










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.2 Buenos Aires mar./jun. 2005
Conodontes de la Formación Rupasca (Ordovícico Inferior) en el Angosto de Chucalezna, Cordillera Oriental de Jujuy: nuevos elementos bioestratigráficos para una localidad clásica del noroeste argentino
Guillermo L. Albanesi1 y Guillermo F. Aceñolaza2
1 CONICET, Museo de Paleontología, Universidad Nacional de Córdoba, C. C. 1598, 5000 Córdoba, Argentina. galbanes@com.uncor.edu
2 CONICET, INSUGEO, Universidad Nacional de Tucumán, Miguel Lillo 205, 4000 Tucumán, Argentina. insugeo@csnat.unt.edu.ar
Resumen. En la presente contribución se da a conocer una asociación de conodontes de la Formación Rupasca expuesta en localidad del Angosto de Chucalezna, Quebrada de Humahuaca, Cordillera Oriental jujeña. En esta localidad aflora la parte inferior a media de la Formación Rupasca, cuyos estratos portadores de conodontes corresponden a areniscas y pelitas pardo-amarillentas, altamente fosilíferas, estrato y granocrecientes depositadas en un ambiente submareal ( shoreface transicional / off-shore superior). Estas rocas constituyen un intervalo estratigráfico característico, con representación regional en la Cordillera Oriental. Los microfósiles se recuperaron de coquinas carbonáticas politáxicas de origen tempestítico, sobre las que se efectúa un análisis petrográfico y tafonómico. Entre las especies de conodontes descriptas se incluye a Paltodus deltifer pristinus Viira y se diagnostican dos especies nuevas: Drepanoistodus chucaleznensis n. sp. y Utahconus humahuacensis n. sp. La fauna de conodontes registrada se asigna a Zona de Paltodus deltifer (Subzona de Paltodus deltifer pristinus ) del Tremadociano superior bajo, según el esquema de biozonación del reino Atlántico. Finalmente, una evaluación paleoambiental y paleobiogeográfica de la nueva asociación de conodontes revela un intervalo de baja diversidad y sugiere su asignación al reino faunístico de Transición, que comprende faunas mixtas en latitudes medias.
Abstract. Conodonts from the Rupasca Formation (Lower Ordovician ) at Angosto de Chucalezna , Jujuy Oriental Cordillera : new biostratigraphical evidence for a classic locality of northwestern argentina . This contribution presents a detailed analysis of a conodont fauna from the Angosto de Chucalezna locality, at the Quebrada de Humahuaca, Cordillera Oriental of Jujuy province. At this locality, where the lower and middle parts of the Rupasca Formation crop out, the conodont bearer strata correspond to yellowish sandstones and shales. They integrate a grain and strata coarsening upwards highly fossiliferous sequence deposited on a subtidal setting (shoreface transition / upper offshore). This sequence represents a characteristic interval in the Cordillera Oriental, and the microfossils were recovered from the interbedded polytaxic carbonate coquinas. Petrographic and taphonomic analyses are carried out on these storm generated rocks. Described taxa include Paltodus deltifer pristinus Viira and two new species: Drepanoistodus chucaleznensis n. sp. and Utahconus humahuacensis n. sp. The conodont fauna corresponds to the Paltodus deltifer Zone ( Paltodus deltifer pristinus Subzone) of the Atlantic realm, which is early late Tremadocian in age. Finally, a paleoenvironmental and paleobiogeographical evaluation of this new conodont association reveals a low diversity interval and suggests assignment to the Transitional faunal realm, which involves mixed faunas in middle latitudes.
Key words. Conodonts; Rupasca Formation; Lower Ordovician; Eastern Cordillera; Argentina.
Palabras clave. Conodontes; Formación Rupasca; Ordovícico Inferior; Cordillera Oriental; Argentina.
Introducción
La Cordillera Oriental del noroeste argentino constituye una región clásica en la bibliografía paleontológica del eopaleozoico sudamericano. El amplio desarrollo de las secuencias cambro-ordovícicas del área, con espesores que en numerosas oportunidades superan los miles de metros, junto a una relativa facilidad de accesos a sus afloramientos, ha generado una importante información paleontológica a partir de los estudios de Kayser (1925), Kobayashi (1937), Harrington (1937, 1938), y Harrington y Leanza (1957), entre otros.
Si bien el particular estilo estructural de la Cordillera Oriental condiciona el desarrollo estratigráfico de las secuencias, se reconocen numerosas unidades litológicas correlacionadas tanto por sus caracteres sedimentarios como por su extensión bioestratigráfica ( e.g ., Moya, 1988, 1999, 2002). La Formación Rupasca (Harrington, 1957, emend. López y Nullo, 1969) se incluye dentro de esta categoría ya que resulta poco frecuente observarla con un desarrollo estratigráfico normal en toda la comarca, encontrándose generalmente asomos esporádicos, parciales y discontinuos, con espesores variables. Ante la mencionada situación, se destaca la importancia que posee el detallado conocimiento tanto del marco estratigráfico como de la sucesión bioestratigráfica en los estratos cambro-ordovícicos de la región (síntesis recientemente publicadas en: Aceñolaza, ed., 2002 y Benedetto, ed., 2003).
En esta oportunidad se presentan nuevos datos paleontológicos, referidos a la fauna de conodontes de la Formación Rupasca (Tremadociano superior) que aflora en el tramo norte del corte del ferrocarril del Angosto de Chucalezna, Quebrada de Humahuaca, provincia de Jujuy (figura 1). La presente contribución aporta información novedosa al conocimiento de una unidad solo recientemente tratada en sus aspectos micropaleontológicos (Zeballo et al ., 2003, 2005a, 2005b), dando a conocer en detalle la asociación de conodontes que fuera oportunamente adelantada en una comunicación breve (Aceñolaza y Albanesi, 1996).

Figura 1. Mapa de ubicación de los afloramientos de la Formación Rupasca en el Angosto de Chucalezna, Quebrada de Humahuaca, provincia de Jujuy. El asterisco marca la localidad de procedencia de los fósiles / Location map showing outcrops of the Rupasca Formation at Angosto de Chucalezna, Quebrada de Humahuaca, Jujuy Province. The asterisk marks the fossil locality.
Estratigrafía y paleontología
La nominación de Formación Rupasca constituye la categorización formal dada por López y Nullo (1969) a la denominación informal de lutitas Rupasca (Rupasca shales), aplicada por Harrington (1957) para describir una potente secuencia arenosopelítica aflorante en la quebrada homónima, sobre el flanco oeste de la sierra de Tilcara (figura 2).
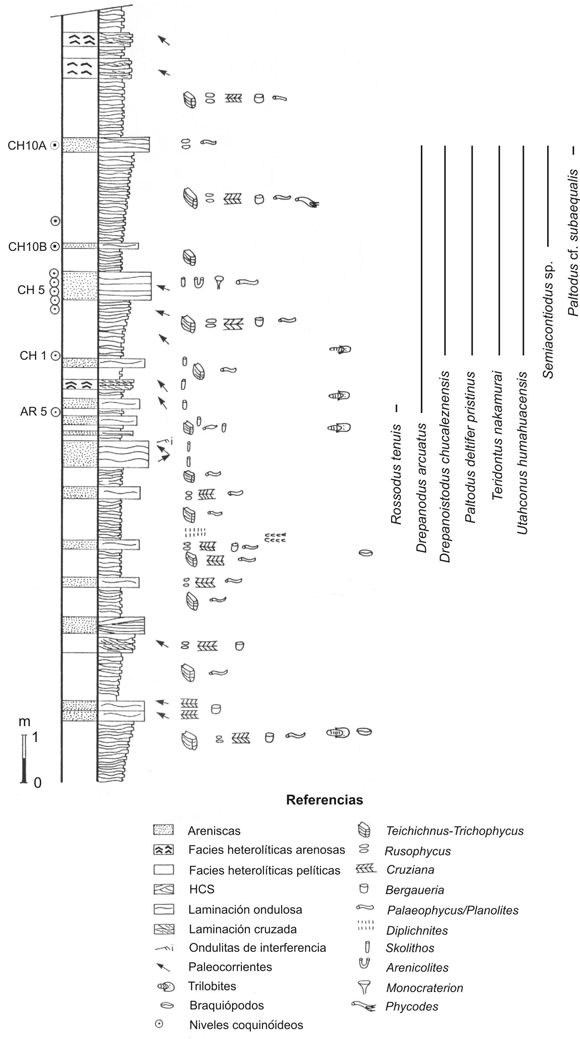
Figura 2. Perfil estratigráfico de la secuencia estudiada en el corte del ferrocarril. Se destacan en la parte superior los niveles portadores de la fauna de conodontes y los rangos estratigráficos de las especies registradas (modificado de Aceñolaza et al ., 2003a) / Stratigraphic profile of the studied sequence from the rail cut. Bearer levels are located in the upper part of the column and species ranges are depicted (modified from Aceñolaza et al. , 2003a).
En su localidad tipo, la unidad se encuentra litológicamente representada por casi 100 metros de una sucesión mayoritariamente heterolítica, de areniscas y lutitas fosilíferas con coloraciones pardo amarillentas (Harrington, 1957; Aceñolaza, 1996; Zeballo et al ., 2003, 2005a, 2005b). En un trabajo reciente sobre las facies sedimentarias y evolución depositacional de la Formación Santa Rosita, Buatois y Mángano (2003) interpretaron a la Formación Rupasca como uno de los miembros superiores de la Formación Santa Rosita, que incluye una serie de eventos transgresivo-regresivos del Tremadociano superior, con amplia extensión regional.
El afloramiento tratado en esta oportunidad, en el Angosto de Chucalezna, constituye un bloque de 39 metros de espesor, limitado por fallas, que se corresponde parcialmente al sector inferior y medio de la unidad. Esta secuencia posee la particularidad de presentar seis niveles destacables de coquinas con cementación carbonática, las cuales resultaron portadoras de una variada asociación de conodontes, objeto de la presente contribución. Las areniscas y pelitas que caracterizan a la Formación Rupasca representan un sistema de depositación submareal, de shoreface transicional al offshore , entre los niveles mínimos y máximos de acción de oleaje (Aceñolaza y Poiré, 1999; Aceñolaza et al ., 2003a).
Los estratos aflorantes en el corte del ferrocarril son altamente fosilíferos, con trilobites, braquiópodos inarticulados, equinodermos y trazas fósiles que contribuyen a un mejor entendimiento del esquema bioestratigráfico de la secuencia. La fauna de trilobites corresponde a las zonas de Bienvillia tetragonalis - Shumardia ( Conophrys ) minutula y Notopeltis orthometopa , con las especies Shumardia ( C .) minutula Harrington, Hapalopleura clavata Harrington y Leanza, H. longicornis Harrington y Leanza, Asaphellus jujuanus Harrington, Leptoplastides marianus (Hoek) y Protopeltura mesembria Harrington y Leanza, entre las más frecuentes. El material asignado a equinodermos es siempre fragmentario y se compone por artejos de pelmatozoos aislados, o asociados en grupos, y por segmentos tecales desarticulados. Proporcionalmente constituye el grupo menos frecuente, habiéndose reconocido solamente la presencia del género Macrocystella . Asimismo, el afloramiento ha sido propuesto como una localidad de interés icnológico por su destacable abundancia y preservación de trazas fósiles, así como por su accesibilidad a la vera de la ruta nacional 9 (Aceñolaza, 2001). Las trazas fósiles se caracterizan por una asociación dominada por elementos de la icnofacies de Cruziana (Seilacher, 1964, 1967; Frey et al ., 1990; Pemberton et al ., 1992a, 1992b), destacando entre las formas presentes a los icnogéneros Cruziana , Rusophycus , Trichophycus y Teichichnus (Aceñolaza, 1996, 2001; Aceñolaza y Poiré, 1999; Mángano et al ., 2001; Aceñolaza et al ., 2003a, con referencias).
Tafonomía y petrografía de las coquinas
Los caracteres tafonómicos, texturales y de composición de las coquinas de la Formación Rupasca han permitido recientemente a Aceñolaza y Nieva (2001) incluirlos en su denominado tipo A: niveles arenoso-margosos de origen tempestítico. Si bien este tipo de acumulación ha sido reconocida en numerosas oportunidades en el intervalo de transición cambro/ordovícico, el caso particular del material aquí analizado es destacable, ya que ha resultado portador de una abundante y novedosa asociación de conodontes.
Este tipo de acumulación bioclástica se reconoce como una fuente excepcional de información faunística, por el carácter particular de sedimentación monoepisódica y repentina, que suele preservar materiales que de otra forma no se hubieran fosilizado (Emig, 1986; Emig y Gutiérrez-Marco, 1997; Aceñolaza et al. , 2003b).
Los materiales bioclásticos de las coquinas de la Formación Rupasca presentan una escasa selección, elevada fragmentación, desorganización interna de sus componentes, y se encuentran asociados a areniscas y lutitas en niveles tempestíticos que, en general, no superan los 25-30 cm de espesor. Las capas suelen presentar una base erosiva de granulometría más gruesa y un carácter granodecreciente hacia el techo. Las mismas presentan variaciones laterales tanto en su espesor como en los porcentajes de participación bioclástica. Asimismo, es frecuente reconocer una cementación secundaria margosa-carbonática que le confiere al nivel tintes amarillentos y una apariencia de escasa litificación.
Las areniscas que incluyen estas acumulaciones bioclásticas son psamitas cuarzo-feldespáticas que se presentan texturalmente bastante inmaduras, constituidas por clastos de cuarzos angulosos con extinciones normales y bordes irregulares que denotan disolución y esporádicamente crecimiento autigénico. Los feldespatos calcosódicos se presentan alterados parcialmente a caolinita, pistacita y zoicita. El feldespato potásico es proporcionalmente minoritario, y también puede presentarse alterado a caolinita y sericita. Adicionalmente, también se reconocieron líticos plutónicos mayoritariamente ácidos, metamórficos de bajo grado y sedimentarios. Los escasos filosilicatos reconocidos están representados por muscovita, biotita parda y clorita. El circón constituye un accesorio frecuente, así como los opacos referidos principalmente a óxidos de hierro.
La composición taxonómica de los niveles coquinoideos presenta un carácter politáxico, con bioclastos conformados mayoritariamente por exuvios fragmentados de trilobites, braquiópodos, equinodermos y moluscos, así como los conodontes analizados en esta oportunidad (Aceñolaza y Nieva, 2001).
Dada la naturaleza de este tipo de coquinas, se las considera como acumulaciones del tipo para-autóctonas (Aceñolaza y Nieva, 2001) y de origen mixto ( sensu Kidwell et al ., 1986), siendo producto de uno o más procesos de acumulación, donde se incluye un retrabajo y transporte previo a su depositación definitiva en forma de un evento sedimentario único ( event concentration de Kidwell, 1991). Si bien la asociación de conodontes registrada no incorpora taxones de edades disímiles, la génesis de estas coquinas prevé la posibilidad de que se agreguen elementos alóctonos en su composición, en caso de que parte del material transportado proceda de la erosión de rocas más antiguas en afloramientos contiguos a la cuenca ( cf ., Zeballo et al ., 2003, 2005a,2005b).
Registro de conodontes
Los conodontes recuperados de la Formación Rupasca proceden de la parte media a superior de la sección del corte del ferrocarril, en la localidad de Chucalezna, con un intervalo de muestreo de aproximadamente 1 m (figura 2). Se tomaron 5 muestras de coquinas carbonáticas para conodontes, con masas de 1 a 3 kg cada una; la más alta de las cuales dista ca . 5 m al contacto por falla del techo del afloramiento. Todas las muestras fueron completamente procesadas mediante métodos convencionales de laboratorio (ácido acético concentrado al 10%; Stone, 1987), y produjeron un total de 585 conodontes (figura 3). Estos microelementos presentan un buen estado de conservación, con una coloración marrón oscura correspondiente al índice de alteración del color 3 (CAI 3 sensu Epstein et al ., 1977), que representa paleotemperaturas de soterramiento de hasta 200ºC. Todos los especimenes ilustrados en las figuras 4 y 5 fueron fotografiados mediante microscopio electrónico de barrido, y la colección estudiada se encuentra depositada en el Museo de Paleontología de la Universidad Nacional de Córdoba, bajo la sigla de repositorio CORD-MP.

Figura 3. Distribución de conodontes en las muestras procesadas de la Formación Rupasca, sección de Chucalezna / Conodont distribution in samples of the Rupasca Formation, Chucalezna section.

Figura 4. A-F, Drepanoistodus chucaleznensis n. sp. A, elemento P, vista lateral externa / P element, outer lateral view , CORD-MP 10401, x 70. B, elemento Sd, vista lateral externa / Sd element, outer lateral view , CORD-MP 10402, x 70. C, elemento Sb, vista lateral interna / Sb element, inner lateral view, CORD-MP 10403, x 70. D, elemento M, vista lateral externa / M element, outer lateral view, CORD-MP 10060, x 70. E, elemento Sa, vista lateral externa / Sa element, outer lateral view , CORD-MP 10059, x 70. F, elemento Sc, vista lateral externa / Sc element, outer lateral view, CORD-MP 10404, x 120. G-H, Drepanodus arcuatus Pander, 1856. G, elemento f, vista lateral interna / f element, inner lateral view , CORD-MP 10064, x 120. H, elemento c, vista lateral interna / c element, inner lateral view , CORD-MP 10145, x 70. I-O, Utahconus humahuacensis n. sp. I, elemento f, vista lateral interna / f element, inner lateral view , CORD-MP 10067, x 90. J, elemento f, vista lateral externa / f element, outer lateral view , CORD-MP 10543, x 130. K, elemento e, vista lateral interna / e element, inner lateral view, CORD-MP 10544, x 100. L, elemento c, vista posterior / c element, posterior view, CORD-MP 10545, x 90. M, elemento b, vista lateral / b element, lateral view , CORD-MP 10546, x 100. N, elemento a, vista lateral / a element, lateral view , CORD-MP 10547, x 90. O, elemento a, vista lateral / a element, lateral view , CORD-MP 10548, x 130.

Figura 5. A-H, Paltodus deltifer pristinus (Viira). A, elemento Pb, vista lateral interna / Pb element, inner lateral view , CORD-MP 10204, x 75. B, elemento M, vista lateral externa / M element, outer lateral view , CORD-MP 10063, x 75. C, elemento Pa, vista lateral externa / Pa element, outer lateral view, CORD-MP 10205, x 75. D, elemento Pb, vista lateral externa / Pb element, outer lateral view, CORD-MP 10206, x 75. E, elemento Sd, vista lateral externa / Sd element, outer lateral view , CORD-MP 10207, x 75. F, elemento Sc, vista lateral externa / Sc element, outer lateral view, CORD-MP 10208, x 75. G, elemento Sc, vista lateral externa / Sc element, outer lateral view , CORD-MP 10209, x 75. H, elemento Sa, vista lateral externa / Sa element, outer lateral view, CORD-MP 10210, x 100. I-J, Semiacontiodus sp. I, elemento f, vista lateral interna / f element, inner lateral view , CORD-MP 10297, x 130. J, elemento a, vista póstero-lateral / a element postero-lateral view, CORD-MP 10066, x 130. K, Rossodus tenuis (Miller). elemento b, vista posterior / b element, posterior view , CORD-MP 10061, x 100. L-M, Teridontus nakamurai (Nogami). L, elemento a, vista lateral / a element, lateral view, CORD-MP 10065, x 100. M, elemento b, vista lateral / b element, lateral view , CORD-MP 10306, x 150. N, Paltodus cf. subaequalis (Pander). elemento Pa, vista lateral externa / Pa element, outer lateral view , CORD-MP 10068, x 130.
Bioestratigrafía
La presencia del conodonte guía Paltodus deltifer pristinus en la sección de Chucalezna refiere a la Zona de Paltodus deltifer , de acuerdo a la revisión efectuada por Löfgren (1997) sobre la definición original de Lindström (1971) para el esquema del reino Atlántico. La Zona de Paltodus deltifer fue definida originalmente en Suecia, donde se corresponde con la fauna de Ceratopyge Limestone (Piso AIII, Varangu, de la Serie Oelandiana inferior) de Västergötland y Öland (Bagnoli et al ., 1988). Esta unidad se correlaciona, a su vez, con el Tremadociano superior bajo, en transición entre los Pisos Cressagiano y Migneintiano de la cronoestratigrafía británica (Fortey et al ., 2000). La Zona de Paltodus deltifer fue subdividida por Löfgren (1997) en dos intervalos: la Subzona de Paltodus deltifer pristinus (inferior) y la Subzona de Paltodus deltifer deltifer (superior); unidades caracterizadas individualmente por la presencia excluyente de las respectivas subespecies. Otra característica que diferencia ambas unidades es la ausencia del género Cordylodus en la subzona superior.
Las especies asociadas a Paltodus d. pristinus en el área de Chucalezna también se registran en los niveles equivalentes inferiores de la Formación Rupasca, expuesta en el área de Alfarcito, al este de Tilcara, en el ámbito de la Cordillera Oriental de Jujuy (Zeballo et al ., 2003, 2005a, 2005b). Según los registros actuales, las nuevas especies que se describen son endémicas de esta región. A su vez, otros taxones tales como Teridontus nakamurai (Nogami) y Drepanodus arcuatus Pander, son formas con extensa distribución vertical. En diversas localidades del mundo, la superposición de rangos de estas especies corresponde al registro tardío de la primera y la aparición más temprana de la segunda. Paltodus cf. subaequalis Pander se ha documentado en Suecia, restringida a la Zona de Paltodus deltifer y parte baja de la suprayacente Zona de Paroistodus proteus (Löfgren, 1997). Rossodus tenuis (Miller), una especie originalmente descripta en América del Norte, tendría sus registros tardíos en estas unidades. La edad indicada por la asociación faunística descripta, que corresponde a la Subzona de Paltodus deltifer pristinus , es tremadociana tardía baja.
El espacio estratigráfico representado por la Zona de P. deltifer tiene un amplio registro global. Esta biozona fue descripta en numerosas oportunidades y con particular detalle en Suecia (Lindström, 1971; van Wamel, 1974; Bagnoli et al ., 1988; Löfgren, 1997); también se registró en Estonia (Viira, 1970), Polonia (Szaniawski, 1980), Letonia (Dubinina, 1983), y Rusia (Tolmacheva, 2001). En América del Norte se correlaciona con el Intervalo de Baja Diversidad y Zona de Macerodus dianae (= Fauna D de Ethington y Clark, 1971) establecida por Ross et al ., 1997 ( cf . Ji y Barnes, 1994a).
La Zona de Paltodus deltifer fue identificada por primera vez en Argentina por Keller et al . (1994), en la sección del cerro La Silla, Precordillera de San Juan. Lehnert (1995) registró una serie de taxones en la parte superior de la Formación La Silla, cuya distribución vertical relaciona con esta zona. Heredia (1995) documentó una asociación de conodontes del conglomerado basal de la Formación Empozada, Precordillera de Mendoza, que atribuye a la misma unidad, si bien no se registró la presencia de la especie epónima. Manca et al . (1995) mencionaron esta unidad para la Formación Santa Rosita en la localidad de Nazareno, Cordillera Oriental de Salta. Una fauna compuesta por Paltodus deltifer y especies de Drepanodus , Drepanoistodus , Paroistodus , Teridontus y Utahconus , fue descripta por Rao y Flores (1998) para los niveles superiores de la Formación Acoite en la Sierra de Aguilar, Cordillera Oriental de Jujuy. La Zona de P. deltifer fue formalmente definida para la Precordillera argentina en la Formación La Silla, sección portezuelo Yanso del cerro Potrerillo, provincia de San Juan, por Albanesi et al . (1998). La Formación San Jorge, expuesta en el sector central de la provincia de La Pampa, se desarrolla a través de un intervalo equivalente a la Zona de Paltodus deltifer (Albanesi et al ., 2003). Recientemente, se dio a conocer la presencia de P. deltifer pristinus en la Formación Bordo Atravesado, aflorante en la quebrada de La Alumbrera, Sistema de Famatina, Argentina (Albanesi et al ., 2005).
Consideraciones paleoambientales y paleobiogeográficas
La fauna de conodontes de la Formación Rupasca (o estratos superiores equivalentes de la Formación Santa Rosita) incluye componentes recurrentes de las faunas tremadocianas de la Cordillera Oriental; e.g ., en las secuencias de la quebrada de Incamayo, provincia de Salta (Rao y Tortello, 1998), sierra de Cajas (Rao, 1999), y área de Alfarcito (Zeballo et al ., 2003, 2005a, 2005b), provincia de Jujuy. Esta fauna combina formas endémicas con otras coetáneas de la región Báltica o del reino Atlántico (Löfgren, 1997), y también incluye algunos elementos característicos del reino Midcontinent, como los géneros Rossodus y Utahconus (Ji y Barnes, 1994b). El espacio bioestratigráfico representado por los escasos taxones que se documentan se corresponde con el Intervalo de Baja Diversidad reconocido particularmente en América del Norte (Ethington et al ., 1987). En este lapso se produce un reemplazo faunístico por comunidades con baja diversidad taxonómica, siguiendo la somerización marina progresiva (Evento Regresivo Ceratopyge ) que implica un evento de extinción de conodontes de incidencia global (Albanesi y Bergström, 2004). La composición faunística local sería producto de mezcla de formas propias de los reinos fríos y cálidos propuestos por Miller (1984), que comenzarían a diferenciarse a partir del Tremadociano tardío. Rao (1999) asigna la fauna de conodontes presente en la sierra de Cajas al reino faunístico de Transición, que Dubinina (1991) define para latitudes bajas a medias. Zeballo et al . (2005a, 2005b) arriban a una interpretación similar para las faunas tremadocianas de la misma cuenca. Aparentemente, los paleoambientes coetáneos de las formaciones Volcancito y Bordo Atravesado habrían estado circunscriptos a condiciones paleogeográficas comparables (Albanesi et al ., 1999, 2005).
Si bien la fauna de conodontes de la Zona de Paltodus deltifer en la Precordillera argentina, registra la especie epónima, propia del reino Atlántico, se diferencia de las faunas tremadocianas tardías del noroeste argentino por el predominio de géneros que dominan en ambientes del reino Midcontinent, tales como Ulrichodina (= Colaptoconus ) (Ji y Barnes, 1994b). Por ejemplo, la asociación de conodontes registrada en el tramo superior de la Formación La Silla en la sección del cerro San Roque, incluye las especies Ulrichodina quadraplicata (Branson y Mehl), Laurentoscandodus triangularis (Furnish) y Paltodus deltifer deltifer (Lindström) (Cañas et al ., 2002). Asimismo, la constitución de la fauna contemporánea de la extensión austral del terreno de Cuyania, registrada en la Formación San Jorge, provincia de La Pampa, es representativa del mismo intervalo de baja diversidad, característico del ambiente Midcontinent de Laurentia (Albanesi et al ., 2003).
Paleontología sistemática
Purnell et al. (2000) recientemente propusieron un nuevo esquema de nomenclatura anatómica para los aparatos multielementales de conodontes. No obstante, en razón del escaso entendimiento actual de la arquitectura de los aparatos del Ordovícico Inferior, se mantienen esquemas de orientación y notación convencionales. Para la descripción de las especies con morfotipos oistodiformes (coniformes geniculados) se sigue la clasificación de Sweet (1981, 1988), que incluye un elemento de posición espacial anterior (M), una serie de transición en simetría (Sa -simétrico-, Sb, Sc, Sd) y elementos posteriores (P). Para los morfotipos de aparatos con elementos coniformes non geniculate , de posición análoga al oistodiforme, se aplica la nomenclatura propuesta por Ji y Barnes (1994a), que distingue elementos a, b, c, e y f, donde el morfotipo c es simétrico, y los elementos e y f ocuparían las posiciones extremas anterior y posterior, respectivamente. En la clasificación sistemática se adoptan las categorías propuestas por Sweet (1988) y revisadas por Sweet y Donoghue (2001).
Clase C ONODONTA Pander, 1856
Género Drepanodus Pander, 1856
Especie tipo. Drepanodus arcuatus Pander, 1856.
Drepanodus arcuatus Pander, 1856
Figuras 4.G-H
1856. Drepanodus arcuatus n. sp. Pander, p. 20, pl. 1, figs. 2, 4, 5, 17. Sinonimia multielemental
1994. Drepanodus arcuatus Pander; Dzik, pp. 68-70, pl. 15, figs. 2-6; text-fig. 8a-b ( cum syn. ).
2003. Drepanodus arcuatus Pander; Löfgren y Tolmacheva, pp. 11-15; figs 2, 3 A-C, E-H, 5 K-V, 6 M-U, 8 A-G ( cum syn. ).
Material. 59 elementos. CORD-MP 10064, 10145-10203.
Observaciones. Drepanodus arcuatus Pander y otras formas vinculadas taxonómicamente fueron revisadas detalladamente por Löfgren y Tolmacheva (2003). Las autoras verificaron una estructura septimembrate para el aparato oral de la especie.
Procedencia. Muestras CH10A, CH10B, CH1, AR5.
Distribución. Drepanodus arcuatus presenta una amplia distribución espacial y temporal, habiéndose registrado, virtualmente, en todos los continentes, particularmente en las facies de aguas frías o profundas del Ordovícico Inferior y Medio; en la región baltoescandinava ( e.g ., Pander, 1856; Löfgren y Tolmacheva, 2003), Polonia (Dzik, 1994), China ( e.g ., An, 1987), Argentina ( e.g ., Albanesi et al ., 1998), América del Norte (Landing y Ludvigsen, 1984).
Género Drepanoistodus Lindström, 1971
Especie tipo. Oistodus forceps Lindström, 1955.
Observaciones. El aparato multielemental de Drepanoistodus incorpora cinco morfotipos característicos, de acuerdo con las reconstrucciones proporcionadas por Stouge (1984) y Dzik (1994). La nomenclatura tipológica tradicionalmente empleada en la descripción de los morfotipos (véase Albanesi, 1998) se complementa con la atribución de posiciones espaciales en los aparatos, según el esquema de Sweet (1988). Si bien esta asignación no verifica necesariamente homología con la posición de los elementos ramiformes o pectiniformes en aparatos evolucionados, la presencia del morfotipo oistodiforme, diagnóstico a nivel específico (Stouge y Bagnoli, 1988), permite fijar una posición extrema a partir de la cual ordenar los elementos que constituyen la serie de transición en simetría y otras formas discretas; i.e ., oistodiforme M, suberectiforme Sa, planiforme o acodiforme Sb, homocurvatiforme o drepanodiforme Sc y paltodiforme o scandodiforme P.
Drepanoistodus chucaleznensis n. sp.
Figuras 4.A-F
Holotipo. Elemento oistodiforme modificado (M), CORD-MP 10060 (figura 4.D).
Material. 18 elementos M, 35 elementos Sa, 30 elementos Sb, 43 elementos Sc, 17 elementos P. CORD-MP 10059-60, 10401-10542.
Derivatio nominis . En referencia a la localidad tipo, Chucalezna.
Localidad tipo y estratotipo. Chucalezna, Cordillera Oriental, departamento Humahuaca, provincia de Jujuy, Argentina. Sección de Chucalezna (corte de ferrocarril), nivel CH10B.
Diagnosis. El elemento de posición M es un morfotipo oistodiforme, modificado, en cuanto presenta una suave transición entre el margen oral y el margen posterior de la cúspide; i.e ., non geniculate . En vista lateral, la base es triangular, y el margen anterior del elemento tiene un punto de inflexión a nivel del margen oral, delineando una cúspide reclinada.
The M element is a modified oistodiform morphotype, which is characterized by a gentle transition between the oral margin and the posterior margin of the cusp; i.e., non geniculate. Its lateral view shows a triangular base, and a flexural point at the level of the oral margin, determining a reclined cusp.
Descripción. En el morfotipo oistodiforme, de posición M, el margen oral de la base continúa sin solución de continuidad hacia el margen posterior de la cúspide. Los márgenes anterior, posterior y oral son aquillados. La cúspide es reclinada y se flexiona a nivel del margen oral. La configuración de la base es triangular. El ángulo entre el margen oral y la cúspide es de 45º-50º. El margen aboral es suavemente convexo, y en el flanco externo es convexo y presenta una suave indentación próxima al ángulo ánterobasal. Asimismo, el flanco interno es convexo, si bien presenta sendos surcos paralelos a los márgenes anterior y posterior. La base se expande hacia la parte posterior en su flanco interno. La extensión del margen oral es aproximadamente 1/3 de la longitud del margen basal. La cavidad basal es somera y su ápice se dirige hacia el eje de la cúspide, alcanzando el nivel del margen oral.
El morfotipo scandodiforme, de posición P, se caracteriza por una amplia base acampanada. El ángulo entre el margen oral y la cúspide es de ca. 90º. La cúspide se flexiona ligeramente hacia la parte interior. El flanco interno está ligeramente deprimido hacia el margen anterior.
El morfotipo homocurvatiforme, Sc, tiene márgenes aquillados, y el margen anterior se presenta comprimido en el ángulo ántero-basal.
El morfotipo paltodiforme, Sb, presenta márgenes aquillados, flanco externo convexo, flanco interno plano a ligeramente convexo, y margen basal sinusoide.
El elemento de posición Sa es suberecto, su margen oral forma un ángulo recto a ligeramente obtuso con la cúspide. Los márgenes anterior y posterior son aquillados. El margen ántero-basal es recto y no se proyecta hacia la parte anterior. Los flancos de la cúspide son suavemente convexos y la base es acampanada. El ápice de la cavidad basal se proyecta hacia el eje del elemento.
Observaciones. Los elementos descriptos son hialinos con escasa materia blanca restringida al sector apical de la cúspide. Si bien la asignación genérica de este nuevo taxón se verifica por la presencia del morfotipo suberectiforme Sa, el elemento de posición M no es típicamente oistodiforme como en las especies contemporáneas documentadas para ambientes disímiles de Laurentia, Báltica, China o Corea ( e.g ., Ji y Barnes, 1994a; Stouge y Bagnoli, 1988; Löfgren, 1997; An et al ., 1983; Seo et al ., 1994). La característica forma non geniculate del morfotipo M es diagnóstica para la nueva especie. Estos elementos tienen un aspecto ligeramente pipaforme en estadios ontogenéticos avanzados.
De acuerdo al presente registro, el aparato multielemental de esta nueva especie tendría la siguiente relación numérica entre los morfotipos constituyentes: 2 M, 4 Sa, 4 Sb, 4 Sc, 2 P.
En secciones estratigráficas del área de Alfarcito, Cordillera Oriental de Jujuy, Argentina, se registró una nueva forma de Drepanoistodus , procedente de niveles inmediatamente subyacentes a los de la Zona de Paltodus deltifer , cuyas características morfológicas sugieren la presencia de la forma ancestral a la especie aquí descripta (Zeballo et al ., 2005a, 2005b).
Discusión. Si bien el elemento M de la nueva especie es típicamente non geniculate , presenta características semejantes a las de Drepanoistodus concavus (Branson y Mehl), según la detallada descripción brindada por Kennedy (1980). No obstante, los elementos de la serie de transición en simetría S, y particularmente el elemento scandodiforme P, manifiestan caracteres distintos a los de aquellos elementos ilustrados por el autor citado y por Ji y Barnes (1994a), que permiten una nueva asignación específica.
Procedencia. Muestras CH1, CH5, CH10, diez metros cuspidales de la Formación Santa Rosita en la sección de Chucalezna. Subzona de Paltodus deltifer pristinus de la Zona de Paltodus deltifer , Tremadociano tardío.
Distribución. Tremadociano superior de la Cordillera Oriental del noroeste argentino ( Drepanoistodus n. sp. A, en Zeballo et al ., 2005a, 2005b).
Género Paltodus Pander, 1856
Especie tipo. Paltodus subaequalis Pander, 1856.
Paltodus deltifer pristinus (Viira, 1970)
Figuras 5.A-H
1970. Drepanodus pristinus Viira, p. 227; figs. 5, 6; lám. 1, figs. 7, 8 (morfotipo Sa).
1980. Drepanoistodus deltifer pristinus (Viira); Szaniawski, p. 113; lám. 16-18, figs. 2, 7-11, 13 (taxonomía multielemental).
1997. Paltodus deltifer pristinus (Viira); Löfgren, pp. 263-264; figs. 5 P-Y, 6 A-G (taxonomía multielemental) ( cum syn. ).
Material. 10 elementos M, 8 elementos Sa, 12 elementos Sb, 12 elementos Sc, 16 elementos Sd, 21 elementos Pa, 14 elementos Pb. CORD-MP 10062, 10063, 10204-10296.
Observaciones. El aparato multielemental de esta especie fue descripto por Löfgren (1997), reconociendo 7 morfotipos: M, Pa, Pb, Sa, Sb, Sc y Sd. El morfotipo M caracteriza a la especie, por la disposición de la cúspide, suberecta, sin vértice definido en la transición al margen oral de la base (elemento no geniculado). Se registran todos los morfotipos, a excepción del elemento varanguensiforme descripto por la autora citada y por Szaniawski (1980).
El elemento M de Paltodus deltifer pristinus (Viira) se diferencia del homólogo de P. d . Deltifer (Lindström) por ser este último típicamente geniculado, y con un margen oral mas extendido. Hasta el presente, la única especie del género con el elemento M no geniculado es P. d. pristinus , cuyas características lo diferencian de otras especies, como por ejemplo: P. subaequalis Pander, P. perrii Stouge y Bagnoli, P . peracutus (Lindström) y P. Jemtlandicus Löfgren.
Procedencia. Muestras CH1, CH5, CH10B, CH10A.
Distribución. Paltodus d. pristinus se registra en la región baltoescandinava (Viira, 1970; Löfgren, 1997) y en las cuencas gondwánicas correspondientes al Sistema de Famatina (Albanesi et al ., en prensa), y Cordillera Oriental (Zeballo et al ., 2005a, 2005b) del noroeste argentino. La distribución estratigráfica de esta especie se limita a la parte inferior de la Zona de Paltodus deltifer , en el Tremadociano superior.
Paltodus cf. subaequalis (Lindström, 1955)
Figura 5.N
cf. 1856. Paltodus subaequalis Pander, p. 24; fig. 4a, lám. 1, fig. 24.
1985. Paltodus cf. subaequalis (Pander); Löfgren, p. 127, figs. 4 R-U.
1997. Paltodus cf. subaequalis (Pander); Löfgren, p. 265, figs. 5 AHAN.
Material. 1 elemento. CORD-MP 10068.
Observaciones. El único elemento registrado presenta una costilla desarrollada en la región basal del flanco externo, y es similar al morfotipo Pb? Ilustrado por Löfgren (1997). Esta especie presenta características intermedias entre Paltodus deltifer (Lindström) y P. subaequalis Pander.
Procedencia. Muestra CH10A.
Distribución. Esta especie se registra en Suecia (Löfgren, 1997) y en la Cordillera Oriental del noroeste argentino (Zeballo et al ., 2005a), y su rango cronoestratigráfico se restringe al Tremadociano superior.
Género Rossodus Repetski y Ethington, 1983
Especie tipo. Rossodus manitouensis Repetski y Ethington, 1983.
Rossodus tenuis (Miller, 1980)
figura 5.K
1980. Utahconus tenuis n. sp. Miller, pp. 36-37, pl. 2, figs. 5-7, textfig. 4T.
Sinonimia multielemental
1994a. Rossodus tenuis (Miller); Ji y Barnes, p. 56, pl. 17, figs. 10-19 ( cum syn. ).
2002. Rossodus tenuis (Miller); Pyle y Barnes, pp. 102-103, lám. 13, figs. 21-26 ( cum syn. ).
Material. 1 elemento. CORD-MP 10061.
Discusión. Ji y Barnes (1994a) interpretan un aparato multielemental para esta especie integrado por 3 morfotipos: a, subredondeado, c, simétrico suberecto, y e, comprimido. Estos autores ilustran un morfotipo b no descripto, con cúspide erecta a levemente proclinada, el margen anterior redondeado y el posterior aquillado.
El elemento e de Rossodus tenuis se diferencia del homónimo de R . manitouensis Repetski y Ethington (1983) por la breve expansión lateral de la base, la cual no forma un ángulo discernible con la cúspide; es decir, la transición desde la base a la cúspide es suave, según las formas no geniculadas. En el morfotipo c de R. tenuis , la cavidad basal es más profunda y la base se expande hacia posterior.
Procedencia. Muestra AR5.
Distribución. Rossodus tenuis se ha documentado en diversas localidades de América del Norte ( e.g ., Miller, 1980; Ji y Barnes, 1994a; Pyle y Barnes, 2002), y en la Cordillera Oriental del noroeste argentino (Zeballo et al ., 2005a). Su distribución vertical ocupa el Tremadociano medio.
Género Semiacontiodus Miller, 1969
Especie tipo. Acontiodus ( Semiacontiodus ) nogamii Miller, 1969.
Semiacontiodus sp.
Figuras 5.I-J
Material. 10 elementos. CORD-MP 10066, 10297-10306.
Observaciones. Los elementos de esta especie presentan una base relativamente larga y carecen de estrías, surcos, costillas, y otros caracteres ornamentales, que usualmente aparecen en las diversas especies de este género; cf. S. bulbosus (Löfgren), S. Cornuformis (Seergeva). Los escasos especímenes registrados no permiten la reconstrucción completa del aparato multiemelental para una diagnosis específica.
Procedencia. Muestras CH10A y CH10B.
Distribución. Aparentemente Semiacontiodus sp. Es una especie endémica, restringida a la Cordillera Oriental del noroeste argentino ( cf ., Zeballo et al ., 2005a).
Género Teridontus Miller, 1980
Especie tipo. Oneotodus nakamurai Nogami, 1967.
Teridontus nakamurai (Nogami, 1967) sensu lato
Figuras 5.L-M
1967. Oneotodus nakamurai Nogami, pp. 216-217, lám. 1, figs. 9, 12; text. figs. 3 A, B.
Sinomimia multielemental
1994a. Teridontus nakamurai (Nogami); Ji y Barnes, pp. 64-65, lám. 24, figs. 1-9, text.fig. 37 C ( cum syn .).
1994. Teridontus nakamurai (Nogami); Lehnert, pp. 255-256, lám. 1, figs. 5, 11 ( cum syn .).
1999. Teridontus nakamurai (Nogami); Albanesi et al ., lám. 1, figs. 11, 12.
2000. Teridontus nakamurai (Nogami); Tortello y Rao, p. 80, figs. 6 A, B, F, L; 7 D, G.
2002. Teridontus nakamurai (Nogami); Pyle y Barnes, p. 71, lám. 15, figs. 17-19.
Material. 94 elementos. CORD-MP 10065, 10306-10400.
Discusión. En la presente colección se identifican especimenes de Teridontus nakamurai según la interpretación de Ji y Barnes (1994a), quienes proponen para esta especie un aparato multielemental de 4 morfotipos: elementos nakamuriformes a, subredondeado, y b, transicional; c, erectiforme simétrico, y e, monocostodiforme comprimido. T. nakamurai (Nogami) se diferencia de T. obesus Ji y Barnes por su tamaño más pequeño y base menos expandida, y de T. Gracillimus Nowlan por un tamaño general de la corona relativamente mayor; en particular, una cúspide más desarrollada. Se considera a Teridontus nogamii en sentido amplio, en razón de que en la revisión de la especie efectuada por Nicoll (1994), el autor no incluye en su sinonimia a las formas ilustradas por Ji y Barnes (1994a) con la misma denominación. Según Nicoll (1994) el rango de Teridontus nogamii sensu stricto no alcanzaría el Tremadociano superior; por otra parte, su interpretación del aparato de la especie en base a especímenes procedentes del Cámbrico Superior incluye seis morfotipos, que no se verifican en la presente colección. Dadas estas diferencias, en el presente estudio se prefiere mantener una asignación tentativa, sensu lato .
Procedencia. Muestras CH1, CH5, CH10A, CH10B.
Distribución. Teridontus nakamurai sensu lato presenta una distribución cosmopolita (véase sinonimia), generalmente es muy abundante en diversas facies, y su rango cronoestratigráfico se extiende a través del Cámbrico Superior y del Tremadociano.
Género Utahconus Miller, 1980
Especie tipo. Paltodus utahensis Miller, 1969.
Diagnosis enmendada. Utahconus Miller presenta un aparato multielemental quinquimembrate. El elemento a es un morfotipo multicostate, con base subredondeada, ornamentada con carenas o costillas y surcos que se extinguen gradualmente hacia el ápice de la cúspide y el contorno basal. El elemento b es un morfotipo asimétrico, con márgenes ántero-lateral y póstero-lateral aquillados, y cúspide torsionada. El elemento c es un morfotipo staufferiforme, con agudas costillas laterales agudas. El elemento e es un morfotipo bicostate, con cúspide proclinada en el mismo plano que la base. El elemento f es un morfotipo acodiforme, caracterizado por una fuerte costilla en el flanco externo, que se extiende desde el ápice de la cúspide al margen basal. El mismo puede presentar la costilla lateral desplazada hacia posterior.
Utahconus Miller presents a quinquimembrate conodont apparatus. The a morphotype is a mulitcostate element, with subrounded base, and ornamented with carina or ridges and grooves that gradually extinguish to the tip of the cusp and the basal margin. The b element is an asymmetrical form, with sharp antero-lateral and posterolateral margins, and cusp torsion. The c element is staufferiform, with acute lateral costae. The e element is bicostate, with proclined cusp at the same plane of the base. The f element is acodiform, characterized by conspicuous costae on the outer flank, which extend from tip of the cusp to the basal margin. In same element the lateral costae can be displaced to posterior.
Observaciones. En la designación original del género Utahconus , Miller (1980) incluyó dos morfotipos, unicostate y bicostate, que representarían los elementos f y e, según esta enmienda. En una revisión previa de la diagnosis original, Ji y Barnes (1994a) incorporan tres morfotipos al aparato multielemental del género, agregando el elemento staufferiforme c, a los dos anteriores de Miller, que ellos identifican como a y e. Estos autores reconocen una gradación de formas entre los morfotipos que, si bien no precisada, podría incluir a los elementos f y b definidos aquí. Distintos morfotipos de Utahconus guardan similitud con elementos de los géneros coetáneos Rossodus Repetski y Ethington (1983), Variabiloconus Landing et al . (1986) y Polycostatus Ji y Barnes (1994a); no obstante, los planes de sus aparatos multielementales no demostrarían relaciones filogenéticas próximas sino, probablemente, convergencias evolutivas funcionales.
Utahconus humahuacensis n. sp.
Figuras 4.I-O
Holotipo. Elemento coniforme non-geniculado f, CORD-MP 10067 (figura 4.I).
Material. 50 elementos a, 34 elementos b, 11 elementos c, 27 elementos e, 63 elementos f. CORD-MP 10067, 10543-10728.
Derivatio nominis. En referencia a la quebrada de Humahuaca, donde se encuentra la localidad tipo.
Localidad tipo y estratotipo. Chucalezna, Cordillera Oriental, departamento Humahuaca, provincia de Jujuy, Argentina. Sección de Chucalezna (corte de ferrocarril), nivel CH10B.
Diagnosis. El morfotipo acodiforme f se caracteriza por presentar una prominente costilla a través del flanco externo, que recorre el elemento desde el ápice de la cúspide al margen basal. El margen basal es triangular en vista inferior.
The acodiform f element is characterized by the presence of prominent costae throughout the outer flank. This ornament goes over all the element longitude, from tip of cusp to basal margin. The basal margin is triangular on lower view.
Descripción. El elemento f es un morfotipo acodiforme o pastinate primitivo. La cúspide es robusta, proclinada y ligeramente flexionada hacia exterior. La base es corta en relación a la cúspide. El margen anterior es redondeado y el posterior es agudo. Una conspicua costilla se extiende desde el ápice de la cúspide hasta el margen aboral. Ésta ocupa una posición lateral, si bien en algunos elementos aparece desplazada hacia posterior. Los vértices del contorno triangular de la base coinciden con los márgenes anterior, posterior y la costilla lateral. La cavidad basal es amplia y su ápice se proyecta hacia la parte anterior.
El elemento e es un morfotipo coniforme non geniculado asimétrico, póstero-lateralmente comprimido. La cúspide es proclinada, en el mismo plano que la base, y su longitud es aproximadamente igual a la de ésta. Los márgenes ántero-lateral y póstero-lateral son aquillados, y ambos flancos son convexos. El flanco interno está recorrido por sendos surcos paralelos a los márgenes, y puede estar ornamentado con estrías o suaves surcos y costillas. La cavidad basal es cónica, con el ápice dirigido hacia el eje de la cúspide.
El elemento c es un morfotipo staufferiforme, bilateralmente simétrico, comprimido en sentido ántero-posterior, y cúspide proclinada. Los márgenes anterior y posterior son redondeados, y ambas costillas laterales recorren la longitud del elemento, desde el ápice de la cúspide al margen basal. El margen posterior puede presentar estrías o suaves costillas ornamentales. El contorno basal es elíptico, con eje mayor bilateral, y la cavidad basal, cónica, proyecta su ápice hacia el eje de la cúspide.
El elemento b es un morfotipo coniforme non geniculate asimétrico, con cúspide proclinada, torsionada con respecto a la base y de mayor longitud que ésta. Márgenes ántero-lateral y postero-lateral aquillados.
El elemento a es un morfotipo multicostate subsimétrico, con cúspide proclinada y base cónica. Esta forma se caracteriza por costillas y surcos que, en número variable, la ornamentan en todo su contorno. Los caracteres son fuertes, extinguiéndose gradualmente hacia contorno basal y el ápice de la cúspide. El contorno basal es subredondeado, y orlado por efecto de las costillas y surcos.
Observaciones. Los elementos son hialinos, con escasa materia blanca concentrada en el sector apical de la cúspide. En nuestra colección se registran algunas formas similares al elemento f, pero con la costilla lateral desplazada hacia la parte posterior, pudiendo representar una variación ontogenética o un nuevo morfotipo, si bien la colección actual no permite verificar estas alternativas.
Según el registro actual, el aparato multielemental de esta especie nueva incorporaría los diversos morfotipos en la siguiente relación numérica: 6 a, 3 b, 1 c, 3 e, 6 f.
Esta nueva forma de Utahconus se distingue de otras especies bien documentadas en Laurentia ( e.g ., Miller, 1980; Nowlan, 1985; Orndorff, 1988; Ji y Barnes, 1994a), como U. utahensis (Miller) y Utahconus longipinatus Ji y Barnes, por el grado de desarrollo de su base, siendo menor que en la primera y mayor que en la segunda.
Procedencia. Muestras CH1, CH5, CH10, diez metros cuspidales de la Formación Santa Rosita en la sección de Chucalezna. Subzona de Paltodus deltifer pristinus de la Zona de Paltodus deltifer , Tremadociano tardío.
Distribución. Tremadociano superior de la Cordillera Oriental del noroeste argentino ( Utahconus n. sp. A, en Zeballo et al ., 2005a, 2005b).
Agradecimientos
Los autores agradecen al CONICET por el constante apoyo al estudio de las faunas del Paleozoico inferior del noroeste argentino. G. Ortega y M.F. Tortello participaron en las tareas de campo. A. Andrada Barone atendió el microscopio electrónico de barrido del LAMENOA, Tucumán, para la fotografía de conodontes. E. Gómez Hasselrot asistió en la confección de las ilustraciones. Se agradece la revisión del manuscrito original efectuada por los árbitros.
Bibliografía
Aceñolaza, F.G. (ed.). 2002. Aspects on the Ordovician System in Argentina. INSUGEO Tucumán, Serie Correlación Geológica 16: 1-370. [ Links ]
Aceñolaza, G.F. 1996. [ Bioestratigrafía del límite Cámbrico-Ordovícico y Ordovícico basal en la Quebrada de Humahuaca, Provincia de Jujuy, Rep. Argentina. Tesis Doctoral, Facultad de Ciencias Naturales e IML, Universidad Nacional de Tucumán, 245 pp. Inédita]. [ Links ]
Aceñolaza, G.F. 2001. El Angosto de Chucalezna: una localidad clásica para el estudio de icnofósiles en el noroeste argentino. 4º Reunión Argentina de Icnología y 2º Reunión de Icnología del Mercosur. Resúmenes : 19. [ Links ]
Aceñolaza, G.F. y Albanesi, G.L. 1996. Conodont-trilobite biostratigraphy of the Santa Rosita Formation from Chucalezna, Cordillera Oriental, Northern Argentina. Reunión de Comunicaciones de la Asociación Paleontológica Argentina (Córdoba), Ameghiniana 34: 114. [ Links ]
Aceñolaza, G.F. y Nieva, S.M. 2001. Sobre algunas acumulaciones fosilíferas en el Ordovícico Inferior de la Cordillera Oriental Argentina. Boletín Geológico y Minero 112: 35-42. [ Links ]
Aceñolaza, G.F. y Poiré, D. 1999. Trace fossils and sedimentology of Rupasca Formation (Lower Ordovician) in Chucalezna, Jujuy Province, northern Argentina. Acta Universitatis Carolinae - Geologica 43: 159-162. [ Links ]
Aceñolaza, G.F., Aráoz, L., Poiré, D., Vergel, M.M. y Albanesi, G. 2003a. Biostratigraphical and sedimentological aspects of the Cambro-Ordovician strata at the Angosto de Chucalezna: new palynological data (Jujuy, NW Argentina). INSUGEO Tucumán, Miscelánea 9: 13-21. [ Links ]
Aceñolaza, G.F., Emig, C.C. y Gutiérrez-Marco, J.C. 2003b. Lingulid shell beds from the Ordovician of Argentina, with notes on other peri-Gondwanan occurrences. En: G.L. Albanesi, M.S. Beresi y S. Peralta (eds.), Ordovician from the Andes. INSUGEO Tucumán, Serie Correlación Geológica 17: 237-244. [ Links ]
Albanesi, G.L. 1998. Taxonomía de conodontes de las secuencias ordovícicas del cerro Potrerillo, Precordillera Central de San Juan, R. Argentina. Actas 12º Academia Nacional de Ciencias , Córdoba: 101-253. [ Links ]
Albanesi G.L. y Bergström, S.M. 2004. Conodonts: Lower to Middle Ordovician record. En: B.D. Webby, F. Paris, M.L. Droser y I.G. Percival (eds.), The Great Ordovician Biodiversification Event , Columbia University Press, New York: 312-336. [ Links ]
Albanesi, G.L., Hünicken, M.A. y Barnes, C.R. 1998. Bioestratigrafía de conodontes de las secuencias ordovícicas del cerro Potrerillo, Precordillera Central de San Juan, R. Argentina. Actas 12º Academia Nacional de Ciencias , Córdoba: 7-72. [ Links ]
Albanesi, G.L., Esteban, S.B. y Barnes, C.R. 1999. Conodontes del intervalo del límite Cámbrico-Ordovícico en la Formación Volcancito, Sistema de Famatina, Argentina. Temas Geológico-Mineros ITGE , Madrid, 26: 521-526. [ Links ]
Albanesi, G.L., Bergström, S.M. y Melchor, R.N. 2003. The San Jorge Formation, La Pampa Province, Argentina, dated by means of conodonts. Ameghiniana Suplemento Resúmenes 40 : 77-78R. [ Links ]
Albanesi, G.L., Esteban, S.B., Ortega, G., Hünicken, M.A. y Barnes, C.R. 2005. Bioestratigrafía y ambientes sedimentarios de las Formaciones Volcancito y Bordo Atravesado (Cámbrico Superior-Ordovícico Inferior), Sistema de Famatina, Provincia de La Rioja, Argentina. En: J.A. Dalhquist, C.W. Rapela y E. Baldo (eds.), Geología de la Provincia de La Rioja (Precámbrico-Paleozoico Inferior). Edición especial de la Asociación Geológica Argentina. [ Links ]
An, T.-X. 1987. The Lower Paleozoic conodonts of South China. Peking University Publication House , Beijing: 1-238. [ Links ]
An, T.-X.; Zhang, F.; Xiang, W.; Zhang, Y.; Xu, W.; Zhang, H.; Jiang, D.; Yang, C.; Lin, L.; Cui, Z. y Yang, X. 1983. The conodonts of North China and the adjacent regions. Science Press of China , Beijing: 1-223. [ Links ]
Bagnoli, G., Stouge, S. y Tongiorgi, M. 1988. Acritarchs and conodonts from the Cambro Ordovician Furuhall (Köpingsklint) Section (Öland,Sweden). Rivista Italiana de Paleontología Stratigraphia 94: 163-248. [ Links ]
Benedetto, J.L. (ed.). 2003. Ordovician Fossils of Argentina, Secretaría de Ciencias y Tecnología, Universidad Nacional de Córdoba: 560 pp. [ Links ]
Branson, E.B. y Mehl, M.G. 1933. Conodont studies, numbers 1 and 2. University of Missouri Studies 19: 1-535. [ Links ]
Buatois, L.A. y Mángano, G. 2003. Sedimentary facies, depositional evolution of the Upper Cambrian-Lower Ordovician Santa Rosita Formation in northwest Argentina. Journal of South American Earth Sciences 16: 343-363. [ Links ]
Cañas, F.L, Carrera, M.G. y Albanesi, G.L. 2002. El evento eustático Ceratopyge en la Precordillera Argentina y su relación con la radiación ordovícica. Reunión de la Asociación Argentina de Sedimentología (Córdoba). Resumen : 51. [ Links ]
Dubinina, S.V. 1983. K stratigrafii po konodontam niznego i nov srednego Ordovika severo-zapadnoj Latvii (Conodont stratigraphy of the Lower Ordovician and the lower part of the Middle Ordovician in North-West Latvia). Eesti Teaduste Akadeemia, Geoloogia 32: 45-52. [ Links ]
Dubinina, S.V. 1991. Upper Cambrian and Lower Ordovician conodont associations from open ocean paleoenvironments, illustrated by Batyrbay and Sarykum sections in Kazakhstan. In: Barnes, C.R. and Williams, S.H. (eds.). Advances in Ordovician Geology. Geological Survey of Canada, Paper 90-9: 107-124. [ Links ]
Dzik, J. 1994. Conodonts of the Mójcza Limestone. Palaeontologia Polonica 53: 43-128. [ Links ]
Emig, C.C. 1986. Conditions de fossilisation du genre Lingula (Brachiopoda) et implications paléoécologiques. Palaeogeography, Palaeoclimatology, Palaeoecology 53: 245-253. [ Links ]
Emig, C.C. y Gutiérrez-Marco, J.C. 1997. Signification des niveaux á lingulidés á la limite supérieur du Grés armoricain (Ordovicien, Arénig, Sud-Ouest de l´Europe). Geobios 30: 481-495. [ Links ]
Epstein, A.G., Epstein, J.B. y Harris, L.D. 1977. Conodont color alteration - An index to organic metamorphism. United States Geological Survey Professional Paper 995: 1-27. [ Links ]
Ethington, R.L. y Clark, D.L. 1971. Lower Ordovician conodonts in North America. En: W.C. Sweet y S.M. Bergström (eds.), Symposium on Conodont Biostratigraphy. Geological Society of America Memoir 127: 63-82. [ Links ]
Ethington, R.L., Engel, K.M.y Elliott, K.L. 1987. An abrupt change in conodont faunas in the Lower Ordovician of the Midcontinent Province. En: R.J. Aldridge (ed.), Palaeobiology of Conodonts. Ellis Horwood Limited, Chichester: 111-127. [ Links ]
Fortey, R.A., Harper, D.A.T., Ingham, J.K., Owen, A.W., Parkes, M.A., Rushton, A.W.A.y Woodcock, N.H. 2000. A revised correlation of Ordovician rocks in the British Isles. The Geological Society, Special Report 24: 1-83. [ Links ]
Frey, R.W., Pemberton, S.G. y Saunders, T.D. 1990. Ichnofacies and bathymetry: a passive relationship. Journal of Paleontology 64: 155-158. [ Links ]
Harrington, H.J. 1937. On some Ordovician fossils from northern Argentina. Geological Magazine 74, 873: 97-124. [ Links ]
Harrington, H.J. 1938. Sobre las faunas del Ordoviciano inferior del norte argentino. Revista del Museo de La Plata (Paleontología) 4: 209-289. [ Links ]
Harrington, H.J. 1957. Ordovician formations of Argentina. En: H.J. Harrington y A.F. Leanza (eds.), Ordovician Trilobites of Argentina, Department of Geology, University of Kansas, Special Publication 1: 1-59. [ Links ]
Harrington, H.J. y Leanza, A.F. 1957. Ordovician Trilobites of Argentina. Department of Geology, University of Kansas, Special Publication 1: 276 pp. [ Links ]
Heredia, S.E. 1995. Conodontes Cámbricos y Ordovícicos en los bloques alóctonos del conglomerado basal de la Formación Empozada, Ordovícico Medio-Superior, San Isidro, Precordillera de Mendoza, Argentina. Boletín de la Academia Nacional de Ciencias , Córdoba, 60: 235-247. [ Links ]
Ji, Z. y Barnes, C.R. 1994a. Lower Ordovician conodonts of the St. George Group, Port au Port Peninsula, western Newfoundland, Canada. Palaeontographica Canadiana , 11: 1-149. [ Links ]
Ji, Z. y Barnes, C.R. 1994b. Conodont paleoecology of the Lower Ordovician St. George Group, Port au Port Peninsula, western Newfoundland. Journal of Paleontology 68: 1368-1383. [ Links ]
Kayser, E. 1925. Contribuciones a la Paleontología de la República Argentina. Sobre fósiles primordiales e infrasilurianos. Boletín de la Academia Nacional de Ciencias , Córdoba, 9: 297-332. [ Links ]
Keller, M., Cañas, F., Lehnert, O. y Vaccari, N.E. 1994. The Upper Cambrian and Lower Ordovician of the Precordillera (Western Argentina): Some stratigraphic reconsiderations. Newsletter on Stratigraphy 31: 115-132. [ Links ]
Kennedy, D.J. 1980. A restudy of conodonts described by Branson and Mehl, 1933, from the Jefferson City Formation, Lower Ordovician, Missouri. Geologica et Palaeontologica 14: 47-76. [ Links ]
Kidwell, S.M. 1991. The stratigraphy of shell concentration. En: P.A. Allison y D.E.G. Briggs (eds.), Taphonomy: releasing the data locked in the fossil record , Plenum Press, New York: 221-290. [ Links ]
Kidwell, S.M., Fürsich, F.T. y Aigner, T. 1986. Conceptual framework for the analysis and classification of fossil concentrations. Palaios 1: 228-238. [ Links ]
Kobayashi, T. 1937. The Cambrian-Ordovician shelly faunas of South America. Journal of the Faculty of Sciences, Tokyo University (Geology) 4: 369-522. [ Links ]
Landing, E. y Ludvigsen, R. 1984. Classification and conodont-based age of the Ordovician trilobite Ellaspis (Middle Arenigian, Ville Guay, Quebec). Canadian Journal of Earth Sciences 21: 1483-1490. [ Links ]
Landing, E., Barnes, C.R. y Stevens, R.K. 1986. Tempo of earliest Ordovician graptolite faunal succession: conodont-based correlations from the Tremadocian of Quebec. Canadian Journal of Earth Sciences 23: 1928-1949. [ Links ]
Lehnert, O. 1994. A Cordylodus proavus fauna from West-Central Argentina (Los Sombreros Fm., Sierra del Tontal, San Juan Province). Zentralblatt für Geologie und Paläontologie , H. 1/2: 245-261. [ Links ]
Lehnert, O. 1995. Ordovizische Conodonten aus der Präkordillere Westargentiniens: Ihre Bedeutung für Stratigraphie und Paläogeographie. Erlanger Geologische Abhandlungen 125: 1-193. [ Links ]
Lindström, M. 1955. Conodonts from the lower-most Ordovician strata of south-central Sweden. Geologiska i Stockholm, Forhandlingar 76: 517-604. [ Links ]
Lindström, M. 1971. Lower Ordovician conodonts of Europe. En: Sweet, W.C. y Bergström, S.M. (eds.), Symposium on conodont biostratigraphy, Geological Society of America Memoir 127: 21-61. [ Links ]
Löfgren, A. 1985. Early Ordovician conodont biozonation at Finngrundet, south Bothnian Bay, Sweden. Bulletin of the Geological Institute of the University of Uppsala N.S. 10: 115-128. [ Links ]
Löfgren, A. 1997. Conodont faunas from the upper Tremadoc at Brattefors, south-central Sweden, and reconstruction of the Paltodus apparatus. GFF, 119: 257-266. [ Links ]
Löfgren, A. y Tolmacheva, T.J. 2003. Taxonomy and distribution of the Ordovician conodont Drepanodus arcuatus Pander, 1856, and related species. Paläontologische Zeitschrift 77: 203-221. [ Links ]
López, C.R. y Nullo, F.E. 1969. Geología de la margen izquierda de la Quebrada de Humahuaca, de Huacalera a Maimará, Dept. Tilcara, Provincia de Jujuy, República Argentina. Revista de la Asociación Geológica Argentina 24: 173-182. [ Links ]
Manca, N., Heredia, S., Hünicken, M.A. y Rubinstein, C. 1995. Macrofauna, Conodontes y Acritarcos de la Formación Santa Rosita (Tremadociano), Nazareno, Provincia de Salta, Argentina. Boletín Academia Nacional de Ciencias , Córdoba, 60: 267-276. [ Links ]
Mángano, M.G., Buatois, L.A. y Muñiz-Guinea F. 2001. Stop 2B. The Upper Cambrian-Tremadoc Santa Rosita Formation at Angosto de Chucalezna. En: L.A. Buatois y M.G. Mángano (eds.), Field Guide. Fourth Argentine Ichnologic Meeting and Second Ichnologic Meeting of Mercosur 50-52. [ Links ]
Miller, J.F. 1969. Conodont fauna from the Notch Peak Limestone (Cambro-Ordovician), House Range, Utah. Journal of Paleontology 43: 413-439. [ Links ]
Miller, J.F. 1980. Taxonomic revisions of some Upper Cambrian and Lower Ordovician conodonts with comments on their evolution. University of Kansas Paleontological Contributions, Paper 99: 1-39. [ Links ]
Miller, J.F. 1984. Cambrian and earliest Ordovician conodont evolution, biofacies, and provincialism. En: D.L. Clark (ed.), Conodont Biofacies and Provincialism, Geological Society of America Special Paper 196: 43-68. [ Links ]
Moya, M.C. 1988. Lower Ordovician in the southern part of the Argentine Eastern Cordillera. En: H. Bahlburg, Ch. Breitkreuz y P. Giese (eds.), The Southern Central Andes, Lecture Notes in Earth Sciences 17: 55-69. [ Links ]
Moya, M.C. 1999. El Ordovícico de los Andes del norte argentino. En: G. González Bonorino, R. Omarini y J. Viramonte (eds.), Geología del Noroeste Argentino, Relatorio del 14º Congreso Geológico Argentino 1: 134-152. [ Links ]
Moya, M.C. 2002. The Ordovician basin of northern Argentina. En: F.G. Aceñolaza (ed.), Aspects of the Ordovician System in Argentina, INSUGEO Tucumán, Serie Correlación Geológica 16: 281-294. [ Links ]
Nicoll, R.S. 1994. Seximembrate apparatus structure of the Late Cambrian coniform conodont Teridontus nakamurai from the Chatsworth Limestone, Georgina Basin, Queensland. AGSO Journal of Australian Geology and Geophysics 15: 367-379. [ Links ]
Nogami, Y. 1967. Kambrische Conodonten von China, Teil 2. Conodonten aus den hoch oberkambrischen Yencho-Schichten. Memoirs of the College of Science, University of Kyoto, Mineralogy and Geology, Series B 33: 211-218. [ Links ]
Nowlan, G.S. 1985. Late Cambrian and Early Ordovician conodonts from the Franklinian Miogeosyncline, Canadian Arctic Islands. Journal of Paleontology 59: 96-122. [ Links ]
Orndorff, R.C. 1988. Latest Cambrian and Earliest Ordovician conodonts from the Conococheague and Stonehenge limestones of northwestern Virginia. Bulletin of the United States Geological Survey 1837-A: 1-18. [ Links ]
Pander, C.H. 1856. Monographie der fossilen Fische des silurischen Systems der russischbaltischen Gouvernements. Akademie der Wissenschaften St. Petersburg, 91 pp. [ Links ]
Pemberton, S.G., Mac Eachern, J.A. y Frey, R., 1992a. Trace fossil facies models: environmental and allostratigraphic significance. En: R.G. Walker y N.P. James (eds.), Facies models, response to sea level change, Geological Association of Canada, cap. 4: 47-72. [ Links ]
Pemberton, S.G., Frey, R.W., Ranger, M.J. y Mac Eachern, J., 1992b. The conceptual framework of ichnology. En: S.G. Pemberton (ed.), Applications of ichnology to petroleum exploration-a core workshop. Society of Economic Paleontologists and Mineralogists, Core Workshop 17: 1-32. [ Links ]
Purnell, M.A., Donoghue, P.C.J. y Aldridge, R.J. 2000. Orientation and anatomical notation in conodonts. Journal of Paleontology 68: 599-616. [ Links ]
Pyle, L.J. y Barnes, C.R. 2002. Taxonomy, Evolution, and Biostratigraphy from the Kechika Formation, Skoki Formation, and Road River Group (Upper Cambrian to Lower Silurian), Northeastern British Columbia. National Research Council, Research Press , Ottawa: 1-227. [ Links ]
Rao, R.I. 1999. Los conodontes Cambro-Ordovícicos de la Sierra de Cajas y del Espinazo del Diablo, Cordillera Oriental, República Argentina. Revista Española de Micropaleontología 31: 23-51. [ Links ]
Rao, R.I. y Flores, F.J. 1998. Conodontes ordovícicos (Tremadoc superior) de la sierra de Aguilar, provincia de Jujuy, República Argentina. Bioestratigrafía y tafonomía. Revista Española de Micropaleontología 30: 5-20. [ Links ]
Rao, R.I. y Tortello, M.F. 1998. Tremadoc conodonts and trilobites from the Cardonal Formation, Incamayo Creek, Salta Province, northwestern Argentina. Palaentolologia Polonica 58: 31-45. [ Links ]
Repetski, J.E. y Ethington R.L. 1983. Rossodus manitouensis (Conodonta), a new Early Ordovician index fossil. Journal of Paleontology 57: 289-301. [ Links ]
Ross, R.J. Jr., Hintze, L.F., Ethington, R.L., Miller, J.F., Taylor, M.E. y Repetski, J.E. 1997. The Ibexian, Lower most Series in the North American Ordovician. U.S. Geological Survey Professional Paper 1579: 1- 50. [ Links ]
Seilacher, A. 1964. Sedimentological classification and nomenclature of trace fossils. Sedimentology 3: 253-256. [ Links ]
Seilacher, A. 1967. Bathymetry of trace fossils. Marine Geology 5: 413-428. [ Links ]
Seo, K.-S., Lee, H.-Y. y Ethington, R.L. 1994. Early Ordovician conodonts from the Dumugol Formation in the Baegunsan Syncline, Eastern Yeongweol and Samcheog areas, Kangweon-Do, Korea. Journal of Paleontology 68: 599-616. [ Links ]
Stone, J. 1987. Review of investigative techniques used in the study of conodonts. En: R.L. Austin (ed.), Conodonts: Investigative Techniques and Applications, Ellis Horwood Limited, Chichester: 17-34. [ Links ]
Stouge, S. 1984. Conodonts of the Middle Ordovician Table Head Formation, western Newfoundland. Fossils and Strata 16: 1-145. [ Links ]
Stouge, S. y Bagnoli, G. 1988. Early Ordovician conodonts from Cow Head Peninsula, western Newfoundland. Palaeontographia Italica 75: 89-179. [ Links ]
Sweet, W.C. 1981. Morphology and composition of elements. Macromorphology of elements and apparatuses. En: R.A. Robison (ed.), Treatise on Invertebrate Paleontology, Part W, Miscellanea, Supplement 2, Conodonta, The Geological Society of America and the University of Kansas, Boulder and Lawrence: W5-W20. [ Links ]
Sweet, W.C. 1988. The Conodonta: Morphology, taxonomy, paleoecology, and evolutionary history of a long-extinct animal phylum. Oxford monographs on geology and geophysics, Oxford University Press, New York, 10: 1-212. [ Links ]
Sweet, W.C. y Donoghue, P.C.J. 2001. Conodonts: Past, Present, Future. Journal of Paleontology 75: 1174-1184. [ Links ]
Szaniawski, H. 1980. Conodonts from the Tremadocian chalcedony beds, Holy Cross Mountains (Poland). Acta Palaeontologica Polonica 25: 101-121. [ Links ]
Tolmacheva, T.J. 2001. Conodont biostratigraphy and diversity in the Lower-Middle Ordovician of Eastern Baltoscandia (St. Petersburgh region, Russia) and Kazakhstan. Comprehensive summary of doctoral dissertation, Department of Earth Sciences, Uppsala University, 40 pp. [ Links ]
Tortello, M.F. y Rao, R.I. 2000. Trilobites y conodontes del Ordovícico temprano del Angosto de Lampazar (Provincia de Salta, Argentina). Boletín Geológico y Minero 111: 61-84. [ Links ]
Viira, V. 1970. Konodonty Varanguskoj pachki (Verchnij Tremadok Estonii). Eesti NSV Teaduste Academia, Keemia-Geoloogia 19: 224-233. [ Links ]
Wamel, W.A. van. 1974. Conodont biostratigraphy of the Upper Cambrian and Lower Ordovician of north-western Öland, south-eastern Sweden. Utrecht Micropalaeontological Bulletin 10: 1-125. [ Links ]
Zeballo, F.J., Albanesi, G.L., Ortega, G. y Tortello, M.F. 2003. Biostratigraphy of Ordovician sequences from Alfarcito Area, Tilcara, Eastern Cordillera of Jujuy, Argentina. En: G.L. Albanesi, M.S. Beresi y S.H. Peralta (eds.), Ordovician from the Andes, INSUGEO Tucumán, Serie Correlación Geológica 17: 167-171. [ Links ]
Zeballo, F., Albanesi, G.L. y Ortega, G. 2005a. Conodontes y graptolitos de las formaciones Alfarcito y Rupasca (Tremadociano) en el área de Alfarcito, Tilcara, Cordillera Oriental de Jujuy, Argentina. Parte 1: Bioestratigrafía. Ameghiniana 42: 39-46. [ Links ]
Zeballo, F., Albanesi, G.L. y Ortega, G. 2005b. Conodontes y graptolitos de las formaciones Alfarcito y Rupasca (Tremadociano) en el área de Alfarcito, Tilcara, Cordillera Oriental de Jujuy, Argentina. Parte 2: Paleontología sistemática. Ameghiniana 42: 47-66. [ Links ]
Recibido: 7 de mayo de 2004.
Aceptado: 29 de julio de 2004.

