










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.2 Buenos Aires mar./jun. 2005
Miospores and chlorococcalean algae from the Los Rastros Formation, Middle to Upper Triassic of central-western Argentina
Eduardo G. Ottone, Adriana C. Mancuso and Magdalena Resano
Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Pabellón n° 2, Ciudad Universitaria, C1428EHA Buenos Aires, Argentina. ottone@gl.fcen.uba.ar , amancu@gl.fcen.uba.ar, magdaresano@yahoo.com
Abstract. Lacustrine strata of the Los Rastros Formation (Middle to Upper Triassic) at Río Gualo section (La Rioja province), yield a distinctive palynological assemblage of miospores and chlorococcalean algae. The miospore association is characterized by a relative abundance of corystosperm pollen grains with subordinate inaperturates, diploxylonoid disaccates, spores, monocolpates, monosaccates and striate pollen grains. The phytoplankton are mostly represented by Botryococcus but also by Plaesiodictyon , a form probably related to the Hydrodictyaceae. Geological data and variations in phytoplankton content indicate that the lacustrine system probably evolved from a stretcht of freshwater with eutrophic conditions, into a body with oligotrophic conditions through the middle and upper part of the Río Gualo section. The genus Variapollenites is emended in order to amplify its original diagnosis.
Resumen. Miosporas y algas chlorococcales de la Formación Los Rastros , Triásico medio a superior del centro - oeste de Argentina. El estudio de los niveles lacustres de la Formación Los Rastros (Triásico Medio a Superior) en la sección de Río Gualo (provincia de La Rioja), incluye una interesante palinoflora compuesta por miosporas y algas Chlorococcales. Entre las miosporas abundan los granos de polen de Corystospermales, con presencia subordinada de inaperturados, disacados diploxilonoides, esporas, monocolpados, monosacados y polen estriado. En el fitoplancton se destaca Botryococcus , pero también se observa Plaesiodictyon , que es una forma probablemente relacionada con las Hydrodictyaceae. Datos geológicos y variaciones en el contenido fitoplanctológico señalan que el sistema lacustre probablemente evolucionó desde un cuerpo de agua dulce y condiciones eutróficas, a un ambiente de carácter oligotrófico hacia la parte media y superior de la sección de Río Gualo. Así mismo, se enmienda el género Variapollenites a fin de ampliar su diagnosis original.
Key words. Argentina; Triassic; Miospores and chlorococcalean algae; Systematics; Lacustrine environment.
Palabras clave. Argentina; Triásico; Miosporas y algas Chlorococcales; Sistemática; Ambiente lacustre.
Introduction
In the Triassic of southern South America, one of the best represented lacustrine successions is that of the Los Rastros Formation, which is part of the continental infilling of the Ischigulasto-Villa Unión Basin in central-westen Argentina (Stipanicic and Bonaparte, 1979; Stipanicic, 1983; López Gamundí et al ., 1989). The unit is characterized by thick deposits of black shales and fine grained sandstones that represent a cyclic lacustrine-deltaic environment. It crops out at different localities in La Rioja and San Juan provinces (figure 1) with slight lateral facies variation (de la Mota, 1946; Bossi, 1971; Milana and Alcober, 1994; Milana, 1998; Rogers et al ., 2001).
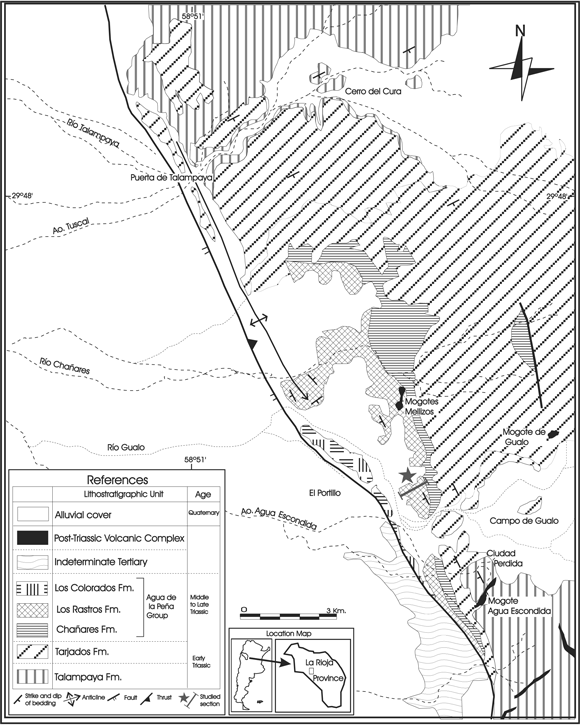
Figure 1. Location map of the Río Gualo section, Ischigualasto-Villa Unión Basin, La Rioja province, Argentina / mapa de ubicación de la sección de Río Gualo, Cuenca de Ischigualasto- Villa Unión, provincia de La Rioja, Argentina. Modified from Caselli (1998)/ modificado de Caselli (1998).
The fossil record of the Los Rastros Formation is mainly represented by floral remains. An abundant macrofloral assemblage, belonging to the Dicroidium - flora, was collected at different levels and localities in the Los Rastros sequence although, the most important records are known from the thick lacustrine black shales, close to the base of the unit (Frenguelli, 1948; Kokogian et al ., 1999; Spalletti et al ., 1999; Stipanicic and Bonetti, 1969; Stipanicic and Bossi, 2002b; Zamuner et al ., 2001). Palynological analysis are known only from selected areas in the basin (Herbst, 1965, 1970, 1972; Yrigoyen and Stover, 1970).
Invertebrate remains exhumed from Los Rastros levels include different groups of conchostracans (clamp-shrimps), insects and bivalves, and they occurr throughout the sequence (Gallego, 1997, 1999). Also, a quite rich invertebrate ichnological data has been mentioned from the unit (Melchor et al ., 2001; Mancuso, 2003). The vertebrate fauna mainly comprises actinopterigian fishes (Cabrera, 1944; Forster et al ., 1995, Mancuso, 2000, 2003), whereas tetrapods are represented only by a temnospondyl amphibian specimen (Contreras et al ., 1997; Mancuso, 2002) and several tetrapod footprints related to different archosaur groups (von Huene, 1931; Arcucci et al ., 1995; Marsicano et al ., 2004).
In the present contribution an interesting association of miospores and Chlorococcalean algae recovered from the Los Rastros sequence outcropping at the Río Gualo area, about 10 km to the southeast of the steep-walled, picturesque canyons of the Puerta de Talampaya, La Rioja province (figures 1, 2), are described. The Río Gualo section is near the western margin of the Ischigulasto-Villa Unión Basin (Kokogian et al ., 2001). The palynoflora, together with sedimentological data, provide new interpretative insights into the basin evolution.
Material and methods
A stratigraphic section of the Los Rastos Formation have been recognized at Rio Gualo. A detailed description of the sedimentology of the section was used to interpret the facies assemblage and to reconstruct the depositional environment. Systematic palynological sampling was conducted; and this paper is based on the study of 15 samples. Laboratory procedures followed conventional practices. Carbonates and silicates were removed by treatment with hydrochloric and hydrofluoric (70%) acids. Residues were sieved on a 25 µm mesh and mounted in unstained glycerine jelly as well as in Cellozise on glass slides. Specimens were examined under a Leitz Orthoplan binocular microscope. Photomicrographs were taken with a Leitz Orthomat camera using AGFAPAN APX25 and ILFORDPAN NF50 films. Palynological slides are prefixed BAFC-Pl. The illustrated specimens are identified with a slide number and England Finder coordinates. The slides are deposited at the Geological Sciences Department, Buenos Aires University. E.G. Ottone prepared the palynological section of the paper. A.C. Mancuso collected the samples and provided geological information. M. Resano collaborated in the study of the Chlorococcales.
Geological setting and age
During the pre-breakup stage of Pangea, during the Late Permian and Early Triassic, several extensional basins were formed along the western margin of Gondwana (Uliana and Biddle, 1988). The Ischigualasto-Villa Unión Basin is situated in centralwestern Argentina, with a NW-SE orientation, between San Juan and La Rioja provinces (figure 1).The sedimentary infilling of the basin consists of up to 4000 m of alluvial, fluvial and lacustrine deposits of Triassic age. The sedimentary succession has been subdivided lithostratigraphically, in ascending order, into the Talampaya and Tarjados formations (Romer and Jensen, 1966), and the Agua de la Peña Group, which comprises five units (de la Mota, 1946; Bossi, 1971; Stipanicic, 1983). The lower Chañares Formation (Bossi and Stipanicic, 2002a) is dominated by tuffaceous sandstones and siltstones deposited in a fluvial-lacustrine environment and grades upward into the lacustrine black shales and deltaic sandstones of the Ischichuca and Los Rastros formations (Bossi et al ., 2002; Stipanicic and Bossi, 2002a). This sequence passes upward into the sandstones, mudstones and tuffs of the Ischigualasto Formation (Stipanicic and Bossi, 2002b), deposited by a moderate- to high-sinuosity river system. The succession culminates in the red beds of the Los Colorados Formation (Bossi and Stipanicic, 2002b). A regional unconformity separates the Triassic succession from the overlying ?Cretaceous Cerro Rajado Formation (Caselli et al ., 2001).
The lacustrine-deltaic Los Rastros Formation is unconformably overlain by the fluvial Ischigualasto Formation and conformably succeeds the fluvial-lacustrine deposits of the Chañares and Ischichuca formations. The Los Rastros Formation deposits are generally interpreted as lacustrine-deltaic cycles with a predominance of lacustrine sedimentation (Stipanicic and Bonaparte, 1979; López Gamundí et al.,1989 ; Milana and Alcober, 1994; Milana, 1998; Rogers et al ., 2001).
The age of the Los Rastros Formation has been referred to the early Late Triassic by some authors, based on its palynological content (Zavattieri and Batten, 1996; Morel et al ., 2001). Nevertheless, an older age (Middle Triassic) has been proposed by other authors due to its stratigraphic relationships (Kokogian et al ., 2001). Thus, a radioisotopid age data from the base of the overlying Ischigualasto Formation (227.8 ± 0.3 Ma) suggests a minimun late Middle Triassic age for the Los Rastros Formation (Rogers et al ., 1993, 2001).
Sedimentology
Fieldwork at the Río Gualo section indicated five lacustrine-deltaic cycles each with two facies assemblages.
Lake facies assemblage
This assemblage is dominated by dark gray to black carbonaceous claystones. Iron mudstones and very fine-grained sandstones are interbedded. The beds are tabular. The clayshales are horizontally laminated, whereas the ironstones may be massive or laminated. Conchostracans, plant debris, insect impressions and rare fish body fossils are found in this assemblage, but with different preservation conditions in the different facies. The clayshales were deposited by suspension in distal-lake conditions whereas the ironstones were deposited by distal turbiditic currents (Mancuso, 2003).
Deltaic facies assemblage
This assemblage includes stream load sediments. The assemblage begins with alternative green siltstones and gray claystones, which form rhythmic packages of massive individual beds containing plant debris. Fine- to medium-grained sandstone beds are intercalated, and are more abundant and thicker upward. Small ripple cross-laminations characterize these sandstones, which also contain plant debris. Medium- to coarse-grained sandstones are dominant upsection. They begin with horizontal lamination, passing to planar cross-bedded and culminate with trough cross-bedded lamination. The sequence coarsens and thickens upwards. Fossil fish bodies and poorly preserved wood fragments occur in the horizontally laminated sandstones while only fossil wood remains were found in the planar and trough cross-bedded sandstones (Mancuso, 2003). In its top, the facies assemblage features dark gray mudstones and very fine-grained sandstones interbedded with planar and trough cross-bedded sandstones. They are internally massive or pass upward from plane to rippled-lamination, and contain a very high percentage of organic matter, mainly carbonized fragments. This facies assemblage represents the progradation of the fluvial system into the lake body (Mancuso, 2003).
Palynological results and discussion
Kerogen is generally abundant and it is composed by a variable amount of wood, cuticle, amorphous organic matter and terrestrial palynomorphs. Fiftyeight species of miospores and three species of Chlorococcalean algae have been recorded at the Río Gualo section (figure 3). Palynomorphs are fairly well preserved, with the exception of those found in samples 1434, 1441 and 1442, which are commonly crushed or corroded.
The palynoflora of the Los Rastros Formation at Río Gualo section is dominated by disaccate pollen grains of corystosperm affinity, especially Alisporites australis de Jersey. Also abundant are inaperturate pollen (mainly Inaperturopollenites reidi de Jersey, a probable Araucariaceae) and diploxylonoid disaccate pollen grains ( Platysaccus spp., Podocarpaceae?), together with spores of pteridophytes ( Cadargasporites spp., Clavatisporites conspicuus Playford, Converrucosisporites cameronii (de Jersey) Playford and Dettmann, Osmundacidites spp., Polypodiisporites ipsviciensis (de Jersey) Playford and Dettmann, Rugulatisporites spp.), lycophytes (mostly Aratrisporites compositus Volkheimer and Zavattieri, of pleuromeian affinity) and sphenophytes ( Calamospora impexa Playford). Ginkgo biloba -like monocolpates ( Cycadopites spp.), monocolpates with alveolar ektexine ( Brachysaccus sp. A), monosaccates ( Variapollenites spp., Coniferales), striate pollen grains ( Lueckisporites spp., Lunatisporites spp., Protohaploxypinus spp., Striatopodocarpites pantii (Jansonius) Balme), ephedroid pollen grains ( Equisetosporites cacheutensis Jain) and Cedrus -like pollen grains ( Cedripites spp.) are also present. Phytoplankton is represented by Botryococcus sp., Plaesiodictyon mosellanum ssp. Perforatum Wille and Plaesiodictyon mosellanum ssp. Symmetricum Brenner and Foster.
Comparison of the Río Gualo fossil palynoflora with local fossil macroflora is hindered by the fact that only scarce and generally unidentifiable plant macrofossil remains are present. However, plant macrofossil assemblages from other localities of the basin has strong affinities with the Río Gualo microflora. The commonest elements from the Los Rastros Formation are the Corystospermales (9 species), together with a consistent representation of Sphenophyta (6 species) and, in decreasing importance, Osmundaceae (3 species), Peltaspermales, Cycadales, Voltziaceae and Czekanowskiales (Zamuner et al ., 2001).
The palynoflora of the Río Gualo section resembles those Late Triassic assemblages grouped into the cool temperate Ipswich microflora (Dolby and Balme, 1976). This microfloral province has been recognized in the south of Australia, New Zealand, Antarctica (Victoria Land), South Africa and Argentina (Zavattieri and Batten, 1996), and is believed to represent plant communities characteristic of high latitudes (40º-70º S lat.).
The palynoflora of the Río Gualo section is, in general, comparable with those described from other Middle to Upper Triassic sequences of Argentina (Zavattieri and Batten, 1996), and particularly, from the Ischigualasto-Villa Unión Basin (Herbst, 1965, 1970, 1972; Yrigoyen and Stover, 1970; Zavattieri and Melchor, 1999; Zavattieri and Milana, 2000). Botryococcus was previously cited in the Triassic, from the fluvio-lacustrine Las Cabras and Casa de Piedra formations, Cuyo Basin (Zavattieri, 1991b; Barredo et al ., 1999; Rodríguez Amenábar and Ottone, 2002, 2003) while Plaesiodictyon is known from lacustrine levels of the Ischichuca Formation, Ischigualasto-Villa Unión Basin (Zavattieri and Melchor, 1999). The presence of planktonic forms, mainly Botryococcus which has extant representatives of known ecological preferences, facilitate palaeoenvironmental interpretations about the lacustrine system at the Río Gualo area (Batten and Grenfell, 1996; Guy-Ohlson, 1992).
Botryococcus Kützing 1849, is an euryhaline, freshwater chlorococcalean that grows preferently in shallow, oligotrophic lakes or ponds (Tyson, 1995; Rodríguez Amenábar and Ottone, 2003). It is relatively abundant (about 5-10% of total palynomorphs content) in levels BAFC-Pl 1433, 1437, 1444. In samples 1431, 1432, 1435, 1436, 1438, 1439, 1440, 1443 and 1445, Botryococcus occurs in moderate quantities (less than 5% of total palynomorphs content). In levels 1441 and 1442, no Botryococcus has been recorded (figure 2). Structureless mass was found in the whole section (figure 5.K) suggesting that the colonies growth either in a stressed environment or with restricted access to oxygen during deposition (Guy-Ohlson, 1992, 1998; Guy-Ohlson and Lindström, 1994; Rodríguez Amenábar and Ottone, 2003). Level 1433 contains simple, globular colonies (40-70 µm in diameter), together with compound colonies showing branching processes and compound colonies with botryoidal form. In level 1437, there are large, compound colonies, with or without branches, with size range of 80-145 µm; and young simple colonies of about 60 µm in diameter (figures 5.G-H). The variable forms found in samples 1433 and 1437 suggest varying seasonal conditions of environment and/or climate (Guy-Ohlson, 1992, 1998; Guy-Ohlson and Lindström, 1994; Rodríguez Amenábar and Ottone, 2003). In level 1444, numerous large, botryoidal compound colonies are present, suggesting a uniform seasonal growth. Botryococcus is unusually abundant in this sample, making up about 99% of the palynoflora. In general, the occurrences of phytoplankton blooms follow occasional freshening stages in the water body, due to heavy rains or changes in water inflow that increased the availability of dissolved phosphorous in the basin (Warren, 1986).

Figure 2. Schematic section of the Los Rastros Formation in Río Gualo indicating palyniferous and other fossiliferous horizons / sección litológica de la Formación Los Rastros en Río Gualo incluyendo horizontes con palinomorfos y otros fósiles. Modified from Mancuso (2003) / modificado de Mancuso (2003).

Figure 3. Stratigraphic distribution of palynomorphs in samples studied / distribución estratigráfica de los palinomorfos en los niveles estudiados.
Plaesiodictyon Wille 1970 is a freshwater form probably related to the Hydrodictyaceae (Wood and Benson, 2000). It is relatively abundant in sample 1431 (about 5% of total palynomorphs content), where is the most consistently represented planktonic form, and is also present, as a minor constituent in samples 1432, 1433 and 1435 (less than 5% of total palynomorphs content) (figure 2, figures 5.E-F, I). The Río Gualo coenobia of Plaesiodictyon lack any dehiscent slit in its cells. This feature, in extant planar coenobia of Hydrodictyaceae, currently reflects a decrease in ability of cell division due to changes in the environment, rising salinity, stressed conditions or competition for nutrients (Brenner and Foster, 1994). Plaesiodictyon is strikingly similar to Pediastrum Meyen 1829, both genera display planar coenobia commonly with holes, and marginal cells usually with lobes and processi. The main difference between this two genera lies in the cell arrangement, which determine the shape of the coenobia. While Pediastrum is circular in outline and has a circular arrangement of cells, Plaesiodictyon displays rectangular coenobia with cells in perpendicular rows (Brenner and Foster, 1994; Komárek and Jankovská, 2001). However, an extant species of Pediastrum , ( P . orbitale Komárek 1983) includes circular and oval coenobia. It is difficult to know, simply by analogy with living Pediastrum , the precise ecological needs of Plaesiodictyon , mostly considering that extant Pediastrum includes many species and subspecies which display different ecological preferences (Komárek and Jankovská, 2001).
Changes in relative abundances of Botryococcus and Pediastrum (Tyson, 1995), or by roughly analogy, Botryococcus and Plaesiodyction , are considered a consequence of ecological changes. Therefore, the variable percentages of these coccal forms throughout the Río Gualo section could indicate that depositional conditions varied through time. The low Botryococcus / Plaesiodictyon ratio at the base of the sequence and its increase towards upsection would presumably reflect changing environmental conditions. Initially, a stretch of freshwater of probably eutrophic conditions would develop, mainly at the level represented by sample 1431, where the eutrophic character appears reinforced by the presence of abundant microscopic, ill preserved filaments of probable algal affinity (A. Godeas, personal communication). This situation would change, through the middle and upper part of the Río Gualo section, to a milieu of oligotrophic conditions.
From a sedimentological viewpoint, each cycle was inferred in the Río Gualo section (figure 2), according to the parasequence criterion (Vail et al ., 1991), as shallowing-upward. Gore (1988) noted that the lakes gradually infilled by sediment from deltaic input and shoreline progradation, produce regressive, coarsening-upward, lacustrine sequences overlain by a fluvial system. This is apparently the case with the Ischigualasto-Villa Unión Basin, wherein each cycle starts with fine sediments deposited in a lacustrine environment and finishes with sandstones of fluvial origin. The palynological analyses indicates that the richest assemblages are those associated with the lacustrine (pelitic) facies. Moreover, the most abundant phytoplankton occurr in the upper distallake facies of each cycle. This facies is characterized by black shales which suggest an anoxic bottom; furthermore, the local absence of grazing trails reinforce this hypothesis. The anoxic conditions might result from stratification of the water column, since the circulation of the oxygen water would be blocked by the stratification, thus preventing the oxygenation of the bottom (Wetzel, 1981). Bottom anoxic conditions are typically produced by a significant lake deepening, however, if there are high organic productivity in saline and shallow waters, the black shales may be accumulated (Gore,1988).
The scarce records of phytoplankton blooms through the section appear to indicate that the lake was probably deficient in dissolved phosphorous (Warren, 1986). The chemical composition of the lake could explain the lack of fish bones in the distal-lake facies.
The conspicuous record of conchostracans through the section indicate the presence of temporary freshwater bodies with development of alternating dry and wet bottom conditions (Vannier et al ., 2003). Adaptations to environmental stress conditions of growth are also reflected in the morphology of the phytoplanctonic forms (structureless mass in Botryococcus , lack of dehiscent slits in Plaesiodictyon ).
Systematic palynology
A complete list of palynomorphs recovered at Río Gualo section is given. Spore and pollen grains genera are listed alphabetically. Taxa are only described systematically, in the case of the emendation of the genus Variapollenites Ottone, or when an additional comment was deemed requisite.
Spores
Anapiculatisporites sandrae Ottone in Ottone et al . 1992
Aratrisporites compositus Volkheimer and Zavattieri 1991
Aratrisporites sp. cf. A. parvispinosus Leschik 1955
Baculatisporites comaumensis (Cookson) Potonié 1956
Biretisporites sp.
Cadargasporites baculatus de Jersey and Paten 1964 emend . Reiser and Williams 1969
Cadargasporites cuyanensis Azcuy and Longobucco 1983
Calamospora impexa Playford 1965
Clavatisporites conspicuus Playford in Playford et al. 1982
Clavatisporites sp.
Converrucosisporites cameronii (de Jersey) Playford and Dettmann 1965
Converrucosisporites sp.
Deltoidospora sp.
Leptolepidites volkheimeri Zavattieri 1986
Leptolepidites sp. cf. L. crassibalteus Filatoff 1975
Leptolepidites sp.
Lundbladispora spp.
Osmundacidites senectus Balme 1963
Osmundacidites wellmanii Couper 1958
Neoraistrickia sp.
Playfordiaspora cancellosa (Playford and Dettmann) Maheshwari and Banerji 1975
Polypodiisporites ipsviciensis (de Jersey) Playford and Dettmann 1965
Punctatisporites sp.
Rugulatisporites nelsonensis Raine in de Jersey and Raine 1990
Rugulatisporites sp.
Uvaesporites verrucosus (de Jersey) Helby in de Jersey 1971
Pollen grains
Alisporites australis de Jersey 1962
Alisporites lowoodensis de Jersey 1963
Alisporites parvus de Jersey 1962
Alisporites spp.
Araucariacites australis Cookson 1947
Brachysaccus sp. A
Cedripites tectus Ottone and Rodríguez Amenábar 2001
Cedripites sp. cf. C. pannellai Jain 1968
Cedripites sp. cf. C. priscus Balme 1970
Chordasporites australiensis de Jersey 1962
Cycadopites sp. cf. C. andrewsii Cornet and Traverse 1975
Cycadopites sp. cf. C . follicularis Wilson and Webster 1946
Cycadopites sp. A
Equisetosporites cacheutensis Jain 1968
Inaperturopollenites nebulosus Balme 1970
Inaperturopollenites reidi de Jersey 1959
Lueckisporites spp.
Lunatisporites sp. cf. L. noviaulensis (Leschik) Foster 1979
Lunatisporites spp.
Platysaccus papilionis Potonié and Klaus 1954
Platysaccus queenslandi de Jersey 1962
Protodiploxypinus sp.
Protohaploxypinus sp. cf. P. limpidus (Balme and Hennelly) Balme and Playford 1967
Protohaploxypinus sp. cf. P. microcorpus (Schaarschmidt) Clarke 1965
Protohaploxypinus sp. cf. P. varius (Bharadwaj) Balme 1970
Protohaploxypinus spp.
Pteruchipollenites spp.
Striatopodocarpites pantii (Jansonius) Balme 1970
Variapollenites curviplicatus Ottone in Ottone et al . 1992
Variapollenites rhombicus Ottone in Ottone et al. 1992
Variapollenites trisulcus Ottone in Ottone et al. 1992
Vitreisporites pallidus (Reissinger) Nilsson 1958
Phytoplankton
Botryococcus sp.
Plaesiodictyon mosellanum ssp. perforatum Wille 1970
Plaesiodictyon mosellanum ssp. symmetricum Brenner and Foster 1994
Genus Brachysaccus Mädler 1964
Type species. Brachysaccus ovalis Mädler 1964.
Remarks. Following Potonié (1970) and de Jersey (1972), it is considered that Brachysaccus includes monosulcate pollen grains having an endoreticulate exine. The monosulcate character of the genus is clearly evident in Mädler's (1964) illustration of the type. Amb and sulcus shape are extremely variable in monosulcate pollen grains displaying a homogeneous or granular ektexine. This is extensively discussed below under the genus Cycadopites Wodehouse ex Wilson and Webster 1946. In Brachysaccus , a monosulcate pollen grain with an alveolar ektexine, amb and sulcus shape are relatively uniform, and its structured ektexine probably controls, in different states of pollen hydration, the contraction or stretching of the wall to a significant extent.
Brachysaccus sp. A
Figures 4.J, L

Figure 4. A-D, Variapollenites trisulcus Ottone; A, BAFC-Pl 1431(14) W42/2; B, BAFC-Pl 1431(10) R37/3; C, BAFC-Pl 1431(12) Z39/1; D, BAFC-Pl 1431(11) Y42/0. E-F, H, Variapollenites curviplicatus Ottone; E, BAFC-Pl 1431(11) C22/2, proximal view/ vista proximal ; F, BAFC-Pl 1431(13) T26/1; H, BAFC-Pl 1431(11) C22/2, distal view/ vista distal . G, I, Variapollenites rhombicus Ottone; G, BAFC-Pl 1431(8) Q30/4; I, BAFC-Pl 1431(3) Q36/4. J, L, Brachysaccus sp. A; J, BAFC-Pl 1431(11) T30/2; L, BAFC-Pl 1431(11) G41/4. K, Equisetosporites cacheutensis Jain BAFC-Pl 1431(11) Z26/2. Scale bar/ escala gráfica = 30 µm.

Figure 5. A, D, Cycadopites sp. A; A, BAFC-Pl 1432(2) Q50/0; D, BAFC-Pl 1433(1) J39/3. B-C, Cycadopites sp. cf. C. follicularis Wilson and Webster; B, BAFC-Pl 1432(2) T41/4; C, BAFC-Pl 1432(2) K53/4. E-F, Plaesiodictyon mosellanum ssp. symmetricum Brenner and Foster; E, 1431(11) N35/2; F, 1431(11) Q24/0. G-H, J-K, Botryococcus sp.; G, BAFC-Pl 1437(1) U55/0, botryoidal colony/ colonia botroidal; H, BAFC-Pl 1437(1) Y52/0, compound colony showing branches/ colonia compuesta con ramas ; J, BAFC-Pl 1443(1) L51/3, botryoidal colony/ c olonia botroidal ; K, BAFC-Pl 1444(1) O41/3, structureless mass/ masa amorfa. I, Plaesiodictyon mosellanum ssp. Perforatum Wille 1970 BAFC-Pl 1431(12) S43/0. Scale bar/ escala gráfica = 30 µm.
Material estudiado. BAFC-PI 1431 (11) T30/2; BAFC-PI 1431 (11) G41/4.
Description. Monosulcate pollen grain. Amb almost circular. Wall endoreticulate (alveolar ektexine). Exine thickness 3.5-5.5 µm. Ektexine infratectal bacula-like elements forming brochi, 1-2.5 µm in diameter, which are perpendicularly disposed all around the grain. Prothallic pole without apertures, lacking folds or splits. Germinal pole defined by a narrow sulcus, ca. about one tenth of the grain diameter. Sulcus margins occasionally accompanied by folds.
Measurements (25 specimens). Equatorial diameter 55(85)106 µm.
Comparison. Judging from its illustration, Brachysaccus triassicus Tripathi et al . 1990 seems to be comparable to B. sp. A, however, the Indian form was originally described as disaccate. B . sp. A differs from B. ovalis Mädler 1964 and B. eskensis de Jersey 1972 by its almost circular amb. B. indicus Kumaran and Maheshwari 1980, originally also described as disaccate, displays an elliptical to oval amb. The specimen figured by Zavattieri (1991a) as Brachysaccus cf. neomundanus (Leschick) Mädler 1964, from the Triassic Las Cabras Formation, Cuyo Basin, Argentina, is disaccate.
Genus Cycadopites Wodehouse ex Wilson and Webster 1946
Type species. Cycadopites follicularis Wilson and Webster 1946.
Discussion. Following Ottone and García (1991), Cycadopites includes monosulcate pollen grains having a fusiform, oval, subpolygonal to subcircular amb, with a homogeneous wall (homogeneous or granulate ektexine), prothallic pole without apertures; germinal pole defined by a sulcus extending over the full grain length, and absent or minute surficial sculpture. Extant pollen grains of Ginkgophytes, Cycadophytes and some Magnoliaceae, together with fossil pollen grains of Bennettitales, Caytoniales, ?Corystospermales and Pentoxylales are entirely comparable to those here observed (Ottone and García, 1991). Figures 6.A-F are different specimens of extant Ginkgo biloba Linnaeus collected in situ from a microsporangiate organ in Buenos Aires, and mounted, as well as the fossil material, in unstained glycerin jelly. See also the different specimens of G. biloba illustrated by Audran and Masure (1978, plate I). In G . biloba , pollen grain shape (and sulcus shape) reflect differences in orientation within the slide or, most probably, variable states of pollen grain hydration. Furthermore, generic and specific distinctions of fossil pollen grains similar to G. biloba are impossible only using transmitted light microscopy (Ottone and García, 1991). In the Triassic of Argentina, Jain (1968), Herbst (1970), Zavattieri (1987, 1991b, 1992) and Zavattieri and Batten (1996) used different genera for monosulcate pollen grains; e.g ., Bharadwajapollenites Jain 1968, Cycadopites Wodehouse ex Wilson and Webster 1946, Monosulcites Cookson ex Couper 1953, Megamonoporites Jain 1968, Pilasporites Balme and Hennelly emend . Jain 1968, Punctamonocolpites Pierce 1961, and Sulcatopites Jain 1968. These forms should be Cycadopites in view of the above discussion.
Cycadopites sp. cf. C. follicularis Wilson and Webster 1946
Figures 5.B-C
Material estudiado. BAFC-PI 1432 (2) T41/4; BAFC-PI 1432 (2) K53/4.
Description. Monosulcate pollen grain, wall smooth, ca. 0.8-1 µm thick.
Measurements (3 specimens). Equatorial diameter 45(46)48 µm.
Comparison. The Argentine specimens are comparable, except for their thinner exine, with the miospores described by Wilson and Webster (1946). Balme (1970) also included in the species Cycadopites follicularis specimens with an exine about 1 µm thick.
Cycadopites sp. A
Figures 5.A, D
Material estudiado. BAFC-PI 1432 (2) Q50/0; BAFC-PI 1433 (1) J39/3.
Description. Monosulcate pollen grain, wall smooth to chagrinate, ca. 1.2 µm thick. In fusiform specimens, sulcus margin occasionally folded.
Measurements (8 specimens). Equatorial diameter 62(77)101 µm.
Comparison. Specific differences within the genus are often subtle and hard to observe; hence, Cycadopites sp. A superficially resembles many species referred to Cycadopites or related genera. Cycadopites sp. A essentially differs from the type of Cycadopites by its larger size. It resembles specimens referred by Jain (1968) to Pilasporites calculus Balme and Hennelly 1956 and P. crateriformis Jain 1968. Cycadopites linae Herbst 1965 and C. magnus Herbst 1965, cited later by Herbst (1970), Jain (1968) and Zavattieri (1987, 1991b), are also similar to Cycadopites sp. A.
Genus Equisetosporites Daugherty emend . Pocock and Jansonius in Pocock, 1964
Type species. Equisetosporites chinleana (Daugherty) Pocock and Jansonius in Pocock, 1964.
Equisetosporites cacheutensis Jain 1968
Figure 4.K
Material estudiado. BAFC-PI 1431 (11) Z26/2.
Discussion. Following Jain (1968), this species is retained in Equisetosporites . Generic re-attribution is not justified for E. cacheutensis because, in acceptance of its probable ephedroid affinity, pollen grains that display a clear separation between the nexine (endexine?) and the sexine (ektexine?) also characterize extant Ephedra americana Humboldt and Bonpland ex Willdenow (Steeves and Barghoorn,1959), and the Triassic Masculostrobus clathratus Ash 1972 (a probable gnetalean). The type species of Equisetosporites , E. chinleana Daugherty 1941, also displays a central endexinal core detached from its banded outer wall. However, attribution of E. cacheutensis to Equitetosporites , implies a slightly expansion of Pocock and Jansonius' (in Pocock, 1964) circumscription of the genus.
Genus Variapollenites Ottone in Ottone et al ., 1992, emend . Ottone
Type species. Variapollenites curviplicatus Ottone in Ottone et al. , 1992.
Discussion. Variapollenites was first described from the Triassic Rancho de Lata Formation, Main Cordillera, Argentina (Ottone et al ., 1992). The genus encompasses monosaccate pollen grains that display a reticulate pattern on the exine of both the saccus and corpus. Several well-preserved specimens of Variapollenites have been found at Río Gualo section, and also in equivalent horizons at the Hoyada de Ischigualasto. The Los Rastros specimens are better preserved than the ones of the type material, thus providing the basis for an emended generic diagnosis. The Los Rastros specimens show differences in ektexine endoreticulum of opposite faces of the corpus not clearly distinguishable in the (lesser preserved) type material due to preservation.
Emended diagnosis. Radial to bilaterally symmetrical monosaccate pollen grains. Tetrad mark absent. Ektexine endoreticulate, detached from endexine toward the equator. The endoreticulum shows a tendency to radial elongation with larger brochi towards the saccus margin (alveolar ektexine). Endexine forms an almost circular or roundly polygonal, mainly triangular or rhombic, corpus (polar view). Corpus margin exhibits a circumpolar fold system. On one face, the ektexine of the corpus is finely structured (ektexine incipiently alveolar?). On the opposite face, corpus ektexine is finely structured towards the equator, becoming smooth and thinner towards the center. Saccus of almost uniform width, not lobed, or constricted; tri- or quadrilobated.
Comparison. Aberrant extant monosaccate grains of Pinaceae (Vishnu-Mittre, 1957, pl. XX, 10) and Podocarpaceae (Pocknall, 1981, fig. 9, I) are comparable to the type species of Variapollenites , as are specimens figured as cf. Walchia by Reissinger (1950, pl. XVII, fig. 22) from the Lias of Germany, and Monosaccate, undet. by Dybkjaer (1988, pl. 6, fig. 7) from the Lias of Denmark. Walchites gradatus Bolkhovitina 1956, from the Jurassic of Russia, seems to be structurally similar to V. curviplicatus Ottone, but its corpus lacks a circumpolar fold system. Picea sp. from the Miocene of Oregon (Gray, 1964, pl. I, fig. 1) exhibits a morphology closely akin to that of V . rhombicus (for further comparison see Ottone et al ., 1992).
Variapollenites curviplicatus Ottone in Ottone et al. 1992
Figures 4.E-F, H
Material estudiado. BAFC-PI 1431 (11) C22/2; BAFC-PI 1431 (13) T26/1; BAFC-PI 1431 (11) C22/2.

Figure 6. A-F, Pollen grains of extant Ginkgo biloba Linnaeus collected in situ from a microsporangiate organ in Buenos Aires. Specimens show variations in the shape of pollen grain and sulcus / granos de polen de Ginkgo biloba actual colectados in situ de un microsporangio en Buenos Aires. Los especímenes muestran variaciones en la forma del grano y del sulco. Scale bar/ escala gráfica = 30 µm.
Discussion. The species, as viewed in polar orientation, includes suboval monosaccates displaying a subcircular corpus. The saccus is not lobed. The specimens from Los Rastros often show, conversely to the ones of the type material, a saccus of variable width.
Variapollenites rhombicus Ottone in Ottone et al . 1992
Figures 4.G, I
Material estudiado. BAFC-PI 1431 (8) Q30/4; BAFC-PI 1431 (3) Q36/4.
Discussion. As viewed in polar orientation, the species includes suboval to subrectangular monosaccates with a roundly rhomboidal corpus. The saccus is quadrilobed and constricted at the apices of the corpus. The Los Rastros material include subrectangular forms with a slightly constricted saccus (figure 13), as well as specimens having a highly incised saccus.
Variapollenites trisulcus Ottone in Ottone et al . 1992
Figures 4.A-D
Material estudiado. BAFC-PI 1431 (14) W42/2; BAFC-PI 1431 (10) R37/3; BAFC-PI 1431 (12) Z39/1; BAFC-PI 1431 (11) Y42/0.
Discussion. The species, as viewed in polar orientation, includes subtriangular monosaccates with a roundly triangular corpus. The saccus is trilobate and often constricted at the corpus apices. The saccus of specimens from Los Rastros is, in some cases, less constricted that those of the type specimens.
Acknowledgements
The authors are indebted to C. Marsicano and S. Vigna whose comments and suggestions improved the manuscript; to A. Godeas for helpful advice; to C.L. Azcuy for providing the slides of Ginkgo biloba ; to A. Arcucci for an enjoyable productive field trip to La Rioja province; and to G.D. Holfeltz for laboratory processing and photographic printing. We also gratefully acknowledge the Buenos Aires University (UBA) and the National Research Council (CONICET) for the use of their facilities during this research. This investigation was mostly supported by UBA (TX 179) and CONICET (PIP 02228) grants.
References
Arcucci, A.B., Forster, C., Abdala, F., May, C. and Marsicano, C., 1995. Teropods tracks from the Los Rastros Formation (Middle Triassic), La Rioja Province, Argentina. Journal of Vertebrate Paleontology 15, supplement 3: 16A. [ Links ]
Ash, S.R. 1972. Late Triassic plants from the Chinle Formation in north-eastern Arizona. Palaeontology 15: 598-618. [ Links ]
Audran, J.C. and Masure, E. 1978. La sculpture et l'infrastructure du sporoderme de Ginkgo biloba comparées à celles des enveloppes polliniques des Cycadales. Review of Palaeobotany and Palynology 26: 363-387. [ Links ]
Azcuy, C.L. and Longobucco, M.I. 1983. Palinología de las Formaciones Potrerillos y Cacheuta, provincia de Mendoza, Argentina. Ameghiniana 20: 297-316. [ Links ]
Balme, B.E. 1963. Plants fossils from the Lower Triassic of Western Australia. Palaeontology 6: 12-40. [ Links ]
Balme, B.E. 1970. Palynology of Permian and Triassic strata in the Salt Range and Surghar Range, West Pakistan. In: B. Kummel and C. Teichert (eds.), Stratigraphic boundary problems: Permian and Triassic of West Pakistan. Department of Geology-University of Kansas, Special Publication 4, pp. 305-453. [ Links ]
Balme, B.E. and Hennelly, J.P.F. 1956. Trilete sporomorphs from Australian Permian sediments. Australian Journal of Botany 4: 240-260. [ Links ]
Barredo, S.P., Ottone, E.G., Marsicano, C.A. and Rodríguez Amenábar, C. 1999. Continental biotic association of the Triassic Rincón Blanco Subasin, Argentina. 7º International Symposium on Mesozoic Terrestrial Ecosystems, (Buenos Aires), Abstracts : 8-9. [ Links ]
Batten, D.J. and Grenfell, H.R. 1996. Chapter 7D. Botryococcus. In: Jansonius, J. and McGregor, D.C. (ed.), Palynology: principles and applications, American Association of Stratigraphic Palynologists Foundation, 1: 205-214. [ Links ]
Bolkhovitina, N.A. 1956. Atlas of spores and pollen from Jurassic and Lower Cretaceous deposits of the Vilyui Depression. Trudy Geologicheskij Institut, Akademija Nauk SSSR 2, 188 p. (in Russian). [ Links ]
Bossi, G. 1971. Análisis de la Cuenca Ischigualasto-Ischichuca. 1º Congreso Hispano-Luso Americano Geología Económica, (Madrid- Lisboa), Actas 2: 611-626. [ Links ]
Bossi, G. and Stipanicic, P.N. 2002a. Formación Chañares. In: P.N. Stipanicic and C.A. Marsicano (ed.), L éxico Estratigráfico de la Argentina. Volumen VIII. Triásico. Asociación Geológica Argentina, Buenos Aires, Serie B (Didáctica y Complementaria) 26, pp. 80-82. [ Links ]
Bossi, G. and Stipanicic, P.N. 2002b. Formación Los Colorados. In: P.N. Stipanicic and C.A. Marsicano (ed.), Léxico Estratigráfico de la Argentina. Volumen VIII. Triásico. Asociación Geológica Argentina, Buenos Aires, Serie B (Didáctica y Complementaria) 26, pp. 178. [ Links ]
Bossi, G., Stipanicic, P.N., Georgieff, S.M. and Milana, J.P. 2002. Formación Ischichuca. In: P.N. Stipanicic and C.A. Marsicano (ed.), Léxico Estratigráfico de la Argentina. Volumen VIII. Triásico. Asociación Geológica Argentina, Buenos Aires, Serie B (Didáctica y Complementaria) 26, pp. 146-147. [ Links ]
Brenner, W. and Foster, C.B. 1994. Chlorophycean algae from the Triassic of Australia. Review of Palaeobotany and Palynology 80: 209-234. [ Links ]
Cabrera, A. 1944. Dos nuevos peces ganoideos del triásico de Argentina. Notas del Museo de La Plata , 9, Paleontología 81: 569-576. [ Links ]
Caselli, A.T. 1998. [ Estratigrafía y sedimentología de las formaciones Patquía (Pérmico) y Talampaya (Triásico inferior) en las Sierras Pampeanas Noroccidentales y Precordillera Central (provincias de La Rioja y San Juan). Universidad de Buenos Aires, Tesis Doctoral, Buenos Aires, 437 pp. Unpublished]. [ Links ]
Caselli, A.T., Marsicano, C.A. and Arcucci, A.B. 2001. Sedimentología y paleontología de la Formación Los Colorados, Triásico superior (provincias de La Rioja y San Juan, Argentina). Revista de la Asociación Geológica Argentina 56: 173-188. [ Links ]
Clarke, R.F.A. 1965. British Permian saccate and monosulcate miospores. Palaeontology 8: 322-354. [ Links ]
Contreras, V.H., Bracco, A. and Sill, W. 1997. Primer registro de restos de tetrápodos en la Formación Los Rastros (Triásico Medio), Parque Provincial Ischigualasto, San Juan, Argentina. Ameghiniana 34: 534. [ Links ]
Cookson, I.C. 1947. Plant microfossils from the lignites of Kerguelen Archipielago . British, Australian and New Zealand Antarctic Research Expedition 1929-31 Reports, Series A, 2: 127-142. [ Links ]
Cornet, B. and Traverse, A. 1975. Palynological contributions to the chronology and stratigraphy of the Hartford Basin in Connecticut and Massachusetts. Geoscience and Man 11: 1-33. [ Links ]
Couper, R.A. 1953. Upper Mesozoic and Cainozoic spores and pollen grains from New Zealand. New Zealand Geological Survey, Paleontological Bulletin 22, 77 pp. [ Links ]
Couper, R.A. 1958. British Mesozoic microspores and pollen grains. A systematic and stratigraphic study. Palaeontographica B 103: 75-179. [ Links ]
Daugherty, L.H. 1941. The Upper Triassic flora of Arizona. Carnegie Institution Washington Publication 526, 108 pp. [ Links ]
de Jersey, N.J. 1959. Jurassic spores and pollen grains from the Rosewood Coalfield. Queensland Government Mining Journal 60: 346-366. [ Links ]
de Jersey, N.J. 1962. Triassic spores and pollen grains from the Ipswich Coalfield. Geological Survey of Queensland, Publication 307: 1-18. [ Links ]
de Jersey, N.J. 1963. Jurassic spores and pollen from the Marburg Sandstone. Geological Survey of Queensland, Publication 313: 1-15. [ Links ]
de Jersey, N.J. and Paten, R.J. 1964. Jurassic spores and pollen grainsfrom the Surat Basin. Geological Survey of Queensland, Publication 322: 1-18. [ Links ]
de Jersey, N.J. 1972. Triassic miospores from the Esk Beds. Geological Survey of Queensland, Publication 357, Palaeontological Papers 32, 40 pp. [ Links ]
de la Mota, H. F. 1946. [ Estudios geológicos en el Cerro Bola, al sur de Villa Unión, Dpto. General Lavalle, provincia de La Rioja. Universidad Nacional de La Plata, Tesis Doctoral 87, La Plata, 145 pp. Unpublished]. [ Links ]
Dolby, J.H. and Balme, B.E. 1976. Triassic palynology of the Camarvon Basin, Western Australia. Review of Palaeobotany and Palynology 22: 105-168. [ Links ]
Dybkjaer, K. 1988. Palynological zonation and stratigraphy of the Jurassic section in the Gassum Nº 1-borehole, Denmark. Danmkarks Geologiske Undergelgerse, DGU Serie A 21, 72 pp. [ Links ]
Filatoff, J. 1975. Jurassic palynology of the Perth Basin, Western Australia. Palaeontographica B 154: 1-113. [ Links ]
Forster, C., Arcucci, A., Marsicano, C., Abdala, F. and May, C. 1995. New vertebrate material from the Los Rastros Formation (Middle Triassic), La Rioja Province, Northwestern Argentina. Journal of Vertebrate Paleontology 15, supplement 3: 29A. [ Links ]
Foster, C.B. 1979. Permian plant microfossils of the Blair Athol Coal Measures, Baralaba Coal Measurs, and basal Rewan Formation of Queensland. Geological Survey of Queensland, Publication 372, Palaeontological Paper 45: 1-244. [ Links ]
Frenguelli, J. 1948. Estratigrafía y edad del llamado Rético en la Argentina. Anales de la Sociedad Argentina de Estudios Geográficos, Sociedad Argentina de Estratigrafía Geológica, GAEA 8: 159-310. [ Links ]
Gallego, O.F. 1997. Hallazgos de Insectos Triásicos en la Argentina. Ameghiniana 34: 511-516. [ Links ]
Gallego, O.F. 1999. [ Estudio sistemático de las faunas de conchostracos triásicos de la República Argentina. Universidad Nacional de Córdoba, Tesis Doctoral, Córdoba, 210 pp. Unpublished]. [ Links ]
Gore, P.J.W. 1988. Chapter 15. Late Triassic and Early Jurassic lacustrine sedimentation in the Culpeper basin Virginia. In: W. Manspeizer (ed.), Triassic-Jurassic rifting. Continental breakup and the origin of the Atlantic Ocean and Passive Margins. Part A. Elsevier Scientific Publications, Amsterdam, pp. 369-396. [ Links ]
Gray, J. 1964. Northwest American Tertiary palynology: The emerging picture. In: L.M. Cranwell (ed.), Ancient Pacific floras. The pollen story . University of Hawaii Press, pp. 21-30. [ Links ]
Guy-Ohlson, D. 1992. Botryococcus as an aid in the interpretation of paleoenvironment and depositional processes. Review of Palaeobotany and Palynology 71: 1-15. [ Links ]
Guy-Ohlson, D. 1998. The use of the microalga Botryococcus in the interpretation of lacustrine environments at the Jurassic-Cretaceous transition in Sweden. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 347-356. [ Links ]
Guy-Ohlson, D. and Lindström, S. 1994. Palaeoecology of the Early Permian strata at Heimefrontfjella, Dronning Maud Land, Antarctica. Antarctic Science 6: 507-519. [ Links ]
Herbst, R. 1965. Algunos esporomorfos del Triásico de Argentina. Ameghiniana 4: 141-155. [ Links ]
Herbst, R. 1970. Estudio palinológico de la Cuenca Ischigualasto-Villa Unión, (Triásico), provincias de San Juan- La Rioja. I. Introducción. II. Monoaperturados. Ameghiniana 7: 83-97. [ Links ]
Herbst, R. 1972. Estudio palinológico de la Cuenca Ischigualasto-Villa Unión (Triásico), provs. San Juan-La Rioja. III. Esporas triletes. Ameghiniana 9: 280-288. [ Links ]
Jain, R.K. 1968. Middle Triassic pollen grains and spores from Minas de Petroleo beds of the Cacheuta Formation (Upper Gondwana), Argentina. Palaeontographica B 122: 1-47. [ Links ]
Kokogian, D.A., Spalletti, L.A., Morel, E.M., Artabe, A.E., Martínez, R.N., Alcober, O.A., Milana, J.P. and Zavattieri, A.M. 2001. Estratigrafía del Triásico argentino. In: A.E. Artabe, E.M. Morel and A.B. Zamuner (ed.), El Sistema Triásico en la Argentina , Fundación Museo de La Plata Francisco Pascasio Moreno, La Plata, pp. 23-54. [ Links ]
Kokogian, D., Spalletti, L., Morel, E., Artabe, A., Martínez, R., Alcober, O., Milana, J., Zavattieri, A. and Papú, O. 1999. Los depósitos continentales triasicos. In: R. Caminos and J. Panza (eds.), Geología Regional Argentina, Instituto de Geología y Recursos Minerales, Anales 29, pp. 377- 398. [ Links ]
Komárek, J. 1983. Contribution to the chlorococcal algae of Cuba. Nowa Edwigia 37: 65-180. [ Links ]
Komárek, J. and Janovská, V. 2001. Review of the green algal genus Pediastrum ; implications for pollen-analytical research. Bibliotheca Phycologica , Band 108, 127 pp. [ Links ]
Kumaran, K.P.N. and Maheshwari, H.K. 1980. Upper Triassic sporae dispersae from the Tiki Formation. 2: miospores from the Janar Nala Section, South Rewa Gondwana Basin, India. Palaeontographica, B 173: 26-84. [ Links ]
Kützing, F.T. 1849. Species algarum. Brockhaus, Leipzing, 922 pp. [ Links ]
Leschik, G. 1955. Die Keuperflora von Neuewelt bei Basel. II. Die Iso- und Mikrosporen. Schweizerischen Paläontologischen Abrhandlungen 72: 5-70. [ Links ]
López Gamundí, O.R., Alvarez, L., Andreis, R., Bossi, G., Espejo, I., Fernandez-Seveso, F., Legarreta, L., Kokogian, D., Limarino, O. and Sesarego, H. 1989. Cuencas Intermontanas. In: G. Chebli and L. Spalletti (eds.), Cuencas Sedimentarias Argentinas, Serie Correlación Geológica 6, Instituto Superior de Correlación Geológica, Universidad Nacional de Tucumán. pp. 123-167. [ Links ]
Mädler, K. 1964. Die geologische Verbreitung von Sporen und Pollen in der Deutschen Trias. Beihefte zum Geologischen Jahrbuch 65, 147 p. [ Links ]
Maheshwari, H.K. and Banerji, J. 1975. Lower Triassic palynomorphs from the Maitur Formation, West Bengal. Palaeontographica B 152: 149-190. [ Links ]
Mancuso, A.C. 2000. Análisis tafonómico de los peces de la Formación Los Rastros (Triásico, La Rioja). Ameghiniana 37: 28R. [ Links ]
Mancuso, A.C. 2002. Vertebrate taphonomy in a Triassic lake: tetrapods of the Los Rastros Formation (Ischigualasto-Villa Unión Basin, central-western Argentina). 8º Congreso Argentino de Paleontología y Bioestratigrafía , (Corrientes), Actas : 35. [ Links ]
Mancuso, A.C. 2003. Continental fish taphonomy: a case study in the Triassic of Argentina. Journal of South American Earth Sciences 16: 275-286. [ Links ]
Marsicano, C.A., Arcucci, A., Mancuso, A.C. and Caselli, A. 2004. Middle Triassic tetrapod footprints of southern South America. Ameghiniana 41: 171-184. [ Links ]
Melchor, R.N., Genise, J.F. and Poiré, D.G. 2001. Icnología de los depósitos continentales triásicos. In: A.E. Artabe, E.M. Morel and A.B. Zamuner (ed.), El Sistema Triásico en la Argentina, Fundación Museo de La Plata Francisco Pascasio Moreno, La Plata, pp. 185-198. [ Links ]
Meyen, F.J.F. 1829. Beobachtungen über einigie niedrige Algenformen. Nova Acta Leopoldina-Carolina Norimbergae 14: 768-778. [ Links ]
Milana, J.P. 1998. Anatomía de parasecuencias en un lago de rift y su relación con la generación de hidrocarburos, cuenca triásica de Ischigualasto, San Juan. Revista de la Asociación Geológica Argentina 53: 365-387. [ Links ]
Milana, J.P. and Alcober, O. 1994. Modelo tectosedimentario de la cuenca triásica de Ischigualasto (San Juan, Argentina). Revista de la Asociación Geológica Argentina 49: 217-235. [ Links ]
Morel, E.M., Artabe, A.E., Zavattieri, A.M. and Bonaparte, J.F. 2001. Cronología del Sistema Triásico. In: A.E. Artabe, E.M. Morel and A.B. Zamuner (ed.), El Sistema Triásico en la Argentina, Fundación Museo de La Plata Francisco Pascasio Moreno, La Plata, pp. 227-253. [ Links ]
Nilsson, T. 1958. Über das Vorkommen eines mesozoischen Sapropelgesteins in Schonen. Acta Universitatis Lundensis 53: 1-112. [ Links ]
Ottone, E.G., Alvarez, P.P., Benoit, S.V. 1992. Late Triassic plant microfossils from the Rancho de Lata Formation, Main Cordillera, Argentina. Micropaleontology 38: 261-278. [ Links ]
Ottone, E.G. and García, G.B. 1991. A Lower Triassic miospore assemblage from the Puesto Viejo Formation, Argentina. Review of Palaeobotany and Palynology 68: 217-232. [ Links ]
Ottone, E.G. and Rodríguez Amenabar, C. 2001. A new disaccate pollen grain from the Triassic of Argentina. Ameghiniana 38: 157-161. [ Links ]
Pierce, R.L. 1961. Lower Upper Cretaceous plant microfossils from Minnesota. Minnesota Geological Survey Bulletin 42, 86 pp. [ Links ]
Playford, G. 1965. Plant microfossils from Triassic sediments near Poatina, Tasmania. Journal of the Geological Society of Australia 12: 173-210. [ Links ]
Playford, G. and Dettmann, M.E. 1965. Rhaeto-Liassic microfossils from the Leigh Creek Coal Measures, South Australia. Senckenbergiana Lethaea 46: 127-181. [ Links ]
Playford, G., Rigby, J.F. and Archibald, D.C. 1982. A Middle Triassic flora from Moolayember Formation, Bowen Basin, Queensland. Geological Survey of Queensland, Publication 380: 1-52. [ Links ]
Pocock, S.A.J. 1964. Pollen and spores of the Chlamydospermidae and Schizaeaceae from Upper Mannville strata of Saskatoon area of Saskatchewan. Grana Palynologica 5: 129-209. [ Links ]
Pocknall, D.F. 1981. Pollen morphology of the New Zealand species of Dacrydium Solander, Podocarpus L'Heritier, and Dacrycarpus Endlicher (Podocarpaceae). New Zealand Journal of Botany 19: 67-95. [ Links ]
Potonié, R. 1956. Synopsis der Gattungen der Sporae dispersae. I Teil: Sporites. Geologischen Jahrbuch 23, 103 pp. [ Links ]
Potonié, R. 1970. Synopsis der Gattungen der Sporae dispersae , V Teil. Nachträge zu allen Gruppen (Turmae). Geologischen Jahrbuch 87, 222 p. [ Links ]
Potonié, R. and Klaus, W. 1954. Einige Sporengattungen des alpinen Salzgebirges. Geologische Jahrbuch 68: 517-546. [ Links ]
Reiser, R.F. and Williams, A.J. 1969. Palynology of the Lower Jurassic sediments of the northern Surat Basin. Geological Survey of Queensland, Publication 399. Palaeontological Paper 15: 1-24. [ Links ]
Reissinger, A. 1950. Die Pollenanalyse ausgedehnt auf alle Sedimentgesteine der geologischen Vergangenheit. Palaeontographica B 90: 99-126. [ Links ]
Rodríguez Amenábar, C. and Ottone, E.G. 2002. Botryococcus como indicador paleoambiental en el Triásico de Argentina. 15º Congreso Geológico Argentino , (El Calafate), Actas 1: 493-494. [ Links ]
Rodríguez Amenábar, C. and Ottone, E.G. 2003. La aplicación de Botryococcus (Chlorococcales) como indicador paleoambiental en el Triásico de Argentina. Revista Española de Micropaleontología 35: 25-35. [ Links ]
Rogers, R.R., Arcucci, A.B., Abdala, F., Sereno, P.C., Forster, C.A. and May, C.L. 2001. Paleoenvironment and taphonomy of the Chañares Formation tetrapod assemblage (Middle Triassic), northwestern Argentina: spectacular preservation in volcanogenic concretions. Palaios 16: 461-481. [ Links ]
Rogers, R.R., Swisher, C.C. III, Sereno, P.C., Monetta, A.M., Forster, C.A. and Martínez, R.N. 1993. The Ischigualasto tetrapod assemblage (Late Triassic, Argentina) and 40 Ar/ 39 Ar dating of dinosaur origins. Science 260: 794-797. [ Links ]
Romer, A.S. and Jensen, J.A. 1966. The Chañares (Argentina) triassic reptile fauna. II Sketch of the geology of the Rio Chañares-Rio Gualo region. Breviora 252: 1-20. [ Links ]
Spalletti, L.A., Artabe, A., Morel, E. and Brea, M. 1999. Biozonación paleoflorística y cronoestratigrafía del Triásico Argentino. Ameghiniana 36: 419- 451. [ Links ]
Steeves, M.W. and Barghoorn, E.S. 1959. The pollen of Ephedra . Journal of the Arnold Arboretum 40: 221-254. [ Links ]
Stipanicic, P. N. 1983. The Triassic of Argentina and Chile. In: M. Moullade and A. E. M. Nairn (eds.), The Phanerozoic Geology of the world. The Mesozoic , Elsevier Scientific Publications, Amsterdam, pp. 181- 199. [ Links ]
Stipanicic, P. and Bonaparte, J. 1979. Cuenca triásica de Ischigualasto-Villa Unión (Provincia de La Rioja y San Juan) . In: J.C.M. Turner (coord.), Geología Regional Argentina, Academia Nacional de Ciencias (Córdoba), 1: 523- 575. [ Links ]
Stipanicic, P.N. and Bonetti, M.I.R. 1969. Consideraciones sobre la cronología de los terrenos triásicos argentinos. In: 1º IUGS Symposium-Gondwana Stratigraphy , (Mar del Plata), Earth sciences 2: 1081-1120. [ Links ]
Stipanicic, P.N. and Bossi, G. 2002a. Formación Ischigualasto. In: P.N. Stipanicic and C.A. Marsicano (ed.), Léxico Estratigráfico de la Argentina. Volumen VIII. Triásico. Asociación Geológica Argentina, Buenos Aires, Serie B (Didáctica y Complementaria) 26, pp. 149-151. [ Links ]
Stipanicic, P.N. and Bossi, G. 2002b. Formación Los Rastros. In: P.N. Stipanicic and C.A. Marsicano (ed.), Léxico Estratigráfico de la Argentina. Volumen VIII. Triásico. Asociación Geológica Argentina, Buenos Aires, Serie B (Didáctica y Complementaria) 26, pp. 184-185. [ Links ]
Tripathi, A., Tiwari, R.S. and Kumar, P. 1990. Palynology of the subsurface Mesozoic sediments in Rajmahal Basin, Bihar. The Palaeobotanist 37: 367-388. [ Links ]
Tyson, R.V. 1995. Sedimentary Organic Matter. Organic facies and palynofacies. Chapman and Hall (eds.), London, 615 p. [ Links ]
Uliana, M. A. and Biddle, K. 1988. Mesozoic-Cenozoic paleogeographic and geodynamic evolution of southern South America. Revista Brasileira de Geociências 18: 172-190. [ Links ]
Vail, P.R., Audermard, F., Bowman, S.A., Eisner, P.N. and Perez Cruz, C. 1991. The stratigraphic signatures of tectonics, eustacy and sedimentology- an overview. In: G. Einsele, G. Ricken and A. Seilacher (eds.), Cycles and events in stratigraphy, Springer-Berlag, Berlin. pp. 617-659. [ Links ]
Vannier, J., Thiéry, A. and Racheboeuf, P.R. 2003. Spinicaudatans and ostracods (Crustacea) from the Montceau Lagerstätte (Late Carboniferous, France): morphology and palaeoenvironmental significance. Palaeontology 46: 999-1030. [ Links ]
Vishnu-Mittre 1957. Abnormal pollen grains in some Indian gymnosperms with remarks on the significance of the abnormalities. The Journal of the Indian Botanical Society 36: 548-563. [ Links ]
Volkheimer, W. and Zavattieri, A.M. 1991. Aratrisporites compositus n. sp., a guide-fossil from the Triassic Cuyo Basin, western Argentina. Neues Jahrbuch für Geologie und Paläontologie, Monastshefte 9, 1991: 564-578. [ Links ]
Von Huene, F. 1931. Die fossilien Fährten im Rhät von Ischigualasto in Nordwest-Argentinien. Palaeobiologica 4: 99-112. [ Links ]
Warren, J.K. 1986. Shallow-water evaporitic environments and their source rock potential. Journal of Sedimentary Petrology 56: 442-454. [ Links ]
Wetzel, R.G., 1981. Limnology . W. B. Saunders Co., Philadelphia, 679 p. [ Links ]
Wille, W. 1970. Plaesiodictyon mosellanum n.g., n.sp., eine mehrzellige Grünalge aus dem Unteren Keuper von Luxembourg. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1970.5: 283-310. [ Links ]
Wilson, R.L. and Webster, R.M. 1946. Plant microfossils from a Fort Union Coal of Montana. American Journal of Botany 33: 271-278. [ Links ]
Wood, G.D. and Benson, D.G. Jr. 2000. The North American occurrence of the algal coenobium Plaesiodictyon : paleogeographic, paleoecologic, and biostratigraphic importance in the Triassic. Palynology 24: 9-20. [ Links ]
Yrigoyen, M.R. and Stover, L.E. 1970. La palinología como elemento de correlación del Triásico en la Cuenca Cuyana. 4° Jornadas Geológicas Argentinas , (Mendoza), Actas 2: 427-447. [ Links ]
Zamuner, A.B., Zavattieri, A.M., Artabe, A.E. and Morel, E.M. 2001. Paleobotánica. In: A.E. Artabe, E.M. Morel and A.B. Zamuner (ed.), El Sistema Triásico en la Argentina, Fundación Museo de La Plata Francisco Pascasio Moreno, La Plata, pp. 143-184. [ Links ]
Zavattieri, A.M. 1986. Estudio palinológico de la Formación Potrerillos (Triásico) en su localidad tipo, Cuenca Cuyana (provincia de Mendoza, Argentina). Parte I. Esporas triletes y monoletes. Revista Española de Micropaleontología 28: 247-294. [ Links ]
Zavattieri, A.M. 1987. Estudio palinológico de la Formación Potrerillos (Triasico) en su localidad tipo, Cuenca Cuyana (provincia de Mendoza, Argentina). Parte II. Granos de polen. Aspectos estadísticos. Correlación palinoestratigráfica. Revista Española de Micropaleontología 19: 173-213. [ Links ]
Zavattieri, A.M. 1991a. Granos de polen de la Formación Las Cabras (Triásico), en su localidad tipo, provincia de Mendoza, Argentina. Parte 1. Ameghiniana 28: 3-29. [ Links ]
Zavattieri, A.M. 1991b. Granos de polen de la Formación Las Cabras (Triásico), en su localidad tipo, provincia de Mendoza, Argentina. Parte 2. Ameghiniana 28: 205-224. [ Links ]
Zavattieri, A.M. 1992. Palinología de la Formación El Tranquilo (Triásico), provincia de Santa Cruz, Argentina. Ameghiniana 29: 305-314. [ Links ]
Zavattieri, A.M. and Batten, D.J. 1996. Miospores from Argentinian Triassic deposits and their potential for intercontinental correlation. In: J. Jansonius and D.C. McGregor (eds.), Palynology: principles and applications, American Association of Stratigraphic Palynologists Foundation, 2: 767-778. [ Links ]
Zavattieri, A.M. and Melchor, R.N. 1999. Estudio palinológico preliminar de la Formación Ischichuca (Triásico), en su localidad tipo (quebrada de Ischichuca Chica), provincia de La Rioja, Argentina. Asociación Paleontológica Argentina, Publicación Especial 6: 33-38. [ Links ]
Zavattieri, A.M. and Milana, J.P. 2000. Zonación palinológica informal para la Formación Ischigualasto (Neotriásico) en la provincia de San Juan, Argentina. 9º Simposio Argentino de Paleobotánica y Palinología, (Tucumán), Resúmenes : 119. [ Links ]
Recibido: 3 de abril de 2003.
Aceptado: 19 de agosto de 2004.

