










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkEcología austral
versão On-line ISSN 1667-782X
Ecol. austral v.19 n.1 Córdoba jan./abr. 2009
TRABAJOS ORIGINALES
Predation on the slug Deroceras reticulatum (Pulmonata: Stylommatophora) by Scarites anthracinus (Coleoptera: Carabidae)
María C Tulli *, Dora M Carmona, Alicia N López, Pablo L Manetti, Ana M Vincini & Gabriela Cendoya
Fac. de Ciencias Agrarias, Univ. Nac. de Mar del Plata, Lab. de Zoología Agrícola, Balcarce, Buenos Aires.
*Facultad de Ciencias Agrarias, Univ. Nac. de Mar del Plata, Ruta 226, km 73.5, Laboratorio de Zoología Agrícola. (7620) Balcarce, Buenos Aires. CC 276. Tel.: +54-2266-439100 ext. 222. Fax +54-2266-439101. Email: maricel_46@hotmail.com
Recibido: 13 de agosto de 2008;
Fin de arbitraje: 10 de noviembre de 2008;
Revisión recibida: 26 de diciembre de 2008;
Aceptado: 26 de diciembre de 2009
ABSTRACT. The grey field slug Deroceras reticulatum has become a pest in no tillage sunflower (Helianthus annuus L.) and soybean (Glycine max L.) crops in the southeast of Buenos Aires province, Argentina. Ground beetles (Coleoptera: Carabidae) are an important component of the arthropod community in agricultural systems and have the potential to reduce pest populations. We tested the predation capacity of the carabid beetle Scarites anthracinus on Deroceras reticulatum individuals of different body mass (10, 20 and 30 mg) and on their eggs, in a controlled laboratory no-choice assay. We also tested its preference for juvenile slugs and the similarly-sized pillbugs (Armadillidium vulgare), under choice conditions. This beetle preyed upon both eggs and juvenile slugs, with the highest consumption rate on 20 mg body mass individuals. While preferring the pillbug during the first 24 h of the choice test, our results suggest that in field conditions, S. anthracinus might be a natural enemy of D. reticulatum, consuming slug eggs and juveniles of different body mass.
Keywords: Carabid beetles; Grey field slug; Armadillidium vulgare; Food preference.
RESUMEN. Predación de la babosa Deroceras reticulatum (Pulmonata: Stylommathophora) por Scarites anthracinus (Coleoptera: Carabidae): La babosa gris grande Deroceras reticulatum se ha convertido en una plaga importante de los cultivos de girasol (Helianthus annuus L.) y soja (Glycine max L.) en siembra directa del sudeste de la provincia de Buenos Aires, Argentina. Por su parte, los carábidos (Coleoptera: Carabidae), son importantes componentes de la comunidad de artrópodos en los agroecosistemas y tienen el potencial para reducir las poblaciones de invertebrados. Determinamos la predación de Scarites anthracinus sobre huevos y juveniles de diferente masa corporal (10, 20 and 30 mg) de D. reticulatum, en experimentos de no elección en condiciones controladas de laboratorio, y la preferencia alimentaria por juveniles de babosas y bichos bolita Armadillidium vulgare de tamaño similar, en experimentos de elección. S. anthracinus consumió huevos y juveniles de babosas con una tasa de consumo mayor de individuos de 20 mg. A pesar de una mayor preferencia por bichos bolita durante las primeras 24 h del ensayo, los resultados sugieren que S. anthracinus podría actuar como enemigo natural de D. reticulatum en condiciones de campo, consumiendo huevos y juveniles de diferente masa corporal.
Palabras clave: Carabidos; Babosa gris grande; Armadillidium vulgare; Preferencia alimentaria.
INTRODUCTION
No tillage is one of the most important improvements introduced in the argentinean cropping systems in the last decades, as a mean to reduce soil degradation (Aragón 1998; Studdert & Echeverría 2000). Nevertheless, crop residues remain on the soil surface throughout the year retaining moisture and maintaining low soil temperatures which favour survival of slugs (Stylommatophora: Pulmonata) and pillbugs (Crustacea: Isopoda). These organisms have become agricultural pests because of increasing numbers in no tillage systems. These species can kill seeds or seedlings, destroy stems and meristems, and reduce leaf area resulting in plant death or suppressed growth (South 1992; Byers & Calvin 1994; Saluso 2001; Hommay 2002).
Information about pillbug as a pest in field crops is scarce and only mentioned as a cause of damage in horticultural crops (García 1999). However research carried out in Argentina in Manfredi (Trumper & Linares 1999), Paraná (Saluso 2001) and Balcarce (Mastronardi 2006), showed that A. vulgare reduces the rate of emergence of seedlings and causes damage during October-December, when sunflower and soybean crops are being established. In addition, preliminary tests carried out by Trumper & Linares (1999) indicated that soybean crops exhibit a period of great susceptibility to A. vulgare during 12 days after sowing.
Slugs have regularly caused severe damage in a wide range of crops over large geographical areas where they were previously unknown as pests (South 1992; Hommay 2002). Although a large number of different slug species are found in crops in Buenos Aires province (Argentina), the major damage is done by Deroceras reticulatum (Clemente 2007). Environmental conditions in this region favour a generation of new individuals of semelparous D. reticulatum during October-December (Costamagna et al. 1999; Clemente 2007), coinciding with the sowing and emergence of winter and summer crops, this coincides with the sowing and the emergence of winter and summer crops when the greatest damage takes place.
In sustainable agricultural systems, pest control by predatory arthropods is crucial to prevent outbreaks. Polyphagous predatory carabid beetles (Coleoptera: Carabidae) are an important component of the arthropod community in agricultural systems, and have the potential to reduce pest populations (Barney & Pass 1986). In most species, both adult and larvae are natural enemies of a wide range of pests including black cutworms, cabbage maggot, true armyworm, corn rootworms, crickets and slugs (Best & Beegle 1977). Most carabid adults use their well developed mandibles to kill and fragment preys into pieces. Specialist species attacking snails seem to paralyse their prey by biting, thus preventing the mucus production that is the slugs defense mechanism (Lovei & Sunderland 1996). Some carabid beetles are known to be slug predators and capable of causing a substantial reduction in slug populations in the Northern Hemisphere. Pterostichus melanarius (Carabidae: Pterostichini) is an abundant generalist carabid known to prey on slugs in laboratory conditions (Symondson 1997; McKemey et al. 2001; Oberholzer & Frank 2003), microcosms and miniplots (McKemey et al. 2003) and under field conditions (Chapman et al. 1997).
In Argentina, more than 679 carabid beetle species have been identified (Roig Juñent 1998), and 32 species have been reported in no-till wheat crops (Marasas 2001). However, there is no information about the potential of common species to consume slugs. The underlying management principle is to reduce if possible, the risk of slug and pigbull damage by cultural practices and to apply molluscicides if necessary. Encouraging biological control may help to reduce molluscicide use, but the efficiency of carabids as natural enemies needs to be evaluated. Scarites anthracinus Dej. (Carabidae: Scaritini) is a dominant carabid beetle in Balcarce county representing 44% and 71% of total catch in no-till maize and pasture crops, respectively (Turienzo et al. 2001). Moreover, Carmona et al. (2004) observed that during October-December, when summer crops are establishing, the dominant species present in no-tillage systems were S. anthracinus, Calosoma retusum and Pachymorphus striatulus. S. anthracinus is a big shiny black carabid beetle (18 and 25 mm) with large mandibles and an elytra grooved separated from a smooth pronotum by a narrow "waist". This beetle is a spring breeder and shows its highest activity in october-november, decreases towards summer (Marasas 2001). Given that in the field, the population dynamics of S. anthracinus is coincident with different developmental stages of D. reticulatum, the predation on eggs and juveniles of different body mass were investigated under laboratory conditions with and without the presence of pillbug (A. vulgare) as an alternative prey.
MATERIALS AND METHODS
Adult grey field slugs were collected from pastures at the Integrated Unit Balcarce (Universidad Nacional de Mar del Plata e Instituto Nacional de Teconología Agropecuaria). Slugs were kept in individual round glass 94 mm diameter by 64 mm deep containers as they showed a surprisingly large mortality rate when kept in groups. The containers with pelleted rabbit food, were kept in a growth chamber (light/dark: 16/8 h) at 20±2 °C. Food was replaced weekly and slug eggs were removed and placed into new containers. When the eggs hatched, juvenile slugs were individually reared to the required body mass of 10, 15, 20 or 30 mg, respectively.
Beetles and pillbug A. vulgare were also collected in the field using pitfall traps, which were emptied daily. Beetles were trapped a day prior to the start of the experiment and kept individually in 6-cm diameter petri dishes without food or water. Pillbugs were held for several days in trays with leaf litter, so as to acclimatize them before their use in the experiments. Both, beetles and pillbugs, were held under the same temperature and light conditions as the slugs. Predation was evaluated in shallow plastic trays (175 x 222 x 70 mm) filled with damp potting soil. To prevent slugs from climbing out, the inner rim of each tray was painted with a mixture of vaseline and salt. Trays were placed randomly in a growth chamber (light/dark: 16/8 h) at 20±2 °C, and watered when necessary to maintain adequate moisture conditions for slug activity.
We designed three experiments in order to evaluate the predation levels: on eggs (experiment 1), on different sized preys (experiment 2), and on preference in the presence of an alternative prey (experiment 3), respectively. Experiment 1 had two treatments: 10 slug eggs with and without the presence of a Scarites anthracinus adult (n=10). Experiment 2 had the following treatments: 10 slugs of each: 10, 20 and 30 mg body mass (n=10), and a control of 15 mg-slugs without the presence of a predator (n=10). Finally, for Experiment 3 we established two treatments of five 15 mg-slugs + five 15 mg-pillbugs with and without the predator (n=8). In all treatments the group of preys was placed in the centre of a shallow plastic tray. Treatments with eggs were evaluated after 24 and 72 h and the number of eggs consumed (eggs missing or only small pieces remaining) were recorded. The rest of the treatments were examined every 24 h for 3 days, and the number of slugs and/or pillbugs consumed (missing, or only small pieces of bodies remaining) were recorded.
No replacement of individuals were performed during the experiments. Data were analysed by repeated measures analysis of variance (ANOVA, PROC GLM) using SAS v.8 software (SAS 2001). When significant effects were detected (P<0.05), differences between means were further compared by Duncan's test.
RESULTS
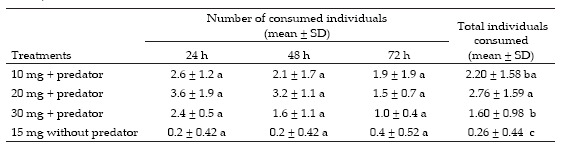
Overall the treatments had a significant effect on egg numbers throughout the incubation period (F1,37=38.5, P<0.001) and among treatments (F1,37=34.2, P<0.001). After 24 h, the number of eggs did not differ from the control without predator (P=0.87), but after 72 h the number of consumed eggs was significantly higher in the treatment including the predator (F1,377=65.8, P<0001, Table 1).
Table 1. Mean number (±Standard Deviation) of Deroceras reticulatum consumed by Scarites anthracinus in a controlled laboratory no-choice assay. Different letter within the same column mean significant differences (Duncan test, P<0.05).
Tabla 1. Número promedio (±Destvío Estándar) de huevos de Deroceras reticulatum consumidos por Scarites anthracinus en experimentos de no elección bajo condiciones controladas de laboratorio. Distinta letra dentro de la misma columna: diferencias significativas (prueba de Duncan, P<0.05).

There were significant differences in the number of slugs consumed at different moments (F2,47=6.8, P=0.002) and among size classes (F2,47=21, P<0.001), but there were no significant differences in the number of slugs consumed per size class after 24, 48 and 72 h (P>0.05). Overall, beetles consumed a greater proportion of 20-mg juvenile slugs than those of 30 or 10 mg (Table 2).
Table 2. Mean number (±Standard Deviation) of different sized Deroceras reticulatum juveniles consumed by Scarites anthracinus in a controlled la1boratory no-choice assay. Different letter within the same column mean significant differences (Duncan test, P<0.05).
Tabla 2. Número promedio (±Desvío Estándar) de juveniles de Deroceras reticulatum de distinta talla consumidos por Scarites anthracinus en experimentos de no elección bajo condiciones controladas de laboratorio. Distinta letra dentro de la misma columna: diferencias significativas (prueba de Duncan, P<0.05).
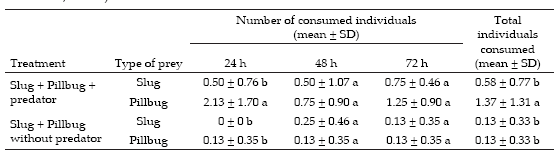
There were not significant differences in the number of slug or pillbugs consumed throughout the incubation period (P=0.42) but during the first 24 h, S. anthracinus consumed significantly more pillbugs than D. reticulatum (P<0.05), although there were no significant differences between the number consumed after 48 and 72 h (P>0.05). Overall, S. anthracinus consumed significantly more pillbugs than D. reticulatum (P<0.05) (Table 3).
Table 3. Mean number (±Standard Deviation) of Deroceras reticulatum (slug) juveniles and Armadillidium vulgare (pillbug) consumed by Scarites anthracinus in controlled laboratory choice assay. Different letter within the same column mean significant differences (Duncan test, P<0.05).
Tabla 3. Número promedio (±Desvío Estándar) de juveniles de Deroceras reticulatum (slug) y Armadillidium vulgare (pillbug) consumidos por Scarites anthracinus en experimentos de elección bajo condiciones controladas de laboratorio. Distinta letra dentro de la misma columna: diferencias significativas (prueba de Duncan, P<0.05).
DISCUSSION
This study demonstrates that S. anthracinus consumes D. reticulatum eggs and juveniles in laboratory experiments and that the individuals of intermediate size were preferred. Moreover, even though S. anthracinus exhibited a preference for other prey (Armadillidium vulgare) it also consumed a moderate proportion of D. reticulatum. This is one of the few studies showing Scaritini species as a slug predator.
Oberholzer & Frank (2003) found that Pterostichus melanarius predation on D. reticulatum eggs was greatest after 24 h. The lower number of eggs consumed by S. anthracinus during this period was probably a consequence of the beetle reduced search activity during an initial acclimatization stage, limiting the probability of finding eggs. Our results support the idea that S. anthracinus has the ability to find slug eggs on the soil surface. However, under conditions of less than 75% soil moisture, eggs may be placed into soil cavities or cracks and covered with earth (Kingston 1996), and so may be less prone to predation.
There were differences in the total consumed after 72 h, with a higher consumption rate of 20 mg-juvenile. It is likely that larger prey satiate carabids with a lower number of slugs. It is also possible that the lower mobility of the 10 mg-juveniles result in a lower encounter rate, and thus, in lower predation. In a miniplot trial, under similar conditions, McKemey et al. (2003) found no significant differences in P. melanarius predation rates on different sizes of D. reticulatum. This beetle has the ability to kill and eat D. reticulatum with body masses smaller than 40 mg (Mckemey et al. 2001), although other species, like Pterostichus madidus and Nebria brevicollis, consume only D. reticulatumwith body masses less than 11 mg (Mair & Port 2001a). Moreover, P. melanarius consumed less number of slugs of 34 mg than of 3.3 mg body mass (Oberholzer & Frank 2003). It is possible that Pterostichini, with a higher mobility, may have a higher encounter rate than Scaritini.
In this experiment, we only recorded dead slugs after 24 h. However, we observed S. anthracinus attack slugs from the onset of the experiment. Early in the period, attacked slugs responded with mucus production, allowing them to slip out of the predator mandibles. Nevertheless, by the end of the day, the slugs produced less mucus and were more susceptible to future attacks. Similar observations were made by Mair & Port (2002), who studied the influence of mucus production on the predation of two carabids, P. melanarius and N. brevicollis, demonstrating that consumption of stressed slugs was greater than healthy slugs.
Adult S. anthracinus consumed both slugs and pillbugs, but preferred pillbugs. It is likely that the greater mobility of pillbugs resulted in a higher encounter rate, and thus increased predation rates. In previous studies Mair & Port (2001b) showed the greater preference of two carabids, P. madidus and N. brevicollis for earthworms and Calliphora fly larvae than for slugs. They concluded that slugs might not be preferred because of their mucus.
Our experiment demonstrated that S. anthracinus might be a natural enemy of D. reticulatum in field condition consuming slug eggs and juveniles of different body mass. Even in the presence of an alternative prey, they did not stop consuming slugs. Alternative preys may help to sustain and retain generalist predators within crops when the target pests are absent or in low density. This dietary flexibility of generalists can, in theory, give them a significant advantage over specialist natural enemies, allowing them to be present within a crop early in the year before other pests arrive in any significant numbers (Murdoch et al. 1985; Symondson et al. 2006). Nevertheless, this simple experiment must be interpreted carefully. It does not necessarily show how many slugs or pillbugs will be consumed under field conditions, or how temporal changes in prey availability affect predator-prey interactions. Moreover, while shallow plastic trays limit predator and prey movement patterns, the choice of available preys may also influence consumption.
ACKNOWLEDGEMENTS
We are most grateful to D. A. Landis for his valuable comments on the manuscript. Many thanks are also due to three anonymous reviewers for their valuable suggestions on an earlier version of the manuscript.
REFERENCES
ARAGÓN, J. 1998. Manejo integrado de plagas relacionadas a la siembra directa. Pp. 163-175 in: PANIGATTI, JL; H MARELLI; D BUSCHIAZZO & R GIL (eds.). Siembra directa, Hemisferio Sur. 333 pp. [ Links ]
BARNEY, RJ & BC PASS. 1986. Foraging behaviour and feeding preference of ground beetles (Coleoptera:Carabidae) in Kentucky alfalfa. J Econ Entomol 17: 1334-1337. [ Links ]
BAILEY, SER. 2002. Molluscicidal baits for control of terrestrial gastropods. Pp. 33-54 in: BARKER, GM (ed.). Molluscs as crop pests, CABI Publishing. 576 pp. [ Links ]
BEST, RL & CC BEEGLE. 1977. Food preferences of five species of carabids commonly found in IOWA corfields. Env Entomol 6:9-12. [ Links ]
BYERS, RA & DD CALVIN. 1994. Economic injury levels to field corn from slug Stylommatophora: Agriolimacidae) feeding. J Econ Entomol 87(5): 1345-1350. [ Links ]
CARMONA, DM & D LANDIS. 1999. Influence of refuge habitats and cover crops on seasonal activity-density of ground beetles (Coleoptera: Carabidae) in field crops. Env Entomol 28:1145-1153. [ Links ]
CARMONA, DM; PL MANETTI; AM VINCINI; A LÓPEZ; HA ÁLVAREZ CASTILLO ET AL. 2004. Abundancia, riqueza y dominancia específica de Carábidos (Insecta: Coleoptera) en agroecosistemas bajo siembra directa y manejo convencional. Pp. 98 in: II Reunión Binacional de Ecología. Mendoza. [ Links ]
CLEMENTE, NL; AJ FABERI; AN LÓPEZ; PL MANETTI & HA ÁLVAREZ CASTILLO. 2007. Biología de Deroceras reticulatum y D. laeve, moluscos de cultivos en siembra directa. RIA 36(2):129-142. [ Links ]
COSTAMAGNA, AC; PL MANETTI; HA ÁLVAREZ CASTILLO & V SADRAS. 1999. Avances en el manejo de babosas en siembra directa. Pp. 101-105 in: Cosecha gruesa. Jornada anual de actualización profesional. Mar del Plata. [ Links ]
CHAPMAN, RB; AS SIMEONIDIS & JT SMITH. 1997. Evaluation of Metallic Green Ground Beetle as predator of slug. Pp. 51-55 in: 50th New Zealands Plant Protection Conference, Palmerston North. [ Links ]
GARCÍA, FRM. 1999. Zoología Agrícola. Manejo Ecológico de Pragas. Porto Alegre. 248 pp. [ Links ]
GLEN, DM & WOC SYMONDSON. 2003. Influence of tillage on slugs and their natural enemies. Pp. 207-227 in: Titi, AE (ed.). Soil Tillage in Agroecosystems. CRC Press. Florida. 384 pp. [ Links ]
HOMMAY, G. 2002. Agriolimacidae, Arionidae and Milacidae as pests in west european sunflower and maize. Pp. 245-254 in: Barker, GM (ed.). Molluscs as crop pests, CABI Publishing. 576 pp. [ Links ]
KINGSTON, N. 1996. Observations on the laboratory rearing of terrestrial molluscs. Am Midland Nat 76(2): 528-532. [ Links ]
KROMP, B. 1999. Carabid beetles in sustainable agriculture: a review on pest control efficacy, cultivation impacts and enhancement. Agr Ecosyst Env 74:187-228. [ Links ]
LOVEI, G & KD SUNDERLAND. 1996. Ecology and behavior of ground beetles (Coleoptera: Carabidae). Ann Rev Entomol 41:231-56. [ Links ]
MAIR, J & GR PORT. 2001a. Predation by the carabid beetles Pterostichus madidus and Nebria brevicollis is affected by size and condition of the prey slug Deroceras reticulatum. Agr Forest Entomol 3(2): 99- 106. [ Links ]
MAIR, J & GR PORT. 2001b. Predation on the slug Deroceras reticulatum by the carabid beetles Pterostichus madidus and Nebria brevicollis in the presence of alternative prey. Agr Forest Entomol 3(3):169-174. [ Links ]
MAIR, J & GR PORT. 2002. The influence of mucus production by the Slug, Deroceras Reticulatum, on predation by Pterostichus madidus and Nebria brevicollis (Coleoptera: Carabidae). Biocontr Sci Technol 12(3):325-335. [ Links ]
MANETTI, PL; AN LÓPEZ; N CLEMENTE; HA ÁLVAREZ CASTILLO; H GIZZI ET AL. 2005. Efecto de los molusquicidas en el control de "babosas", (Pulmonata: Stylomatophora) en el cultivo de soja. Pp. 366 in: XIX Encontro Brasileiro de Malacologia. Río de Janeiro. [ Links ]
MARASAS, ME. 2001. Efectos de distintos sistemas de labranza y diversidad de la coleopterofauna edáfica, con especial referencia a las especies de Carabidae, en un cultivo de trigo y los ambientes naturales circundantes. Tesis doctoral. Facultad de Ciencias Naturales, La Plata, Argentina. [ Links ]
MASTRONARDI, F. 2006. Control químico de isópodos y babosas en un cultivo de girasol bajo siembra directa. Tesis de grado. Facultad de Ciencias Agrarias, Mar del Plata, Argentina. [ Links ]
MCKEMEY, A; W SYMONDSON; D GLEN & P BRAIN. 2001. Effects of slug size on predation by Pterostichus melanarius (Coleoptera: Carabidae). Biocontr Sci Technol 11(1):81-91. [ Links ]
MCKEMEY, A; W SYMONDSON & D GLEN. 2003. Predation and prey size choice by the carabid beetle Pterostichus melanarios (Coleóptera: Carabidae): The dangers of extrapolating from laboratory to field. Bull Entomol Res 93(3):227-34. [ Links ]
MURDOCH, WW; J CHESSON & PL CHESSON. 1985. Biological control in theory and practice. Am Nat 125:344-366. [ Links ]
OBERHOLZER, RF & T FRANK. 2003. Predation by the carabid Beetles Pterostichus melanarius and Poecilus cupreus on Slugs and Slug Eggs. Biocontr Sci Technol 13:99-110. [ Links ]
ROIG-JUÑENT, S. 1998. Carabidae. Pp. 194-209 in: MORRONE, JJ & S COSCARÓN (eds.). Biodiversidad de Artrópodos Argentinos. Ediciones Sur, La Plata. 599 pp. [ Links ]
SALUSO, A. 2001. Isópodos terrestres asociados al cultivo de soja en siembra directa. Soja. Actualización técnica INTA EEA-Paraná 21:80-83. [ Links ]
SAS. 2001. SAS user's guide: statistics. Vers. 8. SAS Institute, Cary. NC. USA. [ Links ]
SOUTH, A.1992. Terrestrial slugs. Biology, ecology and control. Chapman & Hall. London. 428 pp. [ Links ]
STUDDERT, GA & HE ECHEVERRIA. 2000. Maíz, girasol y soja en los sistemas de cultivo del sudeste bonaerense. Pp. 407-437 in: Andrade, FH & V Sadras (eds.). Bases para el manejo del maíz, el girasol y la soja. Advanta Semillas SAIC. Balcarce. 443 pp. [ Links ]
SYMONDSON, WO. 1997. Does Tandonia budapestis (Mollusca : Pulmonata) contain toxins? Evidence from feeding trials withh the slug predator Pterostichus melanarius (Coleoptera: Carabidae). J Moll Studies 63:541-545. [ Links ]
SYMONDSON, WO; S CESARINI; PW DODD; GL HARPER; MW BRUFORD ET AL. 2006. Biodiversity vs. biocontrol: positive and negative effects of alternative prey on control of slugs by carabid beetles. Bull Entomol Res 96:637-645. [ Links ]
TRUMPER, E & M LINARES. 1999. Bicho Bolita, nueva amenaza para la soja. Súper Campo 5:24-27. [ Links ]

