










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.1 Buenos Aires ene./mar. 2005
Pseudoplohophorus absolutus n. sp. (Xenarthra, Glyptodontidae), variabilidad en Sclerocalyptinae y redefinición de una biozona del Mioceno Superior de Uruguay
Daniel Perea 1
1 Departamento de Evolución de Cuencas, Facultad de Ciencias, Iguá 4225, 11400 Montevideo, Uruguay. perea@fcien.edu.uy
Abstract. Pseudoplohophorus absolutus n. sp. (Xenarthra, Glyptodontidae), Sclerocalyptinae variability and a biozone redefinition from the Upper Miocene of Uruguay . A species of glyptodontid, Pseudoplohophorus absolutus n. sp. from the Upper Miocene of Uruguay is described. The holotype is represented by skull, mandible, great part of dorsal and cephalic carapace, caudal tube, humerus, hand bones, some vertebrae, and fragmentary pelvis and caudal ring plates. These materials were compared with very complete specimens of Neogene Sclerocalyptinae from Argentina and Uruguay. On the basis of these comparisons the partial synonymy between Pseudoplohophorus Castellanos and Stromaphoropsis Kraglievich, and the inclusion of genus Hoplophractus Cabrera within Eosclerocalyptyus C. Ameghino are proposed. A biozone characterized by Pseudoplohophorus , included in Camacho Formation deposits, is redefined.
Resumen. Se describe una especie de gliptodonte, Pseudoplohophorus absolutus n. sp. procedente del Mioceno Superior de Uruguay. El holotipo está representado por cráneo, mandíbula, gran parte de la coraza dorsal cefálica, tubo caudal, húmero, huesos de la mano, algunas vértebras, fragmentos de pelvis y placas de anillos caudales. Estos materiales fueron comparados con ejemplares muy completos de Sclerocalyptinae neógenos de Argentina y Uruguay. A base de estas comparaciones se propone la sinonimia parcial de Pseudoplohophorus Castellanos con Stromaphoropsis Kraglievich y la inclusión del género Hoplophractus Cabrera dentro de Eosclerocalyptyus C. Ameghino. Se redefine una biozona caracterizada por el género Pseudoplohophorus incluida en depósitos de la Formación Camacho.
Key words. Glyptodontidae. Upper Miocene. Uruguay. Systematics, Biostratigraphy.
Palabras clave. Glyptodontidae. Mioceno Superior. Uruguay. Sistemática. Bioestratigrafía.
Introducción
Los gliptodontes, por poseer un elevado número de huesos que no siempre fosilizan articulados o en asociación física, han sido objeto de multiplicación taxonómica. Prueba de ello es que se registran en la literatura unos 75 géneros nominados para América del Sur (Mones, 1986). Numerosos taxones se definieron a base de restos muy escasos y fragmentarios como placas sueltas, tubos caudales, fragmentos de coraza, etc. Claros ejemplos de este sobredimensionamiento sistemático pueden verse en las obras de Ameghino (1889) y Castellanos (1940), entre otros. Es probable que parte de lo que se interpreta como categorías taxonómicas discretas, esté reflejando variaciones individuales o poblacionales. Los Sclerocalyptinae neógenos de Argentina y Uruguay no escapan a esta situación. Ellos han sido estudiados principalmente por Kraglievich (1932), Cabrera (1939,1944) y Castellanos (1940, 1949, 1954). El material más completo de Sclerocalyptinae neógenos incluye cráneos asociados a sus respectivas corazas y eventualmente tubos caudales u otras piezas procedentes principalmente de Argentina y Uruguay. Reúnen esta condición ejemplares previamente nominados como Eosclerocalyptus C. Ameghino, Stromaphorus Castellanos, Phlyctaenopyga Cabrera, Plohophorus Ameghino y Hoplophractus Cabrera, pertenecientes al Museo Argentino de Ciencias Naturales, al Museo de La Plata y al Field Museum of Natural History y dos especímenes asignados en el presente trabajo al género Pseudoplohophorus Castellanos, hallados en Uruguay. Uno de ellos es el holotipo de " Stromaphoropsis " benvenutii Castellanos, y el otro constituye la nueva especie aquí descripta, considerada preliminarmente como perteneciente al género Stromaphoropsis Kraglievich ( vide Perea, 1993; Perea et al. , 1994). El análisis pormenorizado de estos especímenes permite enmarcar la información existente sobre material más fragmentario registrado en Uruguay y Argentina.
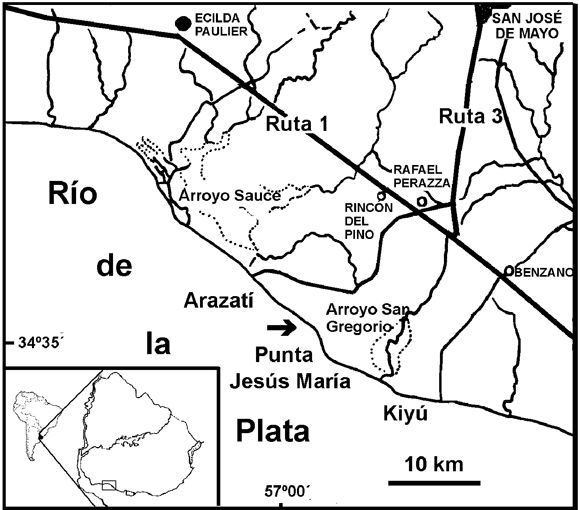
Figura 1. Mapa del lugar de procedencia de Pseudoplohophorus absolutus n. sp. La flecha señala el punto de extracción del espécimen holotipo / map of the site of Pseudoplohophorus absolutus n. sp. The arrow shows the collecting point of the holotype specimen .
La nueva especie, una de las mejor conocidas entre los mamíferos terciarios del Uruguay, permite reformular la identidad taxonómica de los géneros Pseudoplohophorus y Stromaphoropsis , en el marco de la variación observada en la subfamilia Sclerocalyptinae, y redefinir una biozona del Mioceno Superior.
Material y métodos
Para el análisis comparativo se utilizaron ejemplares de Glyptodontidae pertenecientes a las siguientes instituciones: Departamento de Paleontología de la Facultad de Ciencias, Universidad de la República, Uruguay (FC-DPV), Museo de La Plata, Argentina (MLP), Museo Argentino de Ciencias Naturales "B. Rivadavia", Argentina (MACN), Museo "Bautista Rebuffo" de la Plaza Mayor de Colonia del Sacramento, Uruguay (MPM), Museo "Florentino y Carlos Ameghino" del Instituto de Fisiografía y Geología de Rosario, Argentina (MAR), Museo Nacional de Historia Natural de Montevideo, Uruguay (MNHN), Museo Paleontológico "Alejandro Berro" de Mercedes, Uruguay (MB), Colección Aznárez, Maldonado, Uruguay (CA) y Field Museum of Natural History (FMNH). Se observaron también diferentes ejemplares de dasipódidos vivientes pertenecientes a la Colección del Departamento de Zoología de Vertebrados de la Facultad de Ciencias, Universidad de la República, Uruguay (FC-DZV) y de la Colección Comparativa del Departamento de Paleontología de la misma facultad (FC-DPV-Comp).
Los estudios comparados en cráneo fueron complementados por análisis morfométricos numéricos de agrupamiento y ordenación a partir de matrices con medidas craneanas estandarizadas (cuadro 1). Para ello se utilizó el programa NTSYSpc (Rohlf, 2000), siguiendo los postulados teóricos y metodológicos de Sneath y Sokal (1973). Mediante estas técnicas se utilizaron diferentes dimensiones craneanas tomadas entre los Sclerocalyptinae neógenos más completos comparables con el conjunto de medidas de Pseudoplohophorus absolutus n. sp. que se brindan más adelante.
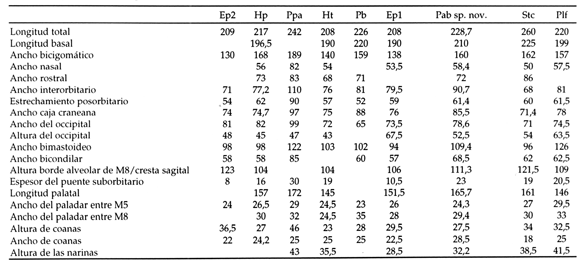
Cuadro 1. Medidas craneanas en mm de diferentes ejemplares de Sclerocalyptinae/ skull measurements in mm of some Sclerocalyptinae specimens. Ep1, holotipo de Eosclerocalyptus planus (MACN-4853); Ep2, E. planus (MLP 29-X-10-24); Hp, holotipo de Hoplophractus proximus Cabrera (MLP 31-XI-12-19); Ht, holotipo de H . tapinocephalus Cabrera (MLP 37-III-7-7); Pab, holotipo de Pseudoplohophorus absolutus n. sp. (FCDPV- 475 y 595); Pb, holotipo de P . benvenuttii (MMC-888); Plf, Plohophorus figuratus Ameghino (MACN-7844); Ppa, holotipo de Phlyctaenopyga ameghini (Ameghino) (MLP 29-X-10-1); Stc, Stromaphorus compressidens Castellanos (MLP 29-X-8-1).
La metodología y nomenclatura estratigráficas se basaron en los conceptos del Comité Argentino de Estratigrafía (1992) y Hedberg (1994).
Sistemática
Superorden Xenarthra Cope, 1889
Orden Cingulata Illiger, 1811
Familia Glyptodontidae Burmeister, 1879
Subfamilia Sclerocalyptinae Ameghino, 1891
Género Pseudoplohophorus Castellanos, 1926
Sinonimia. Plohophorus Ameghino, 1887: 17-18 [ partim ]; Stromaphoropsis Kraglievich, 1932: 281-282 [ partim, nec Stromaphoropsis scavinoi Kraglievich, 1932 ( partim ): 281-282, lám. 47]; Teisseiria Kraglievich, 1932: 306-308 [ partim, nec Teisseiria compressa , Castellanos, 1940: 185-209, figs. 53-61]; Caudaphorus Fariña, 1981: 784.
Especie tipo. Pseudoplohophorus orientalis (Ameghino, 1889: 827- 828).
Procedencia geográfica y temporal. Uruguay, Mioceno tardío.
Diagnosis enmendada. Tamaño intermedio entre Eosclerocalyptus C. Ameghino y Stromaphorus Castellanos y similar a Stromaphoropsis Kraglievich y a Plohophorus Ameghino. Cráneo robusto, con arcos cigomáticos abiertos y muy elevados, tuberculum lacrimae menos acentuado que en Plohophorus y Eosclerocaplyptus (= Hoplophractus ) y bordes alveolares muy divergentes desde el límite M4-M5 hacia adelante y atrás. Cóndilos occipitales sobresalientes. Ramas maxilares del arco cigomático muy robustas. Apófisis paraoccipitales y constricción entre la base del arco cigomático y las apófisis mastoideas muy poco notorias. Ornamentación de las placas de la coraza dorsal de relieve acentuado al igual que Stromaphoropsis , Stromaphorus y Phlyctaenopyga ; placas de las regiones lateral media y posterior subrectangulares y con dos hileras de figuritas periféricas anteriores; placas de la región lumbar subpentagonales o subhexagonales con una sola fila de figuritas periféricas, siempre en número total de 11 o más y muy pequeñas en relación a la figura central. Tubo caudal con figuritas periféricas de las placas muy estrechas transversalmente y a veces total o parcialmente sustituidas por grandes poros al igual que Coscinocercus Cabrera.
Composición. El género incluye, además de la especie tipo Pseudoplohophorus orientalis (Ameghino, 1889) y la nueva que aquí se describe, a P . coloniensis (Kraglievich, 1932) comb. n., P . berroi (Castellanos, 1940) comb. n.; P . benvenutii (Castellanos, 1954) comb. n., P . rebuffoi (Castellanos, 1954) comb. n. y P . francisi Mones, 1970. La denominación específica P . langguthi (Fariña, 1981), constituye un nomen illegitimum , por sustituir arbitrariamente a P . berroi , nombre válido que tiene prioridad ( vide Mones, 1986). Esta última especie sólo incluye un fragmento de tubo caudal (MB-2257) descrito originalmente por Kraglievich (1932) como ? Stromaphorosis scavinoi .
Comentarios. Como se expresa más adelante, las nuevas combinaciones surgen del análisis comparativo de tubos caudales en el caso de P . coloniensis , P . berroi y P . rebuffoi , y del cráneo, la coraza y el tubo caudal en el de P . benvenutii .
Pseudoplohophorus absolutus n. sp.
Figuras 3-5
Holotipo. FC-DPV-475 y 595: cráneo, mandíbula, atlas y vértebra compuesta, húmero y mano izquierdos, algunas placas de anillos caudales, fragmentos de pelvis y de vértebras sacras y caudales; mitad posterior de la coraza dorsal y fragmento anterior de ésta, tubo caudal, placas cefálicas.
Paratipo. FC-DPV-1084: hemimandíbula derecha y húmero izquierdo, placa de anillo caudal.
Derivatio nominis. Del latín absolutus : completo, por ser la especie con mayor cantidad de piezas conocidas dentro del género y uno de los mamíferos terciarios más completos del Uruguay.
Procedencia geográfica y estratigráfica. Arazatí, Departamento de San José, barrancas costeras del Río de la Plata (figura 2), en pelita gris verdosa basal, aproximadamente a 50 cm por encima de la plataforma de abrasión, Formación Camacho, Mioceno Superior (figura 3).

Figura 2. Columna estratigráfica del lugar de procedencia de Pseudoplohophorus absolutus n. sp. La flecha señala del punto de extracción del espécimen holotipo / stratigraphic section of the site of Pseudoplohophorus absolutus n. sp. The arrow shows the collecting point of the holotype specimen.

Figura 3. Pseudoplohophorus absolutus n. sp.: FCDPV-475 y 595 (Holotipo / Holotype ), A, B, C, Cráneo / Skull ; A, Vista lateral / lateral view ; B, Vista dorsal / dorsal view ; C, Vista ventral / ventral view ; D, Mandíbula en vista lateral / mandible in lateral view ; E, F, Coraza dorsal / dorsal carapace ; E, Porción anterior / anterior portion ; F, Porción media-posterior / middle-posterior portion . Escala / scale = 10 cm.
Diagnosis. Tamaño intermedio entre Eosclerocalyptus planus C. Ameghino y Stromaphorus compressidens Castellanos. Región occipital más amplia que en Pseudoplohophorus benvenutti , con los cóndilos sobresalientes y muy separados entre sí y el foramen magnum deprimido y muy ancho. Morfología de M5 a M8 (únicos dientes preservados en este ejemplar) con una mezcla de caracteres de Stromaphoropsis scavinoi (M6 y M7 con último prisma de cara posterior marcadamente convexa) y de P . benvenutii (prisma anterior de M5 a M7 con inclinación hacia adentro y adelante). Patrón general de los molariformes inferiores similar a Stromaphorus y Sclerocalyptus , con una proyección que conforma una saliente en la cara anterior y la superficie de la cara posterior plano-cóncava. Tubo caudal mediano, intermedio entre el de P . benvenutii , más pequeño, y el de P . rebuffoi , más grande. Placas del tubo con límites difusos entre sí y figura central de gran tamaño en relación a las periféricas, entre las cuales hay numerosos orificios, sobre todo en la cara dorsal. Tubo caudal con menor cantidad de orificios, figuritas periféricas más evidentes y numerosas y placas menos convexas que en Pseudoplohophorus orientalis y P . francisi .
Diagnosis. Size intermediate between Eosclerocalyptus planus C. Ameghino and Stromaphorus compressidens Castellanos. Occipital region broader than in Pseudoplohophorus benvenutti , with prominent and separated occipital condils, and very depressed and wide foramen magnum. The morphology of 5th to 8th molariforms (the only preserved in this specimen) shows a mixture of characters of Stromaphoropsis scavinoi (6th and 7th molariforms with posterior prisma having its posterior face markedly convex) and P . benvenutii (anterior prism of 5th to 7th molariforms with forward and inward inclination).
The general pattern of the lower molariforms is similar to Stromaphorus and Sclerocalyptus , with a projection which conforms a prominence in the anterior face and a flat-concave surface of posterior face. Caudal tube medium sized, intermediate between P . benvenutii (smaller), and P . rebuffoi (bigger). The plates which constitute the tube have diffuse limits between them and their central figure is markedly bigger than the peripheral ones, among which there are a lot of foramina, mainly on the dorsal part. It is distinguished from Pseudoplohophorus orientalis and P . francisi caudal tubes because it has fewer foramina, the peripheral figures are more evident and numerous, and has less convex plates.
Observaciones. El ejemplar FC-DPV-475 y 595 fue colectado en dos etapas: la primera en el año 1986, en que se recogió una hemimandíbula, húmero, parte de la mano y fragmentos de anillos caudales (FCDPV- 475). En la segunda etapa, en 1991, se exhumaron los restantes restos del ejemplar (FC-DPV-595) que constituye la mayor parte del holotipo de P . absolutus n. sp. Curiosamente, no se supo que pertenecieran al mismo ejemplar hasta constatar, luego de haber sido ingresados independientemente al catálogo, que las piezas de FC-DPV-475 articulaban perfectamente con las de FC-DPV-595, no habiendo material homólogo repetido. Las dos tandas de huesos estaban al mismo nivel en la vertical y no parece haber mediado gran erosión entre la primera (1986) y segunda (1991) etapas de extracción, a lo sumo los pocos centímetros en la horizontal que separaban la primera tanda de restos de la segunda. El espécimen FC-DPV-1084 fue colectado cerca del holotipo.
Descripción comparativa
El cráneo (figuras 3.A, B, C; cuadro 1) es robusto y se asemeja globalmente al de Plohophorus figuratus Ameghino; tiene los arcos cigomáticos abiertos y muy elevados en norma lateral, a escasos milímetros por debajo del contorno superior craneano. Como sucede en Pseudoplohophorus benvenutii , posee un gran espesor de la rama anterior del arco cigomático a nivel de los forámenes suborbitarios. El rostro es ancho y corto y la región occipital amplia y deprimida. En norma dorsal, más de la mitad de su longitud corresponde a la superficie que se desarrolla a partir de las apófisis post-orbitarias hacia atrás. Presenta la caja craneana amplia y de superficie dorsal rugosa. Las crestas lambdoideas conforman un ángulo marcado con la sagital. La región frontal es abultada, con el tuberculum lacrimae menos desarrollado que en Plohophorus y Eosclerocalyptus (= Hoplophractus ). La constricción posterior al arco cigomático y anterior a la apófisis mastoidea es muy poco marcada al igual que en P . benvenutii . Esto lo diferencia mucho del cráneo de los ejemplares de Sclerocalyptus y bastante del de los de Eosclerocalyptus (= Hoplophractus ). En estos géneros, la región paraoccipital es también más amplia y consecuentemente la apófisis mastoidea más sobresaliente. En estos caracteres Pseudoplohophorus se asemeja más a Stromaphorus y Phlyctaenopyga . El foramen mastoideo está ubicado posteriormente a la apófisis mastoidea, mientras que en el ejemplar FMNHP14501 de Eosclerocapyptus es interno a ésta. Tanto en norma dorsal como ventral muestra una gran similitud con Pseudoplohophorus benvenutii , a pesar de que la región occipital de la nueva especie es más amplia. En norma posterior se observan las coanas de contorno subcuadrado y la amplitud y depresión de la región occipital, en cuyo llano hay una cresta central delimitada lateralmente por dos amplias depresiones para la inserción de la musculatura recta del cuello.
De la dentición superior (figuras 3.C y 4.A, cuadro 2) se preservan las series M5 y M8 derechas y M6-8 izquierda y los alvéolos de los restantes dientes. Ambas series se acercan entre sí a nivel del límite M4-M5 y divergen a partir de ese punto, hacia adelante y hacia atrás, al igual que en P . benvenutii . El M1 es subcircular y pequeño; el M2 más alargado, parece mostrar ya indicios de lobulación; el M3, es claramente trilobulado, los M4 a M8 presentan una conformación y tamaño similares entre sí. La configuración dentaria general y el tamaño coinciden con la descripción de Castellanos (1954). La morfología de los M5 a M8 muestra una mezcla de caracteres de Stromaphoropsis scavinoi Kraglievich (M6 y M7 con el último prisma de la cara posterior marcadamente convexa) y de P . benvenutii (prisma anterior de los M5 a M7 con inclinación hacia adentro y adelante y M8 cóncavo en su cara anterior). Este último carácter es propio de muchos de los representantes de la subfamilia como Sclerocalyptus y Panochthus . El último prisma del M8 posee un surco posteroexterno en su cara posterior, que es principalmente convexa. El mismo patrón se observa en Sclerocalyptus y algunos ejemplares de Glyptodon y Glyptotherium . La dentición superior se asemeja también a la de Eosclerocalyptus (= Hoplophractus ) y difiere notoriamente de la de Phlyctaenopyga (M5 a M8). La dentición superior de Stromaphorus es desconocida; sólo es posible inferirla a partir de los alvéolos, por lo que no se pudieron efectuar comparaciones adecuadas. La mandíbula (figuras 3.D y 4.B, cuadro 2), de aspecto grácil por su escaso espesor, presenta una longitud total de aproximadamente 20 cm y una altura de 16 cm. La rama montante es amplia y su borde anterior asciende en ángulo recto y luego se inclina hacia adelante. La apófisis coronoidea se eleva algo por encima del cóndilo, el que es amplio lateralmente y muy estrecho anteroposteriormente. El margen incisivo está separado del m1 aproximadamente por 2,5 cm; la sínfisis tiene una longitud de unos 8 cm y su límite posterior está a nivel del borde posterior del m3. Las ramas horizontales son subparalelas y de mediana altura, presentando un margen alveolar sub-horizontal y su borde ventral muy convexo. El límite posterior del m8 se localiza al mismo nivel que la escotadura que separa la apófisis coronoides del cóndilo mandibular.
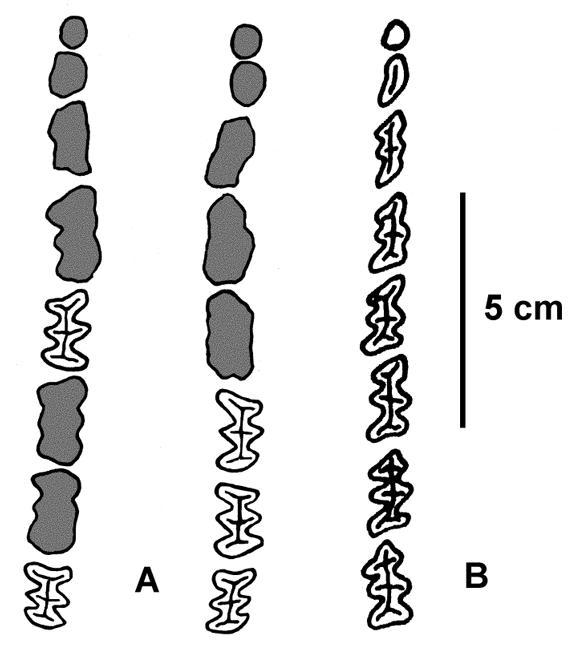
Figura 4. Pseudoplohophorus absolutus n. sp.: FCDPV-475 y 595 (Holotipo / Holotype ), A, series dentarias y alveolares superiores / upper dentary and alveolar series ; B, serie dentaria inferior izquierda / left lower dentary series .

Cuadro 2. Medidas de la mandíbula y dentición (mm) de Pseudoplohophorus absolutus n. sp. FCDPV-475 y 495 / Mandible and dentition measurements (m mm) of Pseudoplohophorus absolutus n. sp. FCDPV-475 y 495.
La dentición inferior (figura 4.B, cuadro 2) presenta un patrón similar a la dentición superior con el m1 de sección subcircular a partir del cual, los elementos de la serie se alargan progresivamente y tienden a hacerse más claramente trilobulados, adquiriendo un tamaño y configuración uniformes a partir del m5. Los m3 - m8 se caracterizan por poseer una proyección del borde anterior que contrasta con la forma trunca del posterior. En el holotipo la dentición inferior está casi completa a excepción del m6 y lóbulo posterior del m5 derechos.
El húmero, de aproximadamente 20 cm de longitud, es algo más corto y grácil que el húmero de P . rebuffoi (MPM-880). Las crestas son más acentuadas. La mano izquierda está casi completa, presentando pisiforme, cuneiforme, lunar, escafoide, trapecio, magno y unciforme. Posee los metacarpianos III, IV y V, la 2a. falange ungueal del dedo II; la 1a. y 2a. falanges del dedo IV, y la falange ungueal del dedo V presenta cinco sesamoideos.
El atlas se asemeja al de Eosclerocalyptus planus , aunque es menos robusto y, al igual que el de Plohophorus figuratus , tiene las facetas cefálicas más desarrolladas (altura: 53,5 mm, diámetro en las facetas cefálicas: 67 mm, diámetro transverso del agujero neural: 31 mm, diámetro vertical del mismo en su parte anterior: 32 mm). El axis tiene una apófisis odontoides sobresaliente y bastante aguda, se asemeja también a la de E . planus aunque es relativamente más larga (longitud: 69 mm; diámetro anteroposterior de la apófisis espinosa: 33 mm; ancho a nivel de la articulación anterior: 53,5 mm; ancho anterior del foramen raquídeo: 23,5 mm; altura anterior del foramen raquídeo: 22,5 mm; ancho posterior del foramen raquídeo: 30 mm; altura posterior del foramen raquídeo: 19 mm). Las vértebras sacras son amplias transversalmente y la pelvis es amplia y constituida por huesos de escaso espesor.
El escudo cefálico está representado por la placa central posterior, 3 placas laterales derechas, la más grande de las cuales articula con la primera, y una placa probablemente anterior. Su configuración difiere notoriamente de la de Phlyctaenopyga ameghini que prácticamente carece de ornamentación y de Plohophorus figuratus que es muy ornamentado. Su patrón se asemeja al de Eosclerocalyptus planus . La placa central posterior es similar en tamaño a la homóloga de P . benvenutii , aunque a diferencia de ésta, posee su mayor diámetro transverso anterior, mientras que en la citada especie los diámetros anterior y posterior son idénticos. Se diferencia de P . rebuffoi porque esta especie posee una clara asimetría en la configuración del escudo cefálico, sobre todo la placa central posterior. Se diferencia de P . benvenutii y se asemeja a P . rebuffoi por poseer plaquitas externas laterales articulando con las mayores laterales a la placa central posterior (Castellanos, 1954). Todas las placas se asemejan a las de E . planus en el patrón general de poseer una superficie porosa rodeando los bordes y zonas circulares centrales rugosas. Estas rugosidades se ven atenuadas a nivel de las placas más pequeñas cuya configuración se asemeja a la de " Hoplophractus " tapinocephalus Cabrera.
Para la coraza (figuras 3.E, F), a pesar de no estar completa, puede estimarse una longitud total de entre 95 cm y 1 m, tamaño intermedio entre Stromaphorus compressidens y Eosclerocalyptus planus . Las placas que la componen se agrandan desde adelante, donde las más pequeñas miden 15,5 mm de diámetro, hacia la zona lumbar, donde adquieren su mayor diámetro (43,5 mm). Desde allí, las últimas filas se van reduciendo, hasta que en la última llegan a 27,5 mm. Las placas dorsales aledañas al plano sagital son subhexagonales, desde las filas más anteriores hasta la zona media-lumbar; en las últimas filas se hacen subcuadradas. La zona anterior lateral posee placas pequeñas y sub-romboidales y la zona pleural placas sub-rectangulares de diámetro mayor promedio de 36 mm. En cada placa la figura central ocupa la mayor parte de la superficie y las figuritas periféricas son pequeñas. En las placas ubicadas sobre el plano sagital la figura central es subcircular y hacia los lados (zona pleural) se va haciendo más elíptica a diámetro mayor anteroposterior, acompañando la forma rectangular de la placa. El máximo número de figuritas periféricas se da en las placas de la zona lumbar (entre 13 y 15) y el mínimo número (entre 3 y 4) de éstas en las primeras filas anteriores, sobre la zona lateral. En la zona anterior dorsal hay entre 12 y 13 de estas figuritas y en la zona media pleural entre 12 y 14. En la última fila están mal definidas, pudiéndose calcular alrededor de 4 sólo sobre el borde anterior y agujeros de regular tamaño entre éstas (o sustituyéndolas). En la zona pleural, las figuritas periféricas anteriores son mayores que las posteriores y las superiores e inferiores muy pequeñas y alargadas. En la parte anterior de estas placas puede haber dos filas de periféricas (figura 3.F). En el conjunto de la coraza pueden distinguirse los siguientes patrones de cambio: 1) un aumento del tamaño de la placa desde adelante y desde atrás hacia las zonas media lumbar y pleural media posterior; 2) la superficie de las figuras y figuritas se hace progresivamente más convexa hacia atrás ; 3) siempre existe una sola fila de figuritas periféricas a excepción de las placas posteriores de la zona pleural, en que puede haber dos y 4) el espesor de la coraza aumenta desde adelante hacia atrás y de los costados hacia la zona media.
Los patrones descritos coinciden con los establecidos previamente para la coraza en las especies definidas como Pseudoplohophorus benvenutti y P . rebuffoi y algunos permiten distinguirlos de otros representantes de la subfamilia: de Eosclerocalyptus (= Hoplophractus ) por el mayor número y menor tamaño relativo de las figuritas periféricas; de Stromaphoropsis scavinoi , Stromaphorus , Phlyctaenopyga y Plohophorus , por poseer una sola fila (parcialmente dos sólo en algunas placas de la zona pleural) de figuritas periféricas.
El tubo caudal (figura 5) posee una longitud total de 34 cm considerando el último anillo soldado, un diámetro transverso a nivel de los límites anteriores de las placas apicales de 64 mm y uno dorso-ventral, al mismo nivel, de 54,5 mm. Posee un máximo diámetro transverso a nivel del último anillo soldado de 89 mm y uno máximo trasverso, al mismo nivel de 102 mm. El tubo parece estar algo comprimido lateralmente a este nivel. Las placas que constituyen el tubo tienen límites difusos entre sí y sus figuras centrales son de gran tamaño en relación al de las periféricas, a veces inexistentes o sustituidas por orificios. Como máximo existe una sola hilera de periféricas entre dos figuras centrales de placas contiguas. Desde atrás hacia adelante se distinguen dos grandes figuras terminales elípticas separadas en la parte apical dorsal por una fila de figuritas periféricas y en la ventral por un surco con pequeños orificios. En vista lateral se observan 4 placas hacia adelante. La más distal dividida en dos, una arriba y otra abajo, las cuales están tan separadas en el lado izquierdo que permiten que la penúltima placa lateral contacte con la placa apical del mismo lado. Esto hace que la longitud ocupada por las placas laterales sea mucho menor que en otras especies del género, ocupando menos de la mitad de la longitud total del tubo. En P . rebuffoi y P . benvenutii esta dimensión es superior a la mencionada longitud total (Castellanos, 1954). Así es que por delante de la primera placa lateral, hay 5 filas de placas por delante del último anillo soldado, mientras que en P . benvenutii hay sólo dos y en P . rebuffoi tres. Otra posibilidad es considerar a la 1a. lateral como doble (figura 5.C), en cuyo caso las placas laterales ocuparían aproximadamente la mitad de la longitud del tubo y las filas de placas ubicadas entre éstas y el último anillo soldado serían 4. Considerando este último esquema, las placas que quedan entre las laterales serían dorsalmente 24 y ventralmente 25. La primera fila anterior a las laterales tiene 12 placas, la 2da.,13, la 3ra., 14 y la 4ta., 13. El último anillo, soldado al tubo, tiene 15 placas en cada fila. A lo largo de la franja media dorsal del tubo se observan abundantes orificios circulares entre las figuritas periféricas. También se pueden observar estos orificios esporádicamente a los lados y en la zona ventral. El diámetro transverso de las figuritas periféricas se hace cada vez más reducido desde el dorso hacia el vientre del tubo, donde los surcos ubicados entre las figuras centrales son muy reducidos, sobre todo hacia el ápex. El tubo caudal tiene similitudes con el de Pseudoplohophorus orientalis y P . francisi por presentar forámenes de gran tamaño, sobre todo en la parte dorsal y la zona ocupada por las placas laterales reducidas (aproximadamente la mitad de la longitud del tubo), pero se diferencia de los de estas especies por poseer menor cantidad de orificios, figuritas periféricas más evidentes y placas menos convexas (Castellanos, 1926; Mones, 1970).

Figura 5. Pseudoplohophorus absolutus n. sp.: FCDPV-475 y 595 (Holotipo / Holotype ), Tubo caudal / caudal tube , A, vista dorsal / dorsal view ; B, vista ventral / ventral view ; C, vista lateral / lateral view .
Estratigrafía y edad
El material descrito proviene de la Formación Camacho (Goso y Bossi, 1966). De acuerdo a su caracterización litoestratigráfica original, ésta coincide en parte con lo que Francis y Mones (1965) denominan Formación Kiyú.
De la revisión de ambas unidades (Perea y Martínez, 2004), lo que se definiera originalmente como "Areniscas de Camacho" (Caorsi y Goñi, 1958), en parte equivalentes a la Formación Camacho (Goso y Bossi, 1966), definida para el Departamento de Colonia, incluye dos miembros, de los cuales el inferior (Miembro San Pedro sensu Martínez, 1994 y Perea y Martínez, 2004) caracterizado por pelitas, se correlaciona litoestratigráficamente con la Formación Kiyú (Francis y Mones, 1965), definida en el limítrofe Departamento de San José. Es así que la Formación Camacho estaría compuesta por el miembro descrito y otro miembro superior (Miembro Cerro Bautista sensu Martínez, 1994 y Perea y Martínez, 2004), más arenoso, cuyas mejores exposiciones se dan en el Cerro Bautista cerca de la ciudad de Carmelo. Atendiendo a esta nueva división, dicha formación estaría abarcando entonces facies pelíticas basales y facies arenosas cuspidales, ambas dispuestas en gran parte de la franja costera de los Departamentos de San José y Colonia (figuras 1 y 2), pero cuyo contacto se observa en las barrancas de San Juan en el Departamento de Colonia (Martínez, 1994; Perea y Martínez, 2004). Esta división fue ya vislumbrada por Serra (1943).
Es de destacar que en el Departamento de San José la Formación Camacho sólo está representada por las facies pelíticas (figura 2) sobre las cuales sobreyace la Formación San José, definida por Francis y Mones (1965) e indistinguible de la Formación Raigón (Goso y Bossi, 1966), de probable edad Plioceno-Pleistoceno inferior y medio (Tambussi et al. , 1999; McDonald y Perea, 2002).
Las facies basales de la Formación Camacho incluyen una biozona cuyo representante mejor conocido es Pseudoplohophorus . Esta biozona, definida como "Biozona de " Stromaphoropsis " (Perea, 1993; Perea et al. , 1994) y redefinida y revisada en el presente artículo, se extiende a lo largo de la franja costera del río de la Plata en parte de los departamentos de San José y Colonia. La abundante mastofauna integrada a esta biozona tiene afinidades "huayqueriense-mesopotamienses" según Perea et al. (1994, 1996) y Perea y Scillato-Yané (1995). La "Edad-mamífero" Huayqueriense ( sensu Pascual et al. ,1966), asignada al Mioceno tardío, está representada clásicamente en sedimentos de las provincias de Buenos Aires, La Pampa, Catamarca y Mendoza en la República Argentina (Pascual et al. , 1966; Zetti, 1972; Marshall et al. , 1983; Marshall y Cifelli, 1991).
Según Cione et al. (2000) el término "Piso Mesopotamiense" o "Mesopotamiense" es considerado inválido y es equivalente al "Conglomerado osífero" de la Formación Ituzaingó, cuya fauna terrestre sugiere una edad Huayqueriense. La base de esta edad se ubica según Flynn y Swisher (1995) en los 9 Ma (Tortoniano).
Discusión
Evolución, sistemática y paleobiogeografía
Los Glyptodontoidea muy probablemente surgieron a partir de Dasypodidae (Hoffstetter, 1958; Paula Couto, 1979) en el Paleoceno-Oligoceno. Los restos más antiguos indudablemente pertenecientes a esta superfamilia datan del Mustersense y Deseadense argentinos (Ameghino, 1897, 1902; Scillato-Yané, 1977). Los cambios que parecen haber mediado entre gliptodontes y sus antecesores dasipodoideos parecen coincidir con patrones macroevolutivos de heterocronía y heterotopía (Alberch et al. , 1979; McKinney y Gittleman, 1995; Zelditch y Fink, 1996). En el marco de un patrón de hipermorfosis (Alberch et al. , 1979; Gould, 1980; McNamara, 1988) el aumento de tamaño, junto con la gran osificación y la particular configuración del aparato masticatorio con un desplazamiento de la dentición hacia atrás y una concomitante elevación del cráneo y mandíbula, son caracteres que en los gliptodontes pueden haber surgido por aceleración de algunas etapas del desarrollo de sus probables ancestros.
Los caracteres craneanos y dentarios son de gran valor para la sistemática de mamíferos como lo demuestran los numerosos trabajos de osteología comparada en grupos actuales y fósiles. En el caso particular de los Cingulata, se suman también los caracteres del exoesqueleto, los cuales tienen un elevado valor diagnóstico (Burmeister, 1879; Ameghino, 1889, Hoffsteter, 1958; Paula Couto, 1979) . Las principales dificultades taxonómicas surgen cuando el grado de variabilidad de los caracteres craneanos no se ve acompañado por el correspondiente de los caracteres exoesqueléticos. Es así que el cambio evolutivo de uno no implica necesariamente la variación del otro, existiendo distintas tasas de diferenciación morfológica o tendencias según la región del esqueleto que se trate. En el caso particular de los Sclerocalyptinae resulta muy difícil proponer hipótesis sistemáticas robustas sin tener material del cráneo asociado al esqueleto externo.
Los Sclerocalyptinae neógenos no se encuentran al margen de esta condición que afecta a los restantes gliptodontes. El agrupamiento de ciertos géneros definidos para la Argentina a base de material craneano asociado a coraza y tubo caudal, surge en forma relativamente clara. Teniendo en cuenta las pocas diferencias observadas se propone la sinonimia de Hoplophractus Cabrera y Eosclerocalyptus C. Ameghino. Las diferencias que existen a nivel craneano (sobre todo altura de las coanas) y a nivel de la coraza (presencia/ ausencia de concavidad de las figuras centrales y figuritas periféricas de las placas) se condicen mejor con una diferenciación de carácter específico. Cabrera (1944), basa parte de su distinción genérica en la altura y estrechez craneana de Eosclerocalyptus respecto de Hoplophractus , sin embargo reconoce que los cráneos del primero que observó para comparar, se encontraban comprimidos (deformados) por la presión de los sedimentos, hecho que fue confirmado por la observación directa del material por parte del autor del presente artículo. Esta homogeneidad entre ambos géneros, que surge principalmente del análisis morfológico cualitativo, se ve confirmada por análisis fenéticos de agrupamiento y ordenación basados en dimensiones craneanas (figuras 6 y 7). " Hoplophractus " proximus Cabrera y Eosclerocalyptus poseen una figura central rodeada de un promedio de 9 periféricas en la franja central de la coraza (8 a 10 en el primero y 7 a 10 en el segundo). " Hoplophractus " tapinocephalus Cabrera presenta de 8 a 11 figuritas en cada placa de la misma región. En todos ellos el relieve de las figuras es poco acentuado, la figura central puede ser plana o cóncava y las figuritas periféricas son proporcionalmente más grandes que las homólogas de Pseudoplohophorus , Stromaphoropsis , Phlyctaenopyga y Stromaphorus , en relación a la central.
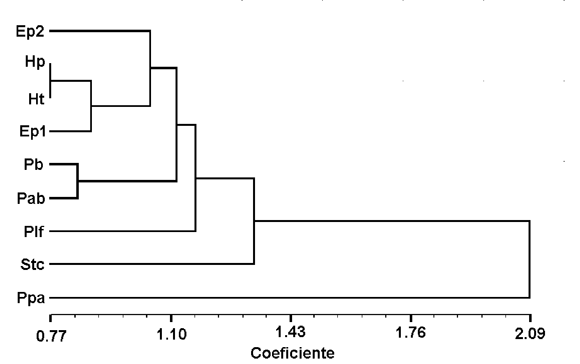
Figura 6. Fenograma basado en una matriz de medidas craneanas (ver Cuadro 1) estandarizadas de ejemplares de Sclerocalyptinae utilizando el método de agrupamiento UPGMA (Rohlf, 2000) / phenogram based on standardized cranial measurements (see Table 1) of specimens of Sclerocalyptinae using the UPGMA clustering method (Rohlf, 2000) . Ep1: Eosclerocalyptus planus (holotipo / holoype , MACN-4853,); Ep2: E . planus (MLP 29-X-10-24); Hp: Hoplophractus proximus (holotipo / holotype , MLP 31-XI-12-19); Ht: H . tapinocephalus (holotipo / holotype , MLP 37-III-7-7); Pab: Pseudoplohophorus absolutus n. sp. (holotipo / holotype , FCDPV-475 y 595); Pb: P . benvenuttii (holotipo / holotype , MMC-888); Plf: Plohophorus figuratus (MACN-7844); Ppa: Phlyctaenopyga ameghini (holotipo / holotype , MLP 29-X-10-1); Stc: Stromaphorus compressidens (MLP 29-X-8-1).
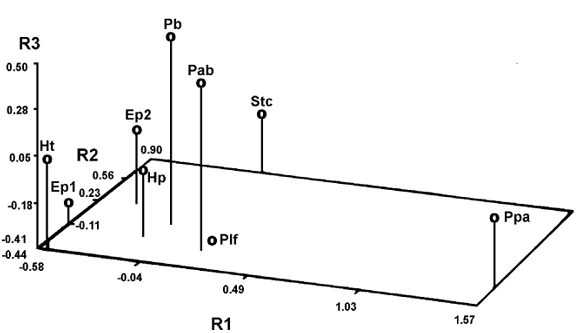
Figura 7. Gráfico mostrando los resultados del análisis de componentes principales basado en una matriz de medidas craneanas (ver Cuadro 1) estandarizadas de algunos ejemplares de Sclerocalyptinae / graphic showing the results of the principal component analysis based on standardized cranial measurements of some Sclerocalyptinae specimens. Ep1: Eosclerocalyptus planus (holotipo / holoype , MACN-4853,); Ep2: E . planus (MLP 29-X-10-24); Hp: Hoplophractus proximus (holotipo / holotype , MLP 31-XI-12-19); Ht: H. tapinocephalus (holotipo / holotype , MLP 37-III-7-7); Pab: Pseudoplohophorus absolutus n. sp. (holotipo / holotype , FCDPV- 475 y 595); Pb: P . benvenuttii (holotipo / holotype , MMC-888); Plf: Plohophorus figuratus (MACN-7844); Ppa: Phlyctaenopyga ameghini (holotipo / holotype , MLP 29-X-10-1); Stc: Stromaphorus compressidens (MLP 29-X-8-1).
El único material asignado a " Hoplophractus " ( vide Marshall y Patterson, 1981) que presenta cráneo asociado a coraza y tubo caudal es el ejemplar FMNH-P14501. Según los citados autores, el tubo caudal de este ejemplar difiere del MLP-29-X-10-32, asignado a H . proximus con dudas por Cabrera (1944), pero no describen las diferencias. La observación por nuestra parte de fotografías de dicho ejemplar permitió identificar muchas similitudes con Eosclerocalyptus .
Las similitudes a nivel craneano y de ornamentación del exoesqueleto observadas entre los ejemplares tipo de Pseudoplohophorus benvenutti y de P . absolutus n. sp., con caracteres exclusivos y diagnósticos, permiten confirmar la robustez taxonómica de este género. Esto se ve apoyado por el análisis numérico efectuado, en donde tanto en las técnicas de agrupamiento, como de ordenación, estos dos ejemplares están más cercanos entre sí, que de cualquier otro del conjunto analizado (figuras 6 y 7).
Los tubos caudales de ambas especies muestran similitudes entre sí y con el neotipo de Pseudoplohophorus orientalis . A su vez, los especímenes decritos por Kraglievich (1932), como ? " Stromaphoropsis " scavinoi (MB-2257) y " Stromaphoropsis " coloniensis y por Castellanos (1954) como S . rebuffoi , poseen caracteres que se condicen con el patrón de variabilidad observado en los tubos caudales aquí considerados como Pseudoplohophorus .
Stromaphoropsis se considera un género válido, con la única especie S . scavinoi Kraglievich, representada por el ejemplar MB-2189, caracterizado por poseer las placas de la región media lumbar con dos filas de figuritas periféricas.
De acuerdo con Cabrera (1944) la diferencia entre Stromaphorus y Phlyctaenopyga , aparte del tamaño, es que desde la octava fila a partir del borde posterior, las figuras centrales son abultadas en el último, mientras que en el primero, son algo menos abultadas y se dan en las 5 ó 6 últimas filas. En Phlyctaenopyga , en la zona media lumbar existen entre 4 y 7 figuritas periféricas entre las figuras centrales, mientras que en la misma región de Stromaphorus hay de 3 a 5 figuritas entre las figuras centrales.
En Plohophorus la coraza tiene una figura pequeña central plana o levemente excavada alrededor de la cual hay 2 ó 3 hileras de figuritas periféricas pequeñas, similar a Nopachthus y a algunas regiones de la coraza de Panochthus . Stromaphoropsis presenta un número de figuritas periféricas intermedio (11 a 18) entre Eosclerocalyptus (= Hoplophractus ) y Pseudophlohophorus , con un número menor por un lado, y Stromaphorus , Phlyctaenopyga y Plohophorus , con mayor número de figuritas periféricas por otro.
La relación del diámetro transverso del promedio de las figuritas periféricas con el diámetro de la figura central, es mayor en Eosclerocalyptus (= Hoplophractus ) y Sclerocalyptus que en Stromaphorus , Phlyctaenopyga , Plohophorus , Pseudoplohophorus y Stromaphoropsis . Esta relación se correlaciona positivamente con la análoga observada para las figuras de tubos caudales en los casos comparables.
La configuración muy variable y frecuentes asimetrías observadas en el escudo cefálico y tubo caudal de Cingulata puede deberse a diferencias individuales. La observación de ejemplares de dasipódidos actuales dio como resultado diferentes grados de asimetrías en todos los ejemplares, sobre todo a nivel de estuche caudal y escudo cefálico. En varios casos hay multiplicación (o división) de placas. Las asimetrías son más perceptibles cuando están cercanas al plano sagital, pero se han observado algunas distales en el caparazón dorsal, incluso en las bandas móviles (6 de un lado y 7 del otro en algunos especímenes de Dasypus ). Los anillos caudales varían en número, sobre todo en Dasypus novemcinctus Linnaeus y Euphractus sexcinctus (Linnaeus). También el número de bandas móviles es variable (sobre todo en Dasypus ). En el único ejemplar de Zaedyus pichi (Desmarest) observado (97-DPFC.COMP), se constataron asimetrías en los escudos cefálico y pélvico. También están citadas este tipo de asimetrías en Chlamyphorus (Yepes, 1931). Mc Bee y Baker (1982) citan un patrón asimétrico de placas óseas dérmicas caracterizando a Dasypodinae.
En Glyptodontidae las asimetrías parecen ser bastante comunes, sobre todo a nivel del tubo caudal. Cabrera (1939, 1944) describe diferentes grados de asimetría en tubos caudales de los géneros de Sclerocalyptinae por él estudiados. En ningún caso considera las asimetrías como carácter taxonómico, sino como producto de la variación individual. Gillette y Ray (1981) mencionan marcada asimetría en el penúltimo anillo caudal de Glyptotherium arizonae Gidley, aunque sólo consideran para la distinción específica a base de caracteres caudales el tamaño y desarrollo de tubérculos. El número de anillos caudales es variable en Glyptotherium . Gillette y Ray (1981) no identifican asimetrías para la diagnosis de las especies de este género ni para la distinción genérica. Kraglievich (1932) no menciona asimetrías de ningún tipo en el momento de diferenciar tubos caudales de diferentes géneros y especies. Fariña (1981) distingue géneros dentro de la tribu Plohophorini a base de la asimetría del tubo caudal. Mones (1987: 501) considera "asimetría más o menos marcada" del tubo caudal para la diagnosis del género Uruguayurus . Castellanos (1926) no incluye asimetrías en la distinción de géneros y especies, a pesar de que la menciona en la descripción del tubo de Urotherium simplex Castellanos. Pascual et al. (1966) sólo mencionan para la diagnosis genérica, asimetría apical de Urotherium simplex . Paula Couto (1979) minimiza el valor sistemático de los tubos caudales en Glyptodontidae.
El carácter intergenéricamente menos variable en tubos caudales de Sclerocalyptinae es la relación entre el diámetro transverso de las figuritas periféricas de las placas respecto del tamaño de la figura central y esto aparenta estar correlacionado con el mismo carácter en la coraza. Géneros con figuritas periféricas de la coraza pequeñas como Stromaphorus , Phlyctaenopyga , Stromaphoropsis y Pseudoplohophorus , poseen tubos caudales con estas figuritas muy estrechas respecto de la figura central. Géneros como Eosclerocalyptus (= Hoplophractus ), Sclerocalyptus y Plohophorus , poseen figuritas periféricas de la coraza más amplias respecto de la figura central y las figuritas del tubo anchas. Como en los tubos caudales no existen figuritas periféricas supernumerarias, resulta especulativo entonces proponer nuevos géneros y especies a base de este tipo de restos aislados.
Bioestratigrafía y paleoambientes
Los depósitos neógenos aflorantes en la costa del Departamento de San José, en particular en las barrancas de Kiyú y Arazatí, han aportado gran cantidad de restos de mamíferos que reflejan muy diversas edades. El primero en efectuar un análisis pormenorizado de estos restos, en particular de aquéllos provenientes de la localidad de Kiyú (Barrancas de San Gregorio), fue Kraglievich (1932), quien identificó esta unidad con el "Piso Mesopotamiense". Muchos de estos restos fueron hallados removidos en la plataforma de abrasión de dichas playas y su procedencia estratigráfica es imprecisa por el hecho de que en esas localidades se superponen por lo menos dos unidades litoestratigráficas. En la base se encuentra la Formación Camacho. De los taxones de mamíferos registrados en esta unidad, Pseudoplohophorus y Lagostomopsis son abundantes y comunes en las áreas más fosilíferas de las localidades de Arazatí y Kiyú. El primer género muestra mayor abundancia de ejemplares y mejor calidad en su registro. Ha sido descrito cráneo, coraza y tubo caudal asociados, procedentes de la localidad de Kiyú (Castellanos, 1954) y aquí se describen elementos homólogos a éstos también asociados, provenientes de Arazatí, además existen otros restos asignables a este taxón procedentes de ambas localidades. Sin duda, se trata del mamífero más representativo de la Formación Camacho, caracterizando una biozona a través de gran parte de esa unidad.
Biozona de Pseudoplohophorus absolutus - Kiyutherium orientalis
Definición. Zona de Asociación caracterizada por la presencia de dichos taxones y Proeuphractus limpidus Ameghino, Kraglievichia paranensis (Ameghino), Stromaphoropsis scavinoi Kraglievich, Pseudoplohophorus benvenutii (Castellanos), P . rebuffoi (Castellanos), Berthawyleria gracilis Castellanos, Prodoedicurus wyleri Castellanos, Pronothrotherium mirabilis (Kraglievich), Pliomorphus ameghinoi Kraglievich, Ranculcus , Anchimys , Isostilomys intermedius Mones y Castiglioni, Lagostomopsis Kraglievich, Dinotoxodon paranensis (d'Orbigny y Laurillard), Toxodontherium , Xotodon , Scalabrinitherium y Saurocetes argentinus Ameghino.
Estratotipo y perfil tipo. Miembro San Pedro de la Fm. Camacho, a 50 cm de la plataforma de abrasión, en la localidad de Arazatí, Departamento de San José.
Edad. Mioceno tardío, Huayqueriense (Tortoniano). La cronozona de Pseudoplohophorus absolutus - Kiyutherium orientalis se extendería a la totalidad del nivel de pelitas inferiores de la Fm. Camacho ( sensu Goso y Bossi, 1966), correspondientes a su Miembro San Pedro ( sensu Perea y Martínez, 2004) y a la Fm Kiyú ( sensu Francis y Mones, 1965), aflorantes en las costas de los Departamentos de Colonia y San José.
Observaciones. Pronothrotherium mirabilis , Scalabrinitherium y Toxodontherium también se registran en en el "Conglomerado osífero" de la Fm. Ituzaingó y en la Fm. Andalhualá (Argentina) (Perea et al. , 1994). Proeuphractus limpidus , Kraglievichia paranensis , Pliomorphus ameghinoi , Dinotoxodon paranensis y Saurocetes argentinus en la Fm. Ituzaingó, y Ranculcus en esta última y dudosamente en sedimentos asignados al "Huayqueriense-Montehermosense" de Brasil (Rancy et al. 1989; Perea y Scillato-Yané, 1990). Por lo tanto se sugiere la correlación temporal con estas unidades.
Pseudoplohophorus , como ya se observó, sinónimo en parte de Stromaphoropsis y de Teisseiria , se reconoce hasta el momento como endémico del Uruguay, aunque muestra afinidades con el material muy fragmentario que tipifica a Coscinocercus de la Formación Epecuén (de "Edad-mamífero" Huayqueriense) de Argentina (Cabrera, 1939; Zetti, 1972), el cual según Castellanos (1939) es sinónimo de Teisseiria . Entre estas similitudes este autor destaca la abundante presencia de poros y las figuritas periféricas estrechas. Lamentablemente, la información que se tiene de éstos es hasta el momento muy escasa como para adelantar hipótesis de sinonimia con los restos de Uruguay. Entre los mamíferos registrados en esta biozona, Kiyutherium orientalis Francis y Mones constituye un argumento para proponer su correspondencia con el Huayqueriense (Mioceno tardío). Fuera del Uruguay este taxón o una forma muy afín ha sido descrito para el Huayqueriense de Argentina (Pascual y Bondesio, 1982; 1985; Dozo et al. , 2002) y la "Fauna Local" Río Acre, Huayqueriense de Perú y Brasil (Frailey, 1986).
De acuerdo a la información general recabada en trabajos de síntesis sobre fauna terciaria (Pascual et al. , 1996; Marshall et al. , 1983; Bondesio, 1986; Flynn y Swisher, 1995; Cione et al. ; 2000), mas los datos que surgen de bibliografía más específica, comentada para cada taxón particular, se desprende una afinidad Huayqueriense-"Mesopotamiense" de los mamíferos registrados en la Formación Camacho (Perea et al. , 1994, 1996). Los datos son aún insuficientes y no se han hallado fósiles guía del Huayqueriense en esta formación, a no ser por la ya comentada probable relación de Coscinocercus con Pseudoplohophorus (Cabrera, 1939; Castellanos, 1939). Lagostomopsis , por su amplia distribución estratigráfica, no aporta datos importantes a los efectos de la cronoestratigrafía de la Formación Camacho. Lagostomopsis es un taxón representado en el Montehermosense y Chapadmalalense de Argentina y L . petrichodactyla (Rovereto) corresponde a los depósitos del "Araucanense s . str. " (Pascual et al. , 1966; Marshall et al. , 1983). Muchos de los taxones son exclusivos o casi exclusivos de la Formación Camacho y del "Mesopotamiense" argentino, unidad informal que se corresponde con la base de la Fm. Ituzaingó según Aceñolaza (1976) y Cione et al. (2000). Entre ellos se destacan Pliomorphus ameghinoi , Proeuphractus limpidus , Kraglievichia paranensis , Anchimys cf. marshi (Ameghino), Dinotoxodon paranensis y Saurocetes argentinus ( vide Mones y Castiglioni, 1979; Perea et al. 1994; Perea y Scillato-Yané, 1995). Otros taxones registrados en la Formación Camacho son Pronothrotherium mirabilis , Scalabrinitherium y ? Toxodontherium ( vide Mones, 1976; Perea, 1988; Verde y Perea, 1992). Estos están citados además de la Formación Ituzaingó, para depósitos "Araucanenses" equivalentes a las Formaciones Andalhualá (Huayqueriense) y Chiquimil (?Chasiquense) (Kraglievich, 1931; Riggs y Patterson, 1939; Simpson, 1974; Marshall y Patterson, 1981; Butler et al. , 1984; Bondesio, 1986; Verde y Perea, 1992; Perea et al. , 1994). Ranculcus , descrito para la Formación Camacho por Perea y Scillato-Yané (1990), se halló además en la Formación Ituzaingó y con dudas en depósitos miopliocénicos de Acre en Brasil (Rancy et al., 1989). Los demás taxones de mamíferos claramente procedentes de la Formación Camacho, son exclusivos del Uruguay. Tales los casos de Pseudoplohophorus , Stromaphoropsis , Berthawyleria gracilis , Prodoedicurus wyleri , Isostilomys intermedius y Xotodon cf. smaltatus ( vide Kraglievich, 1932; Castellanos, 1939, 1954; Mones, 1976; Mones y Castiglioni, 1979). Muchos fósiles hallados rodados en la plataforma de abrasión aledaña a las barrancas costeras del Departamento de San José (figuras 1 y 2) probablemente representen una mezcla de elementos de la Formación Camacho con otros procedentes de depósitos suprayacentes. Entre ellos se destacan Pseudoplohophorus berroi (Kraglievich) (véase Sistemática), Castellanosia establei Kraglievich, Eomegatherium nanum uruguayense Kraglievich, Pliomorphus aff. robustus Ameghino, Menilaus berroi Kraglievich, Toxodontherium listai Kraglievich, Palaeotoxodon paranensis (d'Orbigny y Laurillart), Xotodon smaltatus Kraglievich, Berroia gregoriense Kraglievich, Scalabrinitherium ferreriai Kraglievich, Oxyodontherium zeballosi Ameghino ( vide Kraglievich, 1932), Trigodon aff. gaudryi Ameghino, Pseudoplohophorus francisi Mones ( vide Mones, 1967; 1970). Dados el gran endemismo y el estado fragmentario de gran parte de estos restos, junto a la imprecisa procedencia estratigráfica más arriba señalada, resulta poco conveniente tener en cuenta este conjunto al momento de establecer inferencias bioestratigráficas. Globalmente y considerando en los casos de las especies exclusivas del Uruguay sólo los biocrones genéricos, se observa una mezcla de formas huayquerienses y montehermosenses.
Varios taxones supuestamente provenientes de niveles de la Fm. Camacho en la Bahía de Colonia del Sacramento como Scelidodon corderoi Kraglievich, Prolestodon atavus Kraglievich, Eleutherocercus vilardeboi Kraglievich, Pseudoplohophorus coloniensis y Gyriabrus teisseirei (Kraglievich, 1932) también se encontraron removidos y algunos ( e.g . S . corderoi ) son de dudosa asignación a los sedimentos en cuestión. Todos ellos son especies exclusivas del Uruguay pero considerados a nivel genérico, la mayoría tiene un registro bastante amplio que va del Chasiquense al Montehermosense (Bondesio, 1986, Pascual et al. , 1996), estando Prolestodon también representado en la Formación Ituzaingó (Calcaterra, 1977) y Pseudoplohophorus en los citados depósitos más sudorientales de la Formación Camacho.
El "Mesopotamiense", o más propiamente el "Conglomerado osífero" de la Formación Ituzaingó (Cione et al. , 2000), es una unidad informal cuya geocronología generó cierta confusión. Su fauna se asimiló a aquéllas de las Edades Chasiquense, Huayqueriense y Montehermosense (Pascual, 1970). Pascual y Bondesio (1982) la asocian a la "Edad de las Planicies Australes" (Mioceno Tardío-Plioceno Temprano). Cozzuol (1993), plantea que su fauna se puede interpretar como el producto de un área de contacto entre regiones con historias evolutivas disyuntas. Actualmente se opina que su fauna terrestre representa una edad Huayqueriense (Cione et al. , 2000).
La comentada similitud de la fauna proveniente de sedimentitas neógenas del Uruguay con el "Piso Mesopotamiense", ya observada por Kraglievich (1932), puede tener connotaciones paleozoogeográficas interesantes, relacionadas con el desarrollo de un paleodominio particular en la cuenca del Paraná (Zetti, 1972; Cozzuol, 1993). Parece haber existido durante el Neógeno una clara diferenciación biogeográfica entre las áreas pampeana y del noroeste argentino por un lado y de la mesopotamia por el otro, esta última más estrechamente vinculada a las áreas de Uruguay y de Acre en Brasil (Cione et al. , 2000; Candela y Morrone, 2003).
Esto puede estar estrechamente vinculado a la presencia de un brazo de mar que abarcó la Mesopotamia argentina y cuya extensión allende esta área es motivo de controversia (Räsänen et al. , 1995). Este puede haber actuado como barrera en estrecha relación con corredores zoogeográficos terrestres durante buena parte del Mioceno, lo que puede haber motivado la similitud de la mastofauna de la Formación Camacho con otras más norteñas, en particular la señalada para la Formación Solimões en Acre (Rancy et al. , 1989, Latrubesse et al., 1997).
Los datos paleoautoecológicos y tafonómicos que surgen de los depósitos en cuestión en el Departamento de San José concuerdan con una génesis sedimentaria de tipo llanura de inundación de un sistema fluvial (De Santa Ana et al. , 1988), asociado a lagos o lagunas someros y a ecosistemas terrestres con elevada productividad.
El grado de articulación y de asociación de los elementos óseos hallados permiten sugerir un transporte limitado y probable exposición subaérea previa al sepultamiento definitivo. Se hallaron fósiles con articulaciones muy móviles en posición, v.gr. huesos de las extremidades, y cráneo y mandíbula articulados (Perea, 1988; Perea et al. , 1994) o ejemplares, como el que motiva el estudio central de este trabajo, con huesos en franca asociación y con los elementos de la mano casi en posición original. Los restos de reptiles dulceacuícolas (Perea et al. , 1996), flamencos (Phoenicopteridae) (Ubilla et al. , 1990) y de mamíferos probablemente folívoros (Perea, 1988), permiten proponer los ecosistemas arriba mencionados asociados al sistema principal generador de los sedimentos. Los datos paleoecológicos y paleogeográficos de la región costera uruguaya y zonas aledañas durante el Neógeno (Sprechmann, 1978; Sprechmann et al. , 1994, Cione et al. , 2000) no son contradictorios con esta propuesta.
Agradecimientos
A G.J. Scillato-Yané, A. Carlini, G. McDonald y J.C. Fernicola por sus valiosos comentarios acerca del manuscrito. A M. Aznárez por facilitar el estudio del ejemplar neotipo de Pseudoplohophorus orientalis . A S. Vizcaíno, por proporcionarme fotografías del ejemplar FMNH-P14501. A M. Ubilla, R. Fariña, G. Veroslavsky, S. Martínez y T. Olivera, por colaborar en la extracción de diferentes partes del holotipo de Pseudoplohophorus absolutus n. sp. Contribución al proyecto CSIC-UDELAR "Depósitos cenozoicos continentales y parálicos del Uruguay".
Bibliografía
1. Aceñolaza, F.G. 1976. Consideraciones bioestratigráficas sobre el Terciario marino de Paraná y alrededores. Revista de la Asociación Geológica Argentina 31: 61-62. [ Links ]
2. Ameghino, F. 1887. Apuntes preliminares sobre algunos mamíferos extinguidos del yacimiento de "Monte Hermoso" existentes en el "Museo La Plata". Boletín del Museo La Plata , 1: 1-20, láms. 1-2. Buenos Aires [ Links ]
3. Ameghino, F. 1889. Contribuci ón al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias , Córdoba, 6: 1-1027; Atlas. [ Links ]
4. Ameghino, F. 1897. Mammif ères crétacés de l'Argentine. Boletín del Instituto Geográfico Argentino 18 (4-6): 406-429; (7-9): 431- 521. [ Links ]
5. Ameghino, F. 1902. Notices pr éliminaires sur des mammifères nouveaux des terraines crétacés de Patagonie. Boletín de la Academia Nacional de Ciencias, Córdoba 17: 5-70. [ Links ]
6. Alberch, P., Gould, S.J., Oster, G.F. y Wake, D.B. 1979. Size and shape in ontogeny and phylogeny. Paleobiology 5: 296-317. [ Links ]
7. Bondesio, P. 1986. Lista sistem ática de los vertebrados terrestres del Cenozoico de Argentina. 4º Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 2: 187-190. [ Links ]
8. Burmeister, G. 1879. Description physique de la Repúblique Argentine d'après des observations personnelles et étragères. 3 (animaux vertébrés, 1: Mammifères vivants et éteints) . P.E. Coni, Buenos Aires, 555 pp. [ Links ]
9. Butler, R.F., Marshall, L.G., Drake, R.E y Curtis, G.H. 1984. Magnetic polarity stratigraphy and 40K-40 Ar dating of Late Miocene and Early Pliocene continental deposits, Catamarca Province, NW Argentina. Journal of Geology 92: 623-636. [ Links ]
10. Cabrera, A. 1939. Sobre vertebrados f ósiles del Plioceno de Adolfo Alsina. Revista del Museo de La Plata 2: 3-35. [ Links ]
11. Cabrera, A. 1944. Los Gliptodontoideos del Araucaniano de Catamarca. Revista del Museo de La Plata (N. Serie), Paleontología 3: 5-76. [ Links ]
12. Calcaterra, A. 1977. El g énero Lestodon (Edentata-Mylodontidae). Estudio preliminar. Comunicaciones Paleontológicas del Museo Municipal Real de San Carlos 1: 1-30. Colonia. [ Links ]
13. Candela, A.M. y Morrone, J.J. 2003. Biogeograf ía de puercoespines neotropicales (Rodentia, Hystricognathi): Integrando datos fósiles y actuales a través de un enfoque panbiogeográfico. Ameghiniana 40: 361-378. [ Links ]
14. Caorsi, J.H. y Go ñi, J.C. 1958. Geología Uruguaya. Boletín del Instituto Geológico del Uruguay 37: 1-73. [ Links ]
15. Castellanos, A. 1926. Sobre un nuevo gliptod óntido chapadmalense Urotherium simplex n. gen. y n. sp. y las formas afines. Anales del Museo Nacional de Historia Natural Bernardino Rivadavia 34, Paleontología: Vertebrados 60: 263-278. [ Links ]
16. Castellanos, A. 1939. A prop ósito de los géneros Plohophorus , Nopachthus y Panochthus. Publicaciones del Instituto de Fisiografía y Geología 18: 1-278. [ Links ]
17. Castellanos, A. 1940. A prop ósito de los géneros Plohophorus , Nopachthus y Panochthus. Publicaciones del Instituto de Fisiografía y Geología 20: 279-418. [ Links ]
18. Castellanos, A. 1949. Nuevos restos de Stromaphoropsis Kragl. del Uruguay. Publicaciones del Instituto de Fisiografía y Geología 37: 1-48. [ Links ]
19. Castellanos, A. 1954. Determinaci ón de las especies de Stromaphoropsis Kragl. Anales del Museo de Historia Natural de Montevideo 2, 3: 1-23. [ Links ]
20. Cione, A.L., Azpelicueta, M.M. Bond, M., Carlini, A.A., Casciotta, J.R., Cozzuol, M.A., De la Fuente, M., Gasparini, Z., Goin, F.J., Noriega, J., Scillato-Yan é, G.J., Soilbenzoin, L., Tonni, E.P., Verzi, D., Vucetich, M.G. 2000. Miocene vertebrates from Entre Ríos province, eastern Argentina. En: Aceñolaza, F.G. y Herbst, R. (eds.) El Neógeno de Argentina . INSUGEO, Serie Correlación Geológica, Tucumán. 14: 191-237. [ Links ]
21. Comit é Argentino de Estratigrafía. 1992. Código Argentino de Estratigrafía . Asociación Geológica Argentina, Serie B, Didáctica y Complementaria 20: 1-64. [ Links ]
22. Cozzuol, M. A. 1993. El Mesopotamiense, "Mezcla Temporal o Vía de Dispersión?". Ameghiniana 30: 103. [ Links ]
23. De Santa Ana, H., Veroslavsky, G., Mart ínez, S., Perea, D. y Ubilla, M. 1988. Estudio estratigráfico preliminar de la secuencia mio-pliocena del SW del Uruguay. Actas del 6º Panel de Geología del Litoral y I Reunión de Geología del Uruguay , Actas : 62-67, figs. 1-2. [ Links ]
24. Dozo, M.T., Monti, A., Bouza, P., Vucetich, M.G., Cione A. L., Tonni, E.P. y Scillato-Yan é, G.J. 2002. Geología y Vertebrados Continentales en cercanías de Punta Delgada (Neógeno de Península Valdés, Chubut, Argentina). Actas 15º Congreso Geológico Argentino , Santa Cruz. [ Links ]
25. Fariña, R. 1981. Anatomía comparada de los tubos caudales de Plohophorini (Mammalia, Glyptodontidae), del Montehermosense del Uruguay. 2º Congresso Latino-Americano de Paleontologia (Porto Alegre, 1981), Anais 2: 779-793. [ Links ]
26. Flynn, J.J. y Swisher, C.C. 1995. Cenozoic South American land mammal ages: correlation to global geochronologies. Society for Sedimentary Geology Special Publication 54: 317-333. [ Links ]
27. Frailey, C.D. 1986. Late Miocene and Holocene mammals, exclusive of the Notoungulata, of the Rio Acre Region, Western Amazonia. Contributions in Science 374: 1-46. [ Links ]
28. Francis, J.C. y Mones, A. 1965. Contribuci ón a la Geología y Paleontología de las Barrancas de San Gregorio, Departamento de San José, República Oriental del Uruguay. Kraglieviana 1: 55-85. [ Links ]
29. Gillette, D. y Ray, C. 1981. Glyptodonts of North America. Smithsonian Contributions in Paleobiolog y 40: 1-255. [ Links ]
30. Goso, H. y Bossi, J. 1966. Cenozoico. En: J. Bossi (ed.), Geología del Uruguay , Universidad de la República, Montevideo, pp. 259-301 [ Links ]
31. Gould, S.J. 1980. Ontogeny and Phylogeny . Harvard University Press, 501 pp. [ Links ]
32. Hedberg, H.D. 1994. International Stratigraphic Guide. International Union of Geological Sciences, Amos Salvador, Thondheim, 213 pp. [ Links ]
33. Hoffstetter, R. 1958. Xenarthra. En: J. Piveteau (ed.), Traité de Paléontologie , Masson et Cie. Paris, 2: 535-636. [ Links ]
34. Kraglievich, L. 1931, Sobre la presencia de Toxodontes "Haplodontotéridos" en el Piso Araucanense de Catamarca ( Toxodontotherium andinum n. sp.). Anales de la Sociedad Científica Argentina 3: 93-96. [ Links ]
35. Kraglievich, L. 1932. Nuevos apuntes para la Geolog ía y Paleontología del Uruguay. Anales del Museo de Historia Natural de Montevideo 2: 1-65. [ Links ]
36. Latrubesse, E.M., Bocquentin, J., Santos, J.C.R. y Ramonel, C.G. 1997. Paleoenvironmental Model for the Late Cenozoic of Southwestern Amazonia: Paleontology and Geology. Acta Amazonica 27: 103-118. [ Links ]
37. Marshall. L.G. y Cifelli, R. 1991. Analysis of changing diversity patterns in cenozoic Land Mammal Age Faunas, South America. Palaeovertebrata 19: 169-210. [ Links ]
38. Marshall. L.G. y Patterson, B. 1981. Geology and Geochronology of the mammal-bearing Tertiary of the Valle de Santa Mar ía and río Corral Quemado, Catamarca province, Argentina. Fieldiana Geology 9: 1-80. [ Links ]
39. Marshall, L.G., Hoffstetter, R. y Pascual, R. 1983. Mammals and Stratigraphy: Geochronology of the Continental Mammal-bearing: Tertiary of South America. Palaeovertebrata , Memoire. Extraordinaire 1-90. [ Links ]
40. Martínez, S. 1994. [ Bioestratigrafía (invertebrados) de la Formación Camacho (Mioceno, Uruguay) . Tesis Doctoral, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 346 pp. inédito] [ Links ]
41. McBee, K. y Baker, R.J. 1982. Dasypus novemcinctus . Mammalian Species 162: 1-9. [ Links ]
42. McDonald, H. G. y Perea, D. 2002. The large Scelidothere Catonyx tarijensis (Xenarthra, Mylodontidae) from the Pleistocene of Uruguay. Journal of Vertebrate Paleontology 22: 677- 683. [ Links ]
43. McNamara, K.J. 1988. Patterns of Heterochrony in the Fossil Record. Tree 3: 176-180. [ Links ]
44. McKinney, M. L. y Gittleman, J.L. 1995. Ontogeny and Phylogeny: tinkering with Covariation in Life History, Morphology and Behaviour. En: K.J. McNamara (ed.), Evolutionary Change and Heterochrony , John Wiley & Sons. Pp. 21-47. [ Links ]
45. Mones, A. 1967. Notas paleontol ógicas uruguayas, 1. Trigodon Ameghino, 1882 (Toxodonta, Notoungulata) en la fauna pliocena superior de las barrancas de San Gregorio, Dpto. de San José, Uruguay. Comunicaciones Zoológicas del Museo de Historia Natural de Montevideo 9: 1-4. [ Links ]
46. Mones, A. 1970. Notas paleontol ógicas uruguayas, 2. Un nuevo Pseudoplohophorus Castellanos, 1926 (Hoplophorinae, Edentata) de la R. O. del Uruguay. Pseudoplohophorus francisi n. sp. Comunicaciones Paleontológicas del Museo Nacional de Historia Natural de Montevideo 1: 1-9. [ Links ]
47. Mones, A. 1976. Notas paleontol ógicas uruguayas, III. Vertebrados fósiles nuevos o poco conocidos (Chondrichthyes, Osteichthyes, Amphibia, Mammalia). Ameghiniana 12: 343-349. [ Links ]
48. Mones, A. 1986. Palaeovertebrata Sudamericana. Cat álogo Sistemático de los Vertebrados Fósiles de América del Sur. Parte I. Lista Preliminar y Bibliografía. Courier Forschungsinstitut Senckenberg , 82 Frankfurt a.M.: 1-625. [ Links ]
49. Mones, A. 1987. Sobre tres especies de Gliptodontes del Uruguay: Cingulata: Glyptodontidae). 4º Congreso Latinoamericano de Paleontología (Bolivia, 1987), Memoria 1: 500-515. [ Links ]
50. Mones, A. y Castiglioni, L.R. 1979. Additions to the knowledge on fossil Rodents of Uruguay (Mammalia: Rodentia). Paläontologische Zeitschrifft 53: 77-87. [ Links ]
51. Pascual, R. 1970. Evoluci ón de las comunidades, cambios faunísticos e integraciones biocenóticas de los vertebrados cenozoicos de Argentina. 4º Congreso Latinoamericano de Zoología (Caracas, 1970), Actas 2: 991-1088. [ Links ]
52. Pascual, R., Ortega Hinojosa, E.J., Gondar D. y Tonni. E. 1966. Las Edades del Cenozoico mamal ífero de la Argentina con especial atención a aquellas del territorio bonaerense. Paleontografía Bonaerense , 4 Vertebrata. Comisión de Investigación Científica, Buenos Aires: 3-27. [ Links ]
53. Pascual, R. y Bondesio, P. 1982, Un roedor Cardiatheriinae (Hydrochoeridae) de la Edad Huayqueriense(Mioceno Tard ío) de la Pampa. Sumario de los ambientes terrestres en la Argentina durante el Mioceno. Ameghiniana 19: 19-35. [ Links ]
54. Pascual, R. y Bondesio, P. 1985. Mam íferos terrestres del Mioceno Medio-Tardío de las cuencas de los Ríos Colorado y Negro (Argentina): evolución ambiental. Ameghiniana 22: 133-145. [ Links ]
55. Pascual, R., Ortiz Jaureguizar, E. y Prado, J.L. 1996. Land Mammals: Paradigm for Cenozoic South American Geobiotic Evolution. Münchner Geowissenschaftliche Abhandlungen (A) 30: 265-319. [ Links ]
56. Paula Couto, C. de. 1979. Tratado de Paleomastozoologia . Académia Brasileira de Ciencias, Rio de Janeiro, 590 pp. [ Links ]
57. Perea, D. 1988. Dos Nothrotheriinae (Tardigrada, Megatheriidae) del Mio-Plioceno de Uruguay. Ameghiniana 25: 381-388. [ Links ]
58. Perea, D. 1993. [ Xenarthra del Neógeno del Uruguay: La biozona de Stromaphoropsis Kragl. (Glyptodontidae ), Edad-Mamífero Huayqueriense. Tesis de maestría, Universidad de la República, Uruguay. 139 pp. Inédito] [ Links ].
59. Perea, D. y Mart ínez, S. 2004. Estratigrafía del Mioceno- Pleistoceno en el Litoral Sur-Oeste de Uruguay. En: G. Veroslavsky, M. Ubilla y S. Martínez (eds.), Cuencas sedimentarias de Uruguay, geología, paleontología y recursos minerales , DIRAC, Montevideo, pp. 105-124. [ Links ]
60. Perea, D. y Scillato-Yan é, G.J. 1990. Ranculcus Ameghino (Xenarthra: Mylodontinae) en el Mioceno superior del Uruguay. 1º Congreso Uruguayo de Geología (Montevideo, 1990), Resúmenes Ampliados 2: 1-4. [ Links ]
61. Perea, D. y Scillato-Yan é, G.J. 1995. Proeuphractus limpidus Ameghino, 1886 (Xenarthra, Dasypodidae, Euphractini): Osteología comparada del cráneo y elementos de la coraza asociados (Neógeno del Uruguay). Boletín de la Real Sociedad Española de Historia Natural (Sección Geología) 90: 125-130. [ Links ]
62. Perea, D., Ubilla, M., Mart ínez, S., Piñeiro, G. y Verde, M. 1994. La Edad-Mamífero Huayqueriense (Mioceno Superior) en el Uruguay. Acta Geologica Leopoldensia 17: 375-389. [ Links ]
63. Perea, D., Ubilla, M. y Pi ñeiro, G. 1996. First fossil record (Late Miocene) of Phrynops ( P . geoffroanus complex; Chelidae) from Uruguay; bioestratigraphical and paleoenvironmental context. Copeia 2: 445-451. [ Links ]
64. Rancy, A., Villanueva, J.B., Souza Filho, J.P., Santos, J. C.R. y Negri, F.R. 1989. Lista preliminar da fauna de vertebrados do Neogeno da Regiao oriental do Estado do Acre, Brasil (material depositado em Rio Branco). Ameghiniana 26: 249. [ Links ]
65. Räsänen, M.E., Linna, A.M., Santos, J.C.R. y Negri, F.R. 1995. Late Miocene Tidal Deposits in the Amazonian Foreland Basin. Science 269: 386-390. [ Links ]
66. Riggs, E.S. y Patterson, B. 1939. Stratigraphy of Late Miocene and Pliocene deposits of the Province of Catamarca (Argentina) with notes on the faunae. Physis 14: 143-162. [ Links ]
67. Rohlf, F.J. 2000. NTSYS-pc Numerical Taxonomy and Multivariate Analysis System Version 2.1 . Exeter software. New York. [ Links ] Scillato-Yané, G.J. 1977. Sur quelques Glyptodontidae nouveaux (Mammalia, Edentata) du Déséadien (Oligocène inférieur) de Patagonie (Argentine). Bulletin du Muséum National d' Histoire Naturelle (3) 487 ( Sciences de la Terre , 64): 249-259. [ Links ]
68. Serra, N. 1943. Memoria explicativa del mapa geol ógico del Departamento de Colonia. Boletín del Instituto Geológico del Uruguay 30: 1-50. [ Links ]
69. Simpson, G.G. 1974. Notes on Didelphidae (Mammalia, Marsupialia) from the Huayquerian (Pliocene) of Argentina. American Museum Novitates 2559: 1-15. [ Links ]
70. Sneath, P.H.A. y R. Sokal. 1973. Numerical Taxonomy . W.H. Freeman and Company, San Francisco, 573 pp. [ Links ] Sprechmann, P. 1978. The paleoecology and paleogeography of the Uruguayan coastal area during the Neogene and Quaternary. Zitteliana 4: 3-72. [ Links ]
71. Sprechmann, P., Mart ínez S. y Gaucher, C. 1994. Paleoecología y sedimentología de la Formación Camacho en el Departamento de Colonia (Mioceno Medio-Superior, Uruguay). 1ª Jornada de Paleontología del Uruguay (Montevideo, 1994), Resúmenes. Ampliados , 21-24. [ Links ]
72. Tambussi, C., M. Ubilla y Perea, D. 1999. The Youngest Large Carnassial Bird (Phorusrhacidae, Phorusrhacinae) from South America (Pliocene-Early Pleistocene of Uruguay). Journal of Vertebrate Paleontology 19: 406-408. [ Links ]
73. Ubilla, M., Perea, D., Tambussi, C. y Tonni, E.P. 1990. Primer registro f ósil de Phoenicopteridae (Aves: Charadriiformes) para el Uruguay (Mio-Plioceno). Anais da Académia Brasileira de Ciencias 62: 61-68. [ Links ]
74. Verde, M. y Perea, D. 1992. Mam íferos "Mesopotamienses" (Xenarthra, Litopterna y Cetacea) (Mioceno Superior) de la Bahía de Colonia, Uruguay. Boletín de la Sociedad. Zoológica del Uruguay (2da. Epoca) 7: 31-32. [ Links ]
75. Yepes, J. 1931. El escudete cef álico del pichi ciego menor ( Chlamyphorus truncatus Harl.). Revista Chilena de Historia Natural 35: 107-112. [ Links ]
76. Zelditch, M. L. y Fink, W. L 1996. Heterochrony and heterotopy: stability and innovation in the evolution of form. Paleobiology 22: 241-254. [ Links ]
77. Zetti, J. 1972. [ Los Mamíferos fósiles de edad Huayqueriense (Plioceno Medio) de la Región Pampeana . Tesis Universidad Nacional de La Plata, 86 pp., Inédito] [ Links ].
Recibido: 12 de setiembre de 2003.
Aceptado: 10 de junio de 2004.

