










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.1 Buenos Aires ene./mar. 2005
Distribución de Nonionella auris (d'Orbigny) (Foraminiferida) en el golfo San Matías, provincia de Río Negro, Argentina
Emiliana Bernasconi 1 y Gabriela Cusminsky 1
1 Centro Regional Universitario Bariloche. Universidad Nacional del Comahue. Quintral 1250. 8400 Bariloche / Consejo Nacional de Investigaciones Científicas y Técnicas. Argentina. emibernasconi@yahoo.com.ar gcusmins@crub.uncoma.edu.ar
Resumen. Se estudia la influencia del tipo de sedimento y la profundidad en la distribución y desarrollo del foraminífero bentónico Nonionella auris (d'Orbigny). Se analizaron muestras de sedimentos superficiales provenientes del golfo San Matías, provincia de Río Negro, Argentina. Los estudios cualitativos, cuantitativos y estadísticos realizados sugieren que sitios con mayor profundidad, mayor contenido de fango y bajo nivel de energía propiciarían el desarrollo de esta especie.
Abstract. Distribution of Nonionella auris (D´Orbigny) (Forminiferida) in San Matías gulf, Río Negro province, Argentina. Qualitative, quantitative and statistic analysis on samples of surface sediments from San Matías Gulf, Río Negro Province, Argentina, reveal a relationship between the type of substrate- depth and characteristics and distribution of the benthic foraminifer Nonionella auris (d´Orbigny). The study suggests that the development of this species is favoured in low energy and deep environments with muddy substrates.
Key words. Foraminifera. Nonionella auris . Distribution. San Matías gulf. Argentina.
Palabras clave. Foraminifera. Nonionella auris . Distribución. Golfo San Matías. Argentina.
Introducción
Los foraminíferos bentónicos son organismos que están ampliamente distribuidos en los ambientes marinos. La abundancia y diversidad de sus especies están afectadas por factores ambientales tales como profundidad, tipo de sustrato, temperatura, salinidad y contenido de oxígeno entre otros. En este trabajo se presentan los resultados de los estudios referidos a la distribución de foraminíferos bentónicos, especialmente de Nonionella auris ( d´Orbigny), en un ambiente engolfado.
Estudios anteriores realizados en el área del golfo San Matías y la plataforma continental adyacente verificaron la presencia de N. auris sólo en el ambiente engolfado (Bernasconi et al. , 2001, 2002). Sin embargo, otros investigadores mencionaron a esta especie en el ambiente de plataforma actual (Boltovskoy et al. , 1980). Malumián (1972) la cita para el Oligoceno y Mioceno del sur de la provincia de Buenos Aires y Cusminsky (1986) para sedimentos cuaternarios de un testigo del talud continental argentino. Esto motivó el interés por analizar cómo influyen los factores ambientales, especialmente el tipo de sedimento y la profundidad, en la distribución y desarrollo de N. auris en el golfo San Matías.
Por otra parte el estudio de la distribución de foraminíferos actuales contribuye al conocimiento de la evolución de la plataforma continental argentina, ya que los microorganismos presentes en ella pueden ser utilizados para inferir las variaciones paleoclimáticas, paleoceanográficas y paleoecológicas.
Área de estudio
El área de estudio se encuentra en el sector del golfo San Matías, provincia de Río Negro, aproximadamente entre los 40o 30' y 42o 30' S y los 63o 30'y 65o O (figura 1).
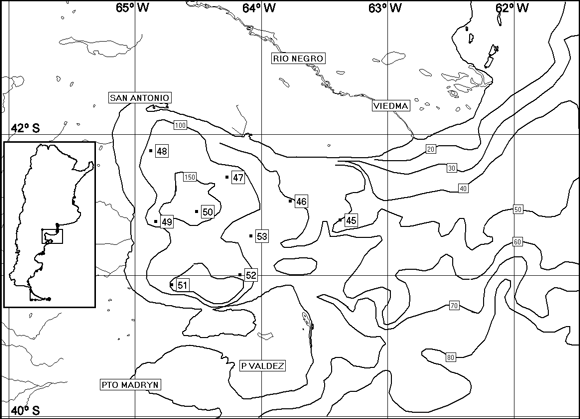
Figura 1. Mapa de ubicación de las muestras 45 a 53, del golfo San Matías / locality map of samples 45 to 53, from San Matías gulf.
Desde el punto de vista morfológico el golfo San Matías fue analizado por Parker et al. (1996). En líneas generales, se puede mencionar que este golfo constituye una verdadera depresión cerrada, presentando profundidades que superan los 190 metros. Una característica del sector bajo estudio la constituye la falta de ríos que viertan caudales significativos y, consecuentemente, la ausencia de sedimentos actuales de tipo fluvial (Gelós et al. , 1988).
Desde el punto de vista sedimentológico la depresión central del golfo está cubierta por sedimentos arcillosos de coloración verde. Su origen puede vincularse a sedimentos actuales transportados en suspensión por el río Negro y llevados hacia el interior del golfo por corrientes de marea que circulan en sentido contrario a las agujas del reloj, o bien a un depósito relíctico correspondiente a un antiguo sistema de drenaje dentro del golfo (Gelós et al. , 1988).
Material y métodos
Se estudiaron 9 muestras extraídas mediante un muestreador de tipo Shipek (con el Buque Oceanográfico Puerto Deseado), en la campaña realizada por el Instituto Argentino de Oceanografía (IADO), en 1984. Sobre la base de los estudios sedimentológicos realizados por Gelós et al. (1988) y Gelós y Spagnuolo (1992) se confeccionó una tabla con los datos de ubicación, profundidad y tipo de sedimento para cada estación analizada (cuadro 1). Se estudió la relación existente entre la profundidad y el tipo de sedimento, o sea entre profundidad y fango, profundidad y arena y profundidad y grava. Debido a que se contaba con pocos datos y al no cumplir los mismos con una distribución normal se debió utilizar el test no paramétrico de Spearman. En este test se ordenan los valores de ambas variables x, y por separado. Luego se suman los cuadrados de las diferencias del orden de las variables x, y (d 1..n 2 ) tomadas de a pares, donde n es el número de muestras.

Cuadro 1. Ubicación, profundidad y tipo de sedimento para cada estación. E =estación, F = % de fango, A = % de arena, G = % de grava, P = profundidad (m) / location, depth and type of sediment in each station. E = station , F = % of mud , A = % of sand , G = % of gravel , P = depth (m) .
El coeficiente de Spearman está dado por (Daniel, 1990):
Rs = 1 - 6 (d 1 2 + d 2 2 + ... + d n 2 )/(n(n 2 -1))
Las muestras fueron lavadas con agua a través de un tamiz de 63 µm de abertura (n o 230 del Tyler Screen System ). Posteriormente se revisaron 5 g de se Nonionella auris en el golfo San Matías, Argentina 169 dimento seco y se extrajo la totalidad de los ejemplares presentes bajo lupa binocular. La determinación sistemática de los organismos a nivel genérico se basó en Loeblich y Tappan (1987) mientras que a nivel específico se usó Boltovskoy (1954a, 1954b, 1959), Boltovskoy y Wright (1976), Boltovskoy et al. (1980) y Cusminsky (1986, 1992). Los ejemplares más representativos fueron fotografiados por medio del microscopio electrónico de barrido (Philips, Modelo SEM 515 del Centro Atómico Bariloche). Los ejemplos se encuentran depositados en el laboratorio de Micropaleontología del Museo de La Plata bajo los números MLP-Mi 1352 a 1359.
Se realizaron estudios cualitativos y cuantitativos determinando la abundancia total, abundancia relativa y diversidad (S) como número de especies por muestra. Se calculó la diversidad de Shannon-Wiener, dada por la ecuación (Buzas y Gibson, 1969):
S
H´(S)= - S pi . ln pi
i=1
donde S es el número de especies y pi la proporción de cada especie en la muestra (cuadro 2).

Cuadro 2. Abundancia relativa de las especies encontradas, abundancia total y diversidad de Shannon-Wiener para cada estación / relative abundance of species, total abundance and Shannon-Wiener diversity in each station.
Para la especie N. auris se realizó un análisis de frecuencia de tamaños para aquellas estaciones donde se encontró más de un ejemplar. Se estableció como hipótesis nula la no existencia de diferencias significativas entre los tamaños en las 5 muestras. Para contrastar dicha hipótesis y dado que las muestras a analizar no presentan una distribución normal debió procederse al uso de contrastes no paramétricos. Se realizaron dos contrastes, primero con el test de Kruskal Wallis (Daniel, 1990) y el segundo con el test de Mann-Whitney (Daniel, 1990). En el primer caso la hipótesis nula fue que no existían diferencias significativas entre las muestras. El estadístico H calculado por este test queda determinado como
k
H= 12 / N (N + 1) S R j 2/ n j -3 (N + 1)
J=1
donde k es el número de muestras, n j el número de casos en la muestra de orden j, N el número de casos de todas las muestras combinadas, R j la suma de rangos en la muestra de orden j (Daniel, 1990).
En el segundo caso el test establece la misma hipótesis nula pero tomando las muestras de a pares. El estadístico calculado por este test es U dado por:
U= n 1 n 2 + (n1 (n 1 + 1)) / 2 - R 1
donde n 1 es el número de casos de la nuestra 1, n 2 el número de casos de la muestra 2 y R 1 es la suma de los rangos de la muestra 1 (Daniel, 1990). Ambos tests ponen a prueba diferencias entre medianas de muestras independientes y en ambos se ordenan los valores como si fuese una sola muestra (Daniel, 1990). Si se rechaza la hipótesis nula planteada en el primer caso, es decir se constata que existe diferencia significativa entre las muestras, se procede a comparar cada grupo con todos los demás con el test de Mann-Whitney. El objetivo es dilucidar si la diferencia de tamaños encontrada en las distintas estaciones es significativa o no.
Resultados
En las estaciones muestreadas la abundancia fluctúa entre 10 y 895 organismos, siendo la de menor abundancia la estación 46 y la de mayor abundancia la estación 50. La diversidad específica de las muestras es baja con valores de H´(S) que van de 0,52 a 1,78 mientras que el S varió entre 3 y 15. Los valores de H´(S) más bajos 0,52 y 0,65 coinciden a su vez con valores muy bajos de S (5), como así también los valores más altos de H´(S) (1, 78 y 1,75) coinciden con los valores más altos de S (12 y 15). También se pudo observar que los valores de H´(S) más bajos se presentaron en las estaciones 50 y 51, donde N. auris se presenta con mayor abundancia, representando más del 80% de la fauna encontrada en ambas estaciones.
Nonionella auris se halló en seis de las nueve estaciones analizadas; es muy abundante en las estaciones 50 y 51 y abundante en las estaciones 47 y 48. Fue encontrada principalmente asociada a Bulimina patagonica d´Orbigny, Buccella peruviana campsi (Boltovskoy) y Bulimina affinis (d´Orbigny) (cuadro 2).
La especie bajo estudio fue encontrada con mayor abundancia en aquellas estaciones donde la profundidad es mayor (estaciones 50 y 51); a su vez dicha abundancia disminuye en general al disminuir la profundidad, no encontrándose ejemplares de esta especie en estaciones con menores profundidades, tales como en las estaciones 45, 46 y 53 (véase cuadro 1).
El coeficiente de Spearman, al analizar la relación de la profundidad con el contenido de fango, fue Rs= 0.86 con p=0,0026; con el porcentaje de arena fue Rs= -0,65 con un p= 0,0539 y con el contenido de grava resultó Rs= -0,48 con un p= 0,1875. De los análisis surge que, en las estaciones analizadas, la profundidad se encuentra asociada positivamente con el contenido de fango y que esta asociación es, desde el punto de vista estadístico, altamente significativa. Sin embargo esto no ocurre cuando se analiza la relación de la profundidad con el porcentaje de arena y menos aún cuando se la relaciona con el contenido de grava, dado que sólo una de las estaciones contenía grava en su sedimento.
El rango de tamaños en N. auris va de 100 a 600 µm (figura 2). Al comparar la diferencia de tamaños de los ejemplares de todas las estaciones se encontró que el coeficiente de Kruskal Wallis (H) fue igual a 59,6 con un nivel de significación del test de p= 0,0001 indicando que existe una diferencia de tamaños estadísticamente significativa. Luego se compararon los tamaños de las estaciones tomadas de a pares con el test de Mann Whitney (U), encontrándose diferencia estadísticamente significativa sólo entre los tamaños de las estaciones 47 y 51 (U = 11427,0 y p= 0,0001); 48 y 51 (U= 24266,0 y p= 0,00493) y entre 50 y 51 (U= 157450,5 y p= 0,0001). La diferencia de tamaños de las estaciones 47 y 51 y 48 y 51 podría ser explicada por las condiciones reinantes en cada estación; ambas diferencias podrían estar determinadas por la profundidad y el contenido de fango. Puede observarse también que las poblaciones de N. auris en las estaciones 50 y 51 son las más parecidas en cuanto al rango de tamaños que poseen. La estación 50 abarca un rango de tamaños de la población mucho más amplio que el resto de las estaciones, incluyendo los tamaños mínimos y máximos de todos los ejemplares hallados. En la estación 51 el rango de tamaños encontrados es también muy amplio aunque no abarca los tamaños más pequeños y más grandes encontrados en la estación 50; ambas estaciones poseen el porcentaje de fango más alto, sin embargo en este caso la profundidad es mayor en la estación 50, siendo esta última la que estaría determinando mejores condiciones aún para el desarrollo de la especie bajo estudio (figura 2). Todo lo antes expuesto estaría corroborando que la diferencia de tamaños entre esas estaciones se debería al contenido de fango, el cual está relacionado con la profundidad.
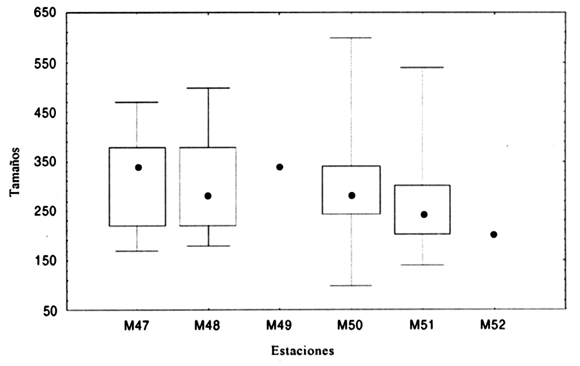
Figura 2. Gráfico mostrando los valores de las medianas (), los tamaños mínimos y máximos ( I ) y el 50 % de la población de N. auris en cada estación (); tamaños en µm / Graph showing medium value (), minimum and maximum values ( I ) a nd 50 % of population of N. auris in each station (); size in µm.
Se presenta una lámina con los ejemplares más representativos de N. auris de cada estación (figura 3).

Figura 3. A-I, Nonionella auris (d´Orbigny). A, vista umbilical / umbilical view , E 47, MLP-Mi 1352. B, vista apertural / apertural view , E 47, MLP-Mi 1353. C, vista umbilical / umbilical view , E 48, MLP-Mi 1354. D, vista espiral / spiral view , E 48, MLP-Mi 1355. E, vista espiral / spiral view , E 50, MLP-Mi 1356. F, vista umbilical / umbilical view , E 50, MLP-Mi 1357. G, detalle de pared / detail of the wall , E 50, MLP-Mi 1356. H, vista apertural / apertural view , E 50, MLP-Mi 1358. I, vista umbilical / umbilical view , E 50, MLP-Mi 1359. Escala = 50 µm (figuras A, B, D, I); 100 µm (figuras C, E, F, H); 5 µm (figura G)/ Scale bar = 50 µm (figures A, B, D, I); 100 µm (figures C, E, F, H); 5 µm
Comentarios
Los factores que influyen en la distribución de los foraminíferos bentónicos son temperatura, salinidad, contenido de nutrientes, profundidad, tipo de sedimento y contenido de oxígeno entre otros (Anderson, 1963; Boltovskoy, 1963; 1965; Boltovskoy y Wright, 1976; Boltovskoy et al. , 1980; Murray, 1969). La combinación de esos factores ambientales afecta la distribución de dichos microorganismos, siendo de mayor influencia los parámetros que se relacionan con la profundidad (Hayward et al. , 2002), y éstos a su vez relacionados con el tipo de sedimento (Murray, 1991).
Concretamente con respecto a este último, Hayward et al. (1996) consideraron al porcentaje de fango como uno de los factores que más afecta la distribución de los foraminíferos bentónicos recientes. A similares conclusiones arribaron Boltovskoy (1965) y Bernhard (1986).
Yanko et al. (1998) han sugerido también que el tipo de sedimento podría afectar tanto el tamaño como la forma de las conchillas de los foraminíferos, mostrando entre otros un crecimiento aberrante de muchos especímenes de Noninella atlantica Cushman. Además en los sedimentos finos se encuentra una mayor cantidad de materia orgánica, lo que significaría mayor disponibilidad de alimento. Como consecuencia de ello la asociación de foraminíferos se desarrollaría con un número mayor de individuos (Boltovskoy, 1963).
Resumiendo las observaciones realizadas por autores como Severin (1983/84) y Corliss y Chen (1988), los foraminíferos pueden ser reunidos en diferentes morfogrupos, pudiéndose inferir a partir de ellos su modo de vida y su modo de alimentación. Estudiando los morfogrupos independientemente del nivel taxonómico se pueden distinguir biofacies, y esto podría tener aplicación en estudios paleoambientales. El género Nonionella se incluye dentro del morfogrupo planoespiral redondeado: la relativa simetría de los especímenes correspondientes a este grupo estaría indicando que se trata de organismos generalistas, con la simetría suficiente para evitar orientaciones desfavorables en sedimentos disturbados, pero con la asimetría necesaria como para tomar una orientación preferencial en sedimentos estables. Las características de Nonionella según su forma y disposición de las cámaras estarían indicando un microhábitat infaunal. Además se trata de un taxón detritívoro de vida libre y se lo encuentra entre los 10-1000 m; puede habitar ambientes marinos de salinidad normal hasta salobres (Murray, 1991).
Nonionella ha sido descripto como un género que vive en un medio fangoso (Murray, 1991), tal como lo encuentran Harloff y Mackensen (1997) para los sitios donde predomina el contenido de fango al de arena o grava. Hayward et al. (2002) realizaron un análisis mediante el cual determinaron que el porcentaje de fango aumenta con la profundidad y que son justamente los factores que tienen relación con la profundidad los más significativos en la influencia de la distribución de los foraminíferos. También sus análisis sugieren que la presencia de N. auris tiene una relación positiva con la mayor concentración de fango.
Los resultados hallados en este trabajo señalan que las estaciones analizadas con una mayor abundancia de ejemplares, poseen una diversidad y diversidad específica baja. Esto podría explicarse teniendo en cuenta las observaciones realizadas por los trabajos previos mencionados. En efecto, la mayor abundancia de ejemplares estaría relacionada con el sedimento de grano fino, el cual propicia el incremento de la materia orgánica y por consiguiente provee un mayor alimento al medio. Si bien hay una gran abundancia de individuos, los mismos son fundamentalmente de N. auris . Esto evidenciaría que el sedimento de tipo fango sería el más favorable para el desarrollo de esta especie.
Nonionella auris es citada como una especie poco frecuente para la costa de América del Sur (Boltovskoy, 1954a; 1957; Cusminsky, 1986; Malumián, 1972) y de Australia (Yassini y Jones, 1995). Fue encontrada tanto en la zona de plataforma (Malumián, 1972; Boltovskoy et al. , 1980; Cusminsky, 1986) como en ambientes engolfados (Boltovskoy, 1954a), de estuarios (Boltovskoy, 1957) y de bahías (Boltovskoy, 1954b). También fue encontrada por Yassini y Jones (1995) en zonas de bahías, estuarios y plataforma interna, y en aguas frías por Boltovskoy (1954a, 1965) y Boltovskoy y Theyer (1970).
Conclusiones
En el presente estudio N. auris fue encontrada en un ambiente engolfado, a profundidades que van desde los 75 a los 150 m. A partir de consideraciones cualitativas y de la aplicación de parámetros cuantitativos y estadísticos se concluye que la mayor abundancia de esta especie se registró en las estaciones ubicadas a mayor profundidad coincidiendo con un substrato de tipo fangoso, el cual sería más propicio para su desarrollo. Estos hallazgos corroboran los estudios realizados por otros autores y llevan a inferir que la distribución de esta especie en el golfo San Matías estaría afectada por la proporción de fango contenida en el sedimento y por la profundidad. Esto tiene una directa relación con la energía del medio. En los ambientes con altos niveles de energía las condiciones no serían las más favorables para la vida de esta especie. Los ambientes con menor nivel de energía, donde predominan los sedimentos finos, serían más propicios para el desarrollo de N. auris .
Agradecimientos
Este trabajo se realizó en el Departamento de Ecología de la Universidad Nacional del Comahue y constituye una contribución a los proyectos financiados por la Agencia de Promoción Científica y Tecnológica (BID 1201/OC- AR PICT 07 - 09659). Las autoras desean agradecer al Instituto Argentino de Oceanografía (IADO) por haber facilitado las muestras analizadas. A los revisores ya que con sus correcciones y sugerencias enriquecieron este trabajo y a A. Farji-Brenner por revisar los análisis estadísticos realizados. Las autoras desean dedicar especialmente este trabajo a la memoria de Alwine Bertels.
Bibliografía
1. Anderson, G. 1963. Distribution patterns of Recent foraminifera of the Bering Sea. Micropaleontology 9: 305-317. [ Links ]
2. Bernasconi, E., Bertels, A. y Cusminsky, G. 2001. Comparación del Orden Foraminiferida en dos paleoambientes en la plataforma continental argentina. 1ra Reunión Binacional de Ecología (20a Reunión Argentina de Ecología - 10a Reunión de la Sociedad de Ecología de Chile) (Bariloche), Resúmenes , p. 57. [ Links ]
3. Bernasconi, E., Bertels, A. y Cusminsky, G. 2002. Foraminíferos cuaternarios bentónicos y su relación con el medio. 8º Congreso Argentino de Paleontología y Bioestratigrafía (Corrientes), Resúmenes , p. 42. [ Links ]
4. Bernhard, J. 1986. Characteristic assemblages and morphologies of benthic foraminifera from anoxic, organic-rich deposits: Jurassic through Holocene. Journal of Foraminiferal Research 16: 207-215. [ Links ]
5. Boltovskoy, E. 1954a. Foraminíferos del Golfo de San Jorge. Revista del Instituto Nacional de Investigación de las Ciencias Naturales y Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". Ciencias Geológicas 3: 85-246. [ Links ]
6. Boltovskoy, E.1954b. Foraminíferos de la Bahía San Blas. Revista del Instituto Nacional de Investigación de las Ciencias Naturales y Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Ciencias Geológicas 3: 247-300. [ Links ]
7. Boltovskoy, E.1957. Los Foraminíferos del estuario del Río de La Plata y su zona de influencia. Revista del Instituto Nacional de Investigación de las Ciencias Naturales y Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Ciencias Geológicas 6: 1-78. [ Links ]
8. Boltovskoy, E. 1959. Recent Foraminifera of southern Brazil and their relation with those of Argentina and West. Revista del Servicio de Hidrografía Naval H. 1005, 124 pp. [ Links ]
9. Boltovskoy, E. 1963. Foraminiferos y sus relaciones con el medio. Revista del Museo Argentino de ciencias Naturales "Bernardino Rivadavia" y Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Hidrobiología 1: 21-107. [ Links ]
10. Boltovskoy, E. 1965. Los foraminíferos recientes . EUDEBA, Buenos Aires, Argentina, 510 pp. [ Links ]
11. Boltovskoy, E y Theyer, F. 1970. Foraminiferos recientes de Chile central. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" e Instituto Nacional de Investigación de las Ciencias Naturales , Hidrobiología 2: 379 pp. [ Links ]
12. Boltovskoy, E.y Wright, R.1976. Recent Foraminifera . W. Junk. Publishers, The Hague, 515 pp. [ Links ]
13. Boltovskoy, E., Giussani, G. Watanabe, S. y Wright, R. 1980. Atlas of benthic shelf foraminifera of the southwest Atlantic . W. Junk. Publishers, The Hague, 147 pp. [ Links ]
14. Buzas, M. A. y Gibson, T. G. 1969. Species diversity: Benthonic foraminifera in Western North Atlantic. Science 163: 72-75. [ Links ]
15. Corliss, B. y Chen, C. 1988. Morphotype patterns of Norwegian Sea deep-sea benthic foraminifers and ecological implications. Geology 16: 716-719. [ Links ]
16. Cusminsky, G. 1986. Foraminíferos de un testigo del talud continental argentino. 4º Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 3: 167-183. [ Links ]
17. Cusminsky, G. 1992. Foraminíferos bentónicos provenientes de testigos del océano Atlántico sudoccidental austral. Revista Española de Micropaleontología 24: 5-32. [ Links ]
18. Daniel, W.W. 1990. Applied Nonparametric Statistics . PWS-Kent publishing company. 510 pp. [ Links ]
19. Gelós, E. y Spagnuolo, J. 1992. Relación área de aporte-ambiente tectónico de las psamitas de la plataforma continental argentina entre los 39º y 43º de latitud sur y el Golfo San Matías. Revista de la Asociación Geológica Argentina 47: 141- 146. [ Links ]
20. Gelós, E., Spagnuolo, J. y Lizasoain, G. 1988. Mineralogía y caracterización granulométrica de sedimentos actuales de la plataforma argentina entre los paralelos 39º y 43º de latitud Sur y del Golfo San Matías. Revista de la Asociación Geológica Argentina 63: 63-79. [ Links ]
21. Harloff, J. y Mackensen, A. 1997. Recent benthic foraminiferal associations and ecology of the Scotia Sea and Argentine Basin. Marine Micropaleontology 31: 1-29. [ Links ]
22. Hayward, B., Grenfell, H., Cairns, G. y Smith, A. 1996. Environmental controls on benthic foraminiferal and thecamoebian asociations in a New Zealand tidal inlet. Journal of Foraminiferal Research 26: 150-171. [ Links ]
23. Hayward, B., Neil, H., Carter, R., Grenfell, H. y Hayward, J. 2002. Factors influencing the distribution patterns of Recent deep-sea benthic foraminifera, east of New Zealand, Southwest Pacific Ocean. Marine Micropaleontology 46: 139-176. [ Links ]
24. Loeblich, A. Jr. y Tappan, H. 1987. Foraminiferal Genera and their Classification . Van Nostrand Reinhold Company, New York, 1-2: 1182 pp. [ Links ]
25. Malumián, N. 1972. Foraminíferos del Oligoceno y Mioceno del subsuelo de la provincia de Buenos Aires. Ameghiniana 9: 97-137. [ Links ]
26. Murray, J.W. 1969. Recent foraminifers from the Atlantic continental shelf of the United States. Micropaleontology 15: 401-419. [ Links ]
27. Murray, J.W. 1991. Ecology and Paleoecology of Benthic Foraminifera . Longman, Wiley, Harlow/Essex, New York, 397 pp. [ Links ]
28. Parker, G., Violante, R. y Paterlini, M. 1996. Fisiografía de la Plataforma Continental. En: V. Ramos y M. Turic (eds.), Geología y Recursos Naturales de la Plataforma Continental Argentina . Buenos Aires, 1-16. [ Links ]
29. Severin, K. 1983/84. Test morphology of benthic Foraminifera as a discriminator of biofacies. Marine Micropaleontology 8: 65-76. [ Links ]
30. Yanko, V., Ahmad, M. y Kaminski, M. 1998. Morphological deformities of benthic foraminiferal tests in response to pollution by heavy metals: implications for pollution monitoring. Journal of Foraminiferal Research 28: 177-200. [ Links ]
31. Yassini, I. y Jones, B. 1995. Foraminiferida and Ostracoda from estuarine and shelf environments on the southeastern coast of Australia . University of Wollongong Press, Australia, 484 pp. [ Links ]
Recibido: 21 de abril de 2003.
Aceptado: 17 de junio de 2004.

