Pérez Leyton ex di Pasquo BAFC-Pl 1256 (1) U43/3. 15, Verrucosisporites premnus (Richardson) Richardson BAFC-Pl 2081 (1) W35/1. 16, 17, Verrucosisporites scurrus (Naumova) McGregor and Camfield BAFC-Pl 1256 (1) Y28/2, BAFC-Pl 2080 (1) G49/3. 18, Verrucosisporites sp. cf. V. loboziakii Marshall and Fletcher BAFCPl 1257 (1) M24. 19, Verrucosisporites tumulentus Clayton and Graham BAFC-Pl 1255 (1) C53/1. Scale bar/ escala gráfica: 1, 3, 4, 6, 8, 10, 11, 13= 20 µm. 2, 5, 7, 12, 15, 18= 15 µm. 9, 14, 16, 17, 19= 10 µm.
Phytoplankton
Ammonidium garrasinoi Ottone 1996 (figure 6.1)
Arkonites bilixus Legault 1973 (figure 6.2)
Crucidia camirense (Lobo Boneta) Ottone 1996 (figure 6.4)
Dactylofusa fastidiona (Cramer) Fensome, Williams, Barss, Freeman and Hill 1990 (figure 6.5)
Dictyotidium munificum (Wicander and Wood) R. Amenábar, di Pasquo, Carrizo and Azcuy 2006 (figure 6.6)
Dorsennidium pastoris (Deunff) Sarjeant and Stancliffe 1994 (figure 6.7)
Duvernaysphaera angelae Deunff 1964 (figure 6.8)
Duvernaysphaera tenuicingulata Staplin 1961 (figure 6.9)
Exochoderma arca Wicander and Wood 1981 (figure 6.10)
Exochoderma triangulata Wicander and Wood 1981 (figure 6.11)
Gorgonisphaeridium discissum Playford in Playford and Dring 1981 (figure 6.12)
Gorgonisphaeridium vesculum Playford in Playford and Dring 1981 (figure 6.13)
Gorgonisphaeridium spp.
Haspidopalla exornata (Deunff) Playford 1977 (figure 6.14)
Hemiruptia legaultii Ottone 1996 (figure 6.15)
Leiofusa sp. (figure 7.1)
Leiosphaeridia spp. (figure 7.2)
Maranhites mosesii (Sommer) Brito emend. Burjack and Oliveira emend. González 2009 (figure 7.3)
Maranhites spp.
Micrhystridium? spinoglobosum (Staplin) Sarjeant and Stancliffe 1994 (figure 7.4)
Multiplicisphaeridium ramispinosum (Staplin) Sarjeant and Vavrdová 1997 (figure 7.5)
Multiplicisphaeridium sp. (figure 6.3)
Navifusa bacilla (Deunff) Playford 1977 (figure 7.6)
Papulogabata sp. A (figure 7.7)
Polygonium barredae Ottone 1996 (figure 7.8)
Pterospermella capitana Wicander 1974 (figure 7.9)
Pterospermella pernambucensis (Brito) Eisenack, Cramer and Diez Rodriguez 1973 (figure 7.10)
Quadrisporites granulatus (Cramer) Ströther 1991 (figure 7.11)
Stellinium micropolygonale (Stockmans and Willière) Playford 1977 (figure 7.12)
Tunisphaeridium caudatum Deunff and Evitt 1968 (figure 7.13)
Veryhachium (Tetraveryhachium) longispinosum (Jardiné et al.) Stancliffe and Sarjeant 1994 (figure 7.14)
Veryhachium (Veryhachium) trispinosum (Deunff) Stancliffe and Sarjeant 1994 (figure 7.15)

Figure 6. 1, Ammonidium garrasinoi Ottone BAFC-Pl 2080 (1) R39. 2, Arkonites bilixus Legault BAFC-Pl 1255 (2) B20. 3, Multiplicisphaeridium sp. BAFC-Pl 2080 (1) F40. 4, Crucidia camirense (Lobo Boneta) Ottone BAFC-Pl 1255 (1) D51. 5, Dactylofusa fastidiona (Cramer) Fensome, Williams, Sedley Barss, Freeman, J and Hill BAFC-Pl 1255 (1) V57/3. 6, Dictyotidium munificum (Wicander and Wood) R. Amenábar, di Pasquo, Carrizo and Azcuy BAFC-Pl 2080 (2) J29/1. 7, Dorsennidium pastoris (Deunff) Sarjeant and Stancliffe BAFC-Pl 2080 (1) T37. 8, Duvernaysphaera angelae Deunff BAFC-Pl 1256 (2) L43/4. 9, Duvernaysphaera tenuincingulata Staplin BAFC-Pl 1255 (1) B54. 10, Exochoderma arca Wicander and Wood BAFC-Pl 1255 (1) M45/4. 11, Exochoderma triangulata Wicander and Wood BAFC-Pl 1255 (2) Y46/2. 12, Gorgonisphaeridium discissum Playford in Playford and Dring BAFC-Pl 1255 (1) Z40/2. 13, Gorgonisphaeridium vesculum Playford in Playford and Dring BAFC-Pl 2080 (1) T39/2. 14, Haspidopalla exornata (Deunff) Playford BAFC-Pl 2080 (1) P25/1. 15, Hemiruptia legaultii Ottone BAFC-Pl 2081 (1) L31. Scale bar / escala gráfica: 1, 3, 4, 5, 6, 14, 15= 15 µm. 2, 8, 9, 12, 13= 10 µm. 7, 10, 11= 20 µm.

Figure 7. 1, Leiofusa sp. BAFC-Pl 1255 (1) O41. 2, Leiosphaeridia sp. BAFC-Pl 2080 (1) X37/2. 3, Maranhites mosesii (Sommer) Brito emend. Burjack and Oliveira BAFC-Pl 1256 (1) U22/4. 4, Micrhystridium? spinoglobosum (Staplin) Sarjeant and Stancliffe BAFC-Pl 2080 (1) L45. 5, Multiplicisphaeridium ramispinosum (Staplin) Sarjeant and Vavrdová BAFC-Pl 1255 (1) M41/4. 6, Navifusa bacilla (Deunff) Playford BAFC-Pl 2080 (1) P35. 7, Papulogabata sp. A. BAFC-Pl 2081 (3) Y33/1. 8, Polygonium barredae Ottone BAFC-Pl 1255 (2) R46/2. 9, Pterospermella capitana Wicander BAFC-Pl 2080 (1) P37/2. 10, Pterospermella pernambucencis (Brito) Eisenack, Cramer and Diez Rodriguez BAFC-Pl 1255 (1) U43/3. 11, Quadrisporites granulatus (Cramer) Ströther BAFC-Pl 1255 (1) Y52. 12, Stellinium micropolygonale (Stockmans and Willière) Playford BAFC-Pl 1255 (1) S47/4. 13, Tunisphaeridium caudatum Deunff and Evitt BAFC-Pl 1255 (2) F20/3. 14, Veryhachium (Tetraveryhachium) longispinosum (Jardiné et al.) Stancliffe and Sarjeant BAFC-Pl 1255 (2) U49. 15, Veryhachium (Veryhachiurn) trispinosum (Deunff) Stancliffe and Sarjeant BAFC-Pl 2080 (1) T46/1. Scale bar / escala gráfica: 1, 6= 20 µm. 2, 3, 4, 5, 7, 9, 10, 11, 13, 14, 15= 15 µm. 8, 12= 10 µm.
Chitinozoans
Alpenachitina matogrossensis Burjack and Paris 1989 (figure 8.1)
Alpenachitina sp. cf. A. eisenacki Dunn and Miller 1964 (figure 8.2)
Ancyrochitina simplex Grahn, Bergamaschi and Pereira 2002 (figure 8.3)
Ancyrochitina spp.
Angochitina katzeri Grahn and Melo 2002 (figure 8.4a,b)
Angochitina mourai Lange 1952 (figure 8.5a,b)
Angochitina spp.
Fungochitina pilosa Collinson and Scott 1958 (figure 8.7)
Fungochitina sp. (figure 8.6)
Ramochitina sp. cf. R. ramosi Sommer and Van Boekel 1964 (figure 8.8)

Figure 8. 1, Alpenachitina matogrossensis Burjack and Paris BAFC-Pl 1257 (1) K32, BAFC-Pl 1257 (1) L52/1. 2, Alpenachitina sp. cf. A. eisenacki Dunn and Miller BAFC-Pl 1258 (2) M36/3. 3, Ancyrochitina simplex Grahn, Bergamaschi and Pereira BAFC-Pl 1257 (2) M54/2. 4a, Angochitina katzeri Grahn and Melo BAFC-Pl 2081 (1) C38/3; b, Detail of ornamentation / detalle de la ornamentación. 5a, Angochitina mourai Lange BAFC-Pl 2081 (1) V38/1; b, Detail of ornamentation / detalle de la ornamentación. 6, Fungochitina sp. BAFC-Pl 2081 (1) O46/2. 7, Fungochitina pilosa Collinson and Scott BAFC-Pl 2081 (1) E37/2. 8, Ramochitina sp. cf. R. ramosi Sommer and Van Boekel BAFC-Pl 1257 (1) Q39/2. Scale bar / escala gráfica: 1, 2, 3, 4, 5, 6= 28 µm. 7= 26 µm. 8= 15 µm.
Systematic descriptions
Trilete spores
Anteturma PROXIMEGERMINANTES Potonié
Turma TRILETES Reinsch emend. Dettmann
Suprasubturma ACAVATITRILETES Dettmann
Subturma AZONOTRILETES Lüber emend. Dettmann
Infraturma LAEVIGATI (Bennie and Kidston) PotoniéGenus Leiotriletes (Naumova) Potonié and Kremp 1954
Type species. L. sphaerotriangulus (Loose) Potonié and Kremp 1954.
Leiotriletes balapucensis di Pasquo 2007a Figure 5.8
Remarks. Of all the studied specimens in these samples, the better preserved one is the one illustrated herein. The lack of margin and exinal foldings, recurrent features in this species, offer a wide range of "apparent" ambs from subtriangular (see di Pasquo 2007a, p. 135, fig. 6. F-I) to round (see Amenábar et al. 2006, p. 354, fig. 5 E-J).
Infraturma MURORNATI Potonié and Kremp
Genus Acinosporites Richardson 1965
Type species. A. acanthomammillatus Richardson 1965.
Acinosporites lindlarensis Riegel 1968 Figure 4.3
Dimensions (6 specimens). 46- 64 µm.
Remarks. The mode for the size of the specimens from Tonono is slightly smaller than the original population.
Acinosporites sp. cf. A. eumammillatus Loboziak, Streel and Burjack 1988 Figure 4.4
Description. Trilete miospore with a subtriangular amb. Thickness of the exine 1.5 µm. Laesurae arms elevated, extending to the equator. Sculptural elements biform, consisting on a wide base surmonted by a small element, barely discernible.
Dimensions (1 specimen). 30 µm.
Remarks. The only specimen found, lacks a consistent thickening of the equatorial region; hence the cf. assignment.
Acinosporites sp. cf. A. macrospinosus Richardson 1965 Figure 4.5
Remarks. Acinosporites sp. cf. A. macrospinosus is slightly smaller and more rounded. The poor preservation prevents a more reliable identification.
Infraturma APICULATI Bennie and Kidston emend. Potonié
Subinfraturma VERRUCATI Dybová and JachowiczGenus Verrucosisporites Ibrahim emend. Smith 1971
Type species. V. verrucosus Ibrahim 1933.
Verrucosisporites sp. cf. V. loboziakii Marshall and Fletcher 2002 Figure 5.18
Description. Spore radial. Amb circular slighlty oval. Suturae not visible. Exine sculptured proximally and distally with verrucae of different sizes, which can be anastomosed.
Dimensions. 70 µm (one specimen).
Remarks. The poor preservation and the lack of additional specimens prevent a more precise assignment.
Paleomicroplankton
Group ACRITARCHA Evitt
Genus Multiplicisphaeridium Staplin emend. Sarjeant and Vavrdová 1997.
Type species. M. ramispinosum Staplin emend. Sarjeant and Vavrdová 1997.
Multiplicisphaeridium sp. Figure 6.3
Description. Vesicle spherical, psilate. Numerous processes, hollow, laevigate as well, homomorphic to slighltly heteromorphic, with circular bases proximally contacting the vesicle and acuminated distal extremities, free communicated with the vesicle.
Dimensions. Vesicle diameter: 40 µm; processes 9.5 µm long (one specimen).
Remarks. Multiplicisphaeridium sp. resembles Baltisphaerosum sp. in Ottone 1996 even though the former does not show any basal plug in the processes.
Genus Dorsennidium Wicander emend. Sarjeant and Stancliffe 1994
Type species. D. patulum Wicander 1974.
Dorsennidium pastoris (Deunff) Sarjeant and Stancliffe 1994 Figure 6.7
Remarks. This specimen resembles Villosacapsula semipunctata (Pöthe de Baldis) Sarjeant and Vavrdová 1997 but it has more processes and these are not ornamented. Genus Leiofusa Eisenack emend. Eisenack emend. Combaz et al. emend. Cramer 1970
Type species. Leiofusa fusiformis Eisenack ex Eisenack 1938.
Leiofusa sp. Figure 7.1
Remarks. Leiofusa sp. resembles L. pyrena Wicander and Wood 1981; however, the missing part of this single specimen prevents specific identification. Leiofusa sp. in Ottone (1996, Plate 7, figure 13) has a thiner wall and longer processes.
Genus Papulogabata Playford in Playford and Dring 1981
Type species. P. annulata Playford in Playford and Dring 1981.
Papulogabata sp. A Figure 7.7
Diagnosis. Circular to subcircular vesicle; surface psilate. Wall presenting three radial thickenings, the one on the margin is 4 µm in width, the following one is 8 µm wide and the one closer to the center, surrounding the cyclopyle (11 µm), is 5 µm.
Comparisons and remarks. Papulogabata annulata Playford in Playford and Dring 1981 is smaller and has only one thickened ring. P. persica Ghavidel-Syooki (2003) is much bigger and the vesicle wall can be scabrated. Because only one specimen of Papulogabata sp. A was found it is placed in open nomenclature.
Chitinozoans
Group CHITINOZOA Eisenack
Order PROSOMATIFERA Eisenack
Family CONOCHITINIDAE Eisenack emend. Paris
Subfamily ANCYROCHITININAE ParisGenus Alpenachitina Dunn and Miller 1964
Type species. A. eisenacki Dunn and Miller 1964.
Alpenachitina sp. cf. A. eisenacki Dunn and Miller 1964 Figure 8.2
Remarks. Despite the poor preservation, the specimen bears processes by the basal margin and by the shoulder, even though the missing collarette and the lack of further specimens prevent a more accurate assignment.
Family LAGENOCHITINIDAE Eisenack emend. Paris
Subfamily ANGOCHITININAE ParisGenus Ramochitina Sommer and van Boekel emend. Paris et al. 1999
Type species. R. ramosi Sommer and van Boekel 1964.
Ramochitina sp. cf. R. ramosi Sommer and Van Boekel 1964 Figure 8.8
Remarks. Because only a fragment of a specimen was found and despite its outstanding ornamentation, a more accurate identification is impossible.
Genus Fungochitina Taugourdeau 1966
Type species. Conochitina fungiformis Eisenack 1931.
Fungochitina sp. Figure 8.6
Remarks. Fungochitina sp. resembles Fungochitina sp. cf. F. pilosa in Grahn and Melo 2002, but the shorter, robust spines, characteristic of this species are difficult to distinguish without a SEM image.
Composition of the assemblages and palaeoenvironmental considerations
The complete microflora recovered along the investigated interval of the Tonono x-1 Borehole comprises 73 relatively well-preserved species represented by diverse palynological groups such as trilete spores (35 species), microplankton including several prasinophycean and acritarch taxa together with chlorophycean algae such as Quadrisporites (30 species) and chitinozoans (8 species) (figures 3- 8). Thermal maturity (TAI) varies between 2 and 3 according to the scale of Utting et al. (in Utting and Wielens, 1992). Phytoclasts, such as tracheids and cuticular fragments, are frequent.
In order to characterize the palynofacies, a matrix was constructed with the relative percentages of the palynomorphs and the PMI, as explained previously (chart 1).
Chart 1. Relative percentage of the phytoclasts and palynomorph groups and values of Palynologycal Marine Index (PMI) of the assemblages registered in the Tonono x-1 well / porcentajes relativos de los fitoclastos y grupos de palinomorfos y valores del Indice Marino Palinológico (IMP) de las asociaciones registradas en el pozo Tonono x-1.

The palynofacies analysis and quantitative data displayed reflect paleoenvironmental changes during the deposition of the succession.
The palynoassemblage of BAFC-Pl 1258 and 1257 is very poorly preserved. There is a high proportion of structured and unstructured phytodebris and a very low diversity of palynomorphs. They have similar PMI values (chart 1) which fall within the lower values range. The high input of USP together with the low values of PMI could reflect a nearshore palaeoenvironment.
The lowest value of PMI is recorded in the palynoassemblage of BAFC-Pl 1256, in coincidence with the highest proportion of spores, structured phytodebris and black phytodebris (chart 1) supporting a very marginal deposition environment.
Both palynofloras of BAFC-Pl 1255 and 2080 yielded the highest PMI values and phytoplankton proportion although AOM is absent in the former and very abundant in the latter. This suggests a closer affinity with the BAFC-Pl 2081 where AOM proportion is high as well as the content of phytoplankton and chitinozoans (chart 1), features that would reinforce the idea of a marine origin for the amorphous organic matter (Batten, 1996). AOM is usually the dominant organic component of sediments accumulated under anoxic conditions, especially in areas removed from significant influence of terrestrial input (e.g., Tyson, 1995). Therefore, a more distal environment is proposed for the levels BAFC-Pl 2080 and 2081.Even though level BAFC-Pl 1255 has a high PMI, meaning a high diversity of microplankton, this value accompanied by a similar proportion of spores and phytoplankton (see chart 1) suggests a more marginal setting -an intermediate deposition environment in between the level BAFC-Pl 1256, representing the nearest shore one- and the top levels (BAFC-Pl 2080, 2081).
Age assessment and correlation
Three assemblages are defined based on the stratigraphic distribution of taxa along the studied interval, presence/absence of key taxa (see figures 3, 9) as well as abundances of different groups of taxa (chart 1) and palynofacies analysis. Unfortunately, the only lithological information available for this study was the one provided by the interpretation of the electric log.
The assemblages are characterized by many species in common with palynofloras from South America and selected ones from other regions (figure 9).

Figure 9. Stratigraphic ranges of selected taxa occuring in the Tonono x-1 Borehole based on the selected literature / rangos estratigráficos de taxa seleccionados del Pozo Tonono x-1: Marhoumi and Rausscher (1984), McGregor and Playford (1992), Turnau (1996), Quadros (1999), Vavrdová and Isaacson (1999), Marshall and Fletcher (2002), Grahn (2005 and references therein), Hashemi and Playford (2005), Grahn et al. (2006), di Pasquo (2007a, and references therein), Al-Ghazi (2007), Amenábar (2007), Amenábar et al. (2006, 2007, and references therein), di Pasquo and Noetinger (2008), di Pasquo et al. (2009, and references therein), González (2009).
Palynoassemblage To1 (3945-3640,5 m interval)
Although the general preservation of the organic matter is very poor, some stratigraphically important species are recognized. The inception and disapearance of some species with a restricted stratigraphic distribution support a late Eifelian to earliest Givetian age for this interval (see figure 9). The co-occurrence of Acinosporites acanthomammillatus and Acinosporites sp. cf. A. macrospinosus allow its correlation with the lower part of the late Eifelian AD (Acinosporites acanthomammillatus-Densosporites devonicus) Zone of the Ardenne-Rhenish region (Streel et al., 1987), and the Calyptosporites velatus-Rhabdosporites langii and Densosporites devonicus-Grandispora naumovii Assemblage Zones of the Old Red Sandstone Continent (Richardson and McGregor, 1986) and the Grandispora pseudoreticulata Zone (Limachi et al., 1996). Occurrence of Grandispora permulta supports the correlation with the Grandispora permulta Interval Zone (Per), of late early Eifelian through the Eifelian/ Givetian transition, for the Amazon Basin (Melo and Loboziak, 2003). Several common species (e.g., Acinosporites acanthomammillatus, A. lindlarensis, A. macrospinosus, Dibolisporites eifeliensis, D. quebecensis, Grandispora douglastownense) support its correlation with assemblage 1 of the Los Monos Formation at Balapuca (di Pasquo, 2007a,2007b) and the assemblage 2 of the Chigua Formation at Del Chaco Creek (Amenábar, 2007). Moreover, Alpenachitina matogrossensis and Ancyrochitina simplex are recorded in the Alpenachitina eisenacki-Spinachitina biconstricta Concurrent Range Zone of late Eifelian?-early Givetian age in the Paraná basin (Grahn et al., 2002), which is correlated to the Alpenachitina eisenacki Biozone for Western Gondwana (Grahn, 2005). Other taxa disappear around the early Givetian on a world-wide basis (e.g., Dibolisporites eifeliensis, Verrucosisporites sp. cf. V. loboziakii, Alpenachitina sp. cf. A. eisenacki, Ramochitina sp. cf. R. ramosi).
Palynoassemblage To2 (3365,5-3289,5 m Interval)
This assemblage is better preserved than the one below. The appearance of Geminospora lemurata, in agreement with the base of the ensensis obliquimarginatus Conodont Zone sensu Weddige (1984; see Loboziak and Streel, 1995) -together with Biharisporites parviornatus recognized as a ?late Eifelian - mid-late Givetian taxon in South America (see di Pasquo, 2007a)- suggests a Givetian age. The appearances within this interval of Leiotriletes balapucensis, Aneurospora greggsii, Acinosporites ledundae, Raistrickia aratra, Arkonites bilixus, Ammonidium garrasinoi, Duvernaysphaera angelae, Exochoderma triangulata, Maranhites mosesii, Verrucosisporites tumulentus among others reinforce this age (see figures 3 and 9).
This assemblage can be correlated with the midlate Givetian assemblage 2 of Los Monos Formation at Balapuca (di Pasquo, 2007a, 2007b) from Bolivia, the assemblage 3 of Chigua Formation at La Cortadera Creek (Amenábar et al., 2006, 2007), partially with the palynoflora of the Los Monos Formation at Galarza Creek (Ottone, 1996) and probably with the microflora of the Punta Negra Formation (Rubinstein, 1999, 2000) from Argentina. In contrast, comparison with the Givetian Verrucosisporites premnus/V. scurrus Zone of Limachi et al. (1996) of Bolivia and with the Geminospora lemurata- Chelinospora ex gr. ligurata Interval Zone (LLi) for the Amazon Basin (Melo and Loboziak, 2003) is difficult because there are few species in common (e.g., Verrucosisporites scurrus, Arkonites bilixus, Geminospora lemurata).
Assemblage III of Hashemi and Playford (2005), attributed to the late Givetian-early Frasnian, and the Frasnian microfloras documented by Balme (1988) and Playford and Dring (1981) for the Gneudna Formation in the Carnarvon Basin (Australia) share with the studied Palynoassemblage To2 only the cosmopolitan age-diagnostic taxa such as Geminospora lemurata, Verrucosisporites tumulentus, V. scurrus and Gorgonisphaeridium discissum.
Palynoassemblage To3 (3133- 3077 m Interval)
Preservation of the palynomorphs is fairly good. Only a few key species are recognized at the top of the interval. These are Acinosporites sp. cf. A. eumammillatus together with some key chitinozoans such as Angochitina katzeri, Fungochitina pilosa and Angochitina mourai. The first occurence of A. mourai indicates the base of the Frasnian in all intracratonic basins of Brazil (Gaugris and Grahn, 2006) and its abundance associated to acritarchs and chitinozoans such as Hoegisphaera glabra and Ancyrochitina sp. render an upper Frasnian age in the Bolivian Devonian (Racheboeuf et al., 1993). The occurrence of Cymbosporites catillus -that first appearsing in the highest levels of the LLi Zone (see Melo and Loboziak, 2003)- is recorded here within this interval. This assemblage shares several species with the wide ranging association documented by Ottone (1996) in the Los Monos Formation attributed to the late Givetian-early Frasnian. The association is correlated to the Concurrent Range Zone of Hoegisphaera glabra and Ramochitina derbyi -which defines the Frasnian beds of Brazil (Grahn et al., 2002)- and the Frasnian Hoegisphaera glabra Biozone (Grahn, 2005) on the basis of the presence of the chitinozoans (see figure 3). Hence, a Frasnian age is supported for this assemblage by the significant increase of marine elements and AOM between To2 and To3, which would probably have occurred as a response to a maximum flooding event, possibly related to major global eustatic changes during this time (see Noetinger and di Pasquo, 2008b).
Concluding remarks
This study presents new palynological data on spores and microplankton for the Tonono x-1 borehole. The stratigraphic distribution of 73 species of trilete spores (35 species), microplankton including several prasinophycean and acritarch taxa together with chlorophycean algae such Quadrisporites (30 species) and chitinozoans (8 species) along the studied interval (between 3945-3077 m) of the Tonono x-1 allow the definition of three palynoassemblages. Diagnostic markers present in these assemblages support a late Eifelian to early Frasnian age to the whole interval. Palynoassemblage To1 ranges from the late Eifelian to earliest Givetian. A Givetian age is proposed for association To2 whilst an early Frasnian is suggested on the basis of few species for the assemblage To3.
The whole interval corresponds to the Tonono Formation. The upper part could be attributed to the Jollín Member and the basal part to the Michicola Formation according to lithology and age of the assemblages (see figure 2).
These assemblages contain many cosmpolitan index species (e.g., Geminospora lemurata, Dibolisporites eifeliensis, Verrucosisporites scurrus, Grandispora douglastownense and Acinosporites acanthomammillatus) that support the correlation mainly with Brazilian and Euro-American zonations. Other age-diagnostic taxa are exclusively recorded from South America (e.g., Grandispora pseudoreticulata, Grandispora brevizonata, Leiotriletes balapucensis, Acinosporites ledundae), belonging to the Mid-Devonian to Frasnian Afro- South American Subrealm (di Pasquo et al., 2009). Assemblages To1 and To2 are interpreted as reflecting nearshore, shallow marine depositional conditions, characterized by a very high terrestrial input, and variable marine influence. This is in agreement with a trend towards a sea-level drop proposed by Albariño et al. (2002), and recorded in this region from the early Eifelian and apparently maintained through the Givetian. Assemblage To3 represents a transgressive period with high diversity and abundance of microplankton along with high AOM content. It seems that this change was gradual from continental conditions during the deposition of To2 (see chart 1) to shelf marine environments in To3, in agreement with the record of a new transgressive cycle in the latest Givetian-early Frasnian according to Albariño et al. (2002) and Noetinger and di Pasquo (2008b).
Acknowledgements
Thanks are given to Mercedes di Pasquo for her critical comments, invaluable support and constant motivation; to C. Rubinstein, J. Marshall and an anonymous reviewer for their careful and constructive reviews and to Repsol-YPF, for providing the core samples of the Tonono x-1 well in 1998 with permission to publish the results G. Holfeltz is thanked for processing the samples. This study is part of the Ph D. thesis of the author and it was supported with funds from the Agencia Nacional de Promoción Científica y Tecnológica (PICTR 00313/03), the Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 5518 CONICET) and the University of Buenos Aires (UBACYT X 428).
References
1. Aceñolaza, F.G., Aceñolaza, G. and García, G. 2000. El Silúrico- Devónico del noroeste Argentino. In: R. Caminos (ed.), Geología Argentina, Instituto de Geología y Recursos Minerales, Anales 29: 205- 214. [ Links ]
2. Albariño, L., Dalenz-Farjat, A., Alvarez, L., Hernandez, R. and Pérez Leyton, M. 2002. Las secuencias sedimentarias del Devónico en el Subandino sur y el Chaco, Bolivia y Argentina. 5º Congreso de Exploración y Desarrollo de Hidrocarburos (Mar del Plata), CD Trabajos Técnicos. [ Links ]
3. Al-Ghazi, A. 2007. New evidence for the Early Devonian age of the Jauf Formation in northern Saudi Arabia. Revue de Micropaléontologie 50: 59-72. [ Links ]
4. Álvarez, L.A., Dalenz-Farjat, A., Hernandez, R.M. and Albariño, L. 2003. Integración de facies y biofacies en un análisis secuencial en plataformas clásticas devónicas del sur de Bolivia y noroeste argentino. Asociación Argentina de Sedimentología, Revista 10: 103-121. [ Links ]
5. Allen, K.C. 1965. Lower and Middle Devonian spores of North and Central Vestspitsbergen. Palaeontology 8: 687-748. [ Links ]
6. Amenábar, C.R. 2007. New palynological assemblage from the Chigua Formation (Early Late-Middle Devonian), at Del Chaco creek, Volcán Range, Precordillera Argentina. In: Field Meeting of the IGCP 499-UNESCO "Devonian Land-Sea Interaction: Evolution of Ecosystems and Climate" (DEVEC), Serie Correlación Geológica, INSUGEO, Universidad Nacional de Tucumán, Special Issue, 92-96. [ Links ]
7. Amenábar, C.R., di Pasquo, M.M., Carrizo, H.and Azcuy, C.L. 2006. Palynology of the Chigua and Malimán Formations in the Sierra del Volcán, San Juan province, Argentina. Part I. Palaeomicroplankton and acavate smooth and ornamented spores. Ameghiniana 43: 339-375. [ Links ]
8. Amenábar, C.R, di Pasquo, M.M., Carrizo, H. and Azcuy, C.L. 2007. Palynology of the Chigua (Devonian) and Malimán (Carboniferous) formations in the Volcán Range, San Juan Province, Argentina. Part II. Cavate, pseudosaccate and cingulizonate spores. Ameghiniana 44: 547-564. [ Links ]
9. Antonelli, J. and Ottone, G. 2006. Palinología de coronas del Devónico y Carbonífero Superior del pozo YPF.SE.EC.X-1, El Caburé, provincia de Santiago del Estero, Argentina. Revista del Museo de Ciencias Naturales 8: 111- 117. [ Links ]
10. Araujo Carvalho de, M., Mendoça Filho, J.G. and Rêgo Menezes, T. 2006. Paleoenvironmental reconstruction based on palynofacies analysis of the Aptian- Albian succession of the Sergipe Basin, Northeastern Brazil. Marine Micropaleontology 59: 56- 81. [ Links ]
11. Azcuy, C.L. and Laffitte, G. 1981. Palinología de la Cuenca Noroeste Argentina, I Caracterísiticas de las asociaciones carbónicas: problemas e interpretación. 8º Congreso Geológico Argentino (San Luis), Actas 4: 823- 838. [ Links ]
12. Balme, B.E. 1962. Upper Devonian (Frasnian) spores from the Carnarvon basin, Western Australia. Palaeobotanist 9: 1-10. [ Links ]
13. Balme, B.E. 1988. Miospores from Late Devonian (early Frasnian) strata, Carnarvon Basin, Western Australia. Palaeontographica, Abteilung B 209: 109-166. [ Links ]
14. Barreda, V.D. 1986. Acritarcos Givetiano-Frasnianos de la Cuenca del Noroeste, Provincia de Salta. Argentina. Revista Española de Micropaleontología 18: 229245. [ Links ]
15. Batten, D.J. 1996. Palynofacies and palaeoenvironmental interpretation. In: J. Jansonius and D.C. McGregor (eds.), Palynology: principles and applications, American Association of Stratigraphica Palynologists Foundation, Utah, pp. 1011-1064. [ Links ]
16. Becker, G., Bless, M.J.M, Streel, M. and Thorez, J. 1974. Palynology and ostracode distribution in the Upper Devonian and basal Dinantian of Belgium and their dependence on sedimentary facies. Mededelingen Rijks Geologische Dienst, Nieuwe Serie 25: 10-99. [ Links ]
17. Böttcher, G., Frigerio, M., Samosiuk, N. and Vistalli, C. 1984. Modelo paleoambiental para la sedimentación de las unidades pre-carbónicas (Siluro- Devónicas) en el subsuelo de la Cuenca Paleozoica del Noroeste. 9º Congreso Geológico Argentino (S.C. de Bariloche), Actas 5: 87- 92. [ Links ]
18. Burjack, M.I and Paris, F. 1989. Chitinozoaires du genre Alpenachitina dans le Dévonien moyen du Brésil, intérêt stratigraphique et relations phylogénétiques. Géobios 22: 197-213. [ Links ]
19. Clayton, G. and Graham., J.R. 1974. Miospore assemblages from the Devonian Sherkin Formation of south-west County Cork, Republic of Ireland. Pollen et Spores 16: 565-588. [ Links ]
20. Collinson, C. and Scott, A.J. 1958. Chitinozoan Faunule of the Devonian Cedar Valley Formation. Illinois State Geological Survey, Circular 247: 1-34. [ Links ]
21. Cramer, F. H. 1970. Distribution of selected Silurian acritarchs. An account of the palynostratigraphy and paleogeography of selected Silurian acritarch taxa. Revista Española de Micropaleontologia, número extraordinario, 1-203, pl. 1-23. [ Links ]
22. Cuerda, A.A. and Baldis, B. 1971. Silúrico-Devónico de la Argentina. Ameghiniana 8: 128- 164. [ Links ]
23. Dalenz Farjat, A., Alvarez, L.A., Hernández, R.M. and Albariño, L.M. 2002. Cuenca Siluro-Devónica del Sur de Bolivia y del Noroeste Argentino: algunas interpretaciones. 5º Congreso de Exploración y Desarrollo de Hidrocarburos (Mar del Plata), CD Trabajos Técnicos. [ Links ]
24. Dávila, J. and Rodríguez, E. 1967. The Devonian System in Bolivia and the problem of its base. Proceedings of the International Symposium on the Devonian System (Calgary) 2: 921-935. [ Links ]
25. Deunff, J. 1964. Le genre Duvernaysphaera Staplin. Grana Palynologica 5: 210-215. [ Links ]
26. Deunff, J. and Evitt, W.R. 1968. Tunisphaeridium, a new acritarch genus from the Silurian and Devonian. Stanford University Publications, Geological Sciences 12: 1-13, pl. 1. [ Links ]
27. di Pasquo, M.M. 2003. Avances sobre palinología, bioestratigrafía y correlación de las asociaciones presentes en los Grupos Macharetí y Mandiyutí, Neopaleozoico de la Cuenca Tarija, provincia de Salta, Argentina. Ameghiniana 40: 3-32. [ Links ]
28. di Pasquo, M.M. 2007a. Asociaciones palinológicas en las formaciones Los Monos (Devónico) e Itacua (Carbonífero Inferior) en Balapuca (cuenca Tarija), sur de Bolivia. Parte 1. Formación Los Monos. Revista Geológica de Chile 34: 97-137. [ Links ]
29. di Pasquo, M.M. 2007b. Asociaciones palinológicas presentes en las Formaciones Los Monos (Devónico) e Itacua (Carbonífero Inferior) en el perfil de Balapuca, sur de Bolivia. Parte 2. Formación Itacua e interpretación estratigráfica y cronología de las formaciones Los Monos e Itacua. Revista Geológica de Chile 34: 163-198. [ Links ]
30. di Pasquo, M.M. and Noetinger, S. 2008. First record of Early Devonian (Lochkovian) flora from the Santa Rosa Formation - Alarache, Southern Bolivia. Geologica Acta 6: 191- 210. [ Links ]
31. di Pasquo, M., Amenábar, C.R., Noetinger, S., 2009. Middle Devonian microfloras and megafloras from western Argentina and southern Bolivia. Their importance in the palaeobiogeographical and palaeoclimatic evolution of western Gondwana. The Geological Society, London, Special Publications 314: 193-213. [ Links ]
32. Disalvo, A. 2002. Cuenca del Noroeste: marco geológico y reseña histórica de la actividad petrolera. Simposio Rocas Reservorio de las Cuencas Productivas de la Argentina. 5º Congreso de Exploración y Desarrollo de Hidrocarburos (Mar del Plata), Actas: 663- 677. [ Links ]
33. Dunn, L. and Miller, T.H. 1964. A distinctive chitinozoan faunule from the Alpena Limestone (Middle Devonian) of Michigan. Journal of Paleontology 38: 725-728. [ Links ]
34. Eisenack, A. 1931. Neue Mikrofossilien des baltischen Silurs. 1. Palaeontologische Zeitschrift 13: 74-118. [ Links ]
35. Eisenack, A. 1938. Hystrichosphaerideen und verwandte Formen im baltischen Silur. Zeitschrift für Geschiebeforschung und Flachlandsgeologie 14: 1-30, pl. [ Links ]
36. Eisenack, A., Cramer, F.H. and Díez, M. del C.R. 1973. Katalog der fossilen Dinoflagellaten, Hystrichosphären und verwandten Mikröfossilien. Band III Acritarcha 1. Teil. E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart, 1104 pp. [ Links ]
37.Fensome , R.A., Williams, G.L., Sedley Barss, M., Freeman, J.M. and Hill, J.M. 1990. Acritarchs and fossil prasinophytes: an index to genera, species and infraspecific taxa. AASP Contribution Series 25: 678 pp. [ Links ]
38. Gaugris, K. de A. and Grahn, Y. 2006. New chitinozoan species from the Devonian of the Paraná Basin, south Brazil, and their biostratigraphic significance. Ameghiniana 43: 293-310. [ Links ]
39. Ghavidel-Syooki, M. 2003. Palynostratigraphy of Devonian sediments in the Zagros Basin, southern Iran. Review of Palaeobotany and Palynology 127: 241- 268. [ Links ]
40. González, F. 2009. Reappraisal of the organic-walled microphytoplankton genus Maranhites: morphology, excystment, and speciation. Review of Palaeobotany and Palynology 154: 6-21. [ Links ]
41. Grahn, Y. 2003. Silurian and Devonian chitinozian assemblages from the Chaco-Paraná Basin, northeastern Argentina and central Uruguay. Revista Española de Micropaleontología 35: 1-8. [ Links ]
42. Grahn, Y. 2005. Devonian chitinozoan biozones of Western Gondwana. Acta Geologica Polonica 55: 211-227. [ Links ]
43. Grahn, Y. and Gutierrez, P.R. 2001. Silurian and Middle Devonian Chitinozoa from the Zapla and Santa Bárbara Ranges, Tarija Basin, northwestern Argentina. Ameghiniana 38: 35-50. [ Links ]
44. Grahn, Y. and Melo, J. H. G. 2002. Chitinozoan biostratigraphy of the Late Devonian formations in well Caima PH-2, Tapajós River area, Amazonas Basin, northern Brazil. Review of Palaeobotany and Palynology 118: 115-139. [ Links ]
45. Grahn, Y., Bergamaschi, S. and Pereira, E. 2002. Middle and Upper Devonian chitinozoan biostratigraphy of the Paraná Basin in Brazil and Paraguay. Palynology 26: 135-165. [ Links ]
46. Grahn, Y., Melo, J.H.G. and Loboziak, S. 2006. Integrated Middle and Late Devonian miospore and chitinozoan zonation of the Parnaíba Basin, Brazil: an update. Revista Brasileira de Paleontologia 9: 283- 294. [ Links ]
47. Hashemi, H. and Playford, G. 2005. Devonian spore assemblages of the Adavale Basin, Queensland (Australia): descriptive systematics and stratigraphic significance. Revista Española de Micropaleontología 37: 317-417. [ Links ]
48. Helenes, J., de-Guerra, C. and Vásquez, J. 1998. Palynology and chronostratigraphy of the Upper Cretaceous in the subsurface of the Barinas area, western Venezuela. The American Association of Petroleum Geologists, Bulletin 82: 1308-1328. [ Links ]
49. Ibrahim, A.C. 1933. [Sporonfermen des ägirhorizontes des Ruhr- Reviers. Würzburg. Ph.D. Dissertation, Berlin, 47 pp. Unpublished. [ Links ]].
50. Kimyai, A. 1983. Palaeozoic microphytoplankton from South America. Revista Española de Micropaleontología 15: 415-426. [ Links ]
51. Lange, F.W. 1952. Quitinozoários do Folhelho Barreirinha, Devoniano do Pará. Dusenia 3: 373-386. [ Links ]
52. Legault, J.A. 1973. Chitinozoa and Acritarcha of the Hamilton Formation (Middle Devonian), Southwestern Ontario. Geological Survey of Canada, Bulletin 221: 1-110. [ Links ]
53. Limachi, R., Goitia, V.H., Sarmiento, D., Arispe, O., Montecinos, R., Diaz Martínez, E., Dalenz Farjat, A., Liachenco, N., Pérez Leyton, M. and Aguilera, E. 1996. Estratigrafía, geoquímica, correlaciones, ambientes sedimentarios y bioestratigrafía del Silúrico-Devónico de Bolivia. 12º Congreso Geológico de Bolivia (Tarija), Memorias 12: 183-197. [ Links ]
54. Loboziak, S. and Streel, M. 1995. West Gondwanan aspects of the Middle and Upper Devonian miospore zonation in North Africa and Brazil. Review of Palaeobotany and Palynology 86: 147- 155. [ Links ]
55. Loboziak, S., Streel, M. and Burjack, M.I.A. 1988. Miospores du Dévonien moyen et supérieur du bassin du Paraná, Brésil: systématique et stratigraphie. Bulletin des Sciences Géologiques 41: 351- 377. [ Links ]
56. Loboziak, S., Streel, M. and Melo, J.H.G. 1999. Grandispora (al. Contagiosporites) permulta (Daemon, 1974) Loboziak, Streel et Melo, comb. nov., a senior synonym of Grandispora riegelli Loboziak et Streel, 1989. Nomenclature and stratigraphic distribution. Review of Palaeobotany and Palynology 106: 97-102. [ Links ]
57. Marhoumi, R. and Rauscher, R.1984. Un plancton Dévonien de la Meseta Orientale au Maroc. Review of Palaeobotany and Palynology 43: 237-253. [ Links ]
58. Marshall, J.E.A. and Fletcher, T.P. 2002. Middle Devonian (Eifelian) spores from a fluvial dominated lake margin in the Orcadian Basin, Scotland. Review of Palaeobotany and Palynology 118: 195- 209. [ Links ]
59. McGregor, D.C. 1973. Lower and Middle Devonian spores of eastern Gaspé, Canada. I. Systematics. Palaeontographica, Abteilung B 142: 1-77. [ Links ]
60. McGregor, D.C. and Camfield, M. 1982. Middle Devonian amiospores from the Cape de Bray, Weatherall, and Hecla Bay Formations of northeastern Melville Island, Canadian Artic. Geological Survey of Canada, Bulletin 348: 1-105. [ Links ]
61. McGregor, D.C. and Playford, G. 1992. Canadian and Australian Devonian spores: zonation and correlation. Geological Survey of Canada Bulletin 438: 1-125. [ Links ]
62. Melo, J.H.G. and Loboziak, S. 2003. Devonian-Early Carboniferous miospore biostratigraphy of the Amazon Basin, Northern Brazil. Review of Palaeobotany and Palynology 124: 131- 202. [ Links ]
63. Noetinger, S. and di Pasquo, M.M. 2008a. Middle-Upper Devonian microflora from a borehole in northwestern Argentina. 12º International Palynological Congress IPC-XII 2008; 8º International Organisation of Palaeobotany Conference IOPC-VIII (Bonn), Terra Nostra: 509 pp. [ Links ]
64. Noetinger, S. and di Pasquo, M.M. 2008b. Givetian- Frasnian microfloras from the Tarija basin, northern Argentina, and their relationship with the global eustasy and palaeoclimate. 17º Congreso Geológico Argentino (Jujuy): 368- 369. [ Links ]
65. Ottone, E.G. 1996. Devonian palynomorphs from the Los Monos formation, Tarija basin, Argentina. Palynology 20: 105- 155. [ Links ]
66. Owens, B. 1971. Miospores from the Middle and Early Upper Devonian rocks of the Western Queen Elizabeth Islands, Artic Archipielago. Geological Survey of Canada, Paper 70: 1-157. [ Links ]
67. Padula, E., Rolleri, E., Mingramm, A., Criado Roque, P., Flores, M. and Baldis, B. 1967. Devonian of Argentina. International Symposium on the Devonian System (Calgary), Proceedings 2: 165-199. [ Links ]
68. Paris, F., Grahn, Y., Nestor, V. and Lakova, I. 1999. Proposal for a revised chitinozoan classification. Journal of Paleontology 73: 549-570. [ Links ]
69. Playford, G. 1977. Lower to Middle Devonian acritarchs of the Moose River Basin, Ontario. Geological Survey of Canada, Bulletin 279: 1-87. [ Links ]
70. Playford, G. and Dring, R.S. 1981. Late Devonian acritarchs from the Carnarvon Basin, Western Australia. Palaeontological Association, Special Papers in Palaeontology 27: 1-78. [ Links ]
71. Potonié, R. and Kremp, G.O. 1954. Die Gattungen der Paläozoischen Sporae dispersae und ihre Stratigraphie. Beihefte zum Geologischen Jahrbuch 69: 111-194. [ Links ]
72. Quadros, L.P. 1999. Silurian-Devonian acritarch assemblages from Paraná Basin: an update and correlation with Northern Brazilian basins. In: M.A.C. Rodrigues, E. Pereira (eds), Ordovician-Devonian palynostratigraphy in Western Gondwana: update, problems and perspectives. Faculdade de Geologia da Universidade Estatal do Rio de Janeiro, Resumos expandidos: 105-145. [ Links ]
73. Racheboeuf, P., Le Hérissé, A., Paris, F., Babin, C., Guillocheau, F., Truyols-Massoni, M. and Suárez-Soruco, R. 1993. Le Dévonien de Bolivie: biostratigraphie et chronostratigraphie. Comptes Rendus de l´Academie de Sciences de Paris 317: 795-802. [ Links ]
74. Richardson, J.B. 1965. Middle Old Red Sandstone spore assemblages from the Orcadian Basin north-east Scotland. Palaeontology 7: 559 605. [ Links ]
75. Richardson, J.B. and Lister, T.R. 1969. Upper Silurian and Lower Devonian spore assemblages from the Welsh Borderland and South Wales. Palaeontology 12: 201-252. [ Links ]
76. Richadson, J.B. and Rasul, S.M. 1978. Palynomorphs in Lower Devonian sediments from the Apley Barn Borehole, southern England. Pollen et Spores 20: 423-462. [ Links ]
77. Richardson, J.B. and McGregor, D.C. 1986. Silurian and Devonian spore zones of the Old Red Sandstone continent and adjacent regions. Geological Survey of Canada, Bulletin 364: 1-79. [ Links ]
78. Riegel, W. 1968. Die Mitteldevon-Flora von Lindlar (Rheinland). 2. Sporae dispersae; Palaeontographica, Abteilung B 123: 76-96. [ Links ]
79. Rubinstein, C.V. 1999. Primer registro palinológico de la Formación Punta Negra (Devónico Medio-Superior), de la Precordillera de San Juan, Argentina. 10º Simposio Argentino de Paleobotánica y Palinología (Buenos Aires). Asociación Paleontológica Argentina, Publicación Especial 6: 13-18. [ Links ]
80. Rubinstein, C.V. 2000. Middle Devonian palynomorphs from the San Juan Precordillera, Argentina: biostratigraphy and paleobiogeography. In: Congreso Ibérico de Paleontología, Jornadas de la Sociedad Española de Paleontología y International Meeting of IGCP 421, Nº 1, Nº 14 y Nº 8 (Evora), Resúmenes: 274-275. [ Links ]
81. Russo, A., Ferello, R. and Chebli, G. 1979. Llanura Chaco Pampeana. 2º Simposio de Geología Regional Argentina (Córdoba). Academia Nacional de Ciencias (Córdoba) 1: 139- 183. [ Links ]
82. Sarjeant, W.A.S. and Stancliffe, R.P.W. 1994. The Micrhystridium and Veryhachium complexes (Acritarcha: Acanthomorphitae and Polygonomorphitae): a taxonomic reconsideration. Micropaleontology 40: 1-77. [ Links ]
83. Sarjeant, W.A.S. and Vavrdová, M. 1997. Taxonomic reconsideration of Multiplicisphaeridium Staplin, 1961 and other acritarch genera with branching processes. Geolines 5: 1-52. [ Links ]
84. Smith, A.H.V. 1971. Microfossiles organiques du Paleozoique. 4. Les spores. 2. Le genre Verrucosisporites par A.H.V. Smith. Commission Internationale de Microflore du Paleozoique, Centre National de la Recherche Scientifique, 35-87. [ Links ]
85. Sommer, F.W. and van Boekel, N.M. 1964. Quitinozoarios do Devoniano de Goiás. Anais da Academia Brasileira de Ciências 36: 423-431. [ Links ]
86. Stancliffe, R.P.W. and Sarjeant, W.A.S. 1994. The acritarch genus Veryhachium Deunff 1954, emend. Sarjeant and Stancliffe 1994: a taxonomic restudy and a reassessment of its constituent species. Micropaleontology 40: 223-241. [ Links ]
87. Staplin, F.L. 1961. Reef-controlled distribution of Devonian microplankton in Alberta. Palaeontology 4: 392-424, pl. 48-51. [ Links ]
88. Starck, D. 1995. Silurian- Jurassic stratigraphy and Basin evolution of northwestern Argentina. In: A.J. Tankard, S.R. Suárez and H.J. Welsink (eds.), Petroleum basins of South America. Tulsa. American Association of Petroleum Geologists, Memoir 62: 251-267. [ Links ]
89. Starck, D. 1999. Los sistemas petroleros de la Cuenca de Tarija. 4º Congreso de Exploración y Desarrollo de Hidrocarburos (Mar del Plata), Actas 1: 63-82. [ Links ]
90. Streel, M. 1967. Associations de spores du Dévonien inférieur belge et leur signification stratigraphique. Annales de la Société Géologique de Belgique 87: 1-30. [ Links ]
91. Streel, M., Higgs, K., Loboziak, S., Riegel, W. and Steemans, P. 1987. Spore stratigraphy and correlation with faunas and floras in the type marine Devonian of the Ardenne-Rhenish regions. Review of Palaeobotany and Palynology 50: 211-229. [ Links ]
92. Ströther, P.K. 1991. A classification schema for the cryptospores. Palynology 15: 219-236. [ Links ]
93. Suárez Soruco, R. 2000. Compendio de Geología de Bolivia. Revista Técnica Yacimientos Petrolíferos Fiscales Bolivianos 18: 1- 213. [ Links ]
94. Taugourdeau, P.H. 1966. Les chitinozoaires, techniques d'études, morphologie et classification. Mémoire de la Société Geólogique de France 45: 64 pp. [ Links ]
95. Turnau, E. 1996. Miospore stratigraphy of Middle Devonian deposits from Western Pomerania. Review of Palaeobotany and Palynology 93: 107-125. [ Links ]
96. Tyson, R.V. 1995. Sedimentary organic matter. Organic facies and palynofacies, Chapman & Hall, London, xviii+615 pp. [ Links ]
97. Utting, J. and Wielens, H. 1992. Organic petrology, thermal maturity, geology, and petroleum source rock potential of Lower Permian coal, Karoo Supersystem, Zambia. Energy Sources 14: 337-354. [ Links ]
98. Vavrdová, M. and Isaacson, P.E. 1999. Late Famennian phytogeography provincialism: Evidence for a limited separation of Gondwana and Laurentia. Abhandlungen der Geologischen Bundesanstalt 54: 453-463. [ Links ]
99. Vistalli, M.C., Hernández, R., Disalvo, A., Starck, D. and Sylwan, C. 2005. Cuencas paleozoicas del noroeste argentino. In: Chebli, G.A., Cortiñas, J.S., Spalletti, L.A., Legarreta, L. and Vallejo, E.L. (eds.). Frontera Exploratoria de la Argentina. 6º Congreso de Exploración y Desarrollo de Hidrocarburos (Mar del Plata): 41-61. [ Links ]
100. Volkheimer, W., Melendi, D.L. and Salas, A. 1986. Devonian chitinozoans from northwestern Argentina. Neues Jahrbuch für Geologie und Paläontologie, Abhandlung 173: 229-251. [ Links ]
101. Weddige, K. 1984. Zur Stratigraphie und Paleobiogeographie des Devons und Karbons von NE-Iran. Senckenbergiana Lethaea 65: 179-223. [ Links ]
102. Wicander, E.R. 1974. Upper Devonian-Lower Mississippian acritarchs and prasinophycean algae from Ohio, U.S.A. Palaeontographica, Abteilung B 148: 9-43, pl. 5-19. [ Links ]
103. Wicander, E.R. and Wood, G.D. 1981. Systematics and biostratigraphy of the organic-walled microphytoplankton from the Middle Devonian (Givetian) Silica Formation, Ohio, U.S.A. American Association of Stratigraphic Palynologists, Contributions Series 8: 1-137. [ Links ]
Recibido: 24 de febrero de 2009.
Adeptado: 6 de octubre de 2009.


{kind=link}