










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.2 Buenos Aires mar./jun. 2005
Mamíferos fósiles y edad de la Formación Salicas (Mioceno tardío) de la sierra de Velasco, La Rioja, Argentina
Adán A. Tauber (h) 1
1 Cátedra de Paleontología, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Av. Vélez Sársfield 299, 5000 Córdoba, Argentina.
Resumen. Se registraron restos óseos de mamíferos fósiles en la Formación Salicas, aflorante en el extremo norte de la sierra de Velasco, unidad que fuera referida previamente al Plioceno con dudas, al Plioceno superior o al Mioceno tardío. Esta unidad está compuesta por areniscas arcillosas, en general friables y de color marrón en los tramos estratigráficos bajos, y marrón rojizo en los niveles superiores. Además, se hallan intercalaciones de paleosuelos y conglomerados osíferos de origen fluvial. Entre los fósiles se encontraron restos de anuros, tortugas y mamíferos. Los principales taxones son: Macrochorobates scalabrinii Scillato Yané, Proeuphractus limpidus Ameghino, Chaetophractus sp., Eosclerocalyptus planus Castellanos, Neophanomys biplicatus Rovereto, Lagostomus ( Lagostomopsis ) cf. pretrichodactyla Kraglievich, Orthomyctera andina Kraglievich, Potamarchus sp., Protypotherium sp., Pseudotypotherium sp., Hemihegetotherium cf. Torresi Zetti y cf. Tremacyllus . Estos taxones indican una edad correspondiente al Mioceno tardío (“Huayqueriense”); sin embargo ésta es una asociación diferente a las registradas en la región pampeana, por lo que se analiza su significado. Para explicar esta diferencia de asociaciones, se propone como hipótesis una retracción en sentido esencialmente latitudinal del género Protypotherium durante el Mioceno tardío, desde la región pampeana hacia el norte. Algunos de estos taxones fueron registrados en el Miembro El Jarillal de la Formación Chiquimil y la parte inferior de la Formación Andalhuala (“Huayqueriense”) de Catamarca, con las cuales se correlacionan los estratos portadores.
Abstract. Fossil mammals and age of the Salicas Formation ( late Miocene ), sierra de Velasco , La Rioja province , Argentina . Bone remains of fossil mammals from the Salicas Formation, appearing in the northern end of Sierra de Velasco, La Rioja province, Argentina, are reported. This stratigraphic unit was previously assigned to the Pliocene, Upper Pliocene and to the Late Miocene. It is made up of clayey sandstone, friable in general, brownish in the lower layers, and reddish brown in the upper levels. There are also intercalated paleosoil layers, and fossiliferous conglomerates of fluvial origin. Among the findings, anuran, turtle and mammal fossils are reported. The main species are: Macrochorobates scalabrinii Scillato Yané, Proeuphractus limpidus Ameghino, Chaetophractus sp., Eosclerocalyptus planus Castellanos, Neophanomys biplicatus Rovereto, Lagostomus ( Lagostomopsis ) cf. pretrichodactyla Kraglievich, Orthomyctera andina Kraglievich, Potamarchus sp., Protypotherium sp., Pseudotypotherium sp., Hemihegetotherium cf. Torresi Zetti and cf. Tremacyllus . All these correspond to the Late Miocene (Huayquerian mammalian age). This association is different from all the other findings in the Pampean region, hence its significance is discussed. Then, the hypothesis of a Late Miocene latitudinal retraction of the genus Protypotherium from the Pampean region towards the North, is proposed. Some of the species were also reported from El Jarillal Member of the Chiquimil Formation and from the lower part of the Andalhuala Formation (Huayquerian) in Catamarca province, with which the bearing sediments are correlated.
Key words. Salicas Formation; Mammals; Late Miocene; La Rioja Province; Argentina.
Palabras clave. Formación Salicas; Mamíferos; Mioceno tardío; La Rioja; Argentina.
Introducción
Los flancos y los valles de la sierra de Velasco en su parte septentrional se encuentran cubiertos por sedimentos que fueron atribuidos al Paleozoico superior (Amos y Zardini, 1962; Aceñolaza y Bortolotti, 1981). Por encima se acumularon pelitas, areniscas y conglomerados del Neógeno y Cuaternario (Turner, 1971; Sosic, 1972, 1973), sin que se conozcan bien las relaciones estratigráficas entre las diferentes unidades.
Entre los depósitos neógenos se encuentran las Formaciones Huaco (De Alba, 1979) y Salicas (Sosic, 1972, 1973; = Schaqui, Turner, 1971). La Formación Los Llanos también fue considerada durante mucho tiempo como neógena (Mioceno tardío o Plioceno) (Machicote, 1946; Amos y Zardini, 1962; Zuzek, 1978; Aceñolaza y Bortolotti, 1981, etc.). Sin embargo, en dos afloramientos del valle de Sanagasta y atribuidos a esta última unidad, se hallaron huevos de dinosaurios saurópodos pertenecientes al Cretácico tardío (Hünicken et al ., 2001, 2002; Tauber, 2002). De esta manera se verificó la hipótesis original de Bodenbender (1911), quien atribuyó con dudas al Cretácico Superior? a la Formación Los Llanos (=Estratos de Los Llanos).
De Alba (1979) definió la Formación Huaco, refiriéndose a los niveles más bajos aflorantes en el semibolsón (o bolsón) del mismo nombre, correlacionándola litoestratigráficamente con la Formación Los Llanos expuesta en las cercanías de Sanagasta.
La Formación Salicas (Sosic, 1973), también llamada Formación Schaqui por Turner (1971), aflora en el extremo norte de la sierra de Velasco, tanto sobre la vertiente occidental como en la oriental, en los departamentos San Blas de Los Sauces, Arauco, Castro Barros, y Capital, provincia de La Rioja. Esta unidad estratigráfica fue asignada al Plioceno con dudas (Araucanense?) (Turner, 1971; Sosic, 1973; Combina y Santiesteban Bové, 2000a), probablemente al Plioceno superior (Groeber, fide , Sosic, 1973). Esta interpretación fue realizada por estos autores sobre la base de correlaciones litoestratigráficas regionales, ya que hasta el presente la Formación Salicas ha brindado material paleontológico muy escaso y fragmentario. Sólo se han citado algunos hallazgos procedentes de los niveles estratigráficamente más bajos, donde se registraron restos atribuidos a Typotheriopsis sp., por lo que fue asignada al Mioceno superior (Chasiquense-Huayqueriense, Mulé et al ., 1997), cf. Tremacyllus , Mesotheriidae indet. y Pachyrukhinae indet. (Huayqueriense, Bossi, et al ., 1999, p. 356). Sin embargo, el género Tremacyllus tiene una distribución que abarca edades desde el Mioceno tardío (Chasiquense) al Plioceno tardío (Marplatense) (Cione y Tonni, 1995; Cerdeño y Bond, 1998; Cerdeño, 2000; Cerdeño y Montalvo, 2001, 2002).
En campañas paleontológicas realizadas a diferentes afloramientos de la Formación Salicas en el sector noroeste de la sierra de Velasco y el borde sur del Salar de Pipanaco durante los años 1998, 2000 y 2001, se han observado numerosos sitios paleontológicos donde se registraron diversos materiales, entre ellos diatomeas, gasterópodos, troncos silicificados con galerías de insectos llamados comúnmente taladros de la Familia Buprestidae, tallos y hojas de vegetales acuáticos, icnitas de invertebrados y de aves, y restos óseos de vertebrados, entre ellos de anuros, quelonios y aves, probablemente de la Familia Ciconidae.
Esta situación obliga a replantear las hipótesis geocronológicas y a realizar nuevos estudios estratigráficos de detalle de estas unidades neógenas, temas esenciales para comprender la evolución geológica de la Sierra de Velasco.
El objetivo de este trabajo es realizar un análisis de una nueva colección de fósiles compuesta por restos óseos de mamíferos, para verificar o contrastar las hipótesis previas sobre la edad de la Formación Salicas, al menos de los niveles fosilíferos prospectados. Sobre la base de estas hipótesis se analizarán los resultados dentro del marco geológico regional.
Abreviaturas. CRILAR-PZ: Centro Regional de Investigaciones Científicas y Transferencia Tecnológica de La Rioja, sección Paleozoología. NF: nivel fosilífero. I, C, P y M dientes superiores e i, c, p y m dientes inferiores. Sm, Fm, Sh, Gm, Gms, Sm, Sh, Sl, St y CH son los códigos de facies reconocidas por Miall (1996).
Geología
La mitad norte de la sierra de Velasco se halla dividida en dos grandes ramas de rumbo aproximadamente meridional, por la acción de fallas inversas muy activas que se encuentran bajo las abruptas laderas orientales. Las vertientes occidentales tienen una pendiente mucho más suave, formando superficies relativamente planas de gran extensión. Éstas probablemente estén vinculadas con las paleosuperficies (Jurásico-Terciario temprano) que originaron las pampas de altura en las sierras de Córdoba (Carignano et al ., 1999). Las rocas del basamento que componen la sierra de Velasco, en su extremo norte, es el granito porfírico de la Formación Paimán y complejo Antinaco (Sosic, 1973) y granitoides porfíricos milonitizados que son parte de los intrusivos de Huaco y de San Blas (Báez et al ., 2002).
Sobre ambos flancos de las dos ramas de la mitad norte de la sierra de Velasco se depositaron los sedimentos predominantemente finos que conforman la Formación Salicas (Terciario) y posteriormente los materiales más gruesos que componen la Formación Las Cumbres (Plio-Pleistoceno). Estos depósitos sobreyacen en inconformidad (en la ladera occidental) y están en contacto tectónico (en la vertiente oriental) con el granitoide porfírico milonitizado del intrusivo de San Blas (figura 1). La Formación Salicas en este sector y en el borde suroeste del salar de Pipanaco alcanzaría, según Sosic (1973), los 600 a 800 m de espesor. En esta región está compuesta principalmente por areniscas finas macizas y limolitas arcillosas de color predominantemente marrón rojizo variando hasta gris amarillento en menor proporción.
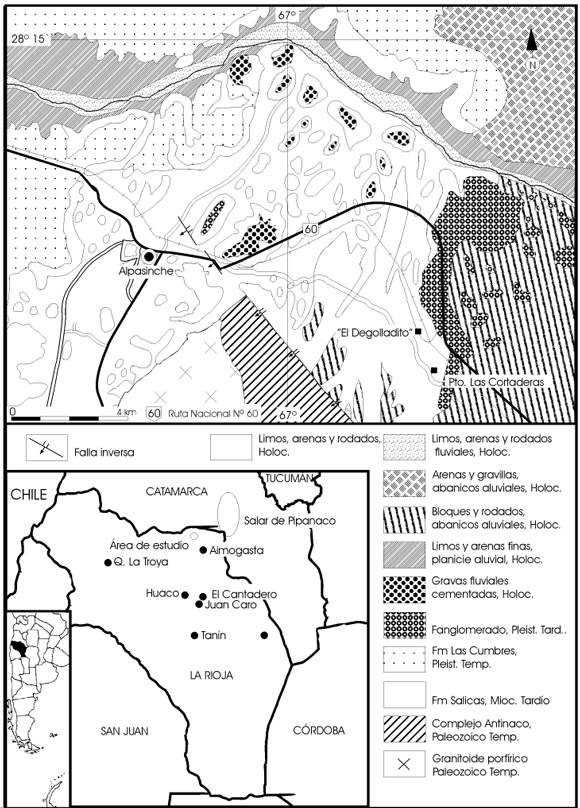
Figura 1. Mapa de ubicación del área de estudio donde se recolectaron los restos fósiles de la Formación Salicas / location map of the locality from which the fossils of the Salicas Formation were collected.
Los fósiles que se analizan en este trabajo proceden del área cercana al antiguo puesto La Cortadera (28º 20´ S y 66º 57´ O) y al paraje El Degolladito (departamento Arauco), ubicado sobre la vertiente oriental de la rama noroccidental de la sierra de Velasco, al oeste de la ruta nacional nº 60, unos 26,5 km al noroeste de la ciudad de Aimogasta (figura 1).
La secuencia sedimentaria que allí aflora tiene un espesor visible de 100 m, conformando un homoclinal con rumbo general Az 48º (variando entre los 16º y los 75º) y 2º 48´ de inclinación promedio hacia el Oeste (medido por taquimetría, n= 10), de tal manera que los niveles estratigráficos más bajos afloran hacia el Este, frente al puesto La Cortadera; no obstante se observan importantes variaciones en el rumbo y buzamiento de los estratos, debido a los movimientos que elevaron la sierra de Velasco, llegando hasta 70º de inclinación de buzamiento hacia el Sur. Esto sumado la alternancia de estratos de areniscas con diferente grado de cementación ha producido un paisaje de cañadones poco profundos.
En el sector noroeste de la sierra de Velasco, en los alrededores de la localidad de Salicas, se puede observar el contacto entre los niveles rojizos de la Formación Salicas, apoyando directamente sobre el basamento granítico que compone el granotoide porfírico. La Formación Salicas está cubierta mediante discordancia angular por gravas gruesas a finas, granodecrecientes, parcialmente cementadas con carbonatos, de probable edad cuaternaria o plio-pleistocena, pertenecientes a la Formación Las Cumbres (Sosic, 1973). Los afloramientos de mayor espesor de esta última unidad se encuentran sobre la Ruta Nacional Nº 60, cerca del puesto La Cortadera.
Sobre la vertiente noreste se halla el frente activo de la sierra de Velasco y el contacto entre la Formación Salicas y el basamento no es visible por estar cubierto por conos aluviales dispuestos en tres unidades, un complejo de dunas hacia el este y barreales (Combina y Santiesteban Bové, 2000b). Sin embargo, este contacto sería de origen tectónico (Sosic, 1973).
En la secuencia estudiada predominan las areniscas finas macizas ( Sm ) y limolitas arcillosas macizas ( Fm ), dispuestas en bancos tabulares amalgamados con estratificación horizontal. El grado de litificación por la presencia de cemento calcáreo varía entre los diferentes bancos, produciendo una erosión diferencial. Las areniscas son en general, friables y sus colores varían desde el marrón o gris en los niveles estratigráficos más bajos y en las proximidades del puesto La Cortadera a marrón rojizo en los tramos estratigráficos altos, hacia el sector noroccidental de la sierra de Velasco.
Estas facies varían lateralmente a areniscas finas laminadas ( Sh ) con icnofósiles de invertebrados ( Taenidium barreti Bradshaw) y aves (Familia Anatidae) (figura 2, NF.4), o areniscas macizas con rizoconcreciones y rellenos de grietas de desecación (28º 22' 113 S y 66º 56' 068 O) (figura 2, NF.7). Asociada a estas facies, especialmente en la parte media de la columna se observaron cuerpos tabulares compuestos por calcretes continuos. Esta asociación de facies se interpreta como depósitos de planicie fluvial y llanuras de inundación, sometida a variaciones periódicas de inundación y desecación, y por la presencia de calcretes y grietas de desecación pudo haberse formado bajo un clima semiárido.
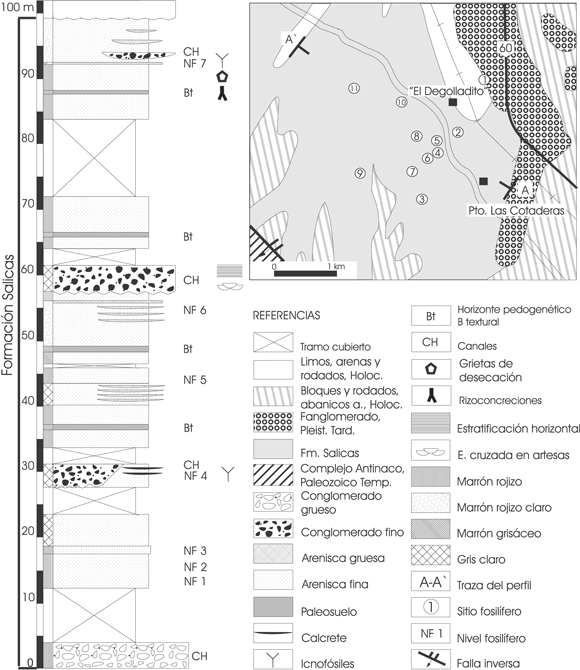
Figura 2. Columna estratigráfica con los niveles fosilíferos de la Formación Salicas y mapa con los sitios paleontológicos / stratigraphic column with fossiliferous levels of Salicas Formation and map with palaeontological sites.
Es importante señalar aquí, que en otras secciones de la Formación Salicas, por ejemplo, en la parte comprendida entre el área de estudio y San Blas de Los Sauces, existen niveles de pelitas que contienen nódulos y delgadas costras de evaporitas. Estos estratos serían más altos estratigráficamente, por lo que es posible que el ambiente haya evolucionado localmente a una playa salina (o lago somero) o playa de bolsón. Sin embargo, se considera más probable que el clima haya variado hacia condiciones de mayor aridez durante la depositación de la Formación Salicas. Esta última hipótesis coincide con la intensa aridización climática interpretada en la sección superior de la Formación Andalhuala (Bossi et al ., 1999: 348), en el río Villavil.
En el área de estudio se observaron 4 grandes canales (CH), con un espesor promedio de 4 m, rellenos en general con gravas macizas ( Gm ) de diferentes granulometrías (ver columna estratigráfica, figura 2). En la base de la columna la grava es más gruesa, llegando a 10 cm de diámetro máximo y en parte clasto soportada ( Gms ). En los cuerpos subsiguientes la granulometría del relleno disminuye a grava mediana y fina, y aumenta la proporción de matriz. Esta última está compuesta por areniscas gruesas y macizas ( Sm ), en parte con estratificación delgada horizontal, entrecruzada de bajo ángulo o entrecruzada en artesas ( Sh, Sl y St ). En algunos sectores se observaron trazas de invertebrados en las facies de areniscas con estratificación horizontal. El relleno de algunos canales es un paraconglomerado osífero, como en el sitio 8 (28º 20' 795 S y 66º 57' 983 O, figura 2, segundo canal de la columna), siendo indeterminables los restos fósiles.
La buena representación de depósitos de canal, los potentes bancos de areniscas amalgamadas y la ausencia de cuerpos de acreción lateral podría interpretarse a estas asociaciones de facies como depositadas en amplias llanuras arenosas ( sand flat ) ( sensu Ramos, 1992) en las que se desarrolló una red de canales entrelazados de baja sinuosidad y poca profundidad, aumentando la carga de fondo arenosa y disminuyendo la grava, hacia el techo de esta formación.
En la zona de El Degolladito se observó la presencia de al menos tres niveles arcillosos de 0,8-1 m de espesor, siendo su color marrón rojizo, más oscuro que los demás. Sus techos son netos y la base es transicional, teniendo una estructura en bloques cúbicos y pequeños. Por la presencia de estas características se interpreta que son horizontes pedogenéticos B texturales (Bt) bien desarrollados en las antiguas zonas de intercanal, siendo muy continuos en el área, por lo que resultan excelentes niveles guías (figura 2).
Paleontología sistemática
Clase M AMMALIA Linneo, 1758
Superorden X ENARTHRA Cope, 1869
Orden C INGULATA Illiger, 1811
Familia D ASYPODIDAE Bonaparte, 1838
Subfamilia E UPHRACTINAE Pocock, 1924
Tribu E UPHRACTINI Pocock, 1924
Género Macrochorobates Scillato Yané, 1980
Macrochorobates scalabrinii (Moreno y Mercerat) Scillato Yané, 1980a
Figura 3.A
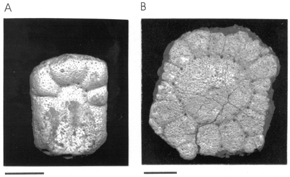
Figura 3. A, Macrochorobates scalabrinii, placa fija (CRILAR-PZ 71) / fixed scute . B, Eoesclerocalyptus planus , placa del caparazón dorsal en vista externa (CRILAR-PZ 61) / plate of dorsal shell in external view. Las escalas representan 1 cm / Scale bars equal 1 cm.
Material y procedencia. Mitad anterior de una placa móvil (CRILAR-PZ 70); placa fija (CRILAR-PZ 71). Estas piezas proceden de los sitios 4 y 6 respectivamente y del nivel fosilífero 2.
Observación. Estas piezas fueron referidas a la especie M. scalabrinii debido a que tienen un tamaño menor y las figuras periféricas de la placa fija son globosas, comparadas con la especie M. Chapalmalensis (Esteban et al ., 2001a).
Medidas (en mm). CRILAR-PZ 70: ancho 19,85; longitud 26,76.
Género Proeuphractus Ameghino, 1886
Proeuphractus limpidus Ameghino, 1886
Figura 4.A
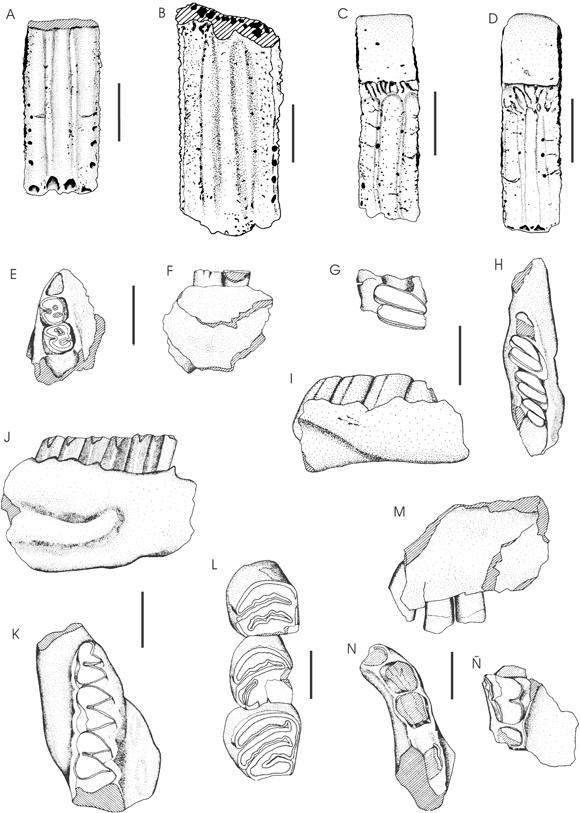
Figura 4. A, Proeuphractus limpidus , fragmento de placa de una banda movible en vista externa (CRILAR-PZ 64) / movable scute fragment in external view. B, Proeuphractus sp., porción de placa de una banda movible en vista externa (CRILAR-PZ 65) / movable scute fragment in external view . C-D, Chaetophractus sp.; C, placa de una banda movible en vista externa (CRILAR-PZ 46) / m ovable scute fragment in external view; D, placa de una banda movible en vista externa (CRILAR-PZ 1) / movable scute fragment in external view . E-F, Neophanomys biplicatus ; E, fragmento de dentario izquierdo con m1 y m2 implantados en vista oclusal (CRILAR-PZ 30) / left mandibular fragment with m1 and m2, in oclusal view ; F, fragmento de dentario izquierdo con m1 y m2 implantados en vista lingual (CRILARPZ 30) / left mandible fragment with m1 and m2, lingual view . G-I, Lagostomus ( Lagostomopsis ) cf. pretrichodactyla ; G, molar inferior en vista labial (CRILAR-PZ 35) / l ower molar in labial view ; H, fragmento de dentario izquierdo con parte del p4 y m1-m3 en vista oclusal (CRILAR-PZ 34) / l eft mandible fragment with m1-m3 in oclusal view ; I, en vista externa / external view . J-K, Orthomyctera andina ; J, porción de dentario derecho con p4-m2 en vista externa (CRILAR-PZ 41) / right dentary fragment with p4-m2 in external view; K, en vista dorsal / in dorsal view. L, Potamarchus sp., maxilar izquierdo con P4-M2 en vista oclusal (CRILAR-PZ 31) / occlusal view of left cheek teeth. M-Ñ, Protypotherium sp.; M, premaxilar izquierdo con las porciones intraalveolares de la serie I1-C en vista labial (CRILAR-PZ 20) / left premaxillary with portions of I1-C, labial view ; N, en vista oclusal / in oclusal view; Ñ, maxilar izquierdo con M2? en vista oclusal (CRILAR-PZ 38 a) / left maxillary with M2?, oclusal view . Las escalas representan 0,5 cm / Scale bars equal 0,5 cm.
Material y procedencia. Fragmentos de tres placas móviles (CRILAR-PZ 64 y 67/1-2). Estas piezas proceden de los sitios 4 y 6 respectivamente y del nivel fosilífero 2.
Observación. Las placas móviles tienen un tamaño reducido y una figura central alargada y recta, delimitada por surcos anchos y de fondo cóncavo. Las perforaciones pilíferas están ubicadas en los márgenes laterales y posteriores. Estas características permitieron asignar este material a Proeuphractus limpidus .
Medidas (en mm). CRILAR-PZ 64: ancho 6,40; CRILAR-PZ 67-2: ancho 6,44; CRILAR-PZ 67-1: ancho 6,34.
Proeuphractus sp.
Figura 4.B
Material y procedencia. Se recolectaron dos fragmentos de placas móviles (CRILAR-PZ 6 y 65). Estas piezas proceden de los sitios 4 y 6 respectivamente y del nivel fosilífero 2.
Observación. Estos fragmentos de placas determinados como Proeuphractus sp. se diferencian de CRILAR-PZ 64 y 67/1-2 solamente por su mayor tamaño, por lo que podrían pertenecer a una posible nueva especie como lo señalan Esteban et al . (2001a, 2003) para algunos materiales procedentes de la Formación Cerro Azul, en la provincia de La Pampa.
Medidas (en mm). CRILAR-PZ 65: ancho 13,6 (figura 4.B); CRILAR-PZ 6: ancho 8,9.
Género Chaetophractus Fitzinger, 1871
Chaetophractus sp.
figura 4.C-D
Material y procedencia. Siete placas móviles. Los osteodermos CRILAR-PZ 3, CRILAR-PZ 46 y CRILAR-PZ 66, proceden de los sitios 4, 5 y 8 respectivamente; las placas CRILAR-PZ 51, CRILARPZ 19-2 y CRILAR-PZ 68 proceden del sitio 6 y todas del nivel fosilífero 2. El osteodermo CRILAR-PZ 1, procede del sitio 9 y del nivel fosilífero 5.
Observación. Talla pequeña, placas móviles con las figuras laterales mejor definidas que en Zaedyus y divididas en figuritas menores, mejor definidas que en cualquier otro Euphractini. La zona de separación con la parte articular más desarrollada y rugosa que en Zaedyus . Numerosas perforaciones pilíferas de tamaño uniforme en el margen posterior. Estos caracteres tomados de Scillato Yané (1982) y Vizcaíno y Bargo, (1993), permitieron asignar estos materiales a Chaetophractus sp.
Medidas (en mm). CRILAR-PZ 51: ancho 5,9; CRILAR-PZ 1: ancho 4,55, longitud 16,45; CRILAR-PZ 3: ancho 5,02, longitud 18,38; CRILAR-PZ 46: ancho 4,7, longitud 15,7.
Superfamilia G LYPTODONTOIDEA (Gray, 1869) Simpson, 1931
Familia G LYPTODONTIDAE Gray, 1869
Subfamilia S CLEROCALYPTINAE Trouessart, 1893
Tribu P LOHOPHORINI Hoffstetter, 1958
Género Eosclerocalyptus Ameghino, 1919
Eosclerocalyptus planus (Rovereto, 1914) Castellanos, 1940
Figura 3.B
Material y procedencia. Una placa de las últimas filas de la región posterior del caparazón dorsal (CRILAR-PZ 61), procedente del sitio 8 y del nivel fosilífero 4.
Descripción. Placa subhexagonal con una figura central de contorno subcircular delimitada por un surco que posee finas perforaciones pilíferas en las intersecciones de éste con los surcos que delimitan cada figurita periférica. La figura central tiene una pequeña y muy suave depresión de contorno circular y concéntrica característica de este género. La figura central está limitada por 15 figuritas periféricas poligonales, la mayoría de las cuales tienen cuatro lados y algunas cinco lados. Hay otras tres figuritas de pequeño tamaño que se intercalan entre las anteriores, en el borde externo de la placa. Todas las figuras periféricas están limitadas entre sí por surcos bien definidos.
Observación. Las placas del caparazón de los Sclercalyptini poseen una morfología muy similar entre sí, dificultándose la determinación con una sola placa aislada, sin embargo este material fue asignado a Eosclerocalyptus planus por el alto número de figuras periféricas y por la presencia de una pequeña depresión dentro de la figura central.
Se infiere que la placa (CRILAR-PZ 61) pertenece a las últimas filas de la región posterior del caparazón dorsal, por la presencia de un mayor número de figuras periféricas, mayor diámetro de la figura central y la presencia de algunas figuras formando una segunda hilera de periféricas (Cabrera, 1944).
Orden R ODENTIA Bowdich, 1821
Suborden H YSTRICOGNATHI Tullberg, 1899
Superfamilia O CTODONTOIDEA Simpson, 1945
Familia O CTODONTIDAE Waterhouse, 1839
Género Neophanomys Rovereto, 1914
Neophanomys biplicatus Rovereto, 1914
Figura 4.E-F
Material y procedencia. Fragmento de dentario izquierdo con m1 y m2 implantados (CRILAR-PZ 30). El material procede del sitio 6 y del nivel fosilífero 2.
Observación. La morfología dentaria de esta especie varía considerablemente con la intensidad del desgaste, según las observaciones de Verzi et al . (1999). Teniendo en cuenta esto, se infiere que el espécimen CRILAR-PZ 30 presenta un nivel de desgaste intermedio. En el extremo labial del mesofléxido del m2 se observa la presencia de un pequeño spur. Este carácter sería variable dentro de esta especie y correspondería a un vestigio de un lófido (Verzi et al . 1999).
Superfamilia C HINCHILLOIDEA Kraglievich, 1940
Familia C HINCHILLIDAE Bennet, 1833
Género Lagostomus Brookes, 1828
Lagostomus ( Lagostomopsis ) cf. Pretrichodactyla (Rovereto, 1914) Kraglievich, 1926
Figura 4.G-I
Material y procedencia. Porción de dentario izquierdo, conteniendo parte del p4 y m1, m2 y m3 (CRILAR 33), procedente del sitio fosilífero 4. Porción de dentario izquierdo, conteniendo parte del p4 y m1, m2 y m3 (CRILAR 34) y un molar inferior aislado (CRILAR-PZ 35), procedentes del sitio 6. Todos estos materiales provienen del nivel fosilífero 2.
Observación. El espécimen CRILAR-PZ 34 es juvenil, por eso es notablemente más pequeño que CRILAR-PZ 33, otra diferencia entre ambos es que en el primero las láminas de los molares se disponen en forma muy oblicua con respecto al plano sagital. Kraglievich (1934) reconoce a Lagostomopsis como un género diferente de Lagostomus , mientras que Francis y Mones (1966) incluyen al primero como un subgénero del segundo. Las diferencias entre ambos taxones se refieren esencialmente a variaciones en la talla y a distintas proporciones craneanas y dentales, por lo tanto en este trabajo se considera a Lagostomopsis un subgénero dentro de Lagostomus , coincidiendo con Francis y Mones (1966) y Vucetich y Verzi (1995).
Superfamilia C AVIOIDEA Kraglievich, 1930
Familia C AVIIDAE Gray, 1821
Subfamilia D OLICHOTINAE Pocock, 1922
Género Orthomyctera
Orthomyctera andina (Rovereto, 1914), Kraglievich, 1934
Figura 4.J-K
Material y procedencia. Porción del dentario derecho con p4-m3 (CRILAR-PZ 50). Fragmentos de ambos maxilares con P4-M1 derecho y la porción intraalveolar del P4 izquierdo y otra porción de diente (CRILAR-PZ 39). Estos materiales proceden del sitio fosilífero 4. Porción del dentario derecho con p4-m2 (CRILAR-PZ 41), que fue recolectado en el sitio 6. Todos estos elementos provienen del nivel fosilífero 2.
Observación. Estos materiales fueron asignados a O. andina, por su talla pequeña, por la forma de la cresta masetérica, que llega hasta la posición del m1 y está bien doblada hacia arriba y el prisma anterior del p4 se prolonga hacia delante y es falciforme.
Medidas (en mm). CRILAR-PZ 39: diámetro mesio distal del M1: 4,1, diámetro transversal del M1: 3,6. CRILAR-PZ 41: diámetro mesio distal del p4: 4,06, diámetro transversal del p4: 3,14; diámetro mesio distal del m1: 3,92, diámetro transversal del m1: 4,08; diámetro mesio distal del m2: 5,46, diámetro transversal del m2: 3,8. CRILAR-PZ 50: diámetro mesio distal del m2: 4,9, diámetro transversal del m2: 3,32.
Familia D INOMYIDAE Alston, 1876
Subfamilia P OTAMARCHINAE Simpson, 1945
Potamarchus sp.
Figura 4.L
Material y procedencia. Maxilar izquierdo con P4-M2 (CRILARPZ 31). El material procede del sitio 6 y nivel fosilífero 2.
Observación. Según la diagnosis de Potamarchus (Pascual et al ., 1966) los pliegues interlaminares originalmente abiertos en ambas caras al comenzar el desgaste, pierden prontamente la comunicación externa, persistiendo sólo la interna. En el espécimen CRILAR-PZ 31 el pliegue posterior del m2 se comunica con la cara externa, atribuyéndose esto a una variación intragenérica.
Medidas (en mm). CRILAR-PZ 31: longitud de la serie P4-M2: 22,2; diámetro mesio distal del P4: 6,48, diámetro transversal del P4: 8,04; diámetro mesio distal del M1: 7,98, diámetro transversal del M1: 7,16; diámetro mesio distal del M2: 7,96, diámetro transversal del M2: 7,43.
Orden N OTOUNGULATA Roth, 1903
Suborden T YPOTHERIA Zittel, 1892
Familia I NTERATHERIIDAE Ameghino, 1887
Subfamilia I NTERATHERIINAE Simpson, 1945
Género Protypotherium Ameghino, 1887
Protypotherium sp.
Figura 4.M-Ñ
Material y procedencia. Fragmento de premaxilar izquierdo con porciones de la serie I1-C (CRILAR-PZ 20) y porción de maxilar izquierdo con M2? (CRILAR-PZ 38a). El material procede del sitio 7 y nivel fosilífero 2. Fragmento de dentario izquierdo con las porciones intraalveolares de m1 y m2 (CRILAR-PZ 13) del sitio 6 y nivel fosilífero 2. Maxilar derecho con la serie P3-M3 (CRILAR-PZ 69), procedente del sitio 4 y nivel fosilífero 2.
Descripción. El fragmento de premaxilar tiene implantadas las porciones intraalveolares de los tres incisivos que se encuentran dispuestos en una serie continua y parte del canino, que está ligeramente separado del I3 por un diastema muy pequeño. En los I2-I3 se observa una gruesa capa de esmalte sólo en la cara labial. Estas mismas se hallan dispuestas en forma alineada, por lo que se interpreta que formaban un solo borde continuo como sucede en Protypotherium australe Ameghino, 1887 (Tauber, 1996). El diámetro mesio distal es similar en los elementos mencionados. A juzgar por la porción conservada del canino, éste habría tenido una forma similar a la de los incisivos, como ocurre en Protypotherium australe .
Observación. Un astrágalo derecho CRILAR-PZ 38b, con rasgos que permiten asignarlo a un Interatheriinae como Protypotherium , fue hallado junto a CRILAR-PZ 38a, siendo altamente probable que represente al mismo espécimen.
Familia M ESOTHERIIDAE Alston, 1876
Subfamilia M ESOTHERIINAE Simpson, 1945
Género Pseudotypotherium Ameghino, 1904
Pseudotypotherium sp.
Figura 5.A-C
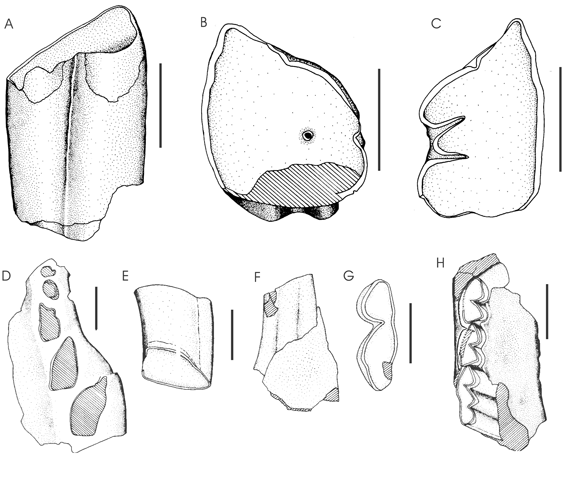
Figura 5. A-C, Pseudotypotherium sp. ; A, porción del P4 izquierdo desde la superficie oclusal hasta la parte media del diente en vista lingual (CRILAR-PZ 55) / left P4 portion from oclusal surface to medial part of teeth in lingual view. B, sección en la parte media del diente / section in middle part of teeth . C, en vista oclusal / in oclusal view. D-G, Hemihegetotherium cf. torresi ; D, maxilar izquierdo con las porciones intraalveolares de C-P4 (CRILAR-PZ 16) / left maxilar with C-P4. E, P3 derecho en vista oclusal y lingual (CRILAR 37) / right P3 in oclusal and labial view. F, m3 izquierdo en vista lingual (CRILAR-PZ 6) / left m3, lingual view. G, en vista oclusal / in oclusal view. H, cf. Tremacyllus sp., parte del dentario derecho con m1-m3 (CRILAR-PZ 57) / right dentary fragment with m1-m3. Las escalas representan 0,5 cm. Scale bars equal 0,5 cm.
Material y procedencia. Porción del P4 izquierdo desde la superficie oclusal hasta la parte media del diente (CRILAR-PZ 55). El material procede del sitio 2 y del nivel fosilífero 1 (28º 20' 527 S y 66º 57' 323 O). Gran porción del cráneo y mandíbula articulados con todos los elementos dentales (CRILAR-PZ 40), este material procede del sitio 1 y del nivel fosilífero 3 (28º 20' 767 S y 66º 57' 575 O).
Descripción. El espécimen CRILAR-PZ 55 tiene los diámetros mesio distal y tranversal constantes a lo largo del diente, sin embargo este diente posee una morfología que varía desde la superfice oclusal hacia la parte media del mismo. La sección tiene un contorno subcuadrangular en el extremo oclusal, haciéndose subtriangular hacia la parte media. En la cara lingual hay un surco longitudinal bien visible, ancho y profundo, estando su fondo cubierto por cemento. Sobre la superficie oclusal se observa que el fondo del surco lingual se halla bifurcado, esbozando un pequeño lóbulo mediano rudimentario, contrariamente a esto en la parte media del diente se observa un repliegue superficial de la capa de esmalte.
Observación. La existencia de un surco lingual en el P4 es el principal rasgo de Pseudotypotherium que permite distinguirlo claramente de Typotheriopsis (Pascual et al ., 1966; Cerdeño y Montalvo, 2001). La bifurcación interna del repliegue de esmalte lingual es un rasgo menos frecuente pero ha sido señalado previamente para Pseudotypotherium (Francis, 1965; Cerdeño y Montalvo, 2001). El bajo grado de imbricación entre los molares superiores, la posicón bien oblicua de los incisivos superiores con respecto al plano sagital son algunos de los rasgos que contribuyeron a determinar a estos restos como Pseudotypotherium sp.
Familia H EGETOTHERIIDAE Ameghino, 1894
Subfamilia H EGETOTHERIINAE Ameghino, 1894
Género Hemihegetotherium Rovereto, 1914
Hemihegetotherium cf. Torresi (Cabrera y Kraglievich, 1931) Zetti, 1972a
Figura 5.D-G
Material y procedencia. Fragmento de maxilar izquierdo con las porciones intraalveolares de C-P4 (CRILAR-PZ 16) y P3 derecho aislado (CRILAR 37), procedentes del sitio 7 y del nivel fosilífero 2. P3 ó P4 derecho aislado (CRILAR 36) y m3 izquierdo (CRILARPZ 6), procedentes del sitio 6 y del nivel fosilífero 2.
Observación. Cabrera y Kraglievich (1931) describieron un hegetotherino del Arroyo Chasicó, provincia de Buenos Aires (Formación Arroyo Chasicó) con el nombre Pseudohegetotherium torresi . Posteriormente este género fue considerado sinónimo de Hemihegetotherium , produciéndose una nueva combinación: H. torresi (Zetti, 1972a; Bond y López, 1997; Cerdeño y Contreras, 2000; Cerdeño, 2000; Cerdeño y Montalvo, 2002). En el último trabajo citado propusieron una sinonimia entre las diferentes especies de Hemihegetotherium , reconociendo solamente a H . Torresi de edad Chasiquense y H . achataleptum Rovereto, 1914 de edad Huayqueriense. Ambos taxones se diferencian por el mayor desarrollo de la quilla distal de los metápodos y por la talla mayor en H. achataleptum .
El material fue comparado con Hemihegetotherium torresi de la Formación Arroyo Chasicó, provincia de Buenos Aires y los especímenes CRILAR-PZ 16, 36 y 37 tienen un tamaño levemente menor, por lo que la asignación a esta especie es dudosa. El espécimen CRILAR-PZ 6, es indiferenciable en forma y tamaño a H. torresi , por lo que se considera que estas diferencias de tamaño podrían ser meras variaciones intraespecíficas, siendo muy fragmentario el material para una determinación más precisa.
Medidas (en mm). CRILAR-PZ 16: longitud de la serie P1-P4: 21,94; longitud cara labial de P3: 6,7; diámetro mesio distal del P2: 5,6; CRILAR-PZ 36: longitud de la cara labial: 7,06; 7,26 en la base; CRILAR-PZ 37: longitud de la cara labial: 7,2.
Subfamilia P ACHYRUKHINAE Kraglievich, 1934
Género Tremacyllus Ameghino, 1891 cf. Tremacyllus sp.
Figura 5.H
Material y procedencia. Porción anterior de una mandíbula con i1 y p3-p4? derechos, e i1-i2 izquierdos (CRILAR-PZ 44). Fragmento mandibular con m1-m2 derecho (CRILAR-PZ 45). Parte del dentario derecho con p3-p4? (CRILAR-PZ 49). Sínfisis mandibular con i1-i2 de ambos lados (CRILAR-PZ 56 a) y astrágalo derecho (CRLIAR-PZ 56 b), todos estos materiales proceden del sitio 4 y del nivel fosilífero 2. Fragmento mandibular derecho con parte de m1 (CRILAR-PZ 9 a). Parte del dentario derecho con m1-m3 (CRILAR-PZ 57), procedente del sitio 6, nivel fosilífero 2. Un fragmento de dentario con parte de dos molares (CRILAR-PZ 6 a) del sitio 2 y nivel fosilífero 1.
Observación. En la provincia de Catamarca (Mioceno tardío) se ha registrado la presencia de dos Pachyrukhinae: Paedotherium minor Cabrera, 1937 (Formación Chiquimil =Loro Huasi) y Tremacyllus incipiens Rovereto, 1914 (= T. latifrons , según Cerdeño y Bond, 1998; Formación Andalhuala). Los materiales aquí estudiados son fragmentarios; sin embargo se observan ciertos caracteres que permiten determinarlos como cf. Tremacyllus sp. Estos elementos son el tamaño pequeño y la gracilidad, la escasa longitud de la sínifisis mandibular con respecto a Paedotherium y la imbricación y longitud proporcionalmente corta de p3-p4? en el espécimen CRILAR-PZ 49.
Para la asignación a cf. Tremacyllus se ha considerado la imbricación entre los premolares ligeramente mayor y la menor longitud proporcional de la sínfisis mandibular comparado con Paedotherium .
Discusión
Edad de la Formación Salicas
Esta unidad litoestratigráfica fue asignada, con dudas, al Plioceno (Sosic, 1973; Combina y Santiesteban Bové, 2000a) o al Plioceno superior (Turner, 1971), sobre la base de las relaciones estratigráficas dentro de un marco regional. Posteriormente Mulé et al . (1997) proponen una edad Chasiquense-Huayqueriense (Mioceno Superior sic) para la Formación Salicas, debido al hallazgo de restos determinados como Typotheriopsis sp. y Pachyrukhinae indet. Según estos autores, la presencia de edentados permite restringir la edad al Huayqueriense, aunque no citan ningún taxón perteneciente a este grupo de mamíferos.
Bossi et al . (1999) citaron la presencia de cf. Tremacyllus , Mesotheriidae indet. y Pachyrukhinae indet., procedentes de la vertiente oriental de la sierra de Ambato, cerca de la localidad de San Pedro. Sobre la base de estos fósiles se atribuyó una probable edad Huayqueriense para la Formación Salicas, aunque en el cuadro 1 se la refiere al Mioceno y Plioceno y en la Figura 10a del mismo trabajo se la asigna al Plioceno (Bossi et al ., 1999 p. 356). No obstante, en ningún caso fueron citados los materiales fósiles, repositorios, procedencias geográficas ni estatigráficas precisas que sustenten las determinaciones de estas edades. Por este motivo las evidencias paleontológicas disponibles hasta el momento son insuficientes para determinar con precisión y fundamentar la edad de los sedimentos portadores. Además, el registro de los taxones citados por Mulé et al. (1997) y Bossi et al . (1999) abarca edades que van desde el Mioceno tardío (Chasiquense) hasta el Plioceno tardío (Marplatense), ya que Typotheriopsis se registró durante el Chasiquense y Huayqueriense, y Tremacyllus durante el Chasiquense-Marplatense (Cione y Tonni, 1995; Cerdeño y Bond, 1998; Cerdeño, 2000; Cerdeño y Montalvo, 2001).
Los términos Chasiquense y Huayqueriense merecen ser redefinidos mediante estudios bioestratigráficos precisos y sobre la base de taxones característicos de distribución bien conocida, de hallazgos frecuentes y que sean fácilmente reconocibles. Se han propuesto biozonas locales sobre la base de la presencia de Chasicotatus ameghinoi Scillato Yané (Dasipodidae, Eutatini), considerado característico del Miembro Las Barrancas de la Formación Chasicó o del Chasiquense superior (Tonni et al ., 1998; Cione et al ., 2000). No obstante, Esteban et al. (2001a, 2001b, 2003) lo registraron en la Formación Cerro Azul (Huayqueriense) de la provincia de La Pampa, dejando de ser un taxón guía del Chasiquense. En la región noroeste de Argentina esta especie fue registrada en los niveles medio y superior del Miembro El Jarillal de la Formación Chiquimil (=Miembro A), en la localidad Villavil, provincia de Catamarca (Esteban y Nasif, 1999). En estos casos los datos de procedencia estratigráfica se refieren al nivel de miembros o formación, resultando imprecisos para establecer una biozona. Algo similar ocurre con las otras especies propuestas como características del Chasiquense y Huayquerienses (Cione et al ., 2000).
En las distintas localidades en las que aflora la Formación Salicas se han registrado icnitas de invertebrados de agua dulce y de aves, diatomitas, troncos con trazas (galerías) de insectos xilófagos de la Familia Buprestidae (Tauber y Mazzoni, 2003), tallos y hojas de plantas acuáticas, restos óseos de quelonios, aves y mamíferos. Sin embargo, son estos últimos los que aportan datos de interés para la determinación de la edad de esta formación.
En la región noroeste de Argentina las unidades que comparten más taxones con los niveles prospectados de la Formación Salicas son las Formaciones Chiquimil y Andalhuala. En efecto, el Miembro Los Baños (= Chiquimil B) es portador de Hemihegetotherium ? sp. (= Pseudohegetotherium ? sp.) y en el suprayecente Miembro El Jarillal (= Chiquimil A) se registraron Neophanomys biplicatus , Lagostomus ( Lagostomopsis ) cf. pretrichotactyla , Orthomyctera andina , Potamarchus sp., Typotheriopsis sp., cf. Tremacyllus sp. (Marshall y Patterson, 1981; Powell et al ., 1998). A esta lista se agrega Proeuphractus limpidus registrada por Esteban et al. (2004), aunque sin especificar de qué miembro proceden los materiales.
El Miembro Los Baños de la Formación Chiquimil es considerado Chasiquense (Bossi et al ., 1998), aunque son muy escasos los mamíferos registrados en esta unidad.
La edad del Miembro El Jarillal es controversial, Simpson (1974) consideró que sus mamíferos son indiferenciables de aquellos del Araucanense, es decir Huayqueriense, aunque los taxones registrados entonces eran muy escasos. Powell et al . (1998) citaron especies consideradas exclusivamente huayquerienses, entre ellas Eosclerocalyptus planus y Orthomyctera andina (véase más adelante). Bossi et al ., (1998: cuadro 1, p. 89) incluyen al Miembro El Jarillal dentro del Huayqueriense, aunque en el mismo trabajo lo consideran de edad transicional entre el Chasiquense y el Huayqueriense (Bossi et al ., 1998: 107), sin que se hayan registrado especies consideradas exclusivas del Chasiquense, por lo que consideramos que esta última hipótesis no tiene sustento. De todos modos, no hay estudios bioestratigráficos de detalle del Miembro El Jarillal, se desconocen por el momento las asociaciones de taxones discriminadas por niveles. Además, aún deben definirse, adecuadamente, los fundamentos bioestratigráficos para distinguir entre las edades Chasiquense o Huayqueriense, cosa que hasta el presente no se ha logrado de manera satisfactoria. En cuanto a los aspectos geocronológicos, Bossi et al . (1987) consideran que el pasaje entre el Miembro El Jarillal y la Formación Andalhuala se produjo en los 7 Ma, sobre la base de dataciones radimétricas en tobas y magnetoestratigrafía asociada.
En la Formación Andalhuala de la provincia de Catamarca, especialmente en su sección media, se han registrado Neophanomys biplicatus (Unidad XVIIIa), Lagostomus ( Lagostomopsis ) cf. pretrichodactyla , Orthomyctera andina , Typotheriopsis [la especie T. internum (Ameghino) Cabrera] y Tremacyllus [la especie T. diminutus (Ameghino) Rovereto, hoy considerada sinónimo de T . impressus por Cerdeño y Bond, 1998] (Marshall y Patterson, 1981). La Formación Andalhuala es considerada de edad Huayqueriense (Bossi et al ., 1998).
Los materiales estudiados proceden en su mayoría del NF 2 con excepciones como Eosclerocalyptus planus del NF 4 (tabla 1). A continuación se analizarán la distribución estratrigráfica y geográfica de los taxones registrados en la Formación Salicas.

Tabla 1. Distribución estratigráfica de los taxones registrados en la Formación Salicas / Stratigraphic distribution of the taxa registered of the Salicas Formation.
Macrochorobates scalabrinii fue registrada en la Formación Epecuén, en una unidad inominada del área del arroyo Chasicó, en la provincia de Buenos Aires, en el Araucanense del valle de Santa María, en la localidad de Chiquimil o Entre Ríos (unidades XVII, XX, Araucanense sensu Stahlecker, 1935; Formación Andalhuala sensu Bossi et al ., 1987) y en el área de Hualfín y Belén, en Puerta de Corral Quemado (unidad 24, Corral Quemado sensu Stahlecker, 1935; Formación Andalhuala sensu Bossi et al ., 1987), estos últimos en la provincia de Catamarca (Riggs y Patterson, 1939; Marshall y Patterson, 1981; Scillato Yané, 1982; Tonni et al ., 1998; Esteban et al ., 2001a). Macrochorobates escalabrinii es considerada como una especie de edad Huayqueriense, habiendo sido propuesta para definir una biozona local de esta edad (Tonni et al ., 1998; Cione et al ., 2000; Esteban et al ., 2001a).
Proeuphractus fue registrado en el Miembro Las Barrancas de la Formación Arroyo Chasicó de la provincia de Buenos Aires (edad Chasiquense), en la Formación Cerro Azul de la provincia de La Pampa (edad Huayqueriense, Montalvo et al ., 1996; Esteban et al ., 2001a, 2003), en el Hermosense típico (Tonni et al ., 1992) y en la Formación Chiquimil (Esteban et al ., 2004), aunque se desconoce el nivel de procedencia de este último registro y su edad. La especie tipo P . limpidus procede de la Formación Ituzaingó, provincia de Entre Ríos (Mesopotamiense, Mioceno tardío, edad Huayqueriense?, Scillato Yané, 1980, 1982; Carlini y Scillato Yané, 1996; Cione et al ., 2000). Recientemente se la registró también en la Formación Toro Negro (Plioceno temprano, Montehermosense), en la sierra de Los Colorados, departamento Vinchina, (Rodríguez Brizuela, 2002), siendo estos los primeros hallazgos documentados en la provincia de La Rioja.
El género Chaetophractus está representado, al menos, por la especie C. villosus registrada desde el Chapadmalalense de la provincia de Buenos Aires hasta la actualidad (Cione y Tonni, 1995).
Eosclercalyptus planus fue registrada en la unidad XVII de la Formación Andalhuala (Riggs y Patterson, 1939; Marshall y Patterson, 1981) y en el Miembro El Jarillal de la Formación Chiquimil de Catamarca (Powell et al ., 1998). El género Eosclerocalyptus en la Formación Epecuén (edad Huayqueriense) de la provincia de Buenos Aires (Tonni et al ., 1992; Scillato Yané et al ., 1995). Estos autores también citan a Eosclerocalyptus sp. en el Montehermosense de Buenos Aires y Mendoza.
Verzi et al . (1999) citan a Neophanomys biplicatus en las provincias de Catamarca, Mendoza y La Pampa. Estos autores describieron materiales procedentes de las localidades Bajo de Giuliani, valle de Quehué y Toay, provincia de La Pampa y de la Formación Cerro Azul, considerada de edad Huayqueriense sensu lato (Verzi et al ., 1991). Además, se conocen restos de depósitos neógenos de la meseta del Guadal, San Carlos, provincia de Mendoza (Tunuyanense sensu Rovereto, 1914 o Formación Tunuyán sensu Dessanti, 1946), cuya edad se ha considerado transicional entre el Araucanense-Montehermosense Huayqueriense-Montehermosense?), Mioceno tardío-Plioceno temprano (Yrigoyen, 1994) (Araucanense sensu Riggs y Patterson, 1939).
En la región noroeste de Argentina, el registro de Neophanomys biplicatus que cuenta con datos estratigráficos más precisos, es de la unidad XVIIIa del perfil de Stahlecker (1935) publicado por Riggs y Patterson (1939) y Marshall y Patterson (1981). Este hallazgo se produjo en la localidad de Chiquimil (=Entre Ríos), Valle de Santa María, Catamarca, por debajo de una toba datada en 6,2 Ma (Marshall y Patterson, 1981). El nivel portador está incluido en la sección media de la Formación Andalhuala (Huayqueriense). Nasif y Esteban (2000) comunicaron el hallazgo de una hemimandíbula en la localidad de Villavil, valle de Hualfín, provincia de Catamarca. El nivel portador está incluido en el Miembro El Jarillal, el que representa la parte superior de la Formación Chiquimil. Según las últimas autoras, este es el registro más antiguo de N. biplicatus , sin embargo se desconoce hasta el presente la posición estratigráfica precisa.
En síntesis la distribución de Neophanomys biplicatus abarcaría desde el Huayqueriense hasta el Montehermosense (Montalvo et al ., 1999; Verzi et al ., 1999), no obstante, es necesario contar con nuevos elementos, debido a que los materiales existentes con datos estratigráficos precisos son escasos.
Los Chinchillidos del subgénero Lagostomus ( Lagostomopsis ) se registran en la región pampeana con dudas desde la edad Chasiquense (Barranquense o Miembro Las Barrancas) hasta el Chapadmalalense (Cione y Tonni, 1995) y la especie L. ( L .) pretrichodactyla es considerada de edad Huayqueriense. Algo similar ocurre con el género Orthomyctera que se registró desde el Chasiquense hasta el Chapadmalalense (Cione y Tonni, 1995) o Ensenadense (Vucetich y Verzi, 1995); sin embargo, la especie Orthomyctera andina se registró en la Formación Andalhuala (unidades XVII, XVIIIb y XVIIIa de Stahlecker, 1935) y se asigna al Huayqueriense.
Los restos determinados como Potamarchus sp., son los primeros registros de la Familia Dinomyidae hallados en la provincia de La Rioja. Este género se registró en el conglomerado osífero de la Formación Ituzaingó (Huayqueriense?, Burmeister, 1885; Cione et al ., 2000). En Catamarca Powell et al . (1998) citaron este género para el Miembro El Jarillal (= Chiquimil A), sin mencionar niveles ni materiales que permitan verificar la determinación. En la región pampeana se lo registró en la Formación Chasicó y en la Formación Epecuén (Huayqueriense) de la provincia de Buenos Aires, aunque estos últimos registros se consideran dudosos en la actualidad (Cione et al ., 2000).
El género Protypotherium está registrado desde el Colhuehuapense y tiene sus últimos registros en el Miembro Vivero de la Formación Arroyo Chasicó, provincia de Buenos Aires (Chasiquense, Cabrera y Kraglievich, 1931; Cione y Tonni, 1995; Bond y López, 1996, 1998) y en Formación Ituzaingó. Además, se han recuperado restos con signos de desgaste por transporte postmortem, de las arenas denominadas Puelchense o Formación Puelche, entre ellos el holotipo de Notopachyrukhos tambuttoi Rusconi (1933, 1948, 1949), considerado sinónimo de Protypotherium (Bond y López, 1998).
En la actualidad se considera que la Formación Puelche es portadora de una mezcla de fósiles de niveles equivalentes al Mesopotamiense, hasta del Pleistoceno tardío-Holoceno temprano (Lujanense). De la provincia de Catamarca fue citado un Interatheriidae en una lista taxonómica por Marshall y Patterson (1981: 66). Sin embargo, estos autores no mencionaron ningún material, procedencia estratigráfica ni geográfica que sustente esta cita. Estos nuevos elementos fósiles permiten verificar la presencia de Interatheriidae en la región noroeste de Argentina y atribuirlos al género Protypotherium .

Tabla 2. Distribución de los mamíferos de la Formación Salicas, provincia de La Rioja / Distribution of the mammals of the Salicas Formation, La Rioja province. Edad Mamífero: CH: Chasiquense, HU: Huayqueriense, MO: Montehermosense, Mp: Mesopotamiense, CHA: Chapadmalalense, MA: Marplatense, EN: Ensenadense, LU: Lujanense. 1, El Género solamente, 2, El Subgénero solamente, 3, L. ( L .) pretrichodactyla , 4, H. torresi , 5, H . achataleptum / Land Mamad Age: CH: Chasicoan, HU: Huayquerian, MO: Montehermosan, Mp: Mesopotamiense, CHA: Chapadmalalan, MA: Marplatan, EN: Ensenadan, LU: Lujanian. 1, Only the genus, 2, Only the subgenus, 3, L. ( L .) pretrichodactyla , 4, H. torresi , 5, H. achataleptum .
En cuanto a los Mesotheriidae, este es el primer registro seguro de Pseudotypotherium en la región noroeste de Argentina, ya que Villarroel (1974) ilustra en un mapa la distribución de este género, incluyendo Catamarca, sin mencionar materiales. Cerdeño y Montalvo (2001) estudiando los Mesotheriinae de la Formación Cerro Azul, verificaron la presencia, en la misma unidad litoestratigráfica, de Pseudotypotherium subinsigne (Rovereto, 1914) y de Typotheriopsis silveyrai Cabrera (1937). Este es el registro más antiguo del género Pseudotypotherium y el más moderno es del Plioceno tardío.
Hemihegetotherium cf. torresi es comparable a una especie originalmente denominada Pseudohegetoherium torresi , la que ha sido registrada en el Miembro Las Barrancas de la Formación Arroyo Chasicó (Bondesio et al ., 1980a; Marshall et al ., 1983; Cione y Tonni, 1995). En el Miembro B de la Formación Chiquimil se citó a Pseudohegetotherium sp. (Riggs y Patterson, 1939; Marshall y Patterson, 1981; Bossi et al ., 1998). La especie de mayor tamaño, H. Achataleptum es de edad Huayqueriense (Cerdeño y Contreras, 2000; Cerdeño y Montalvo, 2002).
El género Tremacyllus ha sido registrado desde el Mioceno tardío (Chasiquense) hasta el Pleistoceno temprano (Marplatense) según Cerdeño y Bond, (1998), de tal modo que en este caso no aporta datos de valor para determinar la edad de la Formación Salicas. Estos hallazgos verifican una vez más la observación de Zetti (1972b) y Cerdeño y Bond (1998), acerca del predominio de Tremacyllus sobre Paedotherium en la región noroeste de Argentina.
Es necesario profundizar los estudios estrictamente bioestratigráficos del Mioceno-Plioceno con la finalidad de realizar nuevas propuestas de biozonas, teniendo en cuenta los aspectos tafonómicos del registro y sedimentológicos de los yacimientos. En la tabla 1 se sintetiza la distribución aceptada de los taxones registrados en la Formación Salicas según el sistema de edades del Cenozoico continental de América del Sur. La asociación de taxones registrados en esta unidad está indicando una edad Mioceno tardío para los niveles prospectados (edad Huayqueriense). De todas las especies sólo Hemihegetotherium torresi es exclusiva de la edad Chasiquense tardío, sin embargo los materiales de la Formación Salicas fueron determinados como H. cf. torresi , mientras que Macrochorobates scalabrinii , Eosclerocalyptus planus y Orthomyctera andina , serían exclusivos de la edad Huayqueriense. Los taxones Neophanomys biplicatus y Pseudotypotherium sp., tienen su primera aparición en el Huayqueriense. En síntesis, la mayoría de las evidencias indican que la hipótesis más plausible sobre la edad de los niveles prospectados (especialmente el NF 2) de la Formación Salicas es Mioceno tardío, a priori Huayqueriense, aunque este último término debería ser redefinido. Una edad Montehermosense para estos niveles es menos probable debido a la presencia de Macrochorobates scalabrinii , Eosclerocalyptus planus , Potamarchus sp., Protypotherium sp. y Hemihegetotherium cf. torresi .
La presencia de Protypotherium sp. durante el Mioceno tardío en el noroeste de Argentina
Se ha interpretado una edad Huayqueriense para los niveles fosilíferos estudiados, sin embargo, es necesario precisar este término ya que se observa una asociación de taxones diferente a las registradas en la región pampeana. En efecto, la presencia de Protypotherium sp. en los niveles prospectados de la Formación Salicas, asociado a los demás taxones citados (de edades Huayqueriense y más modernas), podría explicarse, hipotéticamente, por una probable retracción en dirección latitudinal, desde la región pampeana hacia el norte durante el Mioceno tardío o incluso desde la Patagonia austral a partir del Mioceno temprano a medio. Algunos casos similares estarían ocurriendo con la presencia de Thylatheridium dolgopolae (Huayqueriense de la región pampeana, Goin et al ., 2000) y Xyophorus bondesioi Scillato Yané (Chasiquense), ambas especies presentes en el Miembro Inferior de la Formación Toro Negro del Plioceno temprano, en el departamento Vinchina, provincia de La Rioja (Rodríguez Brizuela, 2002).
En el caso de Protypotherium sp., la retracción latitudinal es una interpretación compatible con los datos conocidos de la distribución del género, sobre todo de los últimos registros analizados por cada región. En el extremo austral de Patagonia el nivel más alto que se ha documentado con P. australe es el techo del Miembro Estancia La Costa, de la Formación Santa Cruz (Mioceno inferior-medio, Tauber, 1996, 1997). En la región noroccidental de Patagonia el registro más moderno es en la Formación Collón Curá (Colloncurense) ( e.g . Bondesio et al ., 1980b; Rolleri et al. , 1984a y 1984b). El material MLP 79-XII-17-1 determinado como P . praerutilum (Rolleri et al ., 1984b), si bien es fragmentario, en realidad posee características de un espécimen juvenil de P . australe , según los caracteres considerados anteriormente (Tauber, 1996). En el sector sur de la región pampeana los últimos registros de Protypotherium sp. son del miembro inferior (Miembro Vivero) de la Formación Chasicó, donde el género está bien representado. En la región mesopotámica la única especie registrada es Protypotherium altum Ameghino, 1887 y los materiales escasos y fragmentarios que representan a este taxón proceden del conglomerado osífero de la Formación Ituzaingó (Mioceno tardío, Huayqueriense?, Cione et al ., 2000).
Una hipótesis alternativa aunque poco probable es que esta asociación podría deberse a un problema tafonómico, que los materiales atribuidos en este trabajo a Protypotherium sp. procedan de niveles más antiguos. Sin embargo, el estrato de donde proceden los materiales fósiles se encuentra en la sección inferior de la Formación Salicas. Esta unidad yace, en está región, sobre el basamento compuesto por el granitoide porfírico milonitizado de San Blas o sobre depósitos del Cretácico tardío en la parte central y sudoriental de la sierra de Velasco ( vide supra ). Por lo tanto, resulta poco probable que esta asociación se deba a una mezcla por procesos tafonómicos.
La falta de estudios bioestratigráficos y tafonómicos de detalle ha imposibilitado analizar estas diferencias en las asociaciones de especies entre la región pampeana y la parte norte de Argentina. Esto aportaría importantes evidencias para reinterpretar hipótesis de correlaciones confusas entre unidades con una diversidad de géneros y especies supuestamente de diferentes edades. Este es el caso de la Formación Ituzaingó (Mesopotamiense, provincia de Entre Ríos, Cione et al ., 2000), portadora de una asociación de mamíferos fósiles que fue considerada como una probable mezcla de materiales de diferentes edades, desde Santacrucense a Montehermosense (Bianchini y Bianchini, 1971; Scillato Yané, 1977, 1980b, 1981; Marshall et al ., 1983), debido a procesos tafonómicos o mezclas artificiales. Al respecto la hipótesis planteada de una retracción latitudinal hacia el norte al menos durante el Mioceno tardío, explicaría la presencia del género Protypotherium en la Formación Ituzaingó (Mesopotamiense, edad Huayqueriense?).
Estas diferencias en las asociaciones de mamíferos fósiles ponen en evidencia la necesidad de definir correctamente unidades bioestratigráficas (biozonas) como sustento para realizar correlaciones más confiables. El establecimiento de un sistema de correlación basado sobre la correcta definición de biozonas requiere la selección de los taxones característicos más adecuados, que tengan una amplia distribución geográfica y corta distribución estratigráfica, que sean frecuentes y estén bien fundamentados taxonómicamente. En tal sentido se destaca Neophanomys biplicatus como uno de los taxones que tiene una buena distribución geográfica, una distribución estratigráfica mejor estudiada con relación a los demás taxones y es fácilmente reconocible. Esta especie podría ser utilizada, cuando se completen los estudios, para establecer una Biozona de Asociación con Macrochorobates scalabrinii , Eosclerocalyptus planus , Potamarchus o Hemihegetotherium .
La Formación Salicas en el contexto regional
La asignación de esta unidad a la parte final del Mioceno tardío tiene un significado especial para la geología regional en el área de la Sierra de Velasco. En la localidad de Tanín, departamento Capital, situada al pie de la vertiente oriental de la sierra de Velasco, unos 37 km al Sur de la ciudad de La Rioja, se pueden abservar algunos afloramientos dispersos, en gran medida cubiertos por depósitos más modernos o por vegetación, de sedimentos asignables a la Formación Salicas. Estos depósitos se encuentran cubriendo mediante una discordancia a niveles de arenisca conglomerádica de color gris que contienen un alto contenido de estructuras calcáreas dispersas de color rosado. Estas areniscas fueron atribuidas por Bodenbender (1911) a los Estratos de Los Llanos o Formación Los Llanos (Zuzek, 1978). Esta última unidad fue referida por Bodenbender (1911) al Cretácico Superior? Con dudas y más tarde otros autores la asignaron al Terciario: al Mioceno medio por Rusconi (1936), al Plioceno por Pascual et al . (1966), al Chasiquense por Bordas (1941), Pascual (1954) y Pascual y Bondesio (1981). El hallazgo de restos de huevos de dinosaurios saurópodos en el valle de Sanagasta en niveles correlacionados con los de los Estratos de Los Llanos ha permitido retomar la hipótesis de Bodenbender sobre la edad de estos depósitos (Hunicken et al ., 2001, Tauber, 2002). El hecho de que los niveles de la Formación Salicas se encuentren sobre la Formación Los Llanos en la localidad de Tanín confirma al menos una edad previa al Mioceno tardío para la última unidad o al menos para los afloramientos de Tanín.
Conclusiones
Los fósiles estudiados indican que los estratos prospectados de la Formación Salicas se depositaron durante la parte final del Mioceno tardío (Huayqueriense). En la Formación Salicas se ha observado una asociación de taxones diferente a las de unidades coetáneas de la región pampeana, poniendo en evidencia la necesidad de lograr definiciones precisas de la base bioestratigráfica que sustente las diferentes edades del Mioceno tardío.
Se ha verificado la presencia de representantes de la Familia Interatheriidae en la región noroeste de Argentina, específicamente del género Protypotherium de amplia distribución en la Patagonia. Se propone como hipótesis una retracción esencialmente latitudinal de este género, desde la región pampeana hacia el norte durante el Mioceno tardío.
Se correlacionan a los estratos estudiados de la Formación Salicas con los niveles altos de la Formación Chiquimil, Miembro El Jarillal (Chiquimil A) y los niveles bajos de la Formación Andalhuala de la provincia de Catamarca.
Se confirma una edad previa miocena tardía para la Formación Los Llanos al menos en los afloramientos de la localidad de Tanín, provincia de La Rioja.
Macrochorobates scalabrinii , Protypotherium sp. Y Pseudotypotherium sp. en la Formación Salicas se suman al conjunto de mamíferos que permitieron proponer la existencia de una conexión biogeográfica de la región noroeste de Argentina, principalmente con la región pampeana (Scillato Yané, 1975).
Agradecimientos
Este trabajo es un homenaje póstumo a la memoria de M. L. Romero de Tauber quien subsidió de manera anónima gran parte de estas y otras investigaciones del autor. El autor agradece a M. Hünicken, A. Sobral y J. Di Ronco por su colaboración en las campañas, tanto en la recolección de fósiles como en el relevantamiento de datos para la columna estratigráfica con brújula taquimétrica. A. Mazzoni colaboró con la redacción del manuscrito. Por los comentarios al manuscrito se agradece a los árbitros G. Esteban y S. Marensi. Este trabajo fue realizado en el Centro Regional de Investigación Científica y Transferencia Tecnológica (CRILAR), Villa de Anillaco, La Rioja, en la Agencia de Cultura, La Rioja, Argentina y en el Museo de Paleontología de la Universidad Nacional de Córdoba.
Bibliografía
Aceñolaza, F.G. y Bortolotti, P. 1981. Estratigrafía y evolución tectónica de la quebrada de La Rioja. 1º Simposio de Geología de La Rioja. Acta Geológica Lilloana 15: 30-39. [ Links ]
Ameghino, C. 1919. Sobre mamíferos fósiles del piso araucanense de Catamarca y Tucumán. 1º Reunión Nacional de la Sociedad Argentina de Ciencias Naturales , (San Miguel de Tucumán): 150-153. [ Links ]
Ameghino, F. 1886. Contribución al conocimiento de los mamíferos fósiles de los terrenos terciarios antiguos de Paraná. Boletín de la Academia Nacional de Ciencias (Córdoba) 9: 3-226. [ Links ]
Ameghino, F. 1887. Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de la Patagonia austral. En: A.J. Torcelli (ed.), Ameghino. Obras completas y correspondencia científica 5: 447-468. [ Links ]
Ameghino, F. 1891. Mamíferos y aves fósiles argentinos: especies nuevas. Adiciones y correcciones. Revista Argentina de Historia Natural 1: 240-259. [ Links ]
Ameghino, F. 1904. Nuevas especies de mamíferos cretáceos terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 56: 162-175. [ Links ]
Amos, A. y Zardini, R. 1962. Geología de algunos depósitos de arcilla de La Rioja. Revista de la Asociación Geológica Argentina 17: 47-84. [ Links ]
Báez, M.A., Rossi de Toselli, J.N. y Sardi, F. 2002. Consideraciones preliminares sobre granitoides del norte de la sierra de Velasco, La Rioja, Argentina. 15º Congreso Geológico Argentino (El Calafate), Actas 2: 69-74. [ Links ]
Bianchini, L.H. Delupi de y Bianchini, J.J. 1971. Revisión de los Proterotheriinae (Mammalia, Litopterna) del Mesopotamiense. Ameghiniana 8: 1-24. [ Links ]
Bodenbender, G. 1911. Constitución geológica de la parte meridional de La Rioja y regiones limítrofes, República Argentina. Boletín de la Academia Nacional de Ciencias (Córdoba) 19: 5-221. [ Links ]
Bond, M. y López, G. 1996. Las especies de Protypotherium sp. (Notoungulata, Interatheriidae) de la Formación Arroyo Chasicó (Mioceno superior), Provincia de Buenos Aires, Argentina. Ameghiniana 33: 460. [ Links ]
Bond, M. y López, G. 1997. Los Hegetotheriinae (Notoungulata, Hegetotheriidae) de la Formación Arroyo Chasicó (Mioceno superior), Provincia de Buenos Aires, Argentina. Ameghiniana 34: 533. [ Links ]
Bond, M. y López, G.M. 1998. Notopachyrukhos tambuttoi Rusconi, 1933, un Interatheriidae (Mammalia, Notoungulata) del Puelchense de la provincia de Buenos Aires, Argentina. 10º Congreso Latinoamericano de Geología y 6º Congreso Nacional de Geología Económica (Buenos Aires), Actas 1: 280-283. [ Links ]
Bondesio, P., Laza, J.H., Scillato Yané, G.J., Tonni, E.P. y Vucetich, M.G. 1980a. Estado actual del conocimiento de los vertebrados de la Formación Arroyo Chasicó (Plioceno temprano) de la provincia de Buenos Aires. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires), Actas 3: 101-127. [ Links ]
Bondesio, P., Rabassa, J., Pascual, R., Vucetich, M.G. y Scillato Yané, G.J. 1980b. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano (Buenos Aires), Actas 3: 85-99. [ Links ]
Bordas, A.F. 1941. Nuevos restos fósiles de la Formación de Los Llanos (San Luis). Physis 19: 23-27. [ Links ]
Bossi, G.E., Ovejero, R. y Strecker, M. 1987. Correlación entre los perfiles del Terciario superior en la Puerta de Corral Quemado-Hualfín y de Entre Ríos (Chiquimil) provincia de Catamarca, Argentina. 10º Congreso Geológico Argentino (San Miguel de Tucumán), Actas 2: 117-120. [ Links ]
Bossi, G.E., Gavriloff, I.J.C. y Esteban, G. 1998. Terciario (Estratigafía, Bioestratigrafía y Paleontología). En: M. Gianfrancisco, M.E. Puchulu, J. Durango de Cabrera y G.E. Aceñolaza (eds.), Geología de Tucumán. Colegio de Graduados en Ciencias Geológicas de Tucumán, Publicación Especial : 87-110. [ Links ]
Bossi, G.E., Muruaga, C.M. y Gavriloff, I.J.C. 1999. Sierras Pampeanas. En: G. González Bonorino, R. Omarini y J. Viramonte (eds.), 14º Congreso Geológico Argentino (Salta), Geología del Noroeste Argentino, Relatorio : 329-360. [ Links ]
Burmeister, G. 1885. Examen crítico de los mamíferos y los reptiles denominados por Don Augusto Bravard. Annales del Museo Público de Buenos Aires 3: 95-173. [ Links ]
Cabrera, A. 1937. Notas sobre el suborden Typotheria. Notas del Museo de La Plata 2: Paleontología 8: 17-43. [ Links ]
Cabrera, A. 1944. Los Gliptodontoideos del Araucaniano de Catamarca. Revista del Museo de La Plata, ( nueva serie ) 15, Paleontología 3: 5-76. [ Links ]
Cabrera, A. y Kraglievich, L. 1931. Diagnosis previas de los ungulados fósiles del Arroyo Chasicó. Notas del Museo de La Plata 1: 107-113. [ Links ]
Carignano, C., Cioccale, M. y Rabassa, J. 1999. Landscape Antiquity of the Central-Eastern Sierras Pampeanas (Argentina): Geomorphological Evolution since Gondwanic Times. Zeitschrift für Geomorphologie, Annals of Geomorpholog y, Suppl.-Bd. 118: 245-268. [ Links ]
Carlini, A.A. y Scillato Yané, G.J. 1996. Chorobates recens (Xenarthra, Dasypodidae) y un análisis de la filogenia de los Euphractini. Revista del Museo de La Plata ( nueva serie ) 9: 225-238. [ Links ]
Castellanos, A. 1940. A propósito de los géneros Plohophorus , Nopachthus y Panochthus (1º Parte). Publicaciones de Instituto de Fisiografía y Geología, Universidad Nacional del Litoral 6: 1-280. [ Links ]
Cerdeño, E. 2000. Los mamíferos de la Formación Arroyo Chasicó (Mioceno superior), provincia de Buenos Aires, del Museo J. C. Moyano de Mendoza. Ameghiniana 37: 503-508 [ Links ]
Cerdeño. E. y Bond, M. 1998. Taxonomic revision and phylogeny of Paedotherium and Trenacyllus (Pachyrukhinae, Hegetotheriidae, Notoungulata) from the Late Miocene to the Pleistocene of Argentina. Journal of Vertebrate Paleontology 18: 799-811. [ Links ]
Cerdeño, E. y Contreras, V.H. 2000. El esqueleto postcraneal de Hemihegetotherium (Hegetotheriidae, Notoungulata) del Mioceno superior de Puchuzum, San Juan, Argentina. Revista Española de Paleontología 15: 171-179. [ Links ]
Cerdeño, E. y Montalvo, C.I. 2001. Los Mesotheriinae (Mesotheriidae, Notoungulata) del Mioceno superior de La Pampa, Argentina. Revista Española de Paleontología 16: 63-75. [ Links ]
Cerdeño, E. y Montalvo, C.I. 2002. Los Hegetotheriinae (Hegetotheriidae, Notoungulata) del Mioceno superior de la provincia de La Pampa, Argentina. Revista del Museo Argentino de Ciencias Naturales, n. s. 4: 35-43. [ Links ]
Cione, A.L., Azpelicueta, M.M., Bond, M., Carlini, A.A., Casciotta, J., Cozzuol, M., de la Fuente, M., Gasparini, Z., Goin, F.J., Noriega, J., Scillato-Yané, G.J., Soibelzon, L., Tonni, E.P., Verzi, D. y Vucetich, M.G. 2000. Miocene vertebrates form Entre Ríos, eastern Argentina. En: E.G. Aceñolaza y R. Herbst (eds.), El Mio-Plioceno Argentino, INSUGEO, Serie Correlación Geológica 14: 191-237. [ Links ]
Cione, A. y Tonni, E.P. 1995. Chronostratigraphy and Land-Mammal Ages in the Cenozoic of southern South America: principles, practices, and the Uquian problem. Journal of Paleontology 69: 135-159. [ Links ]
Combina, A. y Santiesteban Bové, C. 2000a. Descripción de las unidades estratigráficas de los abanicos cuaternarios de la Sierra de Velasco-La Rioja-Argentina. 2º Congreso Latinoamericano de Sedimentología y 8º Reunión Argentina de Sedimentología (Mar del Plata), Resúmenes : 65-66. [ Links ]
Combina, A. y Santiesteban Bové, C. 2000b. Interpretación tectosedimentaria de los abanicos cuaternarios de la Sierra de Velasco-La Rioja-Argentina. 2º Congreso Latinoamericano de Sedimentología y 8º Reunión Argentina de Sedimentología (Mar del Plata), Resúmenes : 66-67. [ Links ]
De Alba, E. 1979. Descripción geológica de la Hoja 16 d Chilecito, provincia de La Rioja. Servicio Geológico Nacional, Boletín 163: 1-87. [ Links ]
Dessanti, R.N. 1946. Hallazgo de depósitos glaciares en las Huayquerías de San Carlos (Mendoza). Revista de la Asociación Geológica Argentina 1: 270-284. [ Links ]
Esteban, G. y Nasif, N. 1999. Mamíferos fósiles de la Formación Chiquimil (Mioceno tardío), provincia de Catamarca, Argentina. Bioestratigrafía. Ameghiniana , Suplemento Resúmenes 36: 11R. [ Links ]
Esteban, G.I., Nasif, N.L. y Montalvo, C.I. 2001a. Nuevos registros de Dasypodidae (Xenarthra) del Mioceno tardío de la provincia de La Pampa, Argentina. Revista Española de Paleontología 16: 77-87. [ Links ]
Esteban, G.I., Nasif, N.L., Montalvo, C.I. y Visconti, G. 2001b. Nuevos registros de Dasypodidae (Xenarthra) para el Terciario tardío de la provincia de La Pampa (Departamento Caleufú). Ameghiniana , Suplemento Resúmenes 38: 8R. [ Links ]
Esteban, G.I., Nasif, N.L., Montalvo, C.I. y Visconti, G. 2003. Nuevos registros de Dasypodidae (Xenarthra) en la Formación Cerro Azul (Mioceno tardío) de Caleufú, La Pampa, Argentina. Ameghiniana 40: 495-499. [ Links ]
Esteban, G., Nasif, N., Muruaga, C. y Prado, J. 2004. Nuevo registro de dasipódidos de la localidad Puerta de Corral Quemado, provincia de Catamarca. 20º Jornadas Argentinas de Paleontología de Vertebrados (La Plata), Resúmenes p. 25. [ Links ]
Fitzinger, L.J. 1871. Die Natürliche famille der Gürtelthiere (Dasypodes). Sitzunsberichte Akad. Wissench. , Wien, 64: 209-276, 329-390. [ Links ]
Francis, J.C. 1965. Los géneros de la subfamilia Mesotheriinae (Typotheria, Notoungulata) de la República Argentina. Boletín del Laboratorio de Paleontología de Vertebrados (Montevideo) 1: 1-31. [ Links ]
Francis, J.C. y Mones, A. 1966. La presencia de vizcachas [ Lagostomus ( Lagostomopsis ) spicatus (Amegh)] en la Fm. Kiyú, Dto. de San José, R.O. del Uruguay. Kraglieviana 22: 147-168. [ Links ]
Goin, F.J., Montalvo, C.I. y Visconti, G. 2000. Los marsupiales (Mammalia) del Mioceno superior de la Formación Cerro Azul (provincia de La Pampa, Argentina). Estudios Geológicos 56: 101-126. [ Links ]
Hünicken, M., Tauber, A. y Leguizamón, R. 2001. Hallazgo de huevos y nidos de dinosaurios, asociados a restos de vegetales silicificados: asignación al Cretácico de las secuencias portadoras aflorantes en Sanagasta, provincia de La Rioja. Ameghiniana , Suplemento Resúmenes 38: 10R. [ Links ]
Hünicken, M., Tauber, A. y Leguizamón, R. 2002. Parque Geológico Sanagasta. Nuevos hallazgos sobre Estratigrafía y Paleontología: relevancia para el turismo. Seminario sobre investigaciones aplicadas a la producción regional (CRILAR, Villa de Anillaco, 2002), Resúmenes : 39. [ Links ]
Kraglievich, L. 1926. Sobre el conducto humeral en las vizcachas y paquirucos chapadmalenses con descripción del Paedotherium imperforatum . Anales del Museo Nacional de Historia Natural, Paleontología: Vertebrados 58: 45-88. [ Links ]
Kraglievich, L. 1934. La antigüedad pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron. Imprenta El Siglo Ilustrado, pp. 17-133. [ Links ]
Machicote, M.E. 1946. [ Bosquejo geológico de la cuenca de Los Sauces y valle de Sanagasta, provincia de La Rioja. Tesis Doctoral. Facultad de Ciencias Exactas, Físicas y Natulares, Universidad Nacional de Córdoba, 106 pp. Inédita]. [ Links ]
Marshall, L.G. y Patterson, B. 1981. Geology and geochronology of the mammal-bearing Tertiary of the Valle de Santa María and Río Corral Quemado, Catamarca province. Argentina. Fieldiana Geology ( new series ) 9: 1-80. [ Links ]
Marshall, L.G., Hoffstetter, R. y Pascual, R. 1983. Mammals and Stratigraphy: Geochronology of the continental mammal-bearing Tertiary of South America. Palaeovertebrata , Mémoire Extraordinaire 1983: 1-93. [ Links ]
Miall, A.D. 1996. The Geology of Fluvial Deposits. Sedimentary Facies, Basin Analysis, and Petroleum Geology. Springer-Verlag, Berlín, 1-582. [ Links ]
Montalvo, C.I., Cardonatto, M.C., Visconti, G., Verzi, D.H. y Vucetich, M.G. 1996. Vertebrados de la Formación Cerro Azul (Mioceno tardío) del Valle de Quehué, provincia de La Pampa, Argentina. 6º Jornadas Pampeanas de Ciencias Naturales (Santa Rosa), Actas : 159-165. [ Links ]
Montalvo, C.I., Verzi, D.H. y Vucetich, M.G. 1999. Nuevos datos sobre los pequeños Octodontidae (Rodentia, Caviomorpha) del Mioceno tardío de Argentina. 14º Jornadas Argentinas de Paleontología de Vertebrados (Neuquen y Plaza Huincul), Ameghiniana 36: 106. [ Links ]
Mulé, P., Powell, J.E. y Bossi, G. 1997. Notoungulados de la Formación Salicas (Mioceno), La Rioja, Argentina. Aspectos sedimentológicos y cronológicos. 13º Jornadas Argentinas de Paleontología de Vertebrados (La Rioja), Resúmenes . [ Links ]
Nasif, N. y Esteban G. 2000. Nuevo registro de Neophanomys biplicatus (Ocodontidae, Caviomorpha) en el Neógeno del noroeste argentino. Ameghiniana , Suplemento Resúmenes 37: 13R. [ Links ]
Pascual, R. 1954. Adiciones a la fauna de la Formación de Los Llanos de San Luis y su edad. Revista del Museo Municipal de Ciencias Naturales y Tradición de Mar del Plata 1: 113-121. [ Links ]
Pascual, R. y Bondesio, P. 1981. Sedimentitas cenozoicas. 8º Congreso Geológico Argentino (San Luis), Geología y recursos naturales de la provincia de San Luis, Relatorio : 117-154. [ Links ]
Pascual, R., Ortega Hinojosa, E.J., Gondar, D y Tonni, E.P. 1966. IV. Vertebrata. En: A.V. Borrello (eds.), Paleontografía bonaerense. Comisión de Investigación Científica de la Provincia de Buenos Aires 202 pp. [ Links ]
Powell, J., Mulé, P., Duarte, R., Ortiz, P., Sanagua, J. y Muruaga, C. 1998. Mamíferos de la Formación Chiquimil (Mioceno superior) de la vertiente occidental de la sierra de Hualfín, provincia de Catamarca. 11º Jornadas Argentinas de Paleontología de Vertebrados (San Miguel de Tucumán), Acta Geológica Lilloana 18: 178. [ Links ]
Ramos, A. 1992. Sistemas aluviales braided. En: A. Arche (coord.), Sedimentología, Consejo Superior de Investigaciones Científicas , Madrid 1: 67-106. [ Links ]
Riggs, E.S. y Patterson, B. 1939. Stratigraphy of Late-Miocene and Pliocene deposits of the Province of Catamarca (Argentina) with notes on the faunae. Physis 14: 143-162. [ Links ]
Rodríguez Brizuela, R. 2002. [ Estratigrafía y vertebrados fósiles de la Formación Toro Negro (Neógeno) del departamento Vinchina, provincia de La Rioja, Argentina. Trabajo Final, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba., 100 pp. Inédito.]. [ Links ]
Rolleri, E.O., Dellapé, D.A. y Manceñido, M.O. 1984a. Relaciones estratigráficas y correlación de las Formaciones Pichi Picún Leufú y Collón Curá (Miembro Naupa Huen), aflorantes en el curso inferior y medio del cañadón Michihuau, provincia de Río Negro. 9º Congreso Geológico Argentino (San Carlos de Bariloche), Actas 1: 458-477. [ Links ]
Rolleri, E.O., Dellapé, D.A. y Manceñido, M.O. 1984b. Estudio geológico del valle del río Limay entre Piedra del Águila y El Chocón (provincias del Neuquén y Río Negro). 9º Congreso Geológico Argentino (San Carlos de Bariloche), Actas 1: 478-497. [ Links ]
Rovereto, C. 1914. Los estratos Araucanos y sus fósiles. Anales del Museo Nacional de Historia Natural de Buenos Aires 25: 1-247. [ Links ]
Rusconi, C. 1933. Apuntes preliminares sobre las arenas puelchenses y su fauna. Anales de la Sociedad Científica Argentina 116: 169-193. [ Links ]
Rusconi, C. 1936. Restos de mamíferos terciarios de San Luis. Boletín Paleontológico Buenos Aires 6: 1-4. [ Links ]
Rusconi, C. 1948. El puelchense de Buenos Aires y su fauna (Plioceno medio). (Primera parte). Publicaciones del Instituto de Fisiografía y Geología 33: 1-99. [ Links ]
Rusconi, C. 1949. El Puelchense de Buenos Aires y su fauna (Plioceno medio). (Segunda parte). Publicaciones del Instituto de Fisiografía y Geología 36: 100-242. [ Links ]
Scillato Yané, G.J. 1975. Presencia de Macroeuphractus retusus (Xenarthra, Dasypodidae) en el Plioceno del Área Mesopotamia (Argentina). Su importancia bioestratigráfica y paleobiogeográfica. Ameghiniana 12: 322-328. [ Links ]
Scillato Yané, G.J. 1977. Octomylodontinae: una nueva Subfamilia de Mylodontidae (Edentata, Tardigrada). Descripción del cráneo y mandíbula de Octomylodon robertoscagliai n. sp. procedente de la Formación Arroyo Chasicó (Edad Chasiquense, Plioceno temprano) del sur de la provincia de Buenos Aires (Argentina). Algunas consideraciones filogenéticas y sistemáticas sobre los Mylodontoidea. Publicaciones del Museo Municipal de Ciencias Naturales Lorenzo Scalglia 2: 123-140. [ Links ]
Scillato Yané, G.J. 1980a. Catálogo de los Dasypodidae fósiles (Mammalia, Edentata) de la República Argentina. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires), Actas 3: 7-36. [ Links ]
Scillato Yané, G.J. 1980b. Nuevo Megalonychidae (Edentata, Tardigrada) del Mesopotamiense (Mioceno Tardío-Plioceno) de la provincia de Entre Ríos. Ameghiniana 17: 193-199. [ Links ]
Scillato Yané, G.J. 1981. Nuevo Mylodontinae (Edentata, Tardigrada) del Mesopotamiense (Mioceno Tardío-Plioceno) de la provincia de Entre Ríos. Ameghiniana 18: 29-34. [ Links ]
Scillato Yané, G.J. 1982. [ Los Dasypodidae (Mammalia-Edentata) del Plioceno y Pleistoceno de Argentina. Tesis Doctoral 406. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 159 pp. Inédita]. [ Links ]
Scillato Yané, G.J., Carlini, A.A., Vizcaíno, S.F. y Ortiz Jaureguizar, E. 1995. Los Xenartros. En: M. T. Alberdi, G. Leone y E. P. Tonni (eds.), Evolución biológica y climática de la región pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el mediterráneo occidental. Museo Nacional de Ciencias Naturales , Madrid, Monografías 12: 183-209. [ Links ]
Simpson, G.G. 1974. Notes on Didelphidae (Mammalia, Marsupialia) from the Huayquerian (Pliocene) of Argentina. American Museum Novitates 2559: 1-15. [ Links ]
Sosic, M.V.J. 1972. Descripción de la Hoja Geológica 14d, Tinogasta, provincias de Catamarca y La Rioja. Dirección Nacional de Geología y Minería, Boletín 129. [ Links ]
Sosic, M.V.J. 1973. Descripción de la Hoja Geológica 14e, Salar de Pipanaco. Dirección Nacional de Geología y Minería, Boletín 137: 1-47. [ Links ]
Stahlecker, R. von 1935. Stratigraphy of the Araucanian Formation (Pliocene). Proceedings of the Geological Society of America 402-405 pp. [ Links ]
Tauber, A.A. 1996. Los representantes del género Protypotherium (Mammalia, Notoungulata, Interatheriidae) del Mioceno temprano del sudeste de la provincia de Santa Cruz, República Argentina. Academia Nacional de Ciencias (Córdoba), Miscelánea 95: 1-29. [ Links ]
Tauber, A. A. 1997. Bioestratigrafía de la Formación Santa Cruz (Mioceno inferior) en el extremo sudeste de la Patagonia. Ameghiniana 34: 413-426. [ Links ]
Tauber, A.A. 2002. Parque Geológico Sanagasta, una nueva área protegida de la provincia de La Rioja, Argentina. 1º Congreso Internacional: Patrimonio Cultural (Córdoba), Actas CD-R: 471-483. [ Links ]
Tauber, A.A. y Mazzoni, A.F. 2003. Trazas de insectos xilófagos de la Formación Salicas (Mioceno tardío) de la provincia de Catamarca, Argentina. Ameghiniana , Suplemento Resúmenes 40: 94R. [ Links ]
Tonni, E.P., Prado, J.L., Fidalgo, F. y Laza, J.L. 1992. El Piso/Edad Montehermosense (Plioceno) y sus mamíferos. 3º Jornadas Geológicas Bonaerenses (La Plata), Actas : 113-118. [ Links ]
Tonni, E.P., Scillato Yané, G.J., Cione, A.L. y Carlini, A. 1998. Bioestratigrafía del Mioceno continental en el curso inferior del arroyo Chasicó, provincia de Buenos Aires. 7º Congreso Argentino de Paleontología y Bioestratigrafía (Bahía Blanca), 135. [ Links ]
Turner, J.A.C. 1971. Descripción Geológica de la Hoja 15d, Famatina, provincia de La Rioja. Dirección Nacional de Geología y Minería, Boletín 126. [ Links ]
Verzi, D.H., Montalvo, C.I. y Vucetich, M.G. 1991. Nuevos restos de Xenodontomys simpsoni Kraglievich, y la sistemática de los más antiguos Ctenomyinae (Rodentia, Octodontidae). Ameghiniana 28: 325-331. [ Links ]
Verzi, D.E., Montalvo, C.I. y Vucetich, M.G. 1999. Afinidades y significado evolutivo de Neophanomys biplicatus (Rodentia, Octodontidae) del Mioceno tardío-Plioceno temprano de Argentina. Ameghiniana 36: 83-90. [ Links ]
Villarroel, C. 1974. Les Mésothérinés (Notoungulata, Mammalia) du Pliocène de Bolivie. Leurs rapports avec ceux d´Argentine. Annales de Paléontologie ( Vertébrés ) 60, 2: 243-286. [ Links ]
Vizcaíno, S.F. y Bargo, M.S. 1993. Los armadillos (Mammalia, Dasypodidae) de La Toma (Partido de Coronel Pringles) y otros sitios arqueológicos de la provincia de Buenos Aires. Consideraciones paleoambientales. Ameghiniana 30: 435-443. [ Links ]
Vucetich, M.G. y Verzi, D.H. 1995. Los roedores caviomorphos. En: M.T. Alberdi, G. Leone y E.P. Tonni (eds.), Evolución biológica y climática de la región pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el mediterráneo occidental. Museo Nacional de Ciencias Naturales , Madrid, Monografías 10: 211-225. [ Links ]
Yrigoyen, M. 1994. Revisión estratigráfica del Neógeno de las Huayquerías de Mendoza septentrional, Argentina. Ameghiniana 31: 125-138. [ Links ]
Zetti, J. 1972a. [ Los mamíferos fósiles de edad huayqueriense (Plioceno medio) de la región pampeana . Tesis Doctoral, Universidad de La Plata, 86 pp. Inédito.]. [ Links ]
Zetti, J. 1972b. Observaciones sobre los Pachyrukhine (Notoungulata) del Plioceno argentino. Publicaciones del Museo Municipal de Ciencias Naturales de Mar del Plata 2: 41-52. [ Links ]
Zuzek, A.B. 1978. Descripción Geológica de la Hoja 18 f. Chamical, provincia de La Rioja. Servicio Geológico Nacional, Boletín 161*. [ Links ]
Recibido: 27 de marzo de 2003.
Aceptado: 12 de agosto de 2004.

