










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de la Asociación Geológica Argentina
versão impressa ISSN 0004-4822versão On-line ISSN 1851-8249
Rev. Asoc. Geol. Argent. v.61 n.2 Buenos Aires abr./jun. 2006
El Grupo Cabo Domingo, Tierra del Fuego: bioestratigrafía, paleoambientes y acontecimientos del Eoceno-Mioceno marino
Malumián, N.1 y Olivero, E.B.2
1 SEGEMAR y CONICET; Benjamín Lavaissé 1194; (1107) Buenos Aires, Argentina. E-mail: malumian@mpgeo1.gov.ar
2 Centro Austral de Investigaciones Científicas, CADIC-CONICET; B. A. Houssay 200; (9410) Ushuaia, Tierra del Fuego. E-mail: emolivero@ciudad.com.ar
RESUMEN
El Grupo Cabo Domingo, cuenca Austral, Eoceno superior a Mioceno medio, comprende una secuencia episódica de sedimentitas marinas levemente dislocadas y depositadas principalmente al norte de la falla de punta Gruesa en Tierra del Fuego. Se inicia con la profundización de la cuenca, resultado de los niveles de mar alto del Eoceno tardío amplificados por un diastrofismo generalizado (Fase Incaica). En el depocentro de la cuenca está representado por el miembro superior de la Formación Cerro Colorado (140 m, fangolitas y areniscas, con el último registro de Globigerinatheka index, y las apariciones de Praetenuitella insolita e Isthmolithus recurvus, 36-34 Ma); y fuera del depocentro, por el conglomerado Tchat-Chii (70 m, Eoceno cuspidal-Oligoceno basal), y el glauconítico A en el subsuelo en el norte de la Isla. El mínimo contenido de esmectita y la abundancia de niveles con Chiloguembelina reflejan tectonismo activo y caída de temperatura durante la depositación del miembro superior y el glauconítico A. Continúan las capas de la Estancia María Cristina (>75 m, arcillitas y areniscas, Oligoceno temprano, 34-30 Ma) y del puesto Herminita (200 m, arcillitas, Oligoceno, 30-26,5 Ma, principalmente con microfaunas residuales de foraminíferos aglutinados depositadas por debajo de la línea de compensación de la calcita), corresponden a la máxima profundización de la cuenca que habilitó el ingreso de aguas antárticas corrosivas. Seguidas por la Formación Desdémona (250 m, fangolitas tobáceas, bioturbadas, depositadas en las proximidades de la lisoclina, con asociaciones de foraminíferos de aguas abisales), que representa los niveles altos de mar del Oligoceno cuspidal-Mioceno temprano. Seguidas por las capas del Cabo Ladrillero (80 m, areniscas glauconíticas basales y fangolitas con asociaciones de foraminíferos de plataforma externa, y 75 m de fangolitas carbonosas, areniscas finas y arcillitas, Mioceno inferior, con la asociación residual de Spirosigmoilinella compressa-Martinottiella spp.) y por las capas del Cabo San Pablo (30- 40 m de areniscas y arcillitas, Mioceno temprano). Una generalizada regresión da lugar a una extendida discordancia en ca. 21 Ma. Finalmente, las Formaciones Carmen Silva (>50 m, fangolitas y conglomerados deltaicos) y Castillo (50 m, conglomerados fluviales) y capas del Cabo Viamonte (>40 m, brechas gruesas, conglomerados, areniscas y fangolitas) todas con clásticos volcánicos basálticos, representan el nivel de mar alto y óptimo del Neógeno reflejado por un acontecimiento esmectítico del Mioceno medio y la existencia de un episodio basáltico en el Mioceno inferior superior.
Palabras clave: Grupo Cabo Domingo. Cuenca Austral. Tierra del Fuego. Bioestratigrafía. Paleoambientes. Acontecimientos del Eoceno-Mioceno. Foraminifera.
ABSTRACT
The Cabo Domingo Group, Tierra del Fuego: Biostratigraphy, paleoenvironments, and events of the marine Eocene-Miocene. The Cabo Domingo Group, upper Eocene to middle Miocene, includes a gently deformed marine episodic sequence, mainly exposed to the north of the Punta Gruesa fault in Tierra del Fuego. Deposition of the Group started in the late Eocene with the basin deepening, resulting from global high sea level, the effects of which were amplified by regional tectonism (Incaica Phase). The Group is represented in the basin depocenter by the upper member of the Cerro Colorado Formation (140 m, mudstones and sandstones, 36-34 Ma), which records the LAD of Globigerinatheka index (34.3 Ma) and the FAD of Praetenuitella insolita and Isthmolithus recurvus (36.6 Ma). Outside the depocenter this time interval is reflected by the Tchat-Chii Conglomerate (70 m, latest Eocene-earliest Oligocene), and by the Glauconítico A in the subsurface of the northern part of the island. The low smectite content and the abundance of Chiloguembelina ototara indicate active tectonism and falling temperatures during the deposition of the upper member and the Glauconítico A . Deposition of the Cabo Domingo Group was followed by that of the Estancia María Cristina beds (>75 m, claystones and sandstones, earliest Oligocene, 34-30 Ma) and the Puesto Herminita beds (200 m, claystones, Oligocene, 30-26.5 Ma), which mainly contain residual agglutinated foraminifera deposited below the calcite compensation depth. These beds were deposited during the maximum deepening of the basin that allowed for the entrance of corrosive, Antarctic waters. The following Desdémona Formation (250 m, tuffaceous, bioturbated mudstones deposited at or near the lisocline), consisting of the Cabo Ladrillero beds (80 m, basal glauconite sandstones and mudstones with outer neritic foraminiferal assemblages, and 75 m of carbonaceous mudstones, fine-grained sandstones and claystones with residual assemblages of the benthic foraminifera Spirosigmoilinella-Martinottiella), and the Cabo San Pablo beds (30-40 m of sandstones and claystones) represents the latest Oligocene-early Miocene high sea-level conditions. An extended unconformity and generalized regression is documented at c. 21 Ma (Early Miocene) . Finally, the Carmen Silva (>50 m, deltaic mudstones and conglomerates) and Castillo (50 m, conglomerates) Formations, and the Cabo Viamonte beds (>40 m, coarse breccias, conglomerates, sandstones and mudstones), all with basalt clasts, represent the Neogene high sea-level and climatic optimum, the latter reflected by the middle Miocene smectite-rich clay composition.
Key words: Cabo Domingo Group. Austral Basin. Tierra del Fuego. Biostratigraphy. Paleoenvironments. Eocene-Miocene events. Foraminifera.
INTRODUCCIÓN
Las sedimentitas del Cenozoico de la isla Grande de Tierra del Fuego han sido divididas en tres grupos: Río Claro, Paleoceno superior-Eoceno inferior; La Despedida, Eoceno medio superior-superior; y Cabo Domingo, Eoceno cuspidal-Mioceno medio. Esta división básicamente refleja características morfoestructurales de capas fuertemente plegadas para los dos primeros, que ya han sido descritos (Olivero y Malumián 1999, Jannou y Olivero 2001, Olivero et al. 2002, 2003) y, en contraste, capas levemente afectadas a subhorizontales para el Grupo Cabo Domingo (Fig. 1), cuya descripción es motivo del presente trabajo.
Figura 1: Mapa de situación relativa y bosquejo geológico del área, perfiles y localidades estudiadas. 
El Grupo Cabo Domingo apoya sobre una discordancia asignada a la fase Incaica, que separa ciclos sedimentarios mayores en toda la Patagonia; el grupo encuentra su correlato en la megasecuencia 5 de cuenca de Malvinas (Galeazzi 1998) y en la denominada Formación Magallanes Superior incluyendo hasta la Formación Santa Cruz, dentro de la cuenca Austral, en nomenclatura interna de YPF (cf. Calegari y Baldi 1993). En este trabajo se describen los afloramientos más destacados del Grupo y se proponen nuevas unidades estratigráficas formales e informales, del Oligoceno y del Mioceno, particularmente en las cercanías del límite de la faja plegada y corrida de los Andes Fueguinos, situado a la latitud de punta Gruesa (Ghiglione 2002).
En el límite y al norte de la faja plegada y corrida (Figs. 1 y 2), por encima del miembro CCd de la Formación Cerro Colorado se definen o reconocen: a) el Conglomerado Tchat-Chii, Eoceno cuspidal-Oligoceno basal; b) las capas de la Estancia María Cristina, Oligoceno basal; c) las capas Puesto Herminita, Oligoceno; d) la Formación Desdémona, Oligoceno cuspidal- Mioceno inferior inferior; e) las capas Cabo Ladrillero, Mioceno inferior, y f) las capas Cabo San Pablo, Mioceno inferior alto?
Figura 2: Corte geológico esquemático con la correlación, arquitectura y paleoambientes generalizados del Eoceno-Mioceno en dirección paralela al eje de la cuenca. Se indican datos de primera y última aparición de algunos marcadores bioestratigráficos utilizados en la correlación. La situación de localidades se indica en la Figura 1. 
Al SSE de la punta Gruesa, la faja es un mosaico de sedimentitas del Paleoceno- Eoceno, fuertemente plegadas y falladas. En el limbo sur del anticlinal Punta Torcida-Cerro Colorado, por encima del miembro CCd (Eoceno superior, Olivero y Malumián 1999, Fig. 1), las sedimentitas oligocenas referidas preliminarmente como estratos innominados (Malumián y Olivero 2005) se correlacionan parcialmente con el Conglomerado Tchat-Chii, las capas de la Estancia María Cristina y las capas del Puesto Herminita. Sobre los estratos innominados yace en suave discordancia angular el conjunto conformado por los estratos de la María Luisa (Mioceno superior-Plioceno) y la Formación Irigoyen (Mioceno superior?- Plioceno) (Malumián y Olivero 2005).
Objetivos del trabajo
Los objetivos principales de este trabajo son: a) caracterizar la espesa y relativamente completa columna estratigráfica del Eoceno superior-Mioceno de la isla mediante sus mejores exposiciones, que mayormente se encuentran sobre la costa atlántica, y definir las unidades morfoestructurales expresables en un mapeo; b) establecer un esquema bioestratigráfico referido a los acontecimientos de un área de alta latitud en el hemisferio sur; y c) discutir brevemente la evolución geológica, con especial referencia a las edades de las discordancias y del fallamiento en el frente orogénico emergente que incluye el límite de la faja plegada.
Métodos de trabajo
Los relevamientos de mapas, confección de perfiles y muestreos se realizaron en 10 campañas entre los años 1997 y 2002. Sobre un total aproximado de 1.400 m de espesor acumulado, se colectaron 120 muestras para estudios micropaleontológicos, que en promedio resulta aproximadamente en una muestra cada 12 m de espesor. En el interior de la isla, el Grupo Cabo Domingo fue reconocido en sus escasos afloramientos y numerosos asomos, y muestreado mediante barreno de mano en áreas con delgada cobertura de suelo. Los foraminíferos planctónicos y los bentónicos se tomaron en cuenta en función del mapeo, y han sido determinados en forma expeditiva en particular las formas aglutinadas. Se reconocieron las formas de nanoplancton calcáreo más distintivas, y se investigó la existencia de silicoflagelados con resultados negativos. Se analizaron además las arcillas de una decena de muestras en el laboratorio del INTEMIN. Considerando que el color es de importancia para el reconocimiento de los diferentes cuerpos de limoarcilitas, en el interior de la Isla, se definen los colores según el Rock color chart del U.S. Geological Survey. Las edades están referidas a Berggren et al (1995).
MARCO GEOLÓGICO
Entre el Cretácico tardío e inicios del Neógeno, la cuenca Austral evolucionó como una cuenca de antepaís adosada al flanco norte del orógeno fueguino (Yrigoyen 1962, Biddle et al. 1986, Robbiano et al. 1996, Galeazzi 1998). La tectónica compresiva y la migración del frente orogénico, originaron los depocentros sedimentarios desplazándolos sucesivamente hacia el norte, junto con el avance de la deformación (Olivero y Malumián 1999). En la costa atlántica fueguina, se reconocen al menos cuatro depocentros que preservan espesas sucesiones sedimentarias marinas del Cretácico tardío-Daniano; Paleoceno tardío-Eoceno temprano; Eoceno Medio tardío-Oligoceno y Oligoceno-Mioceno, respectivamente (Olivero et al. 2002, Olivero y Malumián 2002). La deformación compresiva avanzó hacia el norte hasta alcanzar la punta Gruesa, donde se expone el frente orogénico emergente fosilizado, que constituye el límite norte de la faja plegada y corrida de los Andes Fueguinos (Ghiglione 2002, Fig. 1).
En general, los depósitos sedimentarios del Paleógeno tienen un complejo arreglo estratigráfico, con estratos de crecimiento y discordancias angulares progresivas (Olivero y Malumián 1999, Ghiglione et al. 2002). Durante parte del Mioceno, la sedimentación próxima al frente orogénico emergente, también tuvo un fuerte control tectónico, pero en este caso relacionado con la zona de falla transcurrente Fagnano, con efectos transtensivos y transpresivos, que constituye el límite entre las placas de Scotia y América del Sur (Klepeis y Austin 1997, Ghiglione 2002, Malumián y Olivero 2005).
GRUPO CABO DOMINGO
(Malumián 1999)
El grupo está expuesto mayormente en el límite y al norte de la faja plegada y corrida fueguina y se compone de un espesor mínimo de 1.000 m de sedimentitas del Eoceno cuspidal al Mioceno medio, constituidas dominantemente por limoarcillitas, con areniscas y conglomerados subordinados. Las sedimentitas se pueden diferenciar en paquetes estratigráficos con expresión morfoestructural al menos en tres ciclos o conjuntos, que caracterizan sus partes inferior, media y superior (Figs. 1 y 2), referidos al Eoceno cuspidal-Oligoceno, Oligoceno cuspidal- Mioceno temprano y Mioceno medio, respectivamente. La parte inferior, que consiste de capas con un grado de plegamiento variable, está expuesta en el límite de la faja plegada y corrida y sus aledaños (Fig. 1). Incluye al Conglomerado Tchat-Chii (Codignotto y Malumián 1981, Malumián 1988); a las capas de la Estancia María Cristina y a las del Puesto Herminita, que son depósitos marinos profundos en su mayor parte de ambientes por debajo de la línea de compensación de la calcita (LCC). La parte media, capas ligeramente plegadas a subhorizontales, está expuesta al norte de la anterior. Incluye a la Formación Desdémona y las capas de los Cabos Ladrillero y San Pablo, que principalmente corresponden a ambientes próximos a la lisoclina y por debajo de la LCC, respectivamente. La gran abundancia de diques clásticos arenosos (Ghiglione 2002) constituye un rasgo característico de la parte media. La parte superior, capas subhorizontales expuestas mayormente en el sector NO del área estudiada (Fig. 1), incluye depósitos marinos marginales y deltaicos de la Formación Carmen Silva y fluviales de la Formación Castillo (Codignotto y Malumián 1981). Se interpreta que estos depósitos del sector NO gradan hacia el SE a depósitos marinos profundos diferenciados como capas del Cabo Viamonte (Figs. 1, 2).La base del grupo se apoya en discordancia angular sobre sedimentitas eocenas (Figs. 1 y 2). La discordancia basal tiene una marcada expresión física en el sector NO dada por el fuerte contraste litológico entre el Conglomerado Tchat-Chii, que porta clastos de gran tamaño de las rocas infrayacentes, y las areniscas del Grupo La Despedida (Malumián 1988). Hacia el SE la expresión física de la discordancia se atenúa y se manifiesta por niveles de conglomerados finos y cambios de facies sedimentarias (Ponce et al. 2004, Malumián y Olivero 2005), concomitantemente decrece el hiato que separa el Grupo Cabo Domingo del de La Despedida y el límite inferior es menos marcado hacia el depocentro. En el subsuelo la base del grupo está expresada por el glauconítico A.
1) CICLO EOCENO CUSPIDAL OLIGOCENO
(ZONAS P15-P18, 36-26,5 MA)
Conglomerado Tchat-Chii
Eoceno cuspidal-Oligoceno basal
Miembro CCd de la Formación Cerro Colorado
P15 superior-P16 inferior, 36-34 Ma)
Antecedentes: El Conglomerado Tchat-Chii fue formalizado por Malumián (1988).
Distribución y litología: La base del Grupo Cabo Domingo tiene su mayor expresión en el Conglomerado Tchat-Chii. El área tipo del Conglomerado comprende al cerro epónimo (Fig. 1, localidad 8) y a los mejores afloramientos que están sobre la margen derecha del río Menéndez, aguas arriba del puente sobre la ruta provincial 8 (o b). Afloramientos aislados se presentan también sobre la cresta que constituye el limbo S del anticlinal La Despedida hasta poco al SE de la Estancia La Despedida. Fuera del área tipo, se le equiparan los niveles de areniscas conglomerádicas de reducido espesor, que yacen en contacto neto sobre areniscas finas del Miembro CCd de la Formación Cerro Colorado (Figs. 1 y 2), en los perfiles de la punta Gruesa (Fig. 3, perfil 409; Ponce et al. 2004) y del cerro Colorado (Fig. 5, perfil 428; Malumián y Olivero 2005).
Figura 3: Perfil geológico compuesto de parte del Grupo Cabo Domingo, miembro CCd de la Formación Cerro Colorado, capas de la Estancia María Cristina y del Puesto Herminita, expuesto en las cercanías de la punta Gruesa. Referencias indicadas en la Figura 4. 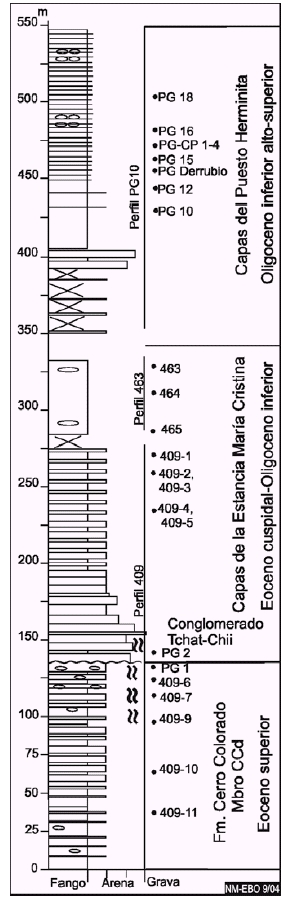
Figura 4: Perfil geológico compuesto de parte del Grupo Cabo Domingo, Formación Desdémona, capas del Cabo Ladrillero y del Cabo San Pablo, expuesto entre la punta Gruesa y el cabo San Pablo. 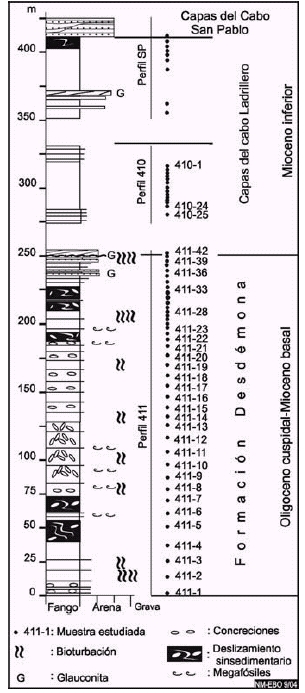
Figura 5: Perfil geológico compuesto de parte del Grupo Cabo Domingo, miembro CCd de la Formación Cerro Colorado, capas de la Estancia María Cristina y del Puesto Herminita, expuesto en las cercanías del cerro Colorado y del río Irigoyen. Referencias indicadas en la Figura 4. Adaptado de Malumián y Olivero (2005). 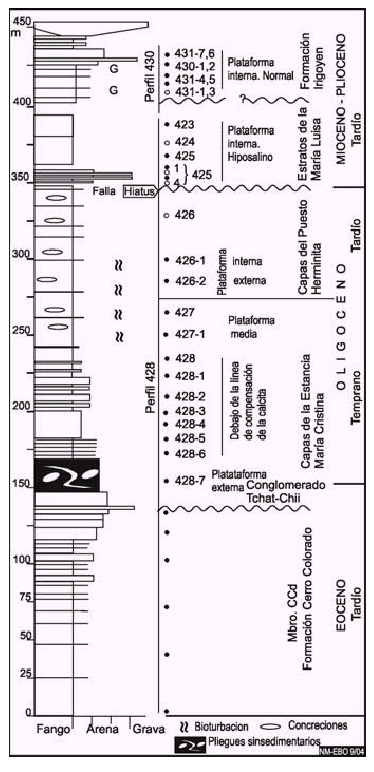
En el área tipo, dominan espesos paquetes de conglomerados gruesos sobre lentes de areniscas gruesas y conglomerados finos con abundantes fragmentos de conchilla, de color de conjunto castaño oscuro. Los conglomerados gruesos son clasto-sostenidos con abundante matriz arenosa, componen capas de estratificación difusa, hasta 2-3 m de espesor, con frecuente gradación inversa y con los clastos mayores, de hasta 1,5 m de diámetro, imbricados y proyectados hacia el techo. Los clastos mayores corresponden en su mayor parte a areniscas subangulosas y concreciones elípticas. El resto de los clastos, con diámetros variables entre 3 y 40 cm, subredondeados a subangulosos, comprenden litologías variadas: riolitas y tobas, en su mayor parte foliadas; pizarras negras, cuarzo de veta; areniscas y concreciones calcáreas. En adición, se mencionaron fragmentos fósiles derivados, que incluyen desde amonites hasta restos óseos (Malumián 1988). La fuente de proveniencia de los clastos comprende a rocas andinas jurásicas, cretácicas y paleógenas, las últimas incluyen clastos de areniscas induradas del Grupo La Despedida.
En el área tipo, el Conglomerado Tchat-Chii tiene un espesor mínimo de 70 m y sus capas inclinan hasta 20º hacia el SO. La base está subexpuesta, pero se interpreta como una marcada discordancia dada la composición clástica y la distinta actitud estructural respecto a las rocas infrayacentes. El techo también está subexpuesto, pero muestras extraídas con barreno en la parte superior del cerro Tchat-Chii, indican que el Conglomerado grada rápidamente a arcillitas arenosas seguidas por las arcillitas de las capas del Puesto Herminita.
En el cerro Colorado, incluye los conglomerados finos y areniscas gruesas (aproximadamente 10 m de espesor, Fig. 5, perfil 428) situados en la base de los estratos innominados de Malumián y Olivero (2005). En la punta Gruesa, incluye las capas basales del ciclo b de Ponce et al. (2004), aproximadamente 20 m de espesor, caracterizadas por conglomerados finos, areniscas conglomerádicas y areniscas medianas a gruesas (Fig. 3, perfil 409). En ambos perfiles, estas capas fuertemente plegadas se apoyan discordantemente sobre areniscas finas y fangolitas del Miembro CCd de la Formación Cerro Colorado y estratigráficamente hacia arriba pasan en forma neta o transicional a la capas de la Estancia María Cristina.
Edad y ambiente de depositación: En el área tipo, la posición estratigráfica intermedia entre el Grupo La Despedida y las capas del Puesto Herminita, acotan la edad al Eoceno tardío-Oligoceno temprano (cf. Malumián 1988). Las capas conglomerádicas en el cerro Colorado están aproximadamente a 135 metros por encima del aparente dato de primera aparición (DPA) de Isthmolithus recurvus Deflandre y los niveles con Praetenuitella insolita (Jenkins) en la punta Gruesa, ocupan una posición estratigráfica intermedia entre el DPA y el dato de ultima aparición (DUA) de I. recurvus y su edad queda restringida entre los 36 y 31,8 Ma. En ambas localidades el DUA de I. recurvus se encuentra a unos pocos metros sobre la base de estas capas (Fig. 5, perfil 428, muestra 428-7 y Fig. 3, perfil 409, muestra PG2). Los conglomerados gruesos del cerro Tchat-Chii corresponden a depósitos de flujos gravitatorios inerciales, del tipo de los flujos hiperconcentrados (Mulder y Alexander 2001), que se interpretan como depósitos de abanico deltaico (fan-delta). Los conglomerados de la punta Gruesa han sido interpretados como flujos gravitatorios submarinos profundos (hiperpicnitas) generados por avenidas fluviales excepcionales (Ponce et al. 2004).
Correlaciones: El Conglomerado Tchat-Chii se correlaciona con el glauconítico A, interpretación ya dada por Manson en 1961 (cf. Malumián 1988). Aparentemente, la edad del glauconítico A, varía desde Eoceno cuspidal hasta el Oligoceno temprano y se ha considerado que incluye dos unidades glauconíticas separadas por una discordancia (cf. Robiano et al. 1996).
En posiciones de depocentro, tal como puede datarse en el pozo Poseidón, el glauconítico A por su contenido de P. insolita, un excelente fósil guía de altas a medias latitudes (cf. Jenkins 1995), es equivalente al miembro CCd de la Formación Cerro Colorado. Una edad oligocena temprana (cf. Masiuk et al. 1990) y al menos post 31,8 Ma por la ausencia de I. recurvus, (cf. Mostajo 1991) es asignable para el glauconítico A en el subsuelo septentrional de la Isla.
Probablemente el glauconítico A incluya dos paquetes separados por una discordancia, uno del Eoceno superior, miembro CCd de la Formación Cerro Colorado; y otro del Oligoceno inferior, Conglomerado Tchat-Chii. En tal sentido se describe brevemente al miembro CCd, con nueva información que avala su distinción como una formación independiente, excluida del Grupo La Despedida, cuya formalización queda pendiente a la espera de una revisión general de la estratigrafía del Cenozoico fueguino.
Miembro CCd de la Formación Cerro Colorado
Antecedentes: El miembro CCd de la Formación Cerro Colorado fue establecido por Olivero y Malumián (1999). Corresponde al "ciclo a" de Ponce et al. (2004).
Distribución y litología: En el perfil tipo de la Formación Cerro Colorado (Figs. 1, 2 y 5), el Miembro CCd comprende 140 m de fangolitas con intercalaciones de areniscas limosas, seguidas en su parte superior por areniscas. También se lo reconoce en la punta Gruesa (Figs. 1 y 3), donde tiene una similar composición litológica. La asociación mineralógica de las arcillas es: esmectita 4%, illita 60% y clorita-caolinita 34%.
Paleontología: El miembro CCd, en cerro Colorado, contiene una típica asociación de tenuitellidos lisos (Olivero y Malumián 1999), junto con I. recurvus, (Olivero y Malumián 2005).
En la punta Gruesa, las muestras PG 2 se caracterizan en sus foraminíferos planctónicos por la aparente última aparición de Globigerinatheka index (Finlay) asociada a I. recurvus, en niveles con abundante y muy grandes ejemplares de Chiloguembelina ototara (Finlay), carentes o con escasos G. index, en una relación como la descrita por Li et al. (1995): la abundancia y hasta la presencia de Chiloguembelina está asociada a ausentes o escasos G. index. Si bien la microfauna es coherente en la autoecología de sus especies componentes que avalan la autoctonía, dada la posición de la muestra PG2, intercalada entre niveles conglomerádicos, existe la posibilidad de que se trate de material retrabajado, que en el caso del nanoplancton calcáreo es sumamente probable.
En los foraminíferos bentónicos, se caracterizan por muy abundante Bolivina sp. asociada con:
Anomalinoides orbiculus Stache
Cibicides sp.
Gyroidinoides sp.
Lagena acuticosta Reuss
Nodosaria longiscata d'Orb.
Stilostomella spp.
Radiolarios abundantes por niveles.
Foraminíferos aglutinados:
Bathysiphon sp.
Reticulophragmium sp.
Rhabdammina sp.
Paleoambiente: El miembro CCd indica una profundización dentro de la Formación Cerro Colorado por su mayor contenido en foraminíferos planctónicos, y puede interpretarse como el inicio de la profundización de la cuenca. Junto con algunos niveles del glauconítico A comparte a P. insolita que ha sido comparada con Parvularugoglobigerina eugubina. Ambas especies son productos de profundos desequilibrios oceánicos; una vinculada al Eoceno terminal, y la otra, al límite Cretácico/Paleógeno, respectivamente (cf. Li et al. 1995).
La abundancia de Chiloguembelina, Bolivina, y Uvigerina, por niveles en el miembro CCd, y los niveles con grandes nódulos fosfáticos reconocidos recientemente en el miembro CCc de la Formación Cerro Colorado, sugieren condiciones ricas en nutrientes. Las microfaunas de las muestras PG 1 y 2 (Fig. 3), de plataforma externa, son las primeras sedimentitas desde el Paleoceno con indiscutibles características micropaleontológicas de ambiente profundo por su relativo alto contenido en foraminíferos planctónicos. La parte basal de la sección de la punta Gruesa (Fig. 3, perfil 409) contiene grafoglíptidos (Ponce et al. 2004), que constituyen trazas fósiles típicas del flysch.
El inusual bajo contenido de esmectita, y muy alto de illita, es el probable resultado de la coincidencia en las condiciones de enfriamiento del Eoceno terminal amplificado por el incremento de la actividad tectónica.
Capas de la Estancia María Cristina Oligoceno basal. Zona P18, 34-30 Ma
Antecedentes: Son las capas mencionadas como La María Cristina beds por Olivero y Malumián (2005). Comprenden parte de las capas referidas como Formación Cerro Colorado de Ghiglione (2002), y a los depósitos del "ciclo b" de Ponce et al. (2004).
Distribución y litología: Los principales afloramientos de referencia se encuentran en un corte artificial sobre la ruta nacional 3 (Figs. 1 y 2, Localidad 4), situado aproximadamente 6 km al N de la Estancia María Cristina, y sobre la costa atlántica, en los aledaños de la punta Gruesa y del cerro Colorado (Figs. 1-5). Por otra parte, se reconocen pequeños asomos de fangolitas (gris mediano 5, secas; gris oscuro 3, húmedas) del Oligoceno inferior (muestras 318-6 y 7) sobre la margen izquierda del río Irigoyen, unos 20 km al O de la costa atlántica (Fig. 1), que se incluyen en las capas de la Estancia María Cristina.
En el corte comprende un espesor mínimo de 100 m de arcillitas gris oliva claro (5y 6/1), muy homogéneas, masivas, con un horizonte de concreciones calcáreas de donde proviene la mayor parte de la microfauna calcárea. En la punta Gruesa, un espesor mínimo de 175 m que consta en su porción inferior de bancos alternantes de areniscas medianas a finas y fangolitas (gris oliva a gris oliva claro, 5y 4/1-6/1) (Fig. 3, perfil 409), seguidas de arcillitas masivas con concreciones calcáreas (Fig. 3, perfil 463). En el cerro Colorado comprende aproximadamente 130 m, su porción basal incluye un paquete alternante de areniscas y fangolitas, con notable plegamiento sinsedimentario, seguido de una sucesión bien estratificada de capas semejantes y culmina con fangolitas y arcillitas masivas con concreciones calcáreas, de igual color que en punta Gruesa (Fig. 5, perfil 428).
En el corte, las capas inclinan 30º hacia el S con la base y el techo ocultos. En la punta Gruesa, las capas, subverticales e invertidas, conforman un anticlinal volcado hacia el N, la base es transicional con el infrayacente Conglomerado Tchat-Chii. En el perfil 409 (Fig. 3) su porción superior está cortada por la falla frontal que limita la faja plegada y corrida fueguina, y en el perfil 463 (Fig. 3) los contactos están dados por fallas. En el cerro Colorado, la porción basal con pliegues sinsedimentarios se asienta en forma neta sobre el Conglomerado Tchat-Chii y su techo pasa en forma transicional a las capas del Puesto Herminita (Fig. 5, perfil 428). En esta localidad las capas basales inclinan aproximadamente 50º al S y estratigráficamente hacia arriba disminuyen la inclinación hasta 15º.
Paleontología: Debido a que las capas se depositaron en su mayor parte por debajo de la LCC, solo se encuentran microfósiles calcáreos en niveles sumamente restringidos, sin que se hayan reconocido megafósiles. En afloramientos próximos a la Estancia María Cristina (Figs. 1 y 2, localidad 4) dentro de un conjunto de conchillas fuertemente seleccionadas hacia tamaños mayores, se reconocieron los siguientes foraminíferos calcáreos:
Foraminíferos bentónicos:
Angulogerina cf. angulosa Williamson
Antarcticella antarctica (Leckie y Webb)
Bolivina sp.
Cibicides sp.
Laticarinina pauperata (Parker y Jones)
Globocassidulina sp.
Parafissurina lateralis carinata Bucher
Oridorsalis umbonatus (Reuss)
Uvigerina subproboscidea Haque
Valvulineria sp.
Foraminíferos planctónicos:
Catapsydrax dissimilis (Cushman y Bermúdez)
Chiloguembelina cubensis (Palmer)
Globigerina ampliapertura Bolli
Subbotina angiporoides angiporoides (Hornibrook)
Zeaglobigerina euapertura (Jenkins)
Z. woodi (Jenkins)
"Globigerina" brevisJenkins
Nanoplancton guía: I. recurvus Deflandre
En el área del cerro Colorado, las muestras 428 a excepción de la 428-7, en más de 150 m de espesor, solo brindaron foraminíferos aglutinados (Fig. 5, perfil 428):
Ammodiscus sp.
Cribrostomoides subglobosus (G.O.Sars)
C. yeffreysii (Williamson)
Haplophragmoides bradyi (Robertson)
Karrerulina sp.
Martinottiella sp.
Psammosphaera sp.
Reticulophragmium acutidorsata (von Hatken)
R. amplectens (Grzybowski)
Rhabdammina spp.
Rhizammina sp.
Sólo en las muestras 428-7 y 427 se encontraron foraminíferos calcáreos. En la 428-7, sobre 160 ejemplares de foraminíferos bentónicos se reconocen los siguientes porcentajes:
Globocassidulina pseudocrassa (Hornibrook) 60%
U. subproboscidea Haque 24%
Stilostomella spp. 10%
menores al 4%:
Cibicides sp.
Fissurina sp.
Gyroidinoides sp.
Hoeglundina elegans (d' Orb.)
Lagena sp.
Lenticulina sp.
Nodosaria longiscata d' Orb.
Oridorsalis sp.
Pullenia spp.
Los foraminíferos planctónicos: S. angiporoides junto con muy abundante Tenuitella gemma (Jenkins).
En la muestra 427 se recuperó escasa microfauna, con Epistominella sp., Martinottiella sp., Haplophragmoides sp., Stilostomella spp., N. longiscata, Cibicides spp. y S. angiporoides.
En 427-1, material muy escaso, muy seleccionado por tamaños menores, con Bulimina cf. mexicana Cushman, Epistominella sp., Gyroidinoides sp., N. longiscata, R. amplectens?, Rhabdammina sp. y Stilostomella sp.
En punta Gruesa, en el Perfil 463 (Fig. 3):
Ammodiscus sp.
Bathysiphon sp.
Haplophragmoides sp.
Psammosphaera sp.
Reticulophragmium sp.
Muy escasos calcáreos:
Heterolepa sp.
Lenticulina alatolimbata (Gümbel)
Nodosaria longiscata d' Orb.
U. subproboscidea Haque
En la muestra 463-4, se encuentra abundante C. dissimilis junto con S. angiporoides acompañados por tenuitellidos relativamente abundantes: T. gemma y T. munda Jenkins.
En la punta Gruesa, las capas basales se caracterizan por horizontes densamente bioturbados, con dominancia de Phymatoderma granulata (Schlotheim), una icnoespecie frecuente en depósitos marinos profundos del flysch europeo y de Ecuador (Olivero et al. 2004).
Edad: La edad de las capas está entre 34,3 y 32 Ma. La ausencia de G. index DUA 34,3 Ma, y la presencia de C. dissimilis indica una edad menor al Eoceno; la de S. angiporoides, mayor a 30 Ma, y la de I. recurvus, mayor a 32 Ma.
La sección del área del cerro Colorado (muestras 427-8 a 428, Fig. 5, perfil 428) se asigna claramente por los foraminíferos planctónicos al Oligoceno temprano, por S. angiporoides y la ausencia de tenuitellidos típicos del Eoceno tardío. Se corresponde con la Zona de S. angiporoides, reconocida en Antártida y Nueva Zelanda, típica de áreas templadas y frías. La muestra 427 contiene la última aparición de S. angiporoides, calibrada en 30 Ma, en la mitad de la Zona P20, levemente por debajo del límite oligoceno inferior/superior.
En general, en las fangolitas del perfil 463 (Fig. 3, muestras 463, 464, 465) se recuperaron frecuentes foraminíferos aglutinados y muy escasos bentónicos calcáreos. En particular, la muestra 463-4 tiene una asociación de foraminíferos planctónicos que resulta ser muy típica de la capas de la Estancia María Cristina. Las fangolitas apoyan sobre las PG 2, del Eoceno cuspidal con G. index y I. recurvus. Entre los foraminíferos bentónicos, las capas basales (Fig. 5, perfil 428, muestra 428-7) brindaron material relativamente abundante, bien conservado, conformando una asociación peculiar por el dominio de G. pseudocrassa y la abundancia de U. subproboscidea. La primera se conoce desde el Oligoceno inferior tardío al Aquitaniano de Nueva Zelanda con una aparición posterior a la última aparición de S. angiporoides (cf. Hornibrook et al. 1989). En los términos superiores se encuentra el registro más antiguo de Spirosigmoilinella compressa Matsunaga, (muestra 318-6), con S. angiporoides pero sin I. recurvus.
Paleoambiente: Son sedimentitas con asociaciones residuales de foraminíferos aglutinados depositadas en casi su totalidad por debajo de LCC. En los eventuales niveles con foraminíferos calcáreos, U. subproboscidea es abundante a dominante y parece ser muy típica de estas capas, es una especie de aguas profundas y también abundante en el Oligoceno del DSDP Site 329 (50°S, 44°O) del plateau de las islas Malvinas (cf. Boersma 1984).
Correlación: En conjunto se correlacionan las capas de la Estancia María Cristina con el glauconítico A del subsuelo de Tierra del Fuego y la parte inferior de las "margas superiores", los que en general tiene una edad levemente menor en la porción septentrional de la Isla, pues allí carecen de I. recurvus, (cf. Mostajo, 1991) y de tenuitellidos lisos. Sólo en situaciones próximas a la región andina comprenden términos con tenuitellidos lisos junto con Ch. ototara, como se presentan en el pozo Poseidón. En este pozo, el glauconitico A y la margosa superior están entre 2.000 y 2.300 mbbp. Riggi (1988) señaló que la parte inferior del glauconítico A se caracteriza, y aún sería identificable, por su bajo contenido en montmorillonita. En este sentido, el miembro CCd, reconoce el valor más bajo en esmectita, y también coincide con la edad del glauconítico A en las partes más profundas de la cuenca.
Capas del Puesto Herminita. Oligoceno inferior alto-Oligoceno superior bajo, dentro de los 30 y 26,5 Ma
Antecedentes: Fueron mencionadas como "limolitas aflorantes en las proximidades de la estancia la Herminita" en estudios micropaleontológicos (Malumián y Náñez 1988, Malumián y Caramés 1989). Corresponden en parte a las capas diferenciadas por Ghiglione (2002) como Mioceno inferior en las cercanías de la punta Gruesa.
Distribución y litología: Estas capas asoman en las crestas de las lomas de las inmediaciones del Puesto Herminita a través de una fina cobertura regolítica (Figs. 1 y 2, localidad 6). Otros afloramientos semicubiertos se sitúan en las inmediaciones de la falla Fagnano, sobre la margen izquierda del río Irigoyen a unos 20 km aguas arriba de su desembocadura y al E del Aserradero Fueguino (Fig. 1). Las mejores exposiciones están sobre la costa atlántica, en las inmediaciones de la punta Gruesa (Fig. 3, perfil PG10) y del cerro Colorado (Fig. 5, perfil 428).
En el Puesto Herminita, las capas alcanzan un espesor mínimo de 200 m, con base y techo ocultos, dominantemente de arcillitas, limolitas arenosas masivas muy subordinadas, en parte con raros horizontes concrecionados y cementados con carbonato (varían entre gris amarillento 5Y 7/2, húmeda; anaranjado amarillento oscuro 5Y 6/4 seca, a anaranjado grisáceo 10Yr 7/4 a pardo amarillento moderado 10Yr 5/4, respectivamente) muy poco expuestas y que conforman un suave anticlinal cuyos limbos inclinan menos de 15º.
En la punta Gruesa comprenden un paquete de 200 m de espesor mínimo (Fig. 3, perfil PG 10) de areniscas y fangolitas en su parte inferior, rítmicamente estratificadas, seguidas de fangolitas grises con concreciones elípticas. El paquete de estratificación rítmica, consta de areniscas finas y medianas, gris claras, en bancos de 0,10-0,60 m de espesor, con gradación normal y laminación paralela, con frecuente deformación convoluta. Las capas conforman un anticlinal asimétrico, con inclinaciones de hasta 60º cerca del eje de la estructura y sobre el limbo N las inclinaciones disminuyen paulatinamente hasta 20º, aproximadamente. El corrimiento Punta Gruesa monta a las capas de la Estancia María Cristina sobre las capas del Puesto Herminita.
En el área del cerro Colorado-río Irigoyen (Fig. 5, perfil 428) comprende una sucesión homogénea, espesor mínimo 75 m, de arcillitas grises con intercalaciones delgadas de fangolitas arenosas, que incluyen niveles con concreciones calcáreas. Las capas inclinan 15º hacia el S. La base es transicional con las capas de la Estancia María Cristina y hacia el techo el contacto con los suprayacentes estratos de la María Luisa está dado por falla (Malumián y Olivero 2005).
Paleontología y edad: Pese al intenso muestreo de las capas en las proximidades del Puesto Herminita mediante barreno, sólo en una localidad se reconocieron microfósiles calcáreos, únicamente los foraminíferos monoloculares fueron ilustrados por Malumián y Caramés (1989).
Los foraminíferos aglutinados son dominantes y conforman la típica asociación residual de Spirosigmoilinella compressa-Martinottiella spp., que son dominantes.
Entre los foraminíferos aglutinados se reconocen:
Eggerella bradyi (Cushman)
Karrerulina sp.
Martinottiella sp.
Pseudobolivina sp.
Reophax sp.
Reticulophragmium spp.
Sacammina sp.
Spirosigmoilinella compressa Matsunaga
Foraminíferos bentónicos calcáreos uniloculares:
Buchnerina orbignyana (Seguenza)
Buchnerina sp. 1330
Fissurina fimbriata (Brady)
F. globosa Bornemann
F. staphyllearia Schwager
Lagena raricosta (d'Orb.)
Lagenosolenia flintiana (Cushman)
Oolina lineata (Williamson)
Parafissurina basicarinata Parr
P. dohrnii (Buchner)
P. textulostoma Loeblich y Tappan
Foraminíferos bentónicos calcáreos multiloculares:
Cassidulinoides mexicanus Cushman
Gyroidinoides neosoldanii (Brotzen)
L. pauperata (Parker y Jones)
Lenticulina sp.
Orthomorphina sp.
Pleurostomella sp.
Proxifrons advena (Cushman)
Sigmomorphina sp.
Stilostomella sp.
Uvigerina cf. topilensis Cushman
Uvigerina cf. ongleyi Finlay
Foraminíferos planctónicos:
Zeaglobigerina connecta (Jenkins)
Z. labiacrassata (Jenkins)
Catapsidrax unicavus Bolli, Loeblich y Tappan
Nanoplancton característico: Contiene el último registro del género Chiasmolithus, con Ch. altus Bukry y Percival reconocido en la muestra PG 16 (Fig. 3, perfil PG10). En las proximidades del Conglomerado Tchat-Chii, que aparentemente corresponden a la base de las capas, contiene muy abundantes microfósiles calcáreos retrabajados del Eoceno, particularmente del Eoceno medio.
En el área del cerro Colorado, se reconocen las capas de la Herminita en el tramo 426-2, a 426 (Fig. 5, perfil 428), que contiene relativamente escasos foraminíferos bentónicos y muy abundantes planctónicos regularmente conservados con dominio del grupo 3 de "grandes" globigerínidos de Spezaferri (1994): "Globigerina" venezuelana Hedberg, "Globigerina" euapertura Jenkins, y además Z. labiacrassata y "Sphaeroidinellopsis" sp., que conforman una asociación típica del Oligoceno superior.
Los foraminíferos bentónicos son:
Anomalinoides macraglabra Finlay
Bulimina sp. (idénticas a las reconocidas en la muestra PG 16)
Glabratella crassa Dorreen
Globobulimina sp.
Globocassidulina sp.
Gyroidinoides subzelandicus Finlay
L. pauperata (Parker y Jones)
Lobatula lobatula (Walker y Jacob)
Nummodiscorbis sp.
Patella corrugata Williamson
Planulina mexicana Cushman
Planulina sp.
Pleurostomella sp.
Trochoelphidiella sp. de Leckie y Webb (1990)
Triseriales calcáreos indeterminados muy pequeños.
La muestra 425-1, correspondiente a los estratos de la María Luisa, contiene microfauna diferente (cf. Malumián y Scarpa 2005).
La ausencia de I. recurvus y S. angiporoides indica una edad menor a 30 Ma, la presencia de Z. labiacrassata (Jenkins), de rango Oligoceno tardío a Mioceno Medio temprano (cf. Spezzaferri 1994) restringe su edad más antigua, el dominio de grandes globigerínidos es típica del Oligoceno superior. Las capas de la Herminita contienen los primeros registros de abundante S. compressa cuya primera aparición se encuentra entre las últimas apariciones de I. recurvus y S. angiporoides.
Entre los foraminíferos bentónicos, una forma comparable a Uvigerina ongleyi Finlay, idéntica a la ilustrada por Boersma (1984), es típica de las capas de la Herminita. En Nueva Zelanda se reconoce desde la base del Oligoceno al Mioceno medio. Ha sido ilustrada como especie indicadora del techo del piso Rosariano (Natland et al. 1974), de manera que se correlaciona con este piso y con la parte superior de la "margosa superior", que en el pozo Poseidón está entre 1.770 y 2.000 mbbp.
Paleoambiente: El absoluto dominio de foraminíferos aglutinados en conjuntos muy pobres en la mayoría de las muestras indica ambientes por debajo de la LCC, la asociación de Spirosigmoilinella-Martinottiella es indicadora de aguas corrosivas de carácter antártico (Malumián y Náñez 1991). Los escasos ejemplares calcáreos son formas resistentes a la disolución y sugieren ambientes profundos de la lisoclina, comprenden típicas formas de carácter antártico, como Antarcticella en los multiloculares, y Fissurina fimbriata en los monoloculares. La relativa abundancia de radiolarios esféricos, dientes de peces e ictiolitos, y niveles con abundante Bolivina, aguas superficiales productivas.
Las areniscas gradadas de la sucesión rítmica inferior se interpretan como turbiditas, y en conjunto con la información paleoecológica, se asimilan a lóbulos arenosos de ambiente marino profundo.
2) CICLO OLIGOCENO CUSPIDALMIOCENO TEMPRANO (P22-N4)
Formación Desdémona
Oligoceno cuspidal-Mioceno basal
Antecedentes: Corresponde al miembro informal A del Mioceno inferior de Ghiglione (2002) y parcialmente a los estratos del cabo Inés (Buatois y Camacho 1993). El origen del nombre alude al barco Desdémona, encallado en la playa cercana al área tipo.
Distribución y litología: La Formación está muy bien expuesta en el acantilado y la restinga al SSE del cabo San Pablo donde se designa el perfil tipo (Fig. 1, Fig. 4, perfil 411).
La Formación Desdémona está caracterizada principalmente por fangolitas tobáceas, inusualmente induradas en cortes frescos para las sedimentitas de similar edad. La Formación, ca. 250 m de espesor, constituye una sucesión dominantemente de arcillitas y fangolitas bioturbadas, gris oliva claro (5Y 6/1) seca y gris oliva (5Y 4/1) húmeda, en parte con marcados rasgos de deformación sinsedimentaria, entre los que se mencionan numerosos diques clásticos de arenisca (Ghiglione 2002), grandes bloques deslizados y replegados de arcillita-fangolita, laminación convoluta y espesos horizontes con concreciones removilizadas, con pérdida de la fábrica sedimentaria original en las fangolitas que las contienen (Fig. 4, perfil 411). En su porción cuspidal, consta de bancos de arenisca muy fina, bioturbadas y con glauconita madura. La asociación de arcillas (sobre el análisis de 7 muestras) se caracteriza por valores constantes de 30% de illita, y variables entre 30 a 50% de esmectita, y de 20 a 30% de clorita.
La base no está expuesta y el techo está cubierto discordantemente por los estratos del cabo Ladrillero. Los estratos están suavemente plegados, con inclinaciones variables entre 5º y 15º (Fig. 1, Ghiglione 2002).
Paleontología y edad: Los megafósiles, muy escasos en general, se reconocieron en seis niveles, con gasterópodos, bivalvos, corales solitarios y equinoideos, mal preservados por disolución.
La bioturbación es relativamente conspicua y en general en toda la Formación se intercalan niveles densamente moteados; en sus niveles basales abundan Zoophycos y Chondrites, hacia el nivel de 200 m (Fig. 4, perfil 411) hay capas totalmente bioturbadas con dominancia de Zoophycos, Taenidium y Teichichnus. Las areniscas cuspidales están intensamente recorridas por galerías de Thalassinoides, en parte con un inusual relleno de láminas curvadas, de tipo chevron.
Los foraminíferos bentónicos calcáreos, y que también se encuentran en las capas de Cabo Ladrillero en el perfil 410, son:
Amphimorphina amchitkaensis Todd
Bolivina malovensis Heron-Allen y Earland
Gyroidina soldanii (d'Orb.)
Pullenia bulloides d'Orb.*
Sphaeroidina bulloides (d'Orb.)
S. compressa Matsunaga
Uvigerina farinosa (Hantken)
Se reconocen además, los foraminíferos bentónicos calcáreos:
Angulogerina angulosa (Williamson)
Anomalinoides spissiformis (Cushman y Staintforth)
Astrononion echolsi Kennett
Buccella peruviana (d'Orb.)
Bueningia sp.
Bulimina inflata Seguenza
Globocassidulina subglobosa (Brady)*
Cassidulina subtumida Cushman
Cibicidoides wuellerstorfi (Schwager)
C. bradyi Trauth
C. mundulus B.P. y Jones
Dentalina consobrina d'Orb.
Elphidium macellum Fitchel y Moll
Epistominella exigua Brady
Gavelinella cagnoni Malumián
Glandulina obtusissima Reuss
Gyroidina neosoldanii (Brotzen)
Heronallenia wilsoni (Heron-Allen y Earland)
Heterolepa sp.
L. pauperata (Parker y Jones)
Lenticulina sp.
Marginulina obliqua d'Orb.
Nodosaria longiscata d'Orb.
N. acuminata Hantken
Nonionella miocenica Cushman
Nummodiscorbis sp.
Oridorsalis umbonatus (Reuss)*
Patellina sp.
Planulinoides norcotti Hedley, Hurdle y Burdett
Pyrgo ringens Lamarck
Quinqueloculina stalkeri Loeblich y Tappan
Proxifrons advena (Cushman)
Stilostomella ex gr. lepidula Schwager
Uvigerina miozea Finlay
Valvulineria laevigata Phleger y Parker
Los foraminíferos aglutinados:
Ammodiscus sp.
Bathysiphon sp.
E. bradyi (Cushman)
Kalamopsis sp.
Karreriella siphonella Reuss
Martinottiella communis d'Orb.
Psammosphaera fusca Schulze
Reticulophragmium sp.
R. rotundidorsata (Hantken)
R. amplectens (Grzybowski)
Reophax pilulifer Brady
R. brevis Parr
R. placenta (Reuss)
Rhabdammina abyssorum M. Sars
Foraminíferos planctónicos:
"Globigerina" euapertura Jenkins
Globorotaloides suteri (Bolli)
Globigerinella obesa (Bolli)
Globigernita juvenilis (Bolli)
Catapsydrax dissimilis (Cushman y Bermúdez)
C. martini Blow y Banner
C. unicavus Bolli, Loeblich y Tappan
Praeglobigerinoides spp.
Sphaeroidinellopsis cf. disjuncta Finlay
Tenuitella clemenciae (Jenkins)
T. gemma (Jenkins)
Zeaglobigerina connecta (Jenkins)
Z. brasieri (Jenkins)
Z. woodi (Jenkins)
Catapsydrax dissimilis, cuyo DUA es 17,3 Ma; y la ausencia de Globorotalia incognita, una especie ampliamente distribuida en medianas altas latitudes y altas latitudes cuyo DPA es de 21,6 Ma, restringe la menor edad de la Formación Desdémona al Mioceno más temprano, edad coherente con el conjunto de foraminíferos planctónicos. La posición estratigráfica, por encima de las capas del Puesto Herminita, indica una edad no mayor al Chattiano medio. La Formación contiene "G." euapertura, que en la zonación subantártica (Berggren 1992) su última aparición fue utilizada como una aproximación del límite Oligoceno/Mioceno, pero es una especie que se reconoce hasta el Mioceno inferior en Australia (cf. Li et al. 2003).
Paleoambiente: Los foraminíferos bentónicos son indicativos de aguas profundas, las especies listadas con un asterisco, tienen su límite de distribución superior en el ambiente nerítico profundo. Entre otras especies batimétricamente diagnósticas, S. bulloides comúnmente se encuentra en ambiente batial superior; C. mundulus, en batial-abisal; L. pauperata, desde 300 metros de profundidad; E. bradyi, desde el batial medio (Morkhoven et al. 1986).
Los foraminíferos planctónicos, en las especies del género Zeaglobigerina, han sido consideradas por Chapronier (1992) como habitantes de masas de agua intermedia cálida-templada, donde dominan sobre los globigerínidos espinosos.
Signos de corrosión se manifiestan en casi todas las muestras, resultando altos porcentajes de foraminíferos aglutinados, que sugieren el dominio de condiciones de lisoclina. En los pocos casos (Fig. 4, perfil 411, muestra 28), en que se conservan formas calcáreas sin aparentes signos de disolución, la relación: foraminíferos planctónicos/ bentónicos, alcanza valores de 1,7, indicativos de ambientes de profundidades mayores a los de plataforma. Entre las muestras 411-20 y 43, donde no hay manifiestos efectos de disolución, los aglutinados están entre el 3 y el 50% de los foraminíferos bentónicos. El dominio de foraminíferos aglutinados hasta los 150 m (muestra 411-16), que componen del 76 al 99 % de los foraminíferos bentónicos, indica la posición de la lisoclina.
La Formación comprende depósitos marinos profundos, probablemente del talud depositacional, con frecuente removilización de sedimentos por deslizamientos gravitatorios.
Correlación: En la base de la Formación, se encuentra el registro más antiguo de Sphaeroidina bulloides. Si bien el biocrón reconocido para esta especie bentónica en los fondos oceánicos por Morkhoven et al. (1986) indica su rango a partir de la zona P19 del Oligoceno temprano tardío, tiene una aparición local relativamente tardía pues consistentemente no se ha registrado esta forma resistente a la disolución en las capas de la María Cristina ni en las del Puesto Herminita. Por lo que se correlaciona por su común contenido con la Formación Cabo Peña, avalada por la similar composición de las asociaciones de arcillas (Cuadro 1).
Cuadro 1: Composición de las Asociaciones de arcillas del Eoceno superior-Mioceno Medio. 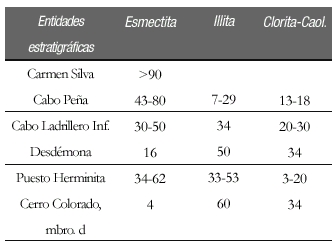
De la Formación Cabo Peña, pese a numerosos muestreos, se han recuperado muy escasos foraminíferos calcáreos. Su edad fue asignada al Eoceno tardío-Oligoceno temprano por Codignotto y Malumián (1981), y correlacionada con la Formación San Julián, (Malumián 1999). La misma edad fue asignada mediante palinología (Guerstein et al. 2002), pero los foraminíferos planctónicos son de grupos escasamente diagnósticos y de pobre preservación, y la probabilidad de retrabajo tanto de los microfósiles calcáreos como del material polínico del Eoceno, es muy grande en la Isla debido a la discordancia de la base del Grupo Cabo Domingo.
La edad de la Formación Desdémona es coincidente con la de la Formación Monte León, y equivalentes en toda la Patagonia y con niveles de mar alto.
Capas del Cabo Ladrillero
Mioceno inferior
Antecedentes: Corresponde al miembro informal A del Mioceno inferior de Ghiglione (2002) y parcialmente a los estratos del cabo Inés (Buatois y Camacho 1993).
Distribución y litología: Estas capas subhorizontales están bien expuestas sobre la costa atlántica, inmediatamente al NNO del área tipo de la Formación Desdémona (Fig. 1, perfil 410), en la base del cabo San Pablo (Figs.1 y 2, localidad 2; Fig. 4, perfil SP), y en el cabo Ladrillero; y también , sobre la ruta nacional 3 y en cortes de meandros del río Candelaria (Fig. 1, localidades 5 y 7, respectivamente).
Se reconocen dos secciones, una inferior, en el perfil 410 (Fig. 4), ca. 80 m de espesor mínimo, también cortada por numerosos diques clásticos, se asienta en suave discordancia angular sobre la Formación Desdémona. Su porción basal consta de areniscas muy finas, glauconíticas, con estratificación interna inclinada que traslapa sucesivamente hacia el SSE sobre el techo de la Formación Desdémona. El resto de la sección, se compone de una alternancia de paquetes espesos, estratificados y masivos, de fangolitas y arcillitas (gris medio nº5 gris oliva claro 5Y 6/1, 5Y seca; y gris oscuro nº3 a gris oliva 5Y 4/1, húmeda) (Fig. 4, perfil 410). La asociación de arcillas es invariable, en análisis sobre seis muestras, está formada por 20% de esmectita, 50% de illita y 30% de clorita, y una muestra proveniente del afloramiento sobre el río Candelaria, próximo a la Ea. El Rodeo, es de idénticos valores. La sección superior, en el perfil SP (Fig. 4) comprende un paquete de 75 m de espesor mínimo, de fangolitas carbonosas (en roca fresca gris verdoso oscuro 5GY 4/1 a gris oliva 5Y 4/1 con intercalación de bancos de arenisca fina (niveles entre 350-375 m) seguida de arcillitas gris violáceo, masivas (niveles entre 375-400 m) que culminan con un conspicuo horizonte con deformación sinsedimentaria. Esta sección se repite en el cabo Ladrillero.
Paleontología y edad: Además de las especies de foraminíferos bentónicos comunes con el perfil 411 de la Formación Desdémona, se reconocen en la sección inferior (perfil 410):
Astacolus gladious Philippi
Alabamina sp.
Bulimina alazanensis Cushman
B. fossa Cushman y Parker
Evolvocassidulina parkeriana (Brady)
Cibicides mckannai (Galloway y Wissler)
Dentalina spinecens Reuss
Duquepsammina cubensis (Cushman y Bermúdez)
Melonis pompilioides Fitchel y Moll
Plectofrondicularia parri Finlay
Pleurostomella elliptica Galloway y Heminway
Uvigerina rugosa d'Orb.
U. farinosa Hantken.
Foraminíferos planctónicos:
Catapsydrax africanus (Blow y Banner)
C. unicavus Bolli, Loeblich y Tappan
Globigerina praebulloides Blow
Globigerinita juvenilis (Bolli)
Globorotaloides suteri Bolli
Praeglobigerinoides sp. sensu Spezaferri
Sphaeroidinellopsis disjuncta Finlay
Tenuitella clemenciae Bermúdez
T. neoclemenciae Li
Tenuitellata pseudoedita (Subbotina)
Zeaglobigerina brasieri (Jenkins)
En la sección superior, en el cabo Ladrillero, las microfaunas están reducidas a abundantes foraminíferos aglutinados de la asociación Spirosigmoilinella-Martinottiella. En solo dos muestras de áreas sumamente localizadas se recuperaron foraminíferos calcáreos fuertemente seleccionados por tamaños menores, las formas bentónicas dominadas por bulimináceos: Bulimina alazanensis y B. fossa. Las formas planctónicas están dominadas por: G. praebulloides y Globigerina aff. praebulloides Bolli sensu Spezaferri y tenuitellidos: Tenuitella clemenciae y T. neoclemenciae.
Numerosas muestras analizadas del perfil SP (Fig. 4) brindaron sólo foraminíferos aglutinados dominados por S. compressa y Reticulophragmium sp. En la base se encuentra un surtido mayor, que incluye además Trochammina sp. y varias especies con la conchilla muy deformada.
El contenido en M. pompilioides y P. parri que según Morkhoven et al. (1986) se reconocen desde la Zona P22 a la N23, y desde la N4 a la N17, respectivamente, acotan la mayor edad de la sección inferior de las capas de Cabo Ladrillero (perfil 411) al Mioceno inferior. La edad de la sección superior, está acotada por las suprayacentes capas del Cabo San Pablo, expuestas en el cabo Santa Inés, al Mioceno inferior.
Paleoambiente: Los foraminíferos bentónicos batimétricamente diagnósticos, listados con un asterisco, tienen su límite superior en la plataforma externa. Entre otras especies significativas: U. rugosa se distribuye en ambientes batiales medios a inferiores (cf. Morkhoven et al. 1986).
Las muestras 410-24 a 18 contienen una microfauna residual de condiciones de disolución similar a las de la Formación Desdémona. En el resto del perfil 410 no se reconocen signos de disolución y la relación planctónicos a bentónicos, no supera a 0,7, indicando condiciones de plataforma externa al igual que los foraminíferos bentónicos. La microfauna residual del perfil SP indica condiciones de disolución por debajo de la LCC. El dominio de formas planctónicos espinosas del grupo de G. bulloides, un grupo considerado de aguas superficiales comúnmente asociado con aguas frías y ricas en nutrientes en el sudeste de Australia (cf. McGowran y Li 1993, Li y McGowran 1994) es coherente con el dominio de condiciones de disolución y la materia carbonosa abundante por niveles.
La asociación de arcillas indica un desmejoramiento climático y/o mayor tectonismo respecto a la asociación de la Formación Desdémona, por el aumento de illita a costa de la esmectita.
Capas del Cabo San Pablo
Mioceno inferior
Antecedentes: Corresponde parcialmente a los estratos del cabo Inés (Buatois y Camacho 1993) y a las capas del Cabo San Pablo (Olivero y Malumián 2005).
Distribución y litología: Constan principalmente de areniscas ocres, medianas y gruesas, sabulíticas, con glauconita relativamente escasa, masivas, con marcada geometría lenticular y frecuentes estructuras de corte y relleno (Fig. 4, perfil SP). Estas areniscas subhorizontales, muy bien expuestas sobre la costa atlántica entre el cabo San Pablo y cercanías del cabo Santa Inés, alcanzan un espesor mínimo entre 30-40 m. Los contactos de base y techo son discordantes. En el cabo Santa Inés, las areniscas están cubiertas concordantemente por ca. 15 m de espesor de areniscas finas y arcillitas, en parte con repliegues sinsedimentarios.
Paleontología y edad: De las areniscas masivas un par de muestras resultó estéril, y otra con S. compressa y foraminíferos aglutinados deformados mayormente indeterminables. En el cabo Santa Inés, las arcillitas superiores contienen dominantemente la asociación residual de Spirosigmoilinella-Martinottiella, solo en niveles muy restringidos se recuperó microfauna calcárea de pobre preservación. Los foraminíferos planctónicos si bien muy abundantes están mayormente aplastados o recristalizados, dominan totalmente Globigerina praebulloides, G. bulloides, G. aff. praebulloides sensu Spezaferri, con un solo ejemplar de C. dissimilis. Los foraminíferos bentónicos están reducidos a formas resistentes a la disolución y representados por grandes ejemplares de: S. bulloides, H. elegans, Cibicides spp., Uvigerina miozea, acompañados por raros ejemplares de U. rugosa. La posición estratigráfica de las capas del Cabo San Pablo y el DUA de C. dissimilis sugieren una edad miocena temprana.
Paleoambiente: El dominio del grupo de G. bulloides sugiere aguas frías, y las asociaciones residuales depositación mayormente por debajo de la LCC. Las areniscas masivas se interpretan como depósitos de flujos gravitatorios marinos profundos.
3) CICLO MIOCENO MEDIO
Formación Carmen Silva
(Codignotto y Malumián 1981)
Mioceno medio bajo
Antecedentes: La Formación fue propuesta por Codignotto y Malumián (1981) designando su perfil tipo en el puesto 26 (Fig. 1, localidad 10).
Distribución y litología: Los principales afloramientos se encuentran en el sector NO del área mapeada, en adición al perfil tipo del puesto 26 otras localidades con exposiciones relativamente buenas se sitúan en los alrededores de la Estancia San Julio (Fig. 1, localidad 9).
Consta de dos miembros, uno inferior de arcillitas y limolitas arenosas y otro superior de conglomerados y tobas, con abundantes invertebrados fósiles. Un rasgo conspicuo de los conglomerados es la abundancia relativa de clastos de volcanitas mesosilíceas y básicas. Las capas tienen actitud horizontal, el espesor mínimo alcanza ca. 50 m, la base no ha sido reconocida y el techo pasa transicionalmente a la Formación Castillo (cf. Codignotto y Malumián 1981). Esta formación está bien identificada en la perforación Aries e-2, donde se encuentra entre 330 a 500 metros bajo boca de pozo con microfauna típica de ambientes deltaicos e hiposalinos, con elevados porcentajes de esmectita en la asociación de arcillas (cf. Malumián et al. 1999).
Paleontología: Los foraminíferos bentónicos fueron ilustrados por Malumián (1982).
Foraminíferos bentónicos
Cyclogira involvens (Reuss)
Pyrgo sp.
P. oblonga (d'Orb.)
P. carinata (d'Orb.)
P. subsphaerica (d'Orb.)
Los foraminíferos planctónicos:
Globorotaloides suteri Bolli
Paragloborotalia pseudocontinuosa (Jenkins)
Tenuitellata pseudoedita (Subbotina)
T. selleyi Li, Reedford y Banner
Los moluscos fueron ilustrados por Malumián et al. (1978).
Edad: Recientemente, se dio a conocer el contenido de foraminíferos planctónicos (Olivero y Malumián 2005) que comprende mayormente tenuitellidos, los que son relativamente abundantes hasta el Mioceno medio (cf. Li et al. 1992). Se corresponde con la extendida transgresión entrerriense, coherente con el elevado contenido esmectítico de las asociaciones de arcillas y el momento de nivel de mar alto.
Paleoambiente: El contenido de foraminíferos es típico de ambientes someros, deltaicos, y marino marginales. La gran dominancia de esmectita en las asociaciones de arcillas, es interpretada como resultado del óptimo del Neógeno y un acontecimiento registrable por toda la Patagonia (cf. Alonso et al. 2001).
Capas del Cabo Viamonte
Mioceno medio
En forma preliminar se diferencian las capas del Cabo Viamonte (Figs. 1 y 2, localidad 3) por un conjunto de rasgos litológicos y faciales que las distinguen netamente del resto del Neógeno. Estas capas componen un paquete de 40 m de espesor mínimo, de brechas gruesas, conglomerados finos, areniscas gruesas y fangolitas, en capas de estratificación gruesa, masivas o con laminación paralela y ondulosa difusas, y con marcada geometría lenticular de gran escala, hasta más de 500 m de ancho, con múltiples superficies erosivas internas. Su base se asienta en marcada discordancia sobre las capas del Cabo San Pablo. Las brechas incluyen grandes bloques autóctonos resedimentados, hasta 3 m de diámetro; y los conglomerados finos, clastos de volcanitas basálticas frescas. La mayor parte de la sucesión clástica más gruesa se interpreta como depósitos de flujos gravitatorios densos y/o hiperconcentrados; y las litologías más finas, depósitos de decantación de fangos en suspensión. La marcada geometría lenticular, con superficies erosivas basales, se interpreta como originada por canales submarinos, rellenos por depósitos de flujos gravitatorios.
La falta de elementos paleontológicos diagnósticos dificulta el establecimiento de sus relaciones estratigráficas. Dada la semejanza en el estilo de estratificación y analogía de facies sedimentarias, es probable que corresponda a las mismas capas expuestas en el acantilado inferior y medio del cabo Domingo. Estas últimas fueron interpretadas como un probable equivalente lateral de la Formación Carmen Silva (cf. Codignotto y Malumián 1981). Se sigue aquí este criterio, con el agregado que probablemente las capas del Cabo Viamonte representen facies marinas profundas, equivalentes en edad al conjunto de Carmen Silva-Castillo, con las que comparte el contenido de clastos frescos de volcanitas.
Formación Castillo (De Ferrariis 1938)
Mioceno medio
Antecedentes: La Formación fue propuesta por De Ferrariis (en Fossa Manccini et al. 1938). El perfil tipo situado en el cerro Castillo (Fig. 1, localidad 9) fue descrito por Malumián et al. (1978) y los antecedentes de la Formación se mencionan en Codignotto y Malumián (1981).
Distribución y litología: Los principales afloramientos se sitúan en el sector NO del área mapeada, en general asociados a la infrayacente Formación Carmen Silva (Fig. 1). En la localidad tipo del cerro Castillo, incluye un espesor mínimo de 6 m de conglomerados y areniscas conglomerádicas con estratificación diagonal, que yacen en transición sobre conglomerados fosilíferos de la parte superior de la Formación Carmen Silva (Malumián et al. 1978). En las adyacencias del cerro Castillo, en el tramo entre las estancias San Julio y El Salvador, tiene mayor espesor (mínimo 20 m) y está compuesta de ciclos alternantes de 3-4 m de espesor, formados por conglomerados finos a medios, con estratificación diagonal, con bases erosivas; siguen areniscas conglomerádicas con estratificación diagonal; y culminan con delgados paquetes de areniscas y fangolitas rítmicamente estratificados, en parte casi totalmente eliminadas por erosión en la base de un nuevo ciclo. En las cercanías del puesto 26 (Fig. 1, localidad 10) consta de una asociación litológica similar a la anterior y alcanza un espesor mínimo de ca. 50 metros.
En todas las localidades estudiadas, los conglomerados incluyen clastos de volcanitas basálticas, de aspecto fresco, y con textura vesicular y amigdaloide.
Edad: Mioceno medio, debido a su pase gradual con la Formación Carmen Silva.
Paleoambiente: La ausencia de fósiles marinos, el estilo de estratificación y el común arreglo de facies en ciclos sedimentarios grano y estrato decreciente, sugiere ambientes fluviales.
Correlación: Se correlaciona con sedimentitas continentales asignadas a la Formación Santa Cruz con edades radimétricas de 16 Ma (cf. Feagle et al. 1995), correlación que ya había sido propuesta en forma generalizada por De Ferrariis (en Fossa Manccini et al. 1938).
DISCUSIÓN Y CONCLUSIONES
Sobre la base de los datos del Grupo Cabo Domingo, resumidos en forma esquemática en las figuras 2 y 6 y enmarcados entre ca. 50 y 15 Ma, se discute la bioestratigrafía y la evolución geológica, subrayando la calibración y el valor de las discordancias, variación de las tasas de sedimentación y la evaluación del control tectónico y/o eustático en la generación de espacio de sedimentación.
Figura 6: Columna estratigráfica. Datos de aparición y desaparición de los principales marcadores bioestratigráficos con las calibraciones de Berggren et al. (1995). Comparación con la curva eustática (Haq et al. 1987), caídas de temperatura reconocidas en áreas antárticas (Stott et al. 1990, Zachos et al. 2001), y diversos acontecimientos australes. 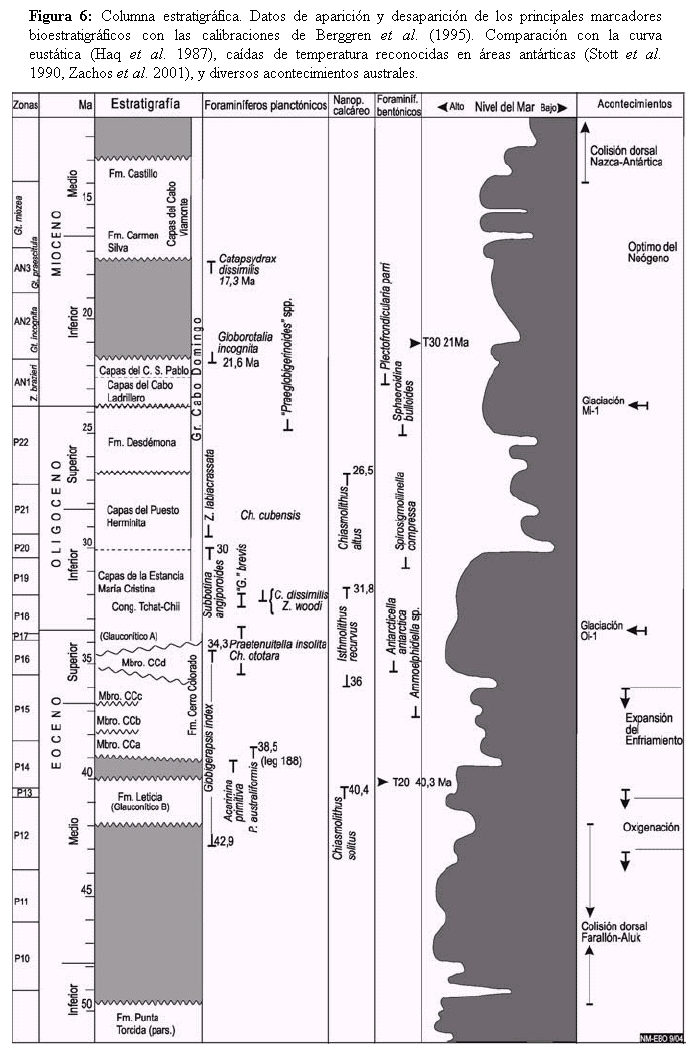
Bioestratigrafía
Antecedentes: Un sistema de pisos para la cuenca Austral, basado mayormente en foraminíferos bentónicos, fue propuesto por Natland et al. (1974). El sistema adolece de dos grandes defectos: uno, la designación de dos localidades tipos para cada piso que crea una indeterminación desde su propia propuesta, y el otro, la muy fuerte apoyatura en los foraminíferos bentónicos que aún en distancias muy cortas presentan dificultades de correlación (cf. Codignotto y Malumián 1981).
Listas de foraminíferos para las sedimentitas del subsuelo fueron provistas por Masiuk et al. (1990), que al estar agrupadas por extensas formaciones, y sin que se detallen las últimas apariciones, resultan de muy escasa resolución. Las listas de planctónicos contienen especies de biocrones incompatibles, y las de bentónicos mayormente son genéricas. El planteo estratigráfico reúne desde las "margas inferiores" hasta las "superiores" en una sola formación, sin tomar en cuenta las discordancias que comprenden, incompatibles con una sola unidad formacional.
Las distribuciones de los nanofósiles calcáreos y las asignaciones cronológicas fueron dadas para el pozo Las Violetas, en el extremo septentrional del sector argentino de la Isla (Mostajo 1991).
Estudios sobre elementos palinológicos han sido realizados para la Formación Cabo Peña y en la sección La Despedida (cf. Guerstein et al. 2002), y la Formación Cullen. Dadas las muy frecuentes superficies erosivas y discordancias de diferentes magnitudes en la Isla, el retrabajo en los microfósiles es común, y puede decirse que casi constante en los microfósiles más pequeños como los elementos palinológicos y de la nanoflora. Edades basadas en los primeros, como en el caso de la Formación Cullen, del extremo septentrional de la Isla, de clara edad post miocena, fueron repetidas veces asignadas al Eoceno (cf. Zamaloa y Romero 1990); de la que se han descrito ricas microfloras del Eoceno medio (Zetter et al. 1998) o del Oligoceno tardío-Mioceno medio (Zamaloa 2000). Curiosamente, se han reconocido abundantes elementos retrabajados, del Cretácico y Paleoceno, pero no se han considerado como retrabajados los de edades menores, que seguramente son los más propicios en ser resedimentados.
Estudios sistemáticos parciales sobre foraminíferos han sido referidos, junto con listados del contenido de las especies recientemente por Olivero y Malumián (2005).
Características generales de las microfaunas cenozoicas de Tierra del Fuego
Las entidades estratigráficas reconocidas desde el Cretácico superior en Tierra del Fuego tienen microfaunas tan distintivas y diferentes entre sí, que sugieren en principio contactos discordantes, que comprenden hiatos de distinta magnitud. En muchos casos, estas diferencias se expresan incluso entre los miembros de las formaciones. Las microfaunas desde el Campaniano pueden diferenciarse en tres grandes conjuntos. El primero, del Campaniano al Eoceno inferior, agrupa asociaciones del tipo flysch con foraminíferos aglutinados dominantes. En general las asociaciones se caracterizan por reflejar en su composición, y en casos como el de la Formación Punta Torcida por su excelente preservación, ambientes deficitarios en oxígeno asociados con abundante materia carbonosa. Las restringidas asociaciones de foraminíferos calcáreos, contienen muy abundantes foraminíferos bentónicos de relativa alta diversidad. El Campaniano se caracteriza por asociaciones de foraminíferos aglutinados de muy baja diversidad, con el género Rzehakina. El Maastrichtiano-Daniano, con faunas de aglutinados de mediana diversidad. El Paleoceno, con faunas de aglutinados de muy baja diversidad dominadas por Spiroplectammina spectabilis, y ocasionalmente contrastantes restringidos niveles con faunas dominadas absolutamente por foraminíferos calcáreos como ocurre en las Formaciones La Barca (Malumián y Caramés 2002) y Punta Noguera.
El Eoceno inferior, comprende faunas de ambientes someros con abundantes elementos endémicos como Antarcticella, y exhibe un recambio dado por ya bien establecidos representantes de la Familia Elphidiidae, en sus dos formas: planoespirales y trocoespirales. Las últimas dan comienzo a un dominio del género Cribro-rotalia en los ambientes someros de Tierra del Fuego y luego reconocible en toda la Patagonia.
El segundo conjunto se debe a un cambio faunístico mayor debido por una parte a la desaparición de ambientes disaeróbicos y típicamente turbidíticos, y por la transgresión representada en Tierra del Fuego por el Grupo La Despedida. El desarrollo del Eoceno medio superior, se realiza en ambientes someros dominantes, con escasos representantes planctónicos, y entre los bentónicos se destacan elementos típicamente antárticos, como Ammoelphidiella.
El tercer conjunto, corresponde al Grupo Cabo Domingo. A partir del Eoceno cuspidal-Oligoceno basal, la súbita profundización, e ingreso de aguas antárticas, propicia ambientes profundos bien oxigenados, con una LCC alta. Si bien los ambientes profundos son adecuados para los foraminíferos planctónicos, principales elementos cronológicos, las extendidas condiciones de ambientes corrosivos se manifiestan por microfaunas residuales conformadas por foraminíferos aglutinados. Estas condiciones de preservación atentan al seguimiento y distribución de los foraminíferos planctónicos, principalmente de sus apariciones y desapariciones, previniendo la postulación de una zonación.
Debido a la alta latitud media de la Isla, y al conocido desmejoramiento climático, marcado a partir del Eoceno medio superior, las especies planctónicas guías típicamente termofílicas están ausentes. Las microfaunas desde el Eoceno superior hasta el Mioceno inferior, se caracterizan por sus foraminíferos planctónicos de altas latitudes, y las zonaciones tropicales de Berggren et al. (1995) no pueden reconocerse directamente a causa de la ausencia de sus marcadores zonales. Ciertas similitudes se denotan con las zonaciones australes de medias latitudes como la propuesta por Jenkins (1985) o esquemas planteados para el sur de Australia (cf. Li et al. 2003). Pero las mayores semejanzas están dadas en las zonaciones que son de latitudes similares a la de Tierra del Fuego como la de Kerguelen (cf. Huber 1991, Berggren 1992) originadas en estudios del ODP (Fig. 7) aunque éstas contienen una mayor diversidad y no han sido afectadas por disolución. Por lo que las zonaciones que resultan ser identificables en la bioestratigrafía de la Isla, son directamente aquellas de las de regiones antárticas (cf. Stott y Kennett 1990).
Figura 7: Posición relativa del área de estudio respecto a los sitios del DSDP y ODP, la Antártida y masas de agua. 
El Eoceno superior, está muy caracterizado por los pretenuitellidos. Particularmente, P. insolita fue descrita para Nueva Zelanda, donde se extiende entre justo por debajo de las últimas apariciones de Acarinina collactea y de Globigerinatheka spp. Idéntico rango se reconoce en el sur de Australia y en Kerguelen, desde aproximadamente el límite entre las zonas P14/P15 hasta los registros más altos de Globigerinatheka (cf. Huber 1991). El corto rango de P. insolita y su abundancia en el Hemisferio Sur dio lugar a la zona homónima. Tiene una tendencia a asociarse con Chiloguembelina cubensis y Pseudohastigerina micra, en una relación inversa de abundancia con Globigerinatheka. Esta relación, sugiere una fluctuación en los nutrientes, siendo Globigerinatheka más oligotrófica. Tal como se ha visto en la Formación Blanche Point asociado con alto contenido de carbono y abundante sílice opalina, y sus faunas de foraminíferos bentónicos ricas en uvigerinidos y bolivinidos (Li et al. 1995). En Tierra del Fuego, se encuentra en similar asociación con Bolivina y abundantes radiolarios, que sumado al reconocimiento de grandes nódulos fosfáticos en el miembro CCc, sugieren condiciones de surgencia.
En suma, en el hemisferio sur los pretenuitellidos se encuentran restringidos a las zonas P15 y P16, revelan la perturbación del acontecimiento terminal del Eoceno dada por un brusco enfriamiento. Asociado se ha encontrado Isthmolithus recurvus, un elemento de la nanoflora sumamente conspicuo cuya aparición y desaparición están calibradas confiablemente, pero la última, debido a que el biocrón de esta especie comprende en Tierra del Fuego a una superficie de discordancia, su dato de desaparición debe tomarse con las prevenciones del caso. El límite Eoceno/Oligoceno, que está definido por la desaparición de las típicamente tropicales hantkeninas; de modo que en altas latitudes se toma como un substituto aproximado la desaparición de G. index. Esta desaparición no es muy aplicable en Tierra del Fuego, pues a diferencia de las regiones australes de medias latitudes como en Nueva Zelanda, G. index se convierte en una especie rara al término de su biocrón, y hasta ausente cuando el grupo de las pretenuitellas es abundante, en condiciones inestables dadas por surgencia. Una sección que contiene acotado al límite Eoceno/ Oligoceno, se encuentra entre los niveles de las muestras PG1, con la última aparición de G. index, y las PG2 dentro del registro de I. recurvus.
El Oligoceno basal, es bien manifiesto por una asociación que comprende la abrupta aparición y abundancia de C. dissimilis, grandes y abundantes ejemplares de S. angiporoides junto con Chiloguembelina cubensis y "Globigerina" brevis. La última especie en Australasia conforma una zona de rango total que comprende el límite Eoceno/Oligoceno, y que mayormente se desarrolla en el Oligoceno basal siendo una aproximación del reconocimiento de la zona P18 (cf. Li et al. 2003). Esta asociación, se encuentra junto con el DUA de I. recurvus por lo que no es más joven que 32 Ma, si es que el nanoplancton no es retrabajado. Esta asociación claramente oligocena basal, contiene típicos ejemplares de Zeaglobigerina woodi, siendo este el registro más antiguo de la especie, y además de Ch. cubensis, diferenciándose así de los niveles eocenos con Ch. ototara. Diferencias en la distribución de estas dos últimas especies, y el reconocimiento como especies independientes, ya fue bien establecido por Hornibrook (1990).
El Oligoceno superior, se denotaría por la aparición de Zeaglobigerina labiacrassata, aparición que puede ser efecto de la disolución ya que su biocrón reconoce una aparición más antigua (cf. Spezaferri 1994).
A partir del Oligoceno superior hasta el Mioceno inferior, se encuentran abundantes "Globigerinoides" y Praeglobigerinoides sensu Spezaferri (1994) o sea formas con la morfología de Globigerinoides pero carentes de aberturas suplementarias.
Si bien C. dissimilis ha sido considerada una especie de aguas frías, es escasa y poco frecuente en el Oligoceno superior y Mioceno inferior fueguino. Las asociaciones se caracterizan por las zeaglobigerinas en el Oligoceno cuspidal-Mioceno inferior bajo, en la Formación Desdémona, y luego por el dominio de las globigerinas espinosas a partir de la sección superior de las capas de Cabo Ladrillero, que aumentan su abundancia asociada a marcadas condiciones de disolución y aguas frías. Un abrupto retiro del mar, sin que se reconozcan facies regresivas someras, se produce antes de 21,6 Ma, dado que no alcanzan a registrarse globorotalias, y algunas especies muy comunes en altas latitudes como G. incognita (Walters) cuya primera aparición está calibrada en esa edad.
El límite Oligoceno/Mioceno, que se corresponde con las zonas P22 y N4, es indeterminable en altas latitudes según los foraminíferos planctónicos, de acuerdo con la opinión de Spezaferri (1994). Opinión que se comparte ya que faltan las formas diagnósticas demarcatorias del límite, todas ellas termofílicas. Una estimación de la posición surge de considerar la primera aparición de Zeaglobigerina brazieri en el Chattiano tardío, que acotaría la máxima edad la Formación Desdémona. Estimándose el Mioceno inferior por el aumento de la abundancia de Z. connecta. La sección inferior de las capas del Cabo Ladrillero sería ya Mioceno inferior por sugerencia de los foraminíferos bentónicos. Por lo que el límite podría encontrarse tanto entre las capas de la Herminita y la Formación Desdémona, y probablemente entre esta Formación y la sección inferior de las capas del Cabo Ladrillero.
Por su edad, resulta la Formación Desdémona el equivalente temporal del "Patagoniano", producto de un momento de nivel de mar alto; pero con contrastantes microfaunas. La primera con típicas especies cosmopolitas de profundidad, y el segundo con especies sumamente someras y típicos géneros endémicos del Hemisferio Sur.
Recientemente, se listaron los foraminíferos planctónicos de la Formación Carmen Silva, y que están dominados por tenuitéllidos, por ser la formación de ambiente somero, solo llegan las formas pequeñas. Los tenuitéllidos son abundantes en altas latitudes y hasta el Mioceno medio inferior. La ausencia por razones latitudinales, del grupo de Orbulina, resta un importante y definitorio componente cronológico del Mioceno medio, incluso en los ambientes profundos de perforaciones situados costa afuera.
Foraminíferos bentónicos: En los foraminíferos bentónicos, la amplia distribución de condiciones por debajo de la LCC, resalta la importancia en su aplicación bioestratigráfica de algunos foraminíferos calcáreos resistentes a la disolución y de los foraminíferos aglutinados.
En especial, el análisis de las primeras apariciones, tema descuidado en los estudios de subsuelo mediante muestras de inyección, es de suma utilidad en superficie. Entre los primeros, la especie cosmopolita Sphaeroidina bulloides en los fondos oceánicos se conoce desde con dudas desde la zona P18, y con seguridad desde la P19, Oligoceno medio (Morkhoven et al. 1986). En Tierra del Fuego, es una de las especies más distribuidas, se reconoce sólo a partir de la Formación Desdémona, o sea tan temprano como Oligoceno cuspidal.
Entre los foraminíferos aglutinados, cabe destacar a S. compressa por su distribución persistente, si bien de aparición mundial en el Eoceno medio superior, el primer registro en Tierra del Fuego se encuentra entre las últimas apariciones de I. recurvus y S. angiporoides. Natland et al. (1974, rectificado en comunicación epistolar del 21/4/75) notaron que en subsuelo tiene dos niveles de abundancia: uno en el techo del Rosariano, y otro en la parte superior del Miradoriano.
Los ambientes por debajo de la LCC: En un intento de interpretar el significado de los ambientes desarrollados en la lisoclina y por debajo de la LCC, se debe considerar que en términos generales dependen de la productividad de las aguas superficiales y la fisiografía de fondo. Así también, se reconocen momentos de niveles bajos de LCC. En particular, el límite Eoceno/Oligoceno es un momento en que la LCC es alta en altas latitudes (cf. Berggren et al. 1995), cuando microfaunas de aglutinados reemplazan las microfaunas calcáreas.
Por otra parte, se pueden comparar las microfaunas residuales reconocidas en el Oligoceno de Tierra del Fuego con las que tienen potencial de fosilización según el criterio de Mackensen et al. (1990). Contándose con el excelente estudio de Harloff y Mackensen (1997) de áreas actuales próximas de la cuenca Argentina.
Estos autores determinaron la relación entre las masas de agua, las asociaciones de foraminíferos vinculadas, y las asociaciones residuales resultantes en las áreas por debajo de la LCC. Solo una, de las tres asociaciones de foraminíferos aglutinados reconocidas con potencial de fosilización, la asociación de Eggerella bradyi-Martinottiella communis, pobre en especies y ejemplares, queda representada en forma de caparazones de ejemplares muertos en condiciones por debajo de la LCC. Esta asociación se encuentra por debajo del agua antártica de fondo y del agua circumpolar profunda inferior, y a más de 1.000 metros de profundidad. En las partes más australes del mar de Weddell en el talud superior, la asociación fósil Martinottiella nodulosa y Miliammina arenacea domina en los 2.000 metros de profundidad. La asociación fósil de Martinottiella nodulosa indica condiciones de talud continental inferior, con un ambiente hemipelágico calmo, en el cual fluye sin inconvenientes la sedimentación fina y la materia orgánica particulada (Mackensen et al. 1990).
Esta última asociación es estrechamente comparable con la asociación de S. compressa- Martinottiella sp., dado el común contenido de Martinottiella, y la homeomorfía entre el género supérstite Miliammina y el extinguido Spirosigmoilinella, que comparten una disposición cameral tipo miliólido y resistencia a la disolución con ácido clorhídrico. Esta comparación sugiere entonces grandes profundidades, sin embargo, el reconocimiento de la asociación de Spirosigmoilinella-Martinottiella hasta la cuenca del Colorado, con registros sobre el nesocratón del Deseado, en ambientes que por razones regionales deben considerarse de plataforma, ha llevado a proponer alguna explicación de carácter fisiográfico del fondo marino que promoviese el remontamiento de agua de características antárticas, corrosivas y densas, sobre la plataforma, como ser una estrecha comunicación entre los océanos Pacífico y Atlántico (cf. Malumián y Náñez 1991).
Un primer episodio de condiciones de fondos por debajo de la LCC iniciadas sincrónicamente con el Oi-1 en el Oligoceno basal y extendidas por lo menos en el inferior (Capas de María Cristina y Herminita), sugieren condiciones propicias para el ingreso de aguas antárticas posiblemente dadas más por condiciones fisiográficas e hidrodinámicas que por grandes profundidades.
Por lo que una entrada pacífica a una conexión pacífica-atlántica debido a la profundización de la cuenca en tiempos oligocenos favorecería el ingreso de aguas corrosivas, dada la circulación de oeste a este de la actual corriente circumantártica. Alternativamente, tomando como ejemplo la actual fosa de Malvinas, donde el flujo hacia el norte de la masa de aguas de fondo originadas en el sur (SOBW, Barker 2001, Barker y Thomas 2004) es derivado hacia el oeste, por el efecto de Coriolis, y es posible que una deriva similar haya ocurrido hacia la antefosa fueguina en el Oligoceno, inundándola con aguas polares corrosivas.
Un segundo episodio de disolución se produce en el Mioceno inferior junto con un cambio hacia asociaciones planctónicas de aguas frías, que comprende la sección superior de las capas de Cabo Ladrillero hasta las arcillas de Cabo Inés.
Evolución geológica: Discordancias, calibración, y significado
La gran transgresión del Eoceno medio superior, se asienta sobre una discordancia de gran amplitud, con un hiato mínimo de 6 Ma (entre 50 y 44 Ma), que sería el hiato de mayor duración dentro del Cenozoico fueguino (Fig. 6) y que separa condiciones de sedimentación contrastantes; disaeróbicas y turbidíticas por debajo, y con fondos ventilados por encima.
En la Isla, la transgresión está representada por el glauconítico B y la Formación Leticia (Olivero y Malumián 1999), la última de 500 m de areniscas glauconíticas depositadas en no más de 3 Ma en condiciones someras, y aproximadamente entre dos caídas de temperatura de las zonas polares, coherente con el registro de pingüinos fósiles (cf. Clarke et al. 2003). En el inicio del correspondiente período, se produce un episodio calcáreo con la Formación Río Bueno depositada en condiciones extratropicales y que representa las calizas más australes de América del Sur (Malumián y Olivero 1998).
La transgresión se corresponde esencialmente con el pico de mar alto de 41,2 Ma, pero como el hiato en su base abarca los niveles de mar muy alto, entre ca. 50 y 43 Ma, la discordancia debe tener un fuerte control tectónico, se corresponde cronológicamente con la colisión de la dorsal Farallón-Aluk y tiene aún mayor amplitud dentro de la Patagonia. Inclusive se manifiesta por estratos de crecimiento y discordancias progresivas (Olivero y Malumián 1999, Ghiglione et al. 2002.) Además, esta discordancia separa composiciones petrográficas contrastantes, marcadamente volcaniclástica en las sedimentitas del Paleoceno- Eoceno inferior, y cuarzo-lítica en el Eoceno medio superior y superior donde se reconocen componentes derivados de rocas andinas jurásicas, cretácicas y paleógenas (Olivero 2002).
La discordancia entre las Formaciones Leticia y Cerro Colorado, expresada en la sección del río Menéndez (Fig. 2, perfil 8) por los términos continentales de la Formación Leticia y por un marcado hiato costa afuera calibrado en 40,3 Ma (T20 en la Fig. 6), abarca un hiato mínimo de 1,5 Ma. Esta discordancia coincide con una pronunciada caída del nivel del mar, que también se refleja por su exposición subaérea y en los depósitos continentales y marinos marginales, según la posición geográfica, del techo de la Formación Leticia (Fig. 2).
La Formación Cerro Colorado, ca. 900 m de sedimentitas (Olivero y Malumián 1999) depositadas en el depocentro de la cuenca en no más de 4 Ma, representa una de las mayores tasas de sedimentación. Sus tres miembros inferiores componen el mayor espesor (ca. 700 m) y si bien sus bases coinciden con tres pulsos eustáticos menores, su depositación transcurre dentro de un intervalo de nivel de mar relativamente bajo. La alta tasa de sedimentación, el aporte de importantes espesores de arena a la cuenca, que conforman típicos lóbulos turbidíticos, sugieren un marcado control tectónico en la generación de espacio de sedimentación. El miembro CCd, que registra las mayores profundidades de depositación de la Formación, se origina durante el pronunciado ascenso del nivel del mar en el Eoceno tardío. La gran caída de temperatura, próxima a los 36 Ma, se refleja por la microfauna típica de condiciones inestables y ricas en nutrientes junto con la aparición de Antarcticella antarctica e I. recurvus, típicos representantes de carácter y origen antártico. El más bajo contenido de esmectita reconocido del cenozoico fueguino, en el miembro CCd y en el glauconítico A, revelan condiciones de tectonismo activo en un clima frío. Se inicia con ellos el nivel de mar alto que, conjuntamente con la antefosa originada por carga tectónica, está representado por sedimentos marinos profundos. Los 300 m de las principalmente limoarcilitas de las capas de la María Cristina y de la Herminita, se depositaron en 5 Ma y constituyen el pico de inundación de la columna sedimentaria terciaria que cambió el escenario deposicional de la cuenca de un ambiente nerítico a otro con profundidades batiales. La inundación es un acontecimiento global que se habría visto amplificado por la profundización tectónica de la cuenca (cf. Galeazzi, 1998). La profundización comienza en el miembro CCd y se establecen condiciones batiales a partir del Oligoceno basal. La amplia distribución de condiciones por debajo de la LCC sugiere más que condiciones abisales, el ingreso de aguas antárticas corrosivas debido a condiciones hidrológicas que permitan incursionar esta agua densa dentro de la cuenca.
En general, los ambientes depositacionales del conjunto eoceno-oligoceno tienen un fuerte control tectónico, relacionado con la propagación de la deformación hacia el norte en la faja plegada y corrida y con la subsidencia por carga tectónica. En función de este control, el relleno sedimentario define una geometría de cuñas asimétricas con mayor espesor hacia la antefosa, y que se adelgazan hacia el norte sobre la rampa del antepaís. Esta geometría se expresa tanto en subsuelo mediante las líneas sísmicas (cf. Robbiano et al. 1996, Galeazzi 1998) como en criterios regionales de superficie (Olivero y Malumián 2002). Un resultado llamativo del presente trabajo, es que a lo largo de la antefosa, i.e. en dirección general ONO-ESE, el relleno sedimentario tiene también una geometría de cuñas sedimentarias, con mayor espesor hacia el ESE (Fig. 2). Estas cuñas reflejan la colmatación progresiva de la antefosa y consecuente extensión de la plataforma hacia el ESE.
El contrastante mayor espesor del Grupo Cabo Domingo, en especial para la Formación Cerro Colorado, hacia el sector SE de la antefosa atenúa la expresión de la discordancia basal. Por el contrario, en el sector NO donde alcanza menor espesor, esta discordancia tiene una pronunciada expresión y el conglomerado Tchat-Chii apoya discordantemente, con apreciable hiato, sobre sedimentitas equivalentes a los miembros CCa y CCb de la Formación Cerro Colorado.
En el sector SE de la antefosa, el conjunto conformado por las capas de la Estancia María Cristina-Puesto Herminita-Formación Desdémona se deposita durante las últimas etapas de la deformación compresiva. El suave plegamiento que afecta a la última es simultáneo con la actividad del corrimiento Punta Gruesa (Ghiglione 2002, 2003), consecuentemente este corrimiento, que limita la faja plegada y corrida fueguina, estuvo activo seguramente hasta el Oligoceno tardío y posiblemente hasta el Mioceno inicial.
El contacto entre las capas de la Estancia María Cristina y del Puesto Herminita, comprende facies y paleoambientes diferentes en los perfiles del Cerro Colorado y del límite de la faja plegada y corrida. En el primero (Perfil 1, Fig. 2) y dentro de una sucesión litológicamente homogénea depositada en el techo de corrimientos activos (cf. Olivero y Malumián 2002, Malumián y Olivero 2005), el contacto está indicado por pronunciados cambios en las microfaunas y de condiciones de depositación. En las capas de la Estancia María Cristina, las condiciones de depositación por debajo de la LCC son seguidas por las de plataforma media, y las del Puesto Herminita comienzan con condiciones de plataforma externa (Fig. 5). Estos cambios son coherentes con la brusca caída y recuperación del nivel del mar hacia los 30 Ma (Fig. 6). En el segundo perfil (Perfil 2, Fig. 2), las microfaunas indican condiciones de depositación homogéneas por debajo de la LCC, y el límite entre estas capas está marcado por la abrupta aparición de bancos potentes de areniscas en los ambientes marinos profundos de la antefosa, que reflejarían la pronunciada caída del nivel del mar de los 30 Ma.
Existiría una relación discordante entre las capas del Puesto Herminita y la Formación Desdémona con un pequeño hiato, vinculable en parte a los bajos niveles de mar del Oligoceno superior. Los 300 m de principalmente limoarcilitas de la Formación Desdémona se depositaron en 3 Ma.
El aumento del nivel del mar iniciado en el Oligoceno cuspidal, que en la Patagonia está representado por sedimentos someros del Patagoniano, se refleja en la Formación Desdémona, que correspondería a facies de talud depositacional, con frecuente removilización de sedimentos por deslizamientos gravitatorios e inyección de numerosos diques clásticos arenosos, probablemente relacionados con actividad tectónica. Los foraminíferos contenidos son indicativos de aguas profundas y signos de corrosión se manifiestan en casi todas sus muestras, consecuentemente con altos porcentajes y dominio de foraminíferos aglutinados. Aparentemente han dominado mayormente condiciones de lisoclina o condiciones diagenéticas que llevaron a la disolución de las conchillas calcáreas. Asociaciones residuales muy similares se reconocen en la actualidad en ambientes profundos en la cuenca Argentina.
La suave discordancia angular que separa a la Formación Desdémona de las capas del Cabo Ladrillero, refleja la finalización de la deformación compresiva, la abundante inyección de diques clásticos en estas últimas reflejaría transcurrencia activa (cf. Ghiglione 2002, 2003). La pronunciada discordancia en el techo de las capas del Cabo San Pablo se correspondería con el fulminante retiro del mar en 21,8 Ma, y con una discordancia regional (T30 de 21 Ma, Fig. 6). Estas últimas capas reflejan en su parte superior ambientes marinos profundos, por debajo de la LCC. Por un tiempo no hay sedimentación marina, la que se reconoce nuevamente en el sector NO de la cuenca, con el óptimo climático y nivel de mar alto del Mioceno medio representado por los sedimentos deltaicos de la Formación Carmen Silva. Formación que representa una transgresión bien documentada en el área antártica (cf. Bartek et al. 1991). La asociación de arcillas, con dominio de esmectita, sería coherente con el óptimo climático del Neógeno (Cuadro 1). En mares de la plataforma continental argentina aledaños a la Isla, si bien se reconocen elementos del nanoplancton calcáreo indicadores de aguas cálidas como Sphenolithus, la temperatura de las aguas no alcanzó la necesaria para las orbulinas, los foraminíferos característicos del Mioceno medio. El retiro del mar del Mioceno medio, coincide con la discordancia de 13,8 Ma, ampliamente reconocida costa afuera (cf. Galeazzi 1998).
Luego del cese de la deformación compresiva y de la consecuente subsidencia por carga tectónica, las sedimentitas marinas marginales y continentales de las Formaciones Carmen Silva y Castillo, representan la colmatación en el sector NO de la antefosa. La Formación Carmen Silva, constituiría además el primer establecimiento de una plataforma de extensión moderada, con abundantes fósiles de invertebrados, hacia el este fueguino. Hacia el sector SE de la cuenca, se mantienen las condiciones marinas profundas, representadas por las capas del Cabo Viamonte. Los clastos de volcanitas basálticas frescas de las sedimentitas del Mioceno medio atestiguan un episodio efusivo todavía no reconocido en Tierra del Fuego.
AGRADECIMIENTOS
Los análisis de arcillas fueron realizados por G. Cozzi (INTEMIN) e interpretados por Susana Alonso (CONICET). Las preparaciones de nanoplancton, por G. Jannou (SEGEMAR). Las determinaciones de los microfósiles de la Formación Desdémona, en colaboración de Romina Scarpa. Gran parte de muestreos, con la colaboración de Carolina Náñez. Financiado en parte por PICT 12257 y PICT 8675 Agencia Nacional de Promoción Científica y Tecnológica-FONCYT y por el PIP 2756 CONICET.
TRABAJOS CITADOS EN EL TEXTO
Alonso, M.S., Cozzi, G. y Malumián, N. 2001. Asociaciones de arcillas de la Formación Barranca Final, Mioceno, Cuenca del Colorado. Argentina. 2º Congreso Latinoamericano de Geología, Actas (CD Rom: Trabajo 176) Montevideo. [ Links ]
Barker, P.F. 2001. Scotia Sea regional tectonic evolution: implications for mantle flor and palaeocirculation. Earth-Science Reviews 55: 1-39. [ Links ]
Barker, P. F. y Thomas, E. 2004. Origin, signature and palaeoclimatic influence of the Antarctic Circumpolar Current. Earth-Science Reviews 66: 143-162. [ Links ]
Bartek, L.R., Vail, P.R.; Anderson, J.B.; Emmet, P.A. y Wu, S. 1991. Effect of Cenozoic Ice Sheet Fluctuation in Antarctica on the Stratigraphic Signature of the Neogene. Journal of Geophysical Research 96(B4): 6753-6778. [ Links ]
Berggren, W.A. 1992. Paleogene planktonic foraminifer magnetobiostratigraphy of the Southern Kerguelen Plateau (Sites 747-749). En Wise, S.W., Jr., Schlich, R., et al., Proc. ODP, Sci. Results, 120 (Pt. 2): College Station, TX (Ocean Drilling Program): 551- 568. [ Links ]
Berggren, W.A., Kent, D.V., Swisher, C.C., III, y Aubry, M-P. 1995. A revised Cenozoic geochronology and chronostratigraphy. En Berggren, W.A., Kent, D.V., Aubry, M.-P., y Hardenbol, J. (eds.), Geochronology, Time Scales and Global Stratigraphic Correlation. Special Publication-SEPM (Soc. Sediment. Geol.) 54:129-212. [ Links ]
Biddle, K.T., Uliana, M.A., Mitchum, R.M. Jr., Fitzgerald, M. y Wright, R.C. 1986. The stratigraphic and structural evolution of the central and eastern Magallanes Basin, southern South America. En Allen, P.A. y Homewood, P. (eds.) Foreland Basins. International Association of Sedimentologists, Special Publication 8: 41-61. [ Links ]
Boersma, A. 1984. Handbook of common Tertiary Uvigerina. Microclimates Press, 207 p. Stony Point, New York. [ Links ]
Buatois, L.A. y Camacho, H.H. 1993. Geología del sector nororiental del lago Fagnano, Isla Grande de Tierra del Fuego. Revista de la Asociación Geológica Argentina 48(2): 109-124. [ Links ]
Calegari, R. y Baldi, J. 1993. Análisis secuencial para el Terciario inferior de cuenca Austral mediante estudios sismoestratigráficos. 12º Congreso Geológico Argentino y 2º Congreso de Exploración de Hidrocarburos, Actas 1: 87-95. [ Links ]
Chapronier, G.C.H. 1992. The distribution and development of Late Oligocene and Early Miocene reticulate globigerines in Australia. Marine Micropaleontology 18: 279-305. [ Links ]
Clarke, J., Olivero, E.B. y Puerta P. 2003. Report of the earliest fossil penguin from South America and first fossil vertebrate from the Paleogene of Tierra del Fuego, Argentina. American Museum Novitates 3423: 1-18. [ Links ]
Codignotto, J.O., Malumián, N. 1981. Geología de la región al Norte del paralelo 54º S. de la Isla Grande de Tierra del Fuego. Revista de la Asociación Geológica Argentina 36 (1): 44-88. [ Links ]
De Ferrariis, C., en Fossa Mancini et al. 1938. Una reunión de geólogos de YPF y el problema de la terminología estratigráfica. Boletín de Informaciones Petroleras 171: 43-44: 95-96. [ Links ]
Feagle, J.G., Bown, T.M., Swisher, C. y Buckley, G. 1995. Age of the Pinturas and Santa Cruz Formations. 6° Congreso Argentino de Paleontología y Bioestratigrafía, Actas: 129-135, Trelew. [ Links ]
Galeazzi, J.S. 1998. Structural and stratigraphic evolution of the Western Malvinas basin, Argentina. American Association of Petroleum Geologists, Bulletin 82 (4): 596-636. [ Links ]
Ghiglione, M. 2002. Diques clásticos asociados a deformación transcurrente en depósitos sinorogénicos del Mioceno inferior de la Cuenca Austral. Revista de la Asociación Geológica Argentina 57(2): 103-118. [ Links ]
Ghiglione, M. 2003. Estructura y Evolución Tectónica del Cretácico-Terciario de la costa atlántica de Tierra del Fuego. Tesis Doctoral, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, (inédito) 225 p., Buenos Aires. [ Links ]
Ghiglione, M., Ramos, V.A. y Cristalini, E.O., 2002. Estructura y estratos de crecimiento en la faja plegada y corrida de los Andes Fueguinos. Revista Geológica de Chile 29(1): 17-41. [ Links ]
Guerstein, G.R., Chiesa, J.O., Guler, M.V. y Camacho, H.H. 2002. Bioestratigrafía basada en quistes de dinoflagelados de la Formación Cabo Peña (Eoceno terminal- Oligoceno temprano), Tierra del Fuego, Argentina. Revista Española de Micropaleontología 354(1): 105-116. [ Links ]
Haq, B.U., Hardenbol, J. y Vail, P.R. 1987. Chronology of fluctuating sea levels since the Triassic. Science 235: 1156-1167. [ Links ]
Harloff, J. y Mackensen, A. 1997. Recent benthic foraminiferal associations and ecology of the Scotia Sea and Argentine Basin. Marine Micropaleontology 31: 1-29. [ Links ]
Hornibrook, N. de B. 1990. Chiloguembelina cubensis (Palmer) and C. ototara (Finlay), in New Zealand. Journal of Foraminiferal Research 20(4): 368-371. [ Links ]
Hornibrook, N. de B., Brazier, R.C. y Strong, C.P. 1989. Manual of New Zealand Permian to Pleistocene foraminiferal Biostratigraphy. New Zealand Geological Survey Paleontological, Bulletin 56: 175 p. [ Links ]
Huber, B.T. 1991. Paleogene and early Neogene planktonic foraminifer biostratigraphy of Sites 738 and 744, Kerguelen Plateau southern Indian Ocean). College Station, Proceedings of the Ocean Drilling Program, Scientific Results 199: 427-449. [ Links ]
Jannou, G. y Olivero, E.B. 2001. Hallazgo de radiolarios del Paleógeno en la Isla Grande de Tierra del Fuego, Argentina. Ameghiniana 38(3): 317-320. [ Links ]
Jenkins, G. 1985. Southern mid latitude Paleocene to Holocene planktic foraminifera. En Bolli H.M., Saunders, S.B. y Perch-Nielsen, K. (eds.) Plankton Stratigraphy. Cambridge University Press: 263-282. [ Links ]
Klepeis, K.A. y Austin, J.A. Jr. 1997. Contrasting styles of superposed deformation in the southernmost Andes. Tectonics 16: 755-776. [ Links ]
Leckie, R. M. y Webb, P.T. 1990. Late Paleogene and Early Neogene Foraminifers of Deep Sea Drilling Project Site 270, Ross Sea, Antarctica. Initial Reports DSDP 90: 1093-1142. [ Links ]
Li, Q. y McGowran, B. 1994. Miocene upwelling events: foraminiferal evidence from southern Australia. Australian Journal Earth Sciences 41: 593-603. [ Links ]
Li, Q., McGowran, B. y Brunner Ch. 2003. Eocene-Oligocene Planktonic Foraminiferal Biostratigraphy of Sites 1126, 1130, 1132, and 1134, ODP Leg 182. Great Australian Bight. Proceedings of the Ocean Drilling Program 182. pag. web. [ Links ]
Li, Q.; McGowran, B. y Boersma, A. 1995. Early Palaeocene Parvularugoglobigerina and late Eocene Praetenuitella: does evolutionary convergence imply similar habitat?. Journal of Micropalaeontology 14: 119-134. [ Links ]
Li, Q., Radford, S.S. y Banner, F.T. 1992. Distribution of microperforate tenuitellid planktonic foraminifers in Holes 747A and 749B, Kerguelen Plateau. En Wise, S.W. Jr., Schilch, R., et al., Proc. ODP, Sci. Results, 120: College Station, TX (Ocean Drilling Program) 569-594. [ Links ]
Mackensen, A., Grobe, H., Kuhn, G. y Fütterer, D.K. 1990. Benthic foraminiferal assemblages from the eastern Weddell Sea between 68 and 73°S: Distribution, ecology and fossilization potential. Marine Micropaleontology 16: 241-283. [ Links ]
Malumián, N. 1982. Foraminíferos bentónicos de la Formación Carmen Silva, Mioceno, Isla Grande de Tierra del Fuego. Ameghiniana 19(1-2): 37-66. [ Links ]
Malumián, N. 1988. Foraminíferos bentónicos de la localidad tipo de la Formación La Despedida (Eoceno, Isla Grande de Tierra del Fuego). Parte I. Textulariina y Miliolina: Ameghiniana 25 (4): 341-356. [ Links ]
Malumián, N. 1999. La sedimentación en la Patagonia extrandina. En: Geología Argentina, R. Caminos (ed.). Servicio Geológico Minero Argentino, Anales 29(18): 557-612. [ Links ]
Malumián, N., Camacho, H.H. y Gorroño, R. 1978. Moluscos del Terciario inferior ("Magallanense" de la Isla Grande de Tierra del Fuego (República Argentina). Ameghiniana 15 (3-4): 265-284. [ Links ]
Malumián, N. y Caramés, A. 1989. Foraminíferos uniloculares de ornamentación no reticulada (Eoceno-Oligoceno), Tierra del Fuego, Argentina. Ameghiniana 26 (3-4): 103-137. [ Links ]
Malumián, N. y Caramés, A. 2002. Foraminíferos de sedimentitas ricas en carbono orgánico: Formación La Barca, Paleoceno superior, Tierra del Fuego, República Argentina. Revista de la Asociación Geológica Argentina 57(3): 219-231 [ Links ]
Malumián, N., Caramés, A. y Martínez, H. 1998. Asociaciones mineralógicas de arcillas del Paleogeno de cuenca Austral, su significado paleoclimático y el límite Paleoceno/Eoceno. Asociación Paleontológica Argentina. Publicación Especial 5, Paleógeno de América del Sur y de la Península Antártica: 85-94. [ Links ]
Malumián, N. y Náñez, C. 1988. Asociaciones de foraminíferos del Terciario medio de Cuenca Austral: sus relaciones con eventos eustáticos globales. Revista de la Asociación Geológica Argentina 43(2): 257-264. [ Links ]
Malumián, N. y Náñez, C. 1991. Paleogeografía del Terciario medio del cono sur: avance de aguas antárticas. 6° Congreso Geológico Chileno, Resúmenes Expandidos: 847-851. [ Links ]
Malumián, N. y Olivero, E.B. 1998. La Formación R ío Bueno, relaciones estratigráficas y edad, Eoceno inferior, Isla Grande de Tierra del Fuego, Argentina. 10º Congreso Latinoamericano de Geología y 6º Congreso Nacional de Geología Económica, Actas 1: 120-124. [ Links ]
Malumián, N. y Olivero, E.B. 2005. El Oligoceno-Plioceno marino del río Irigoyen, costa atlántica de Tierra del Fuego, Argentina: Una conexión atlánticopacífica. Revista Geológica de Chile 32(1): 1117-129. [ Links ]
Malumián, N., Palamarczuk, S., Alonso, M.S., Tassone, A.A., Barreda, V. y Náñez, C. 1999. Micropaleontología, palinología y sedimentología del Eoceno-Mioceno del pozo Aires e-2, Plataforma continental de Tierra del Fuego.14º Congreso Geológico Argentino, Actas 1(Salta 1999): 369-372. [ Links ]
Malumián, N. y Scarpa, R. 2005. Foraminíferos de la Formación Río Irigoyen, Neogeno, Tierra del Fuego, Argentina: su significado paleobiogeográfico. Ameghiniana 42(2): 363-376. [ Links ]
Manson, G. 1961. Despedida área report, n°152. Tennesse Argentina S.A. Inédito. [ Links ]
Masiuk, V., Riggi, J.C. y Bianchi, J.L. 1990. Análisis geológico del Terciario del subsuelo de Tierra del Fuego. Boletín de Informaciones Petroleras 21, parte I: 70- 89, 22, parte II: 8-25. [ Links ]
McGowran, B. y Li, Q. 1993. Miocene planktonic foraminifera from Lakes Entrance in Gippsland. Midlatitude neritic signal from a transforming ocean. Memoir of the Association of Australasian Palaeontologists, 15: 395-405. [ Links ]
Morkhoven, van, F.P.C.M., Berggren, W.A. y Edwards, A.S. 1986. Cenozoic Cosmopolitan Deep-Water Benthic Foraminifera. Bulletin de Centres de Recherches Exploration-Production Elf-Aquitaine, Mémoire 11, 421 p. [ Links ]
Mostajo, E.L. 1991. Nanofósiles calcáreos cenozoicos del pozo "Las Violetas 3". Isla Grande de Tierra del Fuego. Argentina. Ameghiniana 28 (3-4):311-315. [ Links ]
Mulder T. y Alexander, J. 2001. The physical character of subaqueous sedimentary density flows and their deposits. Sedimentology 48: 269-299. [ Links ]
Natland, M.L., González, E.P., Cañón, A. y Ernst, M. 1974. A System of Stages for correlation of Magallanes Basin Sediments. Geological Society of America, Memoir 139:1-125. [ Links ]
Olivero, E.B. 2002. Petrografía sedimentaria de sistemas turbidíticos del Cretácico- Paleogeno, Andes Fueguinos: Procedencia, volcanismo y deformación. 15º Congreso Geológico Argentino, Actas: Artículo 15: 2 p. [ Links ]
Olivero, E.B. y Malumián, N. 1999. Eocene stratigraphy of Southern Tierra del Fuego, Argentina. American Association of Petroleum Geologists, Bulletin 83(2): 295-313. [ Links ]
Olivero, E.B. y Malumián, N. 2002. Upper Cretaceous-Cenozoic clastic wedges from the Austral-Malvinas foreland basins, Tierra del Fuego, Argentina: Eustatic and tectonic controls. 3rd European Meeting on the Paleontology and Stratigraphy of Latin America (EMPSLA), Univ. Paul Sabatier, Addendum: 6-9. Toulouse. [ Links ]
Olivero E.B. y Malumián, N. 2005. Stratigraphy. En: Geology and Geophysics of Isla Grande de Tierra del Fuego. Field-Trip Guide Booklet. Bolletino de Geofisica Teorica ed Applicata, (en prensa) 46. Trieste. [ Links ]
Olivero, E.B., Malumián, N. Palamarczuk, S. y Scasso, R.A. 2002. El Cretácico superior- Paleógeno del área del Río Bueno, costa atlántica de la Isla Grande de Tierra del Fuego. Revista de la Asociación Geológica Argentina 57(3): 199-218. [ Links ]
Olivero, E.B., Malumián, N. y Palamarczuk, S. 2003. Estratigrafía del Cretácico superior- Paleoceno del área de bahía Thetis, Andes Fueguinos, Argentina: acontecimientos tectónicos y paleobiológicos. Revista Geológica de Chile 30 (2): 245-263. [ Links ]
Olivero, E.B., Ponce, J.J., López C., M.I. y Martinioni, D.R. 2004. Phymatoderma granulata from the Oligocene-Miocene of Tierra del Fuego: morphology and ethology. First International Congress on Ichnology, Ichnia 2004, Abstract Book: 63. Trelew. [ Links ]
Ponce, J.J., Olivero, E.B. y Martinioni, D.R. 2004. Phymatoderma-bearing turbidites (Oligocene, Tierra del Fuego): ichnologic implications for discrimination of sustained and episodic gravity flow deposits. First International Congress on Ichnology, Ichnia 2004, Abstract Book: 67-68. Trelew. [ Links ]
Riggi, J.C. 1988. Litoestratigrafía terciaria del subsuelo fueguino. Revista de la Asociación Geológica Argentina 43(1):106-120. [ Links ]
Robbiano, J.A.; Arbe, H. y Gangui, A. 1996. Cuenca Austral marina. En Ramos, V.A. y Turic, M.A. (eds.) Geología y Recursos Naturales de la Plataforma Continental Argentina. 13° Congreso Geológico Argentino y 3° de Exploración de Hidrocarburos (Buenos Aires) Relatorio 17: 323-341. [ Links ]
Spezaferri, S. 1994. Planktonic foraminiferal biostratigraphy and taxonomy of the Oligocene and lower Miocene in the oceanic record. An overview. Palaeontographia Italica 81, 187 p. Pisa. [ Links ]
Stott, L.D. y Kennett, J.P. 1990. Antarctic Paleogene planktonic foraminifer biostratigraphy: ODP Leg 113, Sities 689 and 690. En P.F. Barker, J.P. Kennett, et al., College Station, Texas: Ocean Drilling Program, Proceedings of the Ocean Drilling Program, Scientific Results, 113: 549-569. [ Links ]
Stott, L.D., Kennett, J.P., Shackleton, N. y Corfield, R.M. 1990. The evolution of antarctic surface waters during the Paleogene: inferences form the stable isotopic composition of planktonic foraminifers, ODP LEG 113. En P.F. Barker, J.P. Kennett, et al., College Station, Texas: Ocean Drilling Program, Proceedings of the Ocean Drilling Program, Scientific Results, 113:849-863. [ Links ]
Yrigoyen, M.R. 1962. Evolución de la exploración petrolera en Tierra del Fuego. Petrotecnia 28: 28-38. [ Links ]
Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; y Billups, K. 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292: 686-693. [ Links ]
Zamaloa, M. del C. 2000. Palinoflora y ambiente en el Terciario del nordeste de Tierra del Fuego, Argentina. Revista Museo Argentino Ciencias Naturales, n.s. 2(1): 43- 51. [ Links ]
Zamaloa, M. del C. y Romero, E. 1990. Some spores and pollen from the Cullen Formation (Upper Eocene to middle Oligocene), Tierra del Fuego, Argentina. Palynology 14: 123-133. [ Links ]
Zetter, R, Hofmann C-C., Draxler, I.; Durango de Cabrera, J., Vergel, M. del M. y Vervoorst, F. 1998. A rich Middle Eocene Microflora at Arroyo de los Mineros, near Cañadón Beta, NE Tierra del Fuego Province, Argentina. Abhandlungen der Geologischen Bundesanstalt 56/1: 439-460. [ Links ]
Recibido: 16 de junio, 2005
Aceptado: 1 de mayo, 2006

