










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.3 Buenos Aires jun./sept. 2005
Nueva evidencia sobre el modo de vida de los Conulariida
Nora Sabattini 1 y Julio C. Hlebszevitsch 2
1 Consejo Nacional de Investigaciones Científicas y Técnicas-Departamento Paleozoología Invertebrados, Museo de Ciencias Naturales. Paseo del Bosque s/n, 1900 La Plata. Argentina. nsabatti@museo.fcnym.unlp.edu.ar
2 Administración Yacimientos Petrolíferos Fiscales, Avenida del Libertador, Comodoro Rivadavia, 9000 Chubut. Argentina. jchlebszevitschs@repsolypf.com
Abstract. N ew evidence about the mode of life of C onulariida . The occurrence of one specimen of Paraconularia sp. with the apex connected to a crinoid cirrus is described and figured for the first time. This disposition suggests that the conulariid had a primary, apical attachment to the crinoid. The material comes from Puesto La Carlota (Chubut Province, Argentina) and belongs to the Callitomaria tepuelensis-Streblochondria sp. Zone (Early Permian).
Resumen. Por primera vez se describe e ilustra el hallazgo de un ejemplar de Paraconularia sp. con su ápice unido al cirro de un crinoideo. La disposición de los organismos sugiere que el conulárido estuvo originariamente fijo al crinoideo. El material estudiado proviene del Puesto La Carlota (Provincia del Chubut, Argentina) y pertenece a la Zona de Callitomaria tepuelensis-Streblochondria sp. (Pérmico temprano).
Key words. Conulariida. Paleoecology. Early Permian. Chubut. Argentina
Palabras clave. Conulariida. Paleoecología. Pérmico temprano. Chubut. Argentina.
Introducción
No son frecuentes en la literatura los registros de evidencias bien documentadas e ilustradas del modo de vida de los conuláridos. Algunos autores proponen una forma de vida bentónica sésil, habiendo citado hallazgos que indicarían que los conuláridos fueron organismos epifaunales que habrían pasado, al menos parte de sus vidas, fijos por su ápice al fondo del mar o a esqueletos de otros invertebrados. También se conocen algunas observaciones de conuláridos que habrían vivido fijos o inmersos en el esqueleto de otros invertebrados.
El objetivo de esta contribución es dar a conocer, en el Paleozoico superior de la Cuenca Tepuel-Genoa, localidad de Puesto La Carlota (figura 1), el hallazgo de un conulárido fijo a cirrales de crinoideos, lo que aportaría una nueva evidencia a la teoría de un modo de vida bentónico sésil para dicho grupo.
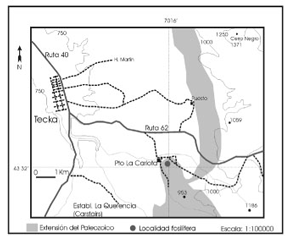
Figura 1. Mapa de ubicación de la localidad Puesto La Carlota, Provincia del Chubut / Location map of La Carlota locality, Chubut Province.
Este hábito, así como la presencia de conuláridos creciendo interrelacionados con calcisponjas, sería para Van Iten (1991a, pág. 153) una evidencia adicional a la relación de este grupo con ciertos Scyphozoa Coronatida que muestran el mismo tipo de relación con poríferos vivientes, así como con los pólipos de Scyphozoa, que son organismos sésiles que viven fijos a un substrato que puede ser el fondo o esqueletos de otros organismos.
Importantes evidencias sobre la relación de los Conulariida con los Scyphozoa son la simetría tetrámera, exhibida mediante la presencia de septos o espesamientos internos a lo largo de la línea media, homólogos a los cuatro septos gástricos de los Scyphozoa (Bischoff, 1978; Van Iten, 1992a; Jerre, 1994), las evidencias de gemaciones (Van Iten y Cox, 1992; McKinney et al ., 1995) y la microestructura y modo de crecimiento del peridermo (Van Iten, 1992b).
El material se halla depositado en el Departamento Paleozoología Invertebrados del Museo de Ciencias Naturales de La Plata, bajo la sigla MLP.
Antecedentes
Ruedemann (1896, lám. 8, figs. 1-3) describe e ilustra especímenes de Conularia gracilis Hall que presentan ejemplares juveniles, fijos a su peridermo, también ejemplares de C . cf. gracilis fijos a un fragmento de conchilla (Ruedemann, 1896, lám. 8, fig. 5) y un espécimen de C. gracilis sobre un gastrópodo: Trochonema cf. ammonius Hall (Ruedemann 1896, lám. 9, fig. 1). Pero todos, excepto uno de los especimenes de C . gracilis en Ruedemann (1896) fueron reasignados por Van Iten (1994, p. 365) a un género de dudosa ubicación sistemática: Sphenothallus may (Van Iten et al. 1992; Neal y Hannibal, 2000).
Finks (1955, p. 832, lám. 82) registró una asociación fósil de esponjas y conuláridos, ilustrando la existencia de dos ejemplares de poríferos con moldes de Conularia leonardensis Finks; debido a la disposición espacial y orientación de los conuláridos, el autor sugiere que fueron sésiles, mostrando una posible cicatriz de fijación en uno de los ejemplares (Finks, 1955, lám. 82, fig. 9). Finks (1960, lám. 12, fig. 6) ilustra moldes de Conularia sp. cf. C. leonardensis rodeados por una esponja: Defordia defuncta King. También ilustra un porífero: Haplistion aeluroglossa Finks conteniendo dos moldes externos de conuláridos de la misma especie (Finks, 1960, lám. 27, fig. 1), mencionando que los moldes son perpendiculares a la superficie superior del porífero y se extienden en su interior, lo cual indicaría que los conuláridos se habrían fijado al porífero cuando éste aún vivía (Finks, 1960, p. 90).
Kozlowski (1968) describe e ilustra varios ejemplares juveniles de Conularia sp., los cuales tienen el ápice abierto y estarían fijos directamente por la extremidad del peridermo. También registra (Kozlowski, 1968, fig. 19) un ejemplar bastante grande con dos fragmentos de ápices, de dos individuos, fijos mediante espinas. Asimismo ilustra varios ejemplares con prolongaciones a modo de espinas en la región apical; no obstante señala que la fijación sería transitoria y los adultos habrían vivido libres (Kozlowski, 1968, p. 521).
Según Babcock y Feldmann (1986a, p. 370) los conuláridos podrían haber sido pseudoplanctónicos, fijándose a algas planctónicas, o haberse fijado a remanentes de plantas previamente asentadas en el fondo del mar, ilustrando (Babcock y Feldmann, 1986b fig. 28.7) dos ejemplares de P. yochelsoni Babcock y Feldmann asociados a remanentes de plantas, tal vez algas. Además, estos mismos autores (1986b, figs. 24.1-3 y 32.5) registran ejemplares de Paraconularia chesterensis (Worthen) con estructuras tubulares, ubicadas cerca de los ápices, a las que interpretan como tallos de fijación.
Harland y Pickerill (1987) analizando un conulárido del Ordovícico de Québec, con epizoicos fijos a su peridermo, concluyen, sobre la base de la inclinación en el sedimento de aguas profundas donde se halló enterrado el fósil, que lo más probable es que los conuláridos fueron organismos filtradores y miembros del bentos sésil.
Babcock et al . (1987a, fig. 4 C; 1987b, fig. 1.3) registran dos ejemplares de Conularia quichua Ulrich adheridos por una especie de tallo al esqueleto de ejemplares de una especie de Hyolithes Eichwald (después de la muerte de los mismos), destacando que algunos conuláridos tuvieron un hábito bentónico sésil cuando fueron adultos. No obstante, Babcock et al . (1987b) manifiestan la posibilidad, debido a la amplia distribución de algunas especies, de que algunos conuláridos lograran la dispersión mediante larvas planctónicas.
Van Iten (1991c), revisa el repositorio y las observaciones de Finks (1955, 1960) concordando con el mismo en que las evidencias sugieren un modo de vida sésil para los conuláridos.
Por otra parte, la presencia de conuláridos con el ápice convexo, truncado, mediante una pared apical, diafragma o schott , llevó a la proposición (Moore y Harrington, 1956, p. F37; Kozlowski, 1968) de un modo vida libre, al menos en algún momento del ciclo de vida, para estos organismos. No obstante, Van Iten (1991b) atribuye la presencia de schott en algunos conuláridos, a daño por las corrientes y representaría una cicatriz producida cuando la parte adapical es seccionada. En el mismo sentido opinan Leme et al . (2003) ya que los estratos que contienen los restos estudiados por ellos fueron depositados bajo condiciones de alta energía y por lo tanto el schott correspondería a una cicatriz dejada al reparar el daño causado por las corrientes.
Van Iten et al . (1996, lám. 1, fig. 8) ilustran dos ejemplares de Climacoconus quadratus asociados a un esqueleto de Sphenothallus , donde los conuláridos están orientados formando el mismo ángulo en relación con el tubo de Sphenothallus . Van Iten et al . (1996, fig. de texto 6) también registra la existencia de tres especímenes incompletos de Conularia splendida Billings preservados en asociación con un nautiloideo; los conuláridos se hallan en el mismo plano, aproximadamente, que el cefalópodo, orientados casi perpendiculares al eje longitudinal del mismo y con sus ápices apuntando hacia el cefalópodo.
Hughes et al. (2000) describen la disposición alineada de ejemplares de Baccaconularia robinsoni Hughes et al ., con sus ápices apuntando en la misma dirección, sugiriendo que habría existido una estructura de fijación en vida de los individuos.
Simões et al . (2000), sobre la base de estudios tafonómicos, analizaron especímenes orientados verticalmente, con la abertura hacia arriba en relación al substrato, los cuales serían elementos autóctonos que indicarían un rápido enterramiento; esta evidencia les permitiría inferir un modo de vida epifaunal bentónico fijo para los conuláridos. Por otra parte Rodrígues et al . (2003) sobre la base de evidencias tafonómicas, sedimentológicas y estratigráficas arriban a la misma conclusión en relación con el modo de vida de este grupo aportando también información sobre el peridermo de los conuláridos el cual habría sido flexible y pasible de deformación.
Descripción del material
Especies del género Paraconularia Sinclair han sido descriptas previamente en la Cuenca Tepuel-Genoa (Mariñelarena, 1970; Cúneo y Sabattini, 1987), pero esta es la primera vez que se da a conocer la presencia de un ejemplar de Paraconularia sp. cuyo ápice se hallaría conectado a cirrales de crinoideos (MLP 31131). El espécimen se halla preservado dentro de una concreción, la que en su interior permite el análisis de los moldes externos y parte del peridermo; éste puede observarse desde la parte media hacia la región adapertural, sin preservación de la abertura. La parte apical del mismo se prolonga llegando al cirro de un crinoideo. El cirro se puede apreciar, como molde externo, desde el interior de la concreción hasta el exterior de la misma.
El fragmento de crinoideo se halla constituido por osículos cilíndricos, homeomórficos, de contorno circular, en donde la faceta articular presenta un crenularium bien desarrollado formado por costillas delgadas y radiales. La areola se halla reducida y rodea un lumen diminuto de contorno circular. La unión entre los osículos es de tipo simplexial, suavemente ondulosa, en vista lateral el latus es liso y suavemente convexo. El diámetro de los osículos es de 1,4 mm y la altura es de 2 mm, siendo la altura mayor al diámetro. La morfología y dimensiones que se observan en este fragmento son características de las estructuras cirrales de los crinoideos, si bien no exclusivas. Sin embargo, la combinación de la morfología antes expuesta y la ausencia de cicatrices cirrales sugieren que se trataría de un fragmento cirral. Desde un punto de vista morfofuncional, la faceta articular ocupada casi exclusivamente por el crenularium y lumen diminuto, junto al elevado índice de relación alto/diámetro de los artejos, permite interpretar que se trata de estructuras poco flexibles y más o menos rígidas.
La concreción estudiada corresponde a la localidad de Puesto La Carlota, 9 km al sureste de Tecka (provincia del Chubut) correspondiendo al nivel de concreciones del perfil de Suero (1953, fig. 2) y pertenece a la Zona de Callitomaria tepuelensis-Streblochondria sp. (Pérmico temprano) según Pagani y Sabattini (2002).
La concreción fue analizada por difracción de rayos X para la identificación de sus componentes, encontrándose cuarzo (mayoritariamente), materiales arcillosos y una especie de fluorapatita en cantidad relativamente baja.
Discusión y conclusiones
Como se dijo anteriormente, la faceta articular ocupada casi exclusivamente por el crenularium y lumen diminuto, junto al elevado índice de relación alto/diámetro de los artejos, permite interpretar a las estructuras cirrales como poco flexibles. Stukalina (1986) demuestra un aumento en la flexibilidad de los tallos como una consecuencia de la reducción de la altura de los columnales. La menor altura de los columnales permite una suma de pequeños movimientos diferenciales entre los componentes del tallo lo cual asegura un mayor combamiento de los tallos o cirros. Contrariamente, en un fragmento de igual longitud pero constituido por osículos de mayor altura existirán muy pocos desplazamientos diferenciales entre los componentes del tallo, y por lo tanto las estructuras serán poco flexibles. Esto es sustentado a su vez por la presencia de un conulárido fijo a esta estructura, sugiriendo una asociación en vida de los conuláridos y crinoideos, en donde estos últimos ofrecían un sustrato para la fijación de los primeros.
La preservación de un fragmento de cirro con sus componentes cirrales articulados entre sí, evidencia un enterramiento súbito a muy rápido debido al bajo potencial de preservación de estas estructuras en forma articulada. Esto es coincidente con la afirmación de Feldmann y Babcock (1986, p. 470) y Babcock y Feldmann (1986a, p. 367): el hallazgo de conuláridos puede ser usualmente atribuido a condiciones inusuales de preservación, particularmente una rápida sedimentación y también una temprana diagénesis. Por otra parte, Babcock et al . (1987b, p. 202) también suponen que la abundancia local de conuláridos en el Devónico de Bolivia se debió a una rápida deposición y varios ejemplares fueron hallados en rocas que indican esas condiciones de sepultamiento. A la misma conclusión arriban Simões et al . (2000, p. 761) fundamentada en estudios tafonómicos.

Figura 2. A-F, Paraconularia sp. y crinoideo, diferentes vistas de la concreción / different views of the concretion. A, MLP 31131a X 2,2. B, MLP 31131b X 1,7. C, 31131b X 5 (molde de látex / latex mould ). D, MLP 31131b X 2,6. E, MLP 31131a X 3 (molde de látex / latex mould ). F, 31131b X 1,7 (molde de látex / latex mould ).
Con respecto a las concreciones que encierran los ejemplares estudiados corresponde señalar que Sprechmann et al . (2001) mencionan para las concreciones del Paleozoico superior de la Formación San Gregorio (Uruguay) anomalías químicas generadas por la presencia de fósiles y la alta disponibilidad de elementos como fósforo, calcio, sílice, nitrógeno y hierro los que dieron lugar a una temprana diagénesis. Según estos autores esto también explica la alta productividad dentro de estos niveles. Esto mismo se observa para los niveles concrecionarios aquí estudiados; la presencia de abundantes restos orgánicos en las concreciones, minerales de fósforo y la excelente preservación del material manteniendo su tridimensionalidad, sugieren una temprana diagénesis que sucedió a un rápido sepultamiento, evitando así la deformación. Es probable que estos eventos de sepultamiento se relacionen con tempestades las cuales dieron lugar a una importante tasa de sedimentación en un corto tiempo. Una variación en las condiciones químicas normales en este ambiente marino de plataforma, vinculado al aporte desde medios continentales de determinados cationes, permitió un rápido nucleamiento de las concreciones. Esto último es sustentado por la presencia de restos vegetales en las mismas.
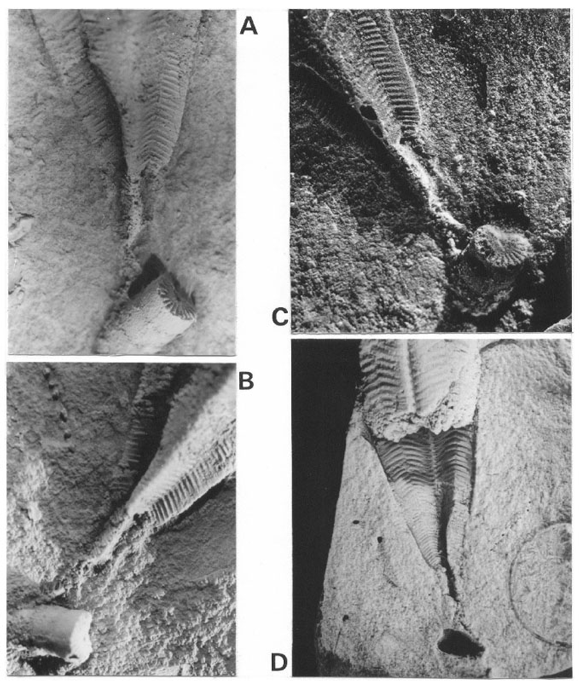
Figura 3. A-D, Paraconularia sp. y crinoideo, MLP 31131b. A-C, Molde de látex / latex mould X 8. D, Concreción / concretion X 4.
En este trabajo se da a conocer el primer hallazgo de conuláridos conectados a cirrales de crinoideos, lo cual sugiere un modo de vida bentónico sésil, al menos para algunos representantes de este grupo. Esta forma de vida sería también, según Van Iten (1991 a, pág. 153), una evidencia suplementaria a la teoría que apoya las relaciones de los Conulariida con los Cnidaria Scyphozoa.
Agradecimientos
A P. Zalba por el análisis por difracción de rayos X realizado a las concreciones. Al B. Pianzola por el trabajo fotográfico. A S. Gomba y C. Martín, encargadas de la Colección Paleozoología Invertebrados del Museo de Ciencias Naturales de La Plata, por la colaboración brindada durante el desarrollo de este trabajo.
Bibliografia
Babcock, L.E. y Feldmann, R.M. 1986a. Devonian and Mississippian conulariids of North America. Part A. General description and Conularia . Annals of Carnegie Museum of Natural History 55: 349-410. [ Links ]
Babcock, L.E. y Feldmann, R.M. 1986b. Devonian and Mississippian conulariids of North America. Part B. Paraconularia , Reticonularia , new genus, and organisms rejected from Conulariida. Annals of Carnegie Museum of Natural History 55: 411-479. [ Links ]
Babcock, L.E., Feldmann, R.M, Wilson, M.T. y Suárez-Riglos, M. 1987a. Devonian Conulariids of Bolivia. National Geographic Research 3: 210-231. [ Links ]
Babcock, L.E., Feldmann, R.M, Wilson, M.T. y Suárez-Riglos, M. 1987b. Conulariids (Phylum Conulariida) from the Devonian of Bolivia. 4° Congreso Latinoamericano de Paleontología I (Santa Cruz de la Sierra, Bolivia), Memorias: 201-215. [ Links ]
Bischoff, G.C.O. 1978. Internal structures of conulariid tests and their functional significance, with special reference to Circonulariina n. suborder (Cnidaria, Scyphozoa). Senckenbergiana lethaea 59: 275-326. [ Links ]
Cúneo, R. y Sabattini, N. 1987. Flora y fauna de la base de la Formación Río Genoa en la localidad de Ferraroti, Pérmico inferior de Chubut, Argentina. 4° Congreso Latinoamericano de Paleontología I (Santa Cruz de la Sierra, Bolivia), Memorias: 283-298. [ Links ]
Feldmann, R.M. y Babcock, L.E. 1986. Exceptionally preserved Conulariids from Ohio-Reinterpretation of their anatomy. National Geographic Research 2: 464-472. [ Links ]
Finks, R.M. 1955. Conularia in a sponge from the west Texas Permian. Journal of Paleontology 29: 831-836. [ Links ]
Finks, R.M., 1960. Late Paleozoic sponge faunas of the Texas region. The siliceous sponges. Bulletin of the American Museum of Natural History 120: 7-120. [ Links ]
Harland, T.L. y Pickerill, R.K. 1987. Epizoic Schizocrania sp. from the Ordovician Trenton Group of Québec, with comments on mode of life of conulariids. Journal of Paleontology 61: 844-849. [ Links ]
Hughes, N.C. Gunderson, G.O. y Weedon, M.J. 2000. Late Cambrian conulariids from Wisconsin and Minnessota. Journal of Paleontology 74: 828-838. [ Links ]
Jerre, F. 1994. Anatomy and phylogenetic significance of Eoconularia loculata , a conulariid from the Silurian of Gotland. Lethaia 27: 97-109. [ Links ]
Kozlowski, R. 1968. Nouvelles observations sur les Conulaires. Acta Palaeontologica Polonica 13: 497-535. [ Links ]
Leme, J.M., Heredia, S., Rodrigues, S.C., Simões, M.G., Aceñolaza, G.F. y Milana, J.P. 2003. Teresoconularia gen. nov. from the Lower Ordovician of the Cordillera Oriental of Salta (NW Argentina): the oldest conulariid (Cnidaria) from South America. Revista Española de Micropaleontología 35: 265-273. [ Links ]
Mariñelarena, M.P. 1970. Algunas especies de Paraconularia Sinclair del Sistema de Tepuel (Chubut) y sus relaciones con faunas del hemisferio austral. Ameghiniana 7: 139-150. [ Links ]
McKinney, F.K., Devolvé, J.-J. y Sobieraj, J. 1995. Conularia sp. from the Pyrénées: further support for scyphozoan affinities of the Conularida. Lethaia 28: 229-236. [ Links ]
Moore, R.C. y Harrington, H.J. 1956a. Scyphozoa. F27-F38. En: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part F. Coelenterata . Geological Survey of America and University of Kansas Press, 498 p. [ Links ]
Neal, M.L. y Hannibal, J.T. 2000. Paleoecologic and taxonomic implications of Sphenothallus and Sphenothallus-like specimens from Ohio and areas adjacent to Ohio. Journal of Paleontology 74: 369-380. [ Links ]
Pagani, M.A. y Sabattini, N. 2002. Biozonas de moluscos del Paleozoico superior de la Cuenca Tepuel-Genoa (Chubut, Argentina). Ameghiniana 39: 351-366. [ Links ]
Rodrigues, S. C., Simões, M. G. y Leme, J. M. de. 2003. Tafonomia comparada dos Conulatae (Cnidaria), Formação Ponta Grossa (Devoniano), Bacia do Paraná, Estado do Paraná, Brasil. Revista Brasileira de Geociências 33: 381-390. [ Links ]
Ruedemann, R. 1896. Note on the discovery of a sessile Conularia . Article I. American Geologist 158-165. [ Links ]
Simões, M.G., Mello, L.H.C., Rodrigues, S.C., Leme, J.M. de y Marques, A.C. Conulariid taphonomy as a tool in paleoenvironmental analysis. Revista Brasileira de Geociências 30: 757-762. [ Links ]
Sprechmann, P., Da Silva, J., Gaucher, C., Montaña, J. y Herrera, Z. 2001. Nuevos hallazgos fósiles en concreciones de la Formación San Gregorio del Uruguay e implicancias paleoecológicas y paleoclimáticas. 2º Simposio Argentino del Paleozoico Superior (Trelew). Resúmenes: p. 28. [ Links ]
Stukalina, G. A. 1986. Zakonomernosti istoricheskogo razvitya krinoidei v rannemi srednem paleozoe SSSR. ( Leyes del desarrollo histórico de los crinoideos en el Paleozoico inferior y medio). Akademiya Nauk SSSR, Paleontological Institut Moskova 1: 1-142 (en ruso). [ Links ]
Suero, T. 1953. Las sucesiones sedimentarias suprapaleozoicas de la zona extraandina del Chubut. Revista de la Asociación Geológica Argentina 8: 37-53. [ Links ]
Van Iten, H. 1991a. Evolutionary affinities of conulariids. En: A.M. Simonetta y Conway S. Morris (eds.), The Early Evolution of Metazoa and the Significance of Problematic Taxa , 145-155. Cambridge University Press. [ Links ]
Van Iten, H. 1991b. Anatomy, patterns of occurrence, and nature of the conulariid schott. Palaeontology 34: 939-954. [ Links ]
Van Iten, H. 1991c. Repositories of and additional comments on Permian sponge specimens containing molds of Paraconularia . Journal of Paleontology 65: 335-337. [ Links ]
Van Iten, H. 1992a. Morphology and phylogenetic significance of the corners and midlines of the conulariid test. Palaeontology 35: 335-358. [ Links ]
Van Iten, H. 1992b. Microstructure and growth of the conulariid test: implications for conulariid affinities. Palaeontology 35: 359-372. [ Links ]
Van Iten, H. 1994. Redescription of Glyptoconularia gracilis (Hall), an ordovician conulariid from North America. New York State Museum Bulletin 481: 363-366. [ Links ]
Van Iten, H. y Cox, R.S. 1992. Evidence of clonal budding in a radial cluster of Paraconularia crustula (White) (Pennsylvanian: ?Cnidaria). Lethaia 25: 421-426. [ Links ]
Van Iten, H., Cox, R.S. y Mapes, R.H. 1992. New data on the morphology of Sphenothallus Hall: implications for its affinities. Lethaia 25: 135-144. [ Links ]
Van Iten, H., Fitzke, J.A. y Cox, R.S. 1996. Problematical fossil cnidarians from the Upper Ordovician of the North-Central USA. Palaeontology 39: 1037-1064. [ Links ]
Recibido: 7 de junio de 2004.
Aceptado: 7 de diciembre de 2004.

