










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.1 Buenos Aires ene./mar. 2005
Análisis palinológico de la Formación Los Molles, Grupo Cuyo, Jurásico medio de la cuenca Neuquina, Argentina
Marcelo A. Martínez 1 , Mirta E. Quattrocchio 1 y Mercedes B. Prámparo 2
1 CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas). Universidad Nacional del Sur, Departamento de Geología, San Juan 670, 8000 Bahía Blanca, Argentina. martinez@criba.edu.ar ; mquattro@criba.edu.ar
2 CONICET. CRICYT (Centro Regional de Investigaciones Científicas y Tecnológicas), Ianigla (Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales), Unidad de Paleopalinología, C.C. 330, 5500 Mendoza, Argentina. mprampar@lab.cricyt.edu.ar
Resumen. Se describen e ilustran en este trabajo esporomorfos y paleomicroplancton identificados en la Formación Los Molles, Grupo Cuyo (Jurásico medio). Los palinomorfos fueron recuperados de 11 muestras de afloramientos pertenecientes a tres de las cinco secciones estratigráficas muestreadas (Lohan Mahuida, Puente Picún Leufú, Puesto Policía, Los Molles y Cerro Lotena) localizadas en cercanías de la ciudad de Zapala, en el ámbito de la Cuenca Neuquina, Argentina. Las microfloras estudiadas están constituídas por 50 especies de esporomorfos (20 esporas y 30 granos de polen) y 31 taxones de microplancton (9 algas verdes, 2 dinoflagelados y 20 acritarcos). Se describen ejemplares pertenecientes a los géneros Divisisporites , Callialasporites , Monosulcites y una espora género y especie indeterminadas. La sección en sistemática incluye prascinofíceas de los géneros Cymatiosphaera , Pleurozonaria y Tasmanites y entre los acritarcos especies pertenecientes a Baltisphaeridium , Filisphaeridium , Micrhystridium , Leiosphaeridia , Polygonium y Veryhachium . Se propone la nueva combinación de Cymatiosphaera volkheimerii (Quattrocchio). Se realiza la primera mención para la Cuenca Neuquina de cuatro especies de acritarcos: Filisphaeridium balmei (Sarjeant) Sarjeant y Stancliffe, F . castaninum (Valensi) Sarjeant y Stancliffe, F . densispinum (Valensi) Sarjeant y Stancliffe y Veryhachium valensii (Valensi) Downie y Sarjeant. Las microfloras analizadas se correlacionan con la Biozona de Callialasporites "complex", Sub-biozonas de Callialasporites trilobatus (Balme) Dev, Antulsporites saevus (Balme) Archangelsky y Gamerro y Klukisporites labiatus (Volkheimer) Baldoni y Archangelsky, propuestas previamente para el área de trabajo. Se sugiere una edad comprendida entre el Aaleniano tardío y Bajociano temprano para las secciones estudiadas.
Abstract. Palynological analysis of the Los Molles Formation, Cuyo Group, Middle Jurassic of the Neuquén Basin, Argentina . Sporomorphs and paleomicroplankton identified from the Middle Jurassic, Los Molles Formation, Cuyo Group, are described and illustrated in this contribution. The palynomorphs were recovered from three of the five sampled surface sections (Lohan Mahuida, Puente Picún Leufú, Puesto Policía, Los Molles and Cerro Lotena) located near Zapala city, Neuquén Basin, Argentina. The studied microfloras include 50 species of sporomorphs (20 spores and 30 pollen grains) and 31 species of paleomicroplankton (9 green algae, two dinoflagellate cysts and 20 acritarchs). Specimens belonging to the genera Divisisporites , Callialasporites , Monosulcites and one spore gen. et sp. indet. are described. Systematics include prasinophyte of the genera Cymatiosphaera , Pleurozonaria and Tasmanites . Also included are acritarchs of the genera Baltisphaeridium , Filisphaeridium , Micrhystridium , Leiosphaeridia , Polygonium and Veryhachium . The new combination Cymatiosphaera volkheimerii (Quattrocchio) is proposed. The occurrence of the acritarchs Filisphaeridium balmei (Sarjeant) Sarjeant and Stancliffe, F. castaninum (Valensi) Sarjeant and Stancliffe, F . densispinum (Valensi) Sarjeant and Stancliffe and Veryhachium valensii (Valensi) Downie and Sarjeant is mentioned for the first time in the Neuquén Basin. The studied palynofloras are correlated with the Biozone of Callialasporites "complex" (Sub-biozone of Callialasporites trilobatus (Balme) Dev, Antulsporites saevus (Balme) Archangelsky and Gamerro and Klukisporites labiatus (Volkheimer) Baldoni and Archangelsky) previously proposed for this area of the Neuquén Basin. A Late Aalenian - Early Bajocian age is suggested for the studied sections.
Key words. Palynology. Systematics. Los Molles Formation. Middle Jurassic. Neuquén Basin. Argentina.
Palabras clave. Palinología. Sistemática. Formación Los Molles. Jurásico medio. Cuenca Neuquina. Argentina.
Introducción
En este trabajo se exponen los resultados del análisis palinológico efectuado sobre muestras provenientes de afloramientos del Jurásico medio, correspondientes a la Formación Los Molles, Grupo Cuyo, en el sector centro-occidental de la cuenca Neuquina. Constituye un aporte al conocimiento de la palinoestratigrafía de la cuenca, ya que se amplía el número de especies citadas en trabajos previos para esta formación.
Se presenta el estudio sistemático de palinomorfos que revisten el carácter de primeras menciones y/o especies nuevas y la lista de especies identificadas. Los resultados del análisis palinofacial y la evaluación paleoambiental y paleoclimática correspondientes a las asociaciones microflorísticas aquí estudiadas serán dados a conocer en futuras publicaciones.
Área de estudio, estratigrafía y edad
La cuenca Neuquina es una cuenca de retroarco, localizada en el sector centro-occidental de Argentina. Se originó durante el Triásico tardío y constituyó una importante área de sedimentación durante el Jurásico y Cretácico temprano. El Grupo Cuyo o "Cuyano" de Groeber (1946) representa el primer episodio importante de deposición después de la configuración de la cuenca (figura 1). En el centro-sur de Neuquén, el "Cuyano" evidencia un arreglo regresivo dada la progradación de las distintas unidades que lo conforman. Para el sector de estudio, y de acuerdo a datos regionales, el Grupo Cuyo se depositó entre el Pliensbaquiano medio hasta el Caloviano temprano (Gulisano et al. , 1984). En este contexto, la sucesión de pelitas oscuras y areniscas intercaladas de interior de cuenca a plataforma, corresponden a la Formación Los Molles (Weaver, 1931). De acuerdo a datos paleontológicos, Volkheimer (1973) sugiere que esta formación se habría depositado entre el Toarciano y Aaleniano, pudiendo extenderse el límite superior hasta el Bajociano basal, con algunas dudas.

Figura 1. Columna estratigráfica de la cuenca Neuquina, válida para el área de estudio / Stratigraphic scheme of the Neuquén Basin, corresponding to the study area.
El área de estudio se ubica en el sector centro-occidental de la cuenca Neuquina, entre los 39º 10' - 39º 21' latitud sur y 69º 39' - 70º 25' longitud oeste y comprende 5 perfiles localizados en cercanías de la ciudad de Zapala (figura 2), e identificados como perfiles Lohan Mahuida (708 metros de espesor estratigráfico), Puesto Policía (513 metros), Puente Picún Leufú (836 metros), Los Molles (403 metros) y Cerro Lotena (928 metros).

Figura 2. Mapa de ubicación de las localidades de estudio / Location map of the studied localities.
Mediante calibración con fauna de amonites y por posición en la secuencia, Zavala (1993, 1996a, 1996b), reconoce para la Formación Los Molles dos secuencias deposicionales de cuarto orden, denominadas JC4.1 y JC4.2. En los perfiles estudiados, la secuencia JC4.1 no cuenta con hallazgos macropaleontológicos de relevancia, sin embargo en sectores cercanos al área de estudio Zavala (1993, 1996a, 1996b) menciona, para la misma secuencia, la presencia de amonites representativos de las Zonas de Asociación de Puchenquia malarguensis y Pseudotoites singularis (Westermann y Riccardi, 1979) y para los niveles superiores, de la Zona de Asociación de Emileia giebeli submicrostoma (Westermann y Riccardi, 1979), sugiriendo un rango de edad entre el Aaleniano tardío y el Bajociano temprano bajo. Riccardi y Gulisano (1990) extienden el límite basal de esta secuencia más allá del Toarciano tardío.
En la secuencia JC4.2, Zavala (1993) cita a los 126 metros de la base del perfil Lohan Mahuida, la presencia de Emileia sp., de la Zona de Asociación de Emileia giebeli (Westermann y Riccardi, 1979) lo cual sugiere una edad bajociana temprana.
Antecedentes palinológicos
Las contribuciones de Volkheimer y Quattrocchio (1981) y Quattrocchio et al. (1996a) constituyen verdaderos trabajos de síntesis con distribuciones estratigráficas de palinomorfos y zonaciones polínicas para el Jurásico y Cretácico de la cuenca Neuquina. Los estudios palinoestratigráficos específicos de la Formación Los Molles son hasta el presente escasos, en comparación con la abundante información proveniente de la Formación Lajas. En Volkheimer (1973, 1974) se sintetizan aspectos relativos a la estratigrafía, y se describen e ilustran palinomorfos extraídos de las formaciones Sierra Chacai Co y Los Molles en Sierra de Chacai Co y adyacencias. Scafati y Morbelli (1984) aportan datos palinológicos de la Formación Lajas y mencionan e ilustran algunos taxones recuperados de la Formación Los Molles. Quattrocchio et al. (1996b), García (1998), Martínez (1999), Martínez et al. (2001) y García et al. (1994, 2000) realizan aportes vinculados a la palinoestratigrafía y palinofacies del Grupo Cuyo. Más recientemente, Martínez (2002), presenta una nueva zonación polínica para el Jurásico medio, en el área de estudio, tomando como base el análisis secuencial y datación con fauna de amonites publicados previamente por otros autores (Zavala, 1993, 1996a, 1996b).
Metodología
A fin de realizar un estudio de estratigrafía por secuencias, Zavala (1993) levantó las 5 secciones estratigráficas de detalle, correspondientes a las formaciones Los Molles, Lajas, Challacó, Lotena y Quebrada del Sapo, con un espesor acumulado de 3388 metros. Dichas secciones fueron medidas banco a banco mediante el uso de báculo de Jacob y muestreadas para estudios macro y micropaleontológicos. Los perfiles completos, con detalle de litofacies ambientes y secuencias deposicionales han sido presentados en Zavala (1993).
Del total de 20 muestras procesadas, correspondientes a la Fm. Los Molles, 11 resultaron palinológicamente fértiles, esto es, con un contenido palinológico que permitió realizar análisis estadísticos en las asociaciones microflorísticas. Las muestras restantes resultaron estériles o con escasa presencia de palinomorfos. En la tabla 1 se consignan las muestras estudiadas, discriminadas según resultaron fértiles o estériles, con la numeración de laboratorio, su ubicación dentro de cada perfil (medido en metros desde la base del perfil), litofacies, ambientes deposicionales y unidades secuenciales correspondientes.

Tabla 1. Distribución de las muestras palinológicas en los perfiles analizados, *muestras fértiles / Distribution of the palynological samples in each analyzed section, * samples with palynomorphs.
La extracción físico-química de las muestras palinológicas se efectuó sometiendo el material al ataque con HCl-HF (ácidos clorhídrico-fluorhídrico) para eliminar carbonatos y silicatos. Cuando fue necesario para el estudio, se oxidó el residuo con HNO 3 (ácido nítrico) concentrado. Los compuestos húmicos fueron removidos con OHNH 4 (hidróxido de amonio) al 5%. Por último los palinomorfos fueron concentrados utilizando tratamiento con ZnCl 2 (cloruro de cinc). Los preparados definitivos se montaron en glicerinagelatina y el residuo palinológico fue preservado en glicerina y solución de formol o fenol. Las muestras están archivadas en el Laboratorio de Palinología de la Universidad Nacional del Sur bajo las siglas UNSP (Universidad Nacional del Sur - Palinología), seguido de las siglas correspondientes a cada perfil: LM (Lohan Mahuida), PL (Puente Picún Leufú), PP (Puesto Policía), M (Los Molles) y CL (Cerro Lotena).
El estudio de las muestras se realizó utilizando un microscopio de luz transmitida Olympus BH 2 Nº 100786. Las fotomicrografías fueron obtenidas en microscopio óptico con una cámara Olympus y película Agfa BN de 100 ASA (figuras 3-5, 8) Para el estudio de parte del microplancton se utilizó un microscopio Nikon Eclipse 600 y se obtuvieron imágenes digitalizadas (figura 9). Las coordenadas corresponden a la escala Vernier del primer microscopio o más frecuentemente a la reglilla England Finder .
El análisis estadístico de las asociaciones microflorísticas se realizó a partir de recuentos polínicos (200 palinomorfos), obteniéndose valores de frecuencias relativas (%) para cada taxón. En las figuras 10- 12 se presentan la ubicación de las muestras palinológicas estudiadas en los perfiles que resultaron fértiles y las frecuencias relativas de cada uno de los taxones identificados como rango de proporciones. Los resultados derivados de este análisis se darán a conocer en una próxima contribución.
Descripciones sistemáticas
Se encontró una variada palinoflora, de la cual sólo se describen aquellas formas nuevas no descriptas antes para la cuenca o con nomenclatura abierta, tanto de esporomorfos como de paleomicroplancton. La lista taxonómica completa se incluye en el Apéndice A.
Esporomorfos
Anteturma Sporites H. Potonié1893
Turma Triletes Reinsch, Potonié y Kremp 1954
Subturma Azonotriletes Luber, emend . Dettmann 1963
Infraturma Laevigati Bennie y Kidston, emend . Potonié 1956
Género Divisisporites Pflug en Thomson y Pflug 1953
Especie tipo. Divisisporites divisus Pflug en Thomson y Pflug, pl. 1, fig. 59.
Divisisporites sp.
Figura 3.A
Material estudiado. UNSP PP 1795c: Y43/4.

Figura 3. A, Divisisporites sp., UNSP PP 1795c: Y43/4, superficie proximal / proximal surface . B, Dictyophyllidites mortoni (de Jersey) Playford y Dettmann, UNSP PP 1795c: C57/5. C, espora gen. et sp. indet., UNSP PP 1790c: L31, superficie proximal / proximal surface . D, Antulsporites varigranulatus (Levet-Carette) Reiser y Williams, UNSP PP 1793c: H60/3, superficie distal / distal surface . E, Rugulatisporites neuquenensis Volkheimer, UNSP PP 1793c: T47/3. F, Classopollis intrareticulatus Volkheimer, UNSP PP 1793c: C57, vista ecuatorial / equatorial view . G, Cerebropollenites cf. C. macroverrucosus (Thiergart) Schulz, UNSP PP 1795c: E39/1. H, Cerebropollenites macroverrucosus (Thiergart) Schulz, UNSP PP 1789c: Y51/4, vista polar con detalle del colpo distal / polar view with detail of the distal colpa. I, Monosulcites sp. cf. M. sp. B (en Volkheimer y Quattrocchio, 1975), UNSP PP 1796c: Y51/1, superficie distal / distal surface . J, L, Callialasporites microvelatus Schulz, J, UNSP PP 1793c: C42/3; L, UNSP PP 1795c: F14/2. K, Callialasporites sp., UNSP PP 1793c: S59. M, Phrixipollenites sp. UNSP PP 1790c: N55/2, vista ecuatorial / equatorial view . N-O, Microcachryidites castellanosii Menéndez; N, UNSP PP 1793c: U11, vista polar, superficie distal / polar view, distal surface ; O, UNSP PP 1793c: F58/3, vista ecuatorial / equatorial view . Todas las fotomicrografías están ampliadas 1000X, excepto K = 400X. Escala gráfica = 10µm / K is 400X, all others are 1000X. Scale bar equals 10µm .
Descripción. Espora trilete de contorno ecuatorial triangular, con un lado cóncavo y dos rectos a levemente convexos; ángulos redondeados. Los rayos de la marca Y son simples y alcanzan 4/5 del radio de la espora. Dos de los rayos de la marca Y presentan bifurcaciones terminales de ángulo agudo. Exina escabrada de 2 µm de espesor.
Dimensiones (1 ejemplar medido). Diámetro ecuatorial máximo: 50 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. Divisisporites sp. A (en Volkheimer 1972) tiene menor tamaño (37 µm) y los rayos de la marca Y en su posición apical son sinuosos. Divisisporites sp. (en Volkheimer 1968) posee exina lisa y de menor espesor (1,5 µm).
Espora gen. et sp. indet.
Figura 3.C
Material estudiado. UNSP PP 1790c: L31.
Descripción. Espora trilete de contorno ecuatorial triangular, con un lado recto, uno levemente cónvexo y otro cóncavo; ángulos redondeados. Los rayos de la marca Y son largos, simples y se bifurcan al llegar al borde interno del ecuador. La exina es lisa y de espesor diferencial mayor en los ápices (2 µm) que en los lados (1 µm). Sobre el hemisferio distal se observan engrosamientos o pliegues en forma de media luna ( "fold-tori" ), cuyos lados convexos se dirigen hacia el polo distal y se van afinando hacia los ápices.
Dimensiones (1 ejemplar medido). Diámetro ecuatorial máximo: 42 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. El ejemplar estudiado presenta rasgos comunes a varios géneros: bifurcaciones terminales de los rayos de la marca Y como en Divisisporites Thomson, engrosamiento diferencial de la exina en la zona ecuatorial mayor en los ápices que en los interradios como en Matonisporites (Couper) Potonié, presencia de fold-tori como en Gleicheniidites (Ross) Skarby. Probablemente se trate de una forma transicional entre los dos últimos géneros mencionados.
Anteturma Pollenites Potonié 1931
Turma Saccites Erdtman 1947
Género Callialasporites Dev 1961
Especie tipo. Callialasporites (Zonalapollenites) trilobatus (Balme) Dev 1961.
Callialasporites sp.
Figura 3.K
Material estudiado. UNSP PP 1793c: S59.
Descripción. Grano de polen monosacado de contorno subcircular. La ornamentación del saco aéreo varía desde microgranulada hasta gránulos del orden del micrómetro, con un espesor de exina de 1,5 µm, numerosos pliegues radiales y alrededor de 20 µm de ancho. Espesor de la exina en el cuerpo central ~2 µm. Sobre el polo distal los pliegues se disponen tan apretadamente que forman un arreglo de apariencia finamente corrugada en el cuerpo central. Se observan vestigios de una posible marca trilete.
Dimensiones (1 ejemplar medido). Diámetro total: 80-91 µm; diámetro del cuerpo central: 49-55 µm; ancho del saco: 14-19 µm.
Distribución. Perfil Puesto Policía.
Observaciones. Es notoria la formación de pliegues más prominentes en el saco, en los sitios que coinciden con la proyección de la marca trilete vestigial sobre el mismo.
Comparaciones. Es similar a Callialasporites sp. A (en Volkheimer, 1972), sin embargo la ornamentación en el presente material es más conspícua. Difiere de Callialasporites dampieri (Balme) Dev 1961, por las dimensiones menores del material de Balme (1957), con un diámetro total: 43-78 µm.
Género Cerebropollenites Nilsson 1958
Especie tipo. Cerebropollenites (Tsugaepollenites) mesozoicus (Couper) Nilsson (en Couper 1958).
Cerebropollenites macroverrucosus (Thiergart) Schulz 1967
Figura 3.H
Material estudiado. UNSP PP 1789c: Y51/4.
Dimensiones (1 ejemplar medido). Diámetro mayor: 42 µm; diámetro menor: 35 µm; colpo distal: 6x19 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. Es similar al material ilustrado en la figura 3.G ( Cerebropollenites cf. macroverrucosus ), sin embargo en este último no se observa tan claramente la presencia del colpo distal.
Observaciones. Según Abbink, 1998 (p. 60), si bien existe incertidumbre en cuanto a la afinidad botánica de este género (taxodiáceas, pináceas, etc.), se han registrado grandes cantidades del mismo inmediatamente después de una regresión, lo cual podría indicar el carácter colonizador de las plantas parentales.
Turma Plicates (Naumova 1939) Potonié 1960
Subturma Monocolpates Wodehouse, Iversen y Troels-Smith 1953
Género Monosulcites (Erdtman 1947, Cookson 1947) ex Couper 1953
Lectotipo. Monosulcites minimus (Cookson 1947) elegido por Couper 1953.
Monosulcites sp. cf. M. sp. B. (en Volkheimer y Quattrocchio 1975)
Figura 3.I
Material estudiado. UNSP PP 1796c: Y51/1.
Descripción. Grano de polen monocolpado. Contorno ecuatorial ovalado. Extremos redondeados. El colpo llega al ecuador y está acompañado por pliegues de hasta 5 µm de ancho; sus bordes se traslapan alrededor de 2/3 de la longitud total del grano. Exina delgada (menos de 1 µm de espesor), escabrada.
Dimensiones (1 ejemplar medido). Largo: 24 µm; ancho: 18 µm; relación largo/ancho: 1,3.
Distribución. Perfil Puesto Policía.
Comparaciones. Se compara y no se asigna a la especie de Volkheimer y Quattrocchio (1975), debido a que en el material original los bordes del colpo están abiertos en toda su extensión sin tocarse.
Paleomicroplancton
División Prasinophyta Round 1971
Clase Prasinophyceae Parke (en Parke y Dixon 1964)
Las Prasinophyceae han merecido escasa consideración en estudios palinológicos del Mesozoico de nuestro país. Muchos palinólogos han ignorado estas algas o sólo se han referido a ellas esporádicamente debido a su escaso valor para correlaciones estratigráficas (Brocke y Riegel, 1996). Guy-Ohlson y Boalch (1992) examinaron distintos estadíos de desarrollo del phycoma y de la fase móvil tanto del fósil Tasmanites Newton como del actual Pachysphaera Ostenfeld y confirmaron la similitud en las características de estas formas ya observadas por otros autores. Llegaron a la conclusión que este género permaneció inalterable desde el Paleozoico hasta nuestros días, por más de 300 millones de años, demostrando su alta capacidad de adaptación al medio ambiente y sus cambios (Guy-Ohlson, 1996).
Fensome et al. (1990) han compilado una lista de 88 especies del género Tasmanites , en comparación con el escaso número de especies de los géneros actuales Pachysphaera (2), Halosphaera (3) y Pterosperma (7), mencionados en Guy-Ohlson (1988). Guy-Ohlson y Boalch (1992) y Guy-Ohlson (1996), consideran que gran parte de estas "especies" en realidad representan morfotipos de una misma especie, que reflejan diferentes estadíos de desarrollo dentro de cada fase del ciclo de vida. Desde este punto de vista, intentar conclusiones palinoestratigráficas, resulta cuanto menos arriesgado sin una re-evaluación previa de estos taxones.
Estas dificultades, sin embargo, no deben restarle valor a las Prasinophyceae como indicadoras paleoambientales. Los phycomas de Prasinophyceae fósiles son frecuentes en ambientes marinos de salinidad reducida, pero no así en depósitos lacustres (Brocke y Riegel, 1996). Prauss (1989, en Prauss, 2001) propone la sucesión: prasinofitas → acritarcos → dinoquistes como indicativa de un gradiente de salinidad creciente desde condiciones costeras hasta marino abiertas. Brocke y Riegel (1996, p. 57) amplían este esquema incorporando otros grupos de algas de agua dulce y salinidad periódica, como Botrycococcus , Plaesiodictyon y Pediastrum entre otros. Prauss y Riegel (1989) y Prauss (1996, 2001) sugirieron que la distribución de estas algas se vió favorecida por masas de agua, no sólo de reducida salinidad, sino también de baja temperatura. Sin embargo, nuevos registros de prasinofitas asociados a depósitos deltaicos del Pleistoceno en latitudes sub-ecuatoriales, confirman el rol de la salinidad como principal factor de control, si no el único, en la distribución de estas algas, restando a la temperatura una influencia directa (Brocke y Riegel, 1996, p. 62). Al-Almery (1983, 1986, en Prauss, 1996, p. 122) propone una sucesión de prasinophytas, desde géneros sin ornamentación conspicua ( Pleurozonaria y Tasmanites ) a otros con retículos superficiales o alas ecuatoriales ( Pterosphaeridia y Pterospermopsis ), sugiriendo el predominio de estos últimos un relativo incremento en la profundidad del agua o condiciones más distales de la costa.
Orden Pterospermatales Parke y Green en Parke y Dixon 1976
Familia Tasmanaceae Sommer 1956 emend . Mädler 1963
Género Pleurozonaria Wetzel 1933, emend . Mädler 1963
Especie tipo. Pleurozonaria globulus Wetzel 1933.
Pleurozonaria cf. P. picunensis Quattrocchio 1980
Figuras 4.M-O
Material estudiado. UNSP PL 1429a: 21,1/110,9; 16,9/122,6; 1429c: 4/160,8; 21,2/161,7; 1429d: 23,8/140,4; 23,3/140,9; 22,4/146,2; 22,8/151,1; 19,9/145; 14,5/156,1; 14,6/149,2; 1429e: 1,3/120, 1429 CAN: E42/4 (figura 4.O), K29/1, K39/4 (figuras 4.M-N), L35, M28/4, M30, M33/2, X48/1, S38/1. UNSP PP 1790c: G14, L19/2. UNSP M 1826c: V50/4.

Figura 4. A, Cymatiosphaera eupeplos (Valensi) Deflandre, UNSP PP 1790c: J44. B-C, Cymatiosphaera cf. C. eupeplos (Valensi) Deflandre, UNSP PL 1429d: M62. D-L, Cymatiosphaera cf. C. volkheimerii (Quattrocchio) n. comb., D, UNSP PL 1429 CAN: O35; E-H, UNSP PL 1429 CAN: P39/1, diferentes enfoques mostrando la superficie reticulada con membranas fuertemente sinuosas vistas en planta / different focuses showing the reticulate surface with strongly undulatory membranes in plan view; H, tomada usando contraste de fase / taken under phase contrast illumination; I, UNSP PL 1429 CAN: E32, tomada usando contraste de interferencia / taken under interference contrast illumination; J, UNSP PL 1429 CAN: D46/2, detalle de membranas con márgenes distales rectos / details of membranes with straight outer margins; K, UNSP PL 1429 CAN: Y29/4; L, UNSP PL 1429 CAN: C37/1. M-O, Pleurozonaria cf. P. picunensis Quattrocchio, M-N, UNSP PL 1429 CAN: K39/4; O, UNSP PL 1429 CAN: E42/4. P, Leiosphaeridia cf. L. sp. F (en Prámparo 1989), UNSP PP 1790c: S49. Q, Botryococcus spp., UNSP PP 1790c: J57/3. Todas las fotomicrografías están ampliadas 1000X, excepto Q = 400X. Escala gráfica= 10 µm / Q is 400X, all others are 1000X. Scale bar equals 10 µm.
Descripción. Cuerpos originalmente esféricos deformados debido al intenso plegamiento secundario ovoidal a subcircularmente. Espesor de la pared: 0,8- 1,7 µm. Los poroides ± poligonales de 0,5-1,2 µm de diámetro, están densa y uniformemente dispuestos. En el contorno se observa que los poroides están limitados hacia afuera por una saliencia hemisférica de la capa externa de la pared; de este modo, la pared aparece densamente "granulada" (alto de los "gránulos": 0,3-1 µm). En el centro de los poroides se observa un punto blanco (u oscuro según el enfoque LO). En algunos ejemplares pudo observarse la presencia de canales porosos de aproximadamente 1 µm de diámetro, sin embargo y debido al regular estado de preservación del material es difícil distinguir a éstos del efecto de corrosión por hongos y bacterias.
Dimensiones (hasta 30 ejemplares medidos). Diámetro máximo del cuerpo: 29-(41,5)-62 µm; diámetro poroides: 0,5-(0,8)-1,2 µm; alto de los "gránulos": 0,3- (0,6)-1 µm; espesor de la pared: 0,8-(1,2)-1,7 µm. Distribución. Perfiles Puente Picún Leufú, Puesto Policía y Los Molles.
Comparaciones. Pleurozonaria media Mädler 1963 posee mayor diámetro del cuerpo (100-130 µm) y de los poroides (2,2-2,7 µm). Pleurozonaria distans Mädler 1963 tiene un diámetro mayor 80-84 µm, y un diámetro de los poroides de 2-2,5 µm.
El presente material es similar a Pleurozonaria picunensis Quattrocchio 1980, citado para la Fm. Vaca Muerta (Tithoniano) en Picún Leufú, sin embargo, luego de una minuciosa observación de los holotipos descriptos por Quattrocchio (1980), surgen algunas diferencias: el rango de dimensiones de esta especie es menor: 19-48 µm; el espesor de la pared (1-1,8 µm) y diámetro de los poroides (1-2 µm), son mayores.
Observaciones. Es posible que tanto el material aquí descripto como el citado por Quattrocchio (1980), representen distintos estadíos evolutivos dentro de una misma especie. Investigaciones de detalle futuras en formaciones de edad intermedia aportarán evidencia para confirmar o descartar esta hipótesis.
Género Tasmanites Newton 1875
Especie tipo. Tasmanites punctatus Newton 1875.
Tasmanites sp.
Figuras 5 y 6.A-J

Figura 5. Características estructurales comparativas de Tasmanites sp. y taxones afines / Comparative structural characteristics of Tasmanites sp. and related taxa. Figura 6. A-J, Tasmanites sp., A-B, UNSP PL 1429a: C32, las flechas muestran canales porosos / the arrows show pore-canals, A, sección óptica / optical section, B, foco alto / high focus; C, UNSP PL 1429e: H26/2; D, UNSP PL 1429a: H31/1, detalle de canales porosos atravesando toda la pared / detail of the pore-canals penetrating the entire wall; E, UNSP PL 14229e: V34/1, vista sub-ecuatorial mostrando el típico aspecto de discos comprimidos / sub-equatorial view showing the tipical compressed disk-shaped aspect; F-J, UNSP PL 1429a: W9/2; G, las flechas muestran finas puntuaciones accesorias entre las prominencias poligonales / the arrows show the accesory fine punctuations among the polygonal prominences; H-J, vista en planta de la estructura de la pared, con foco alto, intermedio y bajo, respectivamente / plan view of the wall structure, with high, intermediate and low focuses, respectively, J, superficie interna de la pared mostrando la finalización de canales porosos / the inner surface of the wall showing the end of pore-canals. K-L, Micrhystridium cf. M. gregarium Sarjeant, UNSP PL 1466a: Q46/4, agrupamiento con alrededor de 40 ejemplares / cluster of about 40 specimens. M, Polygonium sp. cf. ? P. jurassicum Bucefalo Palliani, Riding y Torricelli, UNSP PP 1792c: F45/3. N, Polygonium sp., UNSP PP 1791c: E28/2, tomada usando contraste de fase / taken under phase contrast illumination. O, Veryhachium valensii (Valensi) Downie y Sarjeant, UNSP PL 1429a: G30. P-Q, Filisphaeridium castaninum (Valensi 1953) Sarjeant y Stancliffe, UNSP PP 1793c: E57/2, P, detalle de espinas y contorno / detail of the spines and outline; Q, detalle del piloma subpoligonal / detail of the sub-polygonal pylome. Todas las fotomicrografías están ampliadas 1000X, excepto A-C, F y K = 400X. Escala gráfica = 10 µm / A-C, F and K are 400X, all others are 1000X. Scale bar equals 10 µm.
Material estudiado. UNSP PL 1429a: C32 (figuras 6.A-B), H31/1 (figura 6.D), J30/3, W9/2 (figuras 6.F-J); 1429b: C31/4, D6/2, O31; 1429c: K9, X26, Y40/3; 1429d: D10/2; 1429e: H26/2 (figura 6.C); V33/2; V34/1 (figura 6.E); 1429f: L43/1. UNSP M 1826c: Y11/2.

Tabla 2. Características comparativas de Cymatiosphaera sp. 1, C. sp. 2, C. sp. 3 y C. cf. C. volkheimerii / Comparative characteristics of Cymatiosphaera sp. 1, C. sp. 2, C. sp. 3 and C. cf. C. volkheimerii.
Descripción. Cuerpos originalmente esféricos, generalmente comprimidos a modo de discos. Pared gruesa de hasta 8 µm de espesor. La superficie de la pared está densa y uniformemente ornamentada por gránulos de contorno subcircular hasta perfectamente poligonal (hexágonos) de muy bajo relieve dando la apariencia de un panal de abejas. En el corte óptico los gránulos se presentan como saliencias hemisféricas o aplanados distalmente. El centro de cada gránulo es atravesado por un diminuto canal (observable como un punto blanco u oscuro según el enfoque LO), que no siempre penetra en toda la pared. En algunos ejemplares se han observado puntuaciones accesorias entre los ornamentos que corresponden a canales similares a los anteriores (figura 6.G). En casi todos los ejemplares pudo observarse la presencia de canales porosos de aproximadamente 1µm de diámetro que penetran toda la pared distribuidos ± uniformemente sobre toda la superficie y con un espaciamiento variable. En algunos ejemplares (?juveniles) no se observa la presencia de estos canales. En cortes de determinados ejemplares se observa que estos canales porosos poseen paredes que se van ensanchando hacia el interior del cuerpo, razón por la cual son más fácilmente diferenciables al observar el enfoque más bajo de la pared. La mayoría de los ejemplares exhiben importante daño mecánico: rotura y/o plegamiento. Se observa la presencia de hendiduras posiblemente correspondientes a la liberación del contenido celular.
Dimensiones (hasta 15 ejemplares medidos). Diámetro máximo del cuerpo: 75-(93,2)-115 µm; diámetro de los gránulos: 1-(1,25)-2,5 µm; alto de los gránulos: 0,5-(0,66)-1 µm; espesor de la pared: 3-(5,2)-8 µm; distancia menor entre canales porosos: 5-(6,3)-8 µm; distancia mayor entre canales porosos: 6-(8,7)-13 µm.
Distribución. Perfiles Puente Picún Leufú y Los Molles.
Observaciones. En Volkheimer (1974), se citan e ilustran ejemplares presumiblemente juveniles de características similares a los aquí descriptos asignados a Pleurozonaria suevica (=Tasmanites suevicus Wall 1965), para muestras correspondientes a la Formación Los Molles.
En la figura 5 se observan las características estructurales de Tasmanites sp. (en este trabajo), dibujadas a partir del estudio de un ejemplar incompleto (figuras 6.F-J), en el que puede observarse claramente (por el tamaño de los polígonos) todos sus rasgos.
Comparaciones. El presente material muestra características transicionales (figura 6) entre ciertas especies del género Crassosphaera Cookson y Manum 1960: Crassosphaera concinna Cookson y Manum 1960, Crassosphaera hexagonalis Wall 1965 y ejemplares correspondientes al género Tasmanites Newton 1875: Tasmanites suevicus (Eisenack) Wall 1965 y Tasmanites sp. (en Guy- Ohlson y Boalch, 1992, fig. 2; Guy-Ohlson y Norling, 1994, lám. 2, fig. 12 y Guy-Ohlson, 1996, lám. 1, figs. 6 y 9). Los especímenes descriptos e ilustrados en este trabajo han sido incluidos dentro del género Tasmanites dado que el rasgo citado por Cookson y Manum (1960) para el género Crassosphaera (túbulos radiales dentro de cada prominencia que atraviesan toda la pared), no ha sido observado de manera constante. Siguiendo el criterio de Guy-Ohlson en sucesivos trabajos (Guy- Ohlson, 1988, Guy-Ohlson y Boalch, 1992, Guy-Ohlson, 1996), no se realiza una asignación específica dado que ejemplares que morfológicamente son diferentes pueden corresponder a morfotipos de una misma especie en respuesta a diferentes estadíos de desarrollo dentro de cada fase del ciclo de vida.
Familia Cymatiosphaeraceae Mädler 1963
Género Cymatiosphaera Wetzel, emend. Deflandre 1954
Especie tipo. Cymatiosphaera radiata Wetzel 1933.
"Complejo Cymatiosphaera"
Existe cierta confusión al momento de separar dos géneros reticulados muy próximos morfológicamente entre sí: Cymatiosphaera O. Wetzel, emend. Deflandre 1954 y Pterosphaeridia Mädler1963. Su diferenciación se basa en la presencia (Pterosphaeridia) o ausencia (Cymatiosphaera) de canales porosos en el centro de cada polígono. Sin embargo, al trabajar sólo con microscopía óptica de luz transmitida, dichos poros difícilmente son observables, en cuyo caso la separación resulta casi imposible. Para salvar este inconveniente algunos autores (Prauss et al., 1991, p. 339), incluyen a todas las formas con retículos regulares, elevados, dentro de Pterosphaeridia y formas con retículos irregulares de muy bajo relieve dentro de Cymatiosphaera.
En las prasinofíceas reticuladas estudiadas no se observa la presencia de poro y los retículos muestran toda una gradación en cuanto a regularidad en la distribución de los polígonos y altura de las membranas. El criterio seguido por Prauss et al. (1991), puede llevar a una excesiva separación taxonómica, la que en realidad, puede estar representando sucesivos estadíos en el desarrollo de la misma especie. Por este motivo se ha optado por utilizar el término "Complejo Cymatiosphaera" para las formas reticuladas estudiadas sin indicios evidentes del mencionado poro.
Recientemente Schrank (2003, p. 229), basado en ejemplares del Campaniano y Maastrichtiano de Egipto ha sugerido que el género Cymatiosphaera sería heterogéneo en cuanto a su composición, incluyendo formas de diferentes grupos algales; pudiendo representar incluso quistes de dinoflagelados.
Hasta no disponer de estudios más generalizados, que incluyan distintos especímenes de edad jurásica, se prefiere seguir considerando a este grupo con afinidad a las prasinofíceas.
Cymatiosphaera volkheimerii (Quattrocchio 1980) n. comb.
1980 Pterosphaeridia volkheimerii Quattrocchio 1980, lám. G, H, figs. 58-59, 67
La nueva combinación surge por la necesidad de acotar al género Pterosphaeridia Mädler 1963 a ejemplares que presentan canales porosos en el centro de cada polígono. Al haberse revisado el material original de Quattrocchio (1980) y comprobarse en él la ausencia de canales porosos, se reasigna dicha especie al género Cymatiosphaera. Veáse discusión en "Complejo Cymatiosphera".
Cymatiosphaera cf. C. volkheimerii (Quattrocchio) n. comb.
Figuras 4.D-L
Material estudiado. UNSP PL 1429 CAN: C37/1 (figura 4.L), D46/2 (figura 4.J), E32 (figura 4.I), F39, G30/2, H36/2, O35 (figura 4.D), P39/1 (figuras 4.E-H), V39, Y29/4 (figura 4.K); 1429c: W65; 1429d: D18/1, G44/2. UNSP PP 1790c: G53.
Descripción. Vesícula esferoidal, deformada sub-esferoidal a ovoidalmente por plegamiento secundario, de pared lisa de hasta 2 µm de espesor. Superficie externa dividida en campos poligonales (20-25) por membranas delgadas, lisas, rectas a levemente sinuosas, ocasionalmente fuertemente sinuosas (figuras 4.E-H) que escasamente sobresalen del contorno ecuatorial de hasta 2,2 µm de alto. La malla del retículo presenta una distribución más o menos heterogénea en el tamaño de los polígonos, con diámetros que varían entre 12 y 19 µm. Los márgenes distales de las membranas son discontinuos, rectos a recortados. Los ángulos de los polígonos presentan refuerzos y en vista lateral adquieren la forma de conos, báculas o espinas de 1 µm de ancho por 1,56 µm de alto en promedio. No se observan canales porosos en el interior de los campos poligonales.
Dimensiones (14 ejemplares medidos). Diámetro mayor de la vesícula: 30-(37,4)-47 µm; diámetro menor de la vesícula: 26-(31,8)-42 µm; espesor de la pared: 0,8-(1,5)-2 µm; ancho de los refuerzos de los polígonos: 0,5-(1)-1,5 µm; alto de los refuerzos de los polígonos: 1-(1,56)-2 µm; alto de las membranas: 1- (1,56)-2,2 µm; diámetro máximo de los polígonos: 12- (15)-19 µm.
Distribución. Perfiles Puente Picún Leufú y Puesto Policía.
Comparaciones. El presente material es muy similar a Pterosphaeridia volkheimerii Quattrocchio 1980, del cual pudieron observarse el holotipo así como otros ejemplares en los que se basó la descripción original. Sin embargo difiere levemente en las dimensiones, mayor rango de dimensiones del diámetro total: 24- 65 µm y espesor de la pared: 0,8-3 µm, menor altura de las lamélulas: 0,5-0,8 µm, diámetro de la malla del retículo más pequeña: 5-15 µm.
Cymatiosphaera cf. C. eupeplos (Valensi 1948) Deflandre 1954
Figura 4.B-C
Material estudiado. UNSP PL 1429c: W65; 1429d: M62 (figuras 4.B-C).
Descripción. Vesícula esferoidal, de pared lisa, de ~2 µm de espesor. Superficie externa dividida en campos poligonales (25-30), por membranas delgadas, lisas, levemente sinuosas a sinuosas, de 1µm de alto. El diámetro de los polígonos varía entre 6 y 10 µm. Los márgenes distales de las membranas son rectos a irregulares. Los ángulos de los polígonos se presentan engrosados y en vista lateral adquieren la forma de bastones de 1 µm de ancho y 1,5 µm de alto.
Dimensiones (2 ejemplares medidos). Diámetro de la vesícula: 27-30 µm; diámetro total: 30-32 µm.
Distribución. Perfil Puente Picún Leufú.
Observaciones. La descripción original de Michystridium eupeplos (Valensi 1948) = Cymatiosphaera eupeplos (Valensi 1948) Deflandre 1954 permite incluir ejemplares cuyas dimensiones de la vesícula son: 9- 16 µm (incluyendo las membranas) y 7-12 µm sin las mismas. De Coninck (1965) considera a Cymatiosphaera parva (Sarjeant 1959) como sinónimo taxonómico senior de Cymatiosphaera eupeplos permitiendo de esta manera extender las dimensiones de vesícula de este taxón hasta 25 µm.
Comparaciones. Se compara y no se asigna a Cymatiosphaera eupeplos (Valensi 1948) Deflandre, 1954 porque ésta posee un diámetro menor (25 µm, como máximo) que la del material aquí estudiado.
Cymatiosphaera sp. 1
Figuras 7.A-B y 9.A-B
Material estudiado. UNSP PL 1466a: 9,4/137,2.

Figura 6. A-J, Tasmanites sp., A-B, UNSP PL 1429a: C32, las flechas muestran canales porosos / the arrows show pore-canals, A, sección óptica / optical section, B, foco alto / high focus; C, UNSP PL 1429e: H26/2; D, UNSP PL 1429a: H31/1, detalle de canales porosos atravesando toda la pared / detail of the pore-canals penetrating the entire wall; E, UNSP PL 14229e: V34/1, vista sub-ecuatorial mostrando el típico aspecto de discos comprimidos / sub-equatorial view showing the tipical compressed disk-shaped aspect; F-J, UNSP PL 1429a: W9/2; G, las flechas muestran finas puntuaciones accesorias entre las prominencias poligonales / the arrows show the accesory fine punctuations among the polygonal prominences; H-J, vista en planta de la estructura de la pared, con foco alto, intermedio y bajo, respectivamente / plan view of the wall structure, with high, intermediate and low focuses, respectively, J, superficie interna de la pared mostrando la finalización de canales porosos / the inner surface of the wall showing the end of pore-canals. K-L, Micrhystridium cf. M. gregarium Sarjeant, UNSP PL 1466a: Q46/4, agrupamiento con alrededor de 40 ejemplares / cluster of about 40 specimens. M, Polygonium sp. cf. ? P. jurassicum Bucefalo Palliani, Riding y Torricelli, UNSP PP 1792c: F45/3. N, Polygonium sp., UNSP PP 1791c: E28/2, tomada usando contraste de fase / taken under phase contrast illumination. O, Veryhachium valensii (Valensi) Downie y Sarjeant, UNSP PL 1429a: G30. P-Q, Filisphaeridium castaninum (Valensi 1953) Sarjeant y Stancliffe, UNSP PP 1793c: E57/2, P, detalle de espinas y contorno / detail of the spines and outline; Q, detalle del piloma subpoligonal / detail of the sub-polygonal pylome. Todas las fotomicrografías están ampliadas 1000X, excepto A-C, F y K = 400X. Escala gráfica = 10 µm / A-C, F and K are 400X, all others are 1000X. Scale bar equals 10 µm

Figura 7. Esquema lineal de Cymatiosphaera sp. 1/Sketch of Cymatiosphaera sp. 1. A. Enfoque distal/distal focus. B. Enfoque proximal/proximal focus.
Descripción. Vesícula ovoidal, de pared lisa, de 3 µm de espesor. Superficie externa dividida en campos poligonales (más de 30), por membranas delgadas microgranuladas, rectas a levemente sinuosas, de 2 µm de alto. La malla del retículo presenta una distribución heterogénea en el tamaño de los polígonos, con diámetros que varían entre 13 y 25 µm. Los márgenes distales de las membranas son irregulares. Los ángulos de los polígonos se presentan engrosados y en vista lateral adquieren la forma de conos, conos truncados y báculas de 3,5 µm de alto x 3 µm de ancho.
Dimensiones (1 ejemplar medido). Diámetro máximo de la vesícula: 65 µm; diámetro máximo total: 72 µm.
Distribución. Perfil Puente Picún Leufú. Comparaciones. Es similar a Cymatiosphaera asarota Davey 1970 del Cenomaniano de Canadá. Sin embargo en ésta no se menciona la presencia de membranas microgranuladas y el tamaño es considerablemente menor (diámetro máximo: 53 µm).
Cymatiosphaera sp. 2
Figuras 9.C-D
Material estudiado. UNSP PL 1466a: W59/4.
Descripción. Vesícula eferoidal de pared lisa, de 3,5 µm de espesor. Superficie externa dividida en campos poligonales (aproximadamente 22) por membranas delgadas, microgranuladas, rectas de 4 µm de alto. La malla del retículo presenta una distribución heterogénea en el tamaño de los polígonos, con diámetros que varían entre 15 y 21 µm. En el enfoque proximal, se observan campos poligonales de menor tamaño y en mayor número (± 13), mientras que en el enfoque distal, se observan campos poligonales de mayor tamaño y en menor número (± 9). Los márgenes distales de las membranas son irregulares. Los ángulos de los polígonos se presentan engrosados y en vista lateral adquieren la forma de conos y conos truncados de 3 µm de ancho x 4 µm de alto.
Dimensiones (1 ejemplar medido). Diámetro máximo de la vesícula: 57 µm; diámetro máximo total: 63 µm.
Distribución. Perfil Puente Picún Leufú.
Comparaciones. Es similar a Cymatiosphaera sp. 1 (en este trabajo), sin embargo esta última posee un tamaño mayor, un número de campos poligonales también mayor, que se distribuyen en similar número en ambas vistas y un espesor de la pared ligeramente menor.
Observaciones. Es probable que exista toda una gama de formas transicionales entre Cymatiosphaera sp. 1 y C. sp. 2, sin embargo y debido a que sólo se ha identificado un ejemplar de cada especie, se prefiere profundizar el estudio con más material para intentar llegar a una determinación a nivel específico.
Cymatiosphaera sp. 3
Figuras 9.E-F
Material estudiado. UNSP PL 1466 a: 21,4/139,1 (figura 9.F); 22,2/156,5; D51/2 (figura 9.E).
Descripción. Vesícula ovoidal, de pared microgranulada de ±1,5 µm de espesor. Superficie externa dividida en campos poligonales (aproximadamente 30), por membranas delgadas, puntuadas a microgranuladas, rectas a sinuosas de 2 a 6 µm de alto. La malla del retículo presenta una distribución heterogénea en el tamaño de los polígonos, con diámetros que varían entre 12 y 19 µm. Los márgenes distales de las membranas son rectos a levemente convexos. Los ángulos de los polígonos se presentan engrosados y en vista lateral adquieren la forma de conos truncados, espinas y báculas de 1,5-3 µm de ancho x 3-5 µm de alto.
Dimensiones (3 ejemplares medidos). Diámetro máximo de la vesícula: 52-(56)-62 µm; diámetro máximo total: 56-(64)-70 µm.
Distribución. Perfil Puente Picún Leufú.
Comparaciones. Cymatiosphaera areolata Deflandre 1941, del Kimmeridgiano de Orbagnoux (Francia), posee pared puntuada, pero es considerablemente más pequeña. Cymatiosphaera cf. wetzeli Deflandre 1954, del Terciario de Australia, posee vesícula y pared finamente puntuada, pero tamaño menor (44 µm).
Grupo Acritarcha Evitt 1963
Subgrupo Acanthomorphitae Downie, Evitt y Sarjeant 1963
Género Filisphaeridium Staplin, Jansonius y Pocock 1965, emend. Sarjeant y Stancliffe 1994
Especie tipo. Filisphaeridium setasessitante (Jansonius 1962) Staplin, Jansonius y Pocock 1965.
La enmienda realizada por Sarjeant y Stancliffe (1994, p. 28) para el género Filisphaeridium Staplin, Jansonius y Pocock 1965, permite incluir ejemplares con "vesícula esférica a subesférica, de pared delgada, lisa, levemente granular a sagrinada, con espinas distribuidas más o menos uniformemente. Las espinas son cortas, sólidas y más o menos rígidas, cilíndricas, a veces con una leve expansión proximal y distalmente acuminadas, capitadas o clavadas. Se presentan en un número mayor a 35, a menudo muy superior y constituyen como una densa cubierta pilosa, cuya longitud es menor que el 20% del diámetro del cuerpo de la vesícula. La apertura de la vesícula puede darse por una criptosutura".
Filisphaeridium balmei (Sarjeant 1973) Sarjeant y Stancliffe 1994
Figuras 8.J y 9.M
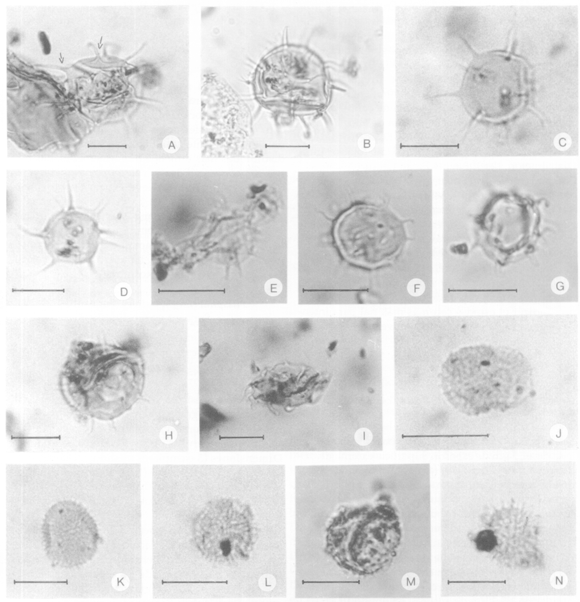
Figura 8. A, Baltisphaeridium sp., UNSP PP 1793c: Z10/4, las flechas muestran un detalle de las espinas huecas sin comunicación con el interior de la vesícula / the arrows show hollow spines without communication with the central body cavity. B, Micrhystridium brevispinosum (Sarjeant) Sarjeant y Stancliffe, UNSP PP 1795c: F48. C, Micrhystridium fragile Deflandre, UNSP PP 1796c: S54/3. D-E, Micrhystridium inconspicuum (Deflandre) Deflandre 1937; emend. Deflandre y Sarjeant, D, UNSP PP 1796c: O20/1; E, UNSP PP 1791c: E20/3. F-G, Micrhystridium cf. M. inconspicuum (Deflandre) Deflandre 1937, emend. Deflandre y Sarjeant, F, UNSP PP 1796c: G62; G, UNSP PP 1796c: E57/2. H-I, Micrhystridium recurvatum Valensi, H, UNSP PP 1789c: D50/2; I, UNSP PP 1792c: C47/1. J, Filisphaeridium balmei (Sarjeant 1973) Sarjeant y Stancliffe, UNSP M 1826: W21/3. K-L, Filisphaeridium densispinum (Valensi 1953) Sarjeant y Stancliffe, K, UNSP PP 1796c: O53/4; L, UNSP PP 1796c: N48/2. M-N, Filisphaeridium castaninum (Valensi 1953) Sarjeant y Stancliffe, M, UNSP PP 1796c: C57; N, UNSP PP 1796c: G54. Todas las fotomicrografías están ampliadas 1000X. Escala gráfica= 10 µm / All are 1000X. Scale bar equals 10 µm.

Figura 9. Todas las fotomicrografías están ampliadas 1000X. Escala gráfica = 10 µm / All are 1000X. Scale bar equals 10µm. A-B, Cymatiosphaera sp. 1, UNSP PL 1466a: 9,4/137,2, A, Enfoque distal / distal focus, B, Enfoque proximal / proximal focus. C-D, Cymatiosphaera sp. 2, UNSP PL 1466a: W59/4, C, Enfoque proximal / proximal focus; D, Enfoque distal / distal focus. E-F, Cymatiosphaera sp. 3, E, UNSP PL 1466 a: D51/2; F, UNSP PL 1466 a: 21,4/139,1. G, Escharisphaeridia pocockii (Sarjeant) Erkmen y Sarjeant, UNSP PL 1429c: L18/3. H, Cleistosphaeridium sp., UNSP PP 1793c:V41/3. I-L, Micrhystridium echinoides cf. M. echinoides forma minor Valensi 1953, UNSP PP 1796c: K46/1. M, Filisphaeridium balmei (Sarjeant 1973) Sarjeant y Stancliffe, UNSP PL 1466a: X44/3. N, Filisphaeridium cf. F. balmei (Sarjeant 1973) Sarjeant y Stancliffe, UNSP PL 1466a: 24,1/143,6.
Material estudiado. UNSP PL 1466a: X44/3 (figura 7.M). UNSP PP 1792c: G38/1. UNSP M 1826c: W21/3 (figura 8.J).
Descripción. Vesícula ovoidal, profusamente ornamentada por un gran número de espinas (100-150), extremadamente cortas y de distribución regular. En un ejemplar se observa la presencia de una hendidura.
Dimensiones (3 ejemplares medidos).
Diámetro máximo de la vesícula: 10,4-(13,1)-16 µm; longitud máxima de las espinas: 0,5-(0,58)-0,65 µm.
Distribución. Perfiles Puente Picún Leufú, Puesto Policía y Los Molles.
Observaciones. Esta especie fue descripta por Sarjeant (1973) para el Triásico del oeste de Pakistán y constituye la primera mención para la cuenca Neuquina. Las dimensiones de los especímenes estudiados se corresponden con el rango de dimensiones del material original de Sarjeant (diámetro de la vesícula: 10-(16,06)-22 µm, longitud de las espinas: 0,33- (0,45)-0,75 µm), si bien los valores promedio son un tanto diferentes.
Filisphaeridium castaninum (Valensi 1953) Sarjeant y Stancliffe 1994
Figuras 6.P-Q y 8.M-N
Material estudiado. UNSP PP 1791c: E27/4; 1793c: E57/2 (figuras 5.P-Q); 1796c: C57 (figura 8.M), G54 (figura 8.N), J49/4.
Descripción. Vesícula esferoidal a subesferoidal, portadora de numerosas espinas pequeñas, uniformemente distribuidas, delgadas, rectas, ocasionalmente flexibles a levemente curvadas, acuminadas, similares a pelos. El número de espinas del material presentado es difícilmente determinable dado el pequeño tamaño tanto de la vesícula como de las espinas. En el borde ecuatorial de los ejemplares medidos se ha determinado un número de espinas promedio de 31, por lo que se asume que el número total de las mismas supera ampliamente ese valor. En ejemplares bien preservados se han contado más de 50 espinas (figura 5.P). La liberación del contenido interno de la vesícula se da por formación de una hendidurra lineal (figura 8.M) o un piloma subpoligonal (figura 5.Q). Los ejemplares estudiados exhiben una coloración castaño medio.
Dimensiones (5 ejemplares medidos). Diámetro mayor de la vesícula: 11-(12,6)-15 µm; diámetro menor de la vesícula: 10-(11,8)-14 µm; longitud máxima de las espinas: 1,5-(1,9)-3 µm; número aproximado de espinas en el contorno ecuatorial: 28-(31)-35.
Distribución. Perfil Puesto Policía.
Observaciones. Constituye la primera mención para la cuenca Neuquina.
Comparaciones. El rango de dimensiones de la vesícula de los ejemplares estudiados es levemente superior al del material tipo de Valensi (1953) (8-13 µm), sin embargo coincide con las dimensiones de los ejemplares presentados por Sarjeant (1973) para el Triásico de Pakistán.
Filisphaeridium densispinum (Valensi 1953) Sarjeant y Stancliffe 1994
Figuras 8.K-L
Material estudiado. UNSP PP 1792c: E53/2-4, E57; 1796c: N48/2 (figura 8.L), O53/4 (figura 8.K).
Descripción. Vesícula esferoidal a subesferoidal, portadora de un gran número de espinas de distribución extremadamente regular, pequeñas, delgadas, rectas y acuminadas, similares a pelos. La longitud de las mismas es del orden del micrómetro. Su número total es difícilmente determinable pero en el borde ecuatorial pueden contarse alrededor de 50 espinas. Los ejemplares estudiados exhiben una coloración castaño claro.
Dimensiones (4 ejemplares medidos). Diámetro mayor de la vesícula: 9-(10,6)-12 µm; diámetro menor de la vesícula: 8-(9,4)-10,5 µm.
Distribución. Perfil Puesto Policía.
Observaciones. Constituye la primera mención para la cuenca Neuquina. Tanto Filisphaeridium castaninum como F. densispinum han sido descriptos por Valensi (1953) para el Jurásico medio (Bajociano-Bathoniano) de Francia.
Género Micrhystridium (Deflandre 1937), emend. Sarjeant y Stancliffe 1994
Especie tipo. Micrhystridium inconspicuum (Deflandre 1935) Deflandre 1937, emend Deflandre y Sarjeant 1970.
Micrhystridium echinoides cf. M. echinoides forma minor Valensi 1953
Figura 9.I-L
Material estudiado. UNSP PP 1796c: K46/1.
Descripción. Vesícula subesferoidal, psilada, portadora de espinas huecas, rectas, flexibles hasta recurvadas, que se ensanchan en sus bases, adoptando formas bulbosas en un número aproximado de 15.
Dimensiones (1 ejemplar medido). Diámetro mayor de la vesícula: 11 µm; diámetro menor de la vesícula: 9 µm; longitud máxima de las espinas: 5 µm.
Distribución. Perfil Puesto Policía.
Observaciones. Courtinat (1983), sinonimizó M. echinoides con Micrhystridium inconspicuum inconspicumm, sin embargo esto no fue aceptado por Sarjeant y Stancliffe (1994). En Martínez et al. (2001) se hace una mención previa de esta especie para el Jurásico medio de la cuenca Neuquina (Fm. Lajas). Se compara y no se asigna, dado el estado de preservación del material y el menor número de espinas determinado en comparación con el material tipo (25-30 espinas).
Micrhystridium cf. M. gregarium Sarjeant 1973
Figuras 6.K-L
Material estudiado. UNSP PL 1466a: Q46/4 (figuras 6.K-L), agrupamiento de M. cf. M. gregarium.
Descripción. Vesícula ovoidal a subesferoidal, ornamentada con espinas (~30) delgadas, huecas, cerradas distalmente, rectas a levemente curvadas. La totalidad de los individuos (~40) se presentan formando un agrupamiento.
Dimensiones (6 ejemplares medidos). El bajo número de ejemplares medidos se debe a la superposición de ejemplares y al mal estado de preservación de los mismos. Diámetro mayor de la vesícula: 18-(20)-21 µm; diámetro menor de la vesícula: 13-(14)-15 µm; longitud de las espinas: 1-(2)-3 µm.
Distribución. Perfil Puente Picún Leufú.
Comparaciones. Se compara y no se asigna con certeza a M. gregarium Sarjeant 1973 debido a que éste posee mayor longitud de las espinas (3,5-7,5 µm). Los ejemplares identificados por Sarjeant (1973) para el Triásico (Anisiano) del oeste de Pakistán, también fueron encontrados bajo la forma de un agrupamiento (18 individuos).
Observaciones. En coincidencia con Sarjeant, 1973 (p. 54), se piensa que los ejemplares de M. cf. M. gregarium que conforman el agrupamiento, fueron liberados como tal y se descarta la posibilidad de una asociación posterior debido a que: a) No existen indicios de coagulación en otros sitios del preparado estudiado; b) El agrupamiento es de tipo monoespecífico y c) No se identificaron ejemplares aislados en el preparado estudiado fuera del mencionado agrupamiento.
Micrhystridium inconspicuum (Deflandre 1935) Deflandre 1937, emend Deflandre y Sarjeant 1970
Figuras 8.D-E
Material estudiado. UNSP PP 1791c: E20/3(figura 8.E); 1796c: G61/4, O20/1(figura 8.D).
Dimensiones (3 ejemplares medidos). Diámetro mayor de la vesícula: 9,6-(10,7)-11,5 µm; diámetro menor de la vesícula: 7-(8,7)-11 µm; longitud máxima de las espinas: 4-(5,2)-6 µm; número de espinas: 15-20
Distribución. Perfil Puesto Policía.
Micrhystridium cf. M. inconspicuum (Deflandre 1935) Deflandre 1937, emend. Deflandre y Sarjeant 1970
Figuras 8.F-G
Material estudiado. UNSP PP 1791c: D26/1, E26/4, C21, C27, C28, C29, D23, D19, 22,2/120,7; 22/115; 22/121,5; 21,9/110. UNSP PP 1793c: D47/1; 1796c: E57/2 (figura 8.G), G62 (figura 8.F).
Descripción. Vesícula esferoidal a subesferoidal, de pared lisa (observada al microscopio óptico), ornamentada con espinas huecas (al menos en sus bases), delgadas, distalmente cerradas y acuminadas, rectas, flexibles, hasta recurvadas. Sus longitudes varían entre 1/5 y 2/5 del diámetro de la vesícula. El número de espinas varía entre 15 y 21, siendo 18 el valor más frecuente. En algunos ejemplares se observan pilomas subpoligonales o hendiduras ("slit").
Dimensiones (16 ejemplares medidos). Diámetro mayor de la vesícula: 7-(9,6)-12 µm; diámetro menor de la vesícula: 6-(8,6)-12 µm; longitud máxima de las espinas: 1,6-(3,05)-4 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. La enmienda de la especie tipo del género Micrhystridium, Micrhystridium inconspicuum (Deflandre 1935) Deflandre 1937 realizada por Deflandre y Sarjeant (1970) permite incluir ejemplares cuyas características son: "vesícula globular o levemente polihedral, portadora de espinas simples cuyo número varía entre 18 y 35, distribuídas en toda la superficie de la misma. Las espinas son huecas y sus cavidades se comunican directamente con el interior de la vesícula. Sus bases pueden estar más o menos engrosadas y disminuir su ancho en forma progresiva distalmente. Pueden ser rectas y rígidas, pero más a menudo levemente arqueadas, recurvadas o flexibles. Su longitud varía entre 1/2 y 2/3 del diámetro de la vesícula. Su superficie carece de otro tipo de ornamentación visible con microscopio óptico. Su diámetro varía entre 5 y 14 µm" (tomado de Sarjeant y Stancliffe 2000). Los ejemplares aquí estudiados coinciden con las características generales de la especie tipo, sin embargo la relación entre longitud de espinas y diámetro de la vesícula es menor (entre 1/5 y 2/5 del diámetro de la vesícula), razón por lo cual se prefiere comparar y no asignar a M. inconspicuum (Deflandre 1935) Deflandre 1937 emend. Deflandre y Sarjeant 1970.
Micrhystridium recurvatum Valensi 1953
Figuras 8.H-I
Material estudiado. UNSP PL 1466a: S25/4, E8/1. UNSP PP 1789c: D50/2 (figura 8.H); 1792c: C47/1(figura 8.I).
Dimensiones (6 ejemplares medidos). Diámetro mayor de la vesícula: 17-(20)-23 µm; diámetro menor de la vesícula: 10-(15,2)-18 µm; longitud de las espinas: 2-(4,3)-8 µm.
Distribución. Perfiles Puente Picún Leufú y Puesto Policía.
Género Baltisphaeridium (Eisenack 1958a), emend. Eiserhardt 1989
Especie tipo. Baltisphaeridium longispinosum (Eisenack 1931) Eisenack 1958a.
Baltisphaeridium sp.
Figura 8.A
Material estudiado. UNSP PP 1793c: Z10/4.
Descripción. Vesícula subesferoidal, psilada (al ser observada con microscopio óptico), de coloración levemente más oscura que las espinas, las que se presentan en un número de aproximadamente 15. Las espinas son delgadas, acuminadas, cerradas distalmente, flexibles hasta totalmente recurvadas, huecas, pero sólidas en sus bases por lo que no se comunican con la cavidad interna de la vesícula.
Dimensiones (1 ejemplar medido). Diámetro mayor de la vesícula: 21µm; diámetro menor de la vesícula: 18 µm; longitud máxima de las espinas: 11 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. El género Baltisphaeridium (Eisenack 1958a), emend. Eiserhardt 1989 ha sido escasamente citado para la cuenca Neuquina. Difiere de Baltisphaeridium eypensis Wall 1965, por la micropuntuación de la pared de la vesícula y menor longitud de las espinas (hasta 7 µm). Es similar a B. jansonii Pocock 1972, sin embargo todos los ejemplares ilustrado por Pocock presentan espinas rectas o flexibles (pero nunca totalmente recurvadas) y vesícula escabrada ("roughened").
Subgrupo Polygonomorphitae Downie, Evitt y Sarjeant 1963
Género Veryhachium Deunff 1954, emend. Sarjeant y Stancliffe 1994
Especie tipo. Veryhachium trisulcum (Deunff 1951) Deunf 1954 ex Downie 1959.
Veryhachium valensii (Valensi 1953) Downie y Sarjeant 1965, emend. Sarjeant 1970
Figura 6.O
Material estudiado. UNSP PL 1429a: G30.
Descripción. Vesícula triangular (triángulo isósceles), de lados rectos a levemente convexos. En cada uno de los vértices se ubica una espina pequeña. Sobre uno de los lados, cercano al vértice se observa una hendidura, con opérculo adherido.
Dimensiones (1 ejemplar medido). Longitud del lado más largo: 11 µm; longitud de las espinas <2 µm.
Distribución. Perfil Puente Picún Leufú.
Observaciones. Constituye ésta la primera mención para la cuenca Neuquina.
Género Polygonium Vavrdová 1966, emend. Sarjeant y Stancliffe 1994
Especie tipo. Polygonium gracile Vavrdová 1966, emend. Jacobson y Achab 1985.
Sarjeant y Stancliffe (1994, p. 43) enmiendan el género Polygonium Vavrdová 1966, incluyendo ejemplares con "vesícula hueca, poligonal a subpoligonal, generalmente con un diámetro superior a 20 µm, de pared lisa a granulada, delgada, ornamentada con 11 o más espinas huecas, simples y homomórficas, distribuídas en más de un plano de la vesícula. Las espinas son distalmente acuminadas, cerradas y a veces sólidas, y proximalmente pueden estar relativamente ensanchadas. Cuando las espinas son huecas, el interior de las mismas se comunica libremente con la cavidad de la vesícula, sin que exista una diferenciación aparente entre la pared de las espinas y la vesícula. La apertura de la vesícula es por criptosutura". Si bien el rango estratigráfico conocido para este género es Cámbrico medio a Jurásico, es más representativo del Paleozoico Temprano (Cámbrico medio- Devónico). Hasta el presente han sido descriptas sólo dos especies formales para el Jurásico: Polygonium ? aster (Sarjeant 1967) Erkmen y Sarjeant 1980, del Jurásico medio de Francia y Polygonium jurassicum Bucefalo Palliani, Riding y Torricelli 1996, del Toarciano temprano de Italia y Grecia. En Martínez et al. (2001) se hace la primera mención del género para la cuenca Neuquina.
Polygonium sp. cf. ?P. jurassicum Bucefalo Palliani, Riding y Torricelli 1996
Figura 6.M
Material estudiado. UNSP PP 1792c: F45/3.
Descripción. Vesícula hueca, subesferoidal, de pared delgada y microescabrada. Las espinas son huecas, simples y homomórficas. Distalmente son cerradas y acuminadas, mientras que proximalmente sus bases se ensanchan hasta 3 µm, permitiendo una libre comunicación entre la cavidad de las espinas y el interior de la vesícula. Las espinas poseen diferente longitud y se distribuyen en diversos planos de la vesícula, por lo que su número exacto es difícilmente determinable, pudiendo contarse más de 20.
Dimensiones (1 ejemplar medido). Diámetro máximo de la vesícula: 21 µm; longitud máxima de las espinas: 12 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. Polygonium sp. (en Martínez et al. 2001, p. 56, lám. 6, figs. 4-5), es muy similar pero de tamaño considerablemente menor (diámetro menor de la vesícula: 11,1-(12,1)-13,1 µm, diámetro mayor de la vesícula: 13,5-(15,1)-16,7 µm y longitud máxima de las espinas: 8,1-(9)-9,8 µm). El presente material podría incluirse perfectamente en Polygonium jurassicum Bucefalo Palliani, Riding y Torricelli 1996 (lám. 1, figs. 1-9) sin embargo estos autores en la descripción de la especie destacan la falta de comunicación entre la cavidad de las espinas y el interior de la vesícula. En el ejemplar de la cuenca Neuquina dicha comunicación es evidente.
Observaciones. La descripción de Polygonium jurassicum Bucefalo Palliani et al. 1996 entra en conflicto con la enmienda realizada por Sarjeant y Stancliffe (1994) para el género Polygonium (p. 43) quienes especifican que "cuando las espinas son huecas, su interior se comunica libremente con el interior de la cavidad de la vesícula".
Polygonium sp.
Figura 6.N
Material estudiado. UNSP PP 1791c: E28/2.
Descripción. Vesícula hueca, subpoligonal, de pared lisa, portadora de espinas (aproximadamente 15), huecas, distalmente cerradas, acuminadas, rectas a levemente curvadas, que se ensanchan en sus bases (hasta 2 µm). Se observa una criptosutura poligonal.
Dimensiones (1 ejemplar medido). Diámetro mayor de la vesícula: 13 µm; diámetro menor de la vesícula: 9 µm; longitud máxima de las espinas: 5 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. Polygonium sp. (en Martínez et al. 2001) presenta vesícula y espinas microgranuladas y un rango de dimensiones levemente mayor.
Subgrupo Sphaeromorphitae Downie, Evitt y Sarjeant 1963
Género Leiosphaeridia Eisenack 1958b, emend. Downie y Sarjeant 1963
Especie tipo. Leiosphaeridia baltica Eisenack 1958b.
El género Leiosphaeridia fue creado por Eisenack (1958b) para incluir formas esféricas de pared lisa o escasamente ornamentada y desde ese momento ha sido motivo de debate y controversia. Pocock (1972, p. 104), considera a este taxón como un grupo misceláneo que incluye quistes de dinoflagelados, algas clorofitas y otros organismos de afinidad desconocida. Dada su afinidad botánica incierta fue considerado como acritarco por varios autores: Lentin y Williams (1993, p. 724), Colbath y Grenfell (1995), Sarjeant (2002), entre otros. Fensome et al. (1990, p. 4), consideran que sólo los géneros Pterospermella Eisenack 1972 y Tasmanites Newton 1875 pueden ser considerados indiscutiblemente como prasinofitas, dado que la inclusión de Leiosphaeridia en este grupo, no ha tenido un consenso universal (Colbath, 1979, 1983).
Tappan (1980) incluyó al género Leiosphaeridia dentro de las algas verdes, como Prasinophyceae basado en la similitud de estas formas fósiles con el ficoma del género actual Halosphaera, criterio que ha sido aceptado por varios autores. Sin embargo, según Sarjeant (2002, p. 299), "la morfología extremadamente simple de las leiosfaeras no da indicaciones convincentes de su afinidad a las prasinofitas. Es probable que representen un grupo incertae sedis y como tal deberían ser tratadas como acritarcos dentro del Subgrupo Sphaeromorphitae".
Colbath y Grenfell (1995, p. 296-297), exponen una interesante discusión acerca de la afinidad incierta de este género basada en diferencias de la ultraestructura de la pared de las formas fósiles y Halosphaera. Estudios de la pared de las formas actuales revelan características estructuras trilaminares, las que no son evidentes en imágenes de ultraestructura de Leiosphaeridia. Concluyen que la afinidad del género Leiosphaeridia a las Prasinophyceae es plausible sólo para algunas especies del género (L. wenlockia Downie 1959 de edad silúrica), y dado su carácter polifilético deberían intensificarse los estudios de composición química y utraestructura tanto de las formas fósiles como las actuales. Talyzina y Moczydlowska (2000), mencionan por primera vez estructuras de tipo tri-laminar para el género Leiosphaeridia, en su estudio del Cámbrico inferior de Estonia; y consideran esta caracterísitica como una evidencia de afinidad botánica con al menos dos clases de algas verdes: Prasinophyceae y Chlorophyceae. Lejos de estar resuelto el problema, las nuevas investigaciones confirman el marcado carácter polifilético del grupo, por lo que nuevos estudios de ultraestructura deberían realizarse con ejemplares de edad mesozoica para poder arribar a una conclusión confiable.
Leiosphaeridia cf. L. sp. F (en Prámparo 1989)
Figura 4.P
Material estudiado. UNSP PP 1790c: N50, S49 (figura 4.P), T50/2.
Descripción. Vesícula originalmente esferoidal, deformada fuertemente por plegamiento secundario, de pared delgada (aproximadamente 0,7 µm de espesor), densamente ornamentada por microgránulos de distribución regular. En algunos ejemplares se observa un cuerpo interno de coloración más oscura.
Dimensiones (4 ejemplares medidos). Diámetro mayor de la vesícula: 22-(32)-38 µm; diámetro menor de la vesícula: 16-(24)-28 µm; diámetro mayor del cuerpo interno: 8-(10)-13 µm; diámetro menor del cuerpo interno: 5-(6,7)-8 µm.
Distribución. Perfil Puesto Policía.
Comparaciones. Leiosphaeridia sp. F (en Prámparo 1989), presenta un rango de dimensiones menor (18- 25 µm) y menor grado de plegamiento.

Figura 10. Muestras palinológicas y frecuencias relativas de los taxones identificados en el perfil Puesto Policía / Palynological samples and relative frequencies of the identified taxa from Puesto Policía section.

Figura 11. Muestras palinológicas y frecuencias relativas de los taxones identificados en el perfil Los Molles / Palynological samples and relative frequencies of the identified taxa from Los Molles section.

Figura 12. Muestras palinológicas y frecuencias relativas de los taxones identificados en el perfil Puente Picún Leufú / Palynological samples and relative frequencies of the identified taxa from Puente Picún Leufú section.
Conclusiones
El estudio palinológico de muestras provenientes de afloramientos de la Formación Los Molles en el sector centro-occidental de la cuenca Neuquina, Argentina (figuras 10-12), permitió identificar 50 taxones de esporomorfos, 20 correspondientes a esporas de briofitas y pteridofitas y 30 a granos de polen. De ellos se describieron ejemplares pertenecientes a los géneros Divisisporites, Callialasporites y Monosulcites, además de una espora cuyos género y especie son indeterminados. Dentro del paleomicroplancton se identificaron 31 taxones distintos, con el siguiente detalle, acritarcos: 20, dinoflagelados: 2, algas verdes (Chlorococcales y Prasinophyceae): 9. Entre las prasinofíceas se describieron especies pertenecientes a Cymatiosphaera (tres nuevas especies), Pleurozonaria y Tasmanites y entre los acritarcos especies de Baltisphaeridium, Filisphaeridium, Micrhystridium, Leiosphaeridia, Polygonium y Veryhachium. Se propone la nueva combinación Cymatiosphaera volkheimerii (Quattrocchio 1980). Se menciona por primera vez para la cuenca Neuquina la presencia de los acritarcos: Filisphaeridium balmei, F. castaninum, F. densispinum y Veryhachium valensii. Si bien en la palinoflora hallada en la Formación Los Molles no se encontraron formas con un gran valor bioestratigráfico, por su composición, se la puede correlacionar con la Biozona de Callialasporites "complex", Sub-biozonas de Callialasporites trilobatus (Balme) Dev, Antulsporites saevus (Balme) Archangelsky y Gamerro y Klukisporites labiatus (Volkheimer) Baldoni y Archangelsky, establecidas por Martínez (2002) para esta misma área de trabajo, determinada a partir del análisis secuencial y datación con fauna de amonites. La edad sugerida para la Formación Los Molles en el centro-oeste de la cuenca Neuquina está comprendida entre el Aaleniano tardío y Bajociano temprano.
Apéndice A. Lista de especies identificadas
Esporomorfos Esporas de briofitas y pteridofitas Antulsporites saevus (Balme) Archangelsky y Gamerro 1966
Antulsporites varigranulatus (Levet-Carette) Reiser y Williams 1969 (figura 3.D)
Auritulinasporites spp.
Biretisporites spp. ?
Camptozonotriletes sp. 1 (en Martínez, Quattrocchio y Sarjeant 2001)
Clavatisporites cf. C. bagualensis (Volkheimer) Martínez, Quattrocchio y Sarjeant 2001
Deltoidospora australis (Couper) Pocock 1970
Deltoidospora minor (Couper) Pocock 1970
Deltoidospora neddeni Pflug 1953
Deltoidospora spp.
Dictyophyllidites mortoni (de Jersey) Playford y Dettmann 1965 (figura 3.B)
Dictyophyllidites spp. Divisisporites sp. (figura 3.A) espora gn. et sp. indet. (figura 3.C)
Gleicheniidites spp.
Klukisporites variegatus Couper 1958
Klukisporites spp.
Leptolepidites spp.
Nevesisporites cf. N. radiatus (Chlonova) Srivastava 1972
Punctatosporites scabratus (Couper) Norris 1965
Retitriletes spp. Rugulatisporites neuquenensis Volkheimer 1972 (figura 3.E)
Verrucosisporites varians Volkheimer 1972
Verrucosisporites spp.
Gimnospermas
Alisporites spp.
Araucariacites australis Cookson 1947
Araucariacites fissus Reiser y Williams 1969
Araucariacites pergranulatus Volkheimer 1968
Callialasporites dampieri (Balme) Dev 1961
Callialasporites microvelatus Schulz 1966 (figuras 3.J y L)
Callialasporites segmentatus (Balme) Srivastava 1963
Callialasporites trilobatus (Balme) Dev 1961
Callialasporites turbatus (Balme) Schulz 1967
Callialasporites sp. (figura 3.K)
Callialasporites spp.
Cerebropollenites macroverrucosus (Thiergart) Schulz 1967 (figura 3.H)
Cerebropollenites cf. C. macroverrucosus (Thiergart) Schulz 1967 (figura 3.G)
Classopollis cf. C. classoides (Pflug) Pocock y Jansonius 1961
Classopollis intrareticulatus Volkheimer 1972 (figura 3.F)
Classopollis itunensis Pocock 1962 Classopollis major Groot y Groot 1962
Classopollis simplex (Danzé, Corsin y Laveine) Reiser y Williams 1969
Classopollis torosus (Reissinger) Burger 1965
Classopollis cf. C. torosus (Reissinger) Burger 1965 (en Volkheimer 1968)
Classopollis spp. Cycadopites adjectus (de Jersey) Volkheimer y Quattrocchio 1975
Cycadopites cf. C. adjectus (de Jersey) Volkheimer y Quattrocchio 1975
Cycadopites nitidus (Balme) de Jersey 1964
Cycadopites spp. Inaperturopollenites indicus Srivastava 1966
Inaperturopollenites microgranulatus Volkheimer 1972
Inaperturopollenites spp. Microcachryidites castellanosii Menéndez 1968 (figuras 3.N-O)
Monosulcites sp. A (en Volkheimer 1972)
Monosulcites sp. cf. M. sp. B (en Volkheimer y Quattrocchio 1975) (figura 3.I)
Monosulcites spp. Phrixipollenites sp. (figura 3.M)
Podocarpidites cf. P. ellipticus Cookson 1947
Podocarpidites spp. Vitreisporites pallidus (Reissinger) Nilsson 1958
Paleomicroplancton
Algas verdes (Chlorococcales y Prasinophyceae)
Botryococcus spp. (figura 4.Q)
Cymatiosphaera eupeplos (Valensi) Deflandre 1954 (figura 4.A)
Cymatiosphaera cf. C. eupeplos (Valensi) Deflandre 1954 (figuras 4.B-C)
Cymatiosphaera cf. C. volkheimerii (Quattrocchio) n. comb. (figuras 4.D-L)
Cymatiosphaera sp. 1 (figuras 7 y 9.A-B)
Cymatiosphaera sp. 2 (figuras 9.C-D)
Cymatiosphaera sp. 3 (figuras 9.E-F)
Cymatiosphaera "complex" Pleurozonaria cf. P. picunensis Quattrocchio 1980 (figuras 4.M-O)
Pleurozonaria spp. Tasmanites sp. (figuras 5 y 6.A-J)
Quistes de dinoflagelados
Cleistosphaeridium sp. (figura 9.H)
Escharisphaeridia pocockii (Sarjeant) Erkmen y Sarjeant 1980 (figura 9.G)
Acritarcos
Baltisphaeridium sp. (figura 8.A)
Filisphaeridium balmei (Sarjeant 1973) Sarjeant y Stancliffe 1994 (figuras 8.J y 9.M)
Filisphaeridium cf. F. balmei (Sarjeant 1973)
Sarjeant y Stancliffe 1994 (figura 9.N)
Filisphaeridium castaninum (Valensi 1953)
Sarjeant y Stancliffe 1994 (figuras 6.P-Q y 8.M-N))
Filisphaeridium densispinum (Valensi 1953)
Sarjeant y Stancliffe 1994 (figuras 8.K-L)
Leiosphaeridia sp. B (en Volkheimer, Caccavari y Sepúlveda 1977)
Leiosphaeridia sp. E (en Prámparo 1989)
Leiosphaeridia cf. L. sp. F (en Prámparo 1989) (figura 4.P)
Leiosphaeridia sp. cf. L. hyalina (Deflandre) Downie 1957 (en Quattrocchio 1980)
Leiosphaeridia spp. Micrhystridium brevispinosum (Sarjeant) Sarjeant y Stancliffe 1994 (figura 8.B)
Micrhystridium echinoides cf. M. echinoides forma minor Valensi 1953 (figuras 9.I-L)
Micrhystridium fragile Deflandre 1947 (figura 8.C)
Micrhystridium cf. M. gregarium Sarjeant 1973 (figuras 6.K-L)
Micrhystridium inconspicuum (Deflandre) Deflandre 1937; emend . Deflandre y Sarjeant 1970 (figuras 8.D-E)
Micrhystridium cf. M. inconspicuum (Deflandre) Deflandre 1937; emend . Deflandre y Sarjeant 1970 (figuras 8.F-G)
Micrhystridium nannacanthum Deflandre 1945
Micrhystridium recurvatum Valensi 1953 (figuras 8.H-I)
Micrhystridium spp. Polygonium sp. cf. ? P. jurassicum Bucefalo Palliani, Riding y Torricelli 1996 (figura 6.M)
Polygonium sp. (figura 6.N)
Veryhachium valensii (Valensi) Downie y Sarjeant 1965 (figura 6.O)
Agradecimientos
Los autores desean expresar su agradecimiento a C. Zavala por proveer las muestras para la realización de este trabajo. Al Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) y la Secretaría General de Ciencia y Tecnología de la Universidad Nacional del Sur (SeGCyT) por el apoyo económico brindado. A J. Helenes Escamilla y al Comité Editor por la lectura y sugerencias recibidas oportunamente.
Bibliografía
1. Abbink, O.A. 1998. Palynological investigations in the Jurassic of the North Sea region. Laboratory of Paleobotany and Palynology Contributions Series 8. Laboratory of Paleobotany and Palynology Foundation, Utrecht, 192 pp. [ Links ]
2. Al-Almery, T.K. 1983. Acid resistant microfossils implemented for the determination of Palaeozoic palaeoenvironments in Libya. Palaeogeography, Palaeoclimatology, Palaeoecology 44: 103-116. [ Links ]
3. Al-Almery, T.K. 1986. Observations in the wall structure and excystment mechanism of acritarchs. Journal of Micropalaeontology 5: 27-38. [ Links ]
4. Balme, B.E. 1957. Spores and pollen grains from the Mesozoic of Western Australia. Commonwealth Scientific and Industrial Research Organization (Australia), Coal Research Section, Reference T.C. 25: 1-48. [ Links ]
5. Brocke, R. y Riegel, W. 1996. Phytoplankton responses to shoreline fluctuations in the Upper Muschelkak (Middle Triassic) of Lower Saxony (Germany). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 200: 53-73. [ Links ]
6. Bucefalo Palliani, R., Riding, J.B. y Torricelli, S. 1996. Polygonium jurassicum sp. nov., a polygonomorph acritarch from the Lower Toarcian (Lower Jurassic) of the Tethyan Realm. Palynology 20: 157-161. [ Links ]
7. Colbath, G.K. 1979. Organic-walled microphytoplankton from the Eden Shale (Upper Ordovician), Indiana, U.S.A. Palaeontographica B 171: 1-38. [ Links ]
8. Colbath, G.K. 1983. Fossil prasinophycean phycomata (Chlorophyta) from the Silurian Bainbridge Formation, Missouri, U.S.A. Phycologia 22: 249-265. [ Links ]
9. Colbath, G.K. y Grenfell, H.R. 1995. Review of biological affinities of Paleozoic acid-resistant, organic-walled eukaryotic algal microfossils (including "acritarchs"). Review of Palaeobotany and Palynology 86: 287-314. [ Links ]
10. Cookson, I.C. 1947. Plant microfossils from the lignites of Kerguelen Arhipelago. B.A.N.Z. Antarctic Research Expedition 1929-1931 , Reports-Ser . A (2), Part. 8: 127-142. [ Links ]
11. Cookson, I. y Manum, S. 1960. On Crassosphaera , a new genus of microfossils from Mesozoic and Tertiary Deposits. Nytt Magasin for Botanikk 8: 5-9. [ Links ]
12. Couper, R.A. 1953. Upper Mesozoic and Cainozoic Spores and Pollen Grains from New Zealand. New Zealand Geological Survey Paleontological Bulletin 22: 3-77. [ Links ]
13. Couper, R.A. 1958. British Mesozoic microspores and pollen grains. Palaeontographica 108 B: 75-179. [ Links ]
14. Courtinat, B. 1983. Evidence ou r éalité chez le genre Micrhystridium . Un exemple par l'étude de quelques formes jurassiques du Jura Français. Cahiers de Micropaléontologie 1: 1-32. [ Links ]
15. Davey, R. 1970. Non-calcareous microplankton from the Cenomanian of England, northern France and North America, Part II. Bulletin of the British Museum (Natural History), Geology 18: 335-397. [ Links ]
16. De Coninck, J. 1965. Microfossiles planctoniques du sable Ypr ésien à Merelbeke. Dinophyceae et Acritarcha. Académie royale des sciences, des lettres et des beaux-arts de Belgique, Classe des sciences, Mémories, Collection in-8, 36: 1-55. [ Links ]
17. Deflandre, G. 1935. Consid érations biologiques sur les microorganismes d'origine planctonique conservés dans les silex de la craie. Bulletin Biologique de la France et de la Belgique 69: 213-244. [ Links ]
18. Deflandre, G. 1937. Microfossiles des silex cr étacés. Deuxième partie. Flagellés incertae sedis . Hystrichosphaeridés. Sarcodinés. Organismes divers. Annales de Paléontologie 26: 51- 103. [ Links ]
19. Deflandre, G. 1941. Le microplankton Kim éridgien d'Orbagnoux et l'origine des huiles sulfurées naturelles. Mémoires de l'Académie des Sciences (de l'Institut de France) 65: 1-32. [ Links ]
20. Deflandre, G. 1954. Syst ématique des Hystrichosphaeridés: sur l'acception du genre Cymatiosphaera O Wetzel. Compte Rendu Sommaire des Séances de la Société Géologique de France 12: 257- 258. [ Links ]
21. Deflandre, G. y Sarjeant, W.A.S. 1970. Nouvel examen de quelques holotypes de Dinoflagell és fossiles et d'Acritarches. Cahiers de Micopaléontologie 2: 1-10. [ Links ]
22. Deunff, J. 1951. Sur la pr ésence de micro-organismes (Hystrichosphères) dans les schistes ordoviciens du Finistère. Compte Rendu Hebdomadaire des Séances de l'Académie des Sciences 233: 321- 323. [ Links ]
23. Deunff, J. 1954. Veryhachium , genre nouveau d'hystrichosphères du Primaire. Compte Rendu Sommaire des Séances de la Société Géologique de France 13: 305-307. [ Links ]
24. Dev, S. 1961. The fossil flora of the Jabalpur series. 3. Spores and Pollen grains. The Palaeobotanist 8: 43-56. [ Links ]
25. Downie, C. 1959. Hystrichospheres from the Silurian Wenlock Shale of England. Palaeontology 2: 56-71. [ Links ]
26. Downie, C. y Sarjeant, W.A.S. 1963. On the interpretation and status of some hystrichosphere genera. Palaeontology 6: 83-96. [ Links ]
27. Downie, C. y Sarjeant, W.A.S. 1965. Bibliography and index of fossil dinoflagellates and acritarchs. Geological Society of America, Memoir 94: 1-180. [ Links ]
28. Downie, C., Evitt, W. y Sarjeant, W.A.S. 1963. Dinoflagellates, hystrichospheres and the classification of the acritarchs. Stanford University Publications, Geological Sciences 7: 1-16. [ Links ]
29. Eisenack, A.E. 1931. Neue Mikrofossilien des baltischen Silurs 1. Palaeontologische Zeitschrift 13: 74-118. [ Links ]
31. Eisenack, A.E. 1958a. Mikroplankton aus dem Ordovizium des Baltikums. 1. Markasitschicht, Dictyonema -Schiefer, Glaukonitsand, Glaukonitkalk. Senckenbergiana Lethaea 39: 389-405. [ Links ]
32. Eisenack, A.E. 1958b. Tasmanites Newton 1875 und Leiosphaeridia n. g. als Gattungen der Hystrichosphaeridia. Palaeontographica A 110: 1-19. [ Links ]
33. Eisenack, A.E. 1972. Kritische Bemerkung zur Gattung Pterospermopsis (Chlorophyta, Prasinophyceae). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 10: 596-601. [ Links ]
34. Eiserhardt, K.H. 1989. Baltisph ären aus Gotländer Öjlemyrflint (Acritarcha, Oberordoviz, Geschiebe, Schweden). Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 68: 79-129. [ Links ]
35. Erdtman, G. 1947. Suggestions for the classification of fossil andecent pollen grains and spores. Svenska Botaniska Tidskrift 41: 104-114.
36. Erkmen, U. y Sarjeant, W.A.S. 1980. Dinoflagellate cysts, acritarchs and tasmanitids from the uppermost Callovian of England and Scotland: with a reconsideration of the " Xanthidium pilosum " problem. Geobios 13: 45-99. [ Links ]
37. Fensome, R.A., Williams, G.L., Barss, M.S., Freeman, J.M. y Hill, J.M. 1990. Acritarchs and fossil prasinophytes: an index to genera, species and infraspecific taxa. American Association of Stratigraphic Palynologists, Contributions Series 25: 1-771. [ Links ]
38. Garc ía, V.M. 1998. [ Reconstrucción paleoambiental en base a palinofacies de las Formaciones Los Molles, Lajas y Challacó (Jurásico medio) en la Sierra de Chacaico y adyacencias. Cuenca Neuquina. Provincia de Neuquén. Argentina. Tesis Doctoral, Departamento de Geología, Universidad Nacional del Sur, Bahía Blanca, Argentina, 204 pp. Inédito] [ Links ].
39. Garc ía, V.M., Zavala, C.A. y Quattrocchio, M.E. 1994. Relación entre análisis palinológico y análisis de facies. Aplicación al Grupo Cuyo (Jurásico medio) en la cuenca Neuquina. Revista de la Asociación Geológica Argentina 49: 184-195. [ Links ]
40. Garc ía, V.M., Quattrocchio, M.E. y Zavala, C.A. 2000. Estudio palinofacial del Grupo Cuyo, Jurásico medio en la Sierra de Chacaico. Parte 1. Formación Los Molles. 11º Simposio Argentino de Paleobotánica y Palinología (Tucumán) Resúmenes , p.32. [ Links ]
41. Groeber, P. 1946. Observaciones geol ógicas a lo largo del meridiano 70. 1. Hoja Chos Malal. Revista de la Asociación Geológica Argentina 1: 177-208. [ Links ]
42. Gulisano, C.A., Guti érrez Pleimling, A.R. y Digregorio, R.E. 1984. Esquema estratigráfico de la secuencia jurásica del oeste de la provincia del Neuquén. 9º Congreso Geológico Argentino (S. C. Bariloche), Actas 1: 236-259. [ Links ]
43. Guy-Ohlson, D. 1988. Developmental stages in the life cycle of Mesozoic Tasmanites . Botanica Marina 31: 447-456. [ Links ] Guy-Ohlson, D. 1996. 7B. Prasinophycean algae. En: J. Jansonius y D.C. McGregor (eds.), Palynology: principles and applications. American Association of Stratigraphic Palynologists Foundation 1: 181-189. [ Links ]
44. Guy-Ohlson, D. y Boalch, G.T. 1992. Comparative morphology of the genus Tasmanites (Pterospermales, Chlorophyta). Phycologia 31: 523-528. [ Links ] Guy-Ohlson, D. y Norling, E. 1994. Jurassic sequences in Sweden. Geobios , M.S. 17: 275-286. [ Links ]
45. Jacobson, S.R. y Achab, A. 1985. Acritarch biostratigraphy of the Dicellograptus complanatus graptolite zone from the Vaureal Formation (Ashgillian), Anticosti Island, Quebec, Canada. Palynology 9: 165-198. [ Links ]
46. Jansonius, J. 1962. Palynology of Permian and Triassic sediments, Peace River area, western Canada. Palaeontographica B 110: 35-98. [ Links ] Lentin, J.K. y Williams, G.L. 1993. Fossil dinoflagellates: Index genera and species. American Association of Stratigraphic Palynologists - Contributions Series 28, 856 pp. [ Links ]
47. Mädler, K.A. 1963. Die figurierten organischen Bestandteile der Posidonienschiefer. Geologisches Jahrbuch Beihefte 58: 287-406. [ Links ]
48. Martínez, M.A. 1999. [ Palinología estratigráfica del Jurásico del centro- oeste de la cuenca Neuquina. Tesis Doctoral, Departamento de Geología, Universidad Nacional del Sur, Bahía Blanca, Argentina, 238 pp. Inédito] [ Links ].
49. Martínez, M.A. 2002. Palynological zonation of the Lajas Formation (Middle Jurassic) of the Neuquén Basin, Argentina. Ameghiniana 39: 221-240. [ Links ]
50. Martínez, M.A., Quattrocchio, M.E. y Sarjeant, W.A.S. 2001. Análisis palinoestratigráfico de la Formación Lajas, Jurásico Medio de la cuenca Neuquina, Argentina. Revista Española de Micropaleontología 33: 31-58. [ Links ]
51. Newton, E.T. 1875. On "Tasmanite" and Australian "white coal". Geological Magazine 12: 337-342. [ Links ]
52. Nilsson, T. 1958. Über das Vorkommen eines mesozoischen Sapropelgesteins in Schonen. Publications, Institutes of Mineralogy, Paleontology and Quaternary Geology , University of Lund 53: 1-111. [ Links ]
53. Pocock, S.A. 1972. Palynology of the Jurassic sediments of western Canada. Palaeontographica B 137: 85-153. [ Links ]
54. Pr ámparo, M.B. 1989. [ Palinología estratigráfica del Cretácico de la cuenca de San Luis. Tesis Doctoral, Universidad Nacional de Río Cuarto, Córdoba, 243 pp. Inédito] [ Links ].
55. Prauss, M. 1989. Dinozysten-Stratigraphie und Palynofazies im Oberen Lias und Dogger von NW-Deutschland. Palaeontographica B 214: 1-124. [ Links ]
56. Prauss, M. 1996. The Lower Toarcian Posidonia Shale of Grimmen, Northeast Germany. Implications from the palynological analysis of a near-shore section. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 200: 107-132. [ Links ]
57. Prauss, M. 2001. Sea-level changes and organic-walled phytoplankton response in a Late Albian epicontinental setting, Lower Saxony basin, NW Germany. Palaeogeography, Palaeoclimatology, Palaeoecology 174: 221-249. [ Links ]
58. Prauss, M. y Riegel, W. 1989. Evidence from phytoplankton associations for causes of black shale formation in epicontinental seas. Neues Jahrbuch für Geologie und Paläontologie , Mh . 1989, 11: 671-682. [ Links ]
59. Prauss, M., Ligouis, B. y Luterbacher, H. 1991. Organic matter and palynomorphs in the "Posidonienschiefer" (Toarcian, Lower Jurassic) of southern Germany. En: R.V. Tyson y T.H. Pearson (eds.), Modern and Ancient Continental Shelf Anoxia. Geological Society Special Publication 58: 335-351. [ Links ]
60. Quattrocchio, M.E. 1980. Contribuci ón al conocimiento de la palinología estratigráfica del Jurásico Superior en la cuenca neuquina. Opera Lilloana 31: 1-59. [ Links ]
61. Quattrocchio, M.E., Sarjeant, W.A.S. y Volkheimer, W. 1996a. Marine and terrestrial Jurassic microfloras of Neuqu én Basin (Argentina): Palynological Zonation. En: A.C. Riccardi, Advances in Jurassic research . Transtec Publications, Switzerland. GeoResearch Forum 1-2: 167-178. [ Links ]
62. Quattrocchio, M., Zavala, C., Garc ía, V. y Volkheimer, W. 1996b. Paleogeographic changes during the Middle Jurassic in the southern part of the Neuquén Basin, Argentina. En: A.C. Riccardi (ed.), Advances in Jurassic research . Transtec Publications, Switzerland. GeoResearch Forum 1-2: 467-484. [ Links ]
63. Riccardi, A.C. y Gulisano, C.A. 1990. Unidades limitadas por discontinuidades, su aplicaci ón al Jurásico Andino. Revista de la Asociación Geológica Argentina 45: 346-364. [ Links ]
64. Sarjeant, W.A.S. 1959. Microplankton from the Cornbrash of Yorkshire. Geological Magazine 96: 329-346. [ Links ]
65. Sarjeant, W.A.S. 1967. Observations on the acritarch genus Micrhystridium (Deflandre). Revue de Micropaléontologie 9: 201- 208. [ Links ]
66. Sarjeant, W.A.S. 1970. Acritarchs and Tasmanitids from the Chhidru Formation, uppermost Permian of West Pakistan. En: B. Kummel, y C. Teichert, (eds.), Stratigraphic boundary problems: Permian and Triassic of West Pakistan . Lawrence, Kansas: Department of Geology Special Publication, University of Kansas 4: 277-304. [ Links ]
67. Sarjeant, W.A.S. 1973. Acritarchs and Tasmanitids from the Mianwali and Tredian Formations (Triassic) of the Salt and Surghar Ranges, West Pakistan. En: The Permian and Triassic Systems and their Mutual Boundary . Canadian Society of Petroleum Geologists, Special Publication 2: 35-73. [ Links ]
68. Sarjeant, W.A.S. 2002. "As chimney-sweepers, come to dust": a history of palynology to 1970. En: D.R. Oldroyd (ed.), The Earth Inside and Out: Some Major Contributions to Geology in the Twentieth Century . Geological Society, London, Special Publications 192: 273-327. [ Links ]
69. Sarjeant, W.A.S. y Stancliffe, R.P.W. 1994. The Micrhystridium and Veryhachium complexes (Acritarcha: Acantomorphitae and Polygonomorphitae): a taxonomic reconsideration. Micropaleontology 40: 1-77. [ Links ]
70. Sarjeant, W.A.S. y Stancliffe, R.P.W. 2000. Acritarch taxonomy: certain controverted questions. Modern Geology 24: 159-176. [ Links ]
71. Scafati, L.H. y Morbelli, M.A. 1984. Nuevos datos palinol ógicos de la Formación Lajas. Jurásico medio de la cuenca Neuquina. 3º Congreso Agentino de Paleontología y Bioestratigrafía (Corrientes), Actas : 73- 105. [ Links ]
72. Schrank, E. 2003. Small acritarchs from the Upper Cretaceous: taxonomy, biological affinities and palaeoecology. Review of Palaeobotany and Palynology 123: 199-235. [ Links ]
73. Schulz, E. 1967. Sporenpal äontologische Untersuchungen rätoliassischer Schichten im Zentralteil des germanischen Beckens. Palaeontographica B 2: 427-633. [ Links ]
74. Staplin, F.L., Jansonius, J. y Pocock, S.A.J. 1965. Evaluation of some acritarchous hystrichosphere genera. Neues Jahrbuch für Geologie und Paläontologie , Abhandlungen 123: 167-201. [ Links ]
75. Talyzina, N.M. y Moczydlowska, M. 2000. Morphological and ultrastructural studies of some acritarchs from the Lower Cambrian L ükati Formation, Estonia. Review of Palaeobotany and Palynology 112: 1-21. [ Links ]
76. Tappan, H. 1980. The paleobiology of plant protists . W.H. Freeman and Company, San Francisco, 1028 p. [ Links ]
77. Thomson, P.W. y Pflug, H.D. 1953. Pollen und Sporen des mitteleurop äischen Tertiärs. Palaeontographica B 94: 1-138. [ Links ] Valensi, L. 1948. Sur quelques micro-organismes planctoniques des silex du Jurassique moyen du Poitou et de Normandie. Bulletin de la Société Géologique de France 18: 537-550. [ Links ]
78. Valensi, L. 1953. Microfossiles des silex du Jurassique moyen. Remarques p étrographiques. Mémoires de la Société Géologique de France 68: 1-100. [ Links ]
79. Vavrdov á, M. 1966. Palaeozoic microplankton from Central Bohemia. Casopis pro Mineralogii a Geologii 11: 409-414. [ Links ]
80. Volkheimer, W. 1968. Esporas y granos de polen del Jur ásico de Neuquén (República Argentina). I. Descripciones sistemáticas. Ameghiniana 5: 333-370. [ Links ]
81. Volkheimer, W. 1972. Estudio palinol ógico de un carbón caloviano de Neuquén y consideraciones sobre los paleoclimas jurásicos de la Argentina. Revista del Museo de La Plata (nueva serie), Paleontología 6: 101-157. [ Links ]
82. Volkheimer, W. 1973. Palinolog ía estratigráfica del Jurásico de la Sierra de Chacai Co y adyacencias (cuenca Neuquina, República Argentina). I. Estratigrafía de las Formaciones Sierra Chacai Co (Pliensbachiano), Los Molles (Toarciano, Aaleniano), Cura Niyeu (Bayociano) y Lajas (Caloviano inferior). Ameghiniana 10: 105-131. [ Links ]
83. Volkheimer, W. 1974. Palinolog ía estratigráfica del Jurásico de la Sierra de Chacai Co y adyacencias (cuenca Neuquina, República Argentina). II. Descripción de los palinomorfos del Jurásico inferior y Aaleniano (Formaciones Sierra Chacai Co y Los Molles). Ameghiniana 11: 135-172. [ Links ]
84. Volkheimer, W. y Quattrocchio, M. 1975. Palinolog ía estratigráfica del Titoniano (Formación Vaca Muerta) en el área de Caichigüe (cuenca Neuquina). Parte A: Especies terrestres. Ameghiniana 12: 193-241. [ Links ]
85. Volkheimer, W. y Quattrocchio, M.E. 1981. Distribuci ón estratigráfica de los palinomorfos jurásicos y cretácicos en la faja andina y áreas adyacentes de América del Sur Austral con especial consideración de la cuenca Neuquina. En: W. Volkheimer y E.A. Musacchio (eds.), Cuencas sedimentarias del Jurásico y Cretácico de América del Sur. Comité Sudamericano del Jurásico y Cretácico , (Buenos Aires) 2: 407-444. [ Links ]
86. Wall, D. 1965. Microplankton, pollen, and spores from the Lower Jurassic in Britain. Micropaleontology 11: 151-190. [ Links ]
87. Weaver, C.E. 1931. Paleontology of the Jurasic and Cretaceous of West Central Argentina . University of Washington, Memoir 1: 1-469, Seattle. [ Links ] Westermann, G.E.G. y Riccardi, A.C. 1979. Middle Jurassic ammonoid fauna and biochronology of the Argentine-Chilean Andes. II.- Bajocian Stephanocerataceae. Palaeontographica A 164: 85-118. [ Links ]
88. Wetzel, O. 1933. Die in organischer Substanz erhaltenen Mikrofossilien des baltischen Kreide-Feuersteins mit einem sediment-petrographischen und stratigraphischen Anhang. Palaeontographica A 77: 141-186; A 78: 1-110. [ Links ]
89. Zavala, C.A. 1993. [ Estratigrafía y análisis de facies de la Formación Lajas (Jurásico medio) en el sector suroccidental de la cuenca Neuquina. Provincia del Neuquén. República Argentina . Tesis Doctoral, Departamento de Geología, Universidad Nacional del Sur, Bahía Blanca, Argentina, 259 pp. Inédito] [ Links ].
90. Zavala, C.A. 1996a. Sequence Stratigraphy in Continental to Marine Transitions. An Example from the Middle Jurassic Cuyo Group, South Neuqu én Basin, Argentina. En: A.C. Riccardi, A dvances in Jurassic research . Transtec Publications, Switzerland. GeoResearch Forum 1-2: 285-294. [ Links ]
91. Zavala, C.A. 1996b. High-Resolution Sequence Stratigraphy in the Middle Jurassic Cuyo Group, South Neuqu én Basin, Argentina. En: A.C. Riccardi, Advances in Jurassic research . Transtec Publications, Switzerland. GeoResearch Forum 1-2: 295-304. [ Links ]
Recibido: 27 de junio de 2003.
Aceptado: 1 de abril de 2004.

