










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana v.42 n.2 Buenos Aires mar./jun. 2005
Morfología caudal de Pterodaustro guinazui (Pterosauria: Ctenochasmatidae) del Cretácico de Argentina
Laura S. Codorniú 1
1 Departamento de Geología, Universidad Nacional de San Luis, Chacabuco y Pedernera, 5700 San Luis, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas. lcodor@unsl.edu.ar
Introducción
La localidad fósil hoy conocida como Loma del Pterodaustro (32° 29' 65'' S; 66° 59' 38'' O) incluye afloramientos de la Formación Lagarcito del Cretácico Inferior de la provincia de San Luis, Argentina. Este yacimiento fue descubierto a principios de la década de 1960 por un equipo dirigido por José Bonaparte, quien describió por primera vez a Pterodaustro guinazui (Bonaparte, 1970). Posteriormente, material adicional, fundamentalmente craneal, permitió ampliar el conocimiento acerca de este taxón (Bonaparte, 1971, 1978; Sánchez, 1973).
Durante la década del 90 varias campañas paleontológicas a la misma área recolectaron numerosos materiales del pterosaurio pterodactiloideo filtrador Pterodaustro guinazui (Chiappe et al ., 1998a; 1998b; 2000), entre ellos se registran ejemplares correspondientes a individuos juveniles y subadultos, cuyas colas se encuentran bien preservadas. Estos especimenes aportan la mejor evidencia conocida de la morfología de la serie caudal de este pterosaurio (Codorniú y Chiappe, 2004). La totalidad de los especimenes asignados preliminarmente a Pterodaustro se encuentran depositados en tres instituciones, a saber: Museo de Ciencias Naturales de Mar del Plata, Instituto Miguel Lillo de la Universidad Nacional de Tucumán y Universidad Nacional de San Luis.
Pocos son los pterodactiloideos para los cuales se conoce la morfología de la serie caudal. Entre éstos se encuentran Pteranodon del Cretácico superior de América del Norte (Bennett, 1987; 2001), Anhanguera piscator del Cretácico inferior de Brasil (Kellner y Tomida, 2000), Dsungaripterus weii (Young, 1964) del Cretácico inferior de China, y Pterodactylus antiquus del Jurásico superior de Alemania (Wellnhofer, 1970; Bennett, 1987). Si bien existe una reconstrucción tentativa de la cola de Pterodaustro guinazui (Bonaparte, 1978), la descripción de especímenes inéditos que incluyen las vértebras caudales bien preservadas adiciona nueva información para el conocimiento de la evolución de la morfología caudal de los pterosaurios.
Abreviaturas. MHIN-UNSL-GEO-V, Museo de Historia Natural de la Universidad Nacional de San Luis, San Luis, Argentina, Geología - Vertebrados; MMP, Museo Municipal de Ciencias Naturales Galileo Scaglia, Mar del Plata, Argentina; PVL, Instituto Miguel Lillo, Universidad Nacional de Tucumán, San Miguel de Tucumán, Argentina.
Nuevos materiales
Material. MHIN-UNSL-GEO-V 243, post-cráneo muy completo y articulado correspondiente a un individuo sub-adulto; MHINUNSL-GEO-V 165, pelvis y sacro con vértebras caudales; MHINUNSL-GEO-V 241, post-cráneo muy completo correspondiente a un individuo juvenil; MMP 1168 huesos del ala y extremidades posteriores con vértebras caudales de un individuo juvenil.
Descripción. La serie caudal se ha preservado en cuatro ejemplares preliminarmente asignados a Pterodaustro (pero al menos uno de ellos, V 243 presenta dientes mandibulares característicos de este taxón). La descripción se basa principalmente en dos de ellos que corresponden a individuos sub-adultos de distintos tamaños, MHIN-UNSL-GEO-V 243 y MHIN-UNSL-GEO-V 165 (referidos de aquí en adelante como V 243 y V 165). Los otros dos especímenes, MHIN-UNSL-GEO-V 241 y MMP 1168 (de 28 cm y 30 cm de envergadura alar, respectivamente) representan estados muy tempranos de desarrollo (Codorniú y Chiappe, 2004).
En todos los ejemplares mencionados anteriormente la cola se encuentra incompleta en su extremo distal, la mayor cantidad de elementos se ha conservado en dos de ellos (V 243 y MMP 1168) y consiste de por lo menos 22 vértebras caudales (figuras 1.AB). De esta manera se interpreta que la serie caudal de Pterodaustro consistía de más de 22 vértebras. La longitud total de la serie caudal es de 110,55 mm en V 243, lo que constituye aproximadamente el 7% de la envergadura alar (envergadura aproximada = 1.590 mm). En el ejemplar juvenil MMP 1168, la longitud de la serie caudal preservada representa un 5,6% de la envergadura alar.
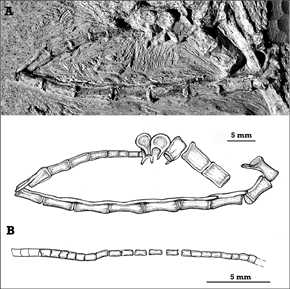
Figura 1. A, fotografía e ilustración de la serie caudal de V 243 que corresponde a un individuo sub-adulto. B, MMP 1168, correspondiente a un individuo juvenil / A, photograph and sketch of the caudal series of V 243, a subadult individual. B, sketch of MMP 1168, juvenile specimen.
Todos los centros vertebrales de la serie caudal son procélicos. No obstante, la morfología de la serie caudal no es uniforme. Los centros de las primeras vértebras son de sección circular pero éstos se van aplanando dorso-ventralmente a medida que se acercan al extremo distal (figuras 2 y 3.A).

Figura 2. Fotografía e ilustración de la serie caudal de V 165. Abreviaturas: cv, centro vertebral; sn, espinas neurales; sv, vértebra sacral; poz, postzigapófisis; prz, prezigapófisis / Photograph and sketch of the caudal series of V 165. Abbreviations: cv, vertebral centre; sn, neural spines; sv, sacral vertebra; poz, postzigapophisis; prz, prezigapophisis.

Figura 3. Comparación de diferentes colas en vista lateral de A, reconstrucción de Pterodaustro guinazui ; B, reconstrucción de Pteranodon (Bennet, 2001); C, Anhanguera piscator (Kellner y Tomida, 2000) y D, Dsungaripterus weii (Young, 1964), en vista lateral y ventral / Comparison of tails in lateral view. A, reconstruction of Pterodaustro guinazui ; B, reconstruction of Pteranodon (Bennet, 2001); C, Anhanguerra piscator (Kellner y Tomida, 2000); D, Dsungaripterus weii (Young, 1964) in lateral and ventral views.
Las longitudes de los centros vertebrales también varían: los dos primeros son los más cortos, a partir de aquí comienzan a alargarse progresivamente, alcanzando la máxima longitud en la 9º vértebra, dimensión que se mantiene casi constante hasta la 12° vértebra, para luego disminuir progresivamente hacia el extremo distal (tabla 1 y figura 3.A). Variaciones en la longitud de los centros vertebrales son comunes en pterosaurios, tanto en no pterodactiloideos como en pterodactiloideos. El patrón de variación de Pterodaustro presenta algunas similitudes con algunos no pterodactiloideos. Por ejemplo, en Campylognathoides liasicus (Campylognathoididae, Kuhn, 1967) la serie caudal aumenta de longitud a partir de la 4° vértebra y los centros vertebrales alcanzan su máxima longitud entre la 9° y la 14° vértebra, para luego disminuir gradualmente hacia el extremo distal (Wellnhofer, 1974). Otro no pterodactiloideo de morfología caudal semejante es Austriadactylus cristatus (Dalla Vecchia et al ., 2002), donde la vértebra que alcanza la máxima longitud es la 9° y donde caudalmente a ésta, la longitud de las subsiguientes vértebras disminuye. Sin embargo la morfología de las vértebras de estos pterosaurios de cola larga es muy diferente ya que presentan las zigapófisis muy alargadas y ramificadas (principalmente las prezigapófisis que son 4-6 veces el largo de la vértebra, hacia anterior; las postzigapófisis son mas cortas y no se ramifican), las hemapófisis también se encuentran extendidas hacia anterior y posterior pero en menor grado; estas estructuras forman como un sistema de tendones osificados que tornan rígida la cola (Wellnhofer, 1978; Dalla-Vecchia et al ., 2002). El patrón de variación de la longitud de los centros entre los pterodactiloideos es menos homogéneo, por ejemplo en Pteranodon las cinco primeras caudales son las más largas y a partir de éstas, las siguientes disminuyen progresivamente para terminar en una varilla caudal (figura 3.B). Mientras que en Anhanguera (Kellner y Tomida, 2000) los dos primeros centros caudales son más pequeños y a partir de la 3° vértebra, los centros aumentan su longitud hasta el 7° y luego disminuyen nuevamente (figura 3.C).

Tabla 1. Medidas de cada una de las vértebras de la serie caudal de Pterodaustro (en mm). Abreviaturas: inc, incompleto; l, longitud del centro vertebral; N°, número de vértebra / Measurements of each caudal vertebra of Pterodaustro (in mm). Abbreviations: inc, incomplete; l, length of vertebral centre; Nº, number of vertebra.
El tamaño de las espinas neurales también varía dependiendo de su posición. En Pterodaustro son altas y dirigidas hacia atrás en las dos vértebras más proximales (figura 2), a partir de las cuales las espinas neurales disminuyen progresivamente en altura, reduciéndose al máximo distalmente.
En casi todas las vértebras de los especimenes descriptos, V 243 y V 165, se preservaron las prezigapófisis y postzigapófisis. Estas articulaciones están muy poco desarrolladas o ausentes en las dos caudales más proximales pero cobran importancia a partir de la tercera caudal donde las prezigapófisis son ligeramente alargadas y proyectadas hacia anterior. Éstas se extienden hacia adelante por encima del borde caudal de la vértebra precedente; las postzigapófisis son más cortas. Este tipo de unión entre las zigapófisis forman como un cordón continuo y lateral a cada lado de la cola (figuras 2 y 3.A). En general las zigapófisis disminuyen de altura desde la región proximal de la cola hacia la distal, estando muy reducidas en las últimas 5-6 vértebras caudales.
Este tipo de diseño caudal sugiere que la cola de Pterodaustro habría sido mucho más flexible proximalmente que distalmente. Es posible que la movilidad lateral de la cola haya estado limitada a las dos vértebras mas proximales y que dichos movimientos laterales se encontraran reducidos en las porciones más distales de la serie caudal, la cual solo habría tenido la capacidad de flexionarse dorsoventralmente. Una inferencia similar fue hecha por Wellnhofer (1974) para la serie caudal de Campylognathoides liasicus (no pterodactiloideo), donde movimientos laterales habrían estado limitados a las primeras cuatro vértebras, ya que dichos movimientos en zonas más distales de la serie caudal habrían estado bloqueados por la presencia de largos tendones osificados.
En su aspecto general, las colas de los ejemplares juveniles (MMP 1168 y V 241) son muy similares entre sí, las vértebras caudales presentan una morfología muy simple probablemente debido a la escasa osificación de las vértebras (figura 1.B).
Discusión
Como se mencionara anteriormente, muy poco se conoce acerca de la morfología de las vértebras caudales de los pterodactiloideos, especialmente dentro de Ctenochasmatidae, taxón al cual pertenece Pterodaustro (Unwin, 1995, 2003; Kellner, 1996, 2003; Chiappe et al ., 2000). Los registros de esta familia incluyen siete géneros, con por lo menos diez especies y la gran mayoría de estos taxones se conocen a partir de material craneal. Esqueletos post-craneanos son conocidos para los holotipos de Huanhepterus quingyangensis y Eosipterus yangi , y para dos especimenes de Ctenochasma gracile . La serie caudal de Huanhepterus quingyangensis preserva las impresiones de solo dos vértebras. Dong (1982) sugirió que la cola de este pterosaurio habría sido probablemente corta y que sus centros vertebrales habrían tenido una morfología simple. Eosipterus yangi fue recientemente clasificado dentro de los Ctenochasmatinae, al igual que Pterodaustro (Unwin et al ., 2000) sin embargo sólo se ha preservado escasamente la serie caudal, deduciéndose que la misma fue corta (Ji y Ji, 1997). Con respecto a Ctenochasma , el género más cercano a Pterodaustro , sólo dos especímenes preservan el esqueleto post-craneal y éstos presentan sólo unas pocas vértebras caudales (Oppel, 1862; Broili, 1936).
Dentro de Pterodactylidae, taxón que comparte una única condición altamente derivada del cráneo con Ctenochamatidae (Unwin, 1995; 2003), Pterodactylus se diferencia de Pterodaustro en que posee un menor número de vértebras caudales y porque los centros son anfiplanos (Bennett, 1991). La serie caudal de Pterodactylus antiquus está integrada por 16 vértebras simples, con centros vertebrales anfiplanos, que carecen de procesos transversos y que disminuyen en tamaño hacia el extremo distal (Bennett, 2001).
La serie caudal de otros pterosaurios de cola corta ha sido sólo descripta en detalle para Anhanguera (Kellner y Tomida, 2000) y Pteranodon (Bennett, 2001). Pteranodon posee numerosas diferencias con Pterodaustro . En primer lugar, la cola de Pteranodon consiste de sólo 11 vértebras, la 1° vértebra tiene muy pequeñas prezigapófisis y carece de postzigapófisis, y ninguna de las caudales subsiguientes tiene zigapófisis (Bennett, 2001). Por otro lado, las cinco anteriores tienen centros amplios y bajos, con los cóndilos articulares anteriores y posteriores dobles, y las siguientes 6 vértebras forman la zona de transición en la cual el arco neural se pierde totalmente y el centro es excavado por el canal neural. Finalmente, estas 11 vértebras están seguidas por una varilla caudal (figura 3.B).
Anhanguera se diferencia de Pterodaustro por que las tres primeras vértebras poseen centros cuyas caras articulares anteriores y posteriores forman un cóndilo con doble articulación y porque a partir de la 4° vértebra y hasta la 7° vértebra, existe una proyección ventral única (Kellner y Tomida, 2000). En este taxón se preservaron 11 vértebras, sin embargo el número total de caudales es desconocido (Kellner y Tomida, 2000). Lo característico de Anhanguera es que las espinas neurales de las 5°-7° vértebras se proyectan cranealmente extendiéndose por encima de la región caudal de la vértebra precedente; formando así un reborde dorsal continuo (figura 3.C). Sin embargo, Pterodaustro es semejante a la forma brasilera en el hecho de que los centros vertebrales son procélicos (a partir del tercero en Anhanguera ).
Además de estos dos pterodactiloideos, donde las vértebras caudales han sido descriptas en detalle, porciones substanciales de la serie caudal son conocidas para Dsungaripterus weii del Cretácico temprano de la China (Young, 1964) y Nyctosaurus del Cretácico tardío de América del Norte. La diferencia fundamental con Dsungaripterus es que las vértebras son opistocélicas, con arcos neurales altos y procesos transversos. Si bien la serie caudal de este pterosaurio no es conocida en su totalidad -un máximo de 9 vértebras se han preservado en 2 series- y se ha considerado (Young, 1964) que la serie caudal de esta forma probablemente consistía de 12 a 14 elementos (figura 3.D). Lo conocido de la serie caudal de Nyctosaurus indica que las vértebras de la parte proximal de la cola de este pterosaurio eran amplias y bajas, y que éstas probablemente se encontraban seguidas por una varilla caudal. Esto último ha llevado a Bennett (1987, 2001) a proponer que la serie caudal de Nyctosaurus podría haber sido muy semejante a la de Pteranodon .
Como se ha observado hasta ahora, Pterodaustro tiene la particularidad de poseer un mayor número de vértebras caudales que cualquier otro pterodactiloideo conocido. Además, morfológicamente, Pterodaustro se distingue claramente de otros pterodactiloideos. De esta manera se concluye que el elevado número de vértebras caudales (más de 22 elementos) y la variación morfológica contenida dentro de la serie caudal de Pterodaustro guinazui constituyen una potencial autopomorfía de este taxón.
Agradecimientos
Agradezco especialmente a L. Chiappe por la lectura y las sugerencias en el manuscrito y a A. Arcucci por sus comentarios. Los trabajos de campo y de laboratorio fueron financiados mediante subsidios de la National Geographic Society, Philip McKenna Foundation, y la Fundación Antorchas otorgados a L. Chiappe, y por una beca del Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) otorgada a L. Codorniú. Este manuscrito se realizó dentro del marco del Proyecto de Ciencia y Técnica de la Universidad Nacional de San Luis (UNSL). Mi especial agradecimiento a los voluntarios que participaron en las expediciones de campo y a Carlos Gardini del Departamento de Geología de la UNSL, por la fotografía.
Bibliografía
Bennett, S.C. 1987. New evidence on the tail of Pterosaur Pteranodon (Archosauria: Pterosauria). Fourth Symposium on Mesozoic Terrestrial Ecosystems . Short Papers: 18-23. [ Links ]
Bennett, S.C. 1991. A large pterodactyloid pterosaur from the Late Cretaceous (Late Mastrichtian) of Western Australia. Records of the Western Australian Museum 15: 435-443. [ Links ]
Bennett, S.C. 2001. The osteology and functional morphology of the late Cretaceous pterosaur Pteranodon . Palaeontographica A, 260: 113-153. [ Links ]
Bonaparte, J.F. 1970. Pterodaustro guiñazui gen. et. sp. nov. Pterosaurio de la Formación Lagarcito, Provincia de San Luis, Argentina y su significado en la geología regional (Pterodactylidae). Acta Geológica Lilloana 10: 207-226. [ Links ]
Bonaparte, J.F. 1971. Descripción del cráneo y mandíbulas de Pterodaustro guiñazui (Pterodactyloidea, Pterodaustriidae nov.) de la Formación Lagarcito, San Luis, Argentina. Publicaciones del Museo Municipal de Ciencias Naturales de Mar del Plata 1: 263-272. [ Links ]
Bonaparte, J.F. 1978. El Mesozoico de América del Sur y sus Tetrápodos . Opera Lilloana 26. [ Links ]
Broili, F. 1936. Weitere Beobachtungen an Ctenochasma . Sitzungsberichte der Mathematisch - Naturwissenschaftlichen Klasse der Bayerischen Akademie der Wissenschaften : 137-156. [ Links ]
Chiappe, L.M., Rivarola, D., Cione, L., Fregenal, M., Sozzi, H., Buatois, L., Gallego, O., Laza, J.H., Romero, E., López, A., Buscalioni, A., Marsicano, C., Adamonis, S., Ortega, P., Mc Gehee, S. y Di Iorio 1998a. Biotic Association and Paleoenvironmental Reconstruction of the Loma del Pterodaustro Fossil Site (Lagarcito Formation, Early Cretaceous, San Luis, Argentina). Geobios 31: 349-369. [ Links ]
Chiappe, L.M., Rivarola, D., Romero, E., Dávila, S. y Codorniú, L.S. 1998 b. Recent advances in the paleontology of the lower Cretaceous Lagarcito Formación (Parque Nacional Sierra de Las Quijadas, San Luis; Argentina). New Mexico Museum of Natural History and Science Bulletin 14: 187-192. [ Links ]
Chiappe, L.M., Kellner, A., Rivarola, D., Dávila, S. and Fox, M. 2000. Cranial morphology of Pterodaustro guinazui (Pterosauria: Pterodactyloidea) from the lower Cretaceous of Argentina. Contributions in Science 483: 1-19. [ Links ]
Codorniú, L.S. and Chiappe, L.M. 2004. Early juvenile pterosaurs (Pterodactyloidea: Pterodaustro guinazui ) from the Lower Cretaceous of central Argentina. Canadian Journal of Earth Sciences 41: 9-18. [ Links ]
Dalla Vecchia, F.M., Wild, R., Hopf, H. y Reitner, J. 2002. A crested Rhamphorynchoid Pterosaur from the Late Triassic of Austria. Journal of Vertebrates Paleontology 22: 196-199. [ Links ]
Dong, Z. 1982. A new pterosaur ( Huanhepterus quingyangensis gen. et sp. nov.) from Ordos, China. Vertebrata PalAsiatica 20: 115-121. [ Links ]
Ji, S. y Ji, Q. 1997. Discovery of a New Pterosaur in Western Liaoning, China. Acta Geologica Sinica 71: 115-123. [ Links ]
Kellner, A. W. A. 1996. [ Description of new material of Tapejaridae and Anhangueridae (Pterosauria, Pterodactyloidea) and discussion of pterosaur phylogeny. Doctoral Disertation. New York, Columbia University: 347 pp. Unpublished.]. [ Links ]
Kellner, A.W.A. 2003. Pterosaurs phylogeny and comments on the evolutionary history of the group. En: E. Buffetaut y J.M. Mazin (eds.), Evolution and Paleobiology of Pterosaurs. Geological Society, Special Publication 217: 105-137. [ Links ]
Kellner, A.W.A. y Tomida, Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian) Northeastern Brazil. National Science Museum Monographs 17: 1-135. [ Links ]
Kuhn, O. 1967. Die fossile Wirbeltierklasse Pterosauria. Oeben. Krailling bei München 52 pp. [ Links ]
Oppel, A. 1862. Ueber Führten im lithographischen Schiefer. Palaeontologische Mittheilungen aus dem Museum des Koenigliche Bayerisches Staates 1: 121-125. [ Links ]
Sánchez, T. 1973. Redescripción del cráneo y mandíbulas de Pterodaustro guiñazui Bonaparte (Pterodaytyloidea, Pterodaustriidae). Ameghiniana 10: 313-325. [ Links ]
Unwin, D.M. 1995. Preliminary results of a phylogenetic analysis of the Pterosauria (Diapsida: Archosauria). 6º Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers : 69-72. [ Links ]
Unwin, D. M. 2003. On the phylogeny and evolutionary history of pterosaurs. En: E. Buffetaut y J.M. Mazin (eds.), Evolution and Paleobiology of Pterosaurs. Geological Society , Special Publication 217: 139-190. [ Links ]
Unwin, D.M., Lü, J. y Bakhurina, N.N. 2000. On the systematic and stratigraphic significance of pterosaurs from the Lower Cretaceous Yixian Formation (Jehol Group) of Liaoning, China. Mitt. Mus. Nat.kd. Geowiss. Reihe 3: 181-206. [ Links ]
Wellnhofer, P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke. Süddeutschlands. Bayerische Akademie Neve Folge Heft 141, 133 pp. [ Links ]
Wellnhofer, P. 1974. Campylognathoides liasicus (Quenstedt) an Upper Liassic pterosaur from Holzmaden the Pittsburgh specimen. Annals of Carnegie Museum 45: 6-34. [ Links ]
Wellnhofer, P. 1978. Pterosauria. En: P. Wellnhofer (ed.), Handbuch der Paläoherpetologie. Stuttgart: Gustav Fischer, Stuttgart, Teil 19, 82 pp. [ Links ]
Young, C. 1964. On a new pterosaurian from Sinkiang, China. Vertebrata PalAsiatica 8: 221-255. [ Links ]
Recibido: 6 de noviembre de 2003.
Aceptado: 6 de julio de 2004.

