










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.2 Buenos Aires mar./jun. 2005
Diconodinium lurense sp. nov., a late Maastrichtian to Danian dinoflagellate cyst from southwest Atlantic basins
G. Raquel Guerstein 1,2 , Gabriela L. Junciel 1 , M.Verónica Guler 1,2 and Gloria Daners 3
1Departamento de Geología, Universidad Nacional del Sur. San Juan 670, B8000ICN Bahía Blanca, Argentina. gmguerst@criba.edu.ar
2 Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina.
3 Departamento de Paleontología, Facultad de Ciencias, Iguá 4225, 11400 Montevideo, Uruguay.
Abstract. Upper Cretaceous and Cenozoic deposits from the Colorado and Austral basins, Argentina and the Punta del Este Basin, Uruguay contain diverse organic-walled dinoflagellate cysts, including Diconodinium lurense sp. nov. which occurs in the late Maastrichtian and Danian throughout the basins. The new species is characterized by an autophragm that is densely covered by short spines and by relatively long apical and antapical horns, and by a variable Ia to IPa archeopyle. On our evidence, the stratigraphical range of Diconodinium lurense sp. nov. is latest Maastrichtian to Danian, and is thus a useful biostratigraphical marker for the Cretaceous-Palaeogene transition in the southwest Atlantic basins.
Resumen. Diconodinium lurense , una nueva especie de quiste de dinoflagelado del M aastrichtiano T ardío- D aniano del A tlántico sudoccidental . Los depósitos del Cretácico Superior y Cenozoico de las cuencas del Colorado y Austral, Argentina y Cuenca Punta del Este, Uruguay son portadores de asociaciones de quistes de dinoflagelados diversas. Diconodinium lurense sp. nov. es un elemento común en las asociaciones provenientes del Maastrichtiano tardío-Daniano en las distintas cuencas. Esta nueva especie se caracteriza por presentar un autofragma densamente cubierto por espinas cortas, cuernos apical y antapical relativamente largos y por un arqueopilo variable Ia-IPa. Nuestras evidencias indican que el rango estratigráfico de Diconodinium lurense sp. nov. se extiende desde al Maastrichtiano tardío hasta el Daniano, constituyendo un importante marcador bioestratigráfico para la transición Cretácico-Paleógeno en las cuencas del Atlántico Sudoccidental.
Key words. Biostratigraphy; Dinoflagellate cysts; Late Maastrichtian-Danian; Southwest Atlantic.
Palabras clave. Bioestratigrafía; Quistes de dinoflagelados; Maastrichtiano tardío-Daniano; Atlántico Sudoccidental.
Introduction
The dinoflagellate cyst (dinocyst) Diconodinium lurense sp. nov., was recovered from wells, boreholes and outcrops from several Southwestern Atlantic basins (figure 1). It is present in the Maastrichtian-Danian Pedro Luro Formation (Kaasschieter, 1963), Colorado Basin, in an onshore borehole and from offshore wells. The species was also recorded as Diconodinium sp. in coeval deposits of the Maastrichtian-Mid Eocene Gaviotín Formation in an offshore well in the Punta del Este Basin, Uruguay (Daners and Guerstein, 2004). In both basins, the distribution of Diconodinium lurense seems to be consistent and characterizes the Cretaceous-Palaeogene (K/P) boundary. And, well preserved specimens of this species from surface section of the Calafate Formation in the Austral Basin, allow determination of its first occurrence datum (FOD) in the latest Maastrichtian (Marenssi et al ., 2004). The aim of the present contribution is to formally describe the new species and to emphasize its regional biostratigraphical potential in the Southwest Atlantic basins.

Figure 1. Map of the southern part of South America that shows localities, wells and outcrops as well as basins mentioned in the text / Mapa de la parte sur de Sudamérica mostrando las localidades, pozos, afloramientos y cuencas mencionadas en el texto.
Geological background
The development of the Colorado Basin, like that of other Southwest Atlantic basins, is related to the Late Jurassic-Early Cretaceous rifting and subsequent break up of Gondwana (Juan et al ., 1996). All these Southwest Atlantic basins were large areas of deposition during the Late Cretaceous and Cenozoic. Fryklund et al . (1996, fig. 5, p. 143) identified tectono-stratigraphic sequences characteristic of a passive margin setting in the Colorado Basin, including rift, sag and drift deposits. The Pedro Luro Formation constitutes the base of the drift sequence and was deposited during a widespread marine transgression of Maastrichtian-Danian age. The formation consists of marine siltstones and claystones with subordinate, sometimes glauconitic sandstones.
In the Punta del Este Basin, the Gaviotín Formation represents partially the passive margin deposits (Ucha et al ., 2003). Gaviotín Formation was defined by Ucha et al . (2003), which consists of a marine succession of dark-greenish to gray claystones, siltstones, and sandstones with occasional intervals of glauconitic beds. The dinocyst assemblages from the lower part of the formation suggest a Maastrichtian to Danian age (Daners and Guerstein, 2004).
In the Austral Basin, the age of the Calafate Formation has been widely discussed. Nullo et al . (1981) and Macellari et al . (1989) suggested a Maastrichtian-Paleocene age for this unit, whereas Malumián and Caramés (1997) indicate that in the area of Lago Argentino the Paleocene deposits were either missing or poorly developed. According to Marenssi et al . (2004), the age of the Calafate Formation south of Lago Argentino is Maastrichtian to late Maastrichtian. The studied section at this area is characterized by a predominance of marine sandstones with minor conglomerate beds and a few mudstone beds.
Materials and methods
Almost all the studied samples proceeding from the Maastrichtian-Danian interval of the Colorado and Punta del Este basins are cuttings, since there are no outcrops of Mesozoic or Palaeogene strata. In the Colorado Basin, we analyzed nine cutting samples from the offshore Cx-1 Well, between 1300 and 1600 metres depth (md) and one core and two cutting samples from the onshore Pedro Luro Borehole, between 1487 and 1550 md (figure 1). Eight cutting samples are derived from the Gaviotín Well, drilled offshore Punta del Este Basin, between 1658 and 1850 md.
The palynological material from the Austral Basin is derived from a surface section at the estancia 25 de Mayo, Southwestern Santa Cruz Province (figure 1). Eleven of the twenty samples processed for palynomorphs contain dinocysts.
The palynological treatment included hydrofluoric and hydrochloric acids, mild oxidation (10% nitric acid for one minute) and a 10% ammonium hydroxide wash for one minute. These procedures were followed by heavy liquid separation of the organic component, differential centrifugation to remove fine particles and screening to concentrate the 10 - 25 and 25-180 µm fractions. The residues from the Cx-1 well were refined, stained with Bismarck C and mounted in Elvacite and Cellosize substitute at the Geological Survey of Canada (Atlantic). The residues from the other sites were mounted in glycerine jelly.
Morphologic studies were made using a Nikon Eclipse 600 microscope at the Universidad Nacional del Sur and specimens were photographed with a Nikon Coolpix 950 digital camera. Coordinates quoted from the Vernier scale of Nikon microscope serial nº 772751 follow the sample and slide number for each specimen illustrated. England Finder (EF) references are provided in the corresponding figure explanations. The types and figured specimens are kept in the Palynological Collection of the Departamento de Geología de la Universidad Nacional del Sur, Bahía Blanca, Argentina. The dinoflagellate cyst nomenclature follows Williams et al . (1998a) and the systematic section uses the classification of Fensome et al . (1993). The geological timescales are from Berggren et al . (1995) and Gradstein et al . (1995).
Palynostratigraphy
The samples from the Colorado, Punta del Este and Austral basins are generally rich in dinocysts, but contain variable amounts of spores, pollen, acritarchs and foraminiferal test linings. Age control is based on the correlation with the dinoflagellate range data of Williams et al . (1998b, 2004) and Brinkhuis et al . (2003). Dinocyst taxa cited in the text are listed in the Appendix 1 and are fully referenced in Williams et al . (1998a).
Colorado Basin
Cx-1 Well. Dinocysts dominate the palynological assemblages throughout the Pedro Luro Formation of the Cx-1 Well. Since all the samples are cuttings, age assignments are based on the highest occurrences of dinoflagellate species.
Danian strata extend from 1340 to 1365 m (figure 2).Ten taxa have their highest ccurrences in this interval, including Areoligera medusettiformis O. Wetzel ex Lejeune-Carpentier, Disphaerogena carposphaeropsis O. Wetzel emend . Sarjeant, Palaeoperidinium pyrophorum (Ehrenberg ex O. Wetzel) Sarjeant, and Senoniasphaera inornata (Drugg) Stover and Evitt. Based on Williams et al. (1998b), Areoligera medusettiformis O. Wetzel ex Lejeune-Carpentier, which appeared during the Late Cretaceous persisted into the Palaeogene (Lutetian). According to Williams et al . (2004) the Last Occurrence Datum (LOD) of Palaeoperidinium pyrophorum (Ehrenberg ex O. Wetzel) Sarjeant, is at about 58 Ma at mid latitudes in the Northern Hemisphere and at high latitudes in the Southern Hemisphere. Moreover, Brinkhuis et al . (2003) found the last abundant occurrence of this taxon in ODP site 1172 East Tasman Plateau (44ºS), at approximately 63 Ma (Danian). Williams et al. (2004) gave a range for Senoniasphaera inornata (Drugg) Stover and Evitt of 64.95 to 62.6 Ma within the Danian at mid latitudes. This species is thus the best stratigraphic marker, suggesting an age no younger than late Danian for this interval.
Maastrichtian or older strata occur below 1380 m, as indicated by the presence of Dinogymnium undulosum Cookson and Eisenack. Williams et al . (2004) placed the LO of the genus Dinogymnium at the Cretaceous/ Palaeogene boundary. Diconodinium lurense sp. nov. has its highest stratigraphical occurrence at 1380 m and is quite common in underlying deposits. Pedro Luro -1 Borehole. Kaasschieter (1963) defined the type section of the Pedro Luro Formation between 1360 and 1508 md (1339 to 1487 m below mean sea level) in the Pedro Luro -1 (figure 2). In our study, the highest productive palynology sample bearing dinocysts proceed from 1487- 1488 m sidewall core sample. The dinocyst assemblages from 1487-1488 m and 1492 m cutting sample are not as diverse as those from the Cx-1 Well but contain taxa such as Alisocysta circumtabulata (Drugg) Stover and Evitt, Diconodinium lurense sp. nov. and Trithyrodinium evittii Drugg. Following Williams et al . (2004), the LODs of Alisocysta circumtabulata (Drugg) Stover and Evitt is recorded in mid latitudes Northern Hemisphere at 58.5 Ma whereas the last common occurrence is placed at 57 Ma in mid-high latitudes Southern Hemisphere; Brinkhuis et al . (2003) found the LOD of Trithyrodinium evittii Drugg in the Tasman Plateau at 57.3 Ma. In this way, the age of the interval 1487-1492 m is believed to be Selandian or older. It is impossible to determine whether Diconodinium lurense sp. nov. extends above 1487 m, because of poor sample control.
Underlying sediments appear to be Campanian or early Maastrichtian in age or older. This determination is based on the presence of Odontochitina sp. At 1505 m. Helby et al . (1987, fig. 40, p. 65) showed the LOD of Odontochitina spp. in their Australasian Isabelidinium korojonense Range Zone, which is mid Campanian to early Maastrichtian. Taking into account the information published by Wilson (1984), Wood and Askin (1992) considered the ranges of Odontochitina spp. in both Australia and New Zealand to be equivalent, and placed its LOD at the end of the Campanian. Williams et al. (2004) considered that the range of Odontochitina operculata (O.Wetzel) Deflandre and Cookson extends into the early Maastrichtian (68.5 Ma) at high latitudes Southern Hemisphere and Odontochitina costata Alberti and Odontochitina operculata (O. Wetzel) Deflandre and Cookson has its LOD at 70 Ma at mid latitudes Northern Hemisphere Thus, the age of the bottom of the Pedro Luro Formation in this borehole is taken to be possibly as young as early Maastrichtian.
Other boreholes. Diconodinium lurense sp. nov. Was previously recorded (as Lejeunia sp.) by Gamerro and Archangelsky (1981) in the two offshore boreholes Ranquel x-1 and Puelche x-1. In the Ranquel x- 1 Borehole, the Pedro Luro Formation extends from 2050 to 2300 m. The top of this unit is marked by the highest occurrences of several species, such us Diconodinium lurense sp. nov. as well as Manumiella ? cretacea (Cookson) Bujak and Davies (as Isabelidinium cretaceum ). The top of Pedro Luro Formation in Puelche x-1 Borehole, at 1600 m corresponds to the highest occurrences of Trithyrodinium evittii Drugg (as T. fragile ), Palaeocystodinium australinum (Cookson) Lentin and Williams and Cerodinium dartmoorium (Cookson and Eisenack) Lentin and Williams (as Deflandrea dartmooria ). Although Diconodinium lurense sp. nov. ranges into the overlying units (dated as Eocene to Oligocene), this species seems to be consistently present only below the top of Pedro Luro Formation. We therefore consider the presence of Diconodinium lurense in the Eocene to Oligocene indicates reworking.
Williams et al . (1998b) placed the LOD of Manumiella ? cretacea (Cookson) Bujak and Davies close to the end of the Danian (at about 61 Ma). A reevaluation of the stratigraphical significance of LODs of selected dinocyst species suggests that the age of the Pedro Luro Formation, and therefore the youngest deposits containing Diconodinim lurense sp. nov. are not younger than Danian. Moreover, Archangelsky et al . (1997) studied the Pejerrey x-1 Well, drilled offshore Colorado Basin, where they recorded our new species in strata assigned to Maastrichtian-Danian age.
Punta del Este Basin
According to Daners and Guerstein (2004), based on dinoflagellate cysts, the transition between Cretaceous and Palaeogene strata in the Gaviotín borehole is placed between 1717 and 1747 md (figure 2). The presence of Damassadinium californicum (Drugg) Fensome et al. between 1582 and 1658 md. suggests a Danian or earliest Selandian age since Williams et al . (2004) recorded the LOD of this species for mid latitudes in the Northern Hemisphere at 60.3 Ma. At 1717 and 1814 md the highest occurrences of Tanyosphaeridium variecalamum Davey and Williams and Alisogymnium euclaense (Cookson and Eisenack) Lentin and Vozzhennikova, confirm an age no younger than Maastrichtian for the lower part of the selected interval (Williams et al ., 1998b). Thus, Diconodinium lurense sp. nov. (as Diconodinium sp.) was recorded from Maastrichtian to lowermost Danian sediments.

Figure 2. Stratigraphical occurrence of Diconodinium lurense sp. nov. in Colorado, Punta del Este and Austral basins, c: cutting sample / Distribución estratigráfica de Diconodinium lurense sp. nov. en las cuencas de Colorado, Punta del Este y Austral, c: muestra de cutting.Austral Basin
The dinocyst assemblages studied in the Austral Basin are recovered from the Calafate Formation, which outcrops south of Lago Argentino, southwestern Santa Cruz Province. Diconodinium lurense sp. nov. is recorded (as Diconodinium sp.) only in the uppermost part of the section (figure 2). The presence of Manumiella druggii (Stover) Bujak and Davies and Alisocysta circumtabulata (Drugg) Stover and Evitt indicate a maximum late Maastrichtian age for the middle part of the section and upwards. Despite the ranges of the recorded dinocyst species extending into the Early Palaeogene, the absence of species first appearing in the Danian is noticeable. Sedimentological studies suggest that the Cretaceous-Palaeogene boundary deposits have been eroded in this area. Marenssi et al . (2002) recognized that southern Lago Argentino, the Eocene Man Aike Formation unconformably overlies the Late Cretaeous Calafate Formation. This is supported by the persistent presence of the megaspore Grapnelispora loncochensis Papú 1997 in the uppermost part of this section. This species, which has hitherto only been recovered from Maastrichtian deposits of Patagonia and would reinforce an age no younger than late Maastrichtian (Marenssi et al ., 2004). This data provide the evidence for considering the FO of Diconodinium lurense to be late Maastrichtian.
Summary
The distribution of Diconodinium lurense sp. nov. in the three basins confirms it as a consistent component of K/P boundary dinocyst assemblages. The new species probably appeared during the late Maastrichtian and disappeared around about the Danian/Selandian boundary. Thus, Diconodinium lurense sp. nov. seems to be a good biostratigraphical marker for deposits related to the Maastrichtian-Danian transition in the Southwest Atlantic Basins.
Systematic palynology
Division DINOFLAGELLATA (Bütschli 1885) Fensome et al . 1993
Subdivision DINOKARYOTA Fensome et al . 1993
Class DINOPHYCEAE Pascher 1914
Subclass PERIDINIPHYCIDAE Fensome et al . 1993
Order PERIDINIALES Haeckel 1894
Suborder PERIDINIINEAE Autonym
Family PERIDINIACEAE Ehrenberg 1831
Genus Diconodinium Eisenack and Cookson 1960 emend . Morgan 1977
Type species. Diconodinium multispinum Deflandre and Cookson, 1955, pl.1, fig.5, as Palaeohystrichophora multispina .
Diconodinium lurense sp. nov.
(Figures 3.A-C, Figure 4.A-I, Figure 5.A-I)
1981 Lejeunia sp. Gamerro and Archangelsky: page 123, pl II, fig.6.

Figure 3. Diconodinium lurense sp. nov., line drawings of external views. For those illustrations indicated as being reversed views, the drawings are a mirror image of the appearance of the specimen in the microscope / Diconodinium lurense sp. nov., dibujos esquemáticos de vistas externas. Para todas las ilustraciones indicadas como vistas reversas, los dibujos son una imagen especular de la observada en el microscopio. A, PL -1 sample 631(2): 31/94.8, D 45/4. Holotype, dorsal surface ( reversed view) / holotipo, superficie dorsal (imagen invertida). B, Cx-1 sample P35295(1): 48/103, N 28/1, dorsal surface ( reversed view) / superficie dorsal (imagen invertida) . C, Cx-1 sample P35295(1): 29/92, A 47/4, dorsal surface (reversed view) / superficie dorsal (imagen invertida).
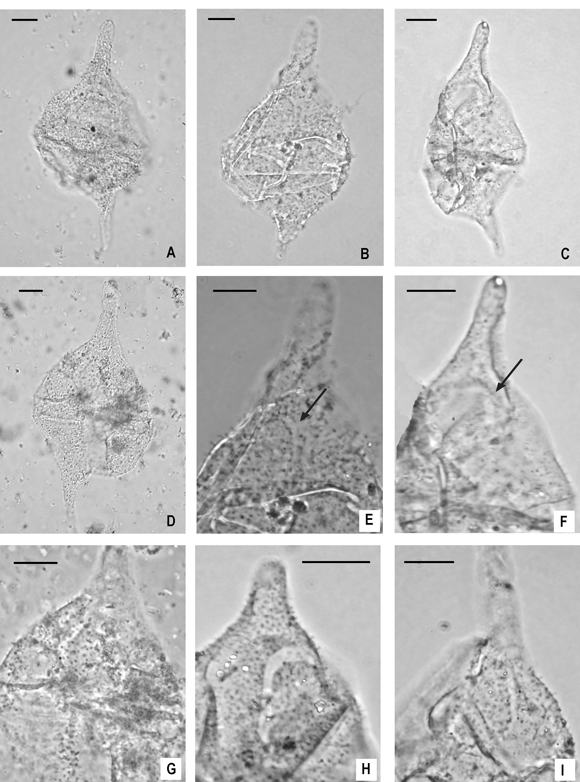
Figure 4. A-I. Diconodinium lurense sp. nov. from the Pedro Luro Formation, Colorado Basin. Scale bar represents 10 µm for all specimens. The digital images were taken using phase contrast, unless otherwise stated / Diconodinium lurense sp. nov. proveniente de la Formación Pedro Luro, cuenca del Colorado. La escala gráfica representa 10 µm para todos los especímenes. Las imágenes digitales fueron tomadas con contraste de fase, excepto cuando es indicado. A, PL -1 sample 631(2): 31/94.8, D 45/4. Holotype, ventral view, dorsal surface, interference contrast / holotipo, vista ventral, superficie dorsal, contraste de interferencia. B, E, Cx-1 sample P35298(1): 48/109, T 28/1: B, ventral view, optical section / vista ventral, sección óptica ; E, ventral view, dorsal surface, archeopyle arrowed / v ista ventral, superficie dorsal, arqueopilo señalado . C, F, Cx-1 sample P35295(1): 48/103, N 28/1. C, ventral view, dorsal surface / vista ventral, superficie dorsal ; F, detail of the archeopyle, arrowed / detalle del arqueopilo, señalado. D,G, PL-1 sample 632(1): 41/109, T 35/0. D, dorsal view, dorsal surface, interference contrast / vista dorsal, superficie dorsal, contraste de interferencia ; G, detail showing archeopyle and cingulum / detalle mostrando el arqueopilo y el cíngulo . H, Cx-1 sample P35295(1): 29/92, A 44/4. detail, ventral view, dorsal surface showing the archeopyle / d etalle, vista ventral, superficie dorsal mostrando el arqueopilo . I, Cx-1 sample P35290(1): 42/104.5, O 34/3. detail, dorsal view, dorsal surface showing the archeopyle / detalle, vista dorsal, superficie dorsal mostrando el arqueopilo.
Holotype. Slide PL-1: 631(2), coordinates: 31/94.8 (England Finder reference: D 45/4) Figure 3.A, Figure 4.A.
Paratype. Slide PL-1: 632(1), coordinates: 41/109 (England Finder reference: T 35/0) Figures 4.D, G.
Repository. Palynological Collection, Departamento de Geología de la Universidad Nacional del Sur, Bahía Blanca, Argentina.
Type locality. Pedro Luro-1 Borehole, Buenos Aires Province, Argentina.
Type stratum. 1487-1488 m depth (core sample).
Derivation of name. After the Pedro Luro Formation, from which this species was first recorded.
Diagnosis. A species of Diconodinium characterized by relatively long apical and antapical horns and with an autophragm densely covered by short nontabular spines.
Description. Cyst fusiform in shape, consisting of a subtriangular epitract slightly larger than the rounded hypotract. The epitract extends into a 20 to 30 µm long apical horn with a rounded distal tip. The hypotract bears a single, pointed, 15 to 25 µm long antapical horn clearly offset to the left from the midline of the cyst. The cingulum is about 4 µm wide, and slightly offset at the sulcus. The autophragm is densely covered by thin, solid spines up to 1.5 µm high. The spines are all of the same type and size. They are distributed randomly except along the margin of the cingulum, where they are always aligned in rows (figures 3.A-C). The archeopyle is rarely discernible but, when visible, is intercalary normally type Ia but occasionally IPa. The former type includes the stenodeltaform plate 2a only with clear apical and lateral sutures (figures 3.B, 4.H). When having a type Ipa archeopyle the lateral sutures of plate 4´´ are partly open but never reach the cingulum (figures 3.A, 4.F, G). In both archeopyle types the operculum remains in situ and is always adnate posteriorly (figures 4.H, I, 5.H, I).

Figure 5. A-I, Diconodinium lurense sp. nov. from the Calafate Formation, which outcrops at the Estancia 25 de Mayo, Austral Basin. Scale bar represents 10 µm for all specimens. The digital images were taken using interference contrast / Diconodinium lurense sp. nov. proveniente de la Formación Calafate aflorante en la Estancia 25 de Mayo, cuenca Austral. La escala gráfica representa 10 µm para todos los especímenes. Las imágenes digitales fueron tomadas con contraste de interferencia. A-C, Sample 3431(1): 40/96.5, U40/4; ventral view / vista ventral ; A, B, ventral surface / superficie ventral; C, dorsal surface / superficie dorsal . D, E, sample 3431(1): 22/108, H22/3, dorsal view / vista dorsal ; D, dorsal surface / superficie dorsal ; E, ventral surface / superficie ventral . F-I, sample 3431(2): 45/91.5, Z45/4, ventral view / vista ventral ; F, G, ventral surface / superficie ventral ; H, dorsal surface / superficie dorsal ; I, close-up of posteriorly attached operculum / detalle mostrando el opérculo adherido posteriormente.
Dimensions . Overall length: 55 (80) 110 µm; width: 30 (41) 50 µm (53 specimens measured).
Stratigraphic distribution. Late Maastrichtian to Danian.
Comparison. Diconodinium lurense differs from most other described species of Diconodinium in having a fusiform shape and relatively long apical and antapical horns. Other species also bearing relatively long horns are Diconodinium martianum Srivastava and Diconodinium longicorne Olaru. The former resembles Diconodinium lurense closely in shape and size, but differs in having the antapical horn located close to the midline and in having a smooth autophragm. Diconodinium longicorne Olaru is characterized by having a subspherical to rhomboidal body, an epitract that is bigger than the hypotract, and an apical horn that is longer than the shorter antapical horn. In addition, its surface is finely granulate and shows tabulation on both epitract and hypotract. Diconodinium davidii Morgan is similar in size to Diconodinium lurense sp. nov. and also has a slightly offset antapical horn, but differs in having coarser, aligned spines that may reveal traces of tabulation. Diconodinium pusillum Singh also has an asymmetrically aligned antapical horn and an epitract that is longer than the hypotract. It differs from the new species by having a shorter, bifid apical horn and finer spines 0.5 µm long, and in being smaller. Diconodinium lurense also resembles Diconodinium multispinum (Deflandre and Cookson) Eisenack and Cookson emend . Morgan, however, the latter shows signs of paratabulation, especially on the epitract. Furthermore, Diconodinium multispinum has ornamentation which is slightly larger and adjacent ornamentation elements may be fused to form discontinuous parasutural ridges. Diconodinium vitricornu Roncaglia et al . differs from Diconodinium lurense in having a smooth autophragm, mostly thin but thickened at the apex to form a solid tip of the horn. Alterbidinium acutulum (Wilson) Lentin and Williams possesses a similar outline and comparable horns to Diconodinium lurense , and also has an epitract larger than the hypotract. However, Alterbidinium acutulum (Wilson) Lentin and Williams is characteristically circumcavate and both endophragm and periphragm are smooth.
Remarks. A few specimens of Diconodinium lurense appear to show cavation, especially below the apical horn (figures 4.F, H). A detailed examination of our specimens revealed that this is an optical effect, produced by folding of the thin autophragm at the anterior margin of the archeopyle, and does not indicate a second wall layer. According to Fensome et al. (1993, p. 131), Diconodinium is morphologically intermediate between the two subfamilies Palaeoperidinioideae and Deflandroideae, since the type species can have either a single plate intercalary archeopyle type 1Ia or a single plate intercalary plus single plate precingular archeopyle type 1I1Pa. Although the archeopyle sutures are not always clearly discernible, most of our specimens have an archeopyle type Ia and just a few of them seem to present an Ipa type. This variation reinforces the comments of Fensome et al . (1993).
Acknowledgments
The authors thank Rob Fensome (Geological Survey of Canada), Henrik Nøhr Hansen (Geological Survey of Denmark and Greenland) and Henk Brinkhuis (Utrecht University, The Netherlands) for critical comments which improved the previous version. To the journal referees Graham Williams (Geological Survey of Canada) and Poul Schiøler (Geological Survey of Denmark and Greenland) for the constructive criticism of the final version of the manuscript. Technical laboratory support by Bernie Crilley (Geological Survey of Canada) is gratefully thanked. Dann Saveanou (Universidad Nacional del Sur) provided helpful assistance regarding the translation from papers published in Romanian language. This contribution has been funded by Fondo para la Investigación Científica y Tecnológica (PICT Nº 07-09659/01), Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 02040) and Secretaría General de Ciencia yTecnología de la Universidad Nacional del Sur.
References
Archangelsky, S., Guerstein, G.R. and Quattrocchio, M.E. 1997. [ Pejerrey x-1 offshore well, Paynological Report. Shell Capsa, Buenos Aires, 19 pp. Unpublished report]. [ Links ]
Berggren, W.A., Kent, D.V., Swisher III, C.C. and Aubry, M.P. 1995. A revised Cenozoic geochronology and chronostratigraphy. In: W.A. Berggren, D.V. Kent, M.P, Aubry and J. Hardenbol (eds.), Geochronology, Timescales and Global Stratigraphic Correlation. Tulsa, SEPM Special Publication 54: 129-212. [ Links ]
Brinkhuis, H., Sengers, S. Sluijs, A., Warnaar, J. and Williams, G.L. 2003. Latest Cretaceous-earliest Oligocene, and Quaternary dinoflagellate cysts, ODP Site 1172, East Tasman Plateau. In: N.F. Exon, J.P. Kennett and M.J. Malone (eds.), Proccedings of the Ocean Drilling Program, Scientific Results Volume 189, 48 pp. [ Links ]
Bütschli, O. 1885. Erster Band. Protozoa. In: Dr. H.G. Bronn's Klassen und Ordnungen des Thier-Reichs. W issenschaftlich dargestellt in Wort und Bild , pp. 865-1088; C.F. Winter'sche Verlagsbuchhandlung, Leipzig and Heidelberg, Germany. [ Links ]
Daners, G. and Guerstein G.R. 2004. Dinoflagelados del Maastrichtienese-Paleogeno en la Formación Gaviotín, cuenca Punta del Este. In: G. Veroslavsky, M. Ubilla and S. Martínez (eds.), Cuencas sedimentarias de Uruguay; Geología, Paleontología y Recursos Naturales, Cenozoico , Edición: División Relaciones y Actividades Culturales de Facultad de Ciencias, Montevideo. 37-62. [ Links ]
Deflandre, G. and Cookson, I.C. 1955. Fossil microplankton from Australian Late Mesozoic and Tertiary sediments. Australian Journal of Marine and Freshwater Research 6: 242- 313. [ Links ]
Ehrenberg, C.G. 1831. Animalia evertebrata. In: P.C. Hamprich and C.G. Ehrenberg, Symbolae physicae....Pars zoological, 10 pl (Plates issued 1828, unpaginated text issued 1831, fide Silva, 1980b, p. 119. Farr et al. cited the authorship of Cryptomonas as Ehrenberg, Symb Phys Anim. Evertebr. I:...1831; Abhandlungen Akademie der Wissenschaften Zu Berlin 1831:....1832, thus implying an 1832 publication date assuming that they were referring to the same publication as that cited herein as Ehrenberg, 1831). [ Links ]
Eisenack, A. and Cookson, I.C. 1960. Microplankton from Australian Lower Cretaceous sediments. Proceedings of the Royal Society of Victoria 72: 1-11. [ Links ]
Fensome, R.A., Taylor, F.J. R., Norris, G., Sarjeant, W.A.S., Wharton, D.I. and Williams, G.L. 1993. A classification of fossil and living dinoflagellates. Micropaleontology Press, Special Publication 7, Pennsylvania, 1-351. [ Links ]
Fryklund, R., Marshall, A. and Steves, J. 1996. La Cuenca del Colorado. In: V.A. Ramos and M.A. Turic (eds.), Geología y Recursos Naturales de la Plataforma Continental Argentina. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos, (Buenos Aires). Relatorio 8: 135-158. [ Links ]
Gamerro, J.C. and Archangelsky, S. 1981. Palinozonas Neocretácicas y Terciarias de la plataforma continental Argentina en la Cuenca del Colorado. Revista Española de Micropaleontología 13: 119-140, pl.1-4. [ Links ]
Gradstein, F.M., Agterberg, F.P., Ogg, J.G., Hardenbol, J., Van Veen, P., Thierry, J. and Huang, Z. 1995. A Triassic, Jurassic, and Cretaceous time scale. In: W.A. Berggren, D.V. Kent, M.P. Aubrey and J. Hardenbol (eds.), Geochronology, Time Scales and Global Stratigraphic Correlation : Society of Economic Paleontologists and Mineralogist (Society for Sedimentary Geology) Special Publication 54, 95-126. [ Links ]
Haeckel, E. 1894. Systematische Phylogenie. Entwurf eines natürlichen Systems der Organismen auf Grund ihrer Stammegeschichte, I. Systematische Phylogenie der Protisten und Pflanzen. XV+400 p.; Reimer, Berlin, Germany. [ Links ]
Helby, R., Morgan, R., and Partridge, A.D. 1987. A palynological zonation of the Australian Mesozoic. In: P.A. Jell (ed.), Studies in Australian Mesozoic Palynology. Memoir of the Association of Australasian Palaeontologists 4: 1-94. [ Links ]
Juan, R., de Jager, J., Russell, J. and Gehard, I. 1996. Flanco norte de la Cuenca del Colorado. In: V.A. Ramos and M.A. Turic (eds.). Geología y Recursos Naturales de la Plataforma Continental Argentina. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos, (Buenos Aires). Relatorio 7: 117-133. [ Links ]
Kaasschieter, J.P.H. 1963. Geology of the Colorado basin. Tulsa Geological Society Digest 31: 177-187. [ Links ]
Macellari, C.E., Barrio, C.A. and Manassero, M.J. 1989. Upper Cretaceous to Paleocene depositional sequences and sandstone petrography of southwestern Patagonia (Argentina and Chile). Journal of South American Earth Sciences 2: 223-239. [ Links ]
Malumián, N. and Caramés, A. 1997. Upper Campanian-Paleogene from the Río Turbio coal measures in southern Argentina: micropaleontology and the Paleocene/Eocene boundary. Journal of South American Earth Sciences 10: 189-201. [ Links ]
Marenssi, S.A., Casadío, S. and Santillana, S.N. 2002. La Formación Man Aike al sur de El Calafate (Provincia de Santa Cruz) y su relación con la discordancia del Eoceno medio en la cuenca Austral. Revista de la Asociación Geológica Argentina 57: 341-344. [ Links ]
Marenssi, S., Guler, M.V., Casadío, S., Guerstein, G.R. and Papú, O. 2004. Sedimentology and bioestratigraphy of Maastrichtian deposits from Austral Basin, Argentina. Cretaceous Research 25: 907-918. [ Links ]
Morgan, R. 1977. Elucidation of the Cretaceous dinoflagellate Diconodinium Eisenack and Cookson, 1960, and related peridinioid species from Australia. Palynology 1: 123-138. [ Links ]
Nullo, F.E., Proserpio, C.A. and Blasco de Nullo, G. 1981. El Cretácico de la Cuenca Austral entre el Lago San Martín y Río Turbio. In: W. Volkheimer and E.A. Musacchio (eds.), Cuencas Sedimentarias del Jurásico y Cretácico de América del Sur, Buenos Aires, pp. 181-220. [ Links ]
Papú, O. 1997. Nueva especie de Grapnelispora del Campaniense-Maastrichtiense, provincia de Mendoza, Argentina. Inferencias filogenéticas, paleobiogeográficas y paleoambientales. Revista Española de Paleontología 12: 197-205. [ Links ]
Pascher, A. 1914. Über Flagellaten und Algen. Deutsche Botanische Gesellschaft : 32, 136-160. [ Links ]
Roncaglia, L., Field, B.D., Raine, J.I., Schiøler, P. and Wilson, G.J. 1999. Dinoflagellate bioestratigraphy of Piripauan-Haumurian (Upper Cretaceaous) sections from northeast South Island, New Zealand. Cretaceous Research 20: 271-314. [ Links ]
Ucha, N., de Santa Ana, H. and Veroslavsky, G. 2003. La cuenca Punta del Este: geología y potencial hidrocarburífero. In: G. Veroslavsky, M. Ubilla, and S. Martínez (eds.), Cuencas Sedimentarias de Uruguay: Geología, Paleontología y Recursos Naturales, Mesozoico . Edición: División Relaciones y Actividades Culturales de Facultad de Ciencias, Montevideo, pp. 171-190. [ Links ]
Williams, G.L., Lentin, J. and Fensome, R.A. 1998a. The Lentin and Williams Index of fossil dinoflagellates 1998 edition. American Association of Stratigraphic Palynologists , Contribution Series 34: 1- 817. [ Links ]
Williams, G.L., Brinkhuis, H., Bujak, J.P., Damassa, S.P., Hochuli, P.A., de Verteuil, L. and Zevenboom, D. 1998b. Dinoflagellates. In: Appendix to Mesozoic and Cenozoic sequence chronostratigraphic framework of European Basins by Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., de Graciansky, P-C. and Vail, P.R. (eds.). In: Mesozoic and Cenozoic Sequence Stratigraphy of European Basins. Society for Sedimentary Geology , Special Publication , 60, page 9 and chart 3. [ Links ]
Williams, G.L., Brinkhuis, H., Pearce, M.A., Fensome, R.A. and Weegink, J.W. 2004. Southern Ocean and global dinoflagellate cyst events compared: index events for the Late Cretaceous-Neogene. In: N.F. Exon, J.P. Kennett and M.J. Malone (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 189: 1-98. [ Links ]
Wilson, G.J. 1984. New Zealand late Jurassic to Eocene dinoflagellate biostratigraphy-a summary. Newsletters in Stratigraphy 13: 104-117. [ Links ]
Wood, S.E. and Askin, R.A. 1992. Dinoflagellate cysts from the Marambio Group (Upper Cretaceous) of Humps Island. Antarctic Science 4: 327-336. [ Links ]
Recibido: 9 de diciembre de 2003.
Aceptado: 29 de julio de 2004.
Appendix 1
List of dinocyst species names cited in the text
(See Williams et al ., 1998a for references, except when indicated).
Alisocysta circumtabulata (Drugg 1967) Stover and Evitt 1978.
Alisogymnium euclaense (Cookson and Eisenack 1971a) Lentin and Vozzhennikova 1990.
Alterbidinium acutulum (Wilson 1967b) Lentin and Williams 1985 emend. Khowaja-Ateequzzaman et al . 1991.
Areoligera medusettiformis Wetzel 1933b ex Lejeune-Carpentier 1938a.
Cerodinium dartmoorium (Cookson and Eisenack 1965) Lentin and Williams 1987.
Damassadinium californicum (Drugg 1967) Fensome et al. 1993b.
Diconodinium davidii Morgan 1975.
Diconodinium longicorne Olaru 1978a.
Dconodinium lurense sp. nov. herein.
Diconodinium martianum Srivastava 1995.
Diconodinium multispinum (Deflandre and Cookson 1955) Eisenack and Cookson 1960 emend . Morgan 1977.
Diconodinium pusillum Singh 1971.
Diconodinium vitricornu Roncaglia et al . 1999.
Dinogymnium undulosum Cookson and Eisenack 1970a.
Disphaerogena carposphaeropsis O. Wetzel 1933b emend . Sarjeant 1985b.
Manumiella ? cretacea (Cookson 1956) Bujak and Davies 1983.
Manumiella druggii (Stover 1974) Bujak and Davies 1983.
Odontochitina costata Alberti 1961 emend . Clarke and Verdier 1967.
Odontochitina operculata (O. Wetzel 1933a) Deflandre and Cookson 1955.
Palaeocystodinium australinum (Cookson 1965b) Lentin and Williams 1976.
Palaeoperidinium pyrophorum (Ehrenberg 1838 ex O. Wetzel 1933a) Sarjeant 1967b.
Senoniasphaera inornata (Drugg 1970b) Stover and Evitt 1978.
Tanyosphaeridium variecalamum Davey and Williams 1966b.
Trithyrodinium evittii Drugg 1967.

