










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.2 Buenos Aires mar./jun. 2005
Patagonichnus : a new trace fossil from the Miocene of Patagonia. A probable fodinichnion of gregarious polychaetes
Eduardo B. Olivero 1 and María I. López Cabrera 1
1 Centro Austral de Investigaciones Científicas (CADIC-CONICET); Bernardo A. Houssay 200; V9410CAB Ushuaia, Tierra del Fuego, Argentina. emolivero@ciudad.com.ar
Abstract. Miocene tidal deposits from Las Grutas, Patagonia, Argentina, superbly preserve cylindrically laminated burrow systems including Asterosoma radiciforme von Otto and three compound ichnospecies of the newly proposed ichnogenus Patagonichnus ; P. calyciformis n. isp.; P. stratiformis n. isp.; and P . thalassiformis n. isp. Together with large, palmate burrows, probably produced by crustacean decapods, they are elite trace fossils. The Patagonichnus burrow systems consist of many distinctive concentrically laminated elementary components including: horizontal and vertical, conical and helicoidal bulbs and swellings; vertical shafts; straight and paired branches; and cone-in-cone structures. Hollow, branched, straight or helicoidal inner lined tubes connect these structures. According to current definitions, isolated components could be misidentified as Asterosoma von Otto, Rosselia Dahmer, Cylindrichnus Toots in Howard, Gyrolithes De Saporta, Helicodromites Berger, or Chondrites von Sternberg. The concentrically laminated bulbs of Patagonichnus represent the feeding behavior of a worm-like organism, spiraling or circling its anterior end within the sediment while its posterior end was temporarily fixed at the terminal position of the inner tube. Spiraling or circling movements, associated with peripheral backfilling and central migration of a probing tube, result in discrete, discordant groups of laminae. In cross-section, these laminae appear as alternate half-spirals, continuous spirals, or concentric circles, which represent the plan view of a helicoidal or conical whole structure. The complete structure is interpreted as a fodinichnion. The connecting, hollow inner tube is interpreted as a dwelling structure and the associated backfilled bulb as a feeding structure, probably made by gregarious polychaetes. Analogies in size, form, geometry, and behavior suggest polychaetes as the trace-makers of the Patagonichnus burrows.
Resumen. Patagonichnus : una nueva traza fósil del Mioceno de Patagonia. Probable fodinichnion de poliquetos gregarios . Las marealitas del Mioceno de Las Grutas, Patagonia, Argentina, preservan una excepcional asociación de trazas con laminación concéntrica que incluyen Asterosoma radiciforme von Otto y tres especies compuestas del nuevo icnogénero propuesto en este trabajo Patagonichnus ; P. calyciformis n. isp.; P. stratiformis n. isp.; y P. Thalassiformis n. isp. Junto a trazas palmadas de gran tamaño, probablemente producidas por crustáceos decápodos, constituyen trazas fósiles de élite. Patagonichnus tiene varias estructuras elementales distintivas con característica laminación concéntrica: bulbos y abultamientos horizontales y verticales, cónicos y helicoidales; columnas verticales; ramas rectas apareadas; y estructuras cono-en-cono. Todas estas estructuras están conectadas por tubos internos huecos, con pared, ramificados, rectos o helicoidales. En forma aislada estos componentes pueden ser confundidos con Asterosoma von Otto, Rosselia Dahmer, Cylindrichnus Toots in Howard, Gyrolithes De Saporta, Helicodromites Berger, o Chondrites von Sternberg. Los bulbos con laminación concéntrica representan la actividad de alimentación de organismos vermiformes, que con movimientos espirales o circulares de su porción anterior penetran en el sedimento, dejando su porción posterior transitoriamente fija en el tubo interno hueco. Los movimientos espirales o circulares, acompañados de retrorrelleno periférico y migración centrípeta, dan como resultado grupos discretos de láminas discordantes. En sección transversal, estos grupos de láminas se disponen formando semiespirales alternantes, espirales continuas, o círculos concéntricos, que representan la expresión en planta de una estructura espacial, con arreglo helicoidal o cónico. Etológicamente, estas estructuras se interpretan como fodinichnia. El tubo conectivo interno conformaría una estructura de habitación y los bulbos asociados una estructura de alimentación, probablemente producidas por organismos vermiformes gregarios. Las analogías de tamaño, forma, geometría y comportamiento, sugieren que los productores podrían ser poliquetos.
Key words. Trace fossils; Patagonichnus; Asterosoma; Miocene; Patagonia; Ethology; Polychaetes.
Palabras clave. Trazas fósiles; Patagonichnus; Asterosoma; Mioceno; Patagonia; Etología; Poliquetos.
Introduction
Single or partially preserved elements of concentrically laminated burrows, as exemplified by the ichnogenera Asterosoma, Rosselia, and Cylindrichnus, all share an apparently simple structural plan consisting of regular to irregular concentric laminae defining swollen bulb-like and/or cylindrical to conical structures, surrounding a central or eccentric lined tube. In general, interpreted ichnotaxobases include spatial orientation, form, and degree of connection of laminated structures, comprising mainly three basic plans: a) several connected bulbs, horizontal or inclined, with radial or branching fan-like arrangements ( Asterosoma ); b) mainly single, vertical or inclined, spindle-shaped or conical bulbs ( Rosselia ); and c) single, mainly inclined, cylindrical or conical forms ( Cylindrichnus ). In spite of this apparent simplicity -or perhaps because of this- the ichnotaxonomy and ethology of concentrically laminated burrows are problematic ( cf. Uchman and Krenmayr, 1995; Goldring, 1996; Schlirf, 2000; Bromley and Uchman, 2003). Seilacher (1969) and Chamberlain (1971) considered Rosselia a synonym or close relative of Asterosoma , but with different interpretations on the origin of the concentric lamination, which is interpreted either as the results of the radial pressure exerted internally by the trace maker (Seilacher written com., 2003; see also Nara, 1995) or of spiraling movements of the organism within the sediments while searching for food (Chamberlain (1971). Frey and Howard (1970; 1982; 1985) and McCarthy (1979) interpreted intergradational forms among Asterosoma , Rosselia and Cylindrichnus . Another problem is that in shallow marine and estuarine environments these ichnogenera are frequently associated and tend to occur in dense numbers (Uchman and Krenmayr, 1995; Nara, 1995, 2002; Schlirf, 2000; Bromley and Uchman, 2003, and references therein), and as a result in most cases it is difficult to understand or reconstruct a complete burrow structure. Moreover, the original definition of Asterosoma includes the mention of closely associated cylindrical branched tubes ( gabelnde Cylinderchen , von Otto, 1854), and a similar association is probably recognized as Asterosoma - Chondrites in the literature ( e.g . Müller, 1971; Schlirf, 2000, Pl. 3, Fig. 9).
Estuarine deposits in the Upper Cretaceous of Antarctica and the Eocene of Tierra del Fuego (Olivero and López C., 2001, 2002) bear distinct horizons fully bioturbated with concentrically laminated burrows, which may be loosely classified either as Asterosoma or Rosselia . They are frequently associated with Chondrites -like delicate branching tubes and partially preserved, conical to cylindrical, laminated shafts similar to Cylindrichnus . The general impression is that these trace fossils could be part of more complicated structures but the dense bioturbated fabric in homogeneous very fine sandstones forming the host rock, prevents the reconstruction of a complete burrow structure.
Luckily, a similar but superbly preserved ichnocoenosis was found in heterolithic tidal deposits of the Patagonia Formation (Early Miocene) at Las Grutas, Río Negro Province (figure 1). In this case, the preservation of whole burrow structures in friable mudstone and sandstone clearly shows that they could consist of large, complicated burrow systems composed of a variety of connected elementary components. Consequently, the new ichnotaxon Patagonichnus n. igen., including three new ichnospecies, is proposed for these complicated burrow systems.
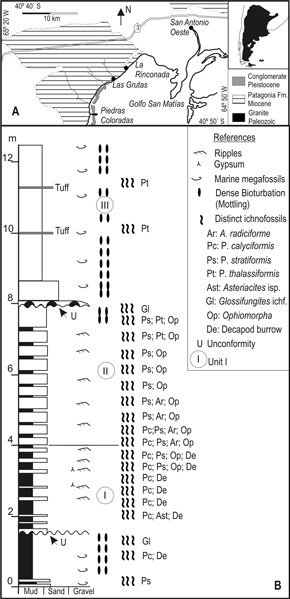
Figure 1. A, Locality map and schematic geology of Las Grutas. Adapted from Angulo et al . (1979)/ Mapa de ubicación y esquema geológico de Las Grutas. Adaptado de Angulo et al. (1979) ; B, Sedimentary log and trace fossil distribution in the Patagonia Formation between Las Grutas and La Rinconada / Perfil sedimentológico y distribución de trazas fósiles de la Formación Patagonia entre Las Grutas y La Rinconada.
The geometry of the Patagonichnus burrow system is variable and closely related to a particular granulometry of the substrate, but in general it consists of an intricate system of vertically and horizontally branched lined tubes surrounded by cylindrically laminated structures. Straight or helicoidal, vertical to horizontal short branches of this tube system are associated with bunches of vertical, horizontal or helicoidal, concentrically laminated bulbs or shafts. If the intricate connecting tube system is not recognized, the elementary bulbs or shafts could be classified by presently recognized ichnotaxobases in part as Asterosoma , Rosselia or Cylindrichnus . The horizontal to vertical helicoidal components of the inner tube could be misidentified as Helicodromites or Gyrolithes , respectively. Moreover, if isolated from bulbs, the branched inner tube could be misidentified with Chondrites . Thus, the main objectives of this paper are to describe and interpret the complicated Patagonichnus n. igen. burrow system from Las Grutas and to briefly discuss the probable biology and gregarious mode of life of the producers. In addition, an associated large, palmate, and concentrically laminated burrow, probably produced by crustacean decapods, is briefly described.
Methodology
Most of the described material was studied in situ during three field seasons in September 2002 and February and May 2003 and from field photographs. The deposits of the Patagonia Formation at Las Grutas are quite friable and the collection of large, relatively complete burrow systems was not possible. More resistant, elementary components were collected and rapidly hardened in the field with glue, and parts of large structures were cast in the field using silicone. In the lab positive molds of the silicone casts were made using plaster of Paris.
The diameter of the inner tube was carefully measured from amplified digital photographs in a PC screen. Part of the terminology concerning the geometry of the backfilling, such as the terms, spiral, helicoidal or conical, used in the systematic ichnology section, is fully explained later in a special section.
The material and molds are housed at the Centro Austral de Investigaciones Científicas (CADIC) under the numbers CADIC PI 21 to 37.
Geologic setting
The general stratigraphy at Las Grutas consists of limited exposures of Paleozoic granitoids at Piedras Coloradas, which are unconformably covered by more extended, shallow marine and tidal, tuffaceous mudstones, siltstones, and fine sandstones of the Patagonia Formation (Miocene). Just south of Las Grutas, a distinctive Pleistocene fossiliferous, hardcemented conglomerate is exposed in the intertidal zone (figure 1.A; Angulo et al ., 1979).
Our study is concentrated on the exposures of the Patagonia Formation along the sea cliff between Las Grutas and La Rinconada (figure 1.A). Here the Patagonia Formation consists of a fully bioturbated succession, ca . 12 m thick (figures 1.B, 2), which is very gently tilted (about 1°) to the NE. The oldest sediments are exposed to the SW, largely in the intertidal zone of the main beach area at Las Grutas, and consist of yellowish, densely bioturbated and mottled mudstones, with minor intercalated thin sandstone beds. Poorly preserved marine gastropods and bivalves are relatively common. They are overlain in sharp unconformity by reddish, bioturbated, heterolithic mudstones and sandstones with frequent ripple crosslamination, flaser bedding, mud drapes, and herringbone cross-stratification, informally referred here to units I and II. These are differentiated by the relative proportion and thickness of mudstones and sandstones; unit I is dominated by mudstones with alternations of thin sandstone beds and unit II is dominated by sandstones with thin mudstone beds (figures 1.B, 2). Some of the sandstone beds of unit II bear a dense concentration of fecal pellets.

Figure 2. The Patagonia Formation at the sea-cliff between Las Grutas and La Rinconada showing the white, tuffaceous, sandy siltstones of unit III (U III), the reddish unit II (U II) with inclined heterolithic stratification, and the intervening unconformity (arrow) / La Formación Patagonia en el acantilado entre Las Grutas y La Rinconada, se observan limolitas arenosas tobáceas de la unidad III (U III) y las capas rojizas de la unidad II (U II) con estratificación heterolítica inclinada, en contacto discordante (flecha).
Part of the bedding in the reddish, heterolithic succession of units I and II is at an angle with respect to the lower and upper strata and represents inclined heterolithic stratification. Unit II is sharply cut in angular relationship by horizontal, whitish beds of unit III, and this angular contact is best seen at the sea cliff just SW of La Rinconada. Unit III consists of a basal, lenticular conglomerate, and sandstone and siltstone beds of contrasting white color (figures 1.B, 2). They are highly bioturbated and record relatively abundant but poorly preserved serpulid tubes, mollusks, bryozoans, and crabs. The unconformities at the base of unit I and unit III are associated with typical components of the Glossifungites ichnofacies (figures 1.B, 2, cf . Frey and Seilacher, 1980), including galleries of Thalassinoides isp.; U-tubes of Diplocraterion isp.; Gastrochaenolites isp.; and distinctive burrows that cut across a dense, mottled bioturbated structure with sharp, neat boundaries indicating that they were excavated in a firm, semiconsolidated substrate.
On the basis of the features described above, the heterolithic beds of units I and II are interpreted as tidal flats and migrating intertidal channel deposits. The unconformity at the top of unit II probably represents a ravinement surface of erosion, covered by shallow marine, fossiliferous sandy siltstones.
Cylindrically laminated burrows are particularly abundant in the reddish, heterolithic deposits. Several, distinct burrow systems are recognized in this paper, and their occurrence is closely tied to a particular grain size of the host sediment (figure 1.B). The mud-dominated unit I bears abundant decapod crustacean trace fossils, Asterosoma radiciforme von Otto, Patagonichnus calyciformis n. isp., and rare Asteriacites isp. The sandier unit II is dominated by Asterosoma radiciforme and Patagonichnus stratiformis n. isp., with some Patagonichnus thalassiformis n. isp. at its top. Large, vertical tubes of Ophiomorpha nodosa , most of them passively filled with concentric laminae, are present in both units I and II. In addition to a characteristic dense, mottled structure, unit III records two levels with dense concentration of Patagonichnus thalassiformis n. isp. in sandy siltstones and tuffs (figure 1.B).
Systematic ichnology
Ichnogenus Asterosoma von Otto 1854
Type ichnospecies. Asterosoma radiciforme von Otto 1854. Original figures are reproduced in part by Häntzschel (1975, fig. 25 1a).
Remarks. The original diagnosis of Asterosoma von Otto (1854) was mostly based on the star-like arrangement of bulbs, typical for A. Radiciforme (Häntzschel, 1975). Subsequently, the ichnogeneric diagnosis was expanded to include the dichotomous to fan-like pattern of branching bulbs typical for A. ludwigae Schlirf (2000). Seilacher (writ. comm., 2003) considers that the radial pushing exerted internally by the tracemaker produces both the concentric lamination and the external longitudinal furrows and striae -interpreted as microfaults- commonly preserved in the German type material. In our material, there is limited evidence for this radial pushing mechanism (see Discussion).
Asterosoma radiciforme von Otto 1854
Figures 3.A4, 4.A-D, 5.A-F
A. radiciforme von Otto, 1854, in Häntzschel (1975, Fig. 25 1a)
A. radiciforme von Otto, Müller, 1971, Pls. 1-3
Material. Many field-specimens, and part of several bulbs, CADIC PI 21 to 26.
Description. Large, concentrically laminated, subhorizontal to inclined bulbs with a star-like arrangement radiating from a central point. The concentric structure of bulbs consists of thick mud-rich laminae alternating with thin sandy laminae, surrounding a central mud-lined tube. The internal structure of bulbs frequently consists of alternated half-spirals (figures 5.A, B, D, E). Bulbs egg-shaped, 10-15 cm long and 3-5 cm wide, connected by an inner lined tube to a central point that continues into a vertical, lined tube encircled by concentric lamination (figure 5.F). Inner tube ranges from 5 to 10 mm (average 7 mm) in width.
Preservation. Endichnial; mud-rich bulbs, commonly present in thin alternations of sandstone and mudstone beds.
Remarks. The construction of the burrow structure in A. radiciforme was interpreted to proceed in a downward direction (Uchman and Krenmayr, 1995). In addition, our material indicates that the tracemaker frequently followed an upward direction during the construction of the burrow as it is indicated by the upward displacement of mud laminae at the contact between mudstone-sandstone beds (figures 5.A-B) and by the conical-concentric backfilling in the central shaft (figure 5.F).
Localities. Relatively common in two or three levels near the top of unit I. Best examples are found between Bajada del ACA ( Bajada is a local name for the access to the beach) and just to the NE of Bajada Los Acantilados at Las Grutas.
Ichnogenus Patagonichnus n. igen.
Type ichnospecies. Patagonichnus stratiformis n. isp.
Derivatio nominis. Patagon -, from Patagonia; ichnus , trace.
Diagnosis. Large, compound, endichnial burrow system characterized by many distinctive concentrically laminated elementary components budding from an intricate system of horizontal to vertical connecting mud-lined tubes. Concentrically laminated elementary components may consist of vertical to horizontal spindle-shaped bulbs, bulb-like swellings, helicoidal bulbs, and backfilled tubular branches. The burrow system tends to cover large, irregular, horizontal layers. Vertical, concentrically laminated shafts or cone-in-cone structures surrounding a lined tube could connect different layers.
Remarks. Two basic taxonomic procedures have been applied to the nomenclature of compound trace fossils. In one case, the trace is named after the most common ichnotaxon or ichnotaxa ( e.g . Pickerill and Narbonne, 1995; Ekdale and Bromley, 2001). In the other case, a new name is given to the whole structure because it is thought that the distinct components, representing different but simultaneous behavioral activities of the same animal, contribute to an ichnological whole (Bromley et al ., 2003).
The complex and compound burrow system of Patagonichnus has many distinct, elementary components that in isolation could be assigned to several already established ichnogenera. In particular, the three newly proposed ichnospecies of Patagonichnus bear concentrically laminated bulbous structures that in isolation resemble the ichnogenus Asterosoma . Also, the constructional mechanism of the bulbs in Asterosoma radiciforme from Las Grutas is similar to that of some bulbs in the three new ichnospecies of Patagonichnus , suggesting that they represent the work of the same organism (see Discussion). However, the burrow system in Patagonichnus is much more complex than in A. radiciforme , or other Asterosoma ichnospecies characterized by discrete bunches of radiating bulbs ( cf . von Otto, 1854; Müller, 1971; Bromley and Uchman, 2003). The large Patagonichnus burrow system (figure 3) is characterized by complex, connected bulbous structures including series of linearly arranged, vertical calyx-like bulbs; sinuous, dense, horizontal branches with lateral bulb-like swellings; helicoidal bulbs; or aggregate, cylindrical structures with isolated or associated bulb-like swellings systematically associated with a variety of structures, which are interpreted as representing different, simultaneous behavioral activities of the same animal. Consequently, we followed the taxonomic procedure of Bromley et al. (2003) and a new name is given to the whole structure.
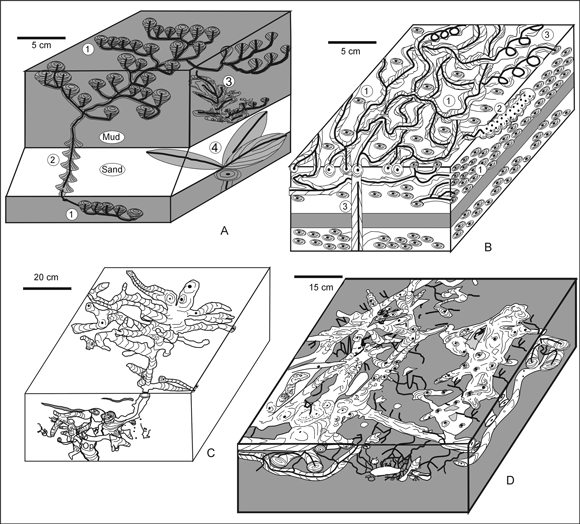
Figure 3. Reconstruction of the Patagonichnus and Asterosoma burrows from Las Grutas. Thick, black lines indicate the connecting, inner tube / Reconstrucción de las trazas de Patagonichnus y Asterosoma de Las Grutas. Las líneas gruesas negras indican los tubos internos conectados. A1-3, P. calyciformis n. isp. (1) linear array of bulbs developed in mud / bulbos alineados en fango , (2) cone-in-cone structures in sand / estructuras cono-en-cono en arena, (3) vertical imbricate bulbs connected to calyx-shaped bulbs / bulbos verticales imbricados conectados con bulbos en forma de cálice , (4) A . radiciforme , large bulbs / bulbos grandes ; B, P. stratiformis n. isp. two layers of sinuous branches with swellings (1) and associated helicoidal bulbs (2) developed in sandstone are connected by a vertical shaft (3) cutting across a mudstone layer / dos niveles con estructuras ramificadas sinuosas y con abultamientos (1) y bulbos helicoidales asociados (2) desarrollados en arenisca, conectados por una columna vertical (3) que corta a un nivel de fangolita; C-D, P. thalassiformis n. isp. paired backfilled branches and bulbs in sandstone (C) and sandy siltstone and tuff (D) , the last with networks of branched, lined tubes / galerías apareadas rellenas y bulbos en arenisca (C) y en limolita arenosa y toba (D) , la última con redes de tubos con pared ramificados .
Patagonichnus calyciformis n. isp.
Figures 3.A1-3, 4.F-I; 5.L-M
Type Material. Holotype: Partially preserved cup-like bulbs (CADIC PI 27, Figure 4.F); connecting cone-in-cone structures (CADIC PI 28, Figure 5.L); and parts of a linear array of bulbs (CADIC PI 29, figure 4.H). Paratypes: several isolated or welded bulbs (CADIC PI 30). Additional material: many field specimens.

Figure 4. A-D, A. radiciforme , field specimens, full relief, upper view / especímenes de campo, relieve completo, vista superior ; A, four bulbs radiating from a central point (arrow) / c uatro bulbos que irradian de un punto central (flecha) ; B, inclined bulbs and several horizontal crosssections of bulbs covered by a layer (arrow) full of P. stratiformis / bulbos inclinados y secciones horizontales de bulbos cubiertos por un nivel (flecha) lleno de P. stratiformis; C, concentrically laminated horizontal bulb with radial backfilling / bulbo horizontal con laminación concéntrica y retrorelleno radial ; D, cross-section of inclined bulbs with concentric laminae arranged in alternate half-spirals or in continuous spirals / sección transversal de bulbos inclinados con laminas concéntricas ordenadas en semiespirales alternantes o en espirales continuas ; E, P. stratiformis n. isp. full relief, lower view, several connected sinuous backfilled galleries surrounding an inner lined tube / relieve completo, vista inferior, varias galerías con retrorelleno, sinuosas y conectadas, que rodean un tubo interno con pared ; F-I, P. calyciformis n. isp. Full relief / relieve completo ; F, detailed upper and lateral views of two calyx-like bulbs showing external oblique cylindrical furrows (arrow), and irregular concentric laminae surrounding an inner lined tube, field photograph of the holotype CADIC PI 27 / vistas superior y lateral detalladas de dos bulbos caliciformes con surcos cilíndricos en la parte externa (flecha), y láminas concéntricas irregulares rodeando el tubo interno con pared, fotografía de campo del holotipo ; G, H, upper view of linear arrays of isolated or welded bulbs, cutting through the main tunnel of the large, palmate burrow (arrow in G), G field specimen, H CADIC PI 29 pars. / vista superior de arreglos lineales de bulbos aislados o soldados, que cortan el tunel principal de la traza palmada de gran tamaño (flecha en G), G espécimen de campo y H CADIC PI 29 pars. I, lateral view of two cone-in-cone structures developed in sandstone, field specimens / vista lateral de dos estructuras cono en cono desarrolladas en arenisca, especímenes de campo . Scale bar 1 cm / escala 1 cm .
Derivatio nominis. It refers to the calyx or cup-like shape of bulbs.
Diagnosis. Series of dominantly linear arrays of vertical, calyx-shaped, concentrically laminated bulbs, budding from a connecting horizontal lined tube.
Description. The burrow system consists of calyxshaped bulbs, forming a linear array of several welded to isolated bulbs connected by a horizontal, mudlined tube. Bulbs vertically elongated, about 5-7 cm in length, maximum diameter 2-3 cm, generally tapering at both ends, with internal regular concentric and/or irregular spiraling or radiating, muddy laminae, alternating with very thin sandy laminae. External part of bulbs bears delicate impressions of oblique to horizontal cylindrical furrows interpreted as bioglyphs (figure 4.F). Inner tube very thin, its diameter ranges from 0.25 to 3.4 mm (average 1.66 mm). In only two specimens, an unusual structure that consists of vertical, imbricate flower-like bulbs (figure 3.A3) was observed in connection to the normal, horizontally aligned calyces.
The burrow system spreads out horizontally covering large areas, in excess of 1 m2, in mudstone beds. Vertical tubes with associated concentric lamination may connect two or more horizontal burrow systems. When the connecting vertical tube cuts through a sandy bed, a distinct cone-in-cone structure is formed around the vertical tube. The structure consists of elongated, vertical to inclined, cylindrical mud packages, spiraling around the vertical inner tube. In cross section, the spiral bands show internal meniscus-like structures (Figures 4.I, 5.L).

Figure 5. Geometry of the internal laminae, illustrations made from photographs, thick black lines and circles represent the inner tube / Geometría de las láminas internas, dibujos de fotografías, las líneas gruesas y círculos en negro representan el tubo interno. A-F, A. radiciforme , horizontal and vertical sections of bulbs / secciones horizontales y verticales de bulbos; A,B,E, half-spiral / semiespiral ; C, continuous spiral / espiral continua ; D, irregular spirals / espirales irregulares ; F, circular / circular ; G-K, P. stratiformis n. isp.; G,H,J, circular / circular; I, half-spiral and continuous spiral / semiespirales y espirales continuas ; K, helicoidal inner tube and bulbs transitional to straight inner tube / bulbos y tubo interno helicoidales transicionales con tubo interno recto ; L-M, P. calyciformis n. isp.; L, mud-filled cone-in-cone structure in sand CADIC PI 28 / estructura cono en cono rellena de fango en arena ; M, half-spirals to irregular spirals / semiespirales y espirales irregulares ; N, P. thalassiformis n. isp., backfilled branches with mostly parallel external, ramified cylindrical tubes / ramas con retrorrelleno y tubos externos ramificados, en su mayoría paralelos a las galerías . Scale bar 1 cm / escala 1 cm .
Preservation. Endichnial in mud-dominated beds.
Remarks. The arrangement of the calyx-shaped bulbs and their internal, complex backfilling patterns indicate a fodinichnion structure. However, the vertical cone-in-cone connection with its elaborated muddy cylinders is difficult to interpret as the result of feeding activity. The introduction of mud within a sandstone bed and the spiral arrangement of the muddy cylinders around the inner tube suggest a kind of very elaborated wall made by the trace-maker for protection of the inner tube.
Localities. Very abundant near the top of unit I. Best examples are just to the NE of Bajada Los Acantilados at Las Grutas.
Patagonichnus stratiformis n. isp.
Figures 3.B, 4.E, 5.G-K, 6.A-F
Type material. Holotype: horizontal, concentrically filled branches (CADIC PI 31 Figure 6.A-B). Paratypes: Several concentrically filled branches (CADIC PI 32, Figure 6.E; CADIC PI 33) and helicoidal bulbs (CADIC PI 34). Additional material: many field specimens.

Figure 6. A-F, P. stratiformis n. isp., full relief / relieve completo ; A-B, general and detailed basal views in longitudinal cross-sections of sinuous backfilled branches with swellings surrounding the inner tube, passively filled with mud, holotype CADIC PI 31 / vistas basales, general y de detalle, de secciones longitudinales, con abultamientos en ramas con retrorrelleno que rodean al tubo interno, con relleno pasivo de fango, holotipo ; C, F, lateral views of inclined (C) and horizontal (F) helicoidal inner tubes and associated backfilled bulbs, field specimens / vistas laterales de tubos internos helicoidales y bulbos asociados con retrorrelleno, inclinados (C) y horizontales (F), especímenes de campo; D, lateral view of concentrically laminated vertical shaft cutting across muddy beds, laterally connected with horizontal layers of sandstone full of bulbs, field specimen / vista lateral de columna vertical con laminación concéntrica que corta a capas fangosas y que se conecta lateralmente con capas horizontales de arenisca, llenas de bulbos, especímenes de campo ; E, vertical section of a typical layer full of connected backfilled branches surrounding the lined inner tube, CADIC PI 32 / sección vertical de una típica capa llena de ramas conectadas con retrorrelleno que rodean al tubo interno con pared ; G, P. thalassiformis n. isp., full relief, upper view, large connected structure with paired backfilled branches and associated concentrically laminated bulbs, holotype CADIC PI 35 / relieve completo, vista superior, estructuras conectadas de gran tamaño, con ramas apareadas con retrorrelleno y bulbos con laminación concéntrica asociados, holotipo. Scale bar 1 cm / escala 1 cm.
Derivatio nominis. It refers to the tendency of the whole burrow system to cover large, stratiform areas.
Diagnosis. Dominantly sinuous, crowded, long horizontal backfilled branches with successive bulb-like lateral swellings surrounding a ramified, central mud-lined tube. Distal ends could be transitional to helicoidal bulbs and associated helicoidal inner tube. Distinct layers could be connected by straight, vertical, conical to helicoidal, concentrically laminated shafts.
Description. Complicated, compound burrow system dominated by long, sinuous horizontal backfilled branches showing successive lateral bulb-like swellings, 1-3 cm wide, with concentric, conical to spiral, backfilling sandy laminae, surrounding a thin, mud-lined tube, ranging from 1.15 to 7.40 mm (average 3.06 mm) in diameter. Large ( ca. 1.5 mm) mica flakes are generally arranged in a vertical position, parallel to the laminae. The inner mud-lined tube may be divided horizontally, giving way to several, sinuous, concentrically laminated branches that generally do not intersect. When two branches are about to meet, mutual intersection is avoided by upwards or downwards deviation movements of the inner tube and the accompanying bulb-like concentrically laminated structures (figure 3.B1). In this way, a large, extensive, horizontal and stratiform structure, covering at least several square meters, is formed. Distal ends of horizontal inner, sinuous tubes could pass transitionally to helicoidal inner tubes (ranging in diameter from 0.7 to 9.2 mm, average 3.3 mm) and associated helicoidal bulbs (figures 3.B3, 5.K, 6.F). Different layers are connected by vertical, conically laminated structures (figures 3.B2, 5.J, 6.D).
Preservation. Endichnial in sandy beds.
Remarks. Vertical or horizontal elementary components with helicoidal inner tube could be mistaken for Gyrolithes or Helicodromites , respectively. However, the associated helicoidal backfilling readily differentiates P . stratiformis from these ichnogenera (figures 3.B3; 5.K, 6.C,F). Vertical, conically laminated shafts (figure 5.J) are similar to Cylindrichnus or Siphonichnus , but in P . stratiformis these vertical shafts are always connected via the inner tube to the stratiform layer of backfilled branches (figures 3.B2; 6.D).
Localities. Very abundant in sandy layers of unit II.
Patagonichnus thalassiformis n. isp.
Figure 3.C-D, 5.N, 6.G, 7.A-E
Type material. Holotype: Silicone cast and mold of paired, filled branches, associated laminated bulbs, and external tubes of part of the burrow system illustrated in figure 6.G (CADIC PI 35).
Paratypes: several partial filled branches, bulbs, and external tubes (CADIC PI 36). Additional material: many field specimens, partly illustrated in figure 7.A-E.

Figure 7. A-D, P. thalassiformis n. isp., full relief, upper view, field specimens / relieve completo, vista superior, especímenes de campo ; AB, large system of backfilled branches and bulbs in sandy siltstone (dark-gray) and external, connected tube networks in tuff (light-gray), arrow in A indicates location of B / sistema de gran tamaño de ramas y bulbos con retrorrelleno en limolita arenosa (gris oscuro) y redes externas de tubos conectados en toba (gris claro), la flecha en A indica la posición de B ; C-D, detailed views of small bulbs and associated dense network of ramified cylindrical tubes of different sizes, sudden expansions in diameter (arrows) probably reflect peristaltic movements of the trace maker / detalle de bulbos pequeños asociados con densas redes de tubos cilíndricos ramificados y de distintos tamaños, la súbita expansión del diámetro (flechas) probablemente refleja movimientos peristálticos del productor; E, P. thalassiformis n. isp., full relief, basal view, field specimen, paired backfilled branches and transversal cross-sections of concentrically laminated bulbs, cutting across Ophiomorpha nodosa (Op) / relieve completo, vista basal, espécimen de campo, ramas apareadas con retrorrelleno y secciones transversales de bulbos con laminación concéntrica cortan a Ophiomorpha nodosa (Op) ; F, full relief, field specimen, large, palmate, crustacean burrow in vertical cross section and upper views, arrow shows vertical section of a branched tunnel with retrusive laminae covered by passively filled concentric laminae / relieve completo, espécimen de campo, traza de crustáceos, palmada y de gran tamaño, en vistas vertical y superior, la flecha indica una sección de túnel ramificado con laminación retrusiva, cubierta por láminas concéntricas de relleno pasivo. Scale bar 1 cm / escala 1 cm .
Derivatio nominis. It refers to the apparent similitude of the burrow system to the galleries of the ichnogenus Thalassinoides .
Diagnosis. Long; dominantly horizontal and straight to curved; single, paired or multiple tubular branches and associated bulbs filled with conical or helicoidal, concentric laminae surrounding an intricate system of lined inner tubes. Horizontal parts could be connected with vertical and shorter backfilled branches. Where the external backfilled structure is missing, the lined inner tube may form intricate, crowded gallery systems with numerous vertical and horizontal branches.
Description. Large, compound, mainly horizontal burrow system, which dominantly consists of up to 1 m long, horizontal, backfilled branches surrounding a complicated system of inner, open lined tubes. Two horizontal, backfilled branches frequently run sideby- side for a long distance and then they detach at acute or right angles, giving the false impression of true branching (figures 3.D, 6.G, 7.B, E). Associated multiple, juxtaposed branches also occur; in most cases they are horizontal and parallel but sometimes they are entangled in joint spiral coils. Crosscutting of the backfilled branches is rare. Horizontal, spindled-shaped, short bulbs could be connected to single, paired or multiple, backfilled branches (figures 6.G, 7.A-C). Backfilling of the tubular branches and bulbs consists of conical or helicoidal laminae. A complex, horizontally and vertically branching network of numerous small tubes, similar to the inner tube, extends outside the backfilled burrow (figures 3.D, 5.N, 7.A-C). In this case, several tubes seem to run in parallel to the surface of the backfilled branches. Short segments of some individual tubes show local expansion of the tube diameter (figures 7.C-D). Single, concentrically backfilled branches are about 1-3 cm wide. The inner open, lined tube ranges from 0.5 to 3.8 mm (average 1.95 mm) in diameter.
Preservation. Endichnial in sandstone beds with abundant fecal pellets of unit II and sandy mudstone beds of unit III. Differentially weathered tuff beds at unit III superbly preserve the complex geometry of the network of lined tubes extending outside the backfilled branches. In part, the external surface of these tubes is covered with ovoid fecal pellets.
Remarks. The geometry of the backfilled branches bears a general resemblance to the gallery system of the ichnogenus Thalassinoides . However, the paired or multiple, closely juxtaposed tubular branches; the concentrically laminated backfilled structures; the associated tapering bulbs; and the inner lined tube are features sufficiently distinctive of P . thalassiformis n. isp. The sub-concentric laminae that may form by passive filling in some galleries of Thalassinoides and Ophiomorpha (Seilacher, 1973; Bromley, 1996) have a different geometry, and are quite different from the delicate concentric lamination of P. thalassiformis n. isp., in which the backfilled material is similar to that of the host rocks. In addition, the associated and connected network of small, branching tubes are not known in Thalassinoides , and this connected tube network is totally different in origin and structure from the composite trace formed by the occupation and reworking of crustacean galleries by polychaetes (Gingras et al ., 2002). In the latter case, the polychaetes used the inner surface of the large crustacean gallery as an extension of the sediment-water interface and thus the threadlike tubes are oriented perpendicular to the gallery inner surface (Gingras et al ., 2002). On the contrary, the tube network in P. thalassiformis tends to be oriented parallel to the surface of the main backfilled branches (figure 5.N).
Localities. Frequent in sandstones bearing abundant reworked fecal pellets at the top of unit II and dominant in sandy siltstones interbedded with thin tuff beds in unit III.
Large, palmate, concentrically laminated burrow
Figure 7.F
Material. Several branched tunnels with retrusive spreite (CADIC PI 37), and many field specimens.
Description. Large, inclined, bundled burrow consisting of a main tunnel from which several minor tunnels branch out in a palmate pattern (figure 7.F). The lower part of the structure consists of an inclined (between 15º-40º) main tunnel up to 40 cm long, about 3-10 cm wide, and with a blind distal end. At its upper part it is divided into several secondary tunnels (minimum 3 and up to 7 observed) forming a broom-like structure. Most of the tunnel wall bears distinctive scratch traces in the form of parallel furrows of similar shape to the bioglyphs made by crustacean decapods (Bromley, 1996). The main and branched tunnels characteristically display one or few sets of retrusive spreite laminae, reaching several centimeters in height, ending with an empty or, more frequently, passively filled, tunnel. The cross sections of these final tunnels are circular, and the passive filling is characterized by concentric laminae with a central or upward displaced, mud-filled tube. The laminae of both the retrusive and the passive fillings consist of different lithologies from that of the host rock. The whole structure is about 70-100 cm in length and 20-50 cm in height.
Remarks. Ethologically the large, palmate burrows are interpreted as both domichnia and equilibrichnia structures. The burrow was probably made by crustacean decapods, which constructed a deep, inclined tube (the main tunnel) and two or more branches connecting to the surface, allowing the animal to maintain water circulation through the main domichnion burrow. The retrusive spreite represents the activity of the producer, packing on the burrow's floor the sediment that accidentally entered the tunnels from outside or from other parts of the burrow. The vertical piling-up of successive, retrusive sets of laminae, probably represents an equilibrium structure, in response to agradational increments of the sedimentary surface and/or to sudden sedimentary infill or collapse of the open tunnel. Near complete infilling of the tunnel promoted the opening of new branches and the abandoned tunnel was passively filled with the concentric laminae. This large, palmate burrow is an elite trace fossil in unit I. It is also present in the layers with fecal pellets characteristic of unit II. Formal systematics of this burrow are delayed, pending results of studies in progress by the authors.
Discussion and conclusions
Internal structure and origin of the concentric lamination
Though not generally accepted, two basic models are used to explain the origin of the obligatory, active filling in concentrically laminated burrows. In one model, the basic features are interpreted as the result of radial backfilling associated with feeding activities (figure 8.A; Seilacher, 1986). In Asterosoma and Rosselia this model was related to the feeding activity of a worm-like organism that systematically reworked the sediment with spiral movements of its distal end, while periodically moving along an inner tube (figure 8.B; Chamberlain, 1971). In the other model, a tube-dwelling organism produces the concentrically laminated structure by adding successive wall layers to the tube and pushing and splitting previous layers outwards, and the resulting concentric laminae are more a thick wall-lining than a backfilled structure (figures 8.C-D; Goldring, 1996; Bromley, 1996 and the bibliography therein). Nara (1995) applied this model to explain the origin of the concentric laminae in Rosselia socialis Dahner, suggesting terebellid polychaetes as the trace markers. The superposition of successive spindle-shaped parts (figure 8.D) was interpreted as an equilibrium structure (Nara, 1995).
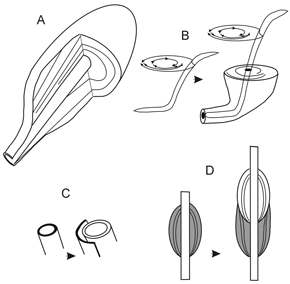
Figure 8. Basic models for the origin of obligatory, concentric lamination / Modelos básicos de génesis de laminación concéntrica obligatoria. A: radial backfilling (Seilacher, 1986) / retrorrelleno radial ; B, radial backfilling as a result of spiral movements of a worm (Chamberlain, 1971) / retrorrelleno radial originado por movimientos espirales de un gusano ; C-D, concentric lamination formed by the pushing and splitting mechanism (Nara, 1995; Bromley, 1996) / laminación concéntrica formada por el mecanismo de empuje y ruptura.

Figure 9. Interpreted burrowing mechanisms of Patagonichnus and Asterosoma from Las Grutas / Interpretación de los mecanismos de excavación en Patagonichnus y Asterosoma de Las Grutas. A, basic structure / estructura básica; B, sequential formation of the helicoidal backfilling from times 0 to 4 / secuencia de formación del retrorrelleno helicoidal para las etapas 0 a 4 ; C, deformation of concentric laminae by the lateral pushing of a worm-like organism / deformación de láminas concéntricas por empuje lateral de un organismo vermiforme. See text for explanation / Explicación en el texto.
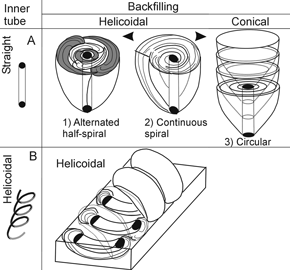
Figure 10. 3-D reconstruction of the backfilling geometry in Patagonichnus and Asterosoma from Las Grutas / Reconstrucción espacial de la geometría del retrorrelleno en Patagonichnus y Asterosoma de Las Grutas . A, straight inner tubes produce helicoidal or conical backfilled bulbs, 1, 2, and 3 indicate the geometry in cross-section / tubos internos rectos producen bulbos con estructuras helicoidales o cónicas, 1, 2, y 3 indican la geometría en sección transversal; B, helicoidal inner tubes produce only helicoidal bulbs / tubos helicoidales sólo producen bulbos con estructura helicoidal.
In Asterosoma radiciforme and the three new ichnospecies of Patagonichnus from Las Grutas, the bioturbated material that composes the concentrically laminated structures is readily differentiated from the host sediments by contrast in color and texture. The reconstruction of the burrow system (figure 3) and the geometry of the internal structure (figure 5) clearly indicate that the concentric lamination is best explained as the radial backfill produced by elongate, worm-like organisms spiraling or circling within the sediment while feeding (figure 9). The basic backfilling mechanism is similar to that proposed by Chamberlain (1971) but significant improvement of this model results from the analysis of the inner structure of the material from Las Grutas.
The analysis of transverse cross-sections shows three recurrent patterns for the arrangement of the internal laminae: 1, discordant groups of laminae defining two half spirals, which are arranged into an alternated pattern (figures 5.A-B); 2, laminae arranged into more continuous spirals (figure 5.C); and 3, circular laminae (figures 5.F, J). Transitions among these patterns are frequent. Patterns 1 and 2 are common in all studied Asterosoma and Patagonichnus from Las Grutas (figures 5.A-E, I, M). Pattern 3 is more frequent in the straight stem of A . radiciforme (figure 5.F); in all compound elements of P. stratiformis (figures 5.G-J); and in the backfilled branches of P . thalassiformis (figure 5.N). In addition, in P . stratiformis these three patterns are transitional with helicoidal bulbs (figures 5.K, 6.F).
By using the case of pattern 1 the alternated half-spiral geometry as an example, the three-dimensional interpretation for the development of the whole structure is shown in figure 9. The basic plan of the structure results from the spiral movement of a worm-like organism with one end in the terminal position of the inner lined tube and the other end extended and spiraling within the sediment (figure 9.A). The interpretation of the sequential development of the structure is shown in figure 9.B. At time 0, during the initial production of the first half-spiral, the organism excavates a conical burrow and then moves the open burrow, excavating towards the center and backfilling the periphery. The process of movement and backfilling of the initial burrow is recorded by a set of slightly discordant, sub-concentric laminae, which in cross-section widens in the direction of progression of the structure and in longitudinal cross-section thins toward the inner lined tube. This process continues after completion of the first half-spiral (time 1) and is then repeated again, but now moving a little bit nearer the inner lined tube, forming the second half-spiral (times 1 and 2). At time 1, during the initiation of the construction of the second half-spiral its point of origin is displaced upwards respect to the similar point of the first halfspiral at time 0, and the resulting three-dimensional geometry is a helicoidal body (i.e. the spiral is stretched in the third dimension). At times 3 and 4, the open burrow now lies in a more central position in the plane containing the cross-section, the inner lined tube has grown in the direction of progression of the structure, and the widest parts of the sets of laminae corresponding to the third and fourth halfspirals were intercalated beside the thinnest parts of the previous sets of the first and second half-spirals. In this way, maximum exploitation of the sediment in the search for food is achieved. The model for the construction of the structure is very similar to the helicoidal phyllotaxis in plants, and in fact it was inspired by the helicoidal arrangement of alternated couples of leafstalks at the basal stem of the celery plant (Stevens, 1989, Fig. 127).
The contact between successive laminae is not even; it frequently is irregular and sinuous (figure 9.C), and this is best seen when the set of laminae consists of alternating thick clay- and thin sandylaminae. This sinuous contact is interpreted as having originated by the pressure exerted by the tracemaker, obviously with a cylindrical body, pushing against the tube-wall.
As shown in figure 10, the sequential development of the structure for the cases of patterns 2 and 3 is similar to that of the described model. In case 2, the continuous spirals partially wrap and the structure is developed much like a gastropod shell, and in case 3, the circular laminae result from the successive displacement of a moving cone. If the inner lined tube is kept straight, the combination of spiraling or circling movements results in helicoidal or conical backfilling, respectively (figure 10.A). If the inner tube is helicoidally twisted, the resulting backfilling is always helicoidal (figure 10.B).
Behavior of the trace-maker and function of the elementary components
All the Patagonichnus material from Las Grutas consists mainly of large, horizontally spread out structures connected by smaller vertical structures, both including an open, mud-lined tube, isolated or surrounded by various concentrically laminated structures (figure 3). The complex backfilling mechanisms clearly indicate mainly fodinichnia structures. However, all of these structures are large, compound burrow systems and the analysis of the elementary components suggests that some of them could be designed to also accomplish different, coordinated functions.
By definition, fodinichnia are characterized by the combined functions of dwelling and feeding. The open, inner mud-lined tube is interpreted to function mainly as a dwelling structure, providing also connection, irrigation and ease of movement of the organisms among different parts of the burrow system. The construction of an extensive mud-lined wall is generally associated with a long occupation of the burrow by the trace maker (Bromley, 1996), and this is consistent with the large area, in excess of 1 m 2 , covered by a single burrow system. The complex strategies used in the development of the backfilled bulbs and swellings indicate that they represent feeding activities; such interpretation is consistent with the restriction, and large horizontal extension, of these backfilled structures to specific horizons, as if the trace maker was differentially mining particular, food-rich layers. The contrast in color and texture of the bioturbated sediment respect to the host rock is also indicative of a feeding activity.
The vertical, mud-filled cone-in-cone structure of P. calyciformis is only present when a sand layer is interposed between mud layers, and it is always found in connection with distinct, horizontally extended backfilled structures, developed only in the lower and upper mud layers (figures 3.A, 4.I, 5.L). The mutual restriction to specific grain-sizes of these different, but connected, structures seems to indicate that the P. calyciformis producer was adapted to actively feed on mud but not on sand. The likely functional interpretation of the cone-in-cone structure is that of an elaborated wall made of mud cylinders. If, when exploring new mining layers, the organism was forced to cross a sandy layer, apparently it built a complex wall, designed mainly for protection of the connecting inner tube.
The vertical, conical laminated shaft of P. Stratiformis (figures 3.B, 5.J, 6.D) is another distinctive structure frequently found in thin alternating sand and mud beds. This vertical shaft is only connected to the horizontally extended laminated bulbs in sandy layers but such bulbs are not developed in the intervening muddy layers (figures 3.B3, 6.D). In this case it seems that the P. stratiformis producer was adapted to only feed in sandy layers. The best functional interpretation for the vertical, conical laminated cylinder is that of a probing structure; and probably it represents a vertical excursion of the organism to detect the preferred grain-size layer for mining.
The intricate tube network projecting outside the backfilled burrow in P. thalassiformis (figures 3.D, 5.N, 7.A-D) is the most intriguing structure. This tube network is frequently found in association with thin, distinctive tuff beds in unit III. Multiple anastomosing and division of the tubes with an irregular pattern is a common feature, but there is a definite tendency for the tubes to follow the surfaces of the backfilled branches at the boundaries of the tuff layers (figures 3.D, 7.B-C). We can only guess that the functional interpretation of this tube-network is that of a probing system searching for food. If the organisms actively mining a food-rich layer encounter a barren tuff bed, and some of them succeed in finding the way to a new food-rich layer, the others try to follow them looking for the successful passage.
The three new ichnospecies are elite burrow systems that distinctively dominate the bioturbated fabric in mud- or sand-rich layers. In P . thalassiformis , P . calyciformis and P . stratiformis the areal distribution of a single burrow system covers at least 1 m2. But as the burrow system consists of several, repeated horizontal layers connected by vertical structures, the total area covered by a single burrow system must be much larger. A single layer, exclusively and fully bioturbated to form P. stratiformis ( e.g . figure 6.E) was followed to at least 15 m2. It is unlikely that a single organism could make such a huge burrow system. On the contrary, it looks as though these large burrow systems, with many different elementary components, were constructed by gregarious organisms, living and working simultaneously. Measurements of the inner, mud-lined dwelling tube seem to confirm this interpretation. The diameter of the inner tube which represents the maximum transversal size of the dwelling organism in any single burrow system of the three new ichnospecies, records a continuous range of variation in size. Continuous range is from 0.25 to 4 mm (average 1.66 mm) in P . calyciformis ; 0.5 to 3.8 mm (average 1.95 mm) in P. thalassiformis ; 1.15 to 7.44 mm (average 3.06 mm) in P . stratiformis with straight inner tube; and 1.47 to 9.2 mm (average 3.3 mm) in P . stratiformis with helicoidal inner tube (figure 11). The fact that this continuous range of sizes is not a representation of some kind of growth vector of a single organism is demonstrated by the frequent joining of inner tubes of different diameters into a single point (figures 7.C-D). This is strong evidence that many similar organisms of different sizes were living and working simultaneously within any single burrow system. Large, fully bioturbated areas exceeding 15 m2, can be explained by the close association of many single burrow systems.

Figure 11. Frequency of range of size of the inner tube diameter plotted every 0.5 mm. n: number of measurements; D: average tube diameter / Frecuencia de distintos rangos de tamaños del diámetro del tubo interno graficados cada 0,5 mm. n: número de mediciones, D: diámetro promedio del tubo.
Regarding the depth of penetration of the burrow system, both direct and indirect evidence indicate that the Patagonichnus producer penetrated several tens of centimeters into the sediment. The conical laminae of the vertical connecting-shaft of P. stratiformis recorded in thin, interbedded sand and mud layers, provide direct evidence that the burrow penetrated the sediment to a depth of at least 30 cm. As the sedimentation of the thin-bedded heterolithic beds was continuous, with no signs of internal erosion surfaces, the downward movement of the burrow is not an equilibrium structure but represents a true minimum depth reached by the organism. Crosscutting relationships with the palmate crustacean burrow and Ophiomorpha provide additional indirect evidence. Patagonichnus calyciformis and P. stratiformis always cut both the palmate branches and the main stem of the crustacean burrow. Even though Ophiomorpha more frequently cuts the Patagonichnus burrows, mutual crosscutting relationships were also observed. Crustacean burrows range from 20 to 50 cm in height, confirming that the Patagonichnus structures penetrated at least about 20-30 cm below the sediment-water interface.
Identification of the trace-makers
In general, biologic identification of trace makers is problematic ( cf. Frey, 1975; Seilacher, 1986). In the material from Las Grutas there is some direct evidence on size and form of the trace maker. Obviously, the cylindrical inner, lined tube indicates that the trace maker must have had an elongate, worm-like body, with only a few millimeters in cross-section (figures 4.E-F, 6.B, 7.B-D). Similar form and size of the producer are also indicated by the sinuous deformation of the laminae within the bulbs, probably produced by the repeated lateral pushing of the organism against the burrow wall (figure 9.C), and by the bioglyphs in the form of cylindrical furrows preserved on the external part of the bulbs in P. calyciformis (figure 4.F). Additional analogies with known burrow geometries, mode of life, and feeding strategies of extant worm-like organisms suggest polychaetes as the most probable architects for the Patagonichnus burrows from Las Grutas.
Irregular, intricate tube systems of burrowing polychaetes are relatively common in modern tidal flats and intertidal channels (Reineck et . al ., 1967; Schäfer, 1972; Barnes, 1974) and the analogies in size and form of some of them with the Patagonichnus burrows from Las Grutas are striking. In particular, Reineck et al . (1967), Schäfer (1972), Powell (1977), Ronan (1977), and Gingras et al . (1999, 2002), described polychaete burrows of similar form and size to those of the Patagonichnus inner tube from Las Grutas. Maldanid and capitellid polychaetes are the known trace-makers of Cylindrichnus -, Gyrolithes -, Trichichnus - or Chondrites -like burrows (Gingras et al . (1999, 2002). Similar forms are found at Las Grutas; in particular the helicoidal inner tube of P . Stratiformis seems to be a common feature developed by capitellid polychaetes (Reineck et al ., 1967; Schäfer, 1972; Powell, 1977; Gingras et al ., 1999, 2002). The capitellid Notomastus magnus Hartman makes large, relatively deep, irregular and meandering tube systems using its proboscis for feeding and locomotion; eversion of the proboscis is recorded in the burrow by a local expansion of the tube diameter (Ronan, 1977) and this structure seems to be present in the tube network of P. thalassiformis (figures 7.C-D). Some relatively deeply burrowing polychaetes commonly develop a complicated burrow system that tends to expand horizontally at certain levels ( e.g some species of Nereis and Heteromastus described by Schäfer, 1972) and this is a common feature of all Patagonichnus from Las Grutas. Different but related species of polychaetes are frequently adapted to burrow different substrates, preferring either fine sand or mud, where they are found in large numbers, usually several hundreds of specimens per square meter (Schäfer, 1972; Barnes, 1974), and the size of the inner tube is a diagnostic species-specific feature in some polychaetes (Gingras et al ., 1999). The Patagonichnus from Las Grutas share all of these polychaete features. Patagonichnus stratiformis is only developed in fine sand and P. calyciformis is mostly developed in mud, and both are characterized by a large number of crowded, associated inner tubes. To a certain extent the inner tube overlaps in size, but when large numbers of specimens are measured, two distinctive size-frequency distributions are clearly shown: the average diameter of the inner tube in P . Calyciformis and P. thalassiformis (mudstone beds) is less than 2 mm, and in P. stratiformis (sandstone beds) is more than 3 mm (figure 11).
Longitudinal cross-sections of P. stratiformis (figures 6.A-B) bear a general similarity to Nereites , which also is produced by a worm-like organism, probably an enteropneust, which sorted out the particles with its protosoma, stowed them in lateral backfilled lobes, and stuffed the ingested material into a median tunnel, behind the animal (Seilacher, 1986). The anterior part of the Patagonichnus animal probably processed the sediments with a similar mechanism but the lack of the stuffed material in its inner, median, tube indicates a different behavior, and probably a different organism. Worm-like organisms in general and deep burrowing polychaetes in particular, use several mechanisms for movement, including bolting, peristaltic and undulatory movements, multiple circular shoveling using specialized parapodia, or some combinations of these (Schäfer, 1972; Buatois et al ., 2002). Their feeding strategies are complex, and they could be selective or non-selective deposit feeders, carnivorous, or omnivorous (Ronan, 1977). Regardless of these different mechanisms and strategies, errant, burrowing polychaetes all have a hollow, and commonly lined, inner tube that connects the horizontal and vertical branches of the burrow system (Schäfer, 1972; Barnes, 1974). As explained before, the main function of the inner tube is that of a dwelling structure, providing both irrigation and ease of movement for the organism among the different parts of the burrow system. In analogy with extant polychaetes, the hollow, lined inner tube of Patagonichnus from Las Grutas probably was also functional to evacuate the fecal material to the surface; explaining why the residual of the ingested material is never found stuffed in the median, inner tunnel as in Nereites .
Acknowledgments
We thank A. Seilacher (University of Tübingen) for helpful comments on Asterosoma , G. Mángano (INSUGEO) for helping with the recognition of Asteriacites , N. Malumián (Servicio Geológico Minero Argentino, SEGEMAR-Consejo Nacional de Investigaciones Científicas y Técnicas, CONICET) for advising on the stratigraphy at Las Grutas, and the reviewers R.G. Bromley and L. A. Buatois for useful and valuable comments. A. Uchman (Jagiellonian University) and S. Thajte (Alfred Wegener Institut) helped with the bibliography, J. Strelin and D. Martinioni (Centro Austral de Investigaciones Científicas, CADIC) with German translations and V. Olivero with the English writing. Part of this research was funded by ANPCYT-FONCYT PICT 8675.
References
Angulo, R., Fidalgo, F., Gómez Peral, M.A. and Schnack, E.J. 1979. Las ingresiones marinas cuaternarias en la bahía de San Antonio y sus vecindades, Provincia de Río Negro. 7º Congreso Geológico Argentino (Neuquén, 1978), Actas 1: 271-283. [ Links ]
Barnes, D. B. 1974. Invertebrate Zoology . Third Edition. W.B. Saunders Company, 870 pp. [ Links ]
Bromley, R.G. 1996. Trace Fossils. Biology, taphonomy and applications. Chapman & Hall, London, 361 pp. [ Links ]
Bromley, R.G. and Uchman, A. 2003. Trace fossils from the Lower and Middle Jurassic marginal marine deposits of the Sorthat Formation, Bornholm, Denmark. Bulletin of the Geological Society of Denmark 52: 185-208. [ Links ]
Bromley, R.G., Uchman, A., Gregory, M.R. and Martin, A. 2003. Hillichnus lobosensis igen. et isp. nov., a complex trace fossil produced by tellinacean bivalves, Paleocene, Monterrey, California, U.S.A. Palaeogeography , Palaeoclimatology , Palaeoecology 192: 157-186. [ Links ]
Buatois, L., Mángano, G. and Aceñolaza, F. 2002. Trazas fósiles. Señales de comportamiento en el registro estratigráfico. Edición Especial MEF 2. Museo Paleontológico Egidio Feruglio, Trelew 382 pp. [ Links ]
Chamberlain, C.K. 1971. Morphology and ethology of trace fossils from the Ouachita Mountains, southeast Oklahoma. Journal of Paleontology 45: 29-32. [ Links ]
Ekdale A.A. and Bromley, R.G. 2001. A day and night in the life of a cleft-foot clam. Lethaia 34: 119-124. [ Links ]
Frey, R.W. 1975. The realm of ichnology, its strengths and limitations. In: R.W. Frey (ed.), The study of trace fossils. A synthesis of principles, problems and procedures in ichnology. Springer-Verlag, New York, pp. 13-38. [ Links ]
Frey, R.W. and Howard, J.D. 1970. Comparison of Upper Cretaceous ichnofaunas from siliceous sandstones and chalk, Western Interior Region, U.S.A. Geological Journal Special Issue 3: 141-166. Liverpool. [ Links ]
Frey, R.W. and Howard, J.D. 1982. Trace Fossils from the Upper Cretaceous of the Western Interior: Potential criteria for Facies Models. The Mountain Geologist 19: 1-10 [ Links ]
Frey, R.W. and Howard, J.D. 1985. Trace Fossils from the Panther Member, Star Point Formation (Upper Cretaceous), Coal Creek Canyon, Utah. Journal of Paleontology 59: 370-404. [ Links ]
Frey, R.W. and Seilacher, A. 1980. Uniformity in marine invertebrate ichnology. Lethaia 13: 183-207. [ Links ]
Gingras, M.K., Pemberton, S.G., Saunders, T.D.A. and Clifton, H.E. 1999. The Ichnology of modern and Pleistocene brackishwater deposits at Willapa Bay, Washington: Variability in estuarine settings. Palaios 14: 352-374. [ Links ]
Gingras, M.K., Pickerill, R. and Pemberton, S.G. 2002. Resin cast of modern burrows provides analogs for composite trace fossils. Palaios 17: 206-211. [ Links ]
Goldring, R. 1996. The sedimentological significance of concentrically laminated burrows from Lower Cretaceous Ca-bentonites, Oxfordshire. Journal of the Geological Society, London 153: 225-263. [ Links ]
Häntzschel, W. 1975. Trace Fossil and Problematica. In: C. Teichert (ed), Treatise on Invertebrate Paleontology, W. Geological Society of America and Kansas University Press, Boulder and Lawrence, 269 pp. [ Links ]
McCarthy, B. 1979. Trace fossils from a Permian shoreface-foreshore environment, Eastern Australia. Journal of Paleontology 53: 345-366. [ Links ]
Müller, A.H. 1971. Zur Kenntnis von Asterosoma (Vestigia invertebratorum). Freiberger Forschungshefte (C) 267: 7-17, Leipzig. [ Links ]
Nara, M. 1995. Rosselia socialis : a dwelling structure of a probable terebellid polychaete. Lethaia 28: 171-178. [ Links ]
Nara, M. 2002. Crowded Rosselia socialis in Pleistocene inner shelf deposits: Benthic paleoecology during rapid sea-level rise. Palaios 17: 268-276. [ Links ]
Olivero, E.B and López C., M.I. 2001. Ichnology of syntectonic turbidites, estuarine and shelf deposits. Eocene, Punta Torcida anticline, Isla Grande de Tierra del Fuego. 4° Reunión Argentina de Icnología y 2° Reunión de Icnología del Mercosur (S. M. de Tucumán). Resúmenes : 62. [ Links ]
Olivero, E.B. y López C., M.I. 2002. Icnocenosis de Terebellina y Neonereites en depósitos mareales del Cretácico superior de Antártida. 9º Reunión Argentina de Sedimentología (Córdoba). Resúmenes : 67. [ Links ]
Otto, E. von, 1854. Additamente zur Flora des Quadergebirges in Sachsen. Heft 2, 53 pp., Leizig (G. Mayer). [ Links ]
Pickerill, R.K. and Narbonne, G.M. 1995. Composite and compound ichnotaxa: a case example from the Ordovician of Québec, eastern Canada. Ichnos 4: 53-69. [ Links ]
Powell, E.N. 1977. The relationship of the Trace Fossils Gyrolithes (= Xenohelix ) to the family Capitellidae (Polychaeta). Journal of Paleontology 51: 552-556. [ Links ]
Reineck, H.E., Gutmann, W.F. and Hertweck, G. 1967. Das Schlickgebiet südlich Helgoland als Beispiel rezenter Schelfablagerungen. Senckenbergiana Letheia 48: 219-275. [ Links ]
Ronan, T.E., Jr. 1977. Formation and paleontologic recognition of structures caused by marine annelids. Paleobiology 3: 389-403. [ Links ]
Schäfer, W. 1972. Ecology and Palaeoecology of Marine Environments . The University of Chicago Press. Chicago, 568 pp. [ Links ]
Schlirf, M. 2000. Upper Jurassic trace fossils from the Boulonnais (northern France). Geologica et Palaeontologica 34: 145-213, Marburg. [ Links ]
Seilacher, A. 1969. Sedimentary rhythms and trace fossils in Paleozoic sandstone of Libya In: W.H. Kannes, Geology, archeology and prehistory of southwestern Fezzan, Libya. 11th Annual Conference: 117-123. Trípoli. [ Links ]
Seilacher, A. 1973. Biostratinomy: The sedimentology of biologically standardized particles. In: R.N Ginsburg (ed), Evolving concepts in sedimentology. The Johns Hopkins University Press, Baltimore and London, pp. 159-177. [ Links ]
Seilacher, A. 1986. Evolution of behavior as expressed in marine trace fossils. In: M.H. Nitecki and J.A. Kitchell (eds.), Evolution of animal behavior. Paleontological and field approaches. Oxford University Press, New York and Oxford, pp. 62-87. [ Links ]
Stevens, P.S. 1989. Patrones y pautas en la naturaleza (Spanish translation of: Patterns in nature). Biblioteca Científica Salvat, Salvat Editores S.A. Barcelona, 293 pp. [ Links ]
Uchman, A and Krenmayr, H.G. 1995. Trace Fossils from the Lower Miocene (Ottnangian) molasse deposits of Upper Austria. Paläontologische Zeitschrift , 69: 503-524. Stuttgart. [ Links ]
Recibido: 6 de noviembre de 2003.
Aceptado: 7 de julio de 2004.

