










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.42 no.2 Buenos Aires Mar./June 2005
Las tafofloras triásicas de la región de los Lagos, Xma Región, Chile
Rafael Herbst 1 , Alejandro Troncoso 2 y Jorge Muñoz 3
1 Instituto Superior de Geología; Consejo Nacional de Investigaciones Científicas y Técnicas, Las Piedras 201 7°/B, T4000BRE S.M. de Tucumán, Argentina. rafaherbst@uolsinectis.com.ar
2 Departamento Biología Vegetal, Universidad de Talca, Casilla de Correo 747, Talca, Chile. aletronc@pehuenche.utalca.cl
3 Servicio Nacional de Geología y Minería, Delegación Sur, La Paz 406, Puerto Varas, Chile. sernageomin@surnet.cl
Resumen. Se da a conocer la composición florística y la descripción de algunas especies de tres tafofloras de la región de los Lagos del sur de Chile, provenientes de las localidades de Licán Ray, Punta Peters y cerro Tralcán (72°15' S - 39°30'/39°45' O), que forman parte de las Formaciones Panguipulli, las dos primeras, y Tralcán, la última. La flora se compone de 27 especies incluidas en los géneros: Hepatica indet., Neocalamites , Asterotheca , Cladophlebis , Gleichenites , Dicroidium , Johnstonia , Lepidopteris , Pterophyllum , Pseudoctenis , Sphenobaiera , Ginkgoites , Phoenicopsis , Rissikia , Heidiphyllum , Gen. et sp. indet., Linguifolium y Taeniopteris ; se describen una nueva especie de Astrerotheca y dos nuevas especies de Pterophyllum . Se señala la composición cuantitativa en las tres localidades y se establece que las asociaciones tafonómicas son notoriamente diferentes, a pesar de ser, cronológica y geográficamente, muy cercanas, concluyéndose que tales diferencias obedecen, principalmente, a razones paleoambientales.
Abstract. The Triassic taphofloras from the Lake district, xth Region, Chile . A list of the fossil plants, in some cases with their description, from the Panguipulli and Tralcán Formations, from the localities Licán Ray, Punta Peters and Cerro Tralcán, from the Lake District (72º15' S and 39º30'/39º45' W), Xth Region, Chile, is presented. The flora is composed of 27 species of the following genera: Hepatica indet., Neocalamites , Asterotheca , Cladophlebis , Gleichenites , Dicroidium , Johnstonia , Lepidopteris , Pterophyllum , Pseudoctenis , Sphenobaiera , Ginkgoites , Phoenicopsis , Rissikia , Heidiphyllum , Gen. et sp. indet., Linguifolium and Taeniopteris ; a new species of Astrerotheca and two new species of Pterophyllum are also described. The quantitative composition of the three localities is analyzed showing that they are quite different, in spite of being of similar age and geographically close to each other; it is suggested that the difference is basically paleoenvironmental.
Key words. Fossil flora; Impressions; Upper Triassic; Southern Chile.
Palabras clave. Tafoflora; Impresiones; Triásico Superior; Chile meridional.
Introducción
Son escasos los trabajos en los que se brindan datos sobre plantas fósiles del Triásico del sur de Chile. En general, éstas aparecen como menciones marginales y en prácticamente ningún caso existen descripciones y/o ilustraciones que permitan justificar los pocos y precarios nombres asignados. De las dos formaciones que nos interesan en este trabajo, se puede mencionar que Tavera (1971) citó Lepidodendron peruvianum (Gothan) para la Formación Panguipulli; posteriormente Minato (1977) señaló, para la ribera del lago Panguipulli, la presencia de Glossopteris sp. y Neuropteris sp. Las determinaciones de ambos autores resultan ser erróneas, y eso los llevó a atribuir los sedimentos portadores al Carbonífero Superior-Pérmico. La observación del material ilustrado por Minato (1977), permite sugerir que se trata de un ejemplar de Dicroidium (señalado como Neuropteris ) y uno de Linguifolium (señalado como Glossopteris ). Arrondo et al. (1988), en una breve contribución, reconocieron en el lago Panguipulli Dicroidium zuberi var. zuberi (Szajnocha) Archangelsky, Ginkgoites sp. y Neocalamites sp., mientras que en la ribera norte del lago Calafquén identificaron Dicroidium lancifolium var. lancifolium (Morris) Retallack y Dicroidium zuberi cf. var. sahnii (Seward) Retallack que, en conjunto, les sugieren el intervalo Triásico Medio a Superior como edad para los estratos portadores. De la Formación Tralcán, Davis y Karzulovic (1961) mencionaron una forma parecida a Cladophlebis tenue Oishi, mientras que Tavera (1971) citó Pterophyllum sp. Solms-Laubach, Equisetites fertilis Frenguelli, Neocalamites carresu (probablemente por carrerei (Zeiller) Halle) y Chiropteris copiapensis Solms-Laubach, provenientes del sector central de la secuencia, probablemente de niveles estratigráficamente comparables con aquéllos de los cuales provienen las formas descritas en este artículo. Sobre la base de tales antecedentes, estos autores le asignaron una edad triásica superior.
En la presente contribución se describen, a veces con otros nombres, algunas de las mismas plantas citadas y se agregan numerosos géneros y especies, tres de estas últimas nuevas para la ciencia. Para el caso de las especies muy conocidas se han obviado las descripciones y solamente se brindan algunos comentarios y/o comparaciones.
El material fósil incluye, además, diversas estructuras reproductivas (algunas de las cuales podrían ser asignadas, tentativamente, al género Umkomasia Thomas), semillas de 2-3 tipos y brácteas, cuya presencia sólo es consignada aquí, y que serán objeto de otro artículo.
Dado que en las tres localidades visitadas las asociaciones tafonómicas aparecían bastante distintas entre sí, hemos intentado un elemental análisis de su composición cuantitativa que, no es exacto ni preciso sino meramente indicativo.
Marco geológico
Formación Panguipulli
Las rocas sedimentarias triásicas que afloran en las riberas de los lagos Panguipulli, Riñihue y Calafquén han sido agrupadas en la Formación Panguipulli (Aguirre y Levi, 1964) (figura 1). Comprende sedimentitas con metamorfismo de bajo grado intruidas por plutones del Jurásico, Cretácico y Mioceno (Parada y Munizaga, 1978; Munizaga et al. , 1988; Rodríguez et al. , 1999). La Formación Panguipulli está localmente cubierta por depósitos fluviales, deltaicos, abanicos aluviales y coluviales del Holoceno y por diamictos glaciares (morrenas) y materiales glaciofluviales del Pleistoceno, representativos de las dos últimas glaciaciones que afectaron la región. Además, los afloramientos más orientales están, en parte, cubiertos por rocas volcánicas cuaternarias y por rocas volcánicas y sedimentarias, continentales, representativas de los Estratos de Lago Ranco, de edad oligocena-miocena (Campos et al. , 1998). Las primeras incluyen lavas, depósitos piroclásticos y lahares que se asocian a los volcanes Villarrica, Sierra de Quinchilca y Complejo Volcánico Mocho-Choshuenco. Las segundas corresponden a areniscas, lutitas, conglomerados, lavas y brechas volcánicas, tobas y tufitas de composición andesítica. Las rocas del Oligoceno-Mioceno se encuentran plegadas y en ellas se han emplazado rocas intrusivas del Mioceno
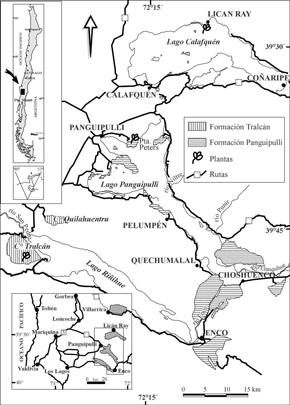
Figura 1. Mapa de ubicación / Location map.
La Formación Panguipulli representa una secuencia rítmica, en parte turbidítica, que tiene una potencia estimada de más de 800 m, compuesta por areniscas cuarcíferas y micáceas, lutitas y conglomerados cuarcíferos, fuertemente plegados y con manteos hasta verticales. Su base no está expuesta. Las columnas estratigráficas parciales de la Formación, levantadas en la ribera norte del Lago Panguipulli (Punta Peters) y en la ribera norte del Lago Calafquén, en las cercanías de la localidad de Licán Ray, se representan en la figura 2. En Punta Peters las capas son verticales, la secuencia expuesta alcanza cerca de 30 m de potencia y está constituida principalmente por conglomerados, desde gruesos a finos, con intercalaciones de areniscas y areniscas conglomerádicas. Los conglomerados se organizan en capas de hasta 5 m de potencia, en casos granodecrecientes o alternando con areniscas gruesas, en capas centimétricas, tanto matriz como clasto soportados, formados por fragmentos de cuarzo y de esquistos, con matriz arenosa de granos de cuarzo, esquistos y muscovita detrítica. Las areniscas son finas a gruesas, localmente lenticulares o conglomerádicas, constituidas por fragmentos de cuarzo, esquistos y muscovita detrítica. La flora fósil se encuentra en tres niveles de lutitas, separados por capas centimétricas de areniscas finas, ubicados en la parte central de la columna (figura 2). En la ribera norte del lago Calafquén, la secuencia expuesta localmente alcanza más de 200 m de potencia, con capas métricas a centimétricas de rumbo N-S a N70O y manteo entre 30 y 60º al este. La secuencia es de carácter rítmico, localmente turbidítico, con alternancia de bancos de areniscas finas a medias y lutitas, métricas a centimétricas, con algunos niveles de conglomerados finos. Las areniscas son de color gris a negro, de granulometría fina a gruesa, en partes conglomerádicas o bandeadas. Las lutitas son negras y en partes alternan con areniscas finas. Los conglomerados son finos, cuarcíferos, y localmente alternan con areniscas bandeadas. La flora fósil, en muchos casos con fragmentos indeterminables, se encuentra en diferentes niveles de la columna, siempre en capas de lutitas. Según Rodríguez et al. (1999) la secuencia completa es de carácter turbíditico-lacustre. Las alternancias rítmicas de lutitas y areniscas representarían facies de abanico subacuático, inferior o distal, en zonas no canalizadas, mientras que las facies de conglomerados corresponderían a facies de canales y pie de talud.

Figura 2. Columnas estratigráficas de Lican Ray y Punta Peters/ Stratigraphic columns of Lican Ray and Punta Peters.
Una edad K-Ar de 291±7 Ma en muscovitas detríticas de una arenisca de la ribera noroccidental del lago Panguipulli (Punta Peters) y, en esa misma localidad, una edad U-Pb de 230 Ma en circones detríticos fueron mencionadas por Rodríguez et al. (1999). La primera edad sería indicativa de un evento de metamorfismo que afectó a las rocas fuentes de los sedimentos de la Formación Panguipulli, probablemente el Complejo Metamórfico Trafún (Illies, 1970; Duhart et al. , 2001), mientras que la segunda señala una edad máxima triásica media para la depositación de los sedimentos de la misma formación.
Formación Tralcán
Esta unidad fue definida en el cerro del mismo nombre y también se expone en el cerro Quilahuentrú, al norte del cerro Tralcán (figura 1). Corresponde a una secuencia sedimentaria compuesta por conglomerados gruesos, localmente de color rojo, con intercalaciones de areniscas, con muscovita detrítica, y fangolitas de colores blancos, gris y amarillo, estas últimas en algunos casos, con flora fósil. La secuencia tiene una actitud monoclinal de suave manteo al sur y espesor mínimo de 800 m; su base se apoya sobre rocas metamórficas del Paleozoico (Complejo Metamórfico Trafún) (Illies, 1970; Rodríguez et al. , 1999) y su techo es la actual superficie de erosión. Lo limitado de los afloramientos no permite levantar una columna estratigráfica relativamente confiable. Según Rodríguez et al. (1999) los conglomerados son gruesos a finos y polimícticos, de hasta 3 m de potencia, clasto soportados y de matriz arenosa, con fragmentos, redondeados y localmente imbricados, de metapelitas, metareniscas y cuarzo provenientes de la erosión de rocas del Complejo Metamórfico Trafún. Estos autores sugieren que las facies de conglomerados de la secuencia representan ambientes fluviales de ríos trenzados, cuya depositación se asocia a corrientes proximales durante estadios de alta carga de sedimentos en el río. Las facies de areniscas y fangolitas representarían depositación a partir de corrientes trenzadas.
Aunque litológicamente distintas, la cercanía geográfica y la estructura geológica regional permiten suponer que estas dos Formaciones (Panguipulli y Tralcán) constituyen un complejo deposicional fluvio-lacustre. Ambas unidades se correlacionan cronoestratigráficamente y, en parte, litológicamente, con los Estratos de Pocillas (Moreno et al. , 1976; Hervé et al. , 1976), Estratos de Santa Juana-Quilacoya (Tavera, 1960; Cucurella, 1978) y con la Formación Llafquentué-Huimpil (Hauser, 1970; Parada y Moreno, 1980).
Material y métodos
Los materiales descriptos están todos preservados como impresiones y no existe materia orgánica preservada, como cutículas. De una sola muestra de la Fm. Panguipulli (marcada en la figura 2) se ha obtenido una asociación polínica, con más de 30 taxones diferentes (Zavattieri et al ., 2003).
Los materiales se integran a la colección del Museo Nacional de Historia Natural (Santiago), cuyo acrónimo es Sgo-Pb.
Descripciones sistemáticas
Hepática indeterminada
Material estudiado. Cerro Tralcán: SGO Pb 1876b, 1879c, 1899.
Descripción. Talo heterotricho, constituido por una especie de rizoma del cual emergen segmentos aplanados, cintiformes, 3 o más veces dicótomos, de 3-6 cm de largo. Cada segmento diferenciado en un cordón central, como vena media, y expansiones membranosas, de 1,5-2,5 mm de ancho, a cada lado.
Comentarios. La singular morfología, así como la posible presencia de estructuras reproductivas, nos han obligado a un estudio más exhaustivo y particular de esta forma, el cual aún se halla en curso.
Orden EQUISETALES Tryon Y Tryon 1982
Familia APOCALAMITACEAE Radczenko 1957
Género Neocalamites Halle 1908
Especie tipo. Neocalamites hoerensis (Schimper) Halle 1908.
Neocalamites sp.
Material estudiado. Licán Ray : Sgo-Pb 1784b, 1786a, 1802b, 1808, 1810, 1812b, 1836b, 1852. Punta Peters: Sgo-Pb 1713, 1714, 1715.
Comentarios. Este género es muy frecuente en todo el Triásico gondwánico, aunque las diversas especies que se le atribuyen deben ser re-estudiadas con detalle. Para la región sudamericana, se suelen incorporar los ejemplares de distintos yacimientos, ya sea a N. carrerei (Zeiller) Halle o bien a la nórdica N. Hoerensis Schimper. Frenguelli (1944b) describió dos especies, N. ischigualastii y N. ramaccionii , que no están muy bien diferenciadas entre sí y que se parecen mucho a los ejemplares adjudicados a N. carrerei . Debido a la escasa cantidad y estado fragmentario de los ejemplares disponibles, optamos por una nomenclatura abierta. En todo caso, este género es un buen indicador paleoambiental aunque no cronológico.
Orden MARATTIALES Tryon y Tryon 1982
Familia ASTEROTHECACEAE sensu Engler 1954
Género Asterotheca Presl in Corda 1845
Especie tipo. Asterotheca sternbergii (Goeppert) Presl, in Corda 1845.
Asterotheca frassinettii Herbst y Troncoso nov. sp.
Figuras 3.E-F, 9.F-G
Holotipo. Sgo-Pb 1876. Localidad: cerro Tralcán, cercanías del lago Riñihue, X Región, Chile. Edad: Triásico Superior.
Paratipos. Sgo-Pb 1920a, b de la misma localidad y edad.
Material estudiado. Cerro Tralcán: Sgo-Pb 1855c, 1857a, 1858a, 1859a, 1861a, 1862a, 1866, 1867a, 1883a, 1889a, 1897, 1925d.

Figura 3. A, Cladophlebis sp. A, Sgo-Pb 1728, Punta Peters; B-C , Gleichenites gallegoi Herbst, B, Sgo-Pb 1836, C, Sgo-Pb 1842, Licán Ray; D, Gleichenites cachivaritensis Herbst, Sgo-PB 1727, Punta Peters; E-F, Asterotheca frassinettii Herbst y Troncoso nov. sp., Sgo-Pb 1876, cerro Tralcán; Escala = 1 cm / Scale bar =1 cm.
Diagnosis. Frondes de forma y tamaño desconocidos; fragmentos de pinna de hasta 7 cm de longitud y 1,5 cm de anchura; raquis de 1 mm de ancho. Pínnulas de unos 7-8 mm de longitud y 1,5 - 1,8 mm de anchura, separadas 1 a 1,5 mm entre sí, insertas por toda la base, la que es ligeramente ensanchada, falcadas, dispuestas a unos 70-80°; los márgenes son paralelos hasta el ápice, que es obtuso-redondeado; vena media presente, la venación lateral no ha sido observada. Sinangios de contorno circular a algo cuadrangular, tetra- a veces pentaesporangiados, de 0,8-0,9 mm de diámetro, en número de 6-8, cubriendo por entero cada hemilámina pinnular.
Diagnosis. Fronds of unknown shape and size. Pinna fragments up to 7 cm long and 1.5 cm wide, rachis 1 mm wide. Pinnules 7-8 mm long and 1.5-1.8 mm wide, remote, separated 1-1.5 mm, inserted at 70-80° by a slightly widened base, falcate; margins parallel to an obtusely-rounded apex. Mid-vein present but lateral venation unknown. Sinangia rounded to somewhat cuadrangular, tetra- to pentasporangiate, 0.8-0.9 mm diameter, 6 to 8 at each side of mid-vein, covering the whole pinnule under surface.
Derivatio nominis. Dedicada al paleontólogo Daniel Frassinetti, por sus aportes a la paleontología de Chile.
Comentarios. Esta pequeña especie de Asterotheca no tiene semejanza cercana con ninguna de las descriptas para el ámbito del Gondwana sudamericano. De las especies argentinas, más cercanas geográficamente, se parece algo a A. hilariensis Menéndez (1957), pero difiere de ésta porque tiene las pínnulas bastante separadas y más angostas; otra especie gondwánica, también parecida, es A. rigbyana Herbst (1977), pero ésta tiene pínnulas contiguas de forma y tamaño diferente.
Incertae sedis
Género Cladophlebis Brongniart 1849
Especie tipo. Cladophlebis albertsii (Dunker) Brongniart 1849.
Cladophlebis kurtzi Frenguelli 1947
Material estudiado. Cerro Tralcán: Sgo-Pb 1854a, 1856, 1863, 1864a. Punta Peters: Sgo-Pb 1820 al 1824. Licán Ray: 1815, 1817.
Cladophlebis mesozoica (Kurzt) ex Frenguelli 1947
Material estudiado. Licán Ray: Sgo-Pb 1812a, 1813, 1814, 1816, 1825, 1847b.
Comentarios. En anteriores contribuciones (Herbst, 1971, 1988; Herbst et al., 1998; Troncoso y Herbst, 2000) se discutió la dificultad de separar especies morfológicamente tan cercanas entre sí como C. Kurtzi y C. mesozoica , habiendo apelado a diferentes modos de designarla. Seguiremos aquí el criterio establecido por Herbst et al. (1998), quienes señalan que ambas no deberían unirse y utilizar eventualmente el epíteto que más se acerque a las morfologías establecidas por Frenguelli (1947) para estas especies. Los ejemplares de cerro Tralcán y de Punta Peters, por su tamaño, forma de las pínnulas y su venación, se pueden incorporar a C. kurtzi , mientras que los de Licán Ray tienen todos los caracteres de C. mesozoica , aunque es muy probable que ambas sean porciones de frondas de una misma especie.
Cladophlebis sp. A
Figura 3.A
Material estudiado. Punta Peters: Sgo-Pb1728, 1729, 1730.
Descripción. Fragmentos de frondes de unos 4 cm de anchura; pínnulas pecopterídeas, insertas por todo el ancho de la base, separadas, dispuestas a 70-75° respecto al raquis; los márgenes son subparalelos, convergiendo al ápice que es agudo; el borde basiscópico es ligeramente decurrente. Vena media bien marcada y venación lateral simple, unas 14-16 venas rectas en cada lado, saliendo a unos 50-60° de la vena media.
Comentarios. Estos ejemplares son demasiado fragmentarios para asignarlos con seguridad a alguna especie conocida. En su amplia discusión sobre el género Cladophlebis , Frenguelli (1947) restringió los caracteres que debían tener las frondes para ser incluidas en éste. Si bien señala que es poco frecuente que las pínnulas tengan venación lateral simple, no las excluye. De las formas conocidas para esta parte del Gondwana, esta sería la primera especie con tal tipo de venación.
Orden FILICALES Tryon y Tryon 1982
Familia GLEICHENIACEAE (R.Br.) Presl 1825
Género Gleichenites Seward ( non Goeppert) 1926
Especie tipo. Gleichenites porsildi Seward (en Andrews, 1955).
Gleichenites cachivaritensis Herbst 1996
Figura 3.D
Material estudiado. Punta Peters: Sgo-Pb 1727.
Comentarios. El material disponible es bastante escaso, pero los caracteres morfológicos coinciden totalmente con los de la especie descripta de la Formación La Ternera de Chile (Herbst, 1996).
Gleichenites gallegoi Herbst 1996
Figuras 3.B-C
Material estudiado. Licán Ray: Sgo-Pb 1803a, 1832, 1833a, 1834, 1835a, 1836a, 1837, 1838.
Comentarios. Igual que en el caso anterior, no cabe ninguna duda sobre la asignación de los ejemplares disponibles a esta especie, que es característica del Triásico Superior de la Formación Cañadón Largo, en la provincia de Santa Cruz, Argentina.
Orden CORYSTOSPERMALES Sternberg 1820
Familia CORYSTOSPERMACEAE Thomas 1933
Género Dicroidium Gothan 1912
Especie tipo. Dicroidium odontopteroides (Morris) Gothan 1912.
Dicroidium odontopteroides (Morris) Gothan 1912
Figuras 4.A-F, 7.A-D

Figura 4. A-F, Dicroidium odontopteroides (Morris) Gothan, A, Sgo-Pb 1846, B, Sgo-Pb 1796, C, Sgo-Pb 1806, D, Sgo-Pb 1795, E, Sgo-Pb 1934, F, Sgo-Pb 1801; todos de Licán Ray/ all from Licán Ray . Escala= 1 cm/ Scale bar= 1cm.
Material estudiado. Licán Ray: Sgo-Pb 1783, 1784a, 1785a, 1791, 1792, 1793a, 1794a-b, 1795 al 1801, 1802a, 1804 al 1807, 1821b, 1824, 1828b, 1833b, 1835b, 1845a, 1846a, 1848a, 1849a, 1850a, 1851a.
Comentarios. El polimorfismo de esta especie, probablemente la más abundante en todo el Gondwana, ha sido señalado en varios trabajos anteriores (Gnaedinger y Herbst, 1998, 2001) en los que se discutió la utilización de rangos taxonómicos infraespecíficos. Dado que en muchos casos existe toda una serie de transiciones entre las formas que en la literatura fueron separadas como subespecies, variedades (Retallack, 1977) o paleodemes (Anderson y Anderson, 1989) se optó en aquellas contribuciones por no utilizar tal tipo de nomenclatura. En el caso presente, vuelve a darse esta transición de formas y tamaños, por lo que reiteramos esta postura. De este modo, puede señalarse que esta especie, con una diversidad de formas, de las que aquí se ilustran los casos extremos (figura 4), está presente en estas floras.
Dicroidium lancifolium (Morris) Gothan 1912
Figuras 6.A-B, 7.E-F
Material estudiado. Licán Ray: Sgo-Pb 1818, 1819, 1820a, 1821a, 1822, 1823.
Comentarios. Esta es una especie con una morfología muy característica y muy difícil de confundir con otra, por lo que no requiere comentarios adicionales.
Dicroidium spp.
Material estudiado. Punta Peters: Sgo-Pb 1725, 1726. Cerro Tralcán: 1906c.
Comentarios. En las localidades Punta Peters y cerro Tralcán se han hallado unos pocos fragmentos que son atribuibles a Dicroidium , pero que aparentemente no pertenecen a ninguna de las dos especies recién citadas. Los pocos caracteres visibles no permiten mayor precisión ni comparaciones.
Género Johnstonia Walkom 1917
Especie tipo. Johnstonia coriacea (Johnston) Walkom, 1917.
Johnstonia sp.
Material estudiado. Cerro Tralcán: Sgo-Pb 1854b.
Comentarios. Unos pocos fragmentos que pertenecen a este género fueron hallados en cerro Tralcán. A pesar de la imposibilidad de asignarlos a una especie concreta, en este trabajo se los cita simplemente para mostrar la diversidad de formas de Pteridospermas presentes en esta región.
Orden PELTASPERMALES Thomas 1933
Familia PELTASPERMACEAE Thomas 1933
Género Lepidopteris Schimper emend Townrow 1956
Especie tipo. Lepidopteris stuttgartiensis (Jaeger) Schimper, 1869.
Lepidopteris stormbergensis (Seward) Townrow 1956
Material estudiado. Punta Peters: Sgo-Pb 1700, 1701, 1702, 1703a.
Comentarios. Si bien la diagnosis emendada de Townrow (1956) implica la descripción de la cutícula, en numerosas oportunidades algunas de las especies fueron incluídas en el género Lepidopteris solamente sobre la base de impresiones. Este es el caso presente. La morfología de estas frondas es bastante característica y se diferencia de otras especies cercanas. Lepidopteris stormbergensis (Seward) Townrow está ampliamente difundida en el Gondwana, aunque con diversos nombres previos (Townrow, 1956). Para la Argentina, Frenguelli (1944a) la cita como Callipteridium ; Baldoni (1972) revisa los materiales del Triásico de este país, con y sin cutículas, y concluye con una buena definición de L. stormbergensis que diferencia de otras especies cercanas. Anderson y Anderson (1989) también describen e ilustran esta especie. En todos los casos hay coincidencia en la morfología por lo que se asigna como tal.
Orden CYCADOPHYTA sensu Strasburger 1981
Género Pterophyllum Brongniart 1828
Especie tipo. Pterophyllum longifolium Brongniart, 1828.
Los alcances del género se tratan con el mismo criterio que en Herbst y Troncoso (2000), esto es, en el sentido que originalmente le diera Brongniart (1828) y luego Arber (1907).
Pterophyllum valdivianum Herbst y Troncoso nov. sp.
Figuras 5.A-C, 8.C-H

Figura 5. A-C, Pterophyllum valdivianum Herbst y Troncoso nov. sp., A, Sgo-Pb 1915a, B, Sgo-Pb 1891, C, Sgo-Pb 1915b, todos de cerro Tralcán/ all from cerro Tralcán ; D-F, Pterophyllum tralcanensis Herbst y Troncoso nov. sp., D, Sgo-Pb 1898, E, Sgo-Pb 1854, F, Sgo-Pb 1898, todos de cerro Tralcán/ all from cerro Tralcán ; G, Pseudoctenis multilineata , (Shirley) Herbst y Troncoso nov. sp., Sgo-Pb 1735; H, Ginkgoites palmata (Ratte) Gnaedinger y Herbst, Sgo-Pb 1772, Punta Peters; I: Gen. et sp. indet., Sgo-Pb 1909, cerro Tralcán. Escala= 1 cm/ Scale bar= 1 cm.
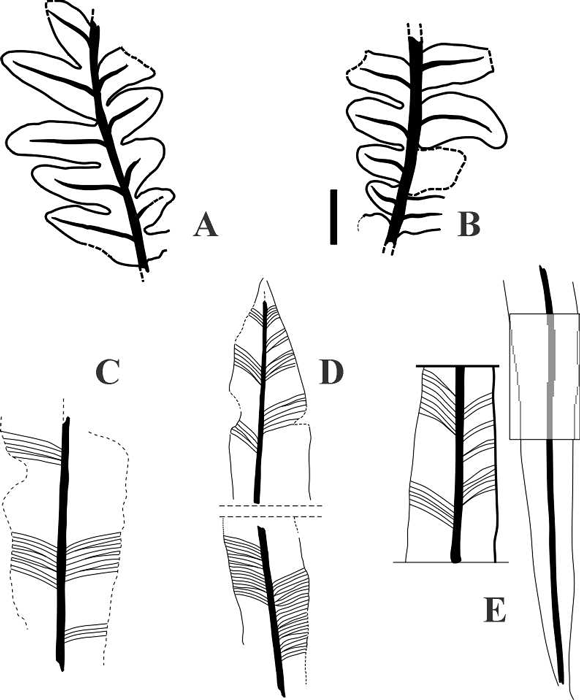
Figura 6. A-B, Dicroidium lancifolium (Morris) Gothan; A, Sgo-Pb 1819; B, Sgo-Pb 1822, Licán Ray; C-D, Taeniopteris parvilocus Anderson y Anderson; C, Sgo-Pb 1918; D, Sgo-Pb 1954, todos de cerro Tralcán/ all from cerro Tralcán ; E, Linguifolium cf. L. tenison-woodsii (Etheridge) Retallack, Sgo-Pb 1780, Punta Peters. Escala = 1 cm/ Scale bar = 1 cm.

Figura 7. A-D, Dicroidium odontopteroides (Morris) Gothan; A, Sgo-Pb 1795; B, Sgo-Pb 1803; C, Sgo-Pb 1801; D, Sgo-Pb 1796, todos de Licán Ray/ all from Lican Ray ; E-F, Dicroidium lancifolium (Morris) Gothan; E, Sgo-Pb 1819; F, Sgo-Pb 1822, todos de Licán Ray/ all from Licán Ray ; G, Linguifolium cf. L. tenison-woodsii (Etheridge) Retallack, Sgo-Pb 1780, Punta Peters; H, Linguifolium lillieanum Arber, Sgo-Pb 1830, Licán Ray; I-J, Gen. et sp. indet. Herbst y Troncoso; I, Sgo-Pb 1909; J, Sgo-Pb 1921, todos de cerro Tralcán/ all from cerro Tralcán . Escalas = 1 cm/ Scale bars = 1 cm.
Holotipo. Sgo-Pb 1915 a,b. Localidad cerro Tralcán, inmediaciones del lago Riñihue, Xma Región, Chile.
Paratipos. Sgo-Pb 1916a, b; 1891. Misma localidad y edad.
Material estudiado. Cerro Tralcán: Sgo-Pb 1853a, 1855d, 1858c, 1859c, 1861b, 1862b, 1864b, 1865a, 1867b, 1868d, 1869a, 1870c, 1872a, 1875, 1876, 1877b, 1878, 1880a, 1882, 1884, 1885a, 1887a, 1890a, 1892a-b, 1898a-b, 1901a, 1902a, 1903b, 1904b,d, 1906b, 1908b, 1909b, 1914b-c, 1915a, 1916a,d, 1920c, 1924, 1925a-c, 1926.
Comentarios. Véase P. tralcanensis.
Diagnosis. Hojas pinnadas de hasta 20 cm de longitud y 7 cm de anchura, de forma elíptica alargada; raquis estriado, de hasta 3 mm de anchura. Pinnas ligeramente falcadas, insertas a 70-80° por todo el ancho de la base, de bordes laterales enteros, paralelos y ápice redondeado; las más grandes miden 3,5-3,8 cm de longitud y 6 mm de anchura; pinnas basales y apicales de 1 cm de longitud y 3-4 mm de anchura. Las venas salen de todo el ancho de la base, son simples o muy ocasionalmente divididas una vez; densidad de unas 45-50 venas/cm.
Diagnosis. Pinnate leaves up to 20 cm long and 7 cm wide; rachis stout, striate, up to 3 mm wide. Pinnae slightly falcate, inserted at 70-80° by their whole width, lateral margins entire, parallel, apex rounded. Largest pinnae 3.5-3.8 cm long and 6 mm wide; basal and apical pinnae reduced to 1 cm long and 3-4 mm wide. Veins arising from the pinna base, simple or occasionally once divided; density about 45-50 veins/cm.
Edad. Triásico Superior
Derivatio nominis. Por la provincia de Valdivia, donde se localiza la Formación Tralcán.
Pterophyllum tralcanensis Herbst y Troncoso nov. sp.
Figuras 5.D-F, 8.A-B

Figura 8. A-B, Pterophyllum tralcanensis Herbst y Troncoso nov. sp.; A, Sgo-Pb 1898a; B, Sgo-Pb 1854, todos de cerro Tralcán/ all from cerro Tralcán ; C-H, Pterophyllum valdiviensis Herbst y Troncoso nov. sp.; C, Sgo-Pb 1872a; D, Sgo-Pb 1915a; E, Sgo-Pb 1915b; F, Sgo-Pb 1891, G, Sgo-Pb 1901; H, Sgo-Pb 1890b, todos de cerro Tralcán/ all from cerro Tralcán . Escalas = 1 cm/ Scale bars = 1 cm.
Holotipo. Sgo-Pb 1898. Localidad: cerro Tralcán, inmediaciones del lago Riñihue, Xma Región, Chile.
Paratipos. Sgo-Pb 1854e, 1872.
Material estudiado. Cerro Tralcán: Sgo-Pb 1854e, 1857b, 1858b, 1859b, 1871, 1872b, 1879a, 1891, 1893, 1898c-d, 1900c, 1911b, 1912b, 1915b, 1916b.
Diagnosis. Hojas pinnadas de hasta 7 cm de longitud y 2-2,5 cm de anchura, oblongas angostas; raquis del orden de 1,5 mm de anchura. Pinnas insertas a 75°-80° por toda su base, de bordes laterales paralelos, enteros, y ápice redondeado; miden 1 cm de longitud y 3 mm de anchura, las apicales y basales de 0,7-0,8 cm de longitud y 2,3 mm de anchura. Venas finas, salen desde toda la base, simples; densidad unas 36-40 venas/cm.
Diagnosis. Pinnate leaves, oblong, up to 7 cm long and 2-2.5 cm wide; rachis about 1.5 mm wide. Pinnae inserted at 75°-80° by their whole base, lateral margins entire, parallel and with rounded apex; pinnae up to 1 cm long and 3 mm wide, apicals and basals 0.7-0.8 cm long and 2.3 mm wide. Veins fine, arising from the whole extension of the base, simple; density about 36-40 veins/cm.
Edad. Triásico Superior.
Derivatio nominis. Por el cerro y la Formación Tralcán, de donde proviene la especie.
Comentarios. Estas dos nuevas especies se diferencian netamente, en sus caracteres morfológicos, de las restantes especies conocidas de la región y de ámbitos geográficos cercanos del Gondwana (Sudáfrica y Australia), de acuerdo con la bibliografía disponible. Ambas se diferencian entre sí por el tamaño de las pinnas (y consiguientemente de las hojas); la densidad de la venación no es muy diferente en ambas, pero, siendo las pinnas de P . tralcanensis bastante más pequeñas, la venación aparenta ser mucho más densa en éstas.
Género Pseudoctenis Seward 1911
Especie tipo. Pseudoctenis eathiensis (Richards) Seward 1911.
Pseudoctenis multilineata (Shirley) Herbst y Troncoso 2000
Figura 5.G
Material estudiado. Punta Peters: Sgo-Pb 1735 al 1750.
Comentarios. Los numerosos ejemplares disponibles coinciden con los descriptos para el Triásico de Chile (Herbst y Troncoso, 2000).
Orden GINKGOALES Engler 1897
Género Sphenobaiera Florin 1936
Especie tipo. Sphenobaiera spectabilis (Nathorst) Florin 1936.
Sphenobaiera calensis Anderson y Anderson 1989
Material estudiado. Licán Ray: Sgo-Pb 1847,1848, 1850, 1851.
Comentarios. No disponemos de ejemplares completos, sólo fragmentarios. Los restos conservados llegan a medir hasta 14 cm de longitud, la anchura máxima varía entre 0,5 y 1 cm. Los ejemplares son aparentemente indivisos, excepto dos que se bifurcan una vez, tal como los ilustrados por Anderson y Anderson (1989, lám. 96, figs. 6, 9, 12-13). Se cuentan 9-10 venas/lacinia. Como señalan Anderson y Anderson (1989) sólo la presencia de esporádicas bifurcaciones de las venas distinguen las láminas indivisas de las de Heidiphyllum .
Sphenobaiera sp.
Material estudiado. Punta Peters: Sgo-Pb 1767b, ? 1778a.
Comentarios. El ejemplar Sgo-Pb 1767b se halla mal preservado, carece de base y ápice. Su longitud inferida debió ser de 12 a 14 cm y anchura máxima alrededor de 4 a 5 cm. La lámina se divide, aparentemente, tres veces; sólo se han preservado 6 lacinias, las que se mantienen muy cerca una de otra, a lo menos hasta tres cuartos de la longitud; el ángulo formado por las lacinias más externas no sobrepasa los 20°. Las lacinias terminales miden 3-4 mm de anchura. Se cuentan alrededor de 8 venas por lacinia.
Estos ejemplares no se corresponden con ninguna de las especies descriptas; las formas más semejantes las encontramos en algunas variaciones de S. Schenkii (Feistmantel) Florin ilustradas por Anderson y Anderson (1989, lám. 98, fig. 16).
Género Ginkgoites Seward emend . Maheshwari y Bajpai 1992
Especie tipo. Ginkgoites obovata (Nathorst) Seward 1919.
Ginkgoites palmata (Ratte) Gnaedinger y Herbst 1999
Figuras 5.H, 9.E
Material estudiado. Punta Peters: Sgo-Pb 1771-1772, 1775-1777.
Comentarios. Sólo disponemos de ejemplares fragmentarios; algunos de ellos deben haber tenido una longitud total de más de 15 cm. Cada división mide, hacia la base, 0,8 cm. Se observan hasta 3 bifurcaciones, pero podría haber más. La densidad de venas es de 22-30 venas/cm.
En el ejemplar ilustrado por Shirley (1898) como Ginkgo simondsi la densidad de venación es similar a la de nuestros ejemplares. Walkom (1917: 10), por su parte, señala que la densidad de venación de su material es de 3 venas/mm, lo que resultaría en alrededor de 30 venas/cm. Anderson y Anderson (1989) dan un valor de 15 venas/cm, a pesar de ilustrar el ejemplar de Shirley. Gnaedinger y Herbst (1999) encuentran valores de 10 venas/cm en la porción distal.
Ginkgoites cf. matatiensis Anderson y Anderson 1989
Figura 9.D

Figura 9. A-B, Taeniopteris parvilocus Anderson y Anderson, A, Sgo-Pb 1918; B, Sgo-Pb 1853d, todos de cerro Tralcán/ all from cerro Tralcán ; C, Heidiphyllum elongatum (Morris) Retallack, Sgo-Pb 1710, Punta Peters; D, Ginkgoites cf. matatiensis Anderson y Anderson, Sgo-Pb 1896, cerro Tralcán; E, Ginkgoites palmata (Ratte) Gnaedinger y Herbst, Sgo-Pb 1772, Punta Peters; F-G, Asterotheca frassinettii Herbst y Troncoso, Sgo-Pb 1876, cerro Tralcán. Escalas = 1 cm/ Scale bars = 1 cm.
Material estudiado. Licán Ray: 1843. Cerro Tralcán: 1896.
Comentario. Disponemos sólo de dos fragmentos basales de lámina, que muestran división en lóbulos y venas bifurcadas. Las venas tienen una densidad de 10-12 venas/cm. Por esta razón y por la comparación con el espécimen ilustrado por Anderson y Anderson (1989: lám. 125, fig. 7) se los asigna tentativamente a esta especie.
Género Phoenicopsis Heer 1876
Especie tipo. Phoenicopsis angustifolia Heer 1876.
cf. Phoenicopsis sp.
Material estudiado. Punta Peters: Sgo-Pb 1752, 1754-1757, 1759-1760. Lican Ray: Sgo-Pb 1993 (cf.). Cerro Tralcán : 1860, 1874, 1994 (todos cf. ).
Descripción. Hojas lineares, angostas, de longitud desconocida, mayor a 9 cm, y de 2-4 cm de anchura, la cual disminuye hacia la base; recorridas por 4 (a veces 5) venas longitudinales, paralelas. Estas hojas se hallan dispuestas, muy imbricadas entre sí, sobre un corto braquiblasto; en nuestro ejemplar se ha conservado un número de 4, pero seguramente había más. El braquiblasto presenta cicatrices que presumiblemente corresponden a bases foliares de hojas desprendidas. El braquiblasto y las hojas se han desprendido como una unidad.
Comentario. Con las características señaladas sólo disponemos de un ejemplar (Sgo-Pb 1757); en cambio, son notablemente abundantes en las muestras, trozos de la parte media de la lámina de estas hojas, en las que no se ha preservado la conexión al braquiblasto.
El género Phoenicopsis fue instituido por Heer (1876: 49) para hojas coriáceas, sésiles o con un breve pecíolo, indivisas, multivenadas, con venas finas, paralelas y densas, en fascículos sobre cortos braquiblastos caducos. Florin (1936) separó géneros de morfología similar sobre la base de caracteres cuticulares, dejando válido el género de Heer para el resto de los materiales. La única cita del género Phoenicopsis para América del Sur es la de P. elongata (Morris) Seward (Jain y Delevoryas, 1967), especie que fue transferida a Heidiphyllum por Retallack (1981). Ya Seward (1926) había incluido con dudas esta especie en Phoenicopsis , toda vez que los materiales originales correponden a hojas dispersas. Los ejemplares de Jain y Delevoryas (1967), de la Formación Potrerillos, de Mendoza (Argentina), que tampoco muestran conexión a braquiblasto ni disposición en fascículos, a su vez, fueron referidos a Desmiophyllum cacheutense (Kurtz) Troncoso, Gnaedinger y Herbst (= Heidiphyllum cacheutense (Kurtz) en Anderson y Anderson, 1989) por estos autores (Troncoso et al., 2000). Los ejemplares aquí descriptos difieren de las otras especies del género por el escaso número de venas presentes en cada hoja.
Orden CONIFERALES Engler 1897
Familia PODOCARPACEAE Endlicher 1847
Género Rissikia Townrow 1967
Especie tipo. Rissikia media (Tenison-Woods) Townrow 1967.
Rissikia media (Tenison-Woods) Townrow 1967
Material estudiado. Punta Peters: Sgo-Pb 1704, 1719.
Comentarios. Los ejemplares disponibles corresponden a porciones pequeñas de ramas con hojas y son asignables sin duda a esta especie. Bajo este nombre se ha incorporado una variedad de formas, ilustradas por ejemplo en Anderson y Anderson (1989). Son muy semejantes a los ilustrados por estos autores en la lámina 269, figura 23; lámina 265, figuras 17 y 24-26, y a los de la lámina 270.
Familia VOLTZIACEAE Arnold 1947
Género Heidiphyllum Retallack 1981
Especie tipo. Heidiphyllum elongatum (Morris) Retallack 1981.
Heidiphyllum elongatum (Morris) Retallack 1981
Figura 9.C
Material estudiado. Punta Peters: Sgo-Pb 1703b, 1705 al 1712, 1716, 1717, 1718, 1768b.
Comentarios. Esta especie está ampliamente difundida en numerosos yacimientos del Triásico de casi todo el Gondwana. Es muy polimórfica, lo que ha generado que bajo este nombre específico se incorporasen numerosos tipos de hojas, desde algunas relativamente angostas hasta bastante más anchas. Esto ya fue señalado por Stpanicic et al. (1995), que también brindan una breve descripción genérica para hojas sin cutícula. Asimismo, Anderson y Anderson (1989), en sus abundantes y bien ilustradas láminas, reflejan muy bien esta diversidad morfológica.
PTERIDOPHYLLA incertae sedis
Género Linguifolium Arber emend. Retallack 1980
Especie tipo. Linguifolium lillieanum Arber 1913.
Linguifolium lillieanum Arber 1913
Figura 7.H
Material estudiado. Licán Ray: Sgo-Pb 1826, 1827, 1828a, 1829, 1830, 1831. Punta Peters: Sgo-Pb 1734, 1780b.
Comentarios. Esta especie, que es muy frecuente en Australia, Sudáfrica y América del Sur, es muy característica en su morfología, por lo que su identificación no implica problemas. La especie fue citada en Chile para el Triásico del Cajón Troncoso (Alta Cordillera del Maule) por Troncoso y Herbst (2000) y también está presente en la Formación La Ternera (Gnaedinger y Herbst, 2004).
Linguifolium sp. cf. L . tenison-woodsii (Etheridge) Retallack 1980
Figuras 6.E, 7.G
Material estudiado. Punta Peters: Sgo-Pb 1734, 1780.
Descripción. Una hoja incompleta, de unos 4,5 cm de longitud y 1,4 cm de anchura en la parte superior, y otra de unos 9 cm de logitud y 1 cm anchura; vena media gruesa, las venas laterales salen arqueadas bajo ángulos del orden de 65-75º, muchas de ellas bifurcadas una vez en el primer tercio de su extensión. Densidad aproximada de 18-20 venas/cm, medido cerca del borde.
Comentarios. Si bien el material es muy fragmentario, estos dos ejemplares se diferencian netamente de L. lillieanum por ser muy angostos, de bordes casi paralelos, y por la venación. Es evidentemente muy similar a lo que en la literatura se denomina L . Tenisonwoodsii ( vide Retallack, 1980, fig. 7.F-H).
Género Taeniopteris Brongniart 1832
Especie tipo. Taeniopteris vittata Brongniart 1832.
Taeniopteris parvilocus Anderson y Anderson 1989
Figuras 6.C-D, 9.A-B
Material estudiado. Cerro Tralcán: Sgo-Pb 1853b, 1870a, 1873, 1874a, 1886, 1894, 1895, 1900b, 1918.
Comentarios. De las muchas especies de Taeniopteris conocidas en América del Sur, esta es la primera vez que aparece esta forma sudafricana. Por sus caracteres morfológicos de tamaño, forma y venación es bastante fácil de identificar. Es lo suficientemente característica para que sus creadores (Anderson y Anderson, 1989) no hayan apelado en este caso al arbitrio de unir especies, como lo hicieran con otras, utilizando una suerte de nomenclatura poco rigurosa: por ejemplo, Taeniopteris cetilocus / anavolans /( homerifolius ) (lámina 228).
Gen. et sp. indet.
Figuras 5.I, 7.I-J
Material estudiado. Cerro Tralcán Sgo-Pb 1865b, 1877a, 1880c, 1892c, 1899a, 1900a, 1902c, 1903a, 1904a,c, 1906a, 1907, a908a, 1909a, 1910, 1911c, 1912a, 1913, 1921.
Descripción. Frondas de forma y tamaño desconocida, con segmentos por lo menos bipinnados de hasta 22 cm de largo x 8 cm de ancho, imparipinnados; raquis de segmento de hasta 3 mm de ancho, estriado; pinnas subopuestas, separadas aproximadamente 1 cm las del mismo lado, insertas por toda la base y decurrentes en el lado acro- y basiscópico; son 2 a 3 veces más largas que anchas, de bordes laterales con algún ensanchamiento en regiones cercanas a la base, convergiendo a ápices redondeados a ligeramente agudos. En porciones medias del segmento miden 5 cm de largo x 1,5 cm de ancho, mientras las apicales y basales son del orden de 1,5-1,8 cm de largo x 0,8-1 cm de ancho. En general están insertas bajo ángulos de 90ª y del orden de 75-80º en porciones apicales y basales. Venas paralelas, simples, saliendo de toda la base, muy ocasionalmente alguna vena dividida; densidad promedio de venas es del orden de 25-40 venas/cm, en pinnas basales y medias de 28-30 venas/ cm y en pinnas apicales de 32-38 venas/cm.
Comentarios. Estas frondes tienen una combinación de caracteres que, por una parte, las acercan a las Pteridospermales, como la bipinnación de los segmentos y aparentemente la bifurcación del raquis en dos pinnas (figura 7.J), con un tipo de venación lateral (paralelo, con pocas dicotomías y sin vena media), que es característico de varios géneros de las Cycadopsida ( sensu lato ) por otra. No hemos podido encontrar en la literatura disponible ninguna forma que se le parezca. No se propone formalmente una nueva entidad genérica, a la espera de especimenes más completos que provean más elementos de juicio; pero se trata, sin duda, de una forma nueva y singular. De entre las formas más o menos frecuentes en nuestras floras, por su contorno y tamaño, estos segmentos y pinnas se parecen lejanamente a las de Diplasiophyllum hughesi (Festmantel) Frenguelli, ilustrado por Gnaedinger y Herbst (1998), pero las diferencia muy claramente la venación y la distancia entre pinnas. Las mismas consideraciones imposibilitan, por ahora, una certera asignación a taxones de mayor rango.
Descripción cuantitativa
Las tres tafofloras estudiadas muestran evidentes diferencias en cuanto a su composición cualitativa, pero la descripción de cada una de éstas no estaría completa si no se agrega la información relativa a la representación cuantitativa de cada taxón en ellas, toda vez que en este aspecto también las diferencias son sumamente notorias (cuadro 2). Habida consideración que los criterios de colecta fueron los mismos en cada caso, los resultados parecen perfectamente comparables.
El significado exacto de este hecho escapa, por ahora, a la comprensión de los autores, pero ello no es óbice para consignar el hecho empírico. Por cierto que, tratándose de una tafoflora de hojas, es claro que la abundancia específica relativa está influida por factores como: la mayor o menor distancia de los individuos fuente a la cuenca de sedimentación, el carácter perennifolio o caducifolio de cada especie, la cantidad de hojas producidas por cada individuo, la energía del medio de sedimentación asociada a la mayor o menor capacidad de fosilizarse de cada tipo foliar (que tiene relación con, entre otras variables, su consistencia), etcétera. Sin embargo, no parece menos cierto que, a composición cualitativa similar entre dos tafofloras (que no es el caso presente), este parámetro podría ser usado en estudios paleoambientales y/o sedimentológicos. De este modo, la información que se provee en este acápite no debe ser interpretada como reflejo de una situación biológica, sino meramente como descripción tafonómica (un espectro tafonómico, podríamos decir), que da cuenta de un hecho. Para hacer esta descripción, hemos contabilizado, por taxón, los ejemplares claramente identificables e individualizables presentes en cada yacimiento. Los resultados así obtenidos han sido llevados a porcentajes y estos últimos expresados en la siguiente escala de abundancia relativa: > 50% 5, > 35%-50% 4, > 25%-35% 3, > 10%-25% 2, 2%-10% 1, < 2% +.

Cuadro 1. Lista de las especies y su distribución en las diferentes tafofloras./ Species list and floral assemblages.

Cuadro 2. Espectros tafonómicos de las tres paleofloras estudiadas / taphonomic spectre of the three studied tafoflora.
La idea de traducir los valores a una escala de abundancia relativa (de algún modo similar a procedimientos utilizados en fitosociología) responde al objetivo de minimizar la incertidumbre inherente a un conteo de esta naturaleza (generada, por ejemplo, por la partición de frondes en fragmentos menores). Es posible que la utilización de análogos modernos permita, a futuro, modelar esta situación y comprender cabalmente su significado. El registro de tal información resulta, así, necesario.
Discusión
Los biocrones conocidos de las especies presentes en cada uno de los yacimientos permiten definir que las tres tafofloras son del Triásico Superior (Cárnico Superior-Nórico), sin mayor precisión. Resulta notable, no obstante, constatar las grandes diferencias entre ellas, tanto desde el punto de la composición florística, como de la descripción basada en las dominancias tafonómicas (cuadro 2). En efecto, entre las tafofloras de Licán Ray y Punta Peters hay una similitud florística (índice de Sorensen) de tan sólo 25,0%, entre la de Licán Ray y cerro Tralcán de 21,4% y entre la de Punta Peters y cerro Tralcán de 23,5%. En los espectros tafonómicos presentados arriba es observable la neta diferencia entre los tres conjuntos, que, en algunos casos, llega al extremo que la especie dominante en una de las tafofloras ni siquiera esté presente en las otras.
No disponemos de argumentos definitorios que nos permitan concluir que las tres tafofloras sean estrictamente contemporáneas o no.
Así, resulta lo más apropiado proponer que las diferencias registradas no son atribuibles a meras diferencias de edad, sino a condiciones paleoambientales, lo cual es congruente con los datos aportados por la sedimentología, asociadas o no a diferencias cronológicas menores.
Agradecimientos
Nos es grato agradecer al Sernageomin (Delegación Sur) por las facilidades logísticas brindadas durante los trabajos de campo. Asimismo, a la National Geographic Society que apoyó económicamente estos trabajos. Los árbitros revisores del trabajo aportaron interesantes sugerencias, que agradecemos.
Bibliografia
Aguirre, L.y Levi, B. 1964. Geología de la Cordillera de los Andes de las Provincias de Cautín, Valdivia, Osorno y Llanquihue. Instituto de Investigaciones Geológicas , Boletín 17, 37 pp. [ Links ]
Anderson, J.M. y Anderson, H.M. 1989. Paleoflora of Southern Africa Molteno Formation (Triassic) . Vol. 2. Gymnosperms (excluding Dicroidium ). A.A. Balkema (Publishers), Rotterdam, 567 pp. [ Links ]
Andrews, H.N. 1955. Index of generic names of fossil plants 1820-1950. United States Geological Survey Bulletin 1013, 263 pp. [ Links ]
Arber, E.A.N. 1907. On Triassic species of the genera Zamites and Pterophyllum : types of fronds belonging to the Cycadophyta. Transactions Linnean Society London, Botany 2nd Ser . 7: 109-127. [ Links ]
Arber, E.A.N. 1913. A preliminary note on the fossil plants of the Mount Potts beds, New Zealand, collected by Mr. D.G. Lillie, biologist to Captain Scott's Antarctic Expedition in the Terra Nova. Proceeding of the Royal Society of London 86: 344-347. [ Links ]
Arrondo, O., Morel, E., Ganuza, D., Hervé, F., Parada, M. Y Muehlebach, W. 1988. La Formación Panguipulli y su contenido paleoflorístico, Chile. 5° Congreso Geológico Chileno, Actas 1: C137-C145. [ Links ]
Baldoni, A.M. 1972. El género Lepidopteris (Pteridospermae) en el Triásico de Argentina. Ameghiniana 9: 1-16. [ Links ]
Brongniart, A. 1828. Histoire des végétaux fossiles ou Recherches botaniques et géologiques sur les végétaux renfermés dans les diverses couches du globe . G. Dufour and Ed. D'Ocagne, Paris. Pars 1: 1-136. [ Links ]
Brongniart, A. 1832. Histoire des végétaux fossiles ou Recherches botaniques et géologiques sur les végétaux renfermés dans les diverses couches du globe . G. Dufour and Ed. D'Ocagne, Paris. Pars 3: 265-288. [ Links ]
Brongniart, A. 1849. Tableau des generes des végétaux fossiles considerés sous le point de vue de leur classification botanique et de leur distribution géologique. Dictionaire Universal Histoire Naturelle vol. 13, pp. 52-176. [ Links ]
Campos, A., Moreno, H., Muñoz, J., Antinao, J., Clayton, J. y Martin, M. 1998. Mapa Geológico Área de Futrono-Lago Ranco, Región de Los Lagos. Servicio Nacional de Geología y Minería, Mapas Geológicos , 8, 1 mapa escala 1: 100.000. [ Links ]
Corda, A.J. 1845. Flora Protogaea. Beiträge zur Flora der Vorwelt. Berlin, S. Calvary and Co., 128 pp. [ Links ]
Cucurella, J. 1978. [ Estudio geológico y radiométrico del valle inferior del río Bío-Bío, Provincia de Concepción, 8a Región . Memoria de Título, Universidad de Chile, Departamento de Geología, 111 pp. Inédito.]. [ Links ]
Davis, S.y Karzulovic, J. 1961. Deslizamientos en el valle del río San Pedro, Provincia de Valdivia, Chile. Universidad de Chile, Instituto de Geología, Publicación 20, 108 pp. [ Links ]
Duhart, P., McDonough, M., Muñoz, J., Martin, M. y Villeneuve, M. 2001. El Complejo Metamórfico Bahía Mansa en la Cordillera de la Costa del centro-sur de Chile (39°30'-42°00'S): geocronología K-Ar, 40Ar/39Ar y U-Pb e implicancias en la evolución del margen sur-occidental de Gondwana. Revista Geológica de Chile 28: 179-208. [ Links ]
Florin, R. 1936. Die Fossilien Ginkgophyten von Franz-Joseph-Land. II. Allgemeiner Teil. Palaeontographica B 81-82: 1-173. [ Links ]
Frenguelli, J. 1944a. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. XVI. Callipteridium argentinum n. sp. N otas del Museo de La Plata, Paleontología 64: 377-382. [ Links ]
Frenguelli, J. 1944b. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. XXI. Neocalamites ramaccioni n. sp. XXII. Neocalamites ischigualastii n. sp. Notas del Museo de La Plata, Paleontología 70-71: 479-492. [ Links ]
Frenguelli, J. 1947. El género Cladophlebis y sus representantes en la Argentina. Anales del Museo de La Plata (n.s). Paleontología B 2: 1-74. [ Links ]
Gnaedinger, S. y Herbst, R. 1998. La flora Triásica del Grupo El Tranquilo, provincia de Santa Cruz (Patagonia). Parte IV. Pteridospermae. Ameghiniana 35: 33-52. [ Links ]
Gnaedinger, S. y Herbst, R. 1999. La flora Triásica del Grupo El Tranquilo, provincia de Santa Cruz, Patagonia. Parte VI: Ginkgoales. Ameghiniana 36: 281-296. [ Links ]
Gnaedinger, S. y Herbst, R. 2001. Pteridospermas triásicas del Norte Chico de Chile. Ameghiniana 38: 281-298. [ Links ]
Gnaedinger, S. y Herbst, R. 2004. Pteridophylla del Triásico del Nort Chico de Chile. II. Los géneros Dejerseya Herbst, Linguifolium (Arber) Retallack y Yabeiella Oishi. Revista Museo Ciencias Naturales B. Rivadavia 6: 49-59. [ Links ]
Gothan, W., 1912. Über die Gattung Thinnfeldia Ettingshausen. Abhandlugen Naturhistorischen Gesellschaft Nüremberg 19: 67-80. [ Links ]
Halle, T.G., 1908. Zur kenntnis Mesozoischen Equisetales Schwedens. Kungliska Svenska Vetenskapakademie Handligar 43, 1: 1-40. [ Links ]
Hauser, A. 1970. Estudio integrado de los recursos naturales, Cautín, Geología. Instituto de Investigación de Recursos Naturales , Publicación 29: 13-72. [ Links ]
Heer, O. 1876. Beiträge zur Jura-Flora Ostsibieriens und des Amurlandes. Mémoirs de l'Académie Impérial des Sciences de Saint Petersbourg ser. 7, vol. 22, 12: 1-122 [ Links ]
Herbst, R. 1971. Palaeophytologia Kurtziana III. 7. Revisión de las especies Argentinas del género Cladophlebis . Ameghiniana 8: 265-281 [ Links ]
Herbst, R. 1977. Sobre Marattiales (Filicopsidae) Triásicas de Argentina y Australia. Parte I. El género Asterotheca . Ameghiniana 14: 1-18. [ Links ]
Herbst, R. 1988. La flora Triásica del Grupo El Tranquilo, provincia de Santa Cruz, Patagonia. Parte II: Filicopsida. Ameghiniana 25: 365-379. [ Links ]
Herbst, R. 1996. New species of Gleichenites (Gleicheniaceae, Filicales) from the Upper Triassic of Argentina and Chile. The Paleobotanist 43: 67-72. [ Links ]
Herbst, R. y Troncoso, A. 2000. Las Cycadophyta del Triásico de las Formaciones La Ternera y El Puquén (Chile). Ameghiniana 37: 283-292. [ Links ]
Herbst, R., Troncoso, A. y Melchor, R. 1998. Las Pteridophyta y el paleoambiente de la parte media de la Formación La Ternera (Triásico Superior) en Quebrada La Cachivarita, III Región, Chile. Revista Geológica de Chile 25: 85-107. [ Links ]
Hervé, F., Thiele, R. y Parada M. 1976. Observaciones Geológicas en el Triásico de Chile Central entre las latitudes 35º30' y 40º00' Sur. 1° Congreso Geológico Chileno, Actas 1: A297-A313. [ Links ]
Illies, H. 1970. Geología de los alrededores de Valdivia y volcanismo y tectónica en márgenes del Pacífico en Chile Meridional . Universidad Austral de Chile, Instituto de Geología y Geografía, 64 pp. [ Links ]
Jain, R.K. y Delevoryas, T. 1967. A Middle Triassic flora from the Cacheuta Formation, Minas de Petróleo, Argentina. Paleontology 10: 564-589. [ Links ]
Maheshwari, H.K. y Bajpai, U. 1992. Ginkgophyte leaves from the Permian Gondwana of the Rajmahal Bassin, India. Palaeontographica B 223: 131-149. [ Links ]
Menéndez, C.A. 1957. Asterotheca hilariensis sp. nov. del Triásico Superior de Hilario, San Juan. Ameghiniana 1: 25-30. [ Links ]
Minato, M. 1977. Carboniferous-Permian plant remains found at the border of lake Panguipulli, Valdivia Province, Chile. En: T. Ishikawa y L. Aguirre (eds.), Comparative Studies on the Geology of the Circum-Pacific Orogenic Belt in Japan and Chile Japan Society for the Promotion of Science, pp. 69-79. [ Links ]
Moreno, H., Hervé, F., Godoy, E. y Parada, M.A. 1976. [ Reconocimiento geológico del área de Pocilllas, Cauquenes, VIII Región. Universidad de Chile, Departamento de Geología, pp. 9. Inédito.]. [ Links ]
Munizaga, F., Hervé, F., Drake, R., Pankhurst, R. and Brook, M. 1988. Geochronology of the Lake Region of South Central Chile (39º-42ºS): Preliminary results. Journal of South American Earth Sciences 1: 309-316. [ Links ]
Parada M.A. y Moreno, H. 1980. Estratigrafía y ambiente de depositación de la formación Triásica Superior Llafquentué-Huimpil, Provincia de Cautín (38º30'S-72º40'W). Departamento de Geología, Universidad de Chile, Comunicaciones , 30: 15-31. [ Links ]
Parada M.A. y Munizaga, F. 1978. El Batolito de Panguipulli: Caracterización del plutonismo jurásico en la Cordillera de Los Andes de la Provincia de Valdivia, X Región (39º30'-40º15'S). Departamento de Geología, Universidad de Chile, Comunicaciones 23: 12-25. [ Links ]
Retallack, G.J. 1977. Triassic vegetation: Microfiche supplement to a paper, Reconstructing Triassic vegetation of Eastern Australia. Alcheringa 1: 253-283. [ Links ]
Retallack, G.J. 1980. Middle Triassic megafossil plants and trace fossils from Tank Gully, Canterbury, New Zealand. Journal Royal Society New Zealand 10: 31-63. [ Links ]
Retallack, G.J. 1981. Middle Triassic megafossil plants from Long Gully, near Otematata, north Otago, New Zealand. Journal Royal Society New Zealand 11: 167-200. [ Links ]
Rodríguez, C., Pérez, Y., Moreno, H., Clayton, J., Antinao, J.L., Duhart, P. y Martin, M. 1999. Área de Panguipulli-Riñihue, Región de Los Lagos. Servicio Nacional de Geología y Minería, Mapas Geológicos, 10, 1 mapa escala 1: 100.000. [ Links ]
Schimper, W. 1869. Traité de Paléontologie Végétale . Tomo 1. J.B. Boilliére et Fils, Paris, 738 pp. [ Links ]
Seward, A.C. 1911. The Jurassic Flora of Yorkshire. The Naturalist 1-8: 85-94. [ Links ]
Seward, A.C. 1919. Fossil Plants. A text book for students of geology and botany . II. Cambridge University Press, Cambridge, 543 pp. [ Links ]
Seward, A.C. 1926. The Cretaceous plant-bearing rocks of western Greenland. Philosophical Transactions Royal Society London 215B: 57-174. [ Links ]
Shirley, J. 1898. Additions to the flora of Queensland. Mainly from the Ipswich Formation, Trias-Jura System. Bulletin Geological Survey of Queensland 7: 1-25. [ Links ]
Stipanicic, P.N., Herbst, R. y Bonetti, M.I.R. 1995. Floras Triásicas. En: Revisión y actualización de la obra paleobotánica de Kurtz en la República Argentina. Actas Academia Nacional Ciencias (Córdoba) 11: 127-184. [ Links ]
Tavera, J. 1960. [ El Triásico del valle inferior del río Biobío. Estudio Geológico . Universidad de Chile, Instituto de Geología, 26 pp. Inédito]. [ Links ]
Tavera, J., 1971. [ Informe-estudio sobre material paleontológico florístico proveniente de la Formación Panguipulli y Cerro Tralcán . Universidad de Chile, Departamento de Geología, 20 pp. Inédito.]. [ Links ]
Townrow, J.A., 1956. The genus Lepidopteris and its southern hemisphere species. Norske Videnskap-Akademi I Oslo. I. Mat. Naturv. Klasse , 2: 3-28. [ Links ]
Townrow, J.A. 1967. On Rissikia and Mataia podocarpaceous conifers from the Lower Mesozoic of southern lands. Papers and Proceedings Royal Society of Tasmania 101: 103-136. [ Links ]
Troncoso, A. y Herbst, R. 2000. La tafoflora triásica del Cajón Troncoso, Alta Cordillera del Maule, 7° Región, Chile. Revista Museo Argentino Ciencias Naturales B. Rivadavia n. s . 2: 137-144. [ Links ]
Troncoso, A., Gnaedinger, S. y Herbst, R. 2000. Heidiphyllum , Rissikia y Desmiophyllum (Pinophyta, Coniferales) en el Triásico del norte chico de Chile y sur de Argentina. Ameghiniana 37: 119-125. [ Links ]
Walkom, A.B. 1917. Mesozoic floras of Queensland. Part I. Concluded. The flora of the Ipswich and Walloon Series (d.) Ginkgoales, (e.) Cycadophyta, (f.) Coniferales. Queensland Geological Survey , Publication 358, 38 pp. [ Links ]
Zavattieri, A.M., Herbst, R. y Muñoz, J. 2003. Microflora de la Formación Panguipulli (Triásico Superior), en Licán Ray, Lago Calfquén, 10° Región, Chile. Ameghiniana 40: 585-600. [ Links ]
Recibido: 26 de mayo de 2003.
Aceptado: 12 de febrero de 2004.

