










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.2 Buenos Aires mar./jun. 2005
Maastrichtian and Danian species of Turkostreini (Ostreidae, Crassostreinae) from the Neuquén Basin, Argentina
Miguel Griffin 1,2 , Silvio Casadío 1 and Ana Parras 1
1 Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, La Pampa, Argentina. miguelgriffin@aol.com, scasadio@cpenet.com.ar, aparras@cpenet.com.ar
2 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Abstract. The record of Turkostrea damboreneae nom. nov. and Cubitostrea primordialis n. sp. in the Maastrichtian Jagüel and Roca Formations and of Turkostrea argentina n. sp., in the Danian beds of the Roca Formation reveals that the two genera involved have a history that reaches back into the Late Cretaceous in northern Patagonia. Turkostrea damboreneae nom. nov. and Cubitostrea primordialis n. sp. are the two oldest representatives of the genera in which they belong. The two species of Turkostrea show clear affinities with other species of this genus that thrived in the Paleogene of the Northern Hemisphere. Cubitostrea primordialis n. sp. is closely allied to the previously known Paleocene species common in the region. While Turkostrea appears to have survived the Cretaceous/Paleogene boundary, it is not found in Patagonia in rocks younger than Danian. Cubitostrea also survived the Cretaceous/Paleogene boundary but, although for diverse reasons absent from Eocene-Oligocene rocks in southern South America, is again common in early and late Miocene sediments exposed at various localities there. Based on these records, the possible South American origin of the Turkostreini is discussed.
Resumen. Especies Danianas y Maastrichtianas de Turkostreini (Ostreidae , Crassostreinae ) from the Neuquén basin , Argentina . El registro de Turkostrea damboreneae nom. nov. y Cubitostrea primordialis n. sp. en sedimentitas maastrichtianas asignadas a las Formaciones Jagüel y Roca, y de Turkostrea argentina n. sp., en niveles danianos de la Formación Roca revela para ambos géneros una historia en el norte de Patagonia que se retrotrae al Cretácico Tardío. Turkostrea damboreneae nom. nov. y Cubitostrea primordialis n. sp. son las representantes más antiguas de los respectivos géneros a los que son asignadas. Las dos especies de Turkostrea muestran claras afinidades con otras especies de este género presentes en el Paleógeno del hemisferio norte, mientras que Cubitostrea primordialis n. sp. está estrechamente vinculada a la especie paleocena que se conocía previamente en el área de estudio. Si bien Turkostrea sobrevivió al límite Cretácico/Paleógeno, en Patagonia no se registra con posterioridad al Daniano. Cubitostrea también sobrevivió a los eventos del límite Cretácico/Paleógeno, aunque por diversas razones se halla ausente en rocas que comprenden el intervalo Eoceno-Oligoceno en el extremo austral de América del Sur, pero vuelve a ser frecuente en las sucesiones del Mioceno expuestas en numerosas localidades de Argentina. Sobre la base de estos nuevos registros se discute el probable origen sudamericano de las Turkostreini.
Key words. Oysters; Systematics; Cretaceous-Paleogene boundary; Neuquén Basin; Argentina.
Palabras clave. Ostras; Sistemática; Límite Cretácico-Paleógeno; Cuenca Neuquina; Argentina.
Introduction
Oysters are amongst the most frequent and characteristic fossils in the sedimentary marine sequences of the Patagonian Mesozoic and Cenozoic. They were particularly diverse and abundant during the Late Cretaceous-Paleogene transition in the Neuquén Basin, where they appear in vast accumulations throughout several lithostratigraphic units. Many nominal species have been described since these rocks were first described during the second half of the nineteenth Century. The great majority of these species were -unfortunately- based on only a limited number of specimens, in many cases with inadequate stratigraphic information. This naturally rendered their use as biostratigraphic tools almost valueless, although a number of authors did in fact base stratigraphic divisions on their presence/absence in certain horizons. However, as a comprehensive taxonomic revision of the taxa involved was missing, such divisions based solely on oyster content could not hold fast or either did so very weakly. This situation -for the Neuquén Basin- was remedied in part by Casadío (1998), who studied large collections and revised most types, setting the foundations for an in depth revision of all oysters in the Maastrichtian-Cenozoic of Patagonia. The large data set available to him allowed a sound biostratigraphic scheme, proving the potential usefulness of this group of bivalves for regional correlation within the southern portion of South America.
In this paper we describe three species of oysters -two of them are new- collected in rocks referred to the Maastrichtian-Danian Jagüel and Roca Formations in the Neuquén Basin. One of these species ( Cubitostrea primordialis n. sp.) was collected in Maastrichtian beds of the Jagüel Formation, being the oldest record of this genus. The other two are Turkostrea damboreneae nom. nov. [= Turkostrea mendozana (Fritzsche, 1919), p. 361], from the Maastrichtian Jagüel Formation and from the Maastrichtian beds of the Roca Formation, and Turkostrea argentina n. sp. from the Danian part of the Roca Formation. All three species suggest a probable austral origin for the Tribe Turkostreini.
Geological setting
During the Cretaceous-Paleogene transition, the southernmost tip of South America was reduced to an archipelago (Urien et al ., 1995; Malumián, 1999). Malumián and Caramés (1995) estimated a minimum area of 507,000 km 2 of present day Patagonia as covered by the Atlantic during the early Paleocene.
This large transgression surface was an important paleogeographic element in the continent. It surely played a paramount role in climatic and evolutionary events, acting as a biogeographic barrier between Patagonia and the rest of South America for many groups (Pascual, 1998; Casadío, 1999). Concomitantly, the Andes also became an important feature in the continent (Uliana and Biddle, 1988; Macellari, 1988; Legarreta et al. , 1989; Barrio, 1990a, 1990b, 1991, Casadío, 1994) and prevented an Atlantic-Pacific connection (Tunik, 2001).
In the Neuquén Basin, in northwestern Patagonia, the sediments that record the Cretaceous-Paleogene transition are included in the Malargüe Group. They are well exposed in southern Mendoza, northeastern Neuquén, western La Pampa and northern Río Negro (figure 1). These rocks were studied by several authors since the nineteenth century (Camacho, 1992) and, during the past few years, were subject to renewed interest. This interest is undoubtedly caused by the fact that they contain the Cretaceous-Paleogene boundary, in both continental and marine facies (Parras and Casadío, 1999). Therefore they provide an excellent opportunity to investigate the environmental and faunal changes in continental and marine ecosystems of the mid-latitudes in the southern hemisphere at the close of the Cretaceous.
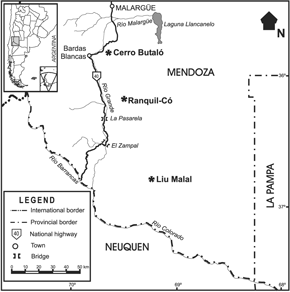
Figure 1. Map of southern Mendoza showing fossil localities / Mapa del sur de Mendoza mostrando las localidades fosilíferas.
In the northeastern region of the basin, the Malargüe Group reaches over 600 m and comprises, from bottom to top, the Loncoche, Jagüel, Roca, and Pircala formations (figure 2). In the southeastern region it reaches up to 200 m thick and comprises the Allen, Jagüel, Roca, and El Carrizo formations.
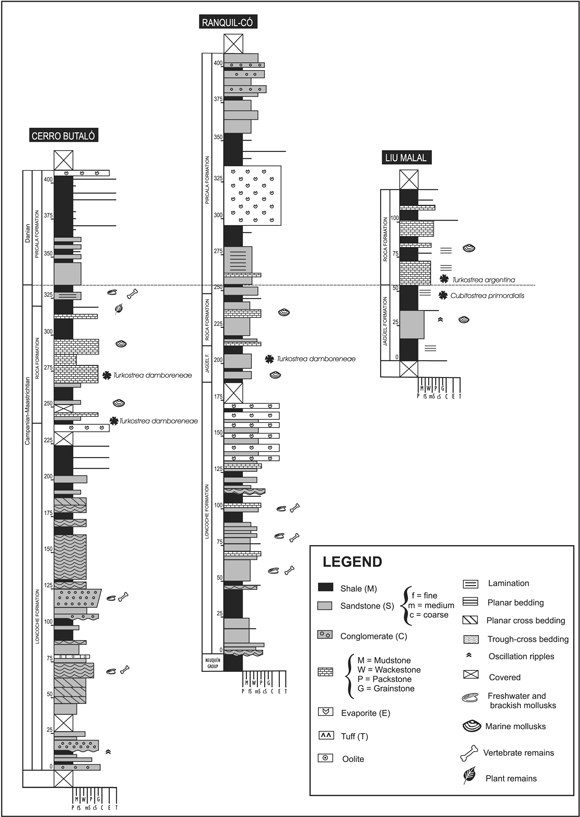
Figure 2. Stratigraphic sections at the three localities studied, showing lithology and fossil-bearing beds; vertical scale in meters / Perfiles columnares de las tres localidades estudiadas, mostrando la litología y los niveles fosilíferos; escala vertical en metros.
Uliana and Dellapé (1981) interpreted the Allen Formation as a littoral to restricted marine environment. At the same time, Barrio (1991) pointed out that, in the eastern portion of the basin, the facies association suggests intertidal conditions, while these were of a tide-dominated estuary in the western part. This difference was attributed to different tide ranges directly related to the basin geometry. The equivalent unit in the Andean area is the Loncoche Formation. The Allen Formation and its equivalent are overlain by marine rocks of the Jagüel Formation, which are mainly siltstone. The Jagüel Formation is transitionally overlain by the limestones of the Roca Formation, of Maastrichtian-Danian age (Casadío et al. , 1998). Overlying the Roca Formation are evaporitic deposits of the El Carrizo Formation, while in the Andean area it gradually gives way to the continental sandstone of the Pircala Formation.
The observed paleoenvironmental differences between the eastern and the Andean sectors of the basin -evidenced by the prevalence of continental and delta facies as well as the abundance of pyroclastic material in the Andean area- suggest the latter was situated near an emergent and volcanically active arc (Parras et al. , 1998; Parras and Casadío, 1999). On the other hand, the eastern region shows a predominance of marine deposits, mainly constituted by siltstone, limestone and evaporites, that suggest a distal position from the supply areas and, at the same time, close oceanic links.
The Cretaceous-Paleogene boundary coincides with a significant decrease of the endemic taxa with austral affinities and an increase in the number of species from low latitudes. This modification in the faunal composition was coeval with an increase of the relative sea level and the beginning of carbonatic sedimentation (Feldmann et al. , 1995; Casadío et al ., 1999).
The studied fossils come from beds referred to the Jagüel and Roca formations exposed in southern Mendoza (figures 1 and 2). All specimens are housed in the Departamento de Ciencias Naturales of the Universidad Nacional de La Pampa (GHUNLPam).
Systematic paleontology
Family O STREIDAE Rafinesque, 1815
Subfamily C RASSOSTREINAE Scarlato and
Starobogatov, 1979 ( ex Crassostreidae)
Tribe T URKOSTREINI Malchus, 1990
Original diagnosis. Gryphaea homoeomorph, thick and many chomata, extremely lens-foliated shell (extracted from Malchus, 1990, p. 197, 1.6.7, lines 5-8).
Emended diagnosis. Shell thick, sickle shaped to subtriangular, or Gryphaea homoeomorph, with many strong chomata, left valve with radial ribs, compact to extremely lens-foliated shell.
Remarks. In a major monograph mainly concerned with Cretaceous oysters published almost two decades after Stenzel's description of the genus, and apparently unaware of Vyalov's ideas, Malchus proposed the Tribe Turkostreini (Malchus, 1990, p. 196-197). He included in his new tribe Turkostrea Vyalov, 1936 (type species Ostrea turkestanensis Romanovsky, 1878), Turkostrea ( Cymbulostrea ) Sacco, 1897b (type species Ostrea cymbula Lamarck, 1806), Cubitostrea Sacco, 1897a (type species Ostrea cubitus Deshayes, 1832), Sokolowia Böhm, 1933 (type species Gryphaea buhsii Grewink, 1853) and Ferganea Vyalov, 1936 (type species Gryphaea swerzowii Romanovsky, 1884). He included this tribe in the Crassostreinae, a subfamily previously defined by Torigoe (1981, p. 303). At the same time (Malchus, 1990, p. 197), he stated that the Crassostreinae and the Turkostreini appeared at some time during the earliest Paleogene as lineages derived probably from the Liostreinae Malchus (1990, p. 150), and within them probably form the Flemingostreini or the Ambigostreini. This subfamily of Ostreidae he proposed in the same paper, surely unaware of Vyalov's previous introduction of the name (Vyalov, 1983, p. 7). On introducing the tribe Turkostreini in his monograph of predominantly Cretaceous oysters, Malchus (1990) gave only a rather brief, informal diagnosis. While we mostly agree with Malchus (1990) in the systematic arrangement he proposed for the oysters he included in his new tribe, we feel that Ferganea belongs better elsewhere. The large, smooth shells with weak chomata and very weak terebratuloid fold suggest that it may fit perhaps in the Flemingostreini as described by Stenzel (1971, p. N1150). On the other hand, as already stated by Stenzel (1971, p. N1150), Sokolowia Böhm appears to be a Gryphaea homeomorph but probably descended from a Turkostrea stock, as revealed by its ribbed left valve, chomata and adductor shell muscle imprint. Thus restricted, the Tribe Turkostreini includes a group of genera sharing the following features: shells ovoid to subtriangular or even sickle shaped, strongly inequivalve, thick, umbos either opisthogyrate to different degrees or else incurved in a Gryphaea -like manner; with strong lathlike chomata at least near the ligament; left valve ornamented by radial ribs, generally strong; right valve smooth or with growth lamellae; adductor muscle scar elongate with an almost straight to concave dorsal margin; shell structure compact foliate to lenticular foliate with mocret.
According to Malchus (1990, p. 196) the stratigraphic range of the Turkostreini spans the Paleocene-Oligocene, although he mentioned that the group may have evolved already in the Cretaceous, possibly from members of the Flemingostreini or Ambigostreini. In Patagonia, at least one clear representative of this group appeared in the Late Cretaceous and at least two species survived in South America into the Miocene, one of them becoming extinct in as late as the Tortonian.
Taxonomic composition. Turkostrea Vyalov, 1936 (type species Ostrea turkestanensis Romanovsky, 1878); Cymbulostrea Sacco, 1897b (type species Ostrea cymbula Lamarck, 1806); Sokolowia Böhm, 1933 (type species Gryphaea buhsii Grewingk, 1853); Cubitostrea Sacco, 1897a (type species Ostrea cubitus Deshayes, 1832).
Stratigraphic range. Late Maastrichtian-late Miocene.
Genus Turkostrea Vyalov, 1936
Type species. Ostrea turkestanensis Romanovsky, 1878, p. 112 (= O. strictiplicata Raulin and Delbos, 1855, p. 1158); original designation.
Emended diagonsis. Gryphaea homeomorph with strong chomata, which can be continuous or else restricted to anterior and posterior margins of valves, radial ribs weak and with rounded crests to strong and with sharp crests, ligament generally opisthogyrous.
Remarks. The subgenus Turkostrea was proposed by Vyalov (1936, p. 18) for a group of oysters characterized by their oval to triangular-ovoid, strongly convex left valves ornamented with strong radial ribs, beak strongly developed and incurved to a larger or lesser degree to a gryphoid appearance. He designated Ostrea turkestanensis Romanovsky, 1878 (p. 112) from Paleocene beds in central Asia (then called Turkestan) as type species of his new subgenus. Almost thirty years later, Vyalov (1965) again dwelled on this genus and its relationship with other oyster genera from central Asia. He pointed out that the apparent similarity of his Turkostrea with Fatina Vyalov, 1936 (type species Gryphaea esterhazyi Pávay, 1873 var. beldersaiensis Gorizdro, 1915) was only superficial as the latter had a concave right valve, inducing him wrongly to place it in the Gryphaeidae together with Costeina Vyalov 1965 (type species Ostrea costei Coquand, 1869). While it is true that Costeina does have radial ribs on the left valve only, its placement in the Gryphaeidae was confirmed by Stenzel (1971, p. N1107). On the other hand, as remarked below, Fatina was considered a junior synonym of Sokolowia Böhm, 1933 by Stenzel (1971, p. N1147). Although both taxa show concave right valves, this character seems to be insufficient to separate any of these two genera from Turkostrea , as intermediate forms such as Turkostrea afghanica Vyalov, 1939 (p. 17) have a right valve that, while still convex, is far less so than in Turkostrea turkestanensis . Another species, Turkostrea khaudaguensis Vyalov, 1948 (p. 63, pl. 26, fig. 7-12) has a flat right valve. All three species, i.e ., Turkostrea turkestanensis , Turkostrea afghanica and Turkostrea khaudaguensis coexist in the Alai Stage (Eocene) in central Asia and are possibly ecophenotypes of the same species, i.e ., of Turkostrea turkestanensis . The other difference between Sokolowia and Turkostrea , i.e ., more strongly gryphaeate shape of Sokolowia , could well be a response to different habitats, such as seen in some species of Odontogryphaea from Patagonia (personal observations). A possible example of this are Vyalov's (1965) illustrated specimens from the middle and late Eocene of Fergana, in central Asia. His Ostrea ( Turkostrea ) turkestanensis angustifrons Vyalov, 1965 (p. 7-8, pl. 1, fig. 1-2), and Ostrea ( Turkostrea ) callipyga Vyalov, 1939 (Vyalov, 1965, p. 8-9, pl. 1, fig. 3, pl. 2, fig. 1-2, pl. 3, fig. 2) are forms that could easily be considered morphologically transitional between Sokolowia and Turkostrea . At the same time, the specimen he illustrated as Fatina beldersaiensis (Gorizdro) (Vyalov, 1965, pl. 3, fig. 1), which is the type species of Fatina , is very close to the type species of Turkostrea and there seems to be no reason to segregate it as a different taxon. Therefore, if Stenzel's conclusion that Fatina is a synonym of Sokolowia is eventually followed, then we should conclude that Sokolowia and Turkostrea are synonyms as well. In this case, the senior synonym would be Sokolowia .
Stenzel (1971, p. N1139-N1140) redescribed Turkostrea and grouped it with the incubatory genera within the Ostreidae and mentioned its affinities with Ostrea , of which he also considered it a subgenus (as did Vyalov, 1936) distinguished by its strong chomata and many strong, continuous, fairly narrow radial ribs on the left valve and a tendency of the ligament area to turn in an opisthogyrous spiral fashion. Stenzel's placement of Turkostrea was later contested by Vyalov (1984, p. 31-36) who, in a general classification of oysters, divided the Ostreoidea into the families Ostreidae, Lophidae and Exogyridae, setting the Gryphaeidae apart as a distinct Superfamily. Within the Ostreidae he identified the Ostreinae, which have left valves with radial sculpture and the Liostreinae, with smooth left valves. He apparently used only this character to subdivide the family, allocating to each one a number of previously described genera. Among the Ostreinae he included his genus Turkostrea . Although this genus was used over the years for a number of species, mainly from the northern hemisphere -especially northern Africa, where it appears to have been very abundant- it has been generally poorly defined and its phylogenetic relationships have not been sufficiently well established. Therefore, species that have been described as Turkostrea may belong in other genera. Conversely, species included originally in other oyster genera may prove to belong in Turkostrea .
Turkostrea damboreneae nom. nov.
Figures 3.A-G, 4.A-G

Figure 3. A-G, Turkostrea damboreneae nom. nov. A-B, left valve / valva izquierda , GHUNLPam 10625: A, interior view / vista interna , B, external view / v ista externa ; C, left valve, internal view / valva izquierda, vista interna, GHUNLPam 15811; D-E, right valve / valva derecha , GHUNLPam 15976: D, interior view / vista interna , E, external view / vista externa ; F-G, left valve / valva izquierda , GHUNLPam 10572: F, external view / vista externa, G, internal view / vista interna . Scale bar 1 cm / escala gráfica 1 cm.
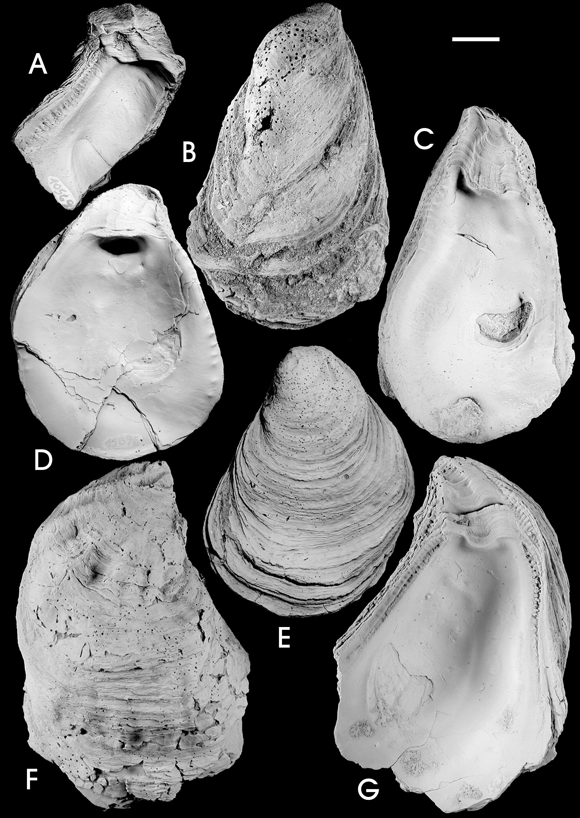
Figure 4. A-G, Turkostrea damboreneae nom. nov. A, left valve, internal view / valva izquierda, vista interna, GHUNLPam 10569; B-C, right valve / valva derecha , GHUNLPam 15664: B, external view / vista externa , C, internal view / vista interna ; D-E, right valve / valva derecha , GHUNLPam 15896: D, internal view / vista interna, E, external view / vista externa ; F-G, left valve / valva izquierda, GHUNLPam 15639: F, external view / vista externa , G, internal view / vista interna . Scale bar 1 cm / escala gráfica 1 cm.
1900 Gryphaea vesicularis Lmk.; Burckhardt, p. 50, pl. 28, fig. 4 only.
1919 Gryphaea mendozana n. sp.; Fritzsche, p. 361. ( non Gryphaea burckhardti mendozana Ihering, 1907, p. 36).
Material. Forty nine specimens, one bivalved shell, partly broken, thirty two left valves and sixteen right valves; twelve come from Cerro Butaló (GHUNLPam 10470, 10478-10479, 10482, 10496, 10554, 10569, 10572, 10581, 10583, 10589) and thirty seven from Ranquil-Có (GHUNLPam 15639-15640, 15644, 15647-15649, 15652, 15664, 15667, 15774-15775, 15777, 15779-15780, 15783, 15793, 15803, 15807, 15811, 15813-15815, 15819-15820, 15827, 15830, 15833, 15853, 15896, 15899, 15903, 15927, 15962, 15976, 15978, 15981, 16009). Additional material (over 800 poorly preserved specimens) was collected by us at various localities other than the three considered herein. These localities include those surveyed by Gerth and from where the syntypes described by Fritzsche come. All the rocks exposed at these localities are Maastrichtian (Parras, 1999; Parras et al. , 1998). The specimen figured by Burckhardt (1900, pl. 50, fig. 4) and housed in the Departamento Científico Paleontología Invertebrados, Museo de La Plata (MLP 10286) is also included in this species.
Diagnosis . Left valve smooth or with four or five radial ribs beginning at one third of shell height and separated by very wide and shallow intercostal spaces.
Description . Shell inequivalve. Left valve ovoid to sub-triangular, expanded towards ventral margin, convex and larger than right valve, surface carrying numerous lamellose commarginal growth lines, some specimens smooth but others with four or five rounded costae, beginning at one third the valve height and diverging towards ventral margin; crest of costae rounded, although most specimens are very worn; attachment area small, near the beak; beak opisthogyrous and incurved in juvenile specimens; growth less gyrate in adults, resulting in an umbo elevated above commissural plane. Right valve: ovoid to sub-triangular, slightly convex on the dorsal half and flat towards the ventral margin; outer surface with commarginal growth lines, ligament area wide, with flat to convex central area. Chomata on margins of both valves, with very conspicuous relict chomata along the anterior and posterior dorsal margins of right valve. Adductor muscle scar large, reniform, with the lower margin convex and the upper one concave, approximately 1.3 cm long and 0.9 cm high and placed slightly posterior to center of valve, closer to the ventral than to the dorsal margin; Quenstedt muscle scar on right valve strongly impressed. Shell microstructure simple compact foliate.
Measurements. The average height for left valves is 58 mm (range 42 to 77 mm); average length is 39 mm (range 30 to 54 mm); average valve thickness near the ventral margin is 7.4 mm (range 4 to 23 mm). The average height for the right valve is 61.5 mm (range 46 to 73 mm); average length is 41 mm (range 34 to 51 mm); average thickness of the valve near ventral margin is 7.8 mm (range 6 to 10 mm).
Discussion. Turkostrea damboreneae nom. nov. Was first described as G. mendozana n. sp. by Fritzsche (1919, p. 361) based on specimens collected by Gerth in four localities in southern Mendoza, all of which were also visited by us. The remaining material, housed in the Goldfuß-Museum für Paläontologie in Bonn, consists of several syntypes. These are all very poorly preserved fragments but all of them agree with the large array of poorly preserved specimens we also found in the type localities. Therefore we have no doubts as to their identity with Fritzsche's species.
Fritzsche stated that his new species was very similar to Gryphaea rothi Böhm, 1903 (p. 71), only distinguishable by its more convex left valve, thick shell, larger ligament area and less sharply incurved umbo. He did not mention the presence of ribs on the left valve, but we must point out here that a large majority (all but just a handful among over eight hundred specimens) of these oysters show no evidence of ribs, in many cases possibly because they are all very worn, as is the case with all the material coming from the type localities. However, a careful search through the sections where he collected his material shows that it is the only gryphaeate oyster present. In the two localities where better preserved material is present -Cerro Butaló and Ranquil-Có- there are also a few specimens with ribs which we illustrate here (figures 3.A-B and 4.F-G). The subspecific name Gryphaea burckhardti mendozana had been previously used by Ihering (1907, p. 36) for a different oyster, and that is the reason why Fritzsche's name had to be replaced (primary homonymy, ICZN Articles 57.2 and 60.3).
Fritzsche compared his very poorly preserved material with the specimens from the Supracrétacique du Roca illustrated by Burckhardt, 1901 (pl. 10) and placed under Gryphaea vesicularis Lamarck, 1806. He correctly assumed that it was not closely related to Lamarck's species, as this material has since been shown to belong in Pycnodonte ( Phygraea ) burckhardti (Böhm, 1903) (Casadío, 1998, p. 4512-452, figs. 2.1-2.2, pl. 2, fig. 16-19). On the other hand, our material, as well as Fritzsche's, is identical to the specimen from Pircala (near Malargüe) illustrated by Burckhardt (1900, pl. 28, fig. 4) and also placed under Gryphaea vesicularis Lamarck, 1806.
Turkostrea damboreneae nom. nov. is very close to Turkostrea turkestanensis (Romanovsky, 1878) and Turkostrea afghanica (Vyalov, 1939), both from the Eocene in Central Asia. The Argentine species differs from them because it has an almost smooth left valve or else ornamented with fewer radial ribs. The specimens illustrated by Perrilliat and Vega (1993, pl. 1, figs. 2, 5; pl. 2, figs. 2, 3) and assigned to Ostrea ( Turkostrea ) strictiplicata (Raulin and Delbos, 1855) –a synonym of Ostrea turkestanensis Romanovsky- and Turkostrea escondida (Perrilliat and Vega, 1993) from the Eocene of Mexico can be easily separated from Turkostrea mendozana because the latter has fewer radial ribs. Turkostrea duvali (Gardner, 1927) from the Eocene of Texas and Nuevo León, can be distinguished because the North American species has more strongly developed chomata. The specimens illustrated by Berizzi Quarto di Palo (1970, pl. 27, fig. 3-5 and pl. 28, fig. 3 and 5) and assigned to Fatina ( Fatina ) beldersaiensis (Vyalov, 1939), collected in the Eocene of Afghanistan, are very similar to Turkostrea argentina n. sp. described below, which can also be separated from Turkostrea damboreneae by their more prominent umbos and larger number of radial ribs.
Localities. Cerro Butaló (69º40'W 35º50'S) and Ranquil-Có (69º30'W 36º12'S), Departament of Malargüe, Province of Mendoza.
Occurrence. Jagüel and Roca formations.
Age. Zone CC 26, late Maastrichtian (Parras et al ., 1998).
Etymology. Honouring S. Damborenea (Museo de La Plata) for her contribution to the knowledge of Mesozoic bivalves from Argentina.
Turkostrea argentina n. sp.
Figures 5.A-J, 6.A-G

Figure 5. A-J, Turkostrea argentina n. sp. A-B, bivalved shell, holotype / ejemplar bivalvado, holotipo, GHUNLPam 17244: A, right external view / vista externa derecha , B, left valve, external view / valva izquierda, vista externa ; C-J, paratypes / paratipos , C, left valve, internal view / valva izquierda, vista interna , GHUNLPam 17465; D-E, left valve / valva izquierda , GHUNLPam 17594: D, external view / vista externa , E, posterior view / vista posterior ; F-G, right valve / valva derecha , GHUNLPam 17438: F, external view / vista externa, G, internal view / vista interna ; H-I, left valve / valva izquierda , GHUNLPam 17481: H, external view / vista externa , I, internal view / vista interna ; J, left valve, internal view / valva izquierda , vista interna, GHUNLPam 17461. Scale bar 1 cm / escala gráfica 1 cm.

Figure 6. A-G, Turkostrea argentina n. sp., paratypes / paratipos . A-C, left valve / valva izquierda , GHUNLPam 17460: A, internal view / vista interna , B, external view / vista externa , C, anterior view / vista anterior ; D-E, right valve / valva derecha , GHUNLPam 17431: D, internal view / vista interna , E, external view / vista externa ; F, left valve, ventral margin broken, external view / valva izquierda, margen ventral roto , vista externa , GHUNLPam 17469; G, right valve, external view / valva derecha, vista externa , GHUNLPam 17428. Scale bar 1 cm / escala gráfica 1 cm.
Material. Holotype: GHUNLPam 17244, a closed specimen (length: 40, height: 52 mm) from Liu Malal; paratypes: one hundred and fifty five specimens, five of which are closed shells, sixty seven are left valves and eighty three are right valves. All come from Liu-Malal and are housed under collection numbers GHUNLPam 17243, 17245-17277, 17387-17498 and 17592-17601.
Diagnosis. Left valve with triangular ligament area, higher than long, ornamented with over 15 strong radial ribs; right valve with ligament area forming an angle larger than 90º to the commissure plane.
Description. Shell subtriangular, inequivalve. Left valve: convex, regularly arched between umbo and ventral margin, slightly inequilateral due to an expansion of the posteroventral area; anterior margin oblique to slightly convex; posterior margin concave dorsally and convex ventrally; attachment area very small; ligament area triangular, higher than long, curved in opisthogyrous spiral fashion; adductor muscle scar elongate, weakly reniform or with a very weakly convex dorsal margin; scar placed near the posterior and ventral shell margins; outer surface with commarginal growth lines and about 15 radial ribs stretching between the umbo and shell margin; some ribs with one or two bifurcations; crest of ribs rounded. Right valve: ovate, flat to slightly convex, and thinner than left valve; ligament area at an angle larger than 90º to commissure plane; Quenstedt muscle scar punctiform, placed near anterior margin; outer shell surface with numerous and weak commarginal lines; lath-like chomata (Malchus, 1990) along anterior and posterior margins, but continuous in some specimens; relict chomata (Stenzel, 1971, p. N1103) along posterior and anterior valve margins. Shell microstructure simple compact foliate to lenticular foliate with mocret chambers (the evolutionary trend from compact to foliate has been observed in Turkostrea species from Africa, Asia and Europe; Malchus, pers. com.).
Measurements. The average height for the left valve is 51 mm (range 24 to 76 mm); average length is 38 mm (range 20 to 57 mm); average thickness near valve margin is 5.3 mm (range 2 to 11 mm). The average height for the right valve is 42 mm (range 24 to 66 mm); average length is 33 mm (range 20 to 46 mm); average thickness near ventral margin is 3.8 mm (range 1 to 6 mm).
Discussion. This species resembles Turkostrea damboreneae nom. nov. in shape, size and thickness of the valves. However, they may be separated by the clearly different radial ornamentation and the more strongly developed chomata in the Danian species. The ribs in Turkostrea argentina n. sp. are more numerous, more strongly marked, covering the entire shell surface and have a sharper crest than in Turkostrea damboreneae nom. nov. They are also present in all available specimens, being evident even if these are fairly worn.
While there is no certainty as to the exact bed from where the specimen illustrated by Camacho (1979, fig. 3-4) comes, the fact that he mentions a pink limestone as the bearing rock suggests that it could have been collected in the Danian part of the section. His specimen is a right valve he called Ostrea lingua Camacho, 1968 (which is in fact a Gyrostrea , as stated by Casadío, 1998, p. 461-462, fig. 5.1-5.6), from the Maastrichtian beds exposed in Huantrai-Co (Neuquén). As the only specimen illustrated by Camacho is a right valve, a positive identification is quite impossible. Thus identity of Camacho's specimen with Turkostrea argentina n. sp is tentative only, as it may be possible that it comes from lower down in the section, where the Maastrichtian Turkostrea damboreneae also occurs.
Turkostrea argentina n. sp. is remarkably similar to Turkostrea beldersaiensis (Gorizdro, 1915) (Vyalov, 1965, pl. 3, fig. 1; Berizzi Quarto di Palo, 1970, pl. 27, fig. 3-5 and pl. 28, fig. 3 and 5). Both species share the same general outline, prominent umbos with a slight posterior expansion, and numerous radial ribs. In the Argentine species, however, these radial ribs are less numerous and the posterior expansion is somewhat better defined than in Turkostrea beldersaiensis .
Locality. Liu Malal (36º 45' S, 68º 43' W), Department of Malargüe, Province of Mendoza.
Occurrence. Roca Formation.
Age. NP1-NP2 Zone, early Danian (Concheyro and Villa, 1996; Pires et al ., 1999).
Etymology. After Argentina.
Genus Cubitostrea Sacco, 1897a
Emended diagnosis [modified from Stenzel, 1971, p. 1141]. Shell subelliptical (dorsoventrally elongated and slightly curved), subtriangular or sickle-shaped; right valve flat to slightly convex, generally flat on inside; left valve keeled; keel generally crescentic, placed nearer the posterior margin of shell; left valve with commarginal growth squamae and low to high, narrowly rounded, dichotomous or intercalating, long radial ribs separated by narrower interspaces; right valve with commarginal growth squamae; margins of left valve crenated by concavities under ribs, right valve margins smooth; chomata restricted to margins immediately adjacent to ligament.
Type species. Ostrea cubitus Deshayes, 1832, p. 365; original designation .
Remarks. Another member of the Turkostreini, Cubitostrea Sacco, 1897, is a genus widely spread in Paleogene rocks of North America, Europe and South America. While obviously related to Turkostrea and Sokolowia , it can be separated from them by its unique sharply triangular to sickle-shaped shell. Like all members of the tribe, it carries strong radial ribs on the left valve and, like in many of them, the left valve also shows a keel running from the umbo to the postero-ventral margin of the valve. In addition they have smooth convex right valves that only show growth lines. The right valve is invariably smaller than the left one; both are relatively thick and composed of compact-foliate to lenticular-foliate shell microstructure.
Stenzel (1971, p. N1141-N1143) believed that Turkostrea was closely allied to Ostrea Linné, 1758, and included it in the Ostreinae, a placement also followed by Vyalov (1984, p. 32). However, as pointed out above, Malchus (1990, p. 196) removed it from the Ostreinae and placed it within the Crassostreinae, proposing for them the tribe Turkostreini. While the exact taxonomic position of these oysters -and all oysters indeed- is prone to change as more material is studied based on a wide range of ecophenotypic variations (Machalski, 1998), the inclusion of Cubitostrea within the Turkostreini seems to be better grounded. Recent studies on larval shells performed by Malchus (1995) appear to confirm this.
The genus has been reported from rocks as early as Paleocene, which was supposed as the moment in which these oysters possibly arose from a member of the Flemingostreini or Ambigostreini (Malchus, 1990, p. 197). However, the new species described herein comes from Maastrichtian rocks (Zone CC 26, Perch-Nielsen, 1985), and is thus the earliest representative of the genus presently known. It appears that this taxon had a longer history in southern South America, as the last representative of this genus – Cubitostrea alvarezii (d'Orbigny, 1842)– lived along the coast of northern Patagonia and even further north in Entre Ríos until the late Miocene, where it forms vast accumulations in the Puerto Madryn Formation. The age of this unit has been subject to debate, but is generally accepted as late Miocene, and 87 Sr/ 86 Sr data more precisely indicate a Tortonian age for it (Scasso et al ., 2001). A slightly older non described species appears in the late Oligocene?-early Miocene Chenque Formation (Bellosi, 1995; Barreda and Palamarczuk, 2000), exposed in the area surrounding Comodoro Rivadavia. The paleogeographic significance of the long range of this genus in Patagonia remains yet to be properly assessed.
Cubitostrea primordialis n. sp.
Figures 7.A-E, 8.A-F

Figure 7. A-E, Cubitostrea primordialis n. sp., holotype / holotipo , GHUNLPam 17157: A, posterior view of conjoined valves / vista posterior de ambas valvas unidas ; B-C, left valve / valva izquierda , B, internal view / vista interna , C, external view / vista externa ; D-E, right valve / valva derecha , D, internal view / vista interna , E, external view / vista externa . Scale bar 1 cm / escala gráfica 1 cm .
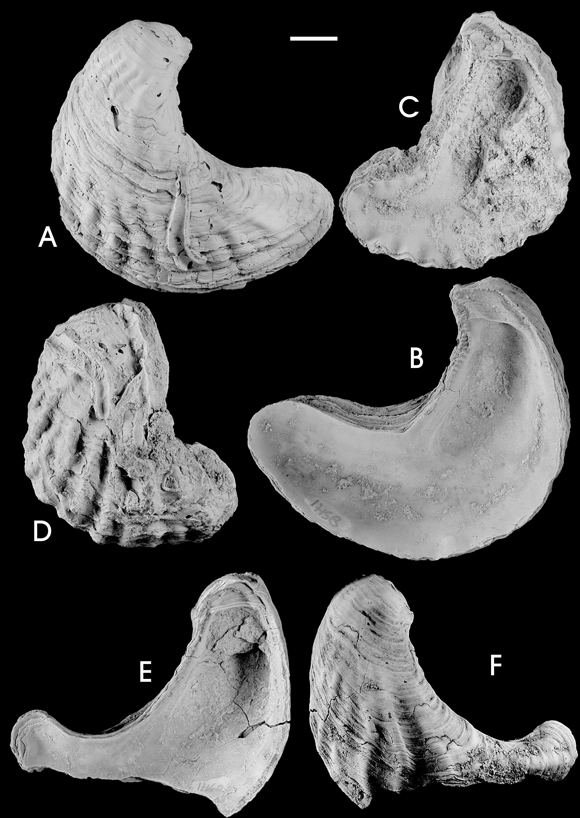
Figure 8. A-F, Cubitostrea primordialis n. sp., paratypes / paratipos. A-B, left valve / valva izquierda , GHUNLPam 17158: A, external view / vista externa , B, internal view / vista interna ; C-D, left valve / valva izquierda , GHUNLPam 17159: C, internal view / vista interna , D, external view / vista externa ; E-F, left valve / valva izquierda , GHUNLPam 17160: E, internal view / vista interna , F, external view / vista externa . Scale bar 1 cm / escala gráfica 1 cm .
Material. Holotype: GHUNLPam 17157, a bivalved specimen (length: 65 mm, height: 73 mm; measurements correspond to left valve) from Liu Malal; paratypes: five specimens, all left valves. All come from Liu Malal and are housed under the collection numbers GHUNLPam 17158-17161 and 17185.
Diagnosis. Outer surface of left valve with fewer than 15 weak, rounded and wavy radial ribs separated by shallow interspaces which are wider than the ribs.
Description. Shell sickle-shaped. Left valve: convex and larger than right valve; outer surface with fewer than 15 rounded and wavy radial ribs separated by shallow interspaces which are wider than ribs; adductor muscle scar reniform and placed very close to posterior shell margin and at half the distance between ligament and ventral margin; umbo opisthogyrous. Right Valve: flat to slightly concave; outer surface with fine commarginal lines; chomata restricted to dorsal third of anterior and posterior margins; relict chomata along posterior and anterior margins. Shell structure simple compact-foliate.
Measurements. Average height of the left valve is 62 mm (range 52 to 73 mm); average length is 54 mm (range 41 to 65 mm); average thickness of shell near ventral margin is 6 mm (range 5 to 7 mm). The only right valve available (GHUNLPam 17157) measures 61 mm high, 60 mm long and 6 mm thick near the ventral margin.
Discussion. Cubitostrea primordialis n. sp. is referred to Cubitostrea because of its sickle shape, its ornamentation, shape and position of the adductor muscle scar and opisthogyrous umbo. This species is very similar to Cubitostrea ameghinoi (Ihering, 1902) from the Danian beds outcropping throughout central and northern Patagonia (Ihering, 1902, p. 114-115, fig. 4-7;, p. 2, pl. 1, fig. 2-4; Ihering, 1903, p. 213; Ihering, 1907, p. 44; Wilckens, 1921, pl. 1, fig. 4a-b; Casadío, 1998, p. 458-461, pl. 2, fig. 8-15). They can be distinguished because the latter shows a larger number of radial ribs and more strongly developed chomata. Wilckens (1921, p. 12) should be credited for being the first to realize that these oysters were somehow related to Ostrea turkestanensis Romanovsky, 1878. As already discussed above, both Turkostrea and Cubitostrea are very close genera, sharing most characters except the tendency of Cubitostrea to be strongly triangular or else sickle-shaped with a strongly opisthogyrous umbo and not being a Gryphaea homeomorph.
The North American Cubitostrea sellaeformis (Conrad, 1832) from the Eocene of Alabama can be distinguished by the wide expansion of the posterodorsal area of the shell, a feature studied in detail by Stenzel (1949). Cubitostrea lisbonensis (Harris, 1919), from the middle Eocene of Alabama also shows auricles, which are not present in our material. The type species, Cubitostrea cubitus (Deshayes, 1832) from the Eocene of France and Cubitostrea mezquitalensis Squires and Demetrion, 1990, from the middle Eocene of Baja California Sur (Mexico), differ from Cubitostrea primordialis n. sp. in that they have a larger number of stronger radial ribs. Moreover, the type species is definitely more clearly triangular than ours. Cubitostrea wemmelensis Glibert 1936, from the Eocene of Belgium and Cubitostrea perplicata (Dall, 1898) from the middle Eocene of the southeastern United States also have more radial ribs and are less strongly curved than Cubitostrea primordialis n. sp.
Localities. Liu Malal (36º 45' S, 68º 43' W), Department of Malargüe, Province of Mendoza.
Occurrence. Jagüel Formation.
Age. Zone CC26, late Maastrichtian (Concheyro and Villa, 1996; Pires et al., 1999).
Etymology. From the Latin primordius (original).
Conclusions
The geographic and stratigraphic distribution of the oysters described above allows some interesting, although as yet preliminary, paleobiogeographic conclusions to be made. In the first place, as observed in figure 9, Turkostrea damboreneae nom. nov. And Cubitostrea primordialis n. sp. occur considerably earlier in southern South America than their close relatives from other parts of the world. Clear morphological differences in the shell show that by the late Maastrichtian they were already well established and that the origin of the Turkostreini should probably be sought for in slightly earlier rocks. Where exactly this could have taken place must necessarily be a matter of speculation, but some clues may be drawn from their presence in northern Patagonia in such comparatively old rocks. In fact, both genera are well known in younger rocks in other parts of the world, particularly the northern hemisphere. Cubitostrea seems to have been quite common throughout the Eocene in the Atlantic and Gulf coast of the United States, while occurring as well in the European Eocene. Turkostrea , although common in central Asia, from where it was originally described, is particularly abundant in the Eocene of northern Africa and Europe (Strougo, 1973, 1976, and references therein), while occurring also in Texas, Trinidad and Mexico according to Perrillat and Vega (1993), and possibly in Jamaica (Trechmann, 1923; Strougo, 1976). In fact, in northern Africa it is known from Paleocene rocks exposed in the Western Desert of Egypt, the age being confirmed by planktonic forams (Luger, 1985; Malchus, pers. com.). In South America, this genus has not been described previously, and the two species described here, although quite common in certain units and localities of the Neuquén Basin, have been largely overlooked. There can be no doubts as to the generic identity of our material and, in such a case, a southern origin for this group of oysters should be seriously considered.

Figure 9. Paleogeographic maps showing the worldwide biogeographic distribution of Turkostrea and Cubitostrea since the Maastrichtian / Mapas paleogeográficos indicando la distribución global de Turkostrea y Cubitostrea desde el Maastrichtiano.
In the case of Cubitostrea , the earliest recorded species of the genus appears also in Maastrichtian rocks in northern and central Patagonia. Cubitostrea primordialis n. sp. clearly belongs in that genus, becoming the oldest species presently identified. Like in the case of Turkostrea , this genus is also quite common in the Northern Hemisphere, particularly in Eocene rocks of Europe (Karagyuleva, 1961; Malchus, 1995) and the Gulf coast and eastern seaboard of the United States (Stenzel, 1949; Dockery and Nystrom, 1992; Willoughby and Nystrom, 1992). This genus shares many characters with Turkostrea and was therefore placed in the Turkostreini by Malchus (1990, p. 196). It seems likely that one of the two genera is ancestral to the other or else that they share a common ancestor not too far back in the Cretaceous. Cubitostrea primordialis n. sp. is, again, the earliest record of Cubitostrea .
While common in Paleocene rocks of Patagonia and western Argentina (Casadío, 1998, and references therein), Cubitostrea seems to be absent in Eocene and Oligocene rocks here. This apparent absence is probably a consequence of various concurrent factors. In the first place, the Eocene is very scantily represented in Argentina. The only reasonably good exposures are those of the Río Turbio and Man Aike Formations, in addition to the poorly fossiliferous Leticia Formation exposed off the coast of Tierra del Fuego. In none of the three units has material referable to Cubitostrea -or Turkostrea - been collected so far, despite the abundant mollusk fauna (including oysters) in some of them. A second cause for the lack of these two genera in Eocene or Oligocene rocks may be the particular ecological requirements that they may have had and about which we can only speculate. The third reason - i.e ., the actual disappearance of Cubitostrea in southern latitudes at the close of the Paleocene while surviving in younger rocks elsewhere- seems likely as well were it not for the fact that species of Cubitostrea , one of them very similar to the Paleocene species, appear again in the widespread early and late Miocene rocks exposed throughout Patagonia and central Argentina (Entre Ríos). In addition to a non described Cubitostrea from the early Miocene Chenque Formation (Bellosi, 1995) in the area surrounding Comodoro Rivadavia (central Chubut), Cubitostrea alvarezii (d'Orbigny, 1842) (d'Orbigny, 1842, p. 134, pl. 7, fig. 19; other references) appears in the late Miocene Puerto Madryn Formation exposed in the area around Peninsula Valdés in northern Patagonia and in the equivalent Paraná Formation in western Entre Ríos (del Río and Martínez, 1998 and references therein). It is also known from several drillings on the Pampas of central Argentina (Wahnish, 1939; Carral Tolosa, 1942; Camacho, 1967). These Miocene occurrences, as young as Tortonian for Cubitostrea alvarezii (Scasso et al ., 2001), are the youngest for any member of this genus in South America. Thus, the time range of Cubitostrea in South America spans the Maastrichtian through the Miocene, albeit the absence of records in the Eocene-Oligocene. Consequently, in this region the Turkostreini appear to have survived at least up to the late Miocene. Malchus (1995, p. 230) stated that Cubitostrea ? latimarginata (Vredenburg, 1908) from Asia and Saudi Arabia and Cubitostrea ? coxi (Gardner, 1945) from Florida were two Miocene species that could be placed in this genus. He also suggested the possibility that the Cubitostrea lineage may be extant in the Indo-West Pacific and Mediterranean.
Summarizing, the available data point towards a southern origin for Cubitostrea and Turkostrea , a fact that would render South America the place or origin of the entire tribe Turkostreini. The presence and abundance of members of this taxon in younger seas of the northern hemisphere must thus be accounted for. In connection with this, the Tethyan affinities of other marine groups during the Maastrichtian-Danian have been documented (Camacho, 1992; Feldmann et al ., 1995; Casadío, 1998). Such affinities, revealed in this case by the oysters of this particular group, are reinforced by the fact that oysters belonging in the Turkostreini have not been recorded from either Maastrichtian, Danian or Eocene (or younger) age in other areas of the southern hemisphere. This is particularly striking in the case of the extremely abundant and well preserved Maastrichtian-Eocene mollusk faunas from Marambio (Seymour) Island in Antarctica (Sharman and Newton, 1894, 1897; Wilckens, 1910, 1911; Zinsmeister, 1984; Zinsmeister and Macellari, 1988; Stilwell and Zinsmeister, 1992) -and also in the case of the Eocene fauna from MacMurdo described by Stilwell (2000)- which have yielded not one specimen that could be even tentatively placed either in Turkostrea or Cubitostrea . Turkostrea also appears to be absent from the very diverse and well known New Zealand Tertiary faunas, which share some particular groups in rocks ranging from the pre Cenozoic to the Pliocene and in some instances share taxa of the present day faunas (Beu and Griffin, 1997; Beu et al ., 1997). Cubitostrea is represented there by the Eocene Cubitostrea gudexi (Suter, 1917), but the generic placement of this species is doubtful, as it does not seem to be a true Cubitostrea and does not resemble the South American species at all.
If the two genera (or either one of them) appeared first in west-central Argentina, then the northward migration must be explained, of course considering the currently accepted patterns of landmass and current distribution in the Atlantic Ocean during the end of the Cretaceous and beginning of the Cenozoic. Such an explanation, as yet premature, would probably be of great heuristic value when dealing with other members of the shallow marine fauna inhabiting the southernmost tip of the American Continent.
Acknowledgments
This study was partially supported by Universidad Nacional de La Pampa and Fundación Antorchas to S.C. and CONICET to M.G. We also thank N. Malchus and M. Machalski for constructive comments. M. Sander kindly allowed access to Fritzsche's material housed in the Goldfuß-Museum, Bonn. A.C. Riccardi granted access to the material housed in the Museo de La Plata, La Plata, Argentina.
References
Barreda, V. and Palamarczuck, S. 2000. Estudio palinoestratigráfico del Oligoceno tardío - Mioceno en secciones de la costa patagónica y plataforma continental argentina. INSUGEO, Serie Correlación Geológica 14: 103-138. [ Links ]
Barrio, C.A. 1990a. Paleogeographic control of Upper Cretaceous tidal deposits, Neuquén Basin, Argentina. Journal of South America Earth Sciences 3: 1-19. [ Links ]
Barrio, C.A. 1990b. Late Cretaceous-Early Tertiary sedimentation in a semi-arid foreland basin (Neuquén Basin, western Argentina). Sedimentary Geology 66: 255-275. [ Links ]
Barrio, C.A. 1991. Controles en la sedimentación de cuencas foreland. El ejemplo del Grupo Malargüe (Campaniano-Paleoceno) en la cuenca Neuquina, Argentina. 6º Congreso Geológico Chileno (Santiago de Chile, 1991), Resúmenes expandidos 1: 597-601. [ Links ]
Bellosi, E.S. 1995. Paleogeografía y cambios ambientales de la Patagonia Central durante el Terciario medio. Boletín de Informaciones Pertroleras 1995: 51-83. [ Links ]
Berizzi Quarto di Palo, A. 1970. Paleogene Pelecypods from Kataghan and Badakhshan. In: Desi, A. (ed.), Scientific Reports, Italian Expeditions to the Karakorum (K2) and Hindu Kush, IV Paleontology-Zoology-Botany, Volume 2, Fossils of North-East Afghanistan, Cretaceous and Paleogene Fossils (by A. Berizzi Quarto di Palo and I. Premoli Silva), pp. 161-240 + 292-323. [ Links ]
Beu, A.G. and Griffin, M. 1997. The South American-Australasian connection in Oligocene-Neogene Mollusca. In: S. Nishimura, P. Utomo, and R. Tsuchi (eds.), Proceedings, Sixth International Congress on Pacific Neogene Stratigraphy and IGCP-355 : Neogene Evolution of the Pacific Ocean: Biotic, Oceanographic and Tectonic Development (Serpong, West Java, Indonesia, 1995), Kyoto Institute of Natural History, Kyoto, pp. 154-167. [ Links ]
Beu, A.G., Griffin, M. and Maxwell, P.A. 1997. Opening of Drake Passage gateway and Late Miocene to Pleistocene cooling reflected in Southern Ocean molluscan dispersal: evidence from New Zealand and Argentina. Tectonophysics 281: 83-97. [ Links ]
Böhm, J. 1903. Über Ostreen von General Roca am Rio Negro. Zeitschrift der Deutschen geologischen Gesellschaft 55: 71-72. [ Links ]
Böhm, J. 1933. Die palaeogene Fauna Ost-Turkestans. Zeitschrift der Deutschen geologischen Gesellschaft 85: 99-118. [ Links ]
Burckhardt, C. 1900. Profils géologiques transversaux de la cordillère Argenino-Chilienne. Stratigraphie et tectonique. Anales del Museo de La Plata, sección Geológica y Mineralógica II, 136 pp. [ Links ]
Burckhardt, C. 1901. Le gisement supracrétacique de Roca (Rio Negro). Revista del Museo de La Plata 10: 207-220. [ Links ]
Camacho, H.H. 1967 [1966]. Fascículo III, (Invertebrados). In: Borrello, A.V. (ed.), Paleontographia bonaerense , pp. I-XXV, 1-159, pl. I-XIX, Comisión de Investigaciones Científicas, La Plata. [ Links ]
Camacho, H.H. 1968. Acerca de la megafauna del Cretácico Superior de Huantraico, Provincia del Neuquén (Argentina). Ameghiniana 5: 321-332. [ Links ]
Camacho, H.H. 1979. Nuevos fósiles del Cretácico Superior (Maestrichtiano) del departamento Malargüe, provincia de Mendoza, Argentina. Ameghiniana 7: 329-334. [ Links ]
Camacho, H.H. 1992. Algunas consideraciones acerca de la transgresión marina paleocena en la Argentina. Academia Nacional de Ciencias en Córdoba , Miscelánea 85: 1-41. [ Links ]
Carral Tolosa, E.W. de 1942. Nota sobre los moluscos procedentes de la perforación de Monte Veloz (Buenos Aires). Anales del Primer Congreso Panamericano de Minas y Geología, Geología 2: 572-579. [ Links ]
Casadío, S. 1994. [ Estratigrafía y paleontología del intervalo Maastrichtiano-Daniano en el occidente de la provincia de La Pampa, Argentina. Tesis doctoral, Universidad Nacional de Córdoba, 420 pp. Unpublished] [ Links ]
Casadío, S. 1998. Las ostras del límite Cretácico-Paleógeno de la cuenca Neuquina (Argentina). Su importancia bioestratigráfica y paleobiogeográfica. Ameghiniana 35: 449-472. [ Links ]
Casadío, S. 1999. Variaciones climáticas durante el Paleógeno en el extremo austral de América del Sur y la Península Antártica. 7º Jornadas Pampeanas de Ciencias Naturales (Santa Rosa, 1999), Actas 1: 19-25. [ Links ]
Casadío, S., Parras, A., Concheyro, A. and Pires, M. 1998. El diacronismo de la sedimentación carbonática en la cuenca Neuquina durante la transición Cretácico-Paleógeno. 7º Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca, 1998), Resúmenes 1: 125. [ Links ]
Casadío, S., Rodríguez, M.F., Reichler, V.A. and Camacho, H.H. 1999. Tertiary nautiloids from Patagonia, southern Argentina. Ameghiniana 36: 182-202. [ Links ]
Concheyro, A. and Villa, G. 1996. Maestrichtian-Danian (K/P) calcareous nannofossils in the Liu Malal section. Northern Patagonia, Argentina. Palaeopelagos 6: 281-297. [ Links ]
Conrad, T. A. 1832. Fossil Shells from the Tertiary Formations of North America, Illustrated by Figures Drawn on Stone, from Nature , volume 1, number 2, Judah Dobson, Philadelphia, pp. 21-28, pl. 7-14. [ Links ]
Coquand, H. 1869. Monographie du Genre Ostrea, Terrain Crétacé , Paris, 215 pp., atlas, 74 pls. [ Links ]
Dall, W.H. 1898. Contributions to the Tertiary fauna of Florida, with special reference to the Silex beds of Tampa and the Pliocene beds of the Caloosahatchie River including in many cases a complete revision of the generic groups treated and their American Tertiary species. Transactions of the Wagner Free Institute of Science 3: 571-947. [ Links ]
del Río, C.A. and Martínez, S. 1998. Moluscos Miocenos de la Argentina y del Uruguay. Monografías de la Academia Nacional de Ciencias Exactas, Físicas y Naturales 15: 1-151. [ Links ]
Deshayes, G.P. 1832. Description des Coquilles Fossiles des Environs de Paris, volume 1:147-290, Chez l'auteur and others, Paris. [ Links ]
Dockery, D.T. and Nystrom, P.G. 1992. The Orangeburg District molluscan fauna of the McBean Formation: a new, diverse, silicified fauna within the Glyptoactis (Claibornicardia) alticostata zone above the occurrence of Cubitostrea sellaeformis , an interval zone of Gosport age. In: V.A. Zullo, W.B. Harris and V. Price (eds.), Proceedings of the Second Bald Head Island Conference on Coastal Plains Geology , The University of North Carolina, pp. 90-94. [ Links ]
d'Orbigny, A. 1842. Voyage dans l'Amérique Méridionale (le Brésil, la République orientale de l'Uruguay, le République Argentine, la Patagonie, le République du Chili,la République de Bolivia, la République du Perou), executé pendant les annés 1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833. Volume 3, part 4, 561 pp., P. Bertrand, Paris [ Links ]
Feldmann, R.M., Casadío, S., Chirino-Gálvez, L. and Aguirre-Urreta, M.B. 1995. Fossil decapod crustaceans from the Jaguel and Roca Formations (Maastrichtian-Danian) of the Neuquén Basin, Argentina. Journal of Paleontology 69, 2: 1-22. [ Links ]
Fritzsche, H. 1919. Eine Fauna aus schichten der Kreide-Tertiärgrenze in der argentinischen Cordillere des südlichen Mendoza. Zentralblatt für Mineralogie, Geologie und Paläontologie 1919: 359-369. [ Links ]
Gardner, J. 1927. New species of mollusks from the Eocene of Texas. Journal of the Washington Academy of Sciences 17: 362-383. [ Links ]
Gardner, J. 1945. Three new species from an Upper Miocene Oyster Reef in Tampa Bay. The Nautilus 59: 37-41. [ Links ]
Glibert, M. 1936. Faune malacologique des Sables de Wemmel, I, Pélécypodes. Musée Royale Histoire Naturelle Belgique, Mémoires 78, 242 pp. [ Links ]
Gorizdro, Z.F. 1915. Materiali k izucheniyu fauny trtichnikh otlozheniy Turkestana. Travaux de la Société Impériale de Naturalistes de Saint Petersbourg, Géologie et Minéralogie 37: 1-56. [ Links ]
Grewink, C. 1853. Die geognostischen und geographischen Verhältnisse des nordlichen Persiens. Verhandlungen der Russisch-Kaiserlichen mineralogischen Gesellschaft zu St. Petersburg : 97-245. [ Links ]
Harris, G.D. 1919. Pelecypoda of the St. Maurice and Claiborne Stages. Bulletins of American Paleontology 6: 1-269. [ Links ]
Ihering, H. von 1902. Historia de las ostras argentinas. Anales del Museo Nacional de Buenos Aires 7: 109-123. [ Links ]
Ihering, H. von 1903. Les mollusques des terrains crétaciques superieurs de l'Argentine orientale. Anales del Museo Nacional de Buenos Aires , serie 3 , 2: 193-229. [ Links ]
Ihering, H. von 1907. Les mollusques fossiles du Tertiaire et du Crétacé superieur de l'Argentine. Anales del Museo Nacional de Buenos Aires, serie 3, 7: 1-611. [ Links ]
Karagyuleva, Y.D. 1961. Predstaviteli na podrod Cubitostrea ot Priobona v Burgasko. Trudove vaeurkhu geologiëiìata na Baeulgariëiìa, Seriëiìa Paleontologiëiìa 3: 237-247. [ Links ]
Lamarck, J.P. de 1806. Mémoire sur les fossiles des environs de Paris. Annales du Muséum d`Histoire naturelle de Paris 8: 156-166. [ Links ]
Legarreta, L. Kokogián, D.A., and Boggetti, D.A. 1989. Depositional sequences of the Malargüe Group (Upper Cretaceous-Lower Tertiary), Neuquén Basin, Argentina. Cretaceous Research 10: 337-356. [ Links ]
Linné, K. von 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. Tomus I iv + 824 pp., Laurentii Salvii, Stockholm. [ Links ]
Luger, P. 1985. Stratigraphie der marinen Oberkreide und des Alttertiärs im südwestlichen Obernil-Becken (Süd Ägyptien) untere besonderer Berüchtigung der Mikropaläontologie, Palökologie und Paläogeographie. Berliner geowissenschaftliche Abhandlugen A 63: 1-153. [ Links ]
Macellari, C.E. 1988. Cretaceous paleogeography and depositional cycles of western South America. Journal South American Earth Sciences 1: 373-418. [ Links ]
Machalski, M. 1998. Oyster life positions and shell beds from the Upper Jurassic of Poland. Acta Paleontologica Polonica 43: 609-634. [ Links ]
Malchus, N. 1990. Revision der Kreide-Austern (Bivalvia: Pteriomorphia) Ägyptens (Biostratigraphie, Systematik). Berliner Geowissentschaftliche Abhandlungen , Reihe A, Band 125, pp. 1-231. [ Links ]
Malchus, N. 1995. Larval shells of Tertiary Cubitostrea Sacco, 1897b, with a review of larval shell characters in the subfamilies Ostreinae and Crassostreinae (Ostreoidea, Bivalvia). Bulletin de l'Institut royal des Sciences naturelles de Belgique, série Sciences de la Terre 64: 109-149. [ Links ]
Malumián, N. 1999. La sedimentación y el volcanismo terciarios en la Patagonia extraandina. In: R. Caminos (ed.), Geología Argentina, Anales del Instituto de Geología y Recursos Minerales 29: 557-612. [ Links ]
Malumián, N. and Caramés, A. 1995. El Daniano marino de Patagonia (Argentina): paleobiogeografía de los foraminíferos bentónicos. In: C. Náñez (ed.), Paleógeno de América del Sur, Publicación Especial de la Asociación Paleontológica Argentina 3: 83-105. [ Links ]
Parras, A. 1999. [ Estratigrafía y Paleontología del Grupo Malargüe (Límite Cretácico-Paleógeno), en la Provincia de Mendoza. Tesis Doctoral Universidad Nacional de Córdoba, 323 pp. Unpublished.] [ Links ]
Parras, A. and Casadío, S. 1999. Paleogeografía del sector septentrional de la cuenca Neuquina, durante el intervalo Campaniano-Daniano. 7º Jornadas Pampeanas de Ciencias Naturales (Santa Rosa, 1999), pp. 261-268. [ Links ]
Parras, A., Casadío, S. and Pires, M. 1998. Secuencias depositacionales del Grupo Malargüe (límite Cretácico-Paleógeno), sur de la provincia de Mendoza, Argentina. In: S. Casadío (ed.), Paleógeno de América del Sur y de la Península Antártica, Publicación Especial de la Asociación Paleontológica Argentina 5: 181-19. [ Links ]
Pascual, R. 1998. The history of South American land mammals: the seminal Cretaceous-Paleocene transition. In: S. Casadío (ed.), Paleógeno de América del Sur y de la Península Antártica, Publicación Especial de la Asociación Paleontológica Argentina 5: 9-18. [ Links ]
Pávay, A. von 1873. Die geologischen Verhältnisse der Ungebung von Klausenburg. Jahresbericht der Königlich Ungarischen geologischen Anstalt 1: 351-441. [ Links ]
Perch-Nielsen, K. 1985. Cenozoic calcareous nannofossils. In: H.M. Bolli, J.B. Saunders and K. Perch-Nielsen (eds.), Plankton Stratigraphy . Cambridge University Press, pp. 427-554. [ Links ]
Perrillat, M. C. and Vega, F. 1993. Early Eocene ostreids from the Adjuntas Formation, Difunta Group, northeastern Mexico. Tulane Studies in Geology and Paleontology 26: 15-25. [ Links ]
Pires, M., Náñez, C., Casadío, S. and Parras, A. 1999. Sedimentología, paleontología y estratigrafía del límite Cretácico-Paleógeno en Liu Malal, Mendoza. 14º Congreso Geológico Argentino, (Salta, 1999), Actas 1: 463-466. [ Links ]
Rafinesque, C.S. 1815. Analyse de la nature ou tableau de l'Univers et des corps organisés, etc. , Palermo, 224 pp. [ Links ]
Raulin, V. and Delbos, J. 1855. Extrait d'une monographie des Ostrea des terrains tertiaires de l'Aquitaine. Bulletin de la Societé Géologique de France, série 2 , 12: 1144-1164. [ Links ]
Romanovsky, G.D. 1878-1890. Materialy dlya geologiy Turkestanskago kraya [Materials for the geology of the Turkestanian region], Academie Impériale des Sciences, St. Petersbourg, vol. 1, viii+167pp., 30 pls. (1878) ; vol. 2, xii+161 pp., 27 pls.(1884) ; vol. 3, x+165 pp., 23 pls. (1890), Saint Petersburg [in Russian]. [ Links ]
Sacco, F. 1897a. I molluschi dei terreni terziarii del Piemonte e della Liguria. Musei Zoologia e Anatomia Comparata, Realle Università de Torino, Bolletino 12(298): 99-102. [ Links ]
Sacco, F. 1897b. Pelecypoda (Ostreidae, Anomiidae e Dimyidae). In: Bellardi, L. and Sacco, F., 1872-1904, I molluschi dei terreni terziarii de Piemote e della Liguria, part 23 (June, 1897), pp. 1-66, Carlo Clausen, Torino. [ Links ]
Scarlato, O.A. and Starobogatov, Ya. L. 1979. Osnovny cherty evoliutsii i sistema klassa Bivalvia. In: Ya. L. Starobogatov (ed.), Morfologiia, sistematika i filogeniia molliuskov, Akademiya NAUK SSSR , Trudy zoologicheskogo Instituta, Leningrad, 80: 5-38 [in Russian]. [ Links ]
Scasso, R.A., McArthur, J.M., del Río, C.J., Martínez, S. And Thirlwall, M.F. 2001. 87 Sr/ 86 Sr Late Miocene age of fossil molluscs in the Entrerriense of the Valdés Peninsula (Chubut, Argentina). Journal of South American Earth Sciences 14: 319-329. [ Links ]
Sharman, G. and Newton, E.T. 1894. Notes on some fossils from Seymour Island, in the Antarctic regions obtained by Dr. Donald. Transactions Royal Society Edinburgh 37: 707-709 [ Links ]
Sharman, G. and Newton, E.T. 1897. Notes on some additional fossils collected at Seymour island ; Graham Land, by Dr. Donald and Captain Larsen. Proceedings Royal Society Edinburgh 22: 58-61. [ Links ]
Squires, R.L. and Demetrion, R. 1990. New Eocene marine bivalves from Baja California Sur, Mexico. Journal of Paleontology 64: 382-391. [ Links ]
Stenzel, H.B. 1949. Successional speciation in paleontology: the case of the oysters of the sellaeformis stock. Evolution 3: 34-50. [ Links ]
Stenzel, H.B. 1971. In: Moore, R.C. (ed.). Treatise on Invertebrate Paleontology, Part N, volume 3 (Oysters), p. N953-N1224. [ Links ]
Stilwell, J.D. 2000. Eocene Mollusca (Bivalvia, Gastropoda and Scaphopoda) from McMurdo Sound: systematics and paleoecologic signicance. In: Stilwell, J. D. and Feldmann, R. (eds.), Paleobiology and Paleoenvironments of Eocene Rocks, McMurdo Sound, East Antarctica. Antarctic Research Series 76: 261-320. [ Links ]
Stilwell, D.A. and Zinsmeister, W.J. 1992. Molluscan Systematics and Biostratigraphy - Lower Tertiary La Meseta Formation, Seymour Island, Antarctic Peninsula. Antarctic Research Series 55, I-xii+1192 [ Links ]
Strougo, A. 1973. Observations sur les Huîtres du groupe multicostata de l'Eocène supérieur d'Egypte. Proceedings Egyptian Academy of Sciences 24: 135-139. [ Links ]
Strougo, A. 1976. Le groupe de Ostrea ( Turkostrea ) multicostata Deshayes, 1832. Géologie méditerranéenne 3: 27-44. [ Links ]
Suter, H. 1917. Descriptions of new Tertiary Mollusca occurring in New Zealand. Accompanied by a few notes on necessary changes in nomenclature. Part 1. New Zealand Geological Survey Palaeontological Bulletin 5: 1-93. [ Links ]
Torigoe, K. 1981. Oysters in Japan. Journal of Science of the Hiroshima University, series B, division 1 (Zoology) 29: 291-347. [ Links ]
Trechmann, C.T. 1923. The yellow limestone of Jamaica and its Mollusca. Geological Magazine 60: 337-367. [ Links ]
Tunik, M.A. 2001. [ Análisis Sedimentológico y Tectónico de la Primera Ingresión Atlántica en la Alta Cordillera de Mendoza. Tesis Doctoral, Universidad de Buenos Aires, 258 pp. Unpublished]. [ Links ]
Uliana, M.A. and Dellapé, D.A. 1981. Estratigrafía y evolución paleoambiental de la sucesión maestrichtiano-eoterciaria del engolfamieto neuquino. (Patagonia septentrional). 8º Congreso Geológico Argentino (San Luis, 1981), 3: 673-711. [ Links ]
Uliana, M.A. and Biddle, K.T. 1988. Mesozoic-Cenozoic paleogeographic and geodynamic evolution of southern South America. Revista Brasileira de Geociências 18: 172-190. [ Links ]
Urien, C.M., Zambrano, J.J. and Yrigoyen, M.R. 1995. Pretroleum basins of southern South America. In: A.J. Tankard, R. Suarez and H.J. Welsink (eds.), Petroleum Basins of South America, American Association of Petroleum Geologist, Memoir 62: 63-77. [ Links ]
Vredenburg, E.W. 1908. Preliminary note on the Indian occurrence of Ostrea multicostata , Deshayes, and other ribbed species of Ostrea . Records of the Geological Survey of India 36: 315-321. [ Links ]
Vyalov, O.S. 1936. Sur la classification des huîtres. Comptes rendus (Doklady) of the Academy of Sciences of the USSR, new series 4(13), nº 1(105): 17-20. [ Links ]
Vyalov, O.S. 1939 (1938). Zamechyania o srednieaziatskich Fatina Vyalov i Turkostrea Vyalov [Remarks on the middle-Asiatic Fatina Vyalov and Turkostrea Vyalov]. Isvestya Akademii Nauk SSSR, otdel matematicheskikh i estestvennykh nauk 1: 15-18. [In Russian] [ Links ]
Vyalov, O.S. 1948. Printsipii klassifikatsii semeystva Ostreidae [Principles of classification of the family Ostreidae]. Trudy lvovskogo geologicheskogo obschestva pri gosudarstviennom universytete im. Ivana Franko, paleontologicheskaya seria 1: 3-40. [In Russian] [ Links ]
Vyalov, O.S. 1965. Nekotorye paleogenovye ustritsi [Some Paleogene oysters]. Palaeontologicheskij Sbornik 2: 5-13. [In Russian]. [ Links ]
Vyalov, O.S. 1983. Zagalna klasifikatsiya ustrits [New general classification of oysters]. Doklady Akademii Nauk Ukkrainskoei SSR, Serieia B, Geologicheskie, Khimicheskie i Biologicheskie Nauki 1983(11): 6-8. [In Ukrainian]. [ Links ]
Vyalov, O.S. 1984. Klasifikatsiya ustrits [Classification of oysters]. Paleontologicheskij Sbornik 21, 31-36. [In Russian]. [ Links ]
Wahnish, E. 1939. Perforación Riachuelo Nº 5, Buenos Aires, perfil geológico y descripción de los fósiles. Revista del Centro de Estudiantes de Ciencias Naturales 2: 1141-156. [ Links ]
Wilckens, O. 1910. Die Anneliden, Bivalven, und Gastropoden der Antarktischen Kreideformation. Wissenschaftliche Ergebnisse der Schwedische Südpolar Expedition 3: 1-132. [ Links ]
Wilckens, O. 1911. Die Mollusken der Antarktischen Tertiärformation. Wissenschaftliche Ergebnisse der Schwedische Südpolar Expedition 3: 1-62. [ Links ]
Wilckens, O. 1921. Beiträge zur Geologie von Patagonien. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie Abhandlungen 1: 1-14. [ Links ]
Willoughby, R.H. and Nystrom, P.G. 1992. Oyster zonation in the Warley Hill Formation and Chapel Branch Member of the Santee Limestone at Cave Hall, Calhoun County, South Carolina and its regional implications. In: V. A. Zullo, W. B. Harris and V. Price (eds.), Proceedings of the Second Bald Head Island Conference on Coastal Plains Geology, pp. 121-126. [ Links ]
Zinsmeister, W.J. 1984. Late Eocene bivalves (Mollusca) from the La Meseta Formation, collected during the 1974-75 joint Argentine-American expedition to Seymour Island, Antarctic Peninsula. Journal of Paleontology 58: 1497-1527. [ Links ]
Zinsmeister, W.J. and Macellari, C. 1988. Bivalvia (Mollusca) from Seymour Island, Antarctic Peninsula. In: R.M. Feldmann and M. O. Woodburne (eds.), Geology and Paleontology of Seymour Island, Antarctic Peninsula. Memoir Geological Society of America 169: 253-284. [ Links ]
Recibido: 19 de abril de 2004.
Aceptado: 8 de julio de 2004.

