










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana v.42 n.1 Buenos Aires jan./mar. 2005
Estudio microflorístico de las formaciones Potrerillos y Cacheuta (Triásico) en el sur del cerro Cacheuta, Mendoza, Argentina. Parte 1
Leandro David Rojo1 y Ana María Zavattieri2
1 Laboratorio de Paleoecología y Paleopalinología, Departamento de Biología, Universidad Nacional de Mar del Plata, Funes 3250, 7600 Mar del Plata, Argentina. ldrojo@mdp.edu.ar
2 Laboratorio de Paleopalinología, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Centro Regional de Investigaciones Científicas y Tecnológicas - Consejo Nacional de Investigaciones Científicas y Técnicas, Casilla de Correo 131, 5500 Mendoza, Argentina. amz@lab.cricyt.edu.ar
Resumen. En dos secciones aflorantes al sur del cerro Cacheuta, provincia de Mendoza, en las sedimentitas triásicas de las Formaciones Potrerillos y Cacheuta, se identificaron 85 especies de esporas, granos de polen y algas clorococales. En esta primera parte se da un resumen del contexto geológico basado en un detallado estudio sedimentológico previo que indica condiciones paleoambientales continentales fluvio/lacustres para ambas unidades en conjunto. Se analiza y reubica la procedencia estratigráfica de los clásicos niveles plantíferos y de las microfloras previamente estudiadas en las "Minas de Petróleo" del cerro Cacheuta. En esta contribución se listan e ilustran 38 especies de esporas de briófitas y pteridófitas de ambas palinofloras, de las cuales se describen sistemáticamente sólo especies que se citan por primera vez para ambas formaciones y/o para el Triásico de Argentina y aquéllas que revisten importancia cronoestratigráfica. Diez especies pertenecientes a los géneros Anapiculatisporites , Antulsporites , Clavatisporites , Densoisporites , Foveosporites , Matonisporites , Neoraistrickia , Raistrickia , Rugulatisporites y Uvaesporites se registran por primera vez para el Triásico de la Cuenca Cuyana. Se presentan las afinidades botánicas conocidas de las mioesporas y se comparan con los grupos vegetales de la megaflora local. La diversidad taxonómica está representada por 38 especies de esporas de briófitas y pteridófitas versus 13 especies conocidas de la megaflora, evidenciando que la biodiversidad microflorística es mayor que la megaflorística.
Abstract. Microfloristic study of the Potrerillos and Cacheuta Formations (Triassic) southern Cacheuta hill, Mendoza, Argentina. Part 1 . At two outcropping sections, south of the Cacheuta hill, Mendoza province, in the Triassic clastic sediments of the Potrerillos and Cacheuta formations, 85 species of spores, pollen grains and chlorococcales algae were identified. In this first part the geologic context is briefly considered and the continental fluvial/lacustrine paleoenvironmental interpretation for both units is based on earlier sedimentologic study. The stratigraphic position of the classic plant levels and previously known microfossil flora from the "Minas de Petroleo", Cacheuta hill, is analized and relocated. In this first contribution thirty eight species of bryophyte and pteridophyte spores from both palynofloras are listed and illustrated, from which those species registered for the first time for both formations and for the Triassic of Argentina and those of chronostratigraphic importance are described systematicaly. Ten species belonging to the genera Anapiculatisporites , Antulsporites , Clavatisporites , Densoisporites , Foveosporites , Matonisporites , Neoraistrickia , Raistrickia , Rugulatisporites and Uvaesporites are new records for the Triassic of the Cuyo Basin. The known botanical affinities of the miospores are presented and compared with the local megaflora species. The taxonomic diversity is represented by 38 species of spores versus 13 species of bryophyte and pteridophyte plant remains known from the megaflora, thus the microfloristic biodiversity is larger than the megafloristic one.
Key words. Palynology. Potrerillos and Cacheuta Formations. Triassic. Mendoza. Argentina.
Palabras clave. Palinología. Formaciones Potrerillos y Cacheuta. Triásico. Mendoza. Argentina.
Introducción
En este trabajo se presenta el estudio sistemático de esporas y granos de polen hallados en las formaciones Potrerillos y Cacheuta que afloran en los perfiles de las secciones de Agua de las Avispas (perfil I) y Puesto Miguez (perfil II), al sur del cerro Cacheuta, en el extremo austral de la Precordillera, provincia de Mendoza (figura 1). Metasedimentitas devónicas forman el núcleo del mencionado cerro, mientras que volcanitas del Grupo Choiyoi se adosan a los flancos oeste y sur del mencionado cerro. Las rocas constitutivas de dicho Grupo fueron acotadas entre el Pérmico Temprano Alto y el Triásico Temprano -o límite Eo-Mesotriásico ( cf. Llambías y Stipanicic, 2002, p. 91). Sobre ellas se apoyan las sedimentitas triásicas continentales de las formaciones Potrerillos, Cacheuta y Río Blanco, faltando en esta localidad, las formaciones Río Mendoza y Cerro de Las Cabras que constituyen las unidades basales del Grupo Uspallata (Stipanicic, 1979; Stipanicic y Zavattieri, 2002) en la cuenca Cuyana. Por encima de la secuencia triásica se dispone localmente y en discordancia, la Formación Mariño del Terciario Medio (Morel, 1994) (figura 1). Dada la profusa literatura sobre la geología y paleontología de las unidades triásicas del sur del cerro Cacheuta, se refiere al lector a los trabajos de Morel (1991 inéd. ; 1994), Stipanicic et al. (1996), Morel et al. (2002) quienes citan los estudios realizados en el área.

Figura 1. Ubicación geográfica y geología de los afloramientos del área sur del cerro Cacheuta y de los perfiles estudiados. Adaptado de Morel, 1994 / Geographic location and outcrop geology of the south of Cacheuta area and the studied profiles. Adapted from Morel, 1994.
Los dos perfiles con los niveles portadores de las microfloras estudiadas distan entre sí aproximadamente 2 km; el perfil I (33º04'52''S; 68º08'56''O) ubicado en la sección Agua de las Avispas (Río Seco de la Mina según Morel, 1994) y el perfil II (33º05'10''S; 69º06'26''O) en Puesto Miguez ("Minas de Petróleo" según Jain, 1968). Ambas secciones fueron detalladamente descriptas por Morel (1994) quien además estudió (Morel, 1991, inéd .) la megaflora contenida en las mismas formaciones y localidades (figuras 1 y 2). Los afloramientos de la Formación Potrerillos en el sur del cerro Cacheuta corresponden al tercio superior de la unidad, teniendo en cuenta el espesor que dicha unidad tiene en su localidad tipo (cerro Bayo de Potrerillos) y en otras partes de la cuenca. El pasaje transicional a la suprayacente Formación Cacheuta se manifiesta por un cambio litológico y de coloración hacia sedimentitas lacustres finamente laminadas y carbonosas; los afloramientos de esta última, en la clásica localidad de Puesto Miguez, constituyen su perfil tipo (Morel et al. , 2002).

Figura 2. Secciones esquemáticas de las unidades aflorantes en el perfil I - Agua de las Avispas y perfil II - Puesto Miguez, sur del cerro Cacheuta, Mendoza con indicación de los niveles muestreados para este estudio. Adaptado de Morel, 1994 / Schematic sections of the outcroping units at the Profile I - Agua de las Avispas and Profile II - Puesto Miquez, at south of cerro Cacheuta, Mendoza, showing palynological sampling levels. Adapted from Morel, 1994.
Hasta la fecha, sólo se cuenta con dos estudios palinológicos realizados en el área. El primero (Orlando, 1954) registró sólo tres "tipos polínicos" en la Formación Cacheuta y el segundo (Jain, 1968) describió una rica asociación microflorística procedente de la Formación Potrerillos en la sección de Puesto Miguez (perfil II). Es importante señalar que la microflora que estudió Jain (1968) es de la Formación Potrerillos y no de la Formación Cacheuta como así lo indica dicho autor en su trabajo "Middle Triassic pollen grains and spores from Minas de Petroleo beds of the Cacheuta Formation (Upper Gondwana), Argentina" . Dichas muestras fueron extraídas de la sección que aflora detrás de la antigua Administración de YPF, denominada por Jain (1968) como "Minas de Petróleo". Este autor (p. 2) indica que las muestras palinológicas "provienen de los intervalos 10-80 m y 90-100 m por encima de la base volcánica de la sección" ... "dentro de los 160 m de espesor que conforma la secuencia triásica aflorante", aunque no presenta ningún perfil que muestre los espesores para ambas formaciones y sus límites. Por su parte, Morel (1991, inéd .; 1994) levantó un perfil litoestratigráfico y sedimentológico de detalle (figura 2) en el que se observa que, dentro de un espesor total aflorante de aproximadamente 155 m, la Formación Potrerillos alcanza cerca de 120 m, mientras que la Formación Cacheuta tiene, por encima de la anterior, unos 35 m, que se constataron en el trabajo de campo realizado. Sobre la base de esta evidencia es que se deduce que la microflora que estudió Jain corresponde a la parte superior de la Formación Potrerillos en lugar de provenir de la Formación Cacheuta. Una consideración adicional a favor de este punto, es que en 1916-17 Wieland recolectó las muestras palinológicas que estudió Jain de los mismos niveles que contenían los restos plantíferos de esta sección (Jain, 1968, p. 2). Por último, en nuestro trabajo de recolección de las muestras para este estudio, comprobamos que los restos de megaflora con excelente estado de preservación en los que identificamos a la "Flora de Kurtz" efectivamente se presentan en los niveles superiores de la primera unidad, mientras que en los de la segunda encontramos escasos restos vegetales fragmentarios y cuando presentes, muy carbonizados.
El estudio de las microfloras procedentes de las formaciones Potrerillos y Cacheuta en la sección de Agua de las Avispas (perfil I) constituye el objetivo principal de este trabajo ya que no se cuenta con estudios palinológicos previos, mientras que el de Puesto Miguez (perfil II) se tomó como complementario por tener ya un estudio palinológico sobre la Formación Potrerillos (Jain, 1968). Por lo tanto, la asociación microflorística hallada en las dos formaciones del perfil I, como en la Formación Cacheuta del perfil II, constituyen registros nuevos.
En esta primera contribución se presenta el estudio de las especies de esporas de briófitas y pteridófitas halladas en ambas formaciones y el análisis de sus afinidades botánicas. En la segunda parte (Zavattieri y Rojo, en prensa) se completará la lista sistemática con los granos de polen de gimnospermas y de algas, se hará el análisis sobre las evidencias cronológicas de los componentes de ambas microfloras, las implicancias bioestratigráficas, paleobiogeográficas y de correlación con asociaciones de otras cuencas triásicas argentinas.
Material y métodos
El presente material fue extraído de 29 niveles de las formaciones Potrerillos y Cacheuta (figura 2). Las muestras palinológicas de la primera provienen de los niveles de arcilitas y limoarcilitas de colores pardo verdosos, gris verdosas a gris negras masivas, niveles de carbón y de lutitas carbonosas que se intercalan entre las areniscas castaño claras y tobáceas en ambos perfiles (figura 2). En el perfil II, Puesto Miguez, las muestras 4423 y 7140 a 7143 corresponden a los niveles plantíferos de Kurtz (1921). De la Formación Cacheuta, fueron extraídas de niveles de lutitas negras carbonosas, laminadas, propias de secuencias de lagos (figura 2).
Las muestras se procesaron según la técnica convencional de extracción palinológica (Volkheimer y Melendi, 1976). Para el estudio del material se utilizó un microscopio Olympus BX50, las fotomicrografías se tomaron con una cámara Olympus PM-C35B con película Agfapan 100 ASA. Los ejemplares se identifican por el número de preparado y las coordenadas correspondientes del microscopio utilizado (con el rótulo a la derecha del observador y/o del objetivo). Los preparados se encuentran en la paleopalinoteca del Laboratorio de Paleopalinología del IANIGLA, CRICYT-Mendoza, con los números 7070-7084, 7140- 7148 y 4423 y la sigla MPLP que identifica el repositorio del citado laboratorio.
En este estudio se describen sistemáticamente las especies de esporas que se citan por primera vez para ambas formaciones, las que constituyen nuevos registros para el Triásico de Argentina, aquellas especies con asignación sistemática abierta y las que revisten interés cronoestratigráfico. Las especies de esporas triletes y monoletes que se hallaron se mencionan en la siguiente lista en orden alfabético.
Lista taxonómica de especies
Esporas de briófitas y pteridófitas
Anapiculatisporites cooksonae Playford 1965. Figura 3.N
Anapiculatisporites cf. pristidentatus Reiser y Williams 1969. Figura 3.O
Antulsporites sp. A (en McKellar 1974). Figuras 4.O y Q
Apiculatisporis sp. cf. A . globosus (Leschik) Playford y Dettmann 1965. Figura 4.I
Aratrisporites parvispinosus (Leschik) Playford 1965. Figura 4.R
Biretisporites sp. 3 (en Zavattieri 1986). Figura 3.G
Cacheutasporites wielandii Jain 1968. Figura 3.B
Cadargasporites cuyanensis Azcuy y Longobucco 1983. Figura 4.F
Cadargasporites verrucosus Reiser y Williams 1969. Figura 4.N
Calamospora tener (Leschik) de Jersey 1962. Figura 3.H
Cirratriradites sp. 1 (en Zavattieri 1990b). Figura 4.G
Clavatisporites sp. Figura 4.A
Convolutispora sp. 1 (en Zavattieri 1986). Figura 3.Q
Deltoidospora directa (Balme y Hennelly) Norris, 1965. Figura 3.C
Densoisporites complicatus Balme 1970. Figura 4.E
Dictyophyllidites harrisii Couper, 1958.
Dictyophyllidites mortonii (de Jersey) Playford y Dettmann 1965. Figura 3.D
Espora trilete gen. et sp. indet. Figura 4.L
Foveosporites mimosae de Jersey y Hamilton 1967. Figura 4.J
Leptolepidites crassibalteus Filatoff 1975. Figura 3.L
Leptolepidites volkheimerii Zavattieri 1986. Figura 3.R
Lophotriletes bauhiniae de Jersey y Hamilton 1967. Figura 3.M
Matonisporites equiexinus Couper 1958. Figura 3.P
Neoraistricka sp. Figura 4.M
Osmundacidites cacheutensis Jain 1968. Figura 4.B
Osmundacidites wellmanii Couper 1953. Figura 4.H
Punctatosporites walkomii de Jersey 1962. Figura 4.S
Raistrickia sp. Figura 4.D
Rogalskaisporites cicatricosus (Rogalska) Danzé-Corsin y Laveine 1963. Figura 3.I
Rugulatisporites permixtus Playford en Playford, Rigby y Archibald 1982. Figura 4.C
Rugulatisporites cf. trisinus de Jersey y Hamilton 1967. Figura 3.K ab
Striatella seebergensis Mädler 1964. Figura 4.K
Thymospora ipsviciencis (de Jersey) Jain 1968. Figura 4.P
Todisporites cinctus (Maljavkina) Orlowska-Zwolinska 1971. Figura 3.A
Todisporites major Couper 1958. Figura 3.J
Todisporites minor Couper 1958.
Uvaesporites glomeratus (Döring) Hiltmann 1967. Figura 3.F
Uvaesporites verrucosus (de Jersey) Helby en de Jersey 1971b. Figura 3.E
Descripciones sistemáticas
Esporas triletes
Género Anapiculatisporites Potonié y Kremp 1954
Especie tipo. Anapiculatisporites isselburgensis Potonié y Kremp 1954.
Anapiculatisporites cooksonae Playford 1965 Figura 3.N
Material estudiado. 7144E: 30,9/108,9 (foto); 7074C: 37,3/107,7 MPLP.

Figura 3. A, Todisporites cinctus (Maljavkina) Orlowska-Zwolinska. 7074C: 36,8/104,5 MPLP (x 650). B, Cacheutasporites wielandii Jain. 7074F: 38,8/114,2 MPLP (x 750). C, Deltoidospora directa (Balme y Hennelly) Norris. 7074C: 56,9/95 MPLP (x 750). D, Dictyophyllidites mortonii (de Jersey) Playford y Dettmann. 7070B: 35,1/108.0 MPLP (x 700). E, Uvaesporites verrucosus (de Jersey) Helby en de Jersey. 4423I: 32/95,2 MPLP; Vista proximal-ecuatorial / Proximal-equatorial view (x 700). F, Uvaesporites glomeratus (Döring) Hiltmann. 4423I: 30,6/104,4 MPLP; vista proximal / proximal view (x 750). G. Biretisporites sp. 3 (en Zavattieri, 1986). 4423I: 40,3/109,1 MPLP (x 800). H, Calamospora tener (Leschik) de Jersey. 4423I: 39,8/101,1 MPLP (x 550). I, Rogalskaisporites cicatricosus (Rogalska) Danzé-Corsin y Laevine. 7075E: 55,9/109,0 MPLP (x 900). J, Todisporites major Couper. 7074C: 42,7/99,0 MPLP (x 700). K, Rugulatisporites cf. trisinus de Jersey y Hamilton. 7074F: 38,9/108,1 MPLP. a, Vista distal, mostrando las rúgulas / Distal view, showing rugulae ; b, Vista proximal mostrando la marca trilete / Proximal view, showing the trilete scar (x 800). L. Leptolepidites crassibalteus Filatoff. 4423I: 37,9/104,3 MPLP (x 750). M, Lophotriletes bauhiniae de Jersey y Hamilton. 7074C: 38,6/108,5 MPLP (x 750). N, Anapiculatisporites cooksonae Playford. 7144E: 30,9/108,9 MPLP; vista ecuatorial, contorno distorsionado por pliegues / equatorial view, outline modified by folds (x 800). O, Anapiculatisporites cf. pristidentatus Reiser y Williams. 7074C: 53,4/104,9 MPLP; Vista proximal, distorsionada por pliegues / Proximal view, distorted by folds (x 700). P, Matonisporites equiexinus Couper. 7074F: 47,2/110,2 MPLP (x 500). Q, Convolutispora sp. 1 (en Zavattieri, 1986). 7074F: 47,0/111,7 MPLP. Vista ecuatorial / equatorial view (x 800). R, Leptolepidites volkheimerii Zavattieri. 4423I: 41,2/103,3 MPLP; vistal distal / distal view (x 800).
Descripción. Espora trilete de contorno circular a subcircular. Marca trilete poco distinguible. Exina de aprox. 2 µm de espesor. La ornamentación está densamente dispuesta, pero con elementos discretos, y se compone de conos y verrugas de 2-4 µm de alto y bases expandidas de aprox. 2 µm de diámetro; las espinas se hallan en menor proporción. En la superficie proximal la ornamentación está marcadamente reducida y en algunos casos se observan áreas de contacto casi lisas.
Dimensiones. Diámetro ecuatorial máximo: 43-47 µm (2 ejemplares).
Observaciones. En la descripción original los elementos esculturales presentan bases con rangos de 1 a 7,5 µm de diámetro y alturas de 2 a 8 µm, mayores a los de este material, aunque el autor de la especie señala que usualmente, los especímenes más grandes poseen elementos esculturales más grandes y viceversa (Playford, 1965). Este material coincide con lo señalado por este último autor, ya que presenta especímenes intermedios en el rango de variabilidad de las dimensiones ecuatoriales (36-59 µm) y de sus elementos ornamentales. En el material ilustrado por Farabee et al. (1989) asignado a esta especie se aprecian los conos con extremos acuminados como en estos ejemplares. Anapiculatisporites sp. 1 (Zavattieri, 1990a) tiene ornamentación similar a éstos, pero difiere en sus menores dimensiones ecuatoriales (25-42 µm), conos más reducidos (bases: 0,7-2 µm; alto: 1,5 µm) y exina más fina (1 µm).
Afinidad botánica. El género presenta afinidad con el grupo de las Hymenophyllaceae (Filatoff, 1975) y a las Lycopsida en general (Balme, 1995).
Distribución. Argentina: Triásico de la Formación Potrerillos y Cacheuta (perfiles I y II, este trabajo), cuenca Cuyana. Australia: Triásico (Playford, 1965). Antártida: Triásico (Farabee et al. , 1989).
Género Antulsporites Archangelsky y Gamerro 1966b
Especie tipo. Antulsporites baculatus (Archangelsky y Gamerro) Archangelsky y Gamerro 1966b.
1966 a. Heliosporites Archangelsky y Gamerro, p. 203.
1966 b. Antulsporites Archangelsky y Gamerro, p. 369.
Observaciones. McKellar, 1974 (p. 28) discute la relación entre los géneros Heliosporites y Antulsporites propuestos por Archangelsky y Gamerro (1966a y 1966b, respectivamente), criterios que se siguen en este trabajo.
Antulsporites sp. A (en McKellar, 1974) Figuras 4.O y Q
Véanse sinonimias en McKellar, 1974, p. 29-30.
Material estudiado. 4423I: 29,8/106,9 (foto); 37,8/96,6; 35,7/96,0; 7074F: 37,5/94,5 MPLP.
Descripción. Espora trilete de contorno subcircular a subtriangular con ángulos redondeados. Marca trilete de rayos algo sinuosos que se extienden casi hasta el ecuador, bordeados por labios finos que se adelgazan hacia los extremos. Exina engrosada ecuatorialmente de manera irregular por la fusión de la escultura en el contorno. Cara proximal lisa; superficie distal con verrugas y gránulos. Los elementos ornamentales son discretos, distribuidos en forma irregular y/o también se fusionan o coalescen de a dos o más. Las verrugas son subcirculares, subpoligonales o de bases irregulares; en perfil tienen cimas rectas, redondeadas, ligeramente cónicas o trapezoidales con bases anchas; en el último caso, los elementos gradan a báculas cortas y gruesas; el tamaño de las verrugas varía entre 0,5 y 3 µm de alto y entre 0,5 y 5 µm de base o más (cuando están fusionados dos o más elementos); los gránulos tienen diámetros y alturas que varían entre 0,5 y 2 µm. Una banda concéntrica o anillo subecuatorial distal se forma por la fusión de las verrugas y gránulos, mientras que el borde externo del mismo es irregular por la proyección de los elementos que lo forman; el anillo o banda tiene un ancho variable de 1 a 3 µm y varía su posición más subecuatorial o más polar de un grano a otro.
Dimensiones. Diámetro ecuatorial máximo: 38-45 µm (2 ejemplares).
Material estudiado. 4423I: 34,1/98,1, 30,0/113,4 MPLP.
Observaciones. Los dos ejemplares hallados en este material se ajustan a la descripción y dimensiones de Antulsporites sp. A (en McKellar, 1974). Antulsporites sp. B, del mismo autor, se diferencia de la primera especie en que aquélla tiene las verrugas de la cara distal distribuidas en forma irregular, mientras que en la segunda las verrugas se ubican dentro del anillo subecuatorial y dispuestas a su vez en forma anular a manera de un engrosamiento concéntrico subpolar; el ecuatorial o anillo a manera de cíngulo es liso, más demarcado y más grueso en A. sp. B, mientras que en A. sp. A puede ser liso, irregular o ligeramente estriado como en este material.
Afinidad botánica. Esporas de briófitas (McKellar, 1974; de Jersey y Raine, 1990).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfil II, este trabajo), cuenca Cuyana. De acuerdo con las sinonimias específicas de A . sp. A dadas por McKellar (1974), este tipo de esporas se distribuye en sedimentos del Jurásico Inferior a Jurásico Medio de Australia (detalles de distribución en el trabajo mencionado, p. 30).
Género Apiculatisporis Potonié y Kremp 1956
Especie tipo (por designación original). Apiculatisporis aculeatus (Ibrahim) Potonié 1956.
Apiculatisporis sp. cf. A . globosus (Leschik) Playford y Dettmann 1965. Figura 4.I
Dimensiones. Diámetro ecuatorial máximo, 48-56 µm (4 ejemplares).
Localidad. Perfiles de Agua de las Avispas y Puesto Miguez.
Observaciones. Este material se caracteriza por una mayor proporción de báculas y conos que de espinas y con exina más fina respecto de A . globosus Playford y Dettmann (2,5-3,5 µm en A . globosus ). Apiculatisporis sp. cf. A . globosus (en de Jersey y Raine, 1990) es comparable a este material y parece coincidir en todas sus características, difiriendo sólo en que estos ejemplares presentan escultura más espaciada y reducida. Apiculatisporis cf. globosus (en Zavattieri, 1990a) posee menores dimensiones (30-43 µm) y mayor proporción de espinas que estos ejemplares. Apiculatisporis clematisi de Jersey 1968 no presenta labios conspicuos que acompañan los rayos de la marca trilete y predominan los conos respecto del resto de los elementos ornamentales, características que también la distinguen de las de este material. Un ejemplar idéntico fue ilustrado por Farabee et al. (1990, figura 4-14) como A . globosus (Leschik) Playford y Dettmann.
Afinidad botánica. Filicales indiferenciadas (de Jersey y Raine, 1990).
Distribución. (para A . globosus ) Argentina: Triásico de las formaciones Cerro de Las Cabras (Zavattieri, 1990a), Potrerillos (perfiles I y II, este trabajo), cuenca Cuyana. Amplia distribución en sedimentos del Triásico Superior y Jurásico de Australia y Nueva Zelanda (véase detalle de los registros en de Jersey, 1971b, p. 7; de Jersey, 1974; Stevens, 1981 y de Jersey y Raine, 1990). En Antártida se encuentra en Fremow Formation (Triásico Inferior a Triásico Medio) (Farabee et al. , 1989).
Género Cirratriradites (Wilson y Coe) Potonié y Kremp 1954
Especie tipo. Cirratriradites saturni (Ibrahim) Potonié y Kremp, 1954.
Cirratriradites sp. 1 (en Zavattieri, 1990b) Figura 4.G
Material estudiado. 7141B: 40,4/112,3 (foto). 4423I: 30,1/112,3; 42,1/95,7; 50/111,9. 7141A: 39,8/93,5. 7141G: 41,9/105,9 MPLP.
Descripción. Espora trilete zonada, de contorno ecuatorial subtriangular, de lados convexos y ápices redondeados. Marca trilete con rayos que llegan hasta el ecuador, con finos labios sinuosos, más engrosados hacia el polo. Cara proximal escabrada, con gránulos; áreas de contacto triangulares. Cara distal densamente granulada-verrucada; elementos pequeños de 1-2 µm de diámetro y aprox. 1 µm de alto. La zona, de aprox. 5-6 µm de ancho, presenta la misma escultura de la cara distal y el contorno parece aserrado por la proyección de la escultura sobre el borde ecuatorial. Algunos especímenes presentan una zona central más oscura donde la exina está engrosada, de aprox. 10 µm de diámetro, subcircular o paralela al contorno de la espora.
Dimensiones. Diámetro ecuatorial máximo, 30-53 µm; diámetro ecuatorial del cuerpo central, 23-42 µm (6 ejemplares).
Afinidad botánica. Esporas de Lycopsida pertenecientes a las familias Selaginellales, Lycopodiales (de Jersey y Raine, 1990).
Distribución. Argentina. Triásico de las formaciones Cerro de las Cabras (Zavattieri, 1990b), Potrerillos (perfil II, este trabajo), cuenca Cuyana.
Género Clavatisporites Kedves y Simoncsics 1964
Especie tipo. Clavatisporites clavus Kedves y Simoncsics 1964.
Clavatisporites sp. Figura 4.A
Material estudiado. 4423I: 44, 0/111,6 MPLP.
Descripción. Espora trilete de contorno circular a subcircular. Marca trilete simple, de rayos rectos que no llegan al ecuador. Exina fina de aprox. 1 µm de espesor, ornamentada en ambas caras por clavas y báculas con cimas más anchas y redondeadas y gemas en menor proporción; la escultura está densamente dispuesta pero los elementos son discretos; bases de 0,5-1,5 µm de diámetro y de 1 a 2 µm de alto.
Dimensiones. Diámetro ecuatorial máximo, 40 µm (1 ejemplar).
Observaciones. Este único ejemplar que se encontró tiene clavas como ornamentación dominante, además de báculas y gemas. No se observa una evidente reducción de los elementos en la cara proximal, pero esta característica podría aseverarse con mayor número de ejemplares. La dominancia de clavas, y en menor proporción báculas de cimas ensanchadas y gemas, distingue este ejemplar de las especies descriptas para el género. Material similar a éste ha sido descripto e ilustrado por Rigby y Hekel (1977) como Clavatisporites spp.
Afinidad botánica. Filicopsida indiferenciada (de Jersey y Raine, 1990).
Distribución. Argentina: Triásico de las Formación Potrerillos (perfil II, este trabajo), cuenca Cuyana.
Género Densoisporites Weyland y Krieger emend . Dettmann 1963
Especie tipo (por designación original). Densoisporites velatus Weyland y Krieger emend. Krasnova en Samoilovich et al. 1961.
Densoisporites complicatus Balme 1970 Figura 4.E
Material estudiado. 4423I: 38,9/92,5 (foto); 44/97,9; 45,1/93,9; 47,1/107,8; 42,5/94,6; 43,5/112,3; 49/102. 7078D: 51/112,3. 7141B: 31,2/95,3 MPLP.
Descripción. Espora trilete cíngulo-cavada, de contorno triangular a subtriangular de lados rectos a ligeramente convexos y ápices redondeados. Marca trilete con labios que se engrosan hacia el polo proximal y llegan hasta el ecuador de la espora. Cara proximal lisa, con áreas de contacto bien distinguibles; cara distal escabrada de aspecto granular. Intexina fuertemente triangular, lisa o con ornamentación muy pequeña. Exoexina más gruesa ecuatorialmente, formando un anillo de aprox. 5 µm de ancho en algunos ejemplares (como en el que se ilustra).
Dimensiones. Diámetro máximo de la exoexina, 28- 42 µm; diámetro máximo de la intexina, 19-29 µm (9 ejemplares).
Observaciones. Como señala Balme (1970), esta especie se diferencia de otras del género por presentar intexina triangular y marca trilete con bordes elevados hacia el centro de la lesura y exoexina engrosada ecuatorialmente.
Afinidad botánica. Lycopsida (Isoetales, Lycopodiales, Selaginellales) (Balme, 1995).
Distribución. Argentina: Triásico de las formaciones Potrerillos y Cacheuta (perfiles I y II, este trabajo), cuenca Cuyana. Permo-Triásico: Salt Range y Surghar Range, Pakistán (Balme, 1970).
Género Foveosporites Balme 1957
Especie tipo. Foveosporites canalis Balme, 1957.
Foveosporites mimosae de Jersey y Hamilton 1967 Figura 4.J
Material estudiado. 7070B: 37,1/95,2 (foto); 37,5/ 107,5; 34,6/110,5 MPLP.
Descripción. Espora trilete de contorno ecuatorial subcircular. Los rayos de la lesura son difíciles de distinguir y no llegan al ecuador de la espora. Exina de aprox. 2 µm de espesor, ornamentada en ambas caras con foveolas de contornos circulares u ovales de 1-2 µm de diámetro, irregularmente distribuidas; en pocos casos se fusionan para formar pequeñas vermículas o canales cortos.
Dimensiones. Diámetro mayor, 63-93 µm (3 ejemplares).
Observaciones. En este material no se ha podido observar la marca trilete de rayos rectos, coincidiendo con la diagnosis original que señala que la lesura puede ser poco conspicua. El aspecto general de los ejemplares ilustrados por de Jersey y Hamilton es muy similar al de Cacheuta, tanto en la forma general -circular-, el tamaño (49-81 µm) y la ornamentación, pero se distingue en que el material australiano tiene exina más gruesa (3 µm) que en estos ejemplares. Foveosporites mimosae de Jersey y Hamilton se diferencia de F . moretonensis de Jersey, 1964 descripta en el Triásico Superior de Australia, por su mayor tamaño, forma circular o casi circular de la espora y por la ornamentación más conspicua. Foveosporites sp. en Ottone et al. (1992) es también similar a estos ejemplares y sólo difiere en que en este material las foveolas están más espaciadamente distribuidas y sólo algunas de ellas son subcirculares a subovales.
Afinidad botánica. Lycopsida (Selaginellales) (Balme, 1995).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfil I, este trabajo), cuenca Cuyana. Australia: Triásico Medio (de Jersey y Hamilton, 1967).
Género Lophotriletes Naumova ex Potonié y Kremp 1954
Especie tipo. Lophotriletes gibbosus (Ibrahim) Potonié y Kremp 1954.
Lophotriletes bauhiniae de Jersey y Hamilton 1967 Figura 3.M
Principal material estudiado: 7074C: 38,6/108,5 (foto); 42,9/107,7. 7070C: 44/103,3. 7074F: 39,5/109; 52,7/108,9; 52,8/113,4. 7075E: 54,4/107,2. 7147D: 37,5/96,1 MPLP.
Descripción. Espora trilete de contorno triangular con lados fuertemente cóncavos y ápices redondeados. Marca trilete en general abierta, con rayos rectos que no se extienden más de 3/4 del radio de la espora. Exina de aprox. 2 µm de espesor. La escultura consiste de conos redondeados no mayores a 1 µm de alto y de aprox. 1 µm de base (isodiamétricos); también se presentan en menor proporción, verrugas y báculas de hasta 1 µm de alto; los elementos esculturales están densa y regularmente distribuidos sobre toda la superficie de la espora, pero en general se reducen en cantidad y tamaños en la cara proximal.
Dimensiones. Diámetro ecuatorial máximo, 41-57 µm (8 ejemplares).
Observaciones. Playford et al. (1982) ampliaron la diagnosis original de la especie incluyendo las características morfológicas de L . novicus Singh, 1964. En este trabajo se sigue la detallada revisión de esta especie de acuerdo con aquellos autores, dentro de la cual, los ejemplares de Cacheuta se circunscriben ampliamente.
Afinidad botánica. Filicopsidas (Balme, 1995; de Jersey y Raine, 1990).
Distribución. Argentina: Triásico de las formaciones Potrerillos y Cacheuta (perfiles I y II, este trabajo) y Triásico Superior de la Formación Santa Clara Arriba (Zavattieri, 1992), cuenca Cuyana; como L . cf. bahuiniae ha sido registrada en la Formación Rancho de Lata, alta Cordillera de San Juan (Ottone et al. , 1992). Australia: Triásico Inferior a Triásico Superior de las cuencas del este y oeste de Australia (véanse detalles de distribución en Playford et al. , 1982).
Género Matonisporites Couper 1958
Especie tipo. Matonisporites phlebopteroides Couper 1958.
Matonisporites equiexinus Couper 1958. Figura 3.P
Material estudiado. 7074F: 47,2/110,2 (foto); 55,8/ 102,1. 7084A: 34,9/99,1 MPLP.
Descripción. Espora trilete de contorno ecuatorial triangular, lados rectos a algo convexos y ápices redondeados. Marca trilete abierta de rayos rectos a levemente curvados que llegan hasta el ecuador, con labios finos de aprox. 1 µm de espesor. Exina levigada de aprox. 2 µm de espesor, poco más engrosada en los ápices; en la cara proximal presenta un leve engrosamiento que bordea a la marca trilete alcanzando 3-4 µm de ancho.
Dimensiones. Diámetro ecuatorial máximo, 64-70 µm (3 ejemplares).
Observaciones. Estos ejemplares tienen la exina un poco más fina (2 µm) que el material de Couper (1958) (2,5-3,5 µm) y el engrosamiento proximal no está tan desarrollado como se puede observar en las figuras (20: 13-14) de dicho autor. Se ajusta en forma estrecha al material de Horowitz (1970) asignado a dicha especie, quien describe un engrosamiento proximal (margo) de 7 µm de ancho hacia los ápices y 10 µm hacia el centro de la espora, valores un poco mayores que en este material. Dettmann (1963) enmienda el género Dictyophyllidites para incluir: "esporas triletes con labios membranosos y elevados y márgenes de la lesura engrosados, exina fina o gruesa, lisa o con un patrón fino", por lo cual transfiere M . equiexinus Couper, 1958 a dicho género. Aquí se prefiere no considerar la citada enmienda y mantener la especie en el género Matonisporites por los engrosamientos de la exina en los ápices, característica no mencionada en el género Dictyophyllidites .
Afinidad botánica. Filicopsida, Matoniaceae (Balme, 1995).
Distribución. Argentina: Triásico de las formaciones Potrerillos y Cacheuta (perfil I, este trabajo), cuenca Cuyana. Europa: Jurásico-Cretácico de Gran Bretaña (Couper, 1958). Asia: Jurásico de Israel (Horowitz, 1970).
Género Neoraistrickia Potonié 1956
Especie tipo. Neoraistrickia truncata (Cookson) Potonié 1956.
Observaciones. De Jersey (1971a, p. 6-7) discute la distinción entre Neoraistrickia y Ceratosporites , remarcando que la diferencia entre ellos está dada por la exina proximal lisa en el segundo. Sin embargo, su especie C . helidonensis de Jersey tiene ornamentación ligera a fuertemente reducida en la cara proximal. Varias especies de Neoraistrickia tienen, a su vez, fuerte reducción de los elementos ornamentales en la cara proximal. En ambos géneros se han descripto especies que presentan escultura con predominio de báculas y algunas muestran en menores proporciones, transiciones hacia espinas. La forma general de este tipo de esporas también varía, desde fuertemente triangular en algunas especies de Neoraistrickia a subcircular o subtriangular redondeada en ambos géneros. Por su parte Bharadwaj y Kumar (1972, p. 214) enmendaron el género Neoraistrickia , para incluir esporas con ámbito triangular con lados convexos, exina distalmente ornamentada con procesos baculados pero lisa en la cara proximal. La alternativa para elegir un género adecuado y satisfactorio para acomodar todas estas variables morfológicas es, consecuentemente, arbitrario. Los ejemplares que se describen a continuación se asignan a este último género de acuerdo al criterio de de Jersey (1971a, p. 6-7) por la presencia de individuos con cara proximal ornamentada. McKellar (1974, p. 9) y de Jersey (1972, p. 6-7), discuten también la sinonimia y distinción del género Neoraistrickia con otros géneros que presentan morfologías similares validando este género. Horriditriletes Bharadwaj y Salujha (1964) conforma también al amplio concepto de Neoraistrickia pero consideramos la prioridad de este último.
Neoraistrickia sp. Figura 4.M
Material estudiado. 7141B: 32,4/105,8 (foto); 40,5/ 109,8; 42,4/108,4; 47,9/111 MPLP.
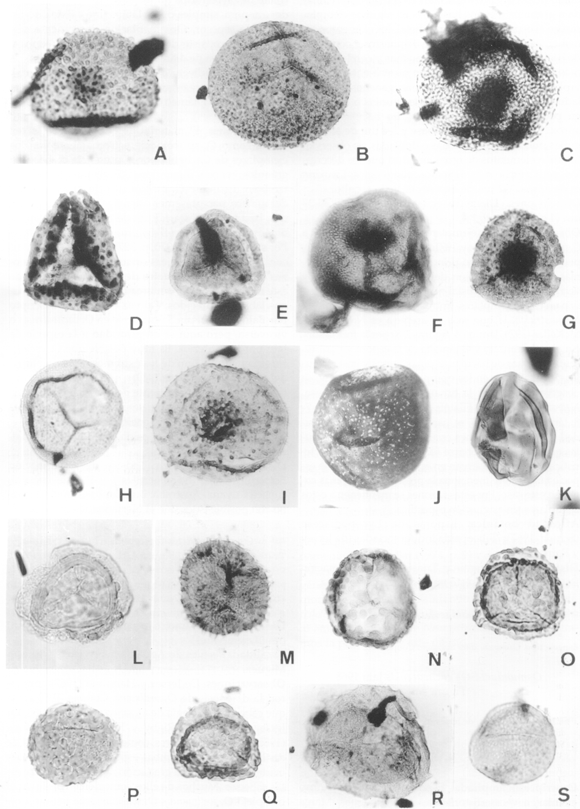
Figura 4. A, Clavatisporites sp. 4423I: 44,0/111,6 MPLP (x 900). B, Osmundacidites cacheutensis Jain. 4423I: 32/112,8 MPLP (x 900). C, Rugulatisporites permixtus Playford, en Playford, Rigby y Archibald. 7074C: 49,9/97,2 MPLP. Vista distal / Distal view (x 1000). D, Raistrickia sp. 7075D: 39,8/107,3 MPLP (x 800). E, Densoisporites complicatus Balme. 4423I: 38,9/92,5 MPLP (x 550). F, Cadargasporites cuyanensis Azcuy y Longobucco. 7074C: 37,0/100,4 MPLP; vista ecuatorial-proximal mostrando la marca trilete y el área de contacto bordeada por pliegues de la exina / Equatorial-proximal view view showing the trilete scar and the contact area bordered by exine foldings (x 900). G, Cirratriradites sp. 1 (en Zavattieri, 1990b). 7141B: 40,4/112,3 MPLP (x 700). H, Osmundacidites wellmanii Couper. 4423I: 39,0/112,3 MPLP (x 750). I, Apiculatisporis sp. cf. A . globosus (Leschik) Playford y Dettmann. 4423I: 29,8/106,9 MPLP; Foco medio / Medium focus (x 900). J, Foveosporites mimosae de Jersey y Hamilton. 7070B: 37,1/95,2 MPLP; Vista ecuatorial / Equatorial view (x 550). K, Striatella seebergensis Mädler. 7147D: 36/100,2 MPLP; Vista ecuatorial / Equatorial view (x 550). L, Espora trilete gen. et sp. indet. 4423I: 29,9/103 MPLP. Vista proximal / Proximal view (x 700). M, Neoraistrickia sp. 7141B: 32,4/105,8 MPLP (x 750). N, Cadargasporites verrucosus Reiser y Williams. 4423I: 47,6/102,4 MPLP (x 700). O, Antulsporites sp. A (en McKellar, 1974). 4423I: 30,0/113,4 MPLP (x 700). P, Thymospora ipsviciencis (de Jersey) Jain. 7074C: 40/94,3 MPLP (x 650). Q, Antulsporites sp. A (en McKellar, 1974). 4423I: 34,1/98,1 MPLP (x 600). R, Aratrisporites parvispinosus (Leschik) Playford. 4423I: 29/99,4 MPLP (x 600). S, Punctatosporites walkomii de Jersey. 7074F: 39,3/107 MPLP (x 650).
Descripción. Espora trilete de contorno ecuatorial subcircular a subtriangular redondeado. Marca trilete poco conspicua o con finos labios; rayos sinuosos que no llegan al ecuador, alcanzando aprox. ¾ o más de la longitud del radio de la espora. Áreas interradiales de la cara proximal con ornamentación reducida a gránulos o conos hasta lisa en algunos ejemplares; en el ecuador y en la cara distal, los elementos ornamentales están densamente dispuestos y consisten de báculas, algunas con bases anchas y con los extremos romos o redondeados, con puntas ensanchadas y planas o bifurcadas que forman ganchos a ambos lados; otras se angostan hacia las puntas o terminan en pequeñas sétulas o espinas; las báculas tienen aprox. 1 µm de ancho en la base y 3-4 µm de alto; algunas pueden estar oblicuas o dobladas. Se observan también elementos de igual longitud que los descriptos, pero mucho más delgados, como pelos. La exina es de aprox. 1-2 µm de espesor entre la escultura.
Dimensiones. Diámetro ecuatorial máximo, 35-37 µm (4 ejemplares).
Observaciones. Los ejemplares descriptos se parecen a C . helidonensis de Jersey 1971a (lámina 1, figuras 8 -11), pero esta especie tiene principalmente espinas y además, presenta reducción del ornamento con conos y verrugas en la cara distal, caracteres que no se observan en estos ejemplares. Entre las especies asignadas a Neoraistrickia , la que más se asemeja es N . suratensis McKellar (1974, lámina 4, figuras 2-8) por la forma subcircular del ámbito y el tamaño de la espora y por presentar además de báculas, espinas, conos y hasta rúgulas. Sin embargo, en los ejemplares de Cacheuta no se observan rúgulas y la ornamentación es más densa. Neoraistrickia densata Filatoff (1975, lámina 6, figuras 7- 8; lámina 7, figura 1), se parece también por la densidad de las proyecciones esculturales, pero tiene báculas que pueden fusionarse formando rúgulas, característica que no se observan en este material; en dicha especie tampoco se menciona la presencia de espinas como en este caso. Por estas razones se mantienen a estos ejemplares sin asignación específica.
Afinidad botánica. Esporas de Lycopodiaceae (Schulz, 1967); ?Filicopsida indiferenciadas (de Jersey y Raine, 1990).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfil II, este trabajo), cuenca Cuyana.
Género Osmundacidites Couper 1953
Especie tipo. Osmundacidites wellmanii Couper 1953.
Osmundacidites cacheutensis Jain 1968. Figura 4.B
1964. Osmundacidites sp. de Jersey, p. 6, lámina 1, figura 6.
Principal material estudiado. 4423I: 32,0/112,8 (foto); 34,0/97,7; 44,3/102,2; 45,7/96,2. 7074C: 37,8/ 95,6. 7141G: 50,4/109 MPLP.
Descripción ampliada. Espora trilete de contorno circular, a veces distorsionado por pliegues. Marca trilete de rayos rectos que se extienden a ¾ del radio de la espora, simples o con labios finos. Exina de 1 a 2 µm de espesor. Ornamentación densamente dispuesta que consiste de pequeños gránulos, conos y báculas de 1 µm de diámetro y de 1 a 2 µm de alto; las báculas pueden llegar a clavas. En la cara proximal, la escultura parece estar algo reducida.
Dimensiones. Diámetro ecuatorial máximo, 44-76 µm (6 ejemplares).
Observaciones. El material descripto por de Jersey (1964) como O . sp. coincide estrechamente con los ejemplares de Cacheuta por la presencia de báculas, gránulos, conos y dimensiones (56-89 µm). Jain (1968) también distingue O . cacheutensis de otras especies del género por presentar además de gránulos y conos, báculas de hasta 1,5 µm de alto aunque con tamaños algo menores (47-56 µm) que el de de Jersey. Por su parte, Balme (1970, p. 327) describe O . senectus con iguales características ornamentales y dimensiones (57-72 µm de diámetro de la espora) que las de este material y con una marca trilete bordeada por finos labios oscurecidos. Tal vez el material descripto por Balme (1970) pueda incluirse en O . cacheutensis o viceversa, pero aquí se da prioridad a la especie de Jain por ser anterior. Es muy posible que O . cacheutensis constituya uno de los extremos de variabilidad del complejo O . wellmanii - O . senectus pero hasta contar con la cantidad suficiente de material para ver la transición de la escultura, se prefiere mantener esta asignación por la razón mencionada.
Afinidad botánica. Filicopsida, Osmundaceae (Balme, 1970; 1995).
Distribución. Argentina: Triásico de la Formación Potrerillos (Jain, 1968; perfiles I y II, este trabajo), cuenca Cuyana. Australia: Bundamba Group, Carniano con posible extensión al Noriano (de Jersey, 1964).
Osmundacidites wellmanii Couper 1953. Figura 4.H
Principal material estudiado. 4423I: 39,0/112,3 (foto); 34,7/112,8; 44,2/109,2; 41,7/102,8. 7074C: 34,1/114,0 MPLP.
Dimensiones. Diámetro ecuatorial máximo, 43-70 µm (9 ejemplares).
Localidad. Perfiles de Agua de las Avispas y Puesto Miguez.
Observaciones. De Jersey y Hamilton (1967) ampliaron la diagnosis de O . wellmanii Couper, 1953 en la que cabe la morfología de O . senectus Balme, 1963. De Jersey (1979, p. 8), Rigby y Hekel (1977), Stevens (1981) entre otros autores, comentaron la dificultad de distinguir y separar ambas especies, lo cual pone en evidencia la continua gradación morfológica entre estas especies. Muchos de ellos coinciden en designar el complejo O . wellmanii - O . senectus para incluir a esporas dispersas que abarquen todas las transiciones entre ellas. Si bien se coincide con este criterio, se incluye dentro de O . wellmanii Couper toda la variabilidad de formas abarcadas por ambas especies y se las asigna a esta última por prioridad, teniendo en cuenta además, que su distinción específica tiene poca o nula aplicación cronoestratigráfica o paleoambiental.
Afinidad botánica. Filicopsida, Osmundaceae (Balme, 1970; 1995).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfiles I y II, este trabajo), cuenca Cuyana. Ampliamente distribuida en sedimentos del Triásico y Jurásico de la Argentina y del Gondwana.
Género Raistrickia (Schopf, Wilson y Bentall) Potonié y Kremp 1954
Especie tipo. Raistrickia grovensis Schopf, en Schopf, Wilson y Bentall 1944.
R aistrickia sp. Figura 4.D
Material estudiado. 7075 D: 39,8/107,3 (foto) MPLP.
Descripción. Espora trilete de contorno triangular, lados rectos o algo cóncavos o convexos y ápices redondeados. Marca trilete simple y abierta llegando al ecuador. Exina de aprox. 1 µm de espesor. Ornamentación poco densa (2-3 µm entre los elementos) en ambas caras; consiste principalmente de báculas y conos en menor proporción. Las primeras son de formas y tamaños variables, de aprox. 2 µm de alto y de 1-2 µm de base, que varían desde formas finas y extremos redondeados hasta tubulares gruesas con ápices truncos. Los conos son de aprox. 0,5-2 µm de base y de 0,5-2,5 µm de alto, con cimas puntiagudas en las formas más pequeñas y redondas en las mayores.
Dimensiones. Diámetro ecuatorial máximo: 50 µm (1 ejemplar).
Observaciones. El único ejemplar encontrado no se pudo asignar a ninguna de las especies del género, aunque es similar a R . ramosa (Balme y Hennelly) Rigby y Hekel, 1977. Sin embargo, esta última presenta dimensiones menores (26-45 µm en la descripción original, Balme y Hennelly, 1956; 18-46 µm en Rigby y Hekel, 1977). El material de estos últimos autores tiene mayor densidad de la escultura y en el de Balme y Hennelly los elementos ornamentales son más grandes (2-5 µm de base y 2-7 µm de alto) y algunos son bifurcados. Neoraistrickia truncata (Cookson) Potonié, 1956 (en Filatoff, 1975) es similar a este de Cacheuta, pero con menores dimensiones ecuatoriales y no posee conos como en este ejemplar.
Afinidad botánica. Filicopsida (Balme, 1995).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfil I, este trabajo), cuenca Cuyana.
Género Rugulatisporites Pflug en Thompson y Pflug, 1953
Especie tipo. Rugulatisporites quintus Thompson y Pflug 1953.
Rugulatisporites cf. trisinus de Jersey y Hamilton 1967. Figuras 3.K a-b
Material estudiado. 7074F: 38,9/108,1 (foto) MPLP.
Descripción. Espora trilete de contorno triangular, con lados rectos y ápices redondeados. Marca trilete recta o levemente sinuosa con labios finos; los extremos de los rayos terminan en el ecuador con curvatura imperfecta. Exina de aprox. 1 µm de espesor, lisa en la cara proximal y ornamentada con muros en la cara distal. Muros bajos, de 2-6 µm de largo, 1 µm de ancho y separados entre sí 1-2 µm, distribuidos irregularmente formando rúgulas de recorrido sinuoso a laberíntico y más difusas sobre el ecuador.
Dimensiones. Diámetro ecuatorial máximo, 36 µm (1 ejemplar).
Observaciones. Rugulatisporites trisinus es semejante a este ejemplar por la forma triangular de la espora y la disposición de los muros en la cara distal. Sin embargo, difiere por tener rúgulas más anchas (1,5-3 µm), menos espaciadas (1 µm) y presentes también en la zona ecuatorial. Muestra además rayos con finos labios y curvatura imperfecta, características que no se mencionan en la especie. Rugulatisporites sp. A (en McKellar, 1974) también es comparable, pero se diferencia por su contorno circular a subcircular, menores dimensiones (25-29 µm) y muros más anchos y altos (1,5 µm en ambos casos) y en algunos casos bifurcados. Aff. Rugulatisporites (en McKellar, 1974) es similar a este espécimen ya que presenta curvatura en los extremos de los rayos y escultura en la cara distal, pero difiere en que se encuentra dispuesta radialmente y se proyecta hacia la zona subecuatorial de la cara proximal en asociación con la curvatura de los rayos.
Afinidad botánica. Filicales indiferenciadas (de Jersey y Raine, 1990).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfil I, este trabajo), cuenca Cuyana. Para R . trisinus : Australia: Triásico Medio (Fm. Moolayember) (de Jersey y Hamilton, 1967).
Género Striatella Mädler emend . Filatoff y Price 1988
Especie tipo (por designación original). Striatella seebergensis Mädler 1964.
Observaciones. En este trabajo, se sigue el criterio y análisis señalados por Zavattieri y Volkheimer (2003), respecto del uso de Striatella seebergensis en lugar de Duplexisporites problematicus .
Striatella seebergensis Mädler 1964. Figura 4.K
1965. Duplexisporites gyratus Playford y Dettmann, p. 141-142; lámina 13, figuras 20-22.
1970. Duplexisporites problematicus (auct. non Couper) Playford y Dettmann; de Jersey, p. 8; lámina 3, figuras 1-3.
1977. Asseretospora gyrata (Playford y Dettmann) Schuurman, p. 198; pl. 10, figs. 5-6.
Material estudiado. 7147D: 36/100,2 (foto) MPLP.
Dimensiones. Diámetro ecuatorial máximo: 61 µm (1 ejemplar).
Observaciones. Por la posición ecuatorial del ejemplar, no se observó el muro ecuatorial que forma el cíngulo, la verruga ubicada en la región polar distal, ni tampoco los muros de la cara proximal como mencionan Playford et al. (1982). Las dimensiones de los elementos esculturales y el tamaño del ejemplar se ajustan a los descriptos para la especie, así como el resto de los rasgos morfológicos y aspecto general ampliamente ilustrado en la literatura.
Afinidad botánica. Filicopsida, Pteridaceae (de Jersey y Raine, 1990).
Distribución. Argentina: Triásico de la Formación Cacheuta (perfil II, este trabajo), cuenca Cuyana; formaciones Paso Flores y Comallo, Macizo Nordpatagónico (Zavattieri, 1997; Zavattieri et al. , 1994); Jurásico Inferior de la Formación Nestares, norte de Patagonia (Zavattieri y Volkheimer, 2003). En Australia y Nueva Zelanda está ampliamente distribuida desde el Triásico Medio a Jurásico Medio; en Antártida en el Triásico Medio y Superior; en el Hemisferio Norte se registra con igual distribución cronológica en Rusia, oeste de Europa y China y en el Jurásico Inferior de Afganistán (de Jersey y Raine, 1990).
Género Uvaesporites Döring, 1965 emend . Helby, en de Jersey 1971b
Especie tipo (por designación original). Uvaesporites glomeratus Döring 1965.
Uvaesporites verrucosus (de Jersey) Helby en de Jersey 1971b. Figura 3.E
Véase sinonimia en de Jersey, 1971b, p. 5-6.
Material estudiado. 4423I: 32/95,2 (foto); 45,5/105,5; 32,1/104,4; 46,5/109,7; 43,9/103,8; 46,4/112; 36,4/106,1; 39/93,6. 7140C: 40,9/106 MPLP.
Dimensiones. Diámetro ecuatorial máximo, 35-43 µm (9 ejemplares).
Observaciones. Dentro de esta asignación se incluyen ejemplares que presentan exinas con gemas, clavas y verrugas densamente dispuestas en la cara distal de hasta 6 µm de alto y de 1-4 µm de base. De Jersey (1971b, p. 5-6) y de Jersey y Raine (1990, p. 25) han discutido los rasgos estructurales y morfológicos y la comparación de este género con otras esporas dispersas afines. Estos autores caracterizan a la especie por presentar una continua variación desde ejemplares con proyecciones verrucosas y con clavas discretas, hasta aquéllos con elementos esculturales ensanchados y más densamente dispuestos.
Afinidad botánica. Lycopsida (Selaginellaceae) (Balme, 1995).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfil II, este trabajo), cuenca Cuyana. Australia y Nueva Zelanda: Triásico Medio a Jurásico Inferior (véase detalle de distribución en de Jersey, 1971b y de Jersey y Raine, 1990).
Espora trilete gen. et sp. indet. Figura 4.L
Material estudiado. 4423I: 29,9/103 (foto); 34,1/98,1; 30,9/102,2; 30/95,3; 30,6/93,7; 34,5/113,4; 40/109,3; 40,7/110,7; 32,1/99,3 MPLP.
Descripción. Espora trilete cingulada (o ?zonada), de contorno subtriangular a subcircular e irregular por la proyección de los elementos esculturales. Marca trilete con rayos sinuosos que llegan casi al ecuador, con labios que acompañan a los rayos de aprox. 1,5 µm de espesor. Cara proximal lisa; áreas de contacto poco delimitadas. Escultura dispuesta desde el ecuador hacia la cara distal, compuesta por verrugas no mayores a 1,5 µm de alto y 2-5 µm de base, de contornos irregulares y en algunos ejemplares se presentan rúgulas sobre la superficie distal. Un anillo ?proximal corre paralelo al borde de la intexina, de 3-4 µm de espesor. La sexina forma un cíngulo o ?zona ecuatorial que tiene aspecto esponjoso a liso dependiendo de la preservación de los ejemplares.
Dimensiones. Diámetro ecuatorial máximo, 36-47 µm; diámetro máximo del anillo, 24-40 µm (9 ejemplares).
Observaciones. Este material presenta caracteres morfológicos que no están circunscriptos en ningún género ni especie conocidos.
Distribución. Argentina: Formación Potrerillos (perfil II, este trabajo), cuenca Cuyana.
Esporas monoletes
Género Aratrisporites (Leschik) Playford y Dettmann 1965
Especie tipo. Aratrisporites parvispinosus (Leschik) Playford 1965.
Aratrisporites parvispinosus (Leschik) Playford 1965. Figura 4.R
Material estudiado. 4423I: 29,0/99,4 (foto); 31,1/ 100,4; 34,7/96,2; 37/99,1. 7074C: 43/102,1; 51,3/98,6 MPLP.
Dimensiones. Exoexina, 65-94 µm x 47-63 µm; intexina, 43-53 µm x 29-40 µm; espinas, 2-5 µm de alto y 2- 3 µm de diámetro basal (6 ejemplares).
Observaciones. Las dimensiones de los ejemplares de Cacheuta son algo mayores a los dados para A . parvispinosus (Leschik) Playford, pero coincide en el resto de las características morfológicas. También se asemeja a A . flexibilis Playford y Dettmann 1965, aunque esta última se diferencia por tener dimensiones menores, intexina más grande en relación con la exoexina y espinas de bases menores (1-2 µm). Aratrisporites tenuispinosus Playford, 1965 tiene un diámetro ecuatorial más reducido, una intexina mayor en relación con la exoexina, mayor densidad de espinas y estas últimas, en general, son más pequeñas (bases de 0,5-2 µm).
Afinidad botánica. Lycopsida (Pleuromeiaceae) (Balme, 1995).
Distribución. Argentina: Triásico de la Formación Potrerillos (perfiles I y II, este trabajo), cuenca Cuyana. Tasmania: Triásico de Brady Formation (Playford, 1965). Suiza: Keuper Medio (Leschik, 1955).
Género Thymospora Wilson y Venkatachala emend . Alpern y Doubinger 1973
Especie tipo. Thymospora thiessenii (Kosanke) Wilson y Venkatachala 1963.
Thymospora ipsviciencis (de Jersey) Jain 1968 Figura 4.P
1962. Verrucososporites ipsviciensis de Jersey, p. 7, figuras 8-10.
1965. Polypodiisporites ipsviciensis (de Jersey) Playford y Dettmann, p. 150-151; lámina 15, figuras. 39-40.
1968. Thymospora ipsviciensis (de Jersey) Jain, p. 15, lámina 2, figuras 29-30.
Principal material estudiado. 7074C: 40/94,3 (foto); 43,9/108,4; 48,5/96,3; 50,5/104,8; 55,6/101,5. 7075E: 55,2/100. 7083B: 44,0/110,4. 7084E: 38,3/99. 7148C: 51,4/105 MPLP.
Dimensiones. Largo x ancho, 26-35 µm x 20-30 µm (9 ejemplares).
Observaciones. El género Polypodiisporites Potonié emend . Khan y Martin, 1971 posee "elementos esculturales apretadamente dispuestos, con intersticios estrechos o interconectados para formar un regular retículo negativo". En cambio, Thymospora incluye esporas con escultura verrucada-rugulada, la cual no forma un retículo negativo (Foster, 1979). En estos ejemplares, las "vermículas" entre los muros o rúgulas no están interconectadas y no se observa que formen tal retículo negativo. En la diagnosis original de la especie tipo ( V . ipsviciensis de Jersey, 1962) se menciona la presencia de papilas que emergen de las verrugas, característica que presentan sólo algunos de los ejemplares estudiados o tienen pequeñas espinas sobre las verrugas-rúgulas.
Afinidad botánica. Filicales de tipo Hymenophyllaceae/ Polypodiaceae (Potonié, 1962). Filicales indiferenciadas (de Jersey y Raine, 1990).
Distribución. Argentina: Triásico de las formaciones Cerro de Las Cabras, Potrerillos y Cacheuta (Jain, 1968; Zavattieri, 1986, 1990b; perfiles I y II, este trabajo). Ampliamente distribuidas en palinofloras del Triásico de Australia (de Jersey, 1979) desde el límite Permo- Triásico (de Jersey, 1979; Foster, 1979) hasta Jurásico Inferior (McKellar, 1974; de Jersey y Raine, 1990).
Conclusiones
Un primer análisis sobre las afinidades botánicas de las especies halladas en estas microfloras del sur del cerro Cacheuta (véase cuadro 1) permite señalar que la diversidad de los grupos vegetales representados en los microfósiles es mayor que los que evidencian las megafloras locales (Morel, 1991 inéd .; 1994; Stipanicic et al. , 1996; Artabe y Zamuner, 1991; Morel y Povilauskas, 2002). La composición microflorística total que se conoce para las Formaciones Potrerillos y Cacheuta en estas dos secciones se presentará en la segunda parte de este estudio. Se registran en este trabajo 15 nuevas especies de esporas de briófitas y pteridófitas para la Formación Potrerillos y 7 para la Formación Cacheuta (incluidas aquéllas en las que no se alcanzó la asignación específica). De ellas, se registran por primera vez para el Triásico de Argentina las siguientes especies: Anapiculatisporites cooksonae , Antulsporites sp. A (en McKellar, 1974), Densoisporites complicatus , Foveosporites mimosae , Matonisporites equiexinus y Uvaesporites verrucosus y en nomenclatura abierta, Clavatisporites sp., Neoraistrickia sp., Raistrickia sp., Rugulatisporites cf. trisinus y una forma que no se pudo asignar a nivel genérico y específico. El género Matonisporites se describe por primera vez para el Triásico de Argentina. Las especies nuevas descriptas aportan elementos adicionales al conocimiento sobre la composición microflorística de estas formaciones y para el Triásico de la cuenca Cuyana.

Cuadro 1. Afinidades botánicas conocidas de las esporas halladas y especies de la megaflora local registrada en las Formaciones Potrerillos y Cacheuta, en las secciones Agua de las Avispas y Puesto Miguez, sur del cerro Cacheuta, Mendoza. / Known botanical affinities of the spores and species of the local megaflora recorded from the Potrerillos and Cacheuta Formations, at Agua de las Avispas and Puesto Miguez sections, south of the cerro Cacheuta, Mendoza.
Agradecimientos
Los autores agradecen al CONICET por los fondos recibidos mediante PIP 4674/96 y PIP 2905/00 para el tratamiento de las muestras en el Laboratorio del IANIGLA, CRICYT (CONICET), Mendoza, y del instrumental óptico utilizado del citado Labo_ratorio. Asimismo a las valiosas críticas de Pedro N. Stipanicic y Eduardo Morel cuyos aportes enriquecieron este trabajo.
Bibliografía
1. Alpern, B. y Doubinger, J. 1973. Microfossiles organiques du Paléozoïque; 6, Les spores. Les miospores monoletes du Paléozoïque . Paris, Commission Internationale de Microflore du Paléozoïque, Éditions C.N.R.S. 103 pp. [ Links ]
2. Archangelsky, S. y Gamerro, J.C. 1966a. Estudio palinológico de la Formación Baqueró (Cretácico), Provincia de Santa Cruz. II. Ameghiniana 4: 201-209. [ Links ]
3. Archangelsky y Gamerro, J.C. 1966b. Estudio palinológico de la Formación Baqueró (Cretácico), Provincia de Santa Cruz. III. Ameghiniana 4: 229-236. [ Links ]
4. Artabe, A.E. y Zamuner, A.B. 1991. Una nueva Equisetal del Triásico de Cacheuta, Argentina, con estructura interna preservada. Ameghiniana 28: 287-294.
Links ] Times, serif"> 5. Azcuy, C.L. y Longobucco, M.I. 1983. Palinología de las formaciones Potrerillos y Cacheuta, provincia de Mendoza, Argentina. Ameghiniana 20: 297-316. [ Links ]
6. Balme, B.E. 1957. Spores and pollen grains from the Mesozoic of Western Australia. Commonwealth Scientific and Industrial Research Organization, Coal Research Section, technical communication 25: 48 p. [ Links ]
7. Balme, B.E. 1963. Plant fossils from the Lower Triassic of Western Australia. Palaeontology 6: 12-40. [ Links ]
8. Balme, B.E. 1970. Palynology of Permian and Triassic strata in the Salt Range and Surghar Range, West Pakistan. En: Kumel, B. y Teichert, C. (eds.), Stratigraphic boundary problems: Permian and Triassic of West Pakistan. University of Kansas, Department of Geology, Special Publication 4: 306-453. [ Links ]
9. Balme, B.E. 1995. Fossil in situ spores and pollen grains: an annotated catalogue. Review of Palaeobotany and Palynology 87: 81-323.
Links ] Times, serif"> 10. Balme, B.E. y Hennelly, J.P.F. 1956. Trilete sporomorphs from Australian Permian sediments. Australian Journal of Botany 4: 240-260. [ Links ]
11. Bharadwaj, D.C. y Kumar, P. 1972. On the status of some miospore genera from the Mesozoic Era. The Palaeobotanist 19: 214- 224.
Links ] Times, serif"> 12. Bharadwaj, D.C. y Salujha, S.K. 1964. Sporological study of seam VIII in Raniganj Coalfield, Bihar (India) - Part 1. Description of sporae dispersae . The Palaeobotanist 12: 181-215. [ Links ]
13. Couper, R.A. 1953. Upper Mesozoic and Caenozoic spores and pollen grains from New Zealand. New Zealand Geological Survey Paleontological Bulletin , 22: 1-77. [ Links ]
14. Couper, R.A. 1958. British Mesozoic microspores and pollen grains. A systematic and stratigraphic study. Palaeontographica B, 103: 75-179. [ Links ]
15. Danzé-Corsin, P. y Laveine, J.P. 1963. Microflore. En: P. Briche, P. Danzé-Corsin y J.P. Laveine, Flore infraliassique du Boulonnais. Société Géologique du Nord Mémoire 13: 57-143. [ Links ]
16. De Jersey, N.J. 1962. Triassic spores and pollen grains from the Ipswich coalfield. Geological Survey of Queensland Publication 307: 1-18. [ Links ]
17. De Jersey, N.J. 1964. Triassic spores and pollen grains from the Bundamba Group. Geological Survey of Queensland Publication 321: 1-21. [ Links ]
18. De Jersey, N.J. 1968. Triassic spores and pollen grains from the Clematis Sandstone. Geological Survey of Queensland Publication 338 , Palaeontological Paper 14: 1-44. [ Links ]
19. De Jersey, N.J. 1970. Triassic miospore from the Blackstone Formation, Aberdare Conglomerate and Raceview Formation. Geological Survey of Queensland publication 348 , Palaeontological Pape r 22: 1-41. [ Links ]
20. De Jersey, N.J. 1971a. Early Jurassic miospores from the Helidon Sandstone. Geological Survey of Queensland Publication 351 , Palaeontological Paper 25: 1-49. [ Links ]
21. De Jersey, N.J. 1971b. Triassic miospores from the Tivoli Formation and Kholo Sub-Group. Geological Survey of Queensland Publication 353 , Palaeontological Paper 28: 1-40. [ Links ]
22. De Jersey, N.J. 1972. Triassic miospores from the Esk Beds. Geological Survey of Queensland Publication 357 , Palaeontological Paper 32: 1-40. [ Links ]
23. De Jersey, N.J. 1974. Palynology and age of the Callide Coal Measures. Queensland Government Mining Journal 75: 249-252. [ Links ]
24. De Jersey, N.J. 1979. Palynology for the Permian-Triassic Transition in the Western Bowen Basin. Geological Survey of Queensland Publication 353 , Palaeontological Paper 23: 1-40. [ Links ]
25. De Jersey, N.J. y Hamilton, M. 1967. Triassic spores and pollen grains from the Moolayember Formation. Geological Survey of Queensland Publication 336 , Palaeontological Paper 10: 1-61. [ Links ]
26. De Jersey, N.J. y Raine, J.I. 1990. Triassic and earliest Jurassic miospores from the Murihiku Supergroup, New Zealand. New Zealand Geological Survey Paleontological Bulletin 62: 1-164. [ Links ]
27. Dettmann, M.E. 1963. Upper Mesozoic microfloras from South- Eastern Australia. Proceedings of the Royal Society of Victoria 77: 1-148. [ Links ]
28. Döring, H. 1965. Die sporenpaläontologische Gliederung des Wealden in Westmecklenburg (Strucktur Werle). Geologie 14: 1-118. [ Links ]
29. Farabee, M.J., Taylor , E.L. y Taylor, T.N. 1989. Pollen and spore assemblages from the Falla Formation (Upper Triassic), Central Transantarctic Mountains, Antarctica. Review of Palaeobotany and Palynology 61: 101-138. [ Links ]
30. Farabee, M.J., Taylor, E.L. y Taylor, T.N. 1990. Correlation and Triassic palynomorph assemblages from the central Transantarctic Mountains, Atnarctica. Review of Palaeobotany an Palynology 65: 257-265. [ Links ]
31. Filatoff, J. 1975. Jurassic palynology of the Perth Basin, Western Australia. Palaeontographica B, 154: 1-113. [ Links ]
32. Filatoff, J. y Price, P.L. 1988. A pteridacean spore lineage in the Australian Mesozoic. Association of Australasian Palaeontologists, Memoir 5: 89-124. [ Links ]
33. Foster, C.B. 1979. Permian plant microfossils of the Blair Athol Coal Measures, Baralaba Coal Measures, and basal Rewan Formation of Queensland. Geological Survey of Queensland Publication 372 , Palaeontological Paper 45: 244 pp. [ Links ]
34. Hiltmann, W. 1967. Über die Sporenführung des Kernprofils der Bohrung Contern FG11 (Unterer Lias, Luxemburg). Service Géologique de Luxemburg Publication 17: 137-206. [ Links ]
35. Horowitz, A. 1970. Jurassic microflora from the Northern Negev, Israel. Israel Journal of Earth Sciences 19: 153-183. [ Links ]
36. Jain, R.K. 1968. Middle Triassic pollen grains and spores from Minas de Petróleo Beds of Cacheuta Formation (Upper Gondwana), Argentina. Palaeontographica B, 122: 1-47. [ Links ]
37. Khan, A.M. y Martin, A.R.H. 1971. A note on genus Polypodiisporites R. Potonié. Pollen et Spores 13: 475-480. [ Links ]
38. Kedves, M. y Simoncsics, P. 1964. Spores nouvelles extraites de minerai manganese Jurassique de la région d'Urkut (Hongrie). Pollen et Spores 6: 605-610. [ Links ]
39. Kurtz, F. 1921. Atlas de Plantas Fósiles de la Repúbica Argentina. Actas de la Academia Nacional de Ciencias , Córdoba 7: 129-153. [ Links ]
40. Leschik, G. 1955. Die Keuperflora von Neuewelt bei Basel. II. Die Iso- und Mikrosporen. Schweizerische Paläontologische Abhandlungen 72:1-70. [ Links ]
41. Llambías. E.J. y Stipanicic, P.N. 2002. Grupo Choiyoi. En: P.N. Stipanicic y C.A. Marsicano (eds.), Léxico Estratigráfico de la Argentina, Volumen VIII: Triásico , Asociación Geológica Argentina, Serie "B" (Didáctica y Complementaria) 26: 88-91. [ Links ]
42. Mädler, K. 1964. Bemerkenswerte Sporenformen aus dem Keuper und unteren Lias: Fortschr. Geologischen Rheinland und Westfalen 12: 169-200. [ Links ]
43. McKellar, J.L. 1974. Jurassic miospores from the upper Evergreen Formation, Hutton Sandstone and basal Injune Creek Group, northeastern Surat Basin. Geological Survey of Queensland Publication 361 , Palaeontological Paper 35: 89 pp. [ Links ]
44. Morel, E. M. 1991. [ Estudio paleoflorístico y paleoambiental de la secuencia triásica en el área de Cacheuta (provincia de Mendoza) . Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de la Plata: 368 pp. Inédita] [ Links ].
45. Morel, E. M. 1994. El Triásico del cerro Cacheuta, Mendoza (Argentina). Parte I: Geología, contenido paleoflorístico y cronoestratigrafía. Ameghiniana 31: 161-176. [ Links ]
46. Morel, E.M. y Povilauskas, L. 2002. Adenda a la flora triásica de la Formación Potrerillos en el cerro Cacheuta, provincia de Mendoza, Argentina. Ameghiniana 39: 501-503. [ Links ]
47. Morel, E.M., Stipanicic, P.N. y Zúñiga, A. 2002. Formación Cacheuta. En: P.N. Stipanicic y C.A. Marsicano (eds.), Léxico Estratigráfico de la Argentina, Volumen VIII: Triásico . Asociación Geológica Argentina, Serie "B" (Didáctica y Complementaria) 26: 50-52. [ Links ]
48. Norris, G. 1965. Triassic and Jurassic miospores and acritarchs from the Beacon and Ferrar Groups, Victoria Land, Antarctica. New Zealand Journal of Geology and Geophysics 8: 236-277. [ Links ]
49. Orlando, H.A. 1954. Acerca de la presencia de esporomorfos fósiles en los "Estratos con Estheria " del Triásico de Cacheuta (Mendoza). Notas del Museo de La Plata (Paleontología) 17: 147- 156. [ Links ]
50. Orlowska-Zwolinska, T. 1971. On several stratigraphically important species of sporomorphs occurring in the Keuper of Poland. Acta Society Botany of Poland 40: 636-651. [ Links ]
51. Ottone, E.G., Alvarez, P.P. y Benoit, S.V. 1992. Late triassic plant microfossils from the Rancho de Lata Formation, Main Cordillera, Argentina. Micropaleontology 38: 261-278. [ Links ]
52. Playford, G. 1965. Plant microfossils from Triassic sediments near Poatina, Tasmania. Journal of the Geological Society of Australi a 12: 173-210. [ Links ]
53. Playford, G. y Dettmann, M.E. 1965. Rhaeto-Liassic plant microfossils from the Leigh Creek Coal Measures, South Australia. Senckenbergiana Lethaea 46: 127-181. [ Links ]
54. Playford, G., Rigby, J.F. y Archibald, D.C. 1982. A Middle Triassic flora from the Moolayember Formation, Bowen Basin, Queensland. Geological Survey of Queensland 380: 1-52. [ Links ]
55. Potonié, R. 1956. Synopsis der Gattungen der Sporae Dispersae. I. Teil: Sporites . Geologischen Jahrbuch 23: 103 pp. [ Links ]
56. Potonié, R. 1962. Sinopsis der Sporae in situ . Geologischen Jahrbuch 52: 1-204. [ Links ]
57. Potonié, R. and Kremp, G. 1954. Die Gattungen der palaozoischen Sporae Dispersae und ihre Stratigraphie. Geologische Jahrbuch 69: 111-194. [ Links ]
58. Potonié, R. y Kremp, G. 1956. Die Sporae Dispersae des Ruhrkarbons, ihre Morphographie und Stratigraphie mit Ausblicken auf Arten anderer Gebiete und Zeitabschnitte. Teil II. Palaeontographica B, 100: 1-136. [ Links ]
59. Reiser, R.F. y Williams, A.J. 1969. Palynology of the Lower Jurassic sediments of the northern Surat Basin. Geological Survey of Queensland 399 , Palaeontological Paper 15: 24 pp. [ Links ]
60. Rigby, J.F. y Hekel, H. 1977. Palynology of the Permian sequence in the Springsure Anticline, central Queensland. Geological Survey of Queensland 363 , Palaeontological Paper 37: 1-76. [ Links ]
61. Samoilovich, S.R., Mchedlishvili, N.D., Rusanova, L.Ya y Yashchurzhinskaya, A.B. 1961. Pyl'tsa i spory zapadnoy Sibiri - paleotsen [Pollen and spores of western Siberia; Jurassic to Palaocene]. Trudy, Vsesoyuznogo Neftyanogo Nauchno/issledovatel'skogo Geologo_razvedochno Instituta (VNIGRI) , 177: 1-657 [en ruso] [ Links ].
62. Schopf, J.M., Wilson, L.R. y Bentall, R. 1944. An annotated synopsis of Paleozoic fossil spores and the definition of generic groups. Report of Investigations of the Illinois State Geological Survey 91, 92 p. [ Links ]
63. Schulz, E. 1967. Sporenpaläontologische Untersuchungen rätoliassischer Schichten im Zentralteil des Germanischen Beckens. Paläontologische Abhandlungen, Paläobotanik B 2: 427- 633. [ Links ]
64. Schuurman, W.M.L. 1977. Aspects of Late Triassic palynology. 2. Palynology of the "Grès et schiste à Avicula contorta " and Argiles de Levallois" (Rhaetian) of northeastern France and southern Luxemburg. Review of Palaeobotany and Palynology 23: 159-253. [ Links ]
65. Singh, H.P. 1964. A miospores assemblage from the Permian of Iraq. Palaeontology 1: 24-265. [ Links ]
66. Stevens, J. 1981. Palynology of the Callide Basin, east-central Queensland. University of Queensland Papers, Department of Geology 9, 35 pp. [ Links ]
67. Stipanicic, P.N. 1979. El Triásico del valle del Río de Los Patos (provincia de San Juan). En: J.C.M. Turner (ed.), Geología Regional Argentina . Academia Nacional de Ciencias, Córdoba 1: 523-575. [ Links ]
68. Stipanicic, P.N. y Zavattieri, A.M. 2002. Grupo Uspallata. En: P.N. Stipanicic y C.A. Marsicano (eds.), Léxico Estratigráfico de la Argentina, Volumen VIII: Triásico . Asociación Geológica Argentina, Serie "B" (Didáctica y Complementaria) 26: 290- 294. [ Links ]
69. Stipanicic, P.N., Herbst, R. y Bonetti, M.I.R. 1996. Floras triásicas. En: P.N. Stipanicic y M. A. Hünicken (Eds.), Contribuciones a la Palaeophytologia Argentina y Revisión de la Obra Paleobotánica de Kurtz en la República Argentina. Actas de la Academia Nacional de Ciencias 11: 127-184, láms. 16 - 37. [ Links ]
70. Thompson, P.W. y Pflug, H. 1953. Pollen und Sporen des mitteleuropäischen Tertiärs. Palaeontographica B, 94:1-138. [ Links ]
71. Volkheimer, W. y Melendi, D.L. 1976. Palinomorfos como fósiles guía. Tercera parte: Técnicas de laboratorio palinológico. Revista Minera de Geología y Mineralogía 34: 119-130. [ Links ]
72. Wilson, L.R. y Venkatachala, B.S. 1963. Thymospora , a new name for Verrucososporites . Oklahoma Geology Notes 23: 75 -79. [ Links ]
73. Zavattieri, A.M. 1986. Estudio palinológico de la Formación Potrerillos (Triásico) en su localidad tipo, Cuenca Cuyana (provincia de Mendoza, Argentina). Parte 1. Esporas triletes y monoletes. Revista Española de Micropaleontología 18: 247-294. [ Links ]
74. Zavattieri, A.M. 1990a. Palinología de la Formación Las Cabras (Triásico) en su localidad tipo, Cuenca Cuyana (provincia de Mendoza, Argentina). Parte 1. Esporas triletes. Ameghiniana 27: 107-129. [ Links ]
75. Zavattieri, A.M. 1990b. Palinología de la Formación Las Cabras (Triásico) en su localidad tipo, Cuenca Cuyana (provincia de Mendoza, Argentina). Parte 2. Esporas triletes y monoletes. Ameghiniana 27: 201-224. [ Links ]
76. Zavattieri, A.M. 1992. Reseña preliminar sobre la palinología del Triásico del área de Santa Clara, norte de la provincia de Mendoza (Argentina). Asociación Paleontológica Argentina, Publicación Especial 2: 101-104. [ Links ]
77. Zavattieri, A.M. 1997. Estudio palinológico de la Formación Paso Flores (Triásico Superior), Cañadón de Pancho, Neuquén, Argentina. Datos Preliminares. 3º Reunión del Triásico del Cono Sur (La Rioja), Resúmenes: 23-26. [ Links ]
78. Zavattieri, A.M. y Rojo, L.D. (en prensa). Estudio microflorístico de las formaciones Potrerillos y Cacheuta (Triásico) en el sur del cerro Cacheuta, Mendoza, Argentina. Parte 2. Ameghiniana . [ Links ]
79. Zavattieri, A.M. y Volkheimer, W. 2003. Palynostratigraphy and paleoenvironments of Early Jurassic strata (Nestares Formation) in northern Patagonia, Argentina. Part 1. Terrestrial species. Ameghiniana 40: 545-558. [ Links ]
80. Zavattieri, A.M., Volkheimer, W. y Rosenfeld, U. 1994. Palynology and facies of the Late Triassic of Comallo (Northern Patagonia, Argentina). Zentralblatt für Geologie und Paläontologie, Teil I (1/2): 133-154. [ Links ]
Recibido: 11 de noviembre de 2003.
Aceptado: 27 de mayo de 2004.

