










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.42 no.1 Buenos Aires Jan./Mar. 2005
El esqueleto postcraneano de Neoaetosauroides (Archosauria: Aetosauria) del Triásico Superior del centro-oeste de Argentina
Julia Brenda Desojo 1 y Ana María Báez 1
1Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Pabellón II, Ciudad Universitaria, 1428, Buenos Aires, Argentina. julideso@gl.fcen.uba.ar
Resumen. Se describe el esqueleto postcraneano del aetosaurio Neoaetosauroides Bonaparte en base a material édito y adicional. El nuevo material consiste en proatlas, vértebras cervicales y placas dérmicas apendiculares colectados de la Formación Los Colorados, Cuenca de Ischigualasto-Villa Unión. Todos estos materiales se comparan con los de otros taxones de aetosaurios, y se discuten particularmente caracteres de la cintura pélvica y miembros posteriores considerados de importancia funcional. Este estudio revela una gran diversidad anatómica ( e.g. , diferencias en el grado de expansión pélvica, curvatura del fémur, morfología del crus y tarso) y sugiere la existencia en Aetosauria de formas semierectas y erectas. Este posible espectro de posturas locomotoras, junto a la amplitud de su rango de tamaño, sustenta propuestas previas de una mayor diversidad de modos de vida entre los Aetosaurios que la reconocida tradicionalmente.
Abstract. The postcranial skeleton of Neoaetosauroides (Archosauria: Aetosauria) from the Upper Triassic of west-central Argentina . The postcranial skeleton of the aetosaur Neoaetosauroides Bonaparte, based on previously studied specimens and additional material, is described herein. The new material consists of proatlas, cervical vertebrae, and appendicular dermal scutes collected in the Los Colorados Formation, Ischigualasto-Villa Unión Basin. All these materials are compared with those of other aetosaur taxa and functionally significant characters of the pelvic girdle and hindlimbs are in particular discussed. This study reveals a great anatomical diversity ( e.g. , differences in degree of pelvic expansion, femoral flexure, and crural and tarsal morphology) and suggests that Aetosauria might have included semi-erect and erect forms. This possible spectrum of locomotor postures, together with their wide size range, support previous propositions of more varied lifestyles among aetosaurs than traditionally recognized.
Key words. Archosauria. Aetosauria. Neoaetosauroides . Triassic. Ischigualasto-Villa Union Basin. Los Colorados Formation. Argentina.
Palabras clave. Archosauria. Aetosauria. Neoaetosauroides . Triásico. Cuenca Ischigualasto-Villa Unión. Formación Los Colorados. Argentina.
Introducción
Los aetosaurios son arcosaurios cuadrúpedos acorazados del Triásico Superior y su registro en el Hemisferio Norte es particularmente abundante, tanto en término de taxones representados como en número de ejemplares. No obstante, recientes hallazgos en América del Sur han ido ampliando nuestro conocimiento del grupo en este continente, inicialmente basado sólo en registros de Argentina (Casamiquela, 1980; Bonaparte; 1969, 1971a). A estos últimos se han sumado materiales de Chile (Casamiquela, 1980; Desojo, 2001, 2003), Brasil (Lucas y Heckert, 2001; Da Rosa y Leal, 2002), y ejemplares adicionales de Argentina (Desojo, 2002). Como consecuencia, se ha incrementado la diversidad de formas conocidas en el continente.
Las primeras menciones de este grupo en América del Sur están basadas en materiales provenientes de secuencias continentales de la Cuenca Ischigualasto- Villa Unión, en el margen occidental de Argentina. Estos restos, provenientes de la Formación Ischigualasto, fueron considerados representantes de los nuevos géneros Aetosauroides y Argentinosuchus (Casamiquela, 1960). Posteriormente, estos materiales fueron descriptos con mayor detalle (Casamiquela, 1961, 1967). Otros restos de aetosaurios fueron hallados más tarde en la Formación Los Colorados, que yace en discordancia sobre la Formación Ischigualasto (Bossi y Stipanicic, 2002). Los primeros registros en la Formación Los Colorados datan de la década de 1960, durante la cual Bonaparte y colaboradores realizaron numerosos trabajos de campo en la sierra Morada, provincia de La Rioja. Estos mate- riales fueron asignados a otro nuevo taxón, Neoaetosauroides engaeus (Bonaparte, 1969, 1971b, 1978). A partir de este registro, conjuntamente con los de arcosaurios basales, dinosaurios y terápsidos, Bonaparte (1966, 1971a, 1997) reconoció una asociación faunística diferente a la conocida en la subyacente Formación Ischigualasto. Debido a la abundancia de restos de prosaurópodos y a la presencia de rasgos presumiblemente derivados en Neoaetosauroides , ese autor propuso inicialmente la edad "Noriano-Rético" de la Formación Los Colorados. En 1973 el mismo autor consideró que el sector superior de la Formación Los Colorados, que contiene entre otros elementos faunísticos al aetosaurio Neoaetosauroides , representa la Edad/Reptil Coloradense superior atribuida al Noriano-Rético (Bonaparte, 1973: 125). Posteriormente fueron recolectados nuevos materiales de aetosaurios en la Formación Los Colorados y depositados en el Museo de Ciencias Naturales de la Universidad Nacional de La Rioja (Caselli et al., 2001).
En el presente trabajo se redescriben los restos postcraneanos del holotipo de Neoaetosauroides engaeus Bonaparte y se dan a conocer nuevos materiales postcraneanos y placas dérmicas apendiculares asignables a la misma especie. Se discuten particularmente caracteres de la cintura pélvica y miembros posteriores considerados de importancia funcional en la literatura.
Contexto geológico y procedencia de los materiales
La cuenca Ischigualasto-Villa Unión constituye una depresión elongada de tipo extensional con una marcada geometría de hemigraben (Milana y Alcober, 1995) que se desarrolló entre las provincias de San Juan y La Rioja. El relleno sedimentario consiste en sedimentitas continentales de edad triásica, las que han sido reunidas en varias unidades estratigráficas: las formaciones Talampaya y Tarjados (Romer y Jensen, 1966), y el Grupo Agua de la Peña (Bossi, 1971). Esta última unidad está integrada, de abajo hacia arriba, por las formaciones Chañares, Ischichuca, Los Rastros, Ischigualasto y Los Colorados (Groeber y Stipanicic, 1953).
La Formación Los Colorados comprende una sucesión de areniscas rojizas de hábito tabular y laminación paralela, con intercalaciones limoarcilíticas rojizas, y cuyo ambiente de depositación ha sido interpretado como fluvial de tipo meandriforme con facies finas de llanura de inundación (Kokogian et al. , 2001; Caselli et al., 2001) o bien como fluvial efímero (López Gamundi et al. , 1989). La secuencia evidenciaría una progresiva aridización del medio (Limarino et al. , 1990).
Los afloramientos de esta unidad se encuentran en todo el flanco este de la sierra Morada, cerro Rajado, cerro Bola y en una estrecha faja al oeste del Mogote del Gualo en el Campo de Talampaya (figura 1). El espesor en esta última zona es de sólo 100 m, mientras que varía entre 900 y 1.000 m en la zona de Ischigualasto-Ischichuca (Stipanicic y Bonaparte, 1979; Casselli et al., 2001; Kokogian et al., 2001).
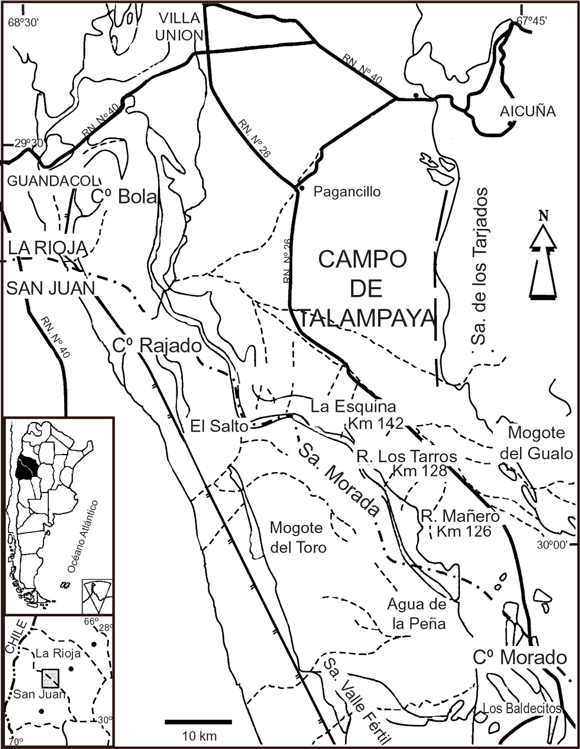
Figura 1. Mapa de la región de Ischigualasto-Villa Unión mostrando la localización de los sitios de donde provienen los restos de aetosaurios. Modificado de Kokogian et al. , 2001 / Map of the Ischigualasto- Villa Union region showing the location of the aetosaur-bearing sites. Modified from Kokogian et al. , 2001 .
Los materiales asignados a Neoaetosauroides provienen de varias localidades a lo largo de los afloramientos de la sierra Morada, desde la cabecera del río Mañero hasta La Esquina, provincia de La Rioja (figura 1). Los tres primeros ejemplares de Neoaetosauroides descriptos por Bonaparte (1969) provienen de los estratos superiores de la Formación Los Colorados. Los ejemplares PVL 3525 (holotipo) y PVL 3528 fueron colectados de una misma capa en la localidad Cabecera del río Mañero, kilómetro 126 de la Ruta Provincial 26, a diferencia del PVL 3842 que proviene de la misma localidad pero "unos 60 metros más arriba...o sea en el tope de la Formación Los Colorados" (Bonaparte, 1971b: 87). Otros materiales coleccionados en la misma área pero en diferentes localidades (figura 1) son los ejemplares PVL 3665 proveniente de la cabecera del río Los Tarros, kilómetro 128 de la Ruta Provincial 26, y el PVL 5698 proveniente de estratos entre La Esquina y la cabecera río Los Tarros, kilómetro 140 de la Ruta Provincial 26 (Caselli et al., 2001).
Abreviaturas. CPE, Coleção Municipal, São Pedro do Sul, Brasil; NMMNH, New Mexico Museum of Natural History and Science, Albuquerque, USA; PVL, Instituto Miguel Lillo, Tucumán; TTUP, Texas Tech University Paleontology, Lubbock, EE.UU.; TMM, Texas Memorial Museum, Austin, EE.UU.; UCMP, University of California Museum of Paleontology, Berkeley, EE.UU.; PULR, Paleontología Museo de Ciencias Naturales, Universidad Nacional de La Rioja, La Rioja.
Paleontología sistemática
ArcosauriA Cope, 1869
Crocodylotarsi Benton y Clark, 1988
Aetosauria Marsh, 1884
Género Neoaetosauroides Bonaparte, 1969
Especie tipo. Neoaetosauroides engaeus Bonaparte, 1969.
Diagnosis enmendada. La misma que para Neoaetosauroides engaeus , la única especie conocida.
Neoaetosauroides engaeus Bonaparte, 1969
Figura 2

Figura 2. Neoaetosauroides engaeus Bonaparte. A, PVL 5669, región cervical en vista lateral; B, dibujo interpretativo de la región cervical en vista lateral; C, PVL 3525, cintura pélvica derecha en vista lateral; D, dibujo interpretativo de la cintura pélvica derecha en vista lateral. Escala: 1,5 cm. Abreviaturas: a, atlas, ax. , axis, c3-c5, centros de cervicales 3 a 5; il, ilión, is, isquión, p, proatlas ; pu, pubis./ Neoaetosauroides engaeus Bonaparte. A, PVL 5669, cervical region in lateral view; B, interpretive drawing of cervical region in lateral view; C, PVL 3525, right half of pelvic girdle in lateral view; D, interpretive drawing of right half of pelvic girdle in lateral view. Scale: 1.5 cm. Abbreviations: a, atlas; ax, axis; c3-c5 , cervical centra 3 to 5; il, ilion; is, ischion; p, proatlas; pu, pubis.
Holotipo. PVL 3525, mandíbula, gran parte del esqueleto postcraneano articulado, coraza dorsal y osteodermos ventrales.
Paratipos. PVL 3528: fragmentos de premaxilar y maxilar con dientes, vértebras y placas dérmicas; huesos indeterminados; PVL 3842: fragmentos de coraza dorsal y ventral, impresión de 8 arcos de placas dérmicas y sus correspondientes vértebras.
Material referido. PVL 4363: cráneo expuesto en vista lateral y mandíbula derecha, impresiones de 4 placas dérmicas paramediales y 1 ventral; PVL 5698: cráneo, proatlas, 6 vértebras cervicales articuladas, 6 placas cervicales izquierdas articuladas; PVL 3665: extremo proximal del fémur y húmero izquierdo, fragmento escapulocoracoideo izquierdo y de 1º vértebra caudal; PULR 108: cráneo en vista palatal y mandíbulas, impresión del fémur, porción distal de la tibia y fíbula izquierdas, molde interno de las placas apendiculares; PULR 109: impresión de la coraza dorsal articulada.
Horizonte y localidad tipo. Formación Los Colorados, sector más superior; kilómetro 126 de la Ruta Provincial 26, de Patquia a Villa Unión, Provincia de La Rioja, Argentina.
Diagnosis enmendada. Aetosaurio que se diferencia de los demás por la presencia de coracoides con un proceso agudo dirigido posteromedialmente, vértebras cervicales con centros más cortos y espinas neurales más bajas que el alto del centro vertebral, cavidad del calcáneo para el cóndilo del astrágalo con bordes poco delimitados, dedo V del pie con dos falanges y estuche caudal corto compuesto por 26 arcos de placas. Además se diferencia de otros aetosaurios excepto Longosuchus por tener mandíbula corta, alta y gruesa y dientes maxilares y mandibulares cónicos y grandes, y de este último por poseer centros vertebrales con quillas ventrales y carecer de cuernos y articulación " tongue and groove " en las placas laterales.
Descripción
Columna vertebral. El ejemplar PVL 3525 (holotipo) es el único disponible en el que se ha preservado parte de la columna vertebral articulada, constituida por 11 vértebras dorsales, 2 sacras y 4 caudales. Las vértebras caudales posteriores a la 4º caudal parecen haberse preservado pero no pueden observarse por no haber sido preparadas. Lo mismo ocurre con las vértebras dorsales, las que sólo pueden ser observadas en vista ventral ya que dorsalmente portan la coraza. Debe destacarse que este material se encuentra en exposición en el Museo del Instituto Miguel Lillo, Tucumán, y la porción del cuello que articula anteriormente con el cráneo y posteriormente con la región dorsal corresponde en realidad a otro ejemplar, el PVL 5698. Las descripciones de las vértebras cervicales así como del proatlas que se presentan a continuación se basan en este último ejemplar (figuras 2.A, B). En cambio, las descripciones de las vértebras dorsales, sacras y caudales se basan en el ejemplar PVL 3525.
Inmediatamente por delante de la primera vértebra cervical (atlas), y contenido en un bloque desplazado anteriormente (figura 2.A), puede observarse la presencia de un par de piezas óseas que forman una V. Las mismas se hallan a la altura de los arcos neurales de las vértebras cervicales (figura 2.B), con el extremo dorsal que corresponde al vértice de la V algo más anterior que los extremos ventrales. El vértice contactaría con una depresión visible en la región occipital del cráneo, dorsal al foramen magno, mientras que los extremos posteriores lo harían con el atlas. Se ha identificado a este elemento como un proatlas, cuya presencia se reconoce por primera vez para el clado Aetosauria.
El atlas es la mitad de largo anteroposterior que el axis. Los pedicelos del arco neural contactan ventralmente con el intercentro y por ello el atlas tiene forma de anillo. Puede observarse en el mismo ejemplar que la apófisis odontoidea del axis lo atraviesa. Las postzigapófisis se sitúan posterodorsalmente, articulando con las prezigapófisis del axis. En vista ventral se observan dos proyecciones lateroposteriores para la articulación con el primer par de costillas cervicales que no se ha preservado. La espina neural del axis posee una apreciable longitud ánteroposterior, generando una amplia superficie que en vista lateral es 2 veces más ancha que las de las espinas de las vértebras siguientes (figura 2.A). El margen superior de la espina está inclinado posteriomente, aumentando su altura en dirección posterior. Los pedicelos del arco neural del axis son más extensos anteroposteriormente que los de las vértebras posteriores; en su base están las postzigapófisis articulando con las prezigapófisis de la 3º cervical. En vista ventral puede observarse una lámina ósea dirigida hacia el atlas que corresponde a la mitad posterior izquierda del piso de la apófisis odontoidea, pero el centro vertebral del axis no se ha preservado.
La espina neural de la tercer vértebra cervical es la más alta de esta serie; igualmente, a partir de esta vértebra se reduce la longitud ánteroposterior de los arcos neurales. El margen posterior de la espina es recto. Los procesos que sostienen a las prezigapófisis se extienden anteriormente en la base de los pedicelos y son anteriores al margen anterior de la espina neural. Contrariamente, el nacimiento de las postzigapófisis se sitúa anteriormente al margen posterior de la espina. El centro vertebral es más alto que la espina neural, y en vista ventral puede observarse que la parapófisis derecha articula con la costilla cervical. En vista ventral se observan los centros de las vértebras cervicales III a V; todos ellos llevan una quilla ventral bien desarrollada, perpendicular a las carillas articulares. Las espinas neurales de las vértebras cervicales IV a VI tienen sus superficies distales más inclinadas posteroventralmente que la vértebra precedente (III). El tamaño de las zigapófisis aumenta notablemente a partir de la articulación entre las vértebras III y IV. El centro vertebral de la vértebra cervical VI no se ha preservado. Entre las cervicales se advierte una posición de las parapófisis cada vez más dorsal y próxima a las diapófisis. Se han preservado tres costillas cervicales bicéfalas derechas correspondientes a las vértebras cervicales III, IV y V, cuyos procesos distales se unen posterolateralmente formando un canal entre éstos y los centros vertebrales para el pasaje de la arteria vertebral.
En el holotipo se han conservado once vértebras dorsales presacras articuladas. Sólo pueden observarse en vista ventral, excepto la primer vértebra dorsal cuya espina neural larga anteroposteriormente es más baja que el alto del cuerpo vertebral. A su vez, el proceso transverso de esta vértebra es más ancho y largo que la espina neural. Las apófisis articulares de las vértebras dorsales no se encuentran bien preservadas, a diferencia de los procesos transversos que articulan con costillas bicéfalas hasta la vértebra presacra IX y disminuyen en longitud posteriormente. Las diápofisis son más anchas que las parapófisis, pero ambas se encuentran en el mismo plano horizontal. Las dorsales anteriores presentan el centro vertebral en forma de carretel, comprimido lateralmente. A partir de la vértebra IV el tamaño del centro aumenta posteriormente. Los centros de las vértebras IX a XI son cilíndricos, cortos anteroposteriormente y muy robustos. A diferencia de lo observado en las vértebras cervicales, no se observan quillas ventrales en los centros vertebrales. Las costillas dorsales son anchas y largas, sobrepasando el borde lateral de la coraza dorsal.
Las costillas sacras derechas de las dos vértebras sacras están fusionadas al ilión derecho, la costilla sacral de la primera de ellas, de sección cuadrangular, es más robusta que la segunda. A diferencia de la primer costilla sacral, la segunda nace más atrás y se dirige anteriormente para fusionarse con la porción posterior del ilión que forma el margen del acetábulo. Los centros de estas vértebras son los más robustos de la columna vertebral. Las cuatro primeras vértebras caudales presentan procesos transversos anchos anteroposteriormente y muy largos. Los centros vertebrales, cuyo tamaño disminuye posteriormente, son cortos y anchos. A partir de la segunda vértebra caudal se observan carillas de articulación para arcos hemales.
Cintura escapular (v. Bonaparte, 1971b: figura 36). La escápula y el coracoides están firmemente unidos, pudiéndose observar la sutura entre ambos ( e.g. , PVL 3525, 3665). La hoja escapular es plana y expandida distalmente, en particular posteriormente, lo que determina que ese extremo termine en punta a la altura del labio coracoideo de la cavidad glenoidea. Por el contrario, su extremo anterior es redondeado. La hoja escapular se continúa ventralmente con el tallo escapular, de sección transversal triangular en su porción basal, en cuyo margen posterior y próximo al labio escapular de la cavidad glenoidea presenta un pequeño tubérculo correspondiente a la zona de inserción del músculo triceps . Ventralmente, la escápula se expande, llevando en su porción posterior el labio escapular de la cavidad glenoide. Su porción anterior presenta un engrosamiento que se extiende dorsalmente a lo largo del margen anterior hasta la base del tallo escapular. Este engrosamiento ha sido interpretado como la porción distal de la clavícula fusionada a la escápula (Bonaparte, 1971b). El coracoides presenta la superficie externa (ventral) convexa y la interna (dorsal) cóncava. El margen anterior es redondeado y de poco espesor, a diferencia del margen posterior que es más grueso. Ventralmente a la cavidad glenoidea se desarrolla en la superficie externa del coracoides una quilla que delimita una superficie plana anterior y otra cóncava de forma triangular muy proyectada posteriormente. El coracoides presenta un foramen coracoideo muy pequeño, difícilmente observable, ubicado en la superficie externa anterior de la cavidad glenoidea y próximo a la sutura escapulocoracoidea.
Miembro anterior (v. Bonaparte, 1971b: figura 38). En líneas generales, el húmero es un hueso robusto y expandido en ambos extremos. La expansión proximal (dorsal), mayor que la distal (ventral), tiene una superficie articular muy ancha y un engrosamiento dorsal que forma la cabeza humeral. Posteriormente a esta última se observa el tubérculo interno que no sobrepasa el nivel del borde lateral del ectepicóndilo. El margen anterior de la expansión proximal presenta una expansión que se proyecta ánteroventralmente, originando la cresta deltoidea, muy desarrollada comparada con la de otros Crurotarsi. Esta cresta se extiende paralelamente al margen lateral del húmero hasta el inicio de la diáfisis, donde desaparece. Entre la cresta y la superficie lateral se observa un área triangular y cóncava. La diáfisis es corta, ancha y de sección transversal triangular. El extremo distal presenta el ectepicóndilo más desarrollado que el entepicóndilo. Este último cóndilo en vista externa presenta una depresión que corresponde a la articulación con el proceso olecraneano de la ulna. En vista interna, la depresión del ectepicóndilo destinada para la articulación con el radio limita, junto con la depresión del entepicóndilo, una superficie cóncava destinada al proceso olecraneano. En el extremo distal del húmero se observa un surco que distalmente termina en el foramen ectepicondilar ( e.g. , PVL 3525), que corresponde al pasaje del nervio radial y vasos sanguíneos (Romer, 1956).
El radio es recto y de menor tamaño que la ulna. El extremo proximal está más expandido láteromedialmente que ánteroposteriormente, y más desarrollado que el distal. En el extremo proximal la carilla articular presenta una superficie plana, mientras que la porción medial es más robusta y se proyecta más ánteromedialmente. La diáfisis, de sección transversal circular, es ancha y se continúa imperceptiblemente con el extremo distal. La carilla articular se halla rota en el único ejemplar en el que se ha conservado el radio (PVL 3525). La ulna es más robusta que el radio, y su extremo proximal, de mayor tamaño que el distal, se halla expandido ánteroposteriormente y comprimido lateralmente. La carilla articular para la recepción del cóndilo medial del húmero es cóncava. El olécranon se halla bien definido. La diáfisis disminuye en tamaño desde el extremo proximal al distal; este último es pequeño y de sección transversal oval. No se ha conservado ningún elemento del autopodio (contra Heckert y Lucas, 1999).
Cintura pélvica. El ilión presenta una hoja ilíaca plana, alta y larga, con una proyección anterior estiliforme que sobrepasa el pedúnculo púbico ( sensu Long y Murry, 1995). Esta proyección presenta el extremo dirigido ventralmente y su punta es roma. El sector posterior de la hoja ilíaca es ancho, de forma cuadrada en vista lateral y dirigido dorsomedialmente, con el borde dorsal recto y posterior cóncavo (figura 2.C). La porción preacetabular ilíaca es más angosta que la postacetabular y contacta con el extremo anterior de la primera costilla sacra, mientras que la costilla de la segunda vértebra sacra lo hace con la expansión postacetabular. El acetábulo, de posición lateroventral, no está perforado, es profundo y de gran tamaño en relación a los huesos que lo forman. Dorsalmente no se observa ninguna estructura a lo largo de su contacto con la hoja ilíaca; ventralmente presenta una depresión limitada por el pubis e isquión (figura 2.D). El pubis es de forma triangular y sección aplanada, y su extremo distal no se ha preservado en el material disponible. Ese hueso forma parte del acetábulo por medio de una fina lámina separada por un reborde óseo de su parte anterior y que contacta con el isquión ventral al acetábulo. En la porción anterior del pubis puede observarse el foramen obturador (figura 2.D). El isquión es de tamaño similar al del pubis; su extremo proximal contacta posteriormente con el ilión y anteriormente forma el acetábulo, el que se halla separado por un reborde óseo. El isquión se proyecta posteroventralmente y finaliza en un extremo distal incompletamente preservado.
Miembro posterior (v. Bonaparte, 1971b: figuras 39 y 40). El fémur es un hueso muy robusto, de forma ligeramente sigmoidea y cuyo largo es casi dos veces el del húmero. Su extremo proximal es más angosto que el distal. La superficie articular del extremo proximal presenta la fosa intertrocantérica muy desarrollada y poco profunda. El margen anterior de la cabeza femoral es redondeado y ancho; se continúa en una lámina ósea que termina cerca del cuarto trocánter, sitio de inserción del músculo caudofemoralis longus . Dicho trocánter se halla a una distancia del extremo proximal equivalente a 1/3 de la longitud femoral. La diáfisis es ancha y corta y de sección transversal triangular. En el extremo distal se halla la fosa intercondilar que es poco profunda. El cóndilo interno es de menor tamaño que el externo y el fibular de posición bien lateral.
La tibia presenta el extremo proximal más ancho que el distal. El extremo proximal presenta una cresta que se extiende por la cara anterior de la diáfisis. La cara articular proximal presenta dos depresiones poco profundas y una proyección lateral con la que contacta la fíbula. En vista anterior se observa la pronunciada cresta cnemial que se extiende a lo largo de la totalidad de este hueso, para la inserción del músculo femoral cuadriceps . La diáfisis, robusta y recta, es de sección transversal triangular. El extremo distal presenta una superficie de articulación para el astrágalo que es plana y ancha en la parte anterior y convexa en su prolongación posterior. La fíbula, menos robusta que la tibia, es casi recta en vista lateral y su diáfisis tiene sección transversal triangular. Su extremo proximal está más expandido anteroposteriomente que láteromedialmente. En vista anterior este hueso presenta un prominente trocanter iliofibular para la inserción del músculo iliofibularis , que se proyecta ánterolateralmente a 1/3 de la longitud del hueso con respecto al extremo proximal. La diáfisis es angosta proximalmente y curvada distalmente. El extremo distal presenta una carilla articular plana para el calcáneo.
El astrágalo (Bonaparte, 1971b: figuras 40 y 41) presenta proximalmente una faceta tibial que se extiende por toda la sección del hueso. Esta faceta es más corta ánteroposteriormente que mediolateralmente y se halla subdividida por un promontorio en dos carillas de diferente orientación, lo que le confiere la característica forma de silla de montar. En cambio, la bien definida faceta fibular es de pequeño tamaño, forma rectangular y está dispuesta casi verticalmente. El cóndilo ventral para la articulación con el calcáneo es de forma redondeada. El calcáneo presenta una cavidad de forma redondeada y no muy profunda para la articulación con el cóndilo del astrágalo. Esta cavidad se continúa medialmente en un plano perpendicular con un área oblonga y cóncava que recibe la superficie convexa ventroposterior del astrágalo. En vista dorsal o ventral ambas concavidades están contenidas en planos que forman un ángulo obtuso. Esta ampliada articulación cóncavoconvexa entre astrágalo y calcáneo caracteriza la articulación cocodrilo-normal ( sensu Chaterjee, 1978). Medialmente se proyecta un proceso denominado sustentaculum (Parrish, 1986). En el área posterior del calcáneo se proyecta la bien desarrollada tuberosidad que constituye un punto de apoyo del músculo gastrocnemius , principal flexorplantar entre el crus y el pie. Sin embargo, no se aprecia un surco dorsoventral bien marcado.
Sólo se han conservado dos tarsales distales en el tarso (v. Bonaparte, 1971b: figura 42). El de menor tamaño se halla en contacto con la base del metatarsal III, mientras que el más grande contacta proximalmente con el calcáneo y distalmente con los metatarsales IV y V. Estos tarsales distales han sido interpretados como III y IV respectivamente (Parrish, 1986). El pie está formado por cinco dígitos, de los cuales los primeros cuatro se superponen parcialmente a través de los extremos proximales expandidos de los metatarsales y el quinto es divergente. Los metatarsales tienen sección triangular y sus extremos proximales más expandidos que los distales. El metatarsal I es el más corto y robusto, siendo el III el más largo. El metatarsal V, con forma de gancho, se halla separado del resto de los metatarsales, por lo que el pie está expandido mediolateralmente. La fórmula falángica es 23452. Todas las falanges terminales preservadas son aguzadas distalmente, mientras que las restantes falanges tienen forma de carretel. La longitud de los dedos decrece IV>III>II>I>V.
Osteodermos. Los osteodermos forman una coraza dorsal, disponiéndose en dos hileras de placas paramediales que se relacionan uno a uno con las vértebras subyacentes, bordeadas lateralmente por una hilera de placas laterales (figura 3). Adicionalmente, se han preservado osteodermos ventrales y apendiculares. Todas estas placas presentan una barra anterior articular, excepto las apendiculares. La ornamentación, consistente en surcos y fosetas, es poco marcada y de patrón radial. La coraza dorsal comienza con dos placas dérmicas cervicales pequeñas, más anchas que largas con el margen lateral redondeado. Posteriormente a ellas se presentan aproximadamente siete placas paramediales cervicales, más anchas que largas, rodeadas lateralmente por una hilera de placas laterales de forma cuadrangular. Las placas dorsales articulan posteriormente con placas caudales. La relación ancho/largo de las placas paramediales dorsales es de 3. Las placas laterales presentan un ángulo agudo y acompañan la constricción en la región sacra, por lo que en esta última región la relación ancho/largo de las placas paramediales es de 2. Posteriormente la coraza se continúa con veintiseis placas caudales. Las placas dérmicas ventrales son de forma cuadrada a hexagonal. En el holotipo se han preservado sólo en la región caudal, formando el estuche caudal, si bien la presencia de placas ventrales aisladas con barra articular en la región anterior, hace presumir la presencia de una coraza ventral (figura 3). Las placas dérmicas apendiculares, preservadas como impresiones en el ejemplar PULR 108, tienen forma romboidal, son pequeñas, y se encontraban inmersas en la dermis. No puede observarse su ornamentación pues sólo se conoce su superficie interna.
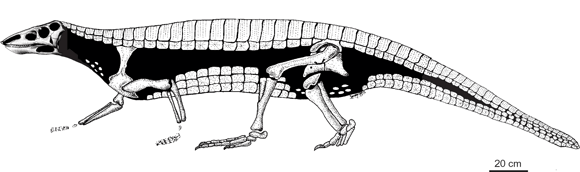
Figura 3. Neoaetosauroides engaeus Bonaparte. Reconstrucción del esqueleto y coraza / Restoration of the skeleton and caparace.
Análisis comparativo
En los últimos 10 años se han realizado numerosos análisis filogenéticos para resolver las interrelaciones de los diversos grupos de arcosaurios, incluyendo los aetosaurios ( e.g. , Sereno y Arcucci, 1990; Sereno, 1991; Parrish, 1993; Juul, 1994; Gower y Wilkinson, 1996; deRicqlès et al. , 2003). En esos trabajos el clado Aetosauria fue incluído dentro de Crurotarsi, un grupo mononofilético que incluye además a Ornithosuchidae, Parasuchia, Rauisuchia, y Crocodylomorpha (Sereno y Arcucci, 1990), aunque no hay un consenso con respecto a los taxones que estos grupos comprenden y a sus interrelaciones. Más aún, la monofilia de Rauisuchia ha sido puesta en duda (Gower y Wilkinson, 1996). Los aetosaurios han sido considerados como grupo hermano de los Rauisuchia, o más recientemente de los Crocodylomorpha (Gower y Walker, 2002). En este contexto filogenético general se discuten algunos caracteres postcraneanos de Neoaetosauroides y otros arcosaurios. A su vez, tampoco las relaciones entre los taxones de aetosaurios están completamente resueltas, habiéndose propuesto hipótesis alternativas (Heckert et al., 1996; Heckert y Lucas, 1999, 2000, 2003; Harris et al. , 2003a, 2003b).
Las comparaciones del esqueleto postcraneano de Neoaetosauroides con otros taxones de aetosaurios se basan en la literatura pertinente: Aetosaurus O. Fraas (Walker, 1961), Aetosauroides Casamiquela (Casamiquela, 1960, 1961, 1967), Coahomasuchus Heckert y Lucas (Heckert y Lucas, 1999), Desmatosuchus Case (Case, 1920; Small, 1985), Longosuchus Hunt y Lucas (Sawin, 1947; Hunt y Lucas, 1990), Lucasuchus Long y Murry (Long y Murry, 1995), Stagonolepis Agassiz (Walker, 1961; Long y Ballew, 1985; Long y Murry, 1995) y Typothorax Cope (Cope, 1875; Long y Murry, 1995). Esa información ha sido complementada por observaciones de una de las autoras (J.B.D.), identificándose en ese caso el ejemplar correspondiente.
El proatlas es un hueso par o impar, aunque siempre originado a partir de un par de cartílagos, intercalado entre la región occipital y el atlas (Goodrich, 1958). Este elemento ha sido descrito en numerosos grupos de tetrápodos vivientes y fósiles, aunque su identificación en Neoaetosauroides en el presente trabajo constituye la primera cita para un aetosaurio. Ha sido descripto en la mayoría de los otros grupos de Crurotarsi, como fitosaurios (Chatterjee, 1978), ornitosúquidos ( e. g. , Bonaparte 1971b) y cocodrilos ( e.g. , Walker, 1990), por lo que presumiblemente su presencia en aetosaurios es plesiomórfica.
Las vértebras cervicales de Neoaetosauroides , que no se conocían, presentan una quilla ventral en sus centros, como ocurre en los aetosaurios del Hemisferio Norte Stagonolepis , Aetosaurus , Coahomasuchus (NMMNH P-3004 J.B.D. obs. pers.) y Typothorax . En Desmatosuchus , en cambio, su presencia varía individualmente (TTUP 10008, UCMP 78705, UCMP 78704, UCMP 177315 J.B.D. obs. pers.), mientras que en Longosuchus (TMM 31185, TMM 31185-84b, CP.E2.168 J.B.D. obs. pers.) los centros vertebrales de las cervicales carecen de quillas (Long y Murry, 1995). Es posible que la presencia de vértebras cervicales con quillas en sus centros sea un carácter plesiomorfo para aetosaurios, dada su presencia en otros Crurotarsi, como fitosaurios (Camp, 1930; Chatterjee, 1978; Lucas et al. , 2002), ornitosúquidos (Walker, 1964; Bonaparte, 1971b), rauisúquidos (Sill, 1974; Cha_tterjee, 1985) y cocodrilos (Nash, 1975; Walker, 1990; Wu et al. , 1997) si bien no es un rasgo constante en todos ellos.
Las proporciones de las vértebras cervicales varían ampliamente entre los distintos géneros de aetosaurios. Así, los centros cervicales en Neoaetosauroides son más altos que anchos, lo que contrasta con las cervicales de Typothorax y Desmatosuchus (UCMP 78705, UCMP 78704, UCMP 139912, UCMP 139913 J.B.D. obs. pers.) cuyos centros son claramente más anchos que altos. Igualmente, las espinas neurales de las vértebras cervicales de Neoaetosauroides son tan o más altas que el centro vertebral, como las de Stagonolepis wellesi Long y Ballew y Longosuchus . En contraste, las cervicales de Typohorax y Desmatosuchus tienen espinas neurales cortas, que en el último nunca exceden el alto del centro vertebral.
En Neoaetosauroides la distancia entre los extremos de los procesos transversos de las vértebras presacrales es dos veces el ancho del centro vertebral; esa distancia es igual o mayor que el doble del ancho del centro en la mayoría de los aetosaurios, excepto Desmatosuchus y Lucasuchus , como fuera señalado en el análisis filogenético de Aetosauria por Heckert y Lucas (1999).
La forma general de la cintura pectoral de Neoaetosauroides es más similar a la de Longosuchus y Typothorax , que a la de Desmatosuchus y Stagonolepis . En estos últimos dos géneros el tallo escapular es comparativamente más ancho y corto que en los primeros. En Neoaetosauroides , en contraste con Stagonolepis robertsoni , Aetosauroides , y posiblemente S. welesi y Desmatosuchus , no hay clavículas como elementos discretos. Sin embargo, como ya se mencionara en la descripción original, se ha interpretado en Neoatosauroides que la clavícula se halla fusionada a la escápula. La forma aguzada del extremo posterior del coracoides, ventralmente al labio coracoideo, es característica de Neoaetosauroides . Ese extremo es de forma redondeada en Desmatosuchus , Stagonolepis , y Longosuchus .
El extremo proximal del húmero está relativamente poco expandido en Neoaetosauroides y Typothorax , a diferencia de Desmatosuchus , y Stagonolepis , en los que está muy expandido y la cresta deltopectoral se extiende hasta la mitad de este hueso. La tuberosidad interna no sobrepasa el margen lateral del ectepicóndilo en Neoaetosauroides y Typothorax a diferencia de Stagonolepis y Desmatosuchus donde el extremo proximal está fuertemente expandido en sentido ánteroposterior. La diáfisis ancha, corta y de sección triangular en Neoaetosauroides y Longosuchus , contrasta con la diáfisis larga, angosta y comprimida dorsoventralmente del húmero de Desmatosuchus . La presencia de un suave surco que termina en el foramen ectepicondilar sobre la cara lateral del cóndilo radial del húmero es un carácter de Neoaetosauroides compartido con Desmatosuchus y Longosuchus , entre los aetosaurios. En otros arcosaurios el nervio radial y varios vasos sanguíneos corren en un surco abierto, sin que el mismo esté techado dando lugar al foramen ectepicondilar (Romer, 1956).
La presencia de una conspicua proyección anterior en la hoja ilíaca no sólo ocurre en Neoaetosauroides sino también en otros aetosaurios como Stagonolepis wellesi Longe y Ballew, Typothorax coccinarum Cope, Desmatosuchus haplocerus Cope, y Aetosauroides scagliai Casamiquela. La proyección ilíaca anterior es algo más corta en Longosuchus y S . robertsoni Walker. Esta expansión anterior de la hoja ilíaca tiene importantes implicancias funcionales, dado que posibilita la modificación de la inserción de los músculos iliotibialis e iliofemoralis que son extensores y protractores del miembro posterior respectivamente y ambos importantes rotadores femorales (Parrish, 1986). El desplazamiento anterior y dorsal con respecto a la articulación de la cadera del origen de esos músculos alarga el brazo de palanca de los mismos, lo que en conjunción con otros rasgos anatómicos posibilita diferentes acciones.
Entre los aetosaurios la posición de los acetábulos varía, lo que debe estar relacionado con una diferente morfología y posición de los fémures. En Neoaetosauroides , y Desmatosuchus (UCMP 25989, TMM 31100-1, TMM 10008, TMM 31100-313, J.B.D. obs. pers.) el acetábulo se halla en un plano oblícuo con respecto al plano sagital y presenta un reborde dorsal marcado que suprayace a la cabeza del fémur cuando éste está articulado. En cambio, en Typothorax , Aetosauroides y Longosuchus (TMM 3100-236, TMM 31185- 40, NMMNH 36075 J.B.D. obs. pers.) el ilión tiene una posición casi perpendicular al plano sagital, por lo que el acetábulo es prácticamente horizontal (Casamiquela, 1961, 1967). De acuerdo a la reconstrucción de Walker (1961), en Stagonolepis el acetábulo sería vertical, una condición similar a la presente en algunos arcosauromorfos basales. El fémur de Neoaetosauroides es distintivamente robusto y, como en Desmatosuchus , se diferencia una cabeza articular cuya inclinación forma con el eje longitudinal de la diáfisis un ángulo agudo (figura 4). En cambio, en Longosuchus y Aetosauroides el ángulo de inclinación de la cabeza femoral con respecto a la diáfisis es menor, lo que estaría correlacionado con la posición horizontal del acetábulo. En este último género el fémur presenta una torsión alrededor de su eje longitudinal, lo que le confiere una configuración netamente sigmoide característica (Casamiquela, 1961: figura 20).
En líneas generales, la tibia de Neoaetosauroides es un hueso delgado, similar al de Stagonolepis , y menos robusto que en Longosuchus , Desmatosuchus y Typothorax . En estos dos últimos taxones la tibia es notoriamente más corta en relación al fémur y más robusta, lo que podría estar relacionado con su gran tamaño con respecto a otros aetosaurios (figura 4). La cresta cnemial está marcada, como en Longosuchus . La articulación entre el astrágalo y el calcáneo en Neoaetosauroides difiere de la de otros aetosaurios en los que conocen estos elementos ( e.g. , Typothorax , Longosuchus ) porque los planos que contienen las concavidades para el astrágalo forman ángulo mayor de 90º. El tarsal distal lateral es de mayor tamaño que el medial, como en Stagonolepis , Aetosauroides y Aetosaurus (Casamiquela, 1967), los únicos taxones en los que se han preservado esos huesos.
Neoaetosauroides retiene un patrón del pie generalizado para Crurotarsi en lo que respecta al menor tamaño de los dedos I y V, y un metatarsiano V con un proceso proximal dirigido medialmente (forma de "gancho"). Difiere del de los demás aetosaurios y otros grupos de Crurotarsi de los que se conoce el pie en la presencia de un dedo IV más largo que los dedos II y III. Otro carácter peculiar del pie de este taxón es la reducción del número de falanges en el dígito V, presentando el menor número de falanges conocida para los aetosaurios (3 en Longosuchus , 4 en Stagonolepis ). Esta tendencia a la reducción de las falanges del dígito V ocurre independientemente en otros Crurotarsi. El tamaño de las falanges ungueales de los dedos I y II generalmente es mayor que los de los dedos más laterales, pero esta diferencia es marcadamente más notoria en Neoaetosauroides que en otros aetosaurios (figura 4). Los metatarsianos de Neoaetosauroides , incluyendo el V recurvado, son relativamente gráciles y en sus proporciones semejantes a los de Stagonolepis y Longosuchus . Contrastan con los más cortos y anchos de Typothorax (NMMNH 36075) y Desmatosuchus (UCMP 34497).

Figura 4. Esquema de los fémures, fíbulas, tibias y autopodios.; A, Desmatosuchus ; B, Typothorax ; C, Longosuchus ; D, Neoaetosauroides ; E, Aetosauroides ; F, Aetosaurus ; G, Stagonolepis robertsoni . A diferentes escalas./ Sketches of femurs, fibulae, tibiae, and pes. Not to the same scale.
Discusión
Neoaetosauroides , junto a Aetosauroides , es uno de los taxones mejor representados de los aetosaurios sudamericanos, constituyendo una de las formas más tardías conocidas a nivel mundial. Por otra parte, este taxón es uno de los pocos representados por ejemplares con postcráneo articulado lo que permite estudios más profundos.
Muchos rasgos de los miembros posteriores de los arcosauromorfos han sido interpretados como parte de complejos de caracteres cuya significación en cuanto a la capacidad locomotora ha sido reiteradamente destacada por diferentes autores. Sin embargo, un análisis del patrón de adquisición de las modificaciones estructurales postcraneanas y su evolución en los aetosaurios en un contexto histórico requieren un marco filogenético explícito y confiable. Precisamente, una revisión de las relaciones filogenéticas de este grupo se halla en marcha por lo que en este trabajo sólo se han interpretado algunos caracteres a la luz de las hipótesis funcionales postuladas por otros autores para los arcosaurios basales.
Diferentes aspectos de la cintura pélvica, del fémur, de la tibia, de la articulación entre zeugopodio y autopodio, y la plantigradía o digitigradía fueron utilizados por Charig (1972) para caracterizar estadios en la evolución de un andar "mejorado" o erecto a partir de una condición primitiva de andar esparrancado. Pero, como bien lo señalara Bonaparte (1984), estas innovaciones locomotoras ocurrieron independientemente en distintos linajes de arcosaurios con diferentes características de acuerdo a su historia filogenética. Varios modelos acerca de las características morfológicas esperadas en formas de arcosaurios con distinto andar, esparrancado, intermedio y erecto, fueron planteados por Parrish (1986), si bien estudios recientes objetan la utilización de los cocodrilos modernos como modelos de una postura locomotora semi erecta intermedia (Reilly y Elias, 1998). En su análisis Parrish (1986) incluyó a los aetosaurios, a los que consideró un grupo de postura plantígrada con articulación crurotarsal cocodrilo-normal ( sensu Chaterjee, 1978) de andar mejorado o erecto. Resulta, por lo tanto, interesante poner a prueba esta aseveración en base a la información que proporciona el reestudio de Neoaetosauroides . En otras palabras, ¿se ajusta este género a todos los aspectos del correspondiente modelo morfofuncional?
1. El acetábulo es profundo, permitiendo sólo la protracción y retracción femoral en un plano parasagital. Cabe señalar que la orientación varía entre los aetosaurios, pudiendo ser por lo menos oblícua, como en Neoaetosauroides , u horizontal, como en Aetosauroides .
2. La hoja ilíaca está expandida anteriormente; isquiones y pubis no están completos distalmente en el material disponible pero parecen tratarse de elementos medianamente alargados. Estos rasgos tenderían a favorecer la orientación vertical del fémur.
3. La morfología crural presenta rasgos esperables en formas de andar erecto. La presencia de dos depresiones correspondientes a un par de meniscos para la articulación con el fémur ayudan a restringir el movimiento de este segmento del miembro a una simple flexión y extensión con respecto a este último hueso. El relieve del extremo distal para la articulación con el astrágalo es moderadamente curvado, pero no tan marcado como en otros aetosarios (e.g. , Desmatosuchus , Typothorax ) ni plana como en formas de andar esparrancado. De todas maneras el relieve complementario del astrágalo y el extremo distal de la tibia favorecen una estrecha articulación. Esto resulta en una columna tibio-astragalar que restringe a un plano parasagital la flexión y extensión del calcáneo con respecto al astrágalo.
4. La articulación astrágalo-calcanear está bien definida como en las formas erectas, aunque la cavidad para el cóndilo del astrágalo no presenta bordes tan bien delimitados como en otros géneros de aetosaurios lo que reduce el nivel de encastre entre estos huesos.
5. El calcáneo presenta una tuberosidad dirigida posteriormente, perpendicular a su eje de rotación.
6. El autopodio se caracteriza por presentar metatarsales superpuestos en las porciones proximales expandidas y dígitos divergentes, no consolidados sensu Parrish (1986). Una consolidación del metatarso, incluyendo un dígito V dirigido anteriormente, provee una base estable para la flexión plantar con respecto al crus . De acuerdo a ello, la morfología pedal de Neoaetosauroides representaría una condición intermedia.
El estudio del esqueleto postcraneano de Neoaetosauroides y su comparación con el de otros aetosaurios ha puesto de manifiesto una gran diversidad anatómica en este grupo. Muchas diferencias en el grado de expansión pélvica, curvatura del fémur, morfología del crus y tarso sugieren que algunos miembros de este grupo pudieron tener una marcha semierecta, mientras que otros habrían tenido una locomoción erecta. Este posible espectro de posturas locomotoras, aunado a la amplitud de su rango de tamaño (0,80-5,00 m), sustenta las propuestas de una mayor gama de modos de vida entre estos arcosaurios acorazados que la tradicionalmente supuesta (Bonaparte, 1971b, 1978; Small, 2002; Desojo, 2003).
Agradecimientos
Las autoras agradecen a M.E. Gonaldi y S. Martin (Museo de La Rioja) y J. Powell (Instituto Miguel Lillo) por permitir el acceso a las colecciones bajo su cuidado. Igualmente agradecen a J. Bonaparte (Museo Argentino de Ciencias Naturales), C. Schultz y A. Arcucci por sus valiosas sugerencias y a J. Gonzáles por las ilustraciones. Hacen extensivo su agradecimiento a B. Small (Denver Museum of Natural Sciences) y a D. Pol (American Museum of Natural History) por sus comentarios. Este trabajo fue parcialmente financiado por The Samuel P. Welles Fund (University of California, Berkeley) a través de un Student Award a J.B.D., por el subsidio PIP CONICET 0535 (Dir. Andrea Arcucci) y por el subsidio UBACyT TX 01 de la Universidad de Buenos Aires a A.M.B.
Bibliografía
1. Benton, M.J. y Clark, J.M. 1988. Archosaur phylogeny and the relationships of the Crocodylia. En: M.J. Benton (ed.), The Phylogeny and Classification of the Tetrapods . Clarendon Press, Oxford, pp. 195-338. [ Links ]
2. Bonaparte, J.F. 1966. Una nueva "fauna" triásica de Argentina (Therapsida: Cynodontia y Dicynodontia). Consideraciones filogenéticas y paleobiogeográficas. Ameghiniana 4: 243-296. [ Links ]
3. Bonaparte, J.F., 1969, Los tetr ápodos triásicos de Argentina. 1º International Symposium of Gondwana (Mar del Plata, 1967): 307-325. [ Links ]
4. Bonaparte, J.F. 1971a. Annotated list of the South American Triassic tetrapods, 2º Gondwana Symposium (South Africa, 1970), Actas 3: 665-682. [ Links ]
5. Bonaparte, J.B. 1971b. Los tetr ápodos del sector superior de la Formación Los Colorados, La Rioja, Argentina (Triásico Superior). Opera Lilloana 22: 87-102. [ Links ]
6. Bonaparte, J.F. 1973. Edades/Reptiles para el Tri ásico de Argentina y Brasil, 5º Congreso Geológico Argentino (Buenos Aires, 1972): 93-128. [ Links ]
7. Bonaparte, J.F. 1978. El Mesozoico de Am érica del Sur y sus tetrápodos. Opera Lilloana (Tucumán), 26, 596 pp. [ Links ]
8. Bonaparte, J.F. 1984. Locomotion in rauisuchid thecodonts. Journal of Vertebrate Paleontology 3: 210-218. [ Links ]
9. Bonaparte, J.F. 1997. El Triásico de San Juan-La Rioja, Argentina, y sus Dinosaurio s. Digital Tech S:R:L, Museo Argentino de Ciencias Naturales, Buenos Aires, 190 pp. [ Links ]
10. Bossi, G.E. 1971. An álisis de la Cuenca Ischigualasto-Ischichuca. 1º Congreso Hispano-Luso-Americano de Geologia Económica (Madrid, Lisboa) Actas 2: 611-626. [ Links ]
11. Bossi, G.E. y Stipanicic, P.N. 2002. Ischigualasto (Formaci ón). En: P.N. Stipanicic y C.A. Marsicano (eds.), Triásico, Léxico Estratigráfico de la Argentina , Asociación Geológica Argentina, Serie B: Didáctica y Complementaria 26, pp. 149-150. [ Links ]
12. Camp, C.L. 1930. A study of the phytosaurs, with description of new material from western North America. Memoirs of the University of California 10: 1-161. [ Links ]
13. Casamiquela, R. 1960. Noticia preliminar sobre dos nuevos estagonolepoideos argentinos. Ameghiniana 2: 3-9. [ Links ]
14. Casamiquela, R. 1961. Dos nuevos Estagonolepoideos Argentinos (de Ischigualasto, San Juan). Revista de la Asociación Geológica Argentina 16: 143-203. [ Links ]
15. Casamiquela, R. 1967. Materiales adicionales y reinterpretaci ón de Aetosauroides scagliai (de Ischigualasto, San Juan). Revista del Museo de La Plata 5: 173-196. [ Links ]
16. Casamiquela, R.M. 1980. Nota sobre restos de un reptil aetosauroideo (Thecodontia, Aetosauria) de Quimal, Cordillera de Domeyko, Antofagasta. Prueba de la existencia del Neotri ásico continental en los Andes del Norte de Chile. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontologí a (Buenos Aires,1978), Anales 1: 135-142. [ Links ]
17. Case, E.C. 1920. Preliminary description of a new suborder of phytosaurian reptiles, with a description of a new species of Phytosaurus . Journal of Geology 28: 524-535. [ Links ]
18. Caselli, A.T., Marsicano, C.A y Arcucci, A.B. 2001. Sedimentolog ía y paleontología de la Formación Los Colorados, Triásico superior (provincias de La Rioja y San Juan, Argentina). Revista de la Asociación Geológica Argentina 56: 173-188. [ Links ]
19. Charig, A. 1972. The evolution of the archosaur pelvis and hindlimbs: an explanation in functional terms. En: K.A. Joysey y T.S. Kemp (eds.), Studies in vertebrate evolution . Oliver & Boyd, Edinburgh, pp. 121-155. [ Links ]
20. Chatterjee, S. 1978. A primitive parasuchid (phytosaur) from the Triassic Maleri Formation of India. Palaeontology 21: 83-127. [ Links ]
21. Chaterjee, S. 1985. Postosuchus , a new thecodontian reptile from the Triassic of Texas and the origin of tyrannosaurs. Philosophical Transactions of the Royal Society of London B 309: 395-460. [ Links ]
22. Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the American Philosophical Society 14: 1-252. [ Links ]
23. Cope, E.D. 1875. Report on the geology of that part of northwestern New Mexico examined during the field-season of 1874 : Annual Report upon the geographical explorations west of the 100th meridian (Wheeler Survery), Appendix LL, Annual Report Chief of Engineers for 1875: 61-97 of separate issue, 981-1017 of full report. [ Links ]
24. Da Rosa, A.A. y Leal, L.A. 2002. New elements of an armored archosaur from the middle to upper Triassic, Santa Maria Formation, South of Brazil. Arquivos do Museu Nacional , Rio de Janeiro, 60: 149-154. [ Links ]
25. de Ricql ès, A. Padian, K. y Horner, J.R. 2003. On the bone histology of some Triassic pseudosuchian archosaurs and related taxa. Annales de Paléontologie 89: 67-101. [ Links ]
26. Desojo, J.B. 2001. Chilenosuchus forttae : un aetosaurio del Triásico de Chile de relaciones filogenéticas y paleogeográficas controvertidas. Ameghiniana 38, Suplemento Resúmenes : 6R. [ Links ]
27. Desojo, J.B. 2002. New data on the Late Triassic aetosaur Neoaetosauroides Bonaparte, 1969 from the Los Colorados Formation, west-central Argentina. Journal of Vertebrate Paleontology 22, Supplement 3: 49A. [ Links ]
28. Desojo, J.B, 2003. Redescripci ón del aetosaurio Chilenosuchus forttae Casamiquela (Diapsida: Arcosauria): presencia de Triásico continental en el norte de Chile. Revista Geológica de Chile 30: 53-63. [ Links ]
29. Desojo, J. B, 2003. H ábitos alimenticios en los aetosaurios: ¿herbívoros, insectívoros o carnívoros?. Ameghiniana 40, Suplemento Resúmenes : 54 R. [ Links ]
30. Goodrich, E.S. 1958. Studies on the structure and development of vertebrates . Dover Publications, INC. New York, 485 pp. [ Links ] Gower, D.J. y Wilkinson, M. 1996. Is there any consensus on basal archosaur phylogeny? Proceedings of the Royal Society of London B 263: 1399-1406. [ Links ]
31. Gower, D.J. y Walker, A.D. 2002. New data on the braincase of the aetosaurian archosaur Stagonolepis robertsoni Agassiz. Zoological Journal of the Linnean Society 136:7-23. [ Links ]
32. Groeber, P.F.C. y Stipanicic, P.N. 1953. Tri ásico. En: P.F.C. Groeber (ed.), Mesozoico. Geografía de la Republica Argentina . Sociedad Argentina de Estudios Geográficos GAEA, Buenos Aires, pp. 13-141. [ Links ]
33. Harris, S.R., Gower, D.J. y Wilkinson, M. 2003a. Intraorganismal homology, character construction, and the phylogeny of aetosaurian archosaurs (Reptilia, Diapsida). Systematic Biology 52: 239-252. [ Links ]
34. Harris, S.R., Gower, D.J. y Wilkinson, M. 2003b. Phylogenetic methods and aetosaur interrelationships: a rejoinder. Systematic Biology 52: 851-852. [ Links ]
35. Heckert, A.B. y Lucas, S.G. 1999. A new aetosaur (Reptilia: Archosauria) from the Upper Triassic of Texas and the phylogeny of aetosaurs. Journal of Vertebrate Paleontology 19: 50-68. [ Links ]
36. Heckert, A.B. y Lucas, S.G. 2000. Taxonomy, phylogeny, biostratigraphy, biochronology, paleobiogeography, and evolution of the Late Triassic Aetosauria (Archosauria: Crurotarsi). Zentralblatt Geologie, Paläontologie I: 1539-1587. [ Links ]
37. Heckert, A.B. y Lucas, S.G. 2003. Clarifying aetosaur phylogeny requires more fossils, not more trees-Reply to intraorganismal homology, character construction, and the phylogeny of aetosaurian archosaurs (Reptilia, Diapsida). Systematic Biology 52: 253-255. [ Links ]
38. Heckert, A.B., Hunt, A.P. y Lucas, S.G. 1996. Redescription of Redondasuchus reseri , a Late Triassic aetosaur (Reptilia: Archosauria) from New Mexico (U.S.A), and the biochronology and phylogeny of aetosaurs. Geobios 29: 619-632. [ Links ]
39. Hunt, A.P. y Lucas, S.G. 1990. Re-evalution of Typothorax meadei , a Late Triassic aetosaur from the United States. Paläontologische Zeitschrift 64: 317-328. [ Links ]
40. Juul, L. 1994. The phylogeny of basal archosaurs. Palaeontologia Africana 31: 287-308. [ Links ]
41. Kokogian, D.A., Spalletti, L.A., Morel, E.M., Artabe, A.E., Mart ínez, R.N., Alcober, O.A., Milana, J.P. y Zavattieri, A.M. 2001. Estratigrafía del Triásico Argentino. En: A.E. Artabe, E.M. Morel y A.B. Zamuner (eds.), El Sistema Triásico en la Argentina , Fundación Museo de La Plata "Fransisco P. Moreno", La Plata, pp. 23-54. [ Links ]
42. Limarino, C., Fauque, L. y Caminos R. 1990. Facies y evoluci ón paleoambiental de los bancos rojos triásicos del norte de la Precordillera Riojana. 3º Reunión Argentina de Sedimentología (San Juan, 1990), Actas : 169-174. [ Links ]Long, R.A. y Ballew, K.L. 1985. Aetosaur dermal armor from the Late Triassic of southwestern North America, with special reference to material from the Chinle Formation of Petrified Forest National Park. Museum of Northern Arizona Bulletin 54. 45-68. [ Links ]
43. Long, R. y Murry, P.A. 1995. Late Triassic (Carnian and Norian) tetrapods from the Southwestern United States. New Mexico Museum of Natural History and Science Bulletin 4. 27-116. [ Links ]
44. López Gamundí, O., Alvarez, L., Andreis, R., Bossi, G.E., Espejos, I., Fernández Seveso, F., Legarreta, L., Kokogian, D.A., Limarino, C.O. y Sessarego, H.L. 1989. Cuencas Intermontanas. En: G.A. Chebli y L.A. Spalletti (eds.), Cuencas Sedimentarias Argentinas . Universidad Nacional de Tucumán, Serie de Correlación Geológica, San Miguel de Tucumán, pp. 123-167. [ Links ]
45. Lucas, S.G. y Heckert, A.B. 2001. The aetosaur Stagonolepis from the Upper Triassic of Brazil and its biochronological significance. Neues Jahrbuch für Geologie und Paläontologie Monatshefte 12: 719-732. [ Links ]
46. Lucas, S.G., Heckert, A.B. y Kahle, R. 2000. Postcranial anatomy of Angistorhinus , a Late Triassic phytosaur from west Texas. New México Museum or Natural History & Science Bulletin , 21: 157- 164. [ Links ]
47. Marsh, O.C. 1884. The classification and affinities of dinosaurian reptiles. Nature , 31: 68-69. [ Links ] Milana, J.P. y Alcober, O.A. 1995. Modelo tectosedimentario de la cuenca triásica de Ischigualasto (San Juan, Argentina). Revista de la Asociación Geológica Argentina 49: 217-235. [ Links ]
48. Nash, D.S. 1975. The morphology and relationships of a crocodilian, Orthosuchus stormbergi , from the Upper Triassic of Lesotho. Annals of the South African Museum 67: 227-329. [ Links ]Parrish, J.M. 1986. Locomotor adaptations in the hindlimb and pelvis of the Thecodontia. Hunteria 1: 1-35. [ Links ]
49. Parrish, J.M. 1993. Phylogeny of the Crocodylotarsi, with reference to archosaurian and crurotarsan monophyly. Journal of Vertebrate Paleontology 13: 287-308. [ Links ]
50. Reilly, S.M. y Elias, J.A. 1998. Locomotion in Alligator mississippiensis : kinematic efects of speed and posture and their relevance to the sprawling-to-erect paradigm. The Journal of Experimental Biology 201: 2559-2574. [ Links ]
51. Romer, A.S. 1956. Osteology of the Reptiles . University of Chicago Press, Chicago, 772 pp. [ Links ]
52. Romer, A.S. y Jensen, J. 1966. The Cha ñares (Argentina). Triassic reptiles fauna, 2 Sketch of the geology of the Rio Chañares-Rio Gualo region. Breviora 252: 1-20. [ Links ]
53. Sawin, H.J. 1947. The pseudosuchian reptile Typothorax meadei . Journal of Paleontology 21: 201-238. [ Links ]
54. Sereno, P.C. 1991. Basal archosaurs: phylogenetic relationships and functional implications. Journal of Vertebrate Paleontology 11, Supplement 4, 53 pp. [ Links ]
55. Sereno, P.C. y Arcucci, A.B. 1990. The monophyly of crurotarsal archosaurs and the origin of bird and crocodile ankle joints. Neues Jahrbuch für Geologie und Paläontologie 180: 21-52. [ Links ]
56. Sill, W.D. 1974. The anatomy of Saurosuchus galilei and the relationships of the rauisuchid thecodonts. Bulletin of the Museum of Comparative Zoology , Harvard University, 146: 317-362. [ Links ]
57. Small, B.J. 1985. [ The Triassic thecodontian reptile Desmatosuchus : osteology and relationships . MSc Thesis, Texas Tech University, Texas, 83 pp. Inédito] [ Links ].
58. Small, B.J. 2002. Cranial anatomy of Desmatosuchus haplocerus (Reptilia: Archosauria: Stagonolepididae). Zoological Journal of the Linnean Society 136: 97-111. [ Links ]
59. Stipanicic, P.N. y Bonaparte, J.F. 1979. Cuenca Tri ásica de Ischigualasto-Villa Unión (Provincia de La Rioja y San Juan). 2º Simposio de Geología Regional Argentina (Córdoba, 1976), Actas 1: 523-575. [ Links ]
60. Walker, A.D. 1961. Triassic reptiles from the Elgin Area: Stagonolepis , Dasygnathus , and their allies. Philosophical Transactions of the Royal Society of London 244: 103-204. [ Links ]
61. Walker, A.D. 1964. Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs. Philosophical Transactions of the Royal Society B. 248: 53-134. [ Links ]
62. Walker, A.D. 1990. A revision of Sphenosuchus acutus Haughton, a crocodylomorph reptile from the Elliott Formation (late Triassic or early Jurassic) of South Africa. Philosophical Transactions of the Royal Society of London B 330: 1-120. [ Links ]
63. Wu, X.C., Sues, H.D. y Dong, Z.M. 1997. Sichuanosuchus shuhanensis , a new ?Early Cretaceous protosuchian (Archosauria: Crocodyliformes) from Sichuan (China) and the monophyly of Protosuchia. Journal of Vertebrate Paleontology 17: 89-103. [ Links ]
Recibido: 12 de febrero de 2004.
Aceptado: 29 de abril de 2004.

