










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.42 n.4 Buenos Aires sept./dic. 2005
The Triassic Insect Fauna from Argentina. Blattoptera from the Los Rastros Formation (Bermejo Basin), La Rioja Province
Rafael Gioia Martins-Neto 1 , Adriana Mancuso 2 and Oscar Florencio Gallego 3
1 Programa de Pós-graduação, Universidade de Juiz de Fora-UFJF. Sociedade Brasileira de Paleoartropodologia-SBPr/: Campus Universitário-Martelos. 36036-900, Juiz de Fora, MG, Brasil. mtnsneto@icb.ufjf.br
2 IANIGLA (CRICYT-CONICET), Adrián Ruiz Leal s/n, Parque Gral. San Martín, 5500 Mendoza, CC 330. amancu@lab.cricyt.edu.ar
3 Paleontología (Facultad de Ciencias Exactas y Naturales y Agrimensura-Universidad Nacional del Nordeste) y Área Paleontología (Centro de Ecología Aplicada del Litoral-Consejo Nacional de Investigaciones Científicas y Técnicas), CC 128, 3400 Corrientes, Argentina. ofgallego@hotmail.com
Abstract. This contribution describes new fossil insect taxa from the Los Rastros Formation (early Late Triassic), La Rioja Province (Argentina). Several new taxa of the order Blattoptera, two families, seven genera and eleven species are proposed: Argentinoblatta herbsti gen. et sp. nov., Condorblatta lutzae gen. et sp. nov. (Argentinoblattidae fam. nov.), Mancusoblatta pulchella gen. et sp. nov., Hermosablatta crassatella gen. et sp. nov., Hermosablatta pectinata sp. nov. ( Mancusoblattidae fam. nov.), Lariojablatta chanarensis gen. et sp. nov. (family uncertain), Triassoblatta argentina sp. nov., Samaroblatta gualoensis sp. nov., S. corrientesina sp. nov. and Pulchellablatta nana gen. et sp. nov. (Mesoblattinidae sensu stricto ), and Argentinoblattula revelata gen. et sp. nov. (Blattulidae Vischnyakova). The presence of the genera Triassoblatta Tillyard and Samaroblatta Tillyard in the Argentinean Triassic considerably extends their known paleobiogeographic distribution.
Resumen. La fauna de insectos triásicos de la Argentina. Blattoptera de la Formación Los Rastros (cuenca del Bermejo) provincia de La Rioja . En este trabajo se describen nuevos insectos fósiles de la Formación Los Rastros (Triásico Superior bajo), provincia de La Rioja (Argentina). Se proponen varios nuevos taxones pertenecientes al orden Blattoptera, dos familias, siete géneros y once especies: Argentinoblatta herbsti gen. et sp. nov., Condorblatta lutzae gen. et sp. nov. (Argentinoblattidae fam. nov.), Mancusoblatta pulchella gen. et sp. nov., Hermosablatta crassatella gen. et sp. nov., Hermosablatta pectinata sp. nov. (Mancusoblattidae fam. nov.), Lariojablatta chanarensis gen. et sp. nov. (familia incierta), Triassoblatta argentina sp. nov., Samaroblatta gualoensis sp. nov., S. corrientesina sp. nov., Pulchellablatta nana gen. et sp. nov (Mesoblattinidae sensu stricto ) y Argentinoblattula revelata gen. et sp. nov. (Blattulidae Vischnyakova). La presencia de los géneros Triassoblatta Tillyard y Samaroblatta Tillyard en el Triásico de la Argentina extiende considerablemente su distribución paleogeográfica.
Key words. Triassic insects; Blattoptera; Los Rastros Formation; Argentina.
Palabras clave. Insectos triásicos; Blattoptera; Formación Los Rastros; Argentina.
Introduction
In this paper we present the first descriptions of Triassic blattopterans from Argentina and the second from southern South America. The only previously described species is Triassoblatta cargnini Pinto and Ornellas, 1974 from the Middle to Upper Triassic (Santa Maria Formation) of southern Brazil. A historical summary and lists of previously described insects can be found in Gallego (1997), Gallego and Martins-Neto (1999), Martins-Neto and Gallego (1999, 2001) and Martins-Neto et al. (2003).
In this contribution we describe new Blattoptera from the Los Rastros Formation (Río Gualo, Picos Gemelos and Los Chañares localities, La Rioja Province, Argentina) (figure 1.A). The samples from the Los Rastros Formation were collected in two field trips, one by OFG, A. Arcucci, C. Forster, C. May, and R. Rogers in 1995 and the other by ACM, A. Arcucci and C. Marsicano in 1999.

Figure 1. A, Geologic map of the Bermejo Basin (modified from Stipanicic and Bonaparte, 1979 and Kokogian et al ., 2001) / Mapa geológico de la Cuenca del Bermejo (modificado de Stipanicic y Bonaparte, 1979 y Kokogian et al ., 2001). B, Schematic section of the Los Rastros Formation indicating fossiliferous horizons / Sección esquemática de la Formación Los Rastros indicando los horizontes fosilíferos.
The terminology adopted here follows Kukalova-Peck (1991), except for the newly proposed abbreviations as follows (figure 2.A):
oMA: the distance from the tegmen base to MA origin;
aml: anal margin length from the tegmen base to CuP distal part;
amh: anal margin hight from CuP origin, at the tegmen base to the anal margin;
d: anal area diagonal, from CuP origin at CuP distal extremity, at the anal margin;
f: height from the main CuP curvature to d, in relation to the tegmen base;
lcm: length from the wing base to the last distal ScP branch;
lrp: length from the distal part of the first RA branch to the last RP branch;
lh: Humeral area length.

Figure 2. Schematic drawing of the tegmen of representatives of different Blattoptera families, showing the main diagnostic characters. The intersection points (black dots) are from the top to bottom, last RA branch extremity, MA origin and CuP distal extremity / Dibujo esquemático de las tegminas de representantes de diferentes familias del Orden Blattoptera, mostrando los principales caracteres diagnósticos. Los punto de intersección (esferas negras) corresponden de arriba hacia abajo, al extremo de la última rama de la RA , y al origen de la MA y al extremo distal de la CuP , respectivamente. A-C, Argentinoblattidae fam. nov. A, Argentinoblatta herbsti sp. nov. drawn from the holotype / dibujado del holotipo, PULR-I 220 . B, Rhipidoblattina geikie Tillyard (Lower Jurassic), modified from Tillyard (1919) / (Jurásico Inferior), modificado de Tillyard (1919). C, Scleroblatta densa Papier et al . (Triassic, France), modified from Papier et al . (1996) / (Triásico, Francia), modificado de Papier et al. (1996). D-G, Mesoblattinidae sensu stricto . D, Samaroblatta reticulata Tillyard (Triassic, Australia), modified from Tillyard (1919) / (Triásico, Australia), modificado de Tillyard (1919). E, Samaroblatta fronda Fujiyama (Triassic, Japan), modified from Fujiyama (1973) / (Triásico, Japón), modificado de Fujiyama (1973). F, Triassoblatta natalensis Riek (Triassic, South Africa), modified from Riek (1974) / (Triásico, Sudáfrica), modificado de Riek (1974). G, Triassoblatta tasmanica Riek (Triassic, Tasmania), modified from Riek (1962) / (Triásico, Tasmania), modificado de Riek (1962). H-K, Mancusoblattinidae fam. nov. H, Pedinoblatta ishidae Fujiyama (Triassic, Japan), modified from Fujiyama (1973) / (Triásico, Japan), modificado de Fujiyama (1973). I, Triassoblatta cargnini Pinto and Ornellas (Triassic, Brazil), modified from Pinto and Ornellas (1974) / (Triásico, Brasil), modificado de Pinto and Ornellas (1974). J, Transitoblatta reticulata Papier et al. (Triassic, France), modified from Papier et al. (1996) / (Triásico, Francia), modificado de Papier et al. (1996). K, Triassoblatta grandis Dodds (Triassic, Australia), modified from Dodds (1949) / (Triásico, Australia), modificado de Dodds (1949). L-N, Blattulidae. L, Blattulopsis popovi Pinto (Lower Cretaceous, Argentina), modified from Pinto (1990) / (Cretácico Inferior, Argentina), modificado de Pinto (1990). M, Blattula intercalata Handlirsch (Lower Jurassic, Germany), modified from Handlirsch (1939) / (Jurásico Inferior, Alemania), modificado de Handlirsch (1939). N, New genus and species, from the Potrerillos Formation (Triassic, Argentina) / Nuevo género y especie, de la Formación Potrerillos (Triásico, Argentina). Terminology: AP, posterior anal; CuA , CuP , anterior and posterior cubitus; MA , MP , anterior and posterior media, respectively; RA , RP , anterior and posterior radius. Scale bar = 2 mm / Terminología: AP , anal posterior; CuA , CuP , cubital anterior y cubital posterior; MA , MP , media anterior y media posterior; RA , RP , radial anterior y radial posterior. Escala = 2 mm.
The repository and institutional abbreviations used here are, PULR-I: Paleontology Collection, Museo de Ciencias Naturales, Universidad Nacional de La Rioja, La Rioja, Argentina. PZ-CTES: Paleozoological Collection, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, Corrientes, Argentina.
Geological setting
Extensional basins were formed along the western margin of Gondwana during the Late Permian and Early Triassic, during the pre-breakup stage of Pangea (Uliana and Biddle, 1988). The Bermejo Basin is located in northwestern Argentina, with a NW-SE orientation along the border between San Juan and La Rioja Provinces (figure 1.A). Alluvial, fluvial and lacustrine deposits (up to 4000 m) of Triassic age constitute the sedimentary infilling of the Bermejo Basin. From a lithostratigraphic point of view, the sedimentary succession has been divided (in ascending order) into the Talampaya and Tarjados Formations (Romer and Jensen, 1966) and the Agua de la Peña Group, which has been divided into five formations (de la Mota, 1946; Bossi, 1971; Stipanicic, 1983). The lower Chañares Formation consists of tuffaceous sandstones and siltstones deposited in a fluvial-lacustrine environment which passes upward into the lacustrine black shales and deltaic sandstones of the Ischichuca and Los Rastros Formations (Rogers et al ., 2001). The sequence continues with sandstones, mudstones and tuffs of the Ischigualasto Formation, deposited by a moderate to high-sinuosity river system. Finally, the red beds of the Los Colorados Formation cap the succession. Following this, a regional unconformity separates the Triassic sucession from the overlying ?Cretaceous Cerro Rajado Formation (Caselli et al ., 2001).
The Los Rastros Formation conformably overlies the Chañares/Ischichuca Formations, whereas it is unconformably overlain by the Ischigualasto Formation (Milana and Alcober, 1994). Generally, the Los Rastros Formation deposits are interpreted as lacustrine-deltaic cycles with a predominance of lacustrine sedimentation (Stipanicic and Bonaparte, 1979; López-Gamundí et al ., 1989; Milana and Alcober, 1994; Milana, 1998; Rogers et al ., 2001).
Fieldwork undertaken at Río Gualo (figure 1.A) allowed Mancuso (1999, 2003) and Rogers et al . (2001) to recognise lacustrine-deltaic cycles with two facies assemblages each. The facies assemblages are characterised by coarsening-upward sequences which consist of (from bottom to top): (1) lake facies assemblage dominated by claystones and iron mudstones interbedded, (2) deltaic facies assemblage characterised by coarsening-upward siltstones and sandstones (figure 1.B). Individual coarsening-upward sequences range from 20 to 40 m.
The lake facies assemblage consists mainly of dark grey to black carbonaceous claystones. Iron mudstones and very fine-grained sandstones are interbedded. The beds are tabular and the claystones are horizontally laminated, whereas the ironstones may be either massive or laminated and rarely show gradation. Conchostracans, plant debris, insect impressions, rare fish body fossils and rare trails are found in this assemblage, but with different preservational conditions in the different facies (Mancuso, 1999, 2000a, 2000b, 2000c, 2003; Mancuso and Gallego, 2000). The claystones were deposited by suspension in distal-lake conditions whereas the ironstones were deposited by distal turbiditic currents (Farquharson, 1982; Talbot and Allen, 1996; Mancuso, 1999, 2000a, 2003).
The deltaic facies assemblage begins with an alternation of green silstones and grey claystones which form rhythmic packages of massive individual beds. Fine to coarse-grained sandstone beds are intercalated, these being more abundant and thicker upwards. They show current-rippled to horizontal lamination and culminate with planar or trough cross-bedding in the upper coarser material. This facies assemblage contains abundant plant remains and well-preserved fish body fossils (Mancuso, 1999, 2000a, 2000b, 2000c, 2003). At the top of the assemblage, dark grey carbonaceous mudstones and very fine-grained sandstones are interbedded with planar and trough cross-bedded sandstones. This facies assemblage represents the progradation of a mouth bar of the fluvial system in the lake body (Farquharson, 1982; Talbot and Allen, 1996; Mancuso, 1999, 2000a, 2003).
At present, the floral record of the Los Rastros Formation is the best represented. The abundant macrofloral assemblage is related to the Dicroidium type flora and has been reported from the lacustrine black shales, which lie at the base of each cycle in the sequence (Frenguelli, 1948; Stipanicic and Bonetti, 1969; Stipanicic and Bonaparte, 1979; Spalletti et al ., 1999; Kokogian et al ., 1999). The few palynological analysis suggest that the rich palynological association is related to the Ipswich-type microflora (Yrigoyen and Stover, 1970; Zavattieri and Melchor, 1999; Ottone et al ., 2005). Conversely, knowledge of the faunal record of the unit has grown during the last years. The invertebrate remains include different groups of conchostracans (clamp shrimps), insects, bivalves and crustacean branchiopods (notostracans), which were collected in different levels throughout the whole sequence in Río Gualo, Los Chañares and Agua de la Peña localities. The vertebrate fauna is mainly represented by actinopterigian fishes, whereas tetrapods are represented only by an external mould of a single temnospondyl amphibian and archosaur footprints.
Systematic paleontology
Order Blattoptera Brunner, 1882
Superfamily Blattoidea sensu Handlirsch, 1906
Remarks on the families Mesoblattinidae Handlirsch and Poroblattinidae Handlirsch
The Family Mesoblattinidae was erected by Handlirsch (1906) for the genus Mesoblattina Giebel from the European Jurassic and the Family Poroblattinidae was erected in the same study by Handlirsch (1906) for the genus Poroblattina Scudder, 1895 from the Palaeozoic of USA. Tillyard (1919) proposed that the Triassic genera Triassoblatta Tillyard, Samaroblatta Tillyard and Austroblattulla Tillyard from Australia belong to the Family Mesoblattinidae. Subsequently, several authors described species of both genera, Triassoblatta and Samaroblatta , from Japan (Fujiyama, 1973), China (Hong, 1980; Lin, 1978, 1989), Tasmania (Riek, 1962), South Africa (Riek, 1974), Brazil (Pinto and Ornellas, 1974) and Australia (Dodds, 1949). There are also specimens attributable to these genera from Germany (Brauckmann and Schlüter, 1993) and from the Russian Jurassic (Martynov, 1937). These authors' interpretations of these two genera at a higher level (subfamily and family), differ greatly. Dodds (1949) referred Triassoblatta and Samaroblatta to the Family Poroblattinidae Handlirsch, removing them from Mesoblattinidae. Rohdendorf (1962) referred them to the Subfamily Mesoblattininae of the Family Blattidae; Riek (1976) interpreted Triassoblatta as Poroblattinidae and Samaroblatta as Mesoblattinidae, and Fujiyama (1973) followed Tillyard's interpretation (Mesoblattinidae). Two recent reviews were made on these related groups: Schneider (1983) focused on Poroblattinidae (not considering Triassoblatta and Samaroblatta as belonging to this family) and Vrsansky (1998) focused on Lower Cretaceous Blattoidea (Family Mesoblattinidae) but mentioning neither Triassic genera nor the taxonomic position of Triassoblatta or Samaroblatta . Vrsansky (1998) also mentioned that several Lower Cretaceous genera attributable to Mesoblattinidae could be transferred to the Family Blattellidae. Apart from the two classical genera Triassoblatta and Samaroblatta , other Triassic genera related to these are: Pedinoblatta Handlirsch, 1912 (with species described from Germany and Japan, interpreted as Mesoblattinidae), Samaroblattella Riek, 1976 from South Africa, interpreted as Mesoblattinidae (later removed to Subioblattidae Schneider by Papier et al ., 1994), Voltziablatta Papier and Grauvogel-Stamm, 1995, Scleroblatta et al ., 1996 and Transitoblatta Papier, et al. , 1996 from France interpreted as uncertain family (although close to Mesoblattinidae).
The Argentinean Triassic furnishes a notably great number of Blattoptera species, with several new taxa, including the presence of the typical genera Triassoblatta and Samaroblatta . We propose simple and consistent morphological parameters (abbreviations presented in the Introduction chapter) which reasonably define the families involved, exemplified in figure 2 (all wings are drawn as right wings, missing if they are left wings, unless their natural position, to facilitate comparisons):
lcm = oMA = aml : Argentinoblattidae fam. nov., described below (Middle Triassic to ?Lower Jurassic) figures 2.A-C; oMA = aml < lcm : Mancusoblattidae fam. nov., described below (Middle Triassic to ?Lower Jurassic) figures 2.H-K; lcm = aml > oMA : Mesoblattinidae Handlirsch, 1906 sensu stricto (for Triassic only) figures 2.D-G; lcm = oMA > aml : Blattulidae Vishnyakova, 1982 (Mesozoic) figures 2.L-N; lcm =aml< oMA : a new Blattoidea family (Triassic) to be described in a forthcoming paper.
Inevitably the identity (=) is approximate because the angle lcm / oMA / aml varies from specimen to specimen as well as the position lcm / oMA / aml varies in relation to the tegmen base (generic or sometimes specifically useful characters). According to this system, both Triassoblatta and Samaroblatta do not belong to the Family Poroblattinidae and are included here in the Family Mesoblattinidae ( sensu stricto ). Furthermore, some species attributed to these genera are reassigned to a new one. We agree with Papier et al . (1996) who consider that Mesoblattinidae Handlirsch needs to be properly reviewed, though that is not the aim of this contribution. Two distinct families, Subioblattidae Schneider and a new one to be proposed in a forthcoming paper have enough synapomorphies to exclude them from the Mesoblattinidae complex although they do not follow the proposed new parameters.
Argentinoblattidae Martins-Neto and Gallego fam. nov.
Genera included. Argentinoblatta gen. nov. And Condorblatta gen. nov. from the Argentinean Triassic. Other possible genera (based on the original published material not examined): Scleroblatta Papier et al ., 1996 from the French Triassic (figure 2.C), Rhipidoblattina Handlirsch, 1906 from the English and Russian Lower Jurassic (figure 2.B), Sogdoblatta Martynov, 1937, Taublatta Martynov, 1937, Samaroblattula Martynov, 1937 and Euryblattula Martynov, 1937, all from the Lower Jurassic of central Asia (Kizil-Kiya and Shurab).
Diagnosis. Tegmen elongated, three times longer than wide, with the distal extremity of the last branch of RA , MA origin and CuP distal extremity perpendicularly aligned to both costal and anal margins ( lcm = oMA = aml ). ScP area notably short (circa 1/6 of the wing length). Anal area restricted between ¼ and 1/3 of the tegmen length; f = 1/2.
Argentinoblatta Martins-Neto and Gallego gen. nov.
1997. Triassoblatta Gallego, p. 513, lám. I, fig. A
1999. Mendozablatta Gallego and Martins-Neto ( nomen nudun ), p. 87, fig. 9.
Type species. Argentinoblatta herbsti Martins-Neto and Gallego sp. nov .
Etymology. Refers to the República Argentina, from which the material originates, and blatta , general suffix for blattoids.
Diagnosis. Tegmen with the costal area extremely reduced, restricted to the base of the wing; notably short ScP (circa 1/6 of the wing length), unbranched. The parameter aml = oMA = d = lcm =4/10 of the total tegmen length.
Discussion. Argentinoblatta gen. nov. is similar to Scleroblatta Papier et al ., 1996 from the French Triassic and Rhipidoblattina Handlirsch, 1906 from the Asiatic Jurassic by having the tegmen with the distal part of the last branch of RA , MA origin and CuP distal part perpendicularly aligned to both costal and anal margins (the same as aml = oMA = lcm , family characters). Argentinoblatta gen. nov. differs from Scleroblatta by having unbranched ScP (multibranched in Scleroblatta ) and multibranched RA (unbranched in Scleroblatta ); M and CuA sligthly sigmoidal in Scleroblatta , straight in Argentinoblatta gen. nov. The new genus is similar to Rhipidoblattina and related genera by having unbranched ScP , however, it differs by the costal area being notably shorter and multibranched RA (unbranched in all known genera). Cross venation absent.
Argentinoblatta herbsti Martins-Neto and Gallego sp. nov.
Figures 2.A; 5.A
1997. Triassoblatta sp. Gallego, p. 513, lám. I, fig. A
1999. Mendozablatta herbsti Gallego and Martins-Neto ( nomen nudun ), pp. 87-88, fig. 9 .
Holotype. PULR-I 220.
Etymology. In honour of Dr. Rafael Herbst (Instituto Miguel Lillo-CONICET, Tucumán, Argentina) for his great contribution to the paleontology of southern South America.
Type locality. Río Gualo, La Rioja Province, Argentina.
Type stratum. 5th cycle from Los Rastros Formation.
Age. Early Late Triassic.
Diagnosis. Tegmen three times longer than wide, around 15 mm long. MA distally forked; threebranched MP ; CuA about five times branched.
Description of the holotype. (Figures 2.A; 5.A). Tegmen 15.2 mm long and 4.7 mm wide. Costal area very small, restricted to the wing base. Short ScP , unbranched; multibranched RA . Radial area wider than the cubital area. Long RP , slightly sigmoid at its base, reaching the apical area a little below the apex; about ten long pectinated secondary branches; MA long and parallel to RP , unbranched; MA origin between 1/4 and 1/3 of the tegmen length. MP branches about three times. Branched CuA , all of them oblique to the anal margin and sometimes forked; CuA1 with distal dichotomy; trichotomous CuA5 . CuP slightly curved towards the anal margins. Anal area relatively small restricted between 1/4 and 1/3 of the wing length occupying around 40% of the tegmen width. Last RA distal secondary branch, MA origin and CuP distal extremity perpendicularly aligned to both costal and anal margins. At least six anal veins, just one with distal dichotomy. No cross-veins detected. Another tegmen fragment possibly attributable to the genus Argentinoblatta is: PZ-CTES 7292a.
Condorblatta Martins-Neto and Gallego gen. nov.
Type species. Condorblatta lutzae Martins-Neto and Gallego sp. nov.
Etymology. Refers to the condor, a large bird, inhabitant of the Andean region, and blatta, common suffix for blattoids.
Diagnosis. Elongated tegmen, with multibranched ScP , with the last distal branches reaching the costal margin at the same level of CuP distal extremity; unbranched RA ; short RP , sigmoid, reaching the costal margin at around 2/3 of the tegmen length; CuA branches bend strongly backwards, running subparallel to each other and the posterior wing border.
Discussion. Similar to Argentinoblatta gen. nov. By having an elongated tegmen (see the reconstruction in figure 3.E), MA origin at 1/3 of the tegmen base, having the last ScP branches restricted to 1/3 of the tegmen length and by the absent of cross venation, however, differing by having CuA secondary branches are bend backward, subparallel to the posterior wing border.
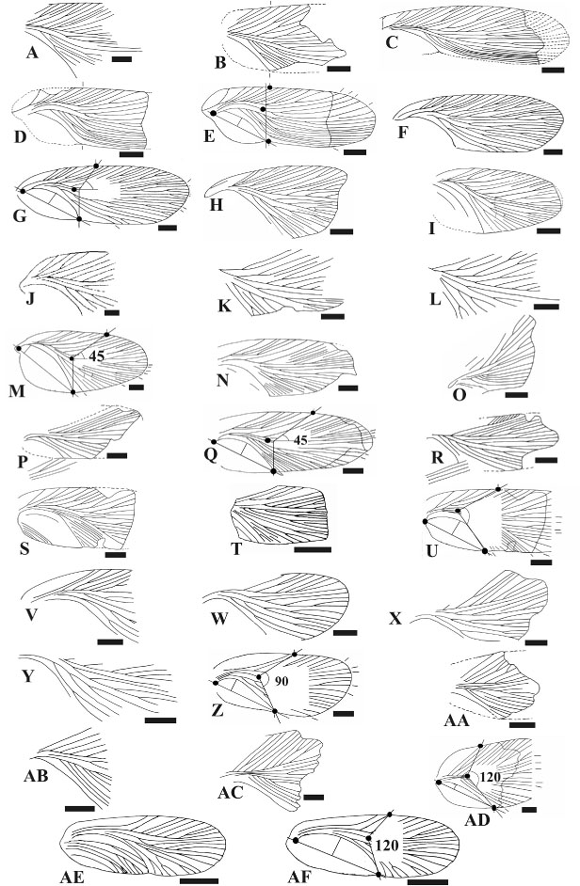
Figure 3. A-E, Condorblatta lutzae sp. nov. A-B, drawn from the paratypes, PULR-I 283, part (B) and PZ-CTES 7298, counterpart (A) / dibujado de los paratipos, PULR-I 283, parte (B) y PZ-CTES 7298, contraparte (A). C, drawn from the paratype, PULR-I 264 / dibujado del paratipo, PULR-I 264, D, drawn from the holotype / dibujado del holotipo, PULR-I 285. E, Wing reconstruction / reconstrucción del ala. F-G, Mancusoblatta pulchella sp. nov. F, drawn from the holotype / dibujado del holotipo, PULR-I 288. G, wing reconstruction / reconstrucción del ala. H-M, Hermosablatta crassatella sp. nov., drawn from holotype, PULR-I 289 (H), paratypes PULR-I 277, part (I) and PZ-CTES 7274, counterpart (J), paratypes PULR-I 273 (K), and PZ CTES 7277 (L), and wing reconstruction (M), respectively / dibujado del holotipo, PULR-I 289 (H), paratipos, PULR-I 277, parte (I) y PZ-CTES 7274, contraparte (J), paratipos, PULR-I 273 (K), y PZ CTES 7277 (L), y reconstrucción del ala (M) respectivamente. N-Q, Hermosablatta pectinata sp. nov., drawn from the holotype, PULR-I 284 (N), paratypes PULRI 276 (O) and PULR-I 291 (P), and wing reconstruction (Q), respectively / dibujado del holotipo, PULR-I 284 (N), paratipos, PULR-I 276 (O) y PULR-I 291 (P), y reconstrucción del ala (Q), respectivamente. R, Hermosablatta ? sp., drawn from the material / dibujado del material, PULR-I 275. S-U, Triassoblatta argentina sp. nov., drawn from the holotype, PULR-I 222 (S), paratype, PULR-I 290 (T) and wing reconstruction (U), respectively / dibujado del holotipo, PULR-I 222 (S), paratipo, PULR-I 290 (T) y reconstrucción del ala (U), respectivamente. V-Z, Samaroblatta gualoensis sp. nov., drawn from holotype, PULR-I 279 (V), paratypes PULR-I 281 (W), PULR-I 280 (X, part), and PZ CTES 7275 (Y, counterpart), and wing reconstruction (Z), respectively / dibujado del holotipo PULR-I 279 (V), paratipos PULR-I 281 (W), PULR-I 280 (X, parte), PZ CTES 7275 (Y, contraparte), y reconstrucción del ala (Z), respectivamente. AA-AD, Samaroblatta corrientesina sp. nov., drawn from paratypes, PULR-I 287 (AA) and PZ-CTES 7296 (AB), holotype, PULR-I 282 (AC), and wing reconstruction (AD), respectively / dibujado de los paratipos, PULR-I 287 (AA) y PZ-CTES 7296 (AB), holotipo, PULR-I 282 (AC), y reconstrucción del ala (AD), respectivamente. AE-AF, Pulchellablatta nana sp. nov., drawn from the holotype / dibujado del holotipo , PULR-I 274 (AE) and wing reconstruction (AF), respectively / y reconstrución del ala (AF), respectivamente. Abbreviations discussed in the text. Scale bar = 2 mm / Abreviaturas discutidas en el texto. Escala = 2 mm.
Condorblatta lutzae Martins-Neto and Gallego sp. nov.
Figures 3.A-E; 5.B
Holotype. PULR-I 285.
Paratypes. PULR-I 264 (part, figure 3.C) and PZ-CTES 7295 (counterpart), PULR-I 283 (part, figure 3.B) and PZ-CTES 7298 (counterpart, figure 3.A), 2 specimens.
Etymology. In honour of Lic. Alicia I. Lutz (FACENA-UNNE and CECOAL-CONICET, Corrientes, Argentina) for her contribution to Argentinian paleobotany.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Other locality. Picos Gemelos (Gualo), La Rioja Province, Argentina.
Diagnosis. As for the genus.
Description of the holotype (figure 3.D). Tegmen 12.6 mm long, as preserved, and 5.6 mm wide. Costal margin parallel to anal margin. Costal area relatively small occupying around 1/3 the length of the tegmen (around 18 mm). ScP , circa 1/3 of the wing length, three-branched. Unbranched RA without distal dichotomy. Radial area relatively narrow. Short RP , sigmoid, reaching the anterior wing margin at 2/3 of the tegmen base; six relatively short pectinated secondary branches; dichotomous RP4 to 6 . MA long and parallel to RP , unbranched as preserved; MA origin at 1/3 to nearly 1/2 of the tegmen length. Two-branched MP as preserved, with MP1 branching again. Six-branched CuA and several secondary branches pectinated and parallel to the anal margin. Base of RP , M and CuA sigmoid and parallel. CuP slightly curved towards the anal margin. Anal area occupying 1/3 of the wing length and 70% of the wing width. RA distal extremity, MA origin and CuP distal extremity perpendicularly aligned ( MA slightly before, however, and very close to the alignment) to both costal and anal margins. Cross-veins and intercalary veins absent. The paratypes PULR-I 283 (part, figure 3.B), and PZ-CTES 7298 (counterpart, figure 3.A) from Picos Gemelos (Gualo, La Rioja Province) consists of a tegmen fragment 10 mm long and 5.4 mm wide, as preserved, exhibiting the same CuA diagnostic characters. Other paratypes, PULR-I 264 (figure 3.C) and PZ-CTES 7295, from the same locality, consist of a similar tegmen fragment 15.0 mm long and 3.7 mm wide.
Family Mancusoblattidae Martins-Neto and Gallego fam. nov.
Genera included. Mancusoblatta gen. nov. And Hermosablatta gen. nov. from the Argentinian Triassic. Other possible constituent genera (based on the original published material not examined, figures 2.J, 2.H): Transitoblatta Papier et al ., 1996 from the French Triassic, Pedinoblatta Handlirsch, 1912 from the Japanese Triassic (Fujiyama, 1973), and Opismoblatta Brauer et al ., 1889 from Lower Jurassic of central Asia (Irkutsk).
Diagnosis. Tegmen with MA origin and CuP distal extremity perpendicularly aligned to both costal and anal margins and the distal part of the last RA branch oblique to the MA / CuP alignment, at an angle around 45º ( oMA = aml < lcm ). CuA secondary branches well marked. Number of MP secondary branches reduced.
Discussion. Similar to Palaeozoic Poroblattinidae Handlirsch, 1906 by having tegmen with MA origin and CuP distal extremity perpendicularly aligned to both costal and anal margins and the distal part of the last RA branch oblique to the MA / CuP alignment, at an angle around 45º ( oMA = aml < lcm ). Poroblattinids are a specialised group with small sclerotized wings. Because of this increasing sclerotization, the basal part of the anal veins disappear, a part CuA veins is replaced by very fine veins, more like the fine veins of the cross venation. The cross venation is not very well expressed, because of sclerotization. All together, the fore wings are more like to beetle elytra.
Mancusoblatta Martins-Neto and Gallego gen. nov.
Type species. Mancusoblatta pulchella Martins-Neto and Gallego sp. nov.
Etymology. In honour to Dra. Adriana Mancuso (Universidad de Buenos Aires, Argentina) who collected this specimen.
Diagnosis. Elongated tegmen with narrow costal area and multibranched ScP , whose last branch reaching 1/3 of the wing length at the same level of CuP distal extremity; no pronounced RA ; branched MA ; twobranched MP multiple forked; CuA branches arise by forking. CuP origin at R , relatively far from the tegmen base. Anal area occupying around 70% of the wing width. No cross venation present.
Discussion. Mancusoblatta gen. nov. differs from Pedinoblatta Handlirsch, 1912, Transitoblatta Papier et al ., 1996 and related genera by having the costal area notably narrower, the CuP origin relatively far from the tegmen base (close to the tegmen base in the mentioned genera) at RP and by the CuA construction: branches originate by multiple forking in Mancusoblatta gen. nov., by branching in related genera.
Mancusoblatta pulchella Martins-Neto and Gallego sp. nov.
Figures 3.F, G; 5.C
Holotype. PULR-I 288.
Etymology. Diminutive of pulcher (Latin), beautiful.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Diagnosis. As for the genus.
Description of the holotype (figures 3.F; 5.C). Male tegmen 18 mm long and 6 mm wide. Costal margin slightly curved and anal margin straight. Costal area relatively large, occupying around 40% of the wing length. ScP relatively long, branched; no pronounced RA . Radial area as wide as the cubital area. R long and sigmoid, reaching the apical area below the apex; about seven long and pectinated secondary branches, sometimes forked. MA long and parallel to RP , unbranched and distally forked; MA origin at 1/3 of the tegmen length. MP multiple forked. CuA forked into two, which are again multiple forked (about eleven twigs). CuP slightly curved towards the posterior margin; CuP origin at R , relatively far from the wing base. Cubital area about 1/3 of the wing length, occupying around 70% of the wing width. Last, MA origin, and CuP distal extremity perpendicularly aligned to both costal and anal margins; RA distal secondary branch and MA origin aligned at an angle around 45º related to MA origin and CuP distal extremity.
Remarks. Mancusoblatta pulchella sp. nov. is similar to Triassoblatta cargnini Pinto and Ornellas, 1974 described from the Brazilian Triassic (figure 2.I) by having a narrow costal area and similar CuP morphology. It differs in having the CuP origin at RP . Additionally T . cargnini has a massive presence of intercalary veins (not present in M. pulchella sp. nov.). The Brazilian species must be removed to the family Mancusoblattidae fam. nov. and a new genus, close to Mancusoblatta gen. nov., is necessary for T . carg nini. Mancusoblatta pulchella sp. nov. differs from Triassoblatta grandis Dodds, 1949 from the Australian Triassic lacking intercalary veins (massive in T . grandis , as for T . cargnini ). Differs from T . triquestra Dodds, 1949, T . deiscens Dodds, 1949 and T . intramedia Dodds, 1949 (all of them from the Australian Triassic), by having no distinct RA .
Hermosablatta Martins-Neto and Gallego gen. nov.
Type species. Hermosablatta pectinata sp. nov.
Etymology. Derivative of Hermes, God of the Greek mythology, the messenger, and blatta , common suffix for blattoids.
Diagnosis. RA reaching the costal margin circa 1/3 of apex. MA origin at 1/3 of the tegmen base. Anal area occupying around 40% of the tegmen length and 50% of its width.
Discussion. Similar to Mancusoblatta gen. nov. By having an elongated tegmen and MA origin at 1/3 of the tegmen base, however, differing by having the last ScP branch over 2/3 of the tegmen length (restricted to 1/3 in Mancusoblatta gen. nov.).
Hermosablatta pectinata Martins-Neto and Gallego sp. nov.
Figures 3.N-Q; 5.D
Holotype. PULR-I 284.
Paratypes. PULR-I 291(figure 3.P), PULR-I 276 (figure 3.O), 2 specimens.
Etymology. Refers to the secondary CuA pectinated branches.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Diagnosis. Male tegmen 14.6 mm long, as preserved, and 5.8 mm wide. Long ScP , threebranched; ScP2 and 3 with long distal dichotomy. Unbranched RA reaching the costal margin circa 1/3 of apex. MA origin at 1/3 of the tegmen base. Anal area occupying around 40% of the tegmen length and 50% of its width.
Description of the holotype (figures 3.N, 5.D). Male tegmen 14.6 mm long, as preserved, and 5.8 mm wide. Costal margin slightly curved. Costal area relatively large, occupying around 1/2 of the wing length. Long ScP , three-branched; ScP2 and 3 with long distal dichotomy; unbranched RA without distal dichotomy. Radial area as wide as the cubital area. RP long, slightly sigmoid at its base, following straight, reaching the apical area little below the apex, seven long, pectinated secondary branches; just perceptibly trichotomous RP5 and 7 . MA long and parallel to RP , unbranched and with distal dichotomy; MA origin at 1/3 of the tegmen length. Twobranched MP , both with distal dichotomy. Fourbranched CuA and with several secondary branches. CuP slightly curved towards the anal margin. Anal area occupying 40% of the wing length and 50% of the wing width. MA origin, and CuP distal extremity perpendicularly aligned to both costal and anal margins; RA distal secondary branch and MA origin aligned with an angle around 45º in relation to MA origin and CuP distal extremity. Cross venation absent and few intercalary veins detected.
Hermosablatta crassatella Martins-Neto and Gallego sp. nov.
Figures 3.H-M; 5.E
Holotype. PULR-I 289.
Paratypes. PULR-I 277 (part, figure 3.I) and PZ-CTES 7274 (counterpart, figure 3.J), PULR-I 273 (figure 3.K), and PZ-CTES 7277 (figure 3.L), 3 specimens.
Etymology. Diminutive of crassus (Latin), fat, in reference to the robust fore wing shape.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Other locality. Picos Gemelos (Gualo), La Rioja, Argentina.
Diagnosis. Female tegmen around two-times longer than wide, with ScP distally multibranched, with last branch reaching the costal margin around 1/3 of the apex, very far from CuP distal extremity. RA last branch reaching the costal margin after the mid-length of the tegmen. MA origin at the mid length of the tegmen. Anal area large, occupying more than 70% of the tegmen width; f at 1/4 of the CuP base.
Description of holotype (figures 3.H, 5.E). Female tegmen 12.1 mm long, as preserved (lacking just the apical margin in this specimen), and 6.1 mm wide. Both the costal and anal margins convex. Costal area very wide, occupying around 50% of the tegmen length. Notably long ScP , multibranched. Radial area wide although not wider than the cubital area. Long RP , slightly curved at its base, following quite straight towards the apex; seven long and pectinated secondary branches. MA origin at the mid-length of the tegmen, unbranched. Two-branched MP . Fivebranched CuA multiple forked. CuP strongly curved towards the anal margin. Cubital area very long, around 1/2 of the wing height ( aml ) occupying more than 80% of the wing ( amh ). Last RA distal secondary branch, oblique to MA origin, reaching the costal margin after the mid-length of the tegmen, MA origin and CuP distal extremity perpendicularly aligned to both costal and anal margins. The paratypes PULR-I 277 (part, figure 3.I), and PZ-CTES 7274 (counterpart, figure 3.J), from Picos Gemelos (Gualo), La Rioja, Argentina have tegmen 13.3 mm long and 6.6 mm wide, ovoid-like, with the same diagnostic characters. Other material attributable to this species, PULR-I 273 (figure 3.K), and PZ-CTES 7277 (figure 3.L), from Picos Gemelos (Gualo), La Rioja, Argentina, consisting of a tegmen fragment 10.3 mm long and 5.3 mm wide, as preserved, virtually identical to the previous material.
Discussion. This new species has the same venation pattern, the same ground plain, especially the multiple forked CuA as for Hermosablatta pectinata sp. nov. The size difference and the elytra shape are indicative of female venation. The proposed new species is justified because exist no proof that this specimens constituent the female of H. pectinata sp. nov. and not of other possible male ones.
Hermosablatta ? sp.
Figures 3.R, 5.F
Material. PULR-I 275.
Locality. Picos Gemelos (Gualo), La Rioja Province, Argentina.
Stratigraphic provenance and age. As for Argentinoblatta herbsti sp. nov.
Description (figures 3.R, 5.F). Hind wing fragment, possibly attributable to Hermosablatta gen. nov., 11.25 mm long, as preserved, and 5 mm wide. RA very long, unbranched with at least eight short pectinated distal secondary branches. RA reaching the costal margin around 1/3 of the apex. Long RP , slightly curved towards the costal margin; five long pectinated secondary branches; just dichotomous RP4 and 5 . MA long and parallel to RP , unbranched, with long distal dichotomy; MA origin between 1/4 and 1/3 of the tegmen length. Two-branched MP , both with long distal dichotomy. Six-branched CuA .
Family uncertain
Lariojablatta Martins-Neto and Gallego gen. nov.
Type species. Lariojablatta chanarensis Martins-Neto and Gallego sp. nov.
Etymology. Refers to La Rioja (Argentina), the province from which the material originates.
Diagnosis. Tegmen between two and three times longer than wide, with unbranched ScP and RA . MA origin at 1/4 of the tegmen base, two-branched, each one polychotomous. Presence of r-m.
Discussion. This peculiar group of blattopterans was recorded by the authors in the Cerro Bayo locality (Potrerillos, Mendoza province), its stratigraphical procedence still uncertain (upper section of Cerro de Las Cabras Formation or lower section of the Potrerillos Formation). This paleoentomofauna, including Blattoptera, will be the focus of a forthcoming paper. Complete tegmens of the group found in this new locality have consistent diagnostic characters for creation of a new family, apparently endemic to the Argentinian Triassic, close to Subioblattidae Schneider, also recorded in the mentioned locality.
Lariojablatta chanarensis Martins-Neto and Gallego sp. nov.
Figure 4.A, B; 5.G

Figure 4. A-B, Lariojablatta chanarensis sp. nov., drawn from the holotype, PULR-I 221 (A) and wing reconstruction (B), respectively / dibujado del holotipo, PULR-I 221 (A) y reconstrucción del ala (B), respectivamente. C-G, Argentinoblattula revelata sp. nov., drawn from the holotype, PULR-I 286 (C), and from the supposed supplementary material, PULR-I 294 (D), PULR-I 295 (E), and PULR-I 293 (F), and wing reconstruction (G), respectively / dibujado del holotipo, PULR-I 286 (C), y del supuesto material suplementario, PULR-I 294 (D), PULIR 295 (E), y PULR-I 293 (F), y reconstrucción del ala (G), respectivamente. H-Z, Blattoptera incertae sedis tegmen fragments / fragmentos de tegminas (R, PULR-I 297; N, PZ-CTES 7275b; others / otros PZ-CTES 7293a-h). H, S, W, Y, Triassoblatta ? sp. (PULR-I 272; PZ-CTES 7292b-c, g); T, U, V, X, Z Samaroblatta ? sp. (PULR-I 296, 286a; PZ-CTES 7292d-f). Abbreviations discussed in the text. Scale bar = 2 mm / Abreviaturas discutidas en el texto. Escala = 2 mm.
Holotype. PULR-I 221.
Etymology. Refers to Los Chañares (La Rioja), geographic unit from which the material came.
Type locality. Los Chañares, La Rioja, Argentina.
Type stratum. 1st cycle from Los Rastros Formation.
Age. Early Late Triassic.
Diagnosis. As for the genus. Tegmen around 17 mm long and 6.5 mm wide.
Description of the holotype (figures 4.A, 5.G). Tegmen 17.1 mm long and 6.4 mm wide, as preserved. Long ScP , with one preserved branch. Unbranched RA without distal dichotomy. Short RP , four-branched; RP4 reaching the costal at the boundary with the apical margin. Long MA , two-branched with polychotomous MA1 and dichotomous MA2 ; MA origin at 1/4 of the tegmen base; presence of a conspicuous cross vein connecting RP4 to MA1 ( rm ). Three-branched MP and all of them with a long distal dichotomy. Two-branched CuA , as preserved; CuA1 branching again into two secondary branches, each of these branching again; CuA2 with a long distal dichotomy.

Figure 5. A, Argentinoblatta herbsti sp. nov., holotype / holotipo , PULR-I 220. B, Condorblatta lutzae sp. nov., paratype / paratipo , PULR-I 264. C, Mancusoblatta pulchella sp. nov., holotype / holotipo , PULR-I 288. D, Hermosablatta pectinata sp. nov., holotype / holotipo , PULR-I 284. E, Hermosablatta crassatella sp. nov., holotype / holotipo , PULR-I 289. F, Hermosablatta ? sp., PULR-I 275. G, Lariojablatta chanarensis sp. nov., holotype / holotipo , PULR-I 221. H, Triassoblatta argentina sp. nov., holotype / holotipo , PULR-I 222. I, Pulchellablatta nana sp. nov., holotype / holotipo , PULR-I 274. J, Samaroblatta gualoensis sp. nov., holotype / holotipo , PULR-I 279. K, Samaroblatta corrientesina sp. nov., holotype / holotipo , PULR-I 282. L, Argentinoblattula revelata sp. nov., holotype / holotipo , PULR-I 286. Scale bar / Escala = 2 mm.
Family Mesoblattinidae Handlirsch, 1906 sensu stricto
Triassic genera included. Austroblattulla Tillyard, 1919, Triassoblatta Tillyard, 1919 and Samaroblatta Tillyard, 1919, with a world-wide Triassic distribution (Argentina, Japan, Tasmania, Africa, Australia and Germany; figures 2.D-G) and Pulchellablatta gen. nov. from the Argentinian Triassic.
Diagnosis. Apart the diagnostic characters listed by Handlirsch, 1906, the follow biometric data, restricted for Triassic genera: lcm +/-= aml > oMA .
Genus Triassoblatta Tillyard, 1919
Type species. Triassoblatta typica Tillyard, 1919, p. 368-370, text-fig. 30a, 31.
Triassoblatta argentina Martins-Neto and Gallego sp. nov.
Figures 3.S-U; 5.H.
1997. Samaroblatta sp. Gallego. p. 513, lám. I, fig. D.
Holotype. PULR-I 222, part (figure 3.S) and PZ-CTES 5730, counterpart.
Paratype. PULR-I 290 (figure 3.T).
Etymology. From the República Argentina, the country from which the material came.
Type locality. Picos Gemelos, La Rioja, Argentina.
Type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Diagnosis. Branched ScP ; multibranched RA ; twobranched MA . The parameters lcm , oMA and aml forming an angle < 90º, lcm slightly longer than aml , MA origin close to the tegmen base.
Description of the holotype (figures 3.S, 5.H). Tegmen 12.9 mm long, as preserved, and 6.5 mm wide. Costal margin parallel to the anal margin. Costal area relatively small, occupying around 1/3 of the wing length. Relatively long ScP , two-branched. Three-branched RA . Radial area relatively wide, although narrower than the cubital area. RP long and sigmoid, reaching the apical area probably at the apex; five long pectinated secondary branches preserved, all of them dichotomous. Long MA , twobranched; MA origin at 1/2 of the anal area length. Three-branched MP , parallel and short. Multibranched CuA . CuP curved towards the anal margin. Cubital area restricted to 1/3 of the wing length occupying around 50% of the wing width. Last RA distal secondary branch, MA origin, and CuP distal extremity forming a quite straight angle. Last RA distal secondary branch and MA origin aligned with an angle around 45º in relation to the latitudinal axis and MA origin and CuP distal extremity origin aligned with an angle around 45º in relation to the anal margin.
Discussion. Triassoblatta argentina sp. nov. differs from all known species in having lcm longer and oMA ; lcm , oMA and aml forms an isosceles triangle, escaleno in the other known species as T. natalensis Riek, 1974, T. robusta Riek from the African Triassic, T. Tasmanica Riek, 1962 from the Tasmanian Triassic and T. Okafuji Fujiyama, 1973 from the Japanese Triassic, and T. Typica Tillyard, 1919 from the Australian Triassic. Brauckmann and Schlüter (1993) reported possible species attributable to Triassoblatta from the Triassic of Germany, although a precise determination has not been made. Another specimen attributable to this genus is PULR-I 290, a tegmen fragment 5.2 mm long, as preserved and 4.2 mm wide (figure 3.T). This specimen is similar to the holotype by having threebranched RA , however, differing in having an extra branch of MA and unbranched MP . Additionally, this specimen is smaller and the anal area is notably smaller, occupying no more than 1/3 of the tegmen length (around 50% in the holotype). Other tegmen fragments possibly attributable to the genus Triassoblatta are: figure 4.H (PZ-CTES 7292b), 4.S (PULR-I 272), 4.W (PZ-CTES 7292g) and 4.Y (PZ-CTES 7292c).
Genus Samaroblatta Tillyard, 1919
Type species. Samaroblatta reticulata Tillyard 1919, p. 374-375, textfig. 30b, 34.
Samaroblatta gualoensis Martins-Neto and Gallego sp. nov.
Figures 3.V-Z; 5.J
Holotype. PULR-I 279.
Paratypes. PULR-I 280 (figure 3.X), part and PZ-CTES 7275 (figure 3.Y), counterpart; PULR-I 281 (figure 3.W), 2 specimens.
Etymology. Refers to Río Gualo, local geographic name where the material came from.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Diagnosis. Tegmen with the anal area occupying 50% of the length and around 80% of the width; f = 1/4; RA distal extremity, MA origin and CuP distal extremity forming an angle around 90º.
Description of the holotype (figures 3.V, 5.J). Tegmen fragment 9.1 mm long, as preserved and 5.8 mm wide. Costal margin slightly curved. Costal area long and narrow around 2/3 the length of the anal area. Long ScP , unbranched. Unbranched RA . Radial area relatively wide although narrower than the cubital area. RP long and slightly curved; five long, pectinated secondary branches preserved, RP2 and 4 dichotomous. MA origin at 1/2 the length of the anal area. MP and CuA with two preserved branches, each branching again. CuP curved toward the anal margin. Cubital area occupying around 80% of the wing width. RA distal extremity, MA origin and CuP distal extremity forming a quite straight angle; RA distal extremity and MA origin aligned with an angle around 45º in relation to the latitudinal axis; MA origin and CuP distal extremity origin aligned with an angle around 45º in relation to the anal margin. Intercalary veins not detected. The paratype PULR-I 280 (figure 3.X, part) and PZ-CTES 7275 (figure 3.Y, counterpart), consisting of a tegmen fragment 11.2 mm long, as preserved and 5.4 mm wide. This specimen differs from the holotype by having the anal area smaller, shared by other known species of the genus such as S. reticulata Tillyard, 1919 from the Australian Triassic and S . fronda Fujiyama, 1973 from the Japanese Triassic and by the MA origin around 2/3 of the anal area length (around 1/2 in S . gualoensis , S. reticulata and S. fronda ). Other paratype, PULRI 281 (figure 3.W) has a tegmen 12.5 mm long, as preserved and 5.4 mm wide and is similar to Samaroblatta reticulata Tillyard, 1919 from the Australian Triassic in its RA and MA morphology, but differs in having unbranched MA (two-branched in S . reticulata ) and MP reaching the apical margin above the apex (below in S . reticulata ).
Discussion. Samaroblatta gualoensis sp. nov. differs from all known species by having a notably large anal area. Although the anal area do not be totally preserved, CuP is, so tracing d and f , we can matemathically estimates with a good precision the size of the anal area.
Samaroblatta corrientesina Martins-Neto and Gallego sp. nov.
Figures 3.AA-AD; 5.K
Holotype. PULR-I 282.
Paratypes. PULR-I 287 (figure 3.AA.), PULR-I 292, PZ-CTES 7296 (figure 3.AB), 2 specimens.
Etymology. Refers to Corrientes Province, Argentina, honouring the team of technician, students and fellows from the Área Paleontología (CECOALCONICET) and Paleontología (FACENA-UNNE) Corrientes, for their friendship and dedication preparing, separating, cleaning and cataloguing the collections.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Diagnosis. Tegmen with both RP and CuA secondary branches oblique, forming a fishbone-like pattern of venation. Costal area as wide as cubital area. MA origin close to the tegmen base. Anal area occupying more than 50% of the tegmen length and width. RA distal extremity, MA origin and CuP distal extremity forming an angle around 120º.
Description of the holotype (figures 3.A.C, 5.K). Tegmen 9.6 mm long, as preserved and 7.3 mm wide. RP long, slightly curved at its base, following quite straight towards the apex, reaching the apical margin above it; eight long, pectinated secondary branches preserved. MA long and slightly divergent from unbranched RP ; MA origin close to the tegmen base. Two-branched MP , and just MP2 with a distal dichotomy. Six-branched CuA , all of them oblique to the anal margin and undichotomous. CuP strongly curved towards the anal margin. Cubital area very long around 1/2 the length and width of the tegmen. RA distal extremity, MA origin and CuP distal extremity, forming an angle around 120º; f = 1/4. PULR-I 287 (figure 3.A.A) and 292, are a tegmen fragment 6.8 mm long, and PZ-CTES 7296 (figure 3.AB) is 5.0 mm long, as preserved and 5.4 mm wide, with a similar venation pattern to previously described specimens. Other tegmen fragments possibly attributable to the genus Samaroblatta are: figure 4.T-X, Z (PULR-I 296, 286a; PZ-CTES 7292d-f).
Discussion. S. corrientesina sp. nov. differs from all previously described species of the genus by having both RP and CuA secondary branches oblique at the same angle, without distal dichotomies, pectinated, forming a fishbone-like pattern of venation. Additionally, S . corrientesina sp. nov. has a notably greater anal area, occupying around 50% of the tegmen length and width.
Pulchellablatta Martins-Neto and Gallego gen. nov.
Type species. Pulchellablatta nana Martins-Neto and Gallego sp. nov.
Etymology. Diminutive of pulcher (Latin), beautiful, and blatta, common suffix for blattoids.
Diagnosis. Small sized tegmen. Anal area occupying more than a half of the total wing length and anal veins multibranched.
Discussion. Pulchellablatta gen. nov. has the new proposed formula for the family: lcm = aml > oMA . Similar to Samaroblatta in the long and narrow costal area by having RA distal extremity (although the specimen only preserves the RA base, the reconstruction of it following a straight line, reaching the costal margin at the same level of CuP distal extremity), MA origin and CuP distal extremity forming an angle around 120º. Pulchellablatta gen. nov. has a large anal area, occupying 50% of the tegmen length and 90% of the width. The small size allied with multibranched anal veins are all characters not present in all of the previously described species, justifying the creation of a new genus for this specimen.
Pulchellablatta nana Martins-Neto and Gallego sp. nov.
Figures 3.AE, AF; 5.I
Holotype. PULR-I 274.
Etymology. Refers to the small size of the species.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Diagnosis. As for the genus. Tegmen around 9 mm long and 3 mm wide.
Description of the holotype (figures 3.D, 5.I). Tegmen 8.8 mm long and 3.3 mm wide. Costal margin slightly curved. Radial area narrow around 1/3 of the cubital area. Long RP , slightly sigmoid, reaching the apical area little below the apex; six relatively short pectinated secondary branches. MA relatively long and parallel to RP , unbranched; MA origin after the mid-length of the tegmen. Four-branched MP , all of them undichotomous. Three-branched CuA , all of them oblique to the anal margin; CuA1 and CuA3 with distal dichotomy; pentachotomous CuA2 . CuP slightly curved towards the anal margin. Cubital area long more than 1/2 of the wing length occupying around 90% of the wing width. At least five anal veins, dichotomous and polychotomous.
Family Blattulidae Vischnyakova, 1982
Emended diagnosis. Apart from the characteristics Triassic blattopteran insects from Argentina 719 discussed by Vischnyakova (1982), tegmen with RP and MA secondary branches restricted to the apical area, above the apex. RA and CuP distal extremity perpendicularly aligned to both costal and anal margin and MA origin after this alignment ( lcm = aml < oMA ). Two-branched MA .
Argentinoblattula Martins-Neto and Gallego gen. nov.
Type species. Argentinoblattula revelata Martins-Neto and Gallego sp. nov.
Etymology. Refers to the República Argentina, the country from which the material originates, and Blattula, the closest genus.
Diagnosis. As for the family. Tegmen three times longer than wide. ScP long and unbranched. RP apparently two-branched.
Discussion. Argentinoblattula gen. nov. differs from the known Jurassic and Cretaceous Blattulidae genera by having both RA distal extremity and MA origin closest to the tegmen base (at the mid-length or further in non Triassic Blattulidae) and both RP and MA secondary branches restricted to the apical margin, above the apex (just the RP ones above the apex and with the MA ones below in non Triassic Blattulidae). Additionally, Argentinoblattula gen. nov. differs from the Jurassic and Cretaceous Blattulidae by having the tegmen with two-branched MA (unbranched in the non Triassic Blattulidae). Argentinoblattula gen. nov. shares with the Jurassic and Cretaceous Blattulidae genera a long and notably narrow tegmen, a generalized reduction of the secondary branches and an apparent division of RP in two main branches and a notably long and unbranched ScP . The apparent branching of RP in non Triassic Blattulidae took place distally, after the tegmen mid-length, when the RP secondary branches of oblique, undichotomous and pectinated are now divided in two clade-like polychotomous branches. In Argentinoblattula gen. nov. the apparent RP division took place close to the tegmen base (around 1/3), with the anterior distal branch following the initial pattern of branching (oblique, undichotomous and parallel) and the posterior one cladelike, polychotomous (a probable autapomorphy of Argentinoblattula gen. nov.).
Argentinoblattula revelata sp. nov.
Figures 4.C-G; 5.L
Holotype. PULR-I 286.
Etymology. From revelatus (Latin), refers to the fact that the specimen was entirely covered by sediment before it was exposed.
Type locality, type stratum, and age. As for Argentinoblatta herbsti sp. nov.
Diagnosis. As for the genus.
Description of the holotype (figures 4.C, 5.L). Tegmen 10.9 mm long and 2.5 mm wide, as preserved. Long ScP , occupying 40% of the tegmen length, unbranched. Unbranched RA . Four-branched RP , RP1 and RP2 the shortest. RP3 + 4 forks at 1/3 of the tegmen length, RP3 with four oblique pectinated secondary branches and three-branched RP4 . The apparent RP division took place close to the tegmen base (around 1/3), with the anterior distal branch following the initial pattern of branching (oblique, undichotomous and parallel) and the posterior one clade-like, polychotomous. Two-branched MA origin, between 1/3 and 1/4 from the wing base. Twobranched MP . Three-branched CuA , with four distal secondary branches Anal area very small, restricted to the wing base. The parameter lcm = aml < oMA ; f =3/4.
Other material possibly attributable to this species are: PULR-I 293 (figure 4.F), a tegmen fragment of cubital area and a turn on anal area, PULR-I 294 (figure 4.D), a tegmen fragment of the central area and PULR-I 295 (figure 4.E), an apical fragment.
Blattoptera incertae sedis tegmen fragments
Several other tegmen fragments were found at the same stratigraphic levels, as shown in figures 4 (HZ), but they are poorly preserved and lack diagnostic elements for a formal description and identification. The Los Rastros Blattoptera thanatocoenosis is formed mainly by fragmented tegmens (just one complete, a few quite complete, and just one hind wing).
Final comments
According to Jarzembowski and Ross (1996), the diversity of Polyneoptera (including Blattoptera) shows a marked increase since Middle Triassic. The family level diversity of the hexapods shows a notable decrease in the Middle Triassic, accompanied by a low turnover, with a recovery in the Upper Triassic. Shear and Kukalova-Peck (1990) mentioned that the blattoid stem group, ancestors of Recent cockroaches, termites and earwigs, are the most abundant insects in nearly all Carboniferous and Lower Permian insect localities, but are often known only from their tough tegmen (fore wings), which survive transport in water and so were readily preserved. Kukalova-Peck (1991) mentioned that the extinct Poroblattinidae were common in the Permian, up to now only known from the Pennsyalvanian (Gehelian) to Lowermost Permian (Asselian-J. Schneider pers. com., 2004) and Triassoblatta is known from the Triassic of Australia, Tasmania and South Africa. Mesoblattinidae were dominant in the northern hemisphere in the Mesozoic and they also lived in the Upper Triassic of South Africa. Now we know that Poroblattinidae were as common as Mesoblattinidae during the Gondwanan Triassic, and Mesoblattinidae are abundant in the Lower Cretaceous, mainly from Brazil (Santana Formation) but also from Spain (Montsech, Lerida). The distribution of the genera Triassoblatta and Samaroblatta during Triassic times, apart from Australia, Tasmania and South Africa, as pointed by Kukalova-Peck (1991), extends now to Argentina, Japan (Fujiyama, 1973 not mentioned by Kukalova-Peck, 1991), China and Germany (Brauckmann and Schlüter, 1993).
Triassic insect records from the Gondwanan continent show that the main orders are Coleoptera (beetles), Auchenorrhyncha (bugs, harvest flies, planthoopers) and Blattoptera (cockroaches). These records came from Australia, South Africa and southern South America (Brazil, Chile and mainly from Argentina). The number of blattopteran species described in each area are: Australia (eleven species), South Africa (ten species), Brazil (one species) and Argentina (fifteen species; eleven from Los Rastros Formation, described here and four from Potrerillos Formation, to be described elsewhere).
The blattopteran record corresponds to nearly 10% of the total insect record of each area: Molteno Formation, 3% of species and Ipswich series, 9%. For the Los Rastros Formation (Argentina) blattopterans constitute around 21% (60 specimens) of the described species. The number of collected specimens varies (maybe paleoecologically distinct cases): Molteno Formation around 46%, Ipswich, 5% (data from Anderson et al ., 1998; Anderson, 1999) and the Los Rastros around 25%. The most significant data in this respect are furnished by Anderson et al . (1998), in which the cockroaches easily outnumber other insect groups in the Molteno Formation. With 956 individuals, mostly isolated tegmens, less common nymphs, abdominal, hind wings or fully articulated adults, from 34 taphocoenoses they comprise nearly half the total faunal count of 2056 individuals. In diversity, however, with a mere three genera and ten species recognized, they only come sixth after the beetles, bugs and three other orders. For the Argentinian taphocoenoses this situation is very different, because blattopterans constitute around 21% of the described species (an explosive diversification rate), but in collected specimens terms the proportion is low (around 25%). By contrast, in the Molteno Formation, the coleopterans have 30 recognized genera (25% of total insect genera) and 161 described species (48% of the total insect species). But in the number of collected specimens the proportion is lower, constituting more than 22%, 99% of which are isolated tegmens (just one hind wing has been detected until now).
Extant cockroach nymphs and adults exhibit the same patterns of habitat and feeding, both being essentially terrestrial omnivores. They are primarily nocturnal, unspecialised scavengers in the leaf litter, feeding largely on decomposing plant material. In Molteno, their relative abundance is marked as the highest among closed-canopy terrestrial habitats- Heidiphyllum thicket, Sphenobaiera closed woodland and the Dicroidium riparian forest-where the leaf litter, ideal for their scavenging would have accumulated (Anderson et al ., 1998).
The abundance and diversity of the Los Rastros blattopterans were probably due to the large Dicroidium forests and the Taeniopteris and Baiera thickets around or next to the Los Rastros lake area, which provided many ecotopes where the blattopterans could survive, consuming litter. According to Shear and Kukalova-Peck (1990), they had a major role in the mechanical reduction of litter. These habitats probably were not very close to the Los Rastros lake, according to preliminary taphonomic evidence, the blattopterans are only preserved as isolated wings, and articulated specimens are still unrecorded, which suggests that they could have been transported from a considerable distance.
Acknowledgements
Thanks are due to Carsten Brauckmann (Institut für Geologie und Paläontologie, Tu Clausthal, Germany) for his friendship, comments and suggestions as one of the journal reviewers. Also to two other reviewers: Andre Nel and Julian Petrulevicius for their interesting comments and suggestions on the manuscript. Francine Papier (Université Louis Pasteur, Strasbourg, France) for her interesting comments and criticism and Jörg Schneider (Freiberg University of Mining and Technology, Institute of Geology, Department of Palaeontology, Freiberg, Germany) for opportune comments suggestions and criticism as one of the journal reviewers. Also to Euan Clarkson (University of Edinburg, Scotland) and Gregory Edgecombe (Australian National Museum, Sydney) for fine suggestions on the manuscript. We also acknowledge Ms. Andrea Arcucci, Drs. Catherine Forster, Cathleen May and Raymond Rogers from the project of the National Geographic Society, Grant 5317-94. Gustavo Barrios for making the digital figures and Ramón Z. Rodríguez for his work in the photographic laboratory. María Elena Gonaldi from the Museo de Ciencias Naturales (Universidad Nacional de La Rioja) for her assistance with the cataloguing of the specimens. This research was partly supported by the Consejo Nacional de Investigaciones Científicas y Técnicas (PIP-5581) and by the Secretaría General de Ciencia y Técnica, Universidad Nacional del Nordeste (PI-64/04) , both to O.F.G.
References
Anderson, J.M. (ed.) 1999. Towards Gondwana Alive. Promoting biodiversity and stemming the Sixth Extinction. Gondwana Alive Society, Pretoria, 140 pp. [ Links ]
Anderson, J.M., Anderson, H.M. and Cruickshank, A.R.I. 1998. Late Triassic ecosystems of the Molteno/Lower Elliot biome of southern Africa. Palaeontology 41: 387-421. [ Links ]
Bossi, G. 1971. Análisis de la Cuenca Ischigualasto-Ischichuca. 1 º Congreso Hispano-Luso-Americano de Geología Económica , (Madrid/Lisboa), Actas 2: 611-626. [ Links ]
Brauckmann, C. and Schlüter, T. 1993. Neue Insekten aus der Trias von Unter-Franker. Geologica et Palaeontologica 27: 181-199. [ Links ]
Brauer, F., Redtenbacher, J. and Ganglbauer, L. 1889. Fossile Insekten aus der Juraformation Ost-Sibiriens. Memoires of Academy Imperial of Sciences of Saint Petersburg, serie 7 , 15: 1-22. [ Links ]
Caselli, A.T., Marsicano, C.A. and Arcucci, A.B. 2001. Sedimentología y paleontología de la Formación Los Colorados, Triásico superior (provincias de La Rioja y San Juan, Argentina). Revista de la Asociación Geológica Argentina 56: 173-188. [ Links ]
de la Mota, H.F. 1946. [ Estudios geológicos en el Cerro Bola, al sur de Villa Unión, Dpto. General Lavalle, provincia de La Rioja. Universidad Nacional de La Plata, Tesis Doctoral 87, La Plata, Argentina, 145 p. Unpublished]. [ Links ]
Dodds, B. 1949. Mid-Triassic Blattoidea from the Mount Crosby insect beds. University of Queensland Department of Geology 3: 1-11. [ Links ]
Farquharson, G.W. 1982. Lacustrine deltas in a Mesozoic alluvial sequence from Camp Hill, Antarctica. Sedimentology 29: 717-725. [ Links ]
Frenguelli, J. 1948. Estratigrafía y edad del llamado Rético en la Argentina. Anales de la Sociedad Argentina de Estudios Geográficos, Sociedad Argentina de Estratigrafía Geológica, GAEA 8: 159-309. [ Links ]
Fujiyama, I. 1973. Mesozoic insect fauna of East Asia. Part 1. Introduction and Upper Triassic faunas. Bulletin of Natural Sciences Museum Tokyo 16: 331-386. [ Links ]
Gallego, O.F. 1997. Hallazgos de Insectos triásicos en la Argentina. Ameghiniana 34: 511-516. [ Links ]
Gallego, O.F. and Martins-Neto, R.G. 1999. La entomofauna mesozoica de la Argentina: Estado actual del conocimiento. Revista de la Sociedad Entomológica Argentina 58: 86-94. [ Links ]
Handlirsch, A. 1906. Die Fossilen Insekten und die Phylogenie der Rezenten Formen. Ein Handbuch für Paläontologie und Zoology. Engelmann (ed.), Leipzig, 640 pp. [ Links ]
Handlirsch, A. 1912. Über Insektenreste aus der Trias Frankens. Abhandlungen der naturhistorischen Gesellschaft Nümberg 18: 79-82 [ Links ]
Handlirsch, A. 1939. Neue Untersuchungen über die fossilen Insekten. II Teil. Annals of Naturhistorischen Museum of Wien 49: 1-240. [ Links ]
Hong, Y. 1980. Fossil insects. In: Institute of Geology (eds.), Mesozoic stratigraphy and palaeontology of basins of Shansi, Gansu and Ningxia. Chinese Academy of Geological Sciences 2: 111-114. [ Links ]
Jarzembowski, E.A. and Ross, A.J. 1996. Insect origination and extinction in the Phanerozoic. In: M.B. Hart (ed.), Biotic Recovery from Mass Extinction Events, Geological Society Special Publication 102: 65-78. [ Links ]
Kokogian, D., Spalletti, L.A., Morel, E.M., Artabe, A.E., Martínez, R.N., Alcober, O.A., Milana, J.P, Zavattieri, A.M. and Papú, O.H. 1999. Los depósitos continentales triasicos. In: R. Caminos and J. Panza (eds.), Geología de la República Argentina. Instituto de Geología y Recursos Minerales, Anales 29: 377-398. [ Links ]
Kokogian, D., Spalletti, L.A., Morel, E.M., Artabe, A.E., Martínez, R.N., Alcober, O.A., Milana, J.P, and Zavattieri, A.M. 2001. Estratigrafía del Triásico argentino. In: A.E Artabe, E.M. Morel and A.B. Zamuner (eds.), El Sistema Triásico en la Argentina. Fundación Museo de La Plata Francisco Pascasio Moreno, La Plata, pp. 23-54. [ Links ]
Kukalova-Peck, J. 1991. Fossil History and the Evolution of Hexapod Structures. In: I.D. Naumann, P.B. Carne, J.F. Lawrence, E.S. Nielsen, J.P. Spradbery, R.W. Taylor, M.J. Whitten and M.J. Littlejohn (eds.), The Insects of Australia. University Press, Melbourne, pp. 141-179. [ Links ]
Lin, Q. 1978. On the fossil Blattodea of China. Acta Entomologica Sinica 21: 335-344 [ Links ]
López Gamundi, O.R., Alvarez, L., Andreis, R., Bossi, G., Espejo, I., Fernandez-Seveso, F., Legarreta, L., Kokogian, D., Limarino, C. and Sesarego, H. 1989. Cuencas Intermontanas. In: G. Chebli and L. Spalletti (eds.), Cuencas Sedimentarias Argentinas. Serie Correlación Geológica, Universidad Nacional de Tucumán 6: 123-167. [ Links ]
Mancuso, A.C. 1999. [ Análisis tafonómico de una asociación fósil continental del Triásico Medio de la Cuenca de Ischigualasto-Villa Unión (Provincia de La Rioja). Ms thesis, Universidad de Buenos Aires, Buenos Aires, 71 pp. Unpublished]. [ Links ]
Mancuso, A.C. 2000a. Caracterización tafonómica del ambiente lacustre del Triásico medio de la Cuenca Ischigualasto-Villa Unión, Argentina. Generalización. 2º Congreso Latinoamericano de Sedimentología-7º Reunión Argentina de Sedimentología (Mar de Plata), Actas : 105-106. [ Links ]
Mancuso, A.C. 2000b. Análisis tafonómico de los peces de la Formación Los Rastros (Triásico, La Rioja). Ameghiniana Suplemento Resúmenes 37: 28R. [ Links ]
Mancuso, A.C. 2000c. Reconstrucción paleoecológica de la biota registrada en la Formación Los Rastros (Triásico, La Rioja). Ameghiniana Suplemento Resúmenes 37: 41R. [ Links ]
Mancuso, A.C. 2003. Continental fish taphonomy: a case study in the Triassic of Argentina. Journal of South American Earth Sciences 16: 275-286. [ Links ]
Mancuso, A.C. and Gallego, O.F. 2000. Primer análisis tafonómico de una asociación fósil de conchóstracos e insectos del triásico medio (Fm. Los Rastros) de la Argentina. First International Meeting on Paleoarthropodology (Ribeirão Preto), Actas : 64. [ Links ]
Martins-Neto, R.G. and Gallego, O.F. 1999. The Triassic Insect Fauna from Argentina. I. Auchenorrhyncha, Miomoptera and Ensifera. Revista Española de Paleontología 14: 191-202. [ Links ]
Martins Neto, R.G. and Gallego, O.F. 2001. The Triassic Insect Fauna from Argentina. IV. Glosselytrodea and complements on Auchenorrhyncha. Acta Geologica Leopoldensia 24: 105-114. [ Links ]
Martins-Neto, R.G., Gallego, O.F. and Melchor, R.N., 2003. The Triassic insect fauna from South America (Brazil, Argentina and Chile): a checklist (except Blattoptera and Coleoptera) and descriptions of new taxa. Acta Zoologica Cracoviensia 46 (suppl.- Fossil Insects): 229-256. [ Links ]
Martynov, A.V. 1937. Liasovye nasekomye Shuraba i Kizil-kii. Trudy Paleontologicheskogo Instituta, SSSR 7: 5-232. [In Russian]. [ Links ]
Milana, J.P. 1998. Anatomía de parasecuencias en un lago de rift y su relación con la generación de hidrocarburos, cuenca triásica de Ischigualasto, San Juan. Revista de la Asociación Geológica Argentina 53: 365-387. [ Links ]
Milana, J.P. and Alcober, O.A. 1994. Modelo tectosedimentario de la cuenca triásica de Ischigualasto (San Juan, Argentina). Revista de la Asociación Geológica Argentina 49: 217-235. [ Links ]
Ottone, E.G., Mancuso, A.C. and Resano, M. (2005). Miospores and Chlorococcales from the Los Rastros Formation, Middle Triassic of central- western Argentina. Ameghiniana 42: 347-362. [ Links ]
Papier, F. and Grauvogel-Stamm, L. 1995. Les Blattodea du Trias: Le genre Voltziablatta n. gen. Du Buntsandstein supérieur des Vósges (France). Palaeontographica A 235: 141-162. [ Links ]
Papier, F., Grauvogel-Stamm, L and Nel, A. 1994. Subioblatta undulata n. sp., une nouvelle blatte (Subioblattidae Schneider) du Buntsandstein supérieur (Anisien) des Vosges (France). Morphologie, systématique et affinités. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 5: 277-290. [ Links ]
Papier, F., Grauvogel-Stamm, L and Nel, A. 1996. Nouveaux Blattodea du Buntsandstein supérieur des Vosgues (France). Paleontologia Lombarda (n. s.) 5: 47-59. [ Links ]
Pinto, I.D. 1990. A new Lower Cretaceous blattoid insect from Argentina. Pesquisas 17: 11-14. [ Links ]
Pinto, I.D. and Ornellas, L. 1974. A new insect Triassoblatta cargnini Pinto et Ornellas, sp. nov., a Triassic Blattoid from Santa Maria Formation, South Brazil. Anais da Academia Brasileira de Ciências 46: 515-521. [ Links ]
Riek, E.F. 1962. Fossil insects from the Triassic at Hobart, Tasmania. Papers and Proceedings of the Royal Society of Tasmania 96: 39. [ Links ]
Riek, E.F. 1974. Upper Triassic insects from the Molteno Formation, South Africa. Palaeontographia Africana 17: 19-31. [ Links ]
Riek, E.F. 1976. A new collection of insects from the Upper Triassic of South Africa. Annals of the Natal Museum 22: 791-820. [ Links ]
Rogers, R.R., Arcucci, A.B., Abdala, F., Sereno, P.C., Forster, C.A. and May, C.L. 2001. Paleoenvironment and Taphonomy of the Chañares Formation Tetrapod Assemblage (Middle Triassic), Northwestern Argentina: Spectacular Preservation in Volcanogenic Concretions. Palaios 16: 461-481. [ Links ]
Rohdendorf, R.R. 1962. Fundamentals of Paleontology; Arthropoda, Tracheata, Chelicerata. Amerind Publishing Co., New Delhi, Vol. 9, 1991, 894 pp. [ Links ]
Romer, A.S. and Jensen, J.A. 1966. The Chañares (Argentina) triassic reptile fauna. II Sketch of the geology of the Rio Chañares-Rio Gualo region. Breviora Cambridge 252: 1-20. [ Links ]
Schneider, J. 1983. Die Blattodea (Insecta) des Paläozoikums. Teil 1: Systematik, Ökologie und Biostratigraphy. Freiberger Forschungshafte 382: 106-145. [ Links ]
Scudder, S. H. 1895. Revision of the American fossil cockroaches with description of new forms. Bulletim of the United State Geological Survey 124: 1-176. [ Links ]
Shear, W.A. and Kukalova-Peck, J. 1990. The ecology of Paleozoic terrestrial artropods: the fossil evidence. Canadian Journal of Zoology 68: 1807-1834. [ Links ]
Spalletti, L.A., Artabe, A., Morel, E. and Brea, M. 1999. Biozonación paleoflorístico y cronoestratigrafía del Triásico argentino. Ameghiniana 36: 419- 451. [ Links ]
Stipanicic, P. N. 1983. The Triassic of Argentina and Chile. In: M. Moullade and A. E. M. Nairn (eds), The Phanerozoic Geology of the World II, The Mesozoic , Elsevier Scientific Publications, Amsterdam, pp. 181- 199. [ Links ]
Stipanicic, P.N. and Bonaparte, J. 1979. Cuenca triásica de Ischigualasto-Villa Unión (Provincia de La Rioja y San Juan). In: J.C.M. Turner (ed.), 2º Simposio de Geología Regional, Academia Nacional de Ciencias 1: 523- 575. [ Links ]
Stipanicic, P.N. and Bonetti, M.I.R. 1969. Consideraciones sobre la cronología de los terrenos triásicos argentinos. In: A.J. Amos (ed.), 1º Coloquio Internacional sobre la Estratigrafía y la Paleontología del Gondwana (Mar del Plata, 1967), UNESCO, Ciencias de La Tierra, 2: 1081-1120. [ Links ]
Talbot, M.R. and Allen, P.A. 1996. Lakes. In: H.G. Reading (ed.), Sedimentary Environments: Processes, Facies and Stratigraphy , Blackwell Science, pp 5-36 [ Links ]
Tillyard, R. J., 1919. Mesozoic Insects of Queensland. N° 6. Blattoidea. Proceedings of the Linnean Society, New South Wales 44: 358-382. [ Links ]
Uliana, M.A. and Biddle, K. 1988. Mesozoic-Cenozoic paleogeographic and geodynamic evolution of southern South America. Revista Brasileira de Geociências 18: 172-190. [ Links ]
Vischnyakova, V.N. 1982. Jurassic Cockroaches of the new family Blattulidae from Siberia. Paleontological Journal 2: 67-77. [ Links ]
Vrsansky, P. 1998. Lower Cretaceous Blattaria. Proceedings of the First Paleoentomological Conference (Moscow): 167-176. [ Links ]
Yrigoyen, M.R. and Stover, L.E. 1970. La palinología como elemento de correlación del Triásico en la Cuenca Cuyana. 4º Jornadas Geológicas Argentinas (Mendoza, 1969), Actas 2: 427-447. [ Links ]
Zavattieri, A.M. and Melchor, R.N. 1999. Estudio palinológico preliminar de la Formación Ischichuca (Triásico), en su localidad tipo (quebrada de Ischichuca Chica), provincia de La Rioja, Argentina. Asociación Paleontológica Argentina, Publicación Especial 6: 33-38. [ Links ]
Recibido: 5 de setiembre de 2003.
Aceptado: 28 de diciembre de 2004.

