










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAmeghiniana
versão On-line ISSN 1851-8044
Ameghiniana v.42 n.3 Buenos Aires jun./set. 2005
Los Litopterna (Mammalia) de la Formación Pinturas, Mioceno Temprano-Medio de Patagonia
Alejandro G. Kramarz 1 y Mariano Bond 2
1 Consejo Nacional de Investigaciones Científicas y Técnicas, Sección Paleontología de Vertebrados. Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Av. Angel Gallardo 470, C1405DJR Buenos Aires, Argentina. agkramarz@macn.gov.ar
2 Consejo Nacional de Investigaciones Científicas y Técnicas, Departamento Paleontología de Vertebrados. Museo de La Plata, Paseo del Bosque s/n, 1900 La Plata, Argentina. constantino1453@yahoo.com.ar
Abstract. Litopterna (Mammalia ) from the Pinturas Formation , Early -middle miocene of Patagonia. The Litopterns from the Pinturas Formation (Ameghino´s Astrapothericulan beds) are examined. Four species referable to the Proterotheriidae are recognized for the lower and middle sequences of this unit: Picturotherium migueli gen. et sp. nov, Lambdaconus lacerum (Ameghino), Tetramerorhinus prosistens (Ameghino), and T. fleaglei Soria. These species retain plesiomorphic dental features, as the labial folds of the paracone and metacone in the upper molars and the little development of the neoparaconid–neoparalophid in lower molars, characteristic of the Deseadan and Colhuehuapian proterotheriids (Soria´s Oligocene Group). Proterotheriids with more derived dental morphotypes, Tetramerorhinus cingulatum ? (Ameghino) and Diadiaphorus ? caniadensis sp. nov., are recorded at those localities referred to the upper sequence. This stratigraphic distribution agree with that of the hystricognath rodents and palaeothentid marsupials, and reinforces the hypothesis that the lower and middle sequence of the Pinturas Formation bear a mammal assemblage different from and older than those derived from the typical Santacrucian localities of the Santa Cruz Formation. Macraucheniids are only recorded in those levels interpreted as belonging to the upper sequence of the Pinturas Formation. The absence of macrauqueniids in the lower and middle sequences of the Pinturas Formation could be the consequence of peculiar and still not well determined palaeoenvironmental conditions during early Pinturas times.
Resumen. Se analizan los Litopterna de la Formación Pinturas (Capas con Astrapothericulus de Ameghino). Se reconocen cuatro especies referidas a Proterotheriidae para las secuencias inferior y media de esta unidad: Picturotherium migueli gen. et sp. nov., Lambdaconus lacerum (Ameghino), Tetramerorhinus prosistens (Ameghino) y T. fleaglei Soria. Estas especies retienen caracteres dentarios plesiomórficos, como los pliegues labiales del paracono y metacono en los molares superiores y el escaso desarrollo del neoparacónido-neoparalófido en los inferiores, características de los proterotéridos deseadenses y colhuehuapenses (Grupo Oligoceno de Soria). Proterotéridos con morfotipos dentarios más derivados, Tetramerorhinus cingulatum ? (Ameghino) y Diadiaphorus ? caniadensis sp. nov., se registran en aquellas localidades referidas a la secuencia superior. Esta distribución estratigráfica coincide con la de los roedores histricognatos y marsupiales paleoténtidos, y refuerza la hipótesis de que las secuencias inferior y media de la Formación Pinturas contiene una asociación de mamíferos diferente y más antigua que aquella derivada de las típicas localidades santacrucenses de la Formación Santa Cruz. Restos de macrauquénidos sólo se han registrado en niveles interpretados como correspondientes a la secuencia superior de la Formación Pinturas. La ausencia de macrauquénidos en las secuencias inferior y media podría ser consecuencia de las peculiares y aún no bien determinadas condiciones paleoambientales durante los primeros tiempos de Pinturas.
Key words. Litopterna. Proterotheriidae. Macraucheniidae. Systematics. Pinturas Formation. Miocene, Patagonia.
Palabras clave. Litopterna. Proterotheriidae. Macraucheniidae. Sitemática. Formación Pinturas. Mioceno. Patagonia.
Introducción
Bown et al. (1988) denominaron Formación Pinturas a aquellos sedimentos continentales del Mioceno expuestos en el valle superior del Río Pinturas, Noroeste de la provincia de Santa Cruz, Argentina (figura 1) portadores de fauna Astrapothericulense (Ameghino, 1906). Sobre la base de esta fauna, Ameghino (1900-1902, 1906) estableció su Étage Astrapothericuléen, como término superior de la Formation Patagonienne. De acuerdo con Ameghino, la Faune Astrapothericuléene es diferente y más antigua que la Faune Notohippidéene (proveniente de la Formación Santa Cruz en la región de Karaiken, figura 1) y que la Faune Santacruzéen (proveniente de las localidades costeras de la Formación Santa Cruz, figura 1), que tipifica la Edad Santacrucense (Simpson, 1940). Esta propuesta fue seguida por Frenguelli (1931) y Castellanos (1937), quien acuñó el término Pinturense para referirse al Astrapothericulense. Sin embargo, autores posteriores (Wood y Patterson, 1959; Pascual et al. , 1965; Pascual y Odreman Rivas, 1971; Marshall, 1976; Marshall et al. , 1977, 1983) consideraron a la fauna Astrapothericulense como una fauna local santacrucense. De Barrio et al. (1984) refirieron estos depósitos a la Formación Santa Cruz, pero señalando que algunos de sus elementos faunísticos mostraban rasgos más primitivos que sus contrapartes derivadas de las típicas localidades de la Formación Santa Cruz.

Figura 1. Mapa de ubicación/ Location map . 1, Area del valle superior del Río Pinturas/ Area of the upper valley of Pinturas river; 2, Estancia La Cañada; 3, Monte León; 4, Monte Observación.
Nuevas y abundantes colecciones de mamíferos fósiles de la Formación Pinturas fueron obtenidas durante expediciones más recientes realizadas conjuntamente por el Museo Argentino de Ciencias Naturales y the State University of New York, lideradas por J.G. Fleagle y, durante los primeros trabajos de campo, por M.F. Soria (MACN). Los primates Platyrrhini, marsupiales Paucituberculata y roedores Hystricognathi fueron examinados (Fleagle et al. , 1987; Fleagle, 1990; Bown y Fleagle, 1993; Rae et al. , 1996; Tejedor, 2002; Dumont et al. , 2000; Kramarz, 2001a, 2001b, 2004). Estudios bioestratigráficos basados en marsupiales paleoténtidos (Bown y Fleagle, 1993) y roedores histricognatos (Kramarz, 2001b; Kramarz y Bellosi, 2005) y recientes dataciones radimétricas (Fleagle et al. , 1995) sugieren que al menos parte de la Formación Pinturas es más antigua que la Formación Santa Cruz en su localidad tipo (Monte León), apoyando la hipótesis original de Ameghino.
Soria (2001) efectuó un extensivo análisis de los Proterotheriidae, donde incluyó un análisis preliminar de aquellos materiales de la Formación Pinturas recolectados antes de su fallecimiento en 1989. En la tabla 1 se listan las especies de Proterotheriidae reconocidas por este autor para el Oligoceno y Mioceno que son analizados o comparados en este estudio y sus respectivas distribuciones estratigráficas y cronológicas. En esta contribución se efectúa un análisis más exhaustivo de la diversidad y sistemática de los Litopterna de la Formación Pinturas sobre la de base los materiales estudiados por Soria y de otros obtenidos con posterioridad a su fallecimiento. Se reconocen aquí nuevas especies y se revisan otras ya conocidas, siguiendo el esquema taxonómico propuesto por Soria (2001). A la luz de las nuevas evidencias paleontológicas y geológicas de la Formación Pinturas, se discuten los significados bioestratigráficos de los Litopterna de esta unidad.

Tabla 1. Lista de especies de Protherotheriidae analizadas o reconocidas en este estudio y sus respectivas distribuciones estratigráficas y cronológicas (ampliado de Soria, 2001)./ List of species of Proterotheriidae analyzed or recognized in this study and their respective stratigraphic and chronological distributions (after Soria, 2001).
Abreviaturas institucionales. MACN A: Colección Nacional Ameghino, Museo Argentino de Ciencias Naturales B. Rivadavia; MACN Pv: Colección Nacional de Paleovertebrados, Museo Argentino de Ciencias Naturales B. Rivadavia; MLP: Museo de La Plata.
Reseña geológica y localidades fosilíferas
La Formación Pinturas es una sucesión de sedimentos eólicos (mayormente tobáceos) de unos 100 m de espesor (Bown y Larriestra, 1990) acumulados en las zonas periféricas elevadas del noroeste de la cuenca de Magallanes. Aflora en numerosas áreas aisladas en el valle superior del río Pinturas, en la zona precordillerana del noroeste de la provincia de Santa Cruz, Argentina. Esta unidad presenta dos discordancias erosivas intraformacionales principales que definen tres secuencias. La secuencia inferior está caracterizada por paleosuelos muy maduros y pelitas piroclásticas; la secuencia media está dominada por uno o dos niveles de psamitas epiclásticas correspondientes a paleodunas barjanoides con paleosuelos inmaduros; la secuencia superior está caracterizada por pelitas piroclásticas masivas con paleosuelos muy inmaduros (Bown y Larriestra, 1990).
Los restos de mamíferos fósiles fueron colectados en ocho principales localidades aisladas distribuidas en el área del valle del río Pinturas y positivamente referidas a la Formación Pinturas: estancia El Carmen, portezuelo Sumich Norte, portezuelo Sumich Sur, cerro de los Monos, loma de la Lluvia, estancia Los Toldos Norte, estancia Los Toldos Sur y estancia Ana María. La posición estratigráfica relativa de estas localidades ha sido determinada por medio de comparaciones litológicas (Bown y Larriestra, 1990). Otros fósiles fueron colectados en localidades ubicadas entre el valle del río Pinturas y la costa atlántica, en la estancia La Cañada, Gobernador Gregores y el lago Cardiel (figura 1), en rocas que se asemejan a las de la Formación Pinturas en su área típica y tentativamente referidas a esta unidad (Bown y Fleagle, 1993).
Los restos de vertebrados fósiles se encuentran mayormente concentrados en paleosuelos relacionados a incisiones fluviales y en paleosuelos pobre a moderadamente desarrollados de las tres secuencias (Bown y Fleagle, 1993). Como el de otros mamíferos, el registro de Litopterna de la Formación Pinturas consiste en numerosos fragmentos mandibulares y maxilares con parte de la dentadura y un gran número de dientes aislados. También se han recuperado algunos restos postcraneanos (astrágalos y calcáneos) asignables a Proterotheriidae, pero ninguno asociado a restos dentarios.
Antecedentes
Los primeros registros de Litopterna procedentes de las capas con Astrapothericulus fueron dados a conocer por F. Ameghino (1899), basados en restos escasos y fragmentarios colectados por C. Ameghino en 1897 que carecen de datos de procedencia estratigráfica y geográfica precisa. F. Ameghino identificó las siguientes especies: Proterotherium prosistens , Heptaconus obcallatus y Diadiaphorus zamius , todas referidas a la familia Proterotheriidae. Licaphrium debile también fue mencionado por Ameghino (1900-1902) para las capas con Astrapothericulus ; el holotipo, extraviado en las colecciones del MACN, fue considerado nomen dubium por Soria (2001) por estar basada en un material muy fragmentario, posiblemente deformado, y que no presenta caracteres diagnósticos. A pesar de la procedencia indicada por Ameghino, Scott (1910) y Soria (2001) consideraron que esta especie provendría de la Formación Santa Cruz. De Barrio et al. (1984) reportaron el hallazgo de unos pocos restos asignados a ? Proterotherium sp. y Proterotheriinae incertae saedis en las Capas con Astrapothericulus . En un detallado y extenso estudio de los Proterotheriidae, Soria (2001) propuso que las tres especies reconocidas por Ameghino para las capas con Astrapothericulus son sinónimas y que el nombre válido es Tetramerorhinus prosistens (Ameghino). Sobre la base de un examen preliminar de las nuevas colecciones de la Formación Pinturas, Soria también reconoció la presencia de Lambdaconus lacerum (Ameghino), Tetramerorhinus mixtum (Ameghino) y Tetramerorhinus cingulatum fleaglei Soria.
Paleontología sistemática
Orden L itopterna Ameghino, 1889
Suborden L opholitopterna Cifelli, 1983
Familia P roterotheriidae , Ameghino, 1887
Picturotherium gen. nov.
Especie tipo. Picturotherium migueli sp. nov.
Etimología. Del latín: pictura = pintura, en referencia al Río Pinturas, y del griego: therios = bestia .
Distribución geográfica y estratigráfica. Formación Pinturas (Mioceno Temprano-Medio, Fleagle et al. , 1995) provincia de Santa Cruz, Argentina.
Diagnosis. Pequeño proterotérido, de tamaño similar al de Paramacrauchenia , pero con los M1 y M2 más anchos y más cortos. Molariformes de corona más baja que en Paramacrauchenia . Estilos labiales más pronunciados que en Lambdaconus . Pliegues labiales del paracono y del metacono muy marcados, como en Lambdaconus , Prolicaphrium , Paramacrauchenia y Proterotherium . Cíngulos linguales bien desarrollados como en Tetramerorhinus . Cíngulo anterolingual del P3 - M3 conectado con la base del protocono. P2 con protocono mucho más desarrollado que en Paramacrauchenia , aislado del cíngulo anterolingual. Hipocono rudimentario en P3 y P4, pero más desarrollado que en Paramacrauchenia . Metacónulo presente y conectado con el protocono en P3 y P4. A diferencia de Tetramerorhinus y Prolicaphrium , el metacónulo del M1 y M2 está relacionado a la pared que une hipocono con protocono, como en Paramacrauchenia y Thoatherium . M3 con metacónulo aislado, sin hipocono pero con postcingulum muy alto como en Thoatherium .
Diagnosis . Small proterotheriid, similar in size to Paramacrauchenia , but the M1 and M2 are wider and shorter. Cheek teeth lower crowned than in Paramacrauchenia . Labial styles more pronounced than in Lambdaconus . Labial folds of the paracone and metacone very conspicuous, as in Lambdaconus , Prolicaphrium , Paramacrauchenia and Proterotherium . Labial cingula well developed as in Tetramerorhinus . Anterolingual cingulum of P3-M3 connected to the base of the protocone. P2 with protocone much more developed than in Paramacrauchenia , isolated from the anterolingual cingulum. Hypocone rudimentary in P3 and P4, but more developed than in Paramacrauchenia . Metaconule present and jointed to the protocone in P3 and P4. In opposition to Tetramerorhinus and Prolicaphrium , the metaconule of M1 and M2 is related to the wall uniting hypocone and protocone, as in Paramacrauchenia and Thoatherium . M3 with isolated metaconule, without hypocone but with a very high postcingulum as in Thoatherium .
Picturotherium migueli sp. nov.
Figuras 2.A-D

Figura 2. Picturotherium migueli sp. nov. A, MACN Pv SC4110 (Holotipo), P2-M2 izquierdos/ (Holotype), left P2-M2. B, idem, M3 derecho (invertido)/ right M3 (reversed) . C, MACN Pv SC120, m1 y m2 en erupción derechos/ right m1 and erupting m2 . D, MACN Pv SC119b, m2-m3 izquierdos (invertidos)/ left m2-m3 (reversed) .
Etimología. En reconocimiento a Miguel F. Soria (h), por sus importantes contribuciones al conocimiento de los ungulados fósiles sudamericanos.
Holotipo. MACN Pv SC4110, fragmento de maxilar derecho con M1 y parte del M2, fragmento de maxilar izquierdo con P2 - M1, M2 izquierdo y M3 derecho aislados, todos correspondientes a un mismo individuo (figura 2.A).
Hipodigma. El holotipo y los ejemplares listados en el apéndice.
Procedencia estratigráfica y localidades. El holotipo y el MACN Pv SC4117 proceden de Estancia Ana María; MACN Pv SC4115 y 4116 de Portezuelo Sumich Norte; MACN Pv SC119 a-d del cerro de los Monos; MACN Pv SC4114 y 4118 de Loma de la lluvia (para la ubicación estratigráfica de las localidades, véase Bown y Larriestra, 1990); MACN Pv SC120-122, 124-126 proceden de niveles indeterminados de las secuencias inferior o media de la Formación Pinturas, en el valle superior del río Pinturas.
Diagnosis. Como la del género por monotipia.
Descripción y comparación. Los molariformes tienen aproximadamente el mismo tamaño que los de Paramacrauchenia inexspectata Soria (Edad Colhuehuapense), pero el diámetro anteroposterior de los M1 y M2 son comparativamente menores y el transverso es mayor (tabla 2). La cuenca del trigono es un poco menos profunda que en las especies de Paramacrauchenia Bordas, pero más que en Lambdaconus lacerum (Ameghino).
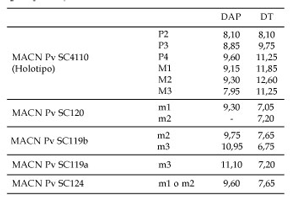
Tabla 2. Medidas dentarias (en mm) de Picturotherium migueli gen. et sp. nov. / Dental measurements (in mm.) for Picturotherium migueli gen. et sp. nov.
Los molariformes superiores (figuras 2.A y B) presentan pliegues labiales del paracono y del metacono muy marcados, como en las especies de Grupo Oligoceno ( e.g. Lambdaconus suinus , L. lacerum , L. inaequifacies , Prolicaphrium specillatum , Paramacrauchenia scamnata y P. inexspectata , sensu Soria, 2001). A diferencia de estas especies, los estilos labiales están bien desarrollados como en Tetramerorhinus . Los cíngulos labiales son muy prominentes como en Thoatherium y Tetramerorhinus , en oposición a las especies del Grupo Oligoceno. El extremo lingual del cíngulo anterolingual del P3 - M3 (figura 2.A) contacta la base del protocono, por lo que la cavidad definida no se abre lingualmente como en Paramacrauchenia .
El P2 (figura 2.A) presenta un gran protocono, como en Thoatherium minusculum , Tetramerorhinus lucarius , T. cingulatum y T. mixtum , pero más alongado transversalmente. Por el contrario, las especies del Grupo Oligoceno carecen de protocono en el P2, menos P. scamnata que sólo presenta una pequeña cúspide labial que podría representar un protocono rudimentario. El cíngulo anterolingual es muy prominente, pero su extremo no llega a contactar al protocono, como en T. prosistens . El cíngulo posterolingual también esta muy desarrollado y contacta con el protocono, a diferencia de T. mixtum . Este cíngulo presenta tres pequeñas cúspulas que decrecen en tamaño hacia el lado labial.
El P3 (figura 2.A) tiene un esbozo de hipocono, que se presenta como una cúspide del cíngulo posterior; éste se extiende hasta el protocono y no se diferencia un flexo lingual, como en T. prosistens y en las especies del Grupo Oligoceno. El metacónulo esta presente, a diferencia de Paramacrauchenia , T. lucarius , Diadiaphorus y Thoatherium ; esta cúspide está unida al protocono, como en L. lacerum , no aislada como en Prolicaphrium . El cíngulo anterolingual es corto y no se extiende sobre la pared lingual del diente, a diferencia de lo observado en algunos ejemplares de Tetramerorhinus , Thoatherium y Diadiaphorus . El P4 (figura 2.A) es de morfología similar al P3 pero con hipocono más desarrollado y flexo lingual entre éste y el protocono más conspicuo, como en T. Cingulatum y Thoatherium , más que en T. prosistens y P. scamnata , pero menos que en T. mixtum , T. lucarius y Diadiaphorus . En oposición a Lambdaconus suinus (= Deuterotherium distichum ) y P. scamnata , el metacónulo está presente y conectado con el protocono, no con el hipocono como en Thoatherium , Diadiaphorus y Neobrachytherium .
Los M1 y M2 (figura 2.A) son similares en morfología, el último es ligeramente mayor. El metacónulo está presente en ambos dientes y presenta una derivación lingual que se conecta con la pared que une protocono e hipocono. Una condición similar está presente en Thoatherium , pero el metacónulo también se extiende labialmente alcanzando al metacono, llegando a formar un metalofo. En P. scamnata el metacónulo está directamente adosado a la pared que une protocono y hipocono, mientras que en P. Inexspectata el metacónulo está adosado al hipocono. El M3 (figura 2.B) carece de hipocono, pero presenta un cíngulo posterolingual muy alto conectado al protocono, como en Thoatherium y P. scamnata . El metacónulo está presente y aislado del protocono y del cíngulo posterolingual. En P. scamnata la presencia de esta cúspide es variable, cuando está presente se halla conectada al cíngulo posterolingual; en Thoatherium el metacónulo raramente está presente y sus relaciones son como en los M1 y M2.
No se han recuperado dientes inferiores y superiores asociados. Los dientes inferiores son asignados aquí tentativamente a Picturotherium migueli porque concuerdan en tamaño con los superiores y difieren de los de todas las otras especies identificadas en la Formación Pinturas. Los m1 y m2 (figuras 2.C y D) presentan el neoparacónido-neoparalófido ( sensu Soria, 2001) poco desarrollado, como en P. scamnata , pero más que en L. lacerum . El entocónido está adosado al hipoconúlido y es ligeramente más pequeño que éste, mientras que en Lambdaconus el entocónido es más grande y, por el otro lado, el entocónido es bastante más pequeño que el hipoconúlido en P. scamnata y Tetramerorhinus , y está completamente reducido en Thoatherium y Diadiaphorus . El m3 (figura 2.D) presenta el hipoconúlido en posición medial, formando un lóbulo posterior bien definido como en Lambdaconus , Paramacrauchenia y Tetramerorhinus . Sin embargo, la hipocrístida contacta tanto al entocónido como al hipoconúlido.
Relaciones filogenéticas de Picturotherium migueli
Ameghino (1904) advirtió que el principal carácter diagnóstico de los molariformes superiores de los proterotéridos es el grado de desarrollo y posición relativa del metacónulo. Sobre esta base, Bianchini y Bianchini (1971) reconocieron dos tipos morfológicos bien definidos para esta familia. Siguiendo esta propuesta, Soria (2001) agrupó a Lambdaconus suinus , Paramacrauchenia , Thoatherium y Neobrachytherium dentro de la Serie Thoatherium , caracterizada por la reducción o fusión del metacónulo al contacto entre el protocono y el hipocono. Picturotherium migueli debe ser referido a la Serie Thoatherium en virtud de que el metacónulo está relacionado al contacto entre el protocono y el hipocono en los M1 y M2. Sin embargo, P. migueli muestra una compleja combinación de caracteres dentarios, dado que difiere de los restantes representantes de la Serie Thoatherium por tener metacónulo no reducido en los P3, P4 y M3, y por otro lado comparte con Thoatherium minusculum cíngulos labiales en los molariformes superiores bien desarrollados, protocono conspicuo en el P2 e hipocono incipiente en el P4. Para evaluar las relaciones filogenéticas entre P. migueli y las especies referidas a la Serie Thoatherium se ha efectuado un análisis cladístico siguiendo el criterio de máxima parsimonia mediante el programa NONA. Se incluyeron en el análisis a Lambdaconus suinus Ameghino, de la Edad Deseadense, Paramacrauchenia scamnata (Ameghino) y P. inexspectata Soria, de la Edad Colhuehuapense, Thoatherium minusculum Ameghino, de la Edad Santacucense, y Neobrachytherium (Moreno y Mercerat), de la Edad Huayqueriense. La polaridad de los caracteres se estableció por comparación con Lambdaconus lacerum (Ameghino), en virtud de que hipótesis previas sugirieren que ésta es la especie más plesiomórfica entre los Proterotheriidae (Soria, 2001). Sólo se analizaron caracteres dentarios, dado que son los únicos que pueden ser evaluados en P. migueli y en la mayoría de los taxones considerados. Los caracteres y sus respectivos estados se listan en la tabla 3. Los caracteres número 11 (condición del metacónulo en el P4) y 12 (condición del metacónulo en los M1 y M2) fueron tratados como caracteres multiestado no ordenados, porque no es posible deducir a priori la serie de transformaciones. La matriz de datos se muestra en la tabla 4.

Tabla 3. Lista de caracteres y estados usados en el análisis cladístico/ List of characters and states used in the cladistic analysis.

Tabla 4. Matriz de caracteres/ Character matrix.
Se obtuvieron tres árboles igualmente parsimoniosos mediante búsqueda exhaustiva, cada uno con 23 pasos, ci = 86 y ri = 81 (figura 3). En todos los árboles obtenidos Picturotherium se agrupa con el clado conformado por Thoatherium y Neobrachytherium por compartir tres sinapomorfías: presencia de cíngulos labiales en los molariformes superiores (carácter 4), protocono en P2 (carácter 6) e hipocono incipiente en P4 (carácter 10, estado 1). M1 y M2 mas anchos que largos (carácter 1) y la presencia de un cíngulo anterolingual en P2 (carácter 7) son interpretados como autapomorfías de Picturotherium . Este género es más plesiomórfico que Thoatherium y Neobrachytherium por tener molariformes de corona más baja (carácter 2), la retención de los pliegues labiales del paracono y metacono (carácter 3), hipocono ausente en P3 (carácter 8) y no completamente desarrollado en P4 (carácter 10), neoparacónido - neoparalófido rudimentario en m1-m3 (carácter 15) y persistencia del lóbulo posterior del m3 (carácter 17), como en las especies del Grupo Oligoceno (Soria, 2001).

Figura 3. Arbol de consenso estricto de los tres árboles más parsimoniosos obtenidos mediante búsqueda exhaustiva. L = 23, ci = 86 y ri = 81. Los números indican las apomorfías correspondientes a los caracteres listados en la tabla 3/ Strict consensus tree from three most parsimonious trees resulting through exhaustive search. Numbers are apomorphies corresponding to characters listed in table 3.
Los caracteres de la dentadura inferior de Picturotherium migueli se evaluaron basándose en molares que se infieren corresponden a esta especie. Si en el análisis se asignan interrogantes a los caracteres de la dentadura inferior de esta especie, los árboles obtenidos presentan las mismas topologías.
Lambdaconus Ameghino, 1897
Especie tipo. Lambdaconus suinus Ameghino, 1897.
Distribución geográfica y estratigráfica. Formación Sarmiento, Edad Deseadense (Oligoceno Tardío?) y Edad Colhuehuapense (Mioceno Temprano), provincia de Chubut; Formación Pinturas (Mioceno Temprano-Medio), provincia de Santa Cruz, Argentina.
Especies referidas. Lambdaconus lacerum (Ameghino), Lambdaconus inaequifacies (Ameghino).
Lambdaconus lacerum (Ameghino, 1902)
Prothoatherium lacerum Ameghino, 1902
Holotipo. MACN A 52-246, fragmento maxilar izquierdo con P3 - M3.
Paratipo. MACN A 52-247, fragmento mandibular izquierdo con p3-m3 completos y parte del alvéolo del p2.
Materiales referidos de la Formación Pinturas. MACN Pv SC119e, M1 o M2 derecho; MACN Pv SC123, M3 izquierdo; MACN Pv SC4112, m3; MACN Pv SC4113, m3 izquierdo incompleto.
Procedencia estratigráfica y localidades. MACN Pv SC4112 y 4113 proceden de Portezuelo Sumich Norte (Bown y Larriestra, 1990); MACN Pv SC119c y 123 proceden de niveles indeterminados de las secuencia inferior o media de la formación Pinturas, próximo al valle superior del río Pinturas.
Comentarios. La mayoría de los ejemplares de la Formación Pinturas referidos por Soria (2001) a L. Lacerum son asignados aquí a P. migueli. Sólo dos de esos ejemplares y otros dos obtenidos en colecciones más recientes concuerdan con los tipos de L. lacerum , confirmando la presencia de esta especie en la Formación Pinturas. Los molares superiores difieren de los de P. migueli por ser ligeramente más pequeños y por tener cíngulos labiales y mesostilo mucho menos desarrollados. El m3 tiene el mismo tamaño y es tan braquiodonte como el del paratipo de L. lacerum . Esta especie también se registra en rocas de Edad Colhuehuapense de Chubut (Ameghino, 1902; Soria, 1981, 2001)
Tetramerorhinus Ameghino, 1894
Especie tipo. Tetramerorhinus fortis Ameghino, 1894.
Distribución estratigráfica y geográfica. Formación Pinturas (Mioceno Temprano-Medio) y Formación Santa Cruz, Edad Santacrucense (Mioceno Medio) (Soria, 2001), provincia de Santa Cruz, Argentina.
Especies referidas. Tetramerorhinus lucarius Ameghino; Tetramerorhinus cingulatum (Ameghino); Tetramerorhinus mixtum (Ameghino). Tetramerorhinus prosistens (Ameghino); Tetramerorhinus fleaglei Soria, 2001.
Tetramerorhinus prosistens (Ameghino, 1899)
Figura 4.A

Figura 4. Tetramerorhinus spp. A, MACN A 11626, T. prosistens (Holotipo), P3 y P4 izquierdos / ( Holotype ), left P3 and P4. B, MACN Pv SC129, T. fleaglei (Holotipo), P4 izquierdo / ( Holotype ), left P4. C, MACN Pv SC4167, T. fleagle i , P3 - P4 izquierdos / left P3 - P4. D, MACN Pv SC183, T. fleaglei , P2 - P3 izquierdos / left P2 - P3.
Proterotherium prosistens Ameghino, 1899
Heptaconus obcallatus Ameghino, 1899
Diadiaphorus zamius Ameghino, 1899
Holotipo. MACN A 11626, fragmento maxilar derecho con P4 -M3, P3 - P4 y M2 - M3 izquierdos, fragmento mandibular derecho con p4 - m3, p2 derecho aislado, fragmento mandibular izquierdo con p3 - p4, otro con m3 y fragmento de un m1 o m2.
Material referido. Véase apéndice.
Procedencia estratigráfica y localidades. El holotipo, los tipos de los sinónimos y MACN A 52-601 proceden de una localidad y nivel indeterminado asignado por Ameghino (1906) a las Capas con Astrapothericulus ; MACN Pv SC177 proviene de cerro de los Monos; MACN Pv SC169 proviene de Los Toldos Norte; MACN Pv SC170 y 4108 provienen de la estancia Ana María; MACN Pv SC149-151, 4125 y 4130 proviene del portezuelo Sumich Sur; MACN Pv SC153, 157, 159, 174-176, 179 y 4131 provienen del portezuelo Sumich Norte (Bown y Larriestra, 1990); MACN Pv SC163-165, 168, 181 y 4124 provienen de niveles indeterminados de la secuencia media o inferior de la Formación Pinturas en el área del valle del río Pinturas.
Comentarios. Soria (2001) señaló que Tetramerorhinus lucarius (la especie de menor talla del género) sólo se registra en la Formación Santa Cruz, pero incluyó cuatro molares aislados procedentes de la Formación Pinturas en el hipodigma de esta especie. Estos molares son ligeramente más pequeños que los del tipo de T. prosistens , pero los superiores presentan remanentes de los pliegues labiales del paracono y metacono, por lo que no corresponden a T. lucarius , y pueden estar incluidos dentro del rango de tamaños de T. prosistens ; los molares inferiores no presentan caracteres diagnósticos. Concluimos que no existen evidencias positivas que indiquen la presencia de T. Lucarius en la Formación Pinturas.
Tetramerorhinus fleaglei Soria, 2001 (nuevo rango)
Figuras 4.B-D
Tetramerorhinus cingulatum fleaglei Soria, 2001.
Holotipo. MACN Pv SC129, fragmentos maxilares izquierdo y derecho asociados con P4 - M2.
Materiales referidos. Véase apéndice.
Procedencia estratigráfica y localidades. El holotipo y MACN Pv SC130 - 134, 159, 183, 4175, 4203 y 4204 proceden del portezuelo Sumich Norte; MACN Pv SC178, 4167,4169, 4170, 4172, 4198, 4200 - 4202, 4205, 4206, 4209, 4212, 4214, 4217, 4219, 4221 y 4222 proceden del cerro de los Monos; MACN Pv SC4177, 4210, 4218 y 4225 proceden de la estancia Ana María; MACN Pv SC143, 144, 4171, 4174, 4176 y 4197 proceden de Los Toldos Norte; MACN Pv SC4196, 4199, 4211, 4224 y 4226 proceden del portezuelo Sumich Sur; MACN Pv SC4173, 4207, 4213 y 4223 proceden de Loma de la Lluvia; MACN Pv SC4168 y 4220 proceden de El Carmen Superior (Bown y Larriestra, 1990); MACN Pv SC136 - 142, 145, 146, 4208, 4215 y 4216 proceden de niveles indeterminados de la secuencia inferior o media de la Formación Pinturas en el valle superior del río Pinturas.
Comentarios. Soria (2001) reconoció este taxón como una subespecie de T. cingulatum , señalando que presentaba molariformes de la misma talla que la subespecie tipo, pero más primitivos por tener conspicuos vestigios de los pliegues labiales del paracono y metacono en los molariformes superiores y por presentar hipocono del P4 y neoparacónido-neoparalófido en los molares inferiores menos desarrollados. Sin embargo, las medidas dentarias dadas por Soria (2001: tablas 8 y 9) indican que el holotipo de T. Fleaglei es más pequeño que T. cingulatum . Otros ejemplares referidos aquí a T. fleaglei son aún más pequeños que el holotipo y apenas difieren en tamaño del de T. prosistens (tabla 5). Soria también indicó que T. fleaglei tiene metacónulo de P3 y P4 comparativamente más grande que en otras especies del género, rasgo aquí interpretado como derivado en función de que el tamaño de esta cúspide es moderado en Lambdaconus lacerum . Por lo tanto, concluimos que T. fleaglei no muestra afinidades en particular con T. cingulatum , lo que justifica su elevación al nivel específico.

Tabla 5. Medidas dentarias comparadas de T. prosistens , T. cingulatum y T. fleaglei (en mm). Los promedios de las medidas de T. Cingulatum se calcularon de los ejemplares referidos por Soria (2001) a esta especie/ Compared dental measurements for T. prosistens , T. Cingulatum and T. fleaglei (in mm). The averages for T. cingulatum calculated from the specimens referred to this species by Soria (2001).
T. fleaglei coincide con T. prosistens (y difiere de las restantes especies del género) en presentar remanentes de los pliegues labiales del paracono y metacono en los molariformes superiores y neoparacónido-neoparalófido poco desarrollado en los molares inferiores. La principal diferencia es el comparativamente mayor desarrollo del hipocono y metacónulo en P3 y P4 de T. fleaglei (figuras 4.B-D). En algunos ejemplares el hipocono es una conspicua cúspide cingular, como en el holotipo (figura 4.B); en otros ejemplares esta cúspide está bien insinuada como un simple cíngulo formando un ángulo posterolingual recto bien definido (figuras 4.C-D). Por lo tanto, el P3 y el P4 tienen contorno cuadrangular, en oposición al subtriangular de T. prosistens (figura 4.A), y pueden ser identificados aún en estados de desgaste avanzados. La mayoría de los P3 y P4 disponibles son referibles a T. fleaglei , por lo que ésta sería la especie de proterotérido más abundante en la Formación Pinturas. Aquellos molariformes sin P3 o P4 asociados y con tamaños intermedios entre los de los holotipos de T. fleaglei y T. prosistens no pudieron ser asignados confiablemente a algunas de estas especies (ver apéndice), pero positivamente no corresponden a alguna de las otras especies del género Tetramerorhinus .
Todos los ejemplares referidos por Soria (2001) a T. fleaglei proceden de la Formación Pinturas, menos el MACN A 206 (un fragmento maxilar con P4 - M2) que procede de la Formación Santa Cruz. Los dientes de este espécimen coinciden aproximadamente en tamaño con los del holotipo de T. fleaglei , pero carecen de los pliegues labiales del paracono y metacono y el metacónulo del P4 es pequeño como en T. cingulatum . Por lo tanto, consideramos que no existen evidencias para referir este ejemplar a T. fleaglei , por lo que esta especie se registra exclusivamente en la Formación Pinturas.
Soria (2001) indicó que T. mixtum se registra en las Formaciones Pinturas y Santa Cruz. El único ejemplar de la Formación Pinturas incluido en el hipodigma de T. mixtum es el MACN Pv SC183, un fragmento maxilar con P2 - P3 (figura 4.D). Este ejemplar es aquí referido a T. fleaglei porque el P3 presenta vestigios de los pliegues labiales y metacónulo grande, en oposición a T. mixtum .
Tetramerorhinus cingulatum ?
Holotipo. MACN A 3065 - 3066, fragmento mandibular derecho con p4 - m3 y fragmento mandibular izquierdo con m2 - m3.
Materiales referidos de la Formación Pinturas. Véase apéndice.
Procedencia estratigráfica y geográfica. Todos los ejemplares proceden de Estancia La Cañada (Bown y Fleagle, 1993)
Comentarios. Estos ejemplares son aquí referidos tentativamente a T. cingulatum (Ameghino, 1891) (= T. fortis Ameghino, 1894) porque se aproximan en tamaño al holotipo de T. cingulatum y a los de las especies sinónimas (Soria, 2001), y son por lo tanto significativamente más grandes que los de T. fleaglei (tabla 6). Los molares inferiores presentan además neoparalófido comparativamente más desarrollado que en T. fleaglei . El único molar superior disponible presenta la corona muy desgastada, por lo que no es posible apreciar caracteres relevantes. T. cingulatum sólo es conocido hasta ahora para la Formación Santa Cruz (Ameghino, 1891; Scott, 1910; Soria, 2001)

Tabla 6. Medidas dentarias comparadas (en mm.) de Tetramerorhinus cingulatum ? de la Formación Pinturas, T. cingulatum y T . fleaglei . Los valores promedio de T. cingulatum y T. fleaglei están tomados de Soria (2001)/ Compared dental measurements for Tetramerorhinus cingulatum ? from the Pinturas Formation, T. cingulatum , and T. fleaglei . The averages for T. cingulatum and T. fleaglei after Soria (2001).
Discusión
Como resultado de esta revisión, concluimos que las únicas especies del género Tetramerorhinus presentes en la Formación Pinturas serían T. prosistens , T. fleaglei y T. cingulatum ? De acuerdo con Soria (2001), T. prosistens es la especie con dentición más primitiva del género por presentar vestigios de los pliegues labiales del paracono y metacono, P3 y P4 sin hipocono y molares inferiores con neoparalófido poco desarrollado, aunque más que en Lambdaconus y Prolicaphrium , por lo que ejemplifica un tipo transicional entre la morfología de las especies del Grupo Oligoceno y las del Grupo Mioceno. La dentición de T. fleaglei también correspondería a un estadio transicional, aunque es más derivado que T. Prosistens por presentar hipocono más desarrollado en los P3 y P4. Soria (2001) interpretó que T. prosistens podría ser la especie antecesora de T. fleaglei , y ésta a su vez de T. cingulatum . Ciertamente, las tres especies estarían representando un gradiente de molarización de los premolares superiores y de desarrollo de un patrón selenodonte en los molares inferiores, asociado con un aumento de tamaño. Sin embargo, al menos T. prosistens y T. fleaglei coexisten en los mismos niveles de la Formación Pinturas, sugiriendo que estas especies no representan un linaje anagenético simple, sino que son el resultado de un evento cladogenético previo a los tiempos de Pinturas.
Las relaciones de las tres especies registradas en la Formación Pinturas con aquellas exclusivas de la Formación Santa Cruz ( T. mixtum y T. lucarius ) son difíciles de establecer. Considerando el tamaño además de los caracteres morfológico mencionados, Soria (2001) propuso que T. mixtum derivaría directamente de T. prosistens , mientras que T. lucarius se habría originado de un ancestro hipotético previo y de menor tamaño que T. prosistens . Sin embargo, T. Mixtum y T. lucarius comparten numerosos rasgos morfológicos derivados (ausencia de pliegues labiales en los molariformes superiores, premolares muy molarizados, hipocono del M3 y neoparalófido de los m1-m3 bien desarrollado) que sugieren una estrecha afinidad. Además, las diferencias de tamaño entre las especies de Tetramerorhinus no son muy significativas y, como se mencionara anteriormente para T. fleaglei , presentan rangos de variaciones importantes. Si bien es válida la apreciación de Soria (2001) de que en los Proterotheriidae se verifica una tendencia evolutiva al aumento de tamaño, este carácter es de utilidad para diferenciar especies cuando está asociado a rasgos morfológicos constantes, pero no puede considerarse decisivo para definir sus relaciones filogenéticas. Considerando los caracteres morfológicos, interpretamos que T. fleaglei es, al menos desde el punto de vista estructural, la especie que se haya más estrechamente relacionada al origen de todas las especies santacrucenses del género ( e.g. T. cingulatum , T. Mixtum y T. lucarius ).
Diadiaphorus Ameghino, 1887
Especie tipo. Diadiaphorus majusculus Ameghino, 1887.
Distribución estratigráfica y geográfica. Formación Pinturas (Mioceno Temprano-Medio) y Formación Santa Cruz, Edad Santacrucense (Mioceno Medio) (Ameghino, 1887, 1906; Scott, 1910; Soria, 2001), provincia de Santa Cruz, Argentina.
Diadiaphorus ? caniadensis sp. nov.
Figura 5.A

Figura 5. Diadiaphorus ? spp. A, MACN Pv SC4241, Diadiaphorus? caniadensis sp. nov. (Holotipo), m1 - m2 y m3 en erupción derechos (el ejemplar incluye p4 y dp4, pero no fueron ilustrados debido a su pobre preservación) / (Holotype), right m1 - m2 and erupting m3 (the specimen includes p4 and dp4, but they are not illustrated due to their poor preservation) . B, MACN Pv SC4239, Diadiaphorus ? sp., P2 izquierdo / left P2.
Etimología. En referencia a Estancia la Cañada, localidad donde se halló el holotipo.
Holotipo. MACN Pv SC4241, fragmento mandibular derecho con raíces del dp4, p4 en erupción, m1 - m2 y m3 en erupción.
Hipodigma. Sólo el holotipo.
Material referido. MACN Pv SC4242, un fragmento mandibular derecho con dp3, raíces del dp4 y porción de la corona de p4.
Procedencia estratigráfica y localidad. El holotipo y el ejemplar referido proceden de Estancia La Cañada (Bown y Fleagle, 1993).
Diagnosis. Proterotérido de tamaño menor que D. majusculus y mayor que T. cingulatum . Molares de corona más bajas que en D. majusculus pero más altas que en Picturotherium migueli y las especies de Lambdaconus y Tetramerorhinus . A diferencia de éstas últimas, los molares inferiores son bastante más largos que anchos, con neoparalófido bien desarrollado y m3 carece de lóbulo posterior, como en D. Majusculus y T. minusculum , pero con entocónido no reducido.
Diagnosis. Proterotheriid smaller than D. Majusculus and larger than T. cingulatum . Molars lower crowned than in D. majusculus , but higher than in Picturotherium migueli and the species of Lambdaconus and Tetramerorhinus . In opposition to the latter ones, the lower molars are much longer than wider, with well-developed neoparalophid, and m3 lacks posterior lobe, as in D. majusculus and Thoatherium minusculum , but with non-reduced entoconid.
Descripción y comparación. Los molares inferiores preservados son algo menores que los de D. Majusculus ( tabla 7 ), de aspecto masivo, más hipsodontes que en las especies de Tetramerorhinus pero menos que en D. majusculus , comparativamente tan largos como en ésta última. El neoparalófido está bien desarrollado, llegando hasta el ángulo anterolingual de la corona; de este modo los molares muestran una condición selenodonte casi tan avanzada como en D. majusculus y Thoatherium minusculum . Sin embargo, a diferencia de estas especies el entocónido se mantiene separado del hipoconúlido en todos los molares. El hipoconúlido del m3 está ubicado en posición lingual y está completamente integrado al hipolofúlido, por lo que la pared posterior del diente no presenta una inflexión que defina un lóbulo posterior, al igual que en D. majusculus y Thoatherium minusculum . Los premolares del holotipo y del ejemplar referido se encuentran pobremente preservados y no se aprecian caracteres morfológicos destacables.

Tabla 7. Medidas de Diadiaphorus ? caniadensis sp. nov. (en mm) / Measurements for Diadiaphorus? caniadensis sp. nov. (in mm) .
Afinidades. D ? caniadensis difiere de las especies de Tetramerorhinus , de Anisolophus y de las del Grupo Oligoceno por presentar molares inferiores de corona comparativamente más alta y más selenodontes y por la ausencia de lóbulo posterior en el m3. Estos caracteres, interpretados como derivados por Soria (2001) lo aproxima a las especies santacrucenses Diadiaphorus majusculus y Thoatherium minusculum . Los molares inferiores de éstos son muy semejantes (Scott, 1910), diferenciándose básicamente por el mayor tamaño y por la condición más masiva de los de Diadiaphorus . D ? caniadensis es aquí referido tentativamente al género Diadiaphorus por aproximarse más en tamaño a D . majusculus y por el aspecto masivo de los molares. La presencia de un entocónido no reducido indicaría una condición más primitiva que la de D . majusculus y T . minusculum (Soria, 2001).
Diadiaphorus? sp.
Figura 5.B
Material referido. MACN Pv SC4239, un fragmento maxilar izquierdo con P2.
Procedencia estratigráfica y localidad. Estancia La Cañada (Bown y Fleagle, 1993).
Descripción y comparación. El único diente preservado se aproxima en tamaño al de T. cingulatum , (DAP = 11,1 mm; DT = 10,2 mm) por lo que corresponde a una especie de talla menor que D ? caniadensis . Este diente se caracteriza por presentar tres cúspides linguales. Las dos cúspides anteriores son subyúgales y están separadas por un valle poco profundo. La cúspide anterior está conectada con el cíngulo anterolingual. La cúspide posterior es la más pequeña y se ubica en el extremo lingual del cíngulo posterior. En la base de la pared lingual del diente se observa un débil cíngulo. La presencia de una estructura oclusal compleja con tres cúspides linguales lo asemeja a algunos ejemplares referidos a Diadiaphorus majusculus y difiere del de todas las especies de Tetramerorhinus .
Familia M acraucheniidae Gill, 1872
Subfamilia C ramaucheniinae Ameghino, 1902
Theosodon Ameghino, 1887
Especies tipo. Theosodon lydekkeri Ameghino, 1887.
Distribución geográfica y estratigráfica. Formación Sarmiento, Edad Colhuehuapense (Mioceno Temprano), provincia de Chubut, Argentina; Formación Pinturas (Mioceno Temprano-Medio) y Formación Santa Cruz, Edad Santacrucense, (Mioceno Medio), provincia de Santa Cruz, Argentina, (Marshall et al. , 1983); ?Formación Chasicó, Edad Chasiquense (Mioceno Tardío) (Bond y López, 1995), provincia de Buenos Aires, Argentina; Formación Chucal, Edad Santacrucense, (Flynn et al. , 2002); Formación Río Frias, Edad Friasense (Mioceno Medio) (Marshall et al. , 1983), Chile; Formación La Victoria, Edad Laventense (Mioceno Medio) (Cifelli y Guerrero, 1997), Colombia.
Especies referidas. Theosodon llalemanti Mercerat; Theosodon fontanae Ameghino; Theosodon gracilis Ameghino; Theosodon patagonicus (Ameghino); Theosodon karaikensis Ameghino; Theosodon garrettorum Scott; Theosodon pozzii Kraglievich y Parodi; Theosodon hystatus Cabrera y Kraglievich; Theosodon ? Frenguellü Soria, 1981.
Theosodon sp.
Material referido. MACN Pv SC4243, corona de un p2 aislada; MACN Pv SC4244, porción anterolingual de un molar inferior derecho; MACN Pv SC4245, porción de un molar superior.
Procedencia estratigráfica y localidades. MACN Pv SC4243 y 4345 proceden de Los Toldos Sur (Bown y Larriestra, 1990); MACN Pv SC4244 procede de Gobernador Gregores, de sedimentos tentativamente referidos a la Formación Pinturas (Bown y Fleagle, 1993).
Comentarios. Las partes preservadas de estos dientes, coinciden en tamaño con los de la especie tipo del género. Sin embargo, los únicos ejemplares disponibles son muy fragmentarios y no permiten una determinación más precisa.
Distribución estratigráfica de los Proterotheriidae de la Formación Pinturas
En aquellas localidades donde los niveles fosilíferos han sido positivamente referidos a las secuencias inferior o media de la Formación Pinturas (Bown y Larriestra, 1990) los Litopterna están representados por Lambdaconus lacerum , Picturotherium migueli , Tetramerorhinus prosistens y Tetramerorhinus fleaglei . En oposición a la interpretación previa de Soria (2001), T. Mixtum y T. lucarius no se registran en esta unidad. P. migueli , T. prosistens y T. fleaglei son hasta ahora registrados exclusivamente en la Formación Pinturas. L. Lacerum también se registra en niveles colhuehuapenses de la Formación Sarmiento, en la provincia de Chubut (Ameghino, 1902; Soria, 1981, 2001) y constituye el más primitivo proterotérido hasta ahora conocido (Soria, 2001). P. migueli está relacionado a la Serie Thoatherium pero comparte con las especies deseadenses y colhuehuahpenses numerosos rasgos dentarios más plesiomórficos que Thoatherium . Tanto T. prosistens como T. fleaglei son más primitivos que sus cogenéricos santacrucenses por tener vestigios de pliegues labiales del paracono y metacono y neoparalófido menos desarrollado. Esta asociación sugiere que al menos las secuencias inferior y media de la Formación Pinturas son más antiguas que la Formación Santa Cruz en sus localidades típicas, coincidiendo con lo observado para los roedores histricognatos y marsupiales paleoténtidos (Kramarz, 2001b, Kramarz y Bellosi, 2005), y apoya la hipótesis original de Ameghino.
Los sedimentos expuestos en La Cañada, al sureste del área del valle del río Pinturas (figura 1), fueron referidos a la parte más alta de la secuencia media de la Formación Pinturas (Bown y Fleagle, 1993). Sin embargo, los roedores histricognatos y los marsupiales paleoténtidos recuperados en La Cañada están representados por especies más derivadas que aquellas positivamente registradas en la parte más alta de la secuencia media en el área tipo (portezuelo Sumich Sur), y muchas de ellas también son conocidas en las típicas localidades costeras de la Formación Santa Cruz (Bown y Fleagle, 1993; Rae et al. , 1996; Kramarz, 2001b). Estas evidencias sugieren que los niveles fosilíferos de La Cañada son más jóvenes que los de la secuencia media de la Formación Pinturas, y que podrían correlacionarse con los de la secuencia superior de esta unidad en su localidad tipo (Kramarz, 2001b). Los proterotéridos recuperados en la estancia La Cañada (aquí referidos tentativamente a taxones santacrucenses Tetramerorhinus cingulatum y Diadiaphorus ) apoyan la hipótesis de una menor antigüedad de estos depósitos, ya que, al igual que los marsupiales y roedores, presentan morfotipos dentarios más derivados que los de las secuencias inferior y media.
Consideraciones sobre los Macraucheniidae de la Formación Pinturas y su paleoambiente
Las evidencias sedimentológicas, paleopedológicas e icnológicas indican que las secuencias inferior y media de la Formación Pinturas se habrían depositado en un ambiente árido a subhúmedo dominado por vegetación herbácea (Bown y Larriestra, 1990; Genise y Bown, 1994; Kramarz y Bellosi, 2005). Sin embargo, los datos palinológicos (Zamaloa, 1993) y a la abundancia y diversidad de primates platirrinos (Fleagle et al. , 1987; Fleagle, 1990; Tejedor, 2002) y roedores eretizóntidos y cavioideos protohipsodontes (Kramarz, 2001b, 2004), sugieren la presencia de un ambiente cálido y húmedo dominado por vegetación arbórea. Estas evidencias contradictorias podrían ser interpretadas como producto de severas fluctuaciones climáticas y/o variaciones locales y ponen en evidencia la complejidad de los ambientes en que se desarrollaron las secuencias inferior y media de la Formación Pinturas (Kramarz y Bellosi, 2005). Por el contrario, todas las evidencias sedimentológicas (Bown y Larriestra, 1990) icnológicas (Genise y Bown, 1994) y paleontológicas (Kramarz, 2001b) son coherentes en indicar una marcado deterioro climático para la secuencia superior. Los Macraucheniidae son extremadamente raros en Pinturas; sólo tres dientes fragmentarios fueron recuperados hasta ahora. Uno procede de Gobernador Gregores y los otros de Toldos Sur. Como en La Cañada, los roedores y los marsupiales paleoténtidos de Gobernador Gregores y Los Toldos Sur sugieren que los sedimentos expuestos en estas localidades serían más modernos que los de las secuencias inferior y media de la Formación Pinturas y podrían corresponder a su secuencia superior (Kramarz, 2001b, Kramarz y Bellosi, 2005). Por lo tanto, ningún macrauquénido se ha registrado hasta ahora en las secuencias inferior y media, mientras que allí los proterotéridos son diversos y muy abundantes (en estos niveles se obtuvieron restos correspondientes a 27 individuos, de acuerdo a la cantidad de los m3 recuperados). El tamaño de la muestra es suficientemente grande como para interpretar que la ausencia de macrauquénidos no se debe a un mero defecto del registro, aun tomando en cuenta que los macrauquénidos usualmente son menos abundantes que los proterotéridos en otras faunas (por ejemplo, las colecciones santacrucenses de Ameghino depositadas en el MACN contienen 459 ejemplares catalogados referidos a Proterotheriidae y 142 a Macraucheniidae). Podría interpretarse que la ocurrencia de macrauquénidos restringidos a la secuencia superior de la Formación Pinturas está relacionada con los ambientes más áridos y abiertos inferidas para esos niveles. Sin embargo, los macrauquénidos están presentes en todas las principales faunas conocidas del Terciario Medio y Tardío y del Cuaternario de América del Sur, sugiriendo que la distribución geográfica de estos mamíferos no habría estado condicionado por restricciones ambientales. Por el contrario, el registro de proterotéridos parece estar relacionado a aquellas faunas asociadas a hábitats más forestados (Cifelli y Guerrero, 1997; Bond et al. , 2001). La asociación de mamíferos de las secuencias inferior y media de la Formación Pinturas es la única del neógeno conocida a través de un gran número de ejemplares en la cual no se registraron macrauquénidos pero sí proterotéridos. La al menos limitada ocurrencia de macrauquénidos podría ser resultado de las peculiares condiciones ambientales, aún no bien establecidas, que caracterizan las secuencias inferior y media de la Formación Pinturas. Ciertamente, no se pueden establecer con certeza inferencias paleoambientales hasta que la paleobiología y los requerimientos ecológicos de los macrauquénidos miocénicos sean mejor conocidos.
Agradecimientos
A G. Vucetich (Museo de La Plata) por su permanente apoyo. A J. Fleagle (Stony Brook University, EE.UU.) y a T. Bown , cuya labor ha resultado en el enriquecimiento de las colecciones de Paleontología de Vertebrados del MACN. A los revisores D. Croft (Case Western Reserve University, EE.UU.) y M. Reguero (Museo de La Plata) por sus valiosos comentarios y sugerencias. Las ilustraciones fueron realizadas por el artista Jorge González.
Bibliografía
Ameghino, F. 1887. Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de Patagonia austral y depositados en el museo de La Plata. Boletín del Museo de La Plata 1: 1-26. [ Links ]
Ameghino, F. 1891. Nuevos restos de mamíferos fósiles descubiertos por Carlos Ameghino en el Eoceno inferior de la Patagonia austral. Nuevas especies, adiciones y correcciones. Revista Argentina de Historia Natural 1: 289-328. [ Links ]
Ameghino, F. 1894. Enumération synoptique des espèces de mammifères fossiles des formations éocènes de Patagonie. Boletín de la Academia Nacional de Ciencias (Córdoba) 13: 259-455. [ Links ]
Ameghino, F. 1897. Mamíferos cretáceos de la Argentina. Segunda contribución al conocimiento de la fauna mastológica de las capas con restos de Pyrotherium. Boletín Instituto Geográfico Argentino 18: 406-429, 431-521. [ Links ]
Ameghino, F. 1899. Sinópsis geológico-paleontológica. Suplemento (Adiciones y correciones) 1: 13 pp., La Plata. [ Links ]
Ameghino, F. 1900-1902. L' age des formations sédimentaires de Patagonie. Anales de la Sociedad Cientítica Argentina 50: 109-131, 145-165, 209-229; 51: 20-39, 65-91; 52: 198-197, 244-240; 54: 161-180, 220-240, 283-342. [ Links ]
Ameghino, F. 1902. Première contribution à la connaissance de la faune mammalogique des couches à Colpodon. Boletín de la Academia Nacional de Ciencias (Córdoba) 17: 71-138. [ Links ]
Ameghino, F. 1904. Recherches de morphologie phylogénétique sur les molaires supérieures des ungulés. Anales del Museo Nacional de Buenos Aires 3: 1-541. [ Links ]
Ameghino, F. 1906. Les formations sédimentaires du Crétacé supérieur et du Tertiaire de Patagonie. Anales del Museo Nacional de Buenos Aires 8: 1-568. [ Links ]
Bianchini, L.H. y Bianchini, J.J. 1971. Revisión de los Proterotheriidae (Mammalia, Litopterna) del Mesopotamiense. Ameghiniana 8: 1-24. [ Links ]
Bond, M. y López, G.M. 1995. Los Macraucheniidae (Mammalia, Litopterna) de la Formación Arroyo Chasicó (Partido de Villarino, Pcía. de Buenos Aires). 4º Jornadas Geológicas y Geofísicas Bonaerenses (Junin), Actas 1: 18-22. [ Links ]
Bond, M., Perea, D., Ubilla, M. y Tauber, A. 2001. Neolicaphrium recens Frenguelli, 1921, the only surviving Proterotheriidae (Litopterna, Mammalia) into the South American Pleistocene. Palaeovertebrata 30: 37-50. [ Links ]
Bown, T.M. y Fleagle, J.G. 1993. Systematics, Bioestratigraphy and Dental Evolution of the Palaeothentidae, Later Oligocene to Early- Middle Miocene (Deseadan-Santacrucian) Caenolestoid Marsupials of South America. The Paleontological Society Memoir 29: 1-76. [ Links ]
Bown, T.M. y Larriestra, C.N. 1990. Sedimentary paleoenvironments of fossil platyrrhine localities, Miocene Pinturas Formation, Santa Cruz province, Argentina. Journal of Human Evolution 19: 87-119. [ Links ]
Bown, T.M., Larriestra, C.N. y Powers, D.W. 1988. Análisis paleoambiental de la formación Pinturas (Mioceno Inferior), Provincia de Santa Cruz. 2º Reunión Argentina de Sedimentología (Buenos Aires) , Actas 1: 31-35. [ Links ]
Castellanos, A. 1937. Ameghino y la antigüedad del hombre sudamericano. Asociación Cultural de Conferencias de Rosario, Ciclo de Carácter General 2: 47- 192. [ Links ]
Cifelli, R.L. y Guerrero, J.G. 1997. Litopterns. En: R. Kay, R. Madden, R. Cifelli y J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene fauna of La Venta, Colombia , Smithsonian Institution Press, Washington and London, pp. 289-302. [ Links ]
de Barrio, R.E., Scillato Yané, G. y Bond, M. 1984. La Formación Santa Cruz en el borde occidental del macizo del Deseado (provincia de Santa Cruz) y su contenido paleontológico. 9º Congreso Geológico Argentino (S. C. de Bariloche), Actas 4: 539-556. [ Links ]
Dumont, E.R., Strait, S.G., y Friscia, A.R. 2000. Abderitid marsupials from the Miocene of Patagonia: an assessment of form, function and evolution. Journal of Paleontology 74: 1161-1172. [ Links ]
Fleagle, J.G. 1990. New fossil platyrrhine from the Pinturas formation, southern Argentina. Journal of Human Evolution 19: 61-85. [ Links ]
Fleagle, J.G., Powers, D.W., Conroy, G.C. y Watters, J.P. 1987. New fossil platyrrhine from Santa Cruz province, Argentina. Folia Primatologica 48: 65-77. [ Links ]
Fleagle, J.G., Bown, T.M., Swisher, C. y Buckley, G. 1995. Age of the Pinturas and Santa Cruz formations. 6º Congreso Argentino de Paleontología y Bioestratigrafía (Trelew), Actas 129-135. [ Links ]
Flynn, J.J., Croft, D.A., Charrier, R., Hérail, G. y Wyss, A.R. 2002. The first Cenozoic mammal fauna from the Chilean Altiplano. Journal of Vertebrate Paleontology 22: 200-206. [ Links ]
Frenguelli, J. 1931. Nomenclatura estratigráfica patagónica. Anales de la Sociedad de Ciencias de Santa Fe 3: 1-115. [ Links ]
Genise, J.F. y Bown, T.M. 1994. New Miocene scarabeid and hymenopterous nest and Early Miocene (Santacrucian) paleoenviromnts, Patagonian Argentina. Ichnos 3: 107-117. [ Links ]
Goloboff, P.A. 1993. NONA 2.0, MS-DOS tree-searching program and documentarion. Publicado por el autor, Argentina. [ Links ]
Kramarz A.G. 2001a. Prostichomys bowni , un nuevo roedor Adelphomyinae (Hystricognathi, Echimyidae) del Mioceno medio - inferior de Patagonia, Argentina. Ameghiniana 38: 163-168. [ Links ]
Kramarz, A.G. 2001b. [ La fauna de roedores de la Formación Pinturas, Mioceno medio-inferior de la provincia de Santa Cruz . Tesis doctoral inédita. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires, 243 pp. Inédita]. [ Links ]
Kramarz, A.G. 2002. Roedores chinchilloideos (Hystricognathi) de la Formación Pinturas, Mioceno temprano-medio de la provincia de Santa Cruz, Argentina. Revista del Museo Argentino de Ciencias Naturales, (nueva serie) 4: 167-180. [ Links ]
Kramarz, A.G. 2004. Octodontoids and erithizontoids (Rodentia, Hystricognathi) from the Pinturas Formation, Early-Middle Miocene of Patagonia, Argentina. Ameghiniana 41: 199-216. [ Links ]
Kramarz, A.G. y Bellosi, E.S. 2005. Hystricognath rodents from the Pinturas Formation, Early-Middle Miocene of Patagonia, biostratigraphic and palaeoenvironmental implications. Journal of South American Earth Sciences 18: 199-212. [ Links ]
Marshall, L.G. 1976. Fossil localities for Santacrucian (early Miocene) mammals, Santa Cruz province, southern Patagonia, Argentina. Journal of Paleontology 50: 1129-1142. [ Links ]
Marshall, L.G., Hoffstetter, R. y Pascual, R. 1983. Mammals and stratigraphy: geochronology of continental mammals-bearing Tertiary of South America. Paleovertebrata , Memoria extraordinaria 1-93. [ Links ]
Marshall, L.G., Pascual, R., Curtis, G.H. y Drake, R.E. 1977. South American geochronology: radimetric time scale for middle to late Tertiary mammals bearing horizons in Patagonia. Science 195: 1325- 1328. [ Links ]
Pascual, R. y Odreman Rivas, O.E. 1971. Evolución de las comunidades de vertebrados del Terciario argentino. Los aspectos paleozoogeográficos y paleoclimáticos relacionados. Ameghiniana 8: 372-412. [ Links ]
Pascual, R., Ortega Hinojosa, E., Gondar, E. y Tonni, E. 1965. Las edades del Cenozoico mamalífero de la Argentina, con especial atención a aquellas del territorio bonaerense. Anales de la Comisión de Investigaciones Científicas 6: 165-193. [ Links ]
Rae, T.C., Bown, T.M. y Fleagle, J.G., 1996. New Palaeothentid Marsupials (Caenolestoidea) from the Early Miocene of Patagonian Argentina. American Museum Novitates 3165: 1-10. [ Links ]
Scott, W.B. 1910. Mammalia of the Santa Cruz beds. Part I: Litopterna. R eports of the Princeton Expedition to Patagonia 7: 1-156. [ Links ]
Simpson, G.G. 1940. Review of the mammal-bearing Tertiary of South America. Proceedings of the American Philosophy Society 83: 649-709. [ Links ]
Soria, M.F. (h). 1981. Los Liptoterna del Colhuehuapense (Oligoceno Tardío) de la Argentina. Revista del Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Serie Paleontología 3: 1-54. [ Links ]
Soria, M.F. (h). 2001. Los Proterotheriidae (Litopterna, Mammalia), sistemática, origen y filogenia. Monografías del Museo Argentino de Ciencias Naturales 1: 167 pp. [ Links ]
Tejedor, M.F. 2002. Primate canines from the early Miocene Pinturas Formation, Southern Argentina. Journal of Human Evolution 43: 127-141. [ Links ]
Wood, A.E. y Patterson, B. 1959. Rodents of the Deseadan Oligocene of Patagonia and the beginnings of South American rodent evolution. Bulletin of the Museum of Comparative Zoology 120: 281-428. [ Links ]
Zamaloa, M.C. 1993. Hallazgos palinológicos en la Formación Pinturas, sección Cerro de los Monos (Mioceno inferior), provincia de Santa Cruz, Argentina. Ameghiniana 30: 353. [ Links ]
Recibido: 19 de febrero de 2004.
Aceptado: 28 de diciembre de 2004.
Apéndice
Lista de ejemplares referidos a Picturotherium migueli gen. et sp. nov.: MACN Pv SC119 a, m3 derecho; MACN Pv SC119 b, fragmento mandibular izquierdo con m2 - m3; MACN Pv SC119 c, P1 derecho; MACN Pv SC119 d, P2 derecho; MACN Pv SC120, fragmento mandibular derecho con m1 y m2 en erupción; MACN Pv SC121, P4 derecho incompleto; MACN Pv SC122, dp4? izquierdo; MACN Pv SC124, m1 o m2 derecho; MACN Pv SC125, p4 derecho incompleto; MACN Pv SC126, M1 o M2 derecho incompleto; MACN Pv SC4114, M1 o M2 derecho incompleto; MACN Pv SC4115, M1 o M2 izquierdo incompleto; MACN Pv SC4116, M3 derecho; MACN Pv SC4117, p4 derecho; MACN Pv SC4118, p3 izquierdo.
Lista de ejemplares referidos a Tetramerorhinus prosistens Ameghino: MACN A 52-601, fragmento maxilar derecho con P4 -M3 de un individuo senil; MACN A 11625 (holotipo de H . Obcallatus Ameghino), fragmento maxilar derecho con DP1 - DP4, fragmento maxilar izquierdo con DP1 - DP2, DP4 izquierdo y ambos M1 asociados; MACN SC149, fragmento mandibular izquierdo con m1 - m2; MACN SC150, fragmento mandibular con m3; MACN SC151, m3 derecho; MACN SC153, fragmento maxilar derecho con M1 - M2; MACN SC157, m3 izquierdo; MACN SC159, P4 derecho; MACN SC163, fragmento mandibular derecho con p4?, MACN SC164, fragmento mandibular derecho con dp3 incompleto, dp4 -m1; MACN SC165, fragmento mandibular derecho con m3; MACN SC168, m1 o m2 izquierdo; MACN SC169, m3 incompleto; MACN SC170, fragmento mandibular izquierdo con p4; MACN Pv SC174, M3 izquierdo; MACN SC175, fragmento mandibular izquierdo con m1 - m2; MACN Pv SC176, M1 o M2 derecho; MACN Pv SC177, m1 o m2 derecho; MACN SC179, fragmento mandibular derecho con m2 - m3; MACN SC181, fragmento mandibular derecho con m3; MACN Pv SC4108, fragmento mandibular derecho con p4, m1 y m3 derechos, m1 y p3 izquierdos, P1, P2 y P3 derechos, P2, P4 y M3 izquierdos asociados; MACN Pv SC4124, P3 izquierdo; MACN Pv SC4125, 3 M3 aislados; MACN Pv SC4130, 3 M3 aislados; MACN Pv SC4131, 3 M3 incompletos aislados; MLP 82-VI-3-2, fragmento mandibular derecho con p3 - m3 y fragmento mandibular izquierdo p3 - m3 asociados.
Lista de ejemplares referidos a Tetramerorhinus fleaglei Soria: MACN Pv SC130, fragmento mandibular izquierdo con m2 - m3; MACN Pv SC131, P4? derecho; MACN Pv SC132, M3 derecho; MACN Pv SC133, P4 izquierdo incompleto; MACN Pv SC134, m1 o m2 derecho; MACN Pv SC136, P4 derecho incompleto; MACN Pv SC137, M3 izquierdo; MACN Pv SC138, m1 o m2; MACN Pv SC139, m3 izquierdo; MACN Pv SC140, m3 izquierdo; MACN Pv SC141, fragmento mandibular derecho con m1 o m2; MACN Pv SC142, fragmento mandibular izquierdo con m1 o m2; MACN Pv SC143, fragmento mandibular izquierdo con p3 - p4; MACN Pv SC144, M3 izquierdo incompleto; MACN Pv SC145, m1 o m2 derecho incompleto; MACN Pv SC146, fragmento mandibular derecho con m3; MACN Pv SC178, fragmento maxilar derecho con P3 - P4; MACN Pv SC183, fragmento maxilar izquierdo con P2 - P3; MACN Pv SC4167, fragmento maxilar izquierdo con P4 - P3; MACN Pv SC4168, fragmento maxilar derecho con M2 - M3 y P4 asociado; MACN Pv SC4169, fragmento maxilar derecho con P3 y raíces del P2; MACN Pv SC4170, fragmento maxilar izquierdo con P2 - P3; MACN Pv SC4171, P3 izquierdo; MACN Pv SC4172, fragmento maxilar izquierdo con P2 - P3; MACN Pv SC4173, P3 derecho; MACN Pv SC4174, P4 derecho; MACN Pv SC4175, P4 izquierdo; MACN Pv SC4176, P4 derecho; MACN Pv SC4177, P4 izquierdo; MACN Pv SC4196, P3? izquierdo; MACN Pv SC4197, P3? izquierdo; MACN Pv SC4198, P3? derecho; MACN Pv SC4199, P3 derecho; MACN Pv SC4200, fragmento maxilar derecho con P3 y raíces del P2; MACN Pv SC4201, fragmento maxilar izquierdo con M1 - M2 y M3 incompleto; MACN Pv SC4202, M3 derecho; MACN Pv SC4203, M1 o M2 izquierdo; MACN Pv SC4204, M3 izquierdo incompleto; MACN Pv SC4205, fragmento maxilar derecho con DP2; MACN Pv SC4206, M3 izquierdo; MACN Pv SC4207, M3 derecho; MACN Pv SC4208, fragmento maxilar derecho con M2 - M3; MACN Pv SC4209, M3 izquierdo; MACN Pv SC4210, M3 izquierdo; MACN Pv SC4211, P2 derecho; MACN Pv SC4212, fragmento maxilar derecho con M1 - M2; MACN Pv SC4213, P2 derecho; MACN Pv SC4214, M3 derecho; MACN Pv SC4215, fragmento mandibular derecho con raíces de los m1 - m2 y m3 incompleto; MACN Pv SC4216, fragmento mandibular izquierdo con m1 - m3; MACN Pv SC4217, fragmento mandibular derecho con m2 - m3; MACN Pv SC4218, fragmento mandibular izquierdo con m1 - m2; MACN Pv SC4219, fragmento mandibular izquierdo con p3 incompleto y p4; MACN Pv SC4220, fragmento mandibular derecho con m1?; MACN Pv SC4221, fragmento mandibular derecho con m2 incompleto; MACN Pv SC4222, fragmento mandibular izquierdo con p3; MACN Pv SC4223, fragmento mandibular izquierdo con p3; MACN Pv SC4224, p3 derecho; MACN Pv SC4225, dos m3 aislados; MACN Pv SC4226, m1 o m2 izquierdo.
Lista de ejemplares referibles a Tetramerorhinus prosistens o a T. fleaglei : MACN SC148, fragmento mandibular izquierdo con m1 o m2; MACN SC180, fragmento maxilar derecho con DP2 incompleto, DP3 y DP4; MACN SC182, fragmento maxilar izquierdo con DP3 - DP4; MACN Pv SC4126, 3 M3 aislados; MACN Pv SC4127, fragmento maxilar derecho con M3; MACN Pv SC4128, M3 derecho; MACN Pv SC4129, M3 izquierdo; MACN Pv SC4132, M1 o M2 izquierdo; MACN Pv SC4133, M1 o M2 derecho; MACN Pv SC4134, dos molares superiores derechos incompletos probablemente asociados; MACN Pv SC4135, M1 o M2 derecho; MACN Pv SC4136, 3 M1 o M2 aislados; MACN Pv SC4137, m3 derecho incompleto; MACN Pv SC4138, 4 m3 aislados; MACN Pv SC4139, 2 m3 aislados; MACN Pv SC4140, 6 m3 aislados; MACN Pv SC4141, m3; MACN Pv SC4142, 4 m3 aislados; MACN Pv SC4143, 3 m3 aislados; MACN Pv SC4144, fragmento mandibular izquierdo con m3; MACN Pv SC4145, m3 izquierdo; MACN Pv SC4146, 3 m3 aislados; MACN Pv SC4147, fragmento mandibular izquierdo con m1; MACN Pv SC4148, m1 o m2 derecho; MACN Pv SC4149, m1 o m2 derecho; MACN Pv SC4150, m1 o m2 derecho; MACN Pv SC4151, 2 m1 o m2 aislados; MACN Pv SC4152, 3 m1 o m2 aislados; MACN Pv SC4153, 4 m1 o m2 aislados; MACN Pv SC4154, p4 derecho; MACN Pv SC4155, 4 p4 aislados; MACN Pv SC4156, 2 p4 aislados; MACN Pv SC4157, p4 izquierdo; MACN Pv SC4158, 4 p3 aislados; MACN Pv SC4159, p3 izquierdo; MACN Pv SC4160, 2 p3 aislados; MACN Pv SC4161, p3 izquierdo; MACN Pv SC4162, p3 derecho; MACN Pv SC4163, 3 p2 aislados; MACN Pv SC4164, p2 derecho; MACN Pv SC4165, p2 izquierdo; MACN Pv SC4166, 4 p2 aislados; MACN Pv SC4178, fragmento mandibular derecho con m1 y raíces de dp3 - dp4; MACN Pv SC4179, fragmento mandibular derecho con m1 y raíces de p3 - p4; MACN Pv SC4180, fragmento mandibular derecho con p4 - m2 y raíces del m3; MACN Pv SC4181, fragmento mandibular izquierdo con p4 - m1; MACN Pv SC4182, fragmento mandibular derecho con p3 y p4 incompleto, alvéolos para el p2 y raíces del m1; MACN Pv SC4183, fragmento mandibular izquierdo con m1; MACN Pv SC4184, fragmento mandibular derecho con p3 - p4; MACN Pv SC4185, fragmento mandibular izquierdo con m2 incompleto y m3; MACN Pv SC4186, fragmento mandibular izquierdo con p2 - p3 y p4 incompleto; MACN Pv SC4187, fragmento mandibular derecho con m3; MACN Pv SC4188, fragmento mandibular izquierdo con m3; MACN Pv SC4189, fragmento mandibular izquierdo con m1 o m2; MACN Pv SC4190, fragmento mandibular izquierdo con p4 - m1; MACN Pv SC4191, fragmento mandibular derecho con m2 y m3; MACN Pv SC4192, fragmento maxilar derecho con M3; MACN Pv SC4193, fragmento maxilar derecho con M2 - M3; MACN Pv SC4194, fragmento maxilar izquierdo con P2 - P4 y M1 incompleto.
Lista de ejemplares referidos tentativamente a Tetramerorhinus cingulatum Ameghino de la Formación Pinturas: MACN Pv SC4232, fragmento mandibular derecho con m3 y talónido del m2; MACN Pv SC4233, fragmento mandibular derecho con m3 y talónido del m2; MACN Pv SC4234, fragmento mandibular izquierdo con m3; MACN Pv SC4235, M1 o M2 izquierdo incompleto; MACN Pv SC4236, p3 izquierdo; MACN Pv SC4237, fragmento mandibular izquierdo con p3.

