










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.42 no.4 Buenos Aires Sept./Dec. 2005
Gymnospermous woods from Jejenes Formation, Carboniferous of San Juan, Argentina: Abietopitys petriellae (Brea and Césari) nov. comb.
Roberto R. Pujana 1
1 Museo Argentino de Ciencias Naturales B. Rivadavia. Ángel Gallardo 470, C1405DJR Buenos Aires, Argentina. rpujana@macn.gov.ar
Abstract. Fossil woods from Carboniferous sediments of the Jejenes Formation are described and a new combination Abietopitys petriellae is proposed based on new and the original material. The wood is pycnoxylic with primitive characters such as abundant pits in radial walls of tracheids in an alternate disposition. The ray cells have peculiar wall thickenings. The primary xylem has a mesarch protoxylem. The species is compared with other Paleozoic woods; its botanical affinity remains uncertain.
Resumen. Maderas gimnospérmicas de la Formación Jejenes, Carbonífero de San Juan, Argentina: Abietopitys petriellae (Brea y Césari) nov. comb . Se describen maderas fósiles del Carbonífero de la Formación Jejenes y se propone una nueva combinación Abietopitys petriellae sobre la base de nuevo material, como así también sobre la revisión del material original. La madera es picnoxílica y posee características primitivas, como abundantes puntuaciones en las paredes radiales de las traqueidas con disposición alterna. Las células radiales tienen engrosamientos particulares. El xilema primario posee protoxilema mesarco. Esta especie es comparada con otras maderas paleozoicas, siendo su afinidad botánica incierta.
Key words. Wood; Anatomy; Carboniferous; Jejenes Formation; Argentina.
Palabras clave. Madera; Anatomía; Carbonífero; Formación Jejenes; Argentina.
Introduction
The record of Upper Paleozoic fossil wood is very scarce in Argentina and up to date there are no Devonian fossil woods described. Several studies of Permian pycnoxylic woods have been carried out (Archangelsky, 1960; Herbst and Crisafulli, 1997; Crisafulli et al ., 2000). There are some reports of Carboniferous fossil woods without detailed studies (Mésigos, 1953; Pujana, 2003) and only two Carboniferous species were described, Medullopitys menendezii Petriella (1982) from Mendoza and Phyllocladopitys petriellae Brea and Césari (1995) from San Juan. The latter is here revised and its description complemented with the study of new specimens from the same fossiliferous bearing strata but from a different locality. The diagnosis of the species is emended, and a new combination is proposed.
The Jejenes Formation was defined by Amos (1954) for sedimentary sequences cropping out in the eastern margin of the Sierra Chica de Zonda, south of San Juan city (figure 1). The fossiliferous locality Quebrada de la Mina was described in detail by Cladera et al . (2000) and is the same locality which Bracaccini (1946) called Quebrada de la Cantera de Mármol. Bracaccini (1946) described, at the base of the sequence, conglomerates of glacial origin, and to the upper part of the sequence he noted the presence of abundant paleoflora (see Vega, 1995; Vega and Archangelsky, 2001). This upper section also bears the material here studied.
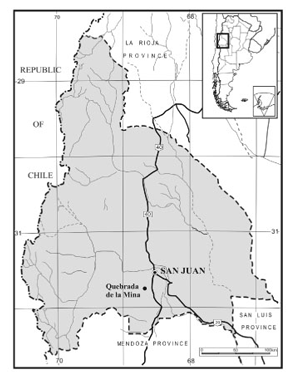
Figure 1. Map showing the fossiliferous locality / Mapa con la ubicación de la localidad fosilífera.
The paleoflora of this unit is referred to the NBG biozone that characterizes the Upper Carboniferous of Argentina (Archangelsky et al ., 1987). There are many species of leaves such as Nothorhacopteris argentinica (Geinitz) Archangelsky, Sphenopteris sarmientoi Vega and Iannuzzi, Fedekurtzia argentina (Kurtz) Archangelsky, Diplothmema bodenbenderi (Kurtz) Césari, Rhacopteris szajnochai Kurtz, Triphyllopteris cuyana Leguizamon and Vega, Eusphenopteris sanjuanina (Kurtz) Césari emend . Césari et al ., Ginkgophyllum diazii Archangelsky and Arrondo and Cordaites sp. Also, there are pteridosperm reproductive structures such as Rinconadia archangelskyi Vega, Austrocalyx jejenensis Vega and Archangelsky, Polycalyx laterale Vega and Archangelsky. The palynological record includes monosaccate pollen grains and spores (Gutiérrez and Césari, 1987; Césari and Bercowski, 1997) that are referred to the biozone Raistrickia densa-Convolutispora muriornata of north-western Argentina. The only wood already described is the one revised in this paper and a polixylic stem was reported by Césari et al . (2005).
According to Cladera et al. (2000) the depositional paleoenvironment of the upper section of the Jejenes Formation corresponds to a lacustrine type with sporadic occurrence of fluvial systems. Recently, Kneller et al . (2004) made a complete paleoenvironmental analysis of all the sequence in the nearby locality Quebrada Grande, and was interpreted as the fill of a glacial valley and its tributaries in a paleofjord environment with catastrophic sedimentation (Kneller et al ., 2004).
Material and methods
Wood taxonomy is based on Lepekhina (1972), who revised the nomenclature and taxonomy of Paleozoic woods worldwide. The wood has been described, when possible, in accordance with the list of microscopic features for softwood identification (Richter et al ., 2004) and the Greguss' glossary of terms (Greguss, 1955).
The new material consists of 5 specimens which were collected in the fossiliferous locality known as Quebrada de la Mina, located at 31°50´34´´ S and 68°37´57´´ W in the eastern margin of the Sierra Chica de Zonda (figure 1). These specimens were compared with the holotype from Las Lajas locality (see Brea and Césari, 1995). Stratigraphically, the fossiliferous bearing strata correspond to the upper part of the Jejenes Formation, which is suggested to be early late Carboniferous in age. All the specimens are silicified (chalcedony) displaying fissures filled with carbonatic material.
Wood fragments were cut in thin sections in the three planes usually used to study their anatomy. Cellulose acetate peels were also made, with poor results compared to thin sections. Good results were obtained from macerations of small wood fragments. Samples were first placed in HF 75%, washed with water, and then mounted in glycerine-jelly. These preparations allowed the observation of fine details such as cross-field pits and ray cell walls thickenings (figures 2.J, 2.K, 2.L).

Figure 2. Abietopitys petriellae nov. comb. A, general view of a transverse section / aspecto general de una sección transversal. Bar: 1 cm (BAPbPm 441). B, transverse section showing distinct growth ring boundaries / sección transversal mostrando los límites de los anillos de crecimiento marcados. Bar: 200 µm (BAPbPm 449). C, transverse section showing an anomalous growth / sección transversal mostrando un crecimiento anómalo . Bar: 500 µm (BAPbPm 441). D, transverse section of the pith / sección transversal de la médula . Bar: 200 µm (BAPbPm 449). E, tracheids of the metaxylem in a radial longitudinal section / traqueidas del metaxilema en una sección radial longitudinal. Bar: 50 µm (BAPbPm 442). F, transverse section showing the mesarch protoxylem / sección transversal mostrando el protoxilema mesarco. Bar: 200 µm (BAPbPm 449). G, transverse section showing the rectangular tracheids and the thickenings in the transverse wall of ray cells / sección transversal mostrando traqueidas rectangulares y engrosamientos en la pared transversal de las células radiales . Bar: 100 µm (BAPb 244 a). H, tangential longitudinal section showing four ray cells, with thickenings or probable pits in the tangential wall (arrows) / sección longitudinal tangencial mostrando cuatro células radiales con engrosamientos o posibles puntuaciones en la pared tangencial (flechas) . Bar: 50 µm (BAPb 244 d). I, radial longitudinal section showing alternate pitting / sección longitudinal radial mostrando puntuaciones de disposición alterna. Bar: 25 µm (BAPbPm 453). J, cross-field pits in a maceration / campos de cruzamiento en un disociado. Bar: 50 µm (BAPbPm 443). K, ray cells in a maceration / células radiales en un disociado . Bar: 50 µm (BAPbPm 443). L, radial longitudinal section showing ray cells with radial wall thickenings in a maceration / sección longitudinal radial mostrando células radiales con engrosamientos en sus paredes en un disociado. Bar: 50 µm (BAPbPm 443).
The studied material is deposited in the Paleobotany collection of the Museo Argentino de Ciencias Naturales (BAPb for fossil wood fragments and BAPbPm for thin sections): BAPb 12906 (with slides BAPbPm 441 to 445), BAPb 12907 (with slides BAPbPm 446 to 449), BAPb 12908 (with slides BAPbPm 450 and 451), BAPb 12909 (with slides BAPbPm 452 and 453) and BAPb 12905. The holotype, BAPB 244 (from Las Lajas locality) includes thin sections (BAPb 244a to 244f, 244l and 244m) and cellulose acetate peels (BAPb 244g to 244j).
Systematic paleontology
Genus Abietopitys Kräusel, in Kräusel and Range, 1928
Type species. Abietopitys perforata Kräusel, in Kräusel and Range, 1928.
Abietopitys petriellae (Brea and Césari) nov. comb.
Figures 2.A-L
Basionym. Phyllocladopitys petriella Brea and Césari, 1995
Holotype. BAPb 244 (with slides 244a to 244m).
Other material. BAPb 12905 to 12909 (with their slides).
Emended diagnosis. Parenchymatous homocellular pith. Primary xylem prominent to the pith, adjacent to the secondary xylem with mesarch protoxylem. Small leaf traces diverge as single bundles. Secondary xylem and tracheids with bordered pits exclusively in radial walls, usually biseriate and with an alternate arrangement. Rays homocellular, mostly uniseriate with a medium average height and with thickenings on the cell walls. Cross-field pits with usually 1 to 5 circular simple pits. Growth ring boundaries distinct.
Description. Because of the similar characteristics of the specimens here described and those of the original specimen, they are regarded as a single taxonomic unit. The maximum width of the specimens is 14 cm and the maximum length is 25 cm, in a secondary xylem fragment (BAPb 12909). The wood is pycnoxylic and resembles that of gymnosperms with tracheids aligned in radial rows (figure 2.G). Only two of the five new specimens have the pith preserved (figure 2.A). The phloem and cortex were not preserved, and there were only two leaf traces observed in one specimen (BAPbPm 449).
The woods present distinct growth ring boundaries (figure 2.B). Ring width varies between specimens from 1.5 to 5 mm. However, the ring widths in the same specimen do not vary very much. The transition from earlywood to latewood is gradual.
The pith is cylindrical, not septated, and has a diameter of 2 cm in the specimen BAPb 12906; minerals usually replace the contact zone with the xylem. In the specimen BAPb 12907 the diameter is 2.5 cm and presents moderate lobes. It is homocellular and composed of polygonal to circular parenchymatic cells, with a diameter in transverse section of 121 µm (55-75 µm). There are intercellular spaces between cells. The cell wall is thin, less than 4 µm of width (figure 2.D).
The primary xylem differentiation is mesarch; individual bundles are prominent into the pith and adjacent to the secondary xylem (figure 2.F). There are up to 11 rows of metaxylem tracheids with a radial diameter of 33 µm (25-45 µm) showing scalariform pits. This is followed by 2 or 3 rows of transition tracheids with similar radial diameter and opposite two-three seriate pits (figure 2.E). Four primary xylem bundles were observed in specimen BAPb 12906, while BAPb 12907, that has a slightly bigger diameter, presents eight. Both specimens have the pith and primary xylem very well preserved.
The secondary xylem is composed of tracheids of rectangular shape with a radial diameter of 30 µm (22-45 µm) and a tangential diameter of 28 µm (17-40 µm) (figure 2.G). Bordered pits are present exclusively in the radial walls. The apertures are elliptical to circular with a diameter of ca . 10 µm. The tracheid pitting is usually biseriate and is arranged in an alternate pattern (figure 2.I). Uniseriate or triseriate pitting is also present. Pitting is always continuous and fills completely the radial wall.
One specimen (BAPb 12906) has an anomalous growth having tracheids in a radial longitudinal view in transverse section (figure 2.C). The anomaly is 9 mm long by 2 mm wide mith centripetal development, and it starts in the second ring (figure 2.C).
The rays are homocellular, exclusively uniseriate and have a medium average height within 1 to 19 cells tall (figure 2.H). There are 4 (3-6) rays per mm. The ray cells are radially very long, which precludes the observation of their endings; when these are discerned, they are oblique (figure 2.K). The ray cells have peculiar thickenings; in the radial walls they have regularly spaced protrusions arising from the upper and lower walls showing a nodular appearance (figures 2.K, 2.L). In the tangential (figure 2.H) and horizontal walls (figure 2.G) the thickenings extend from wall to wall.
The cross-fields have 1 to 5 simple pits, with a mean diameter of ca . 10 µm (figure 2.J) and apparently without any arrangement.
Comparisons
Kräusel, in Kräusel and Range (1928) described the morphospecies of fossil wood Abietopitys perforata from the Permian of the Karoo Supergroup. Later, Kräusel (1956) and Bangert and Bamford (2001) found new specimens of this species, but they did not add details and only reported its presence in a different locality. Archangelsky (1960) added two new species to the genus from the Permian of Chubut, Argentina, A. patagonica and A. crassiradiata . He assigned these two new species to the genus based on the particular thickenings of the ray cell walls. However, following Lepekhina (1972) proposal, these two species should be assigned to a new morphogenus of exclusively secondary xylem of the Abietopitys type, which Lepekhina named Type A. Lepekhina proposed different morphogenera to include those specimens with secondary xylem with pith and primary xylem, and those without it. Her reasoning was that is not unusual to find specimens with the same secondary xylem and different primary xylem or pith and vice versa.
The most particular diagnostic feature of this genus is the presence of thickenings in the ray cell walls, and pits in the tangential ray cell walls (Kräusel, in Kräusel and Range, 1928) which distinguish it from other morphogenus of secondary xylem such as Protophyllocladoxylon Kraüsel with short ray cell walls with smooth walls. The thickenings in the tangential ray cell walls were interpreted by Kräusel (in Kräusel and Range, 1928) as pits, but Archangelsky (1960) suggests that they do not reach such grade of specialization. These thickenings could be localized thickenings of the primary wall, as in the transverse walls of axial parenchyma in some Cupressaceae (Richter et al ., 2004), and not due to pitting. However, the ray cells are not the typical parenchymatic ray cells, and apparently they have some degree of specialization (Archangelsky, 1960). The thickenings on the radial wall with a nodular appearance, slightly resemble the dentate wall of ray tracheids, which are common in some extant conifers of the genus Pinus , among other genera. In the transverse walls, where the thickenings extend from wall to wall, they resemble the ray tracheids of some Callixylon Zalessky (see Chitaley and Cai, 2001, plate I, figures 7 and 8), a Devonian and Carboniferous progymnosperm wood. In the tangential walls the thickenings also extend from wall to wall. Whether they are pits or just thickenings of the tangential walls is difficult to determine. Kräusel (in Kräusel and Range, 1928) compared these thickenings to the pits of the tangential walls of the ray elements of the extant Pinaceae genera Abies and Larix . Based on the similarities he found with Abies , he named the fossil specimens Abietopitys . However, this could lead to confusion because Abies has opposite pitting arrangement on the radial walls of the tracheids and Abietopitys has an alternate arrangement.
The comparison of this new combination with the other species of Abietopitys (table 1) allows to conclude that this is a new species for this genus. The most similar is Abietopitys perforata but the woods described here have different ray cell height, different shape of primary xylem in transverse sections (subtriangular in A. perforata and rounded in A . petriellae ) and different cross-field pitting. They also differ in the age (Permian in A. perforata ).

Table 1. Comparison of diagnostic features of Abietopitys petriellae nov. comb. with the other species assigned to Abietopitys / Comparación de las características diagnósticas de Abietopitys petriellae nov. comb. con las otras especies asignadas al género Abietopitys .
All features described for the new specimens from Quebrada de la Mina are also present in Phyllocladopitys petriellae Brea and Césari (1995); although in the original diagnosis and description the thickenings of the ray cell walls were neither mentioned nor illustrated. The original material (BAPb 244 and slides) was revised, and although this is not as finely preserved as the new material, the thickenings of the ray cell walls were observed (figures 2.G, 2H). Locally biseriate rays were mentioned for the holotype and not observed in the new material. As a result, a new combination to the genus Abietopitys is proposed because of the thickenings that the genus Phyllocladopitys lack. In addition, original and new specimens show pitting of the tracheids exclusively in the radial walls (figure 2.I).
Conclusions
The Abietopitys petriellae woods here described have primitive gymnospermous characters, such as narrow rays, alternate pitting and crowded continuous bordered pits that completely fill the radial wall. These features are already present in some Devonian fossil woods (Beck, 1970).
The genus Abietopitys is characterized by the peculiar thickenings in the walls of ray cells. This type of thickenings is unique in the Paleozoic woods, although some woods referred to the genus Callixylon have ray tracheids with secondary thickenings that slightly resemble these thickenings. The Abietopitys genus was originally recognized for the Permian Ecca Group of South Africa (Kräusel, in Kräusel and Range, 1928; 1956), and then two fragments of secondary xylem from the Permian of Chubut province, Argentina, were assigned to the genus by Archangelsky (1960). Therefore, based on the present results the range of the genus is extended to the Upper Carboniferous in sediments from Argentina.
The leaf traces observed, only two in a new specimen, and three in the holotype, are insufficient to make a mapping. Unfortunately, in the other transverse sections, no leaf traces were found.
Because fossil woods are not usually found in organic connection with other parts such as leaves or reproductive organs, the botanical affinity of most Paleozoic woods, such as those described in this paper, is difficult to determine. However, it was possible to compare with the plants of known affinity present in the same beds. Among them, Cordaites and Pteridospermales are recognized.
The axes are similar in the pitting, among other features, to woods of Cordaites, although the pith is not septated, as in most northern Cordaites. Much less is known about the southern Cordaites, and an affinity to this group cannot be discarded. It is also different from other typical pteridosperm stems in having only one stele. In addition, they could also be related to primitive pteridosperms with one stele like Eristophyton Zalessky, (see Galtier and Scott, 1990, for a description of Eristophyton and comparisons with other Paleozoic gymnosperms).
In any case, the presence of the same association of leaves and fertile structures with woods in different localities should be taken into account to infer the probable affinity of the woods.
Acknowledgements
The author thanks S.N. Césari and P.R. Gutiérrez for collecting the new material and S.N. Césari and J. Garcia Massini for reading the manuscript. He is also grateful to J. Galtier and B. Meyer-Berthaud for their valuable comments as reviewers. The funding for this work was provided by the Agencia Nacional de Promoción Científica y Tecnológica (PICT 0482).
References
Amos, A.J. 1954. Estructuras de las formaciones paleozoicas de La Rinconada, pie oriental de la Sierra Chica de Zonda (San Juan). Revista de la Asociación Geológica Argentina 9: 5-38. [ Links ]
Archangelsky, S. 1960. Estudio anatómico de 2 especies del género Abietopitys Kräusel, procedentes de la serie Nueva Lubecka prov. Chubut, Argentina. Acta Geológica Lilloana 3: 331-342. [ Links ]
Archangelsky, S., Azcuy, C.L., González, C.R. and Sabattini, N. 1987. Edad de las biozonas. In: S. Archangelsky (ed.), El Sistema Carbonífero en la República Argentina , Academia Nacional de Ciencias (Córdoba), pp. 293-301. [ Links ]
Bangert, B. and Bamford, M.K. 2001. Carboniferous pycnoxylic woods from the Dwyka group of southern Namibia. Palaeontologia Africana 37: 13-23. [ Links ]
Beck, C.B. 1970. The appearence of gymnospermous structure. Biological Reviews 45: 379-400. [ Links ]
Bracaccini, O. 1946. Contribución al conocimiento de la Precordillera sanjuanino-mendocina. 4. Faldeo oriental de la Sierra Chica de Zonda. Boletín de Informaciones Petroleras 261: 361-384. [ Links ]
Brea, M. and Césari, S.N. 1995. An anatomically preserved stem from the Carboniferous of Gondwana: Phyllocladopitys petriellae Brea and Césari, sp. nov. Review of Palaeobotany and Palynology 86: 315-323. [ Links ]
Césari, S.N. and Bercowski, F. 1997. Palinología de la Formación Jejenes (Carbonífero) en la quebrada de Las Lajas, provincia de San Juan, Argentina. Nuevas evidencias paleoambientales. Ameghiniana 34: 497-509. [ Links ]
Césari, S.N., Archangelsky, S. and Vega, J.C. 2005. Anatomy of a new probable pteridosperm stem from the Late Carboniferous of Argentina. Revista del Museo Argentino de Ciencias Naturales, nueva serie 7: 7-15. [ Links ]
Chitaley, S. and Cai, C. 2001. Permineralized Callixylon woods from the late Devonian Claveland Shale of Ohio, USA. And that of Kettle Point, Ontario, Canada. Review of Palaeobotany and Palynology 114: 127-144. [ Links ]
Cladera, G., Archangelsky, S. and Vega, J.C. 2000. Precisiones geográficas, estratigráficas y paleoambientales sobre los niveles portadores de cúpulas pteridospérmicas de la Formación Jejenes, Carbonífero de San Juan, Argentina. Ameghiniana 37: 213-219. [ Links ]
Crisafulli, A., Lutz, A. and Melchor, R. 2000. Maderas gimnospérmicas de la Formación Carapacha (Pérmico), provincia de La Pampa. Ameghiniana 37: 181-191. [ Links ]
Galtier, J. and Scott, A.C. 1990. On Eristophyton and other gymnosperms from the Lower Carboniferous of Castelton bay, east Lothian, Scotland. Geobios 23: 5-19. [ Links ]
Greguss, P. 1955. Identification of living Gymnosperms on the basis of xylotomy. Akadémiai Kiamó. Budapest, 263 pp. [ Links ]
Gutiérrez, P.R. and Césari, S.N. 1987. Nuevos elementos microflorísticos de la Formación Jejenes (Carbónico) provincia de San Juan. Asociación Geológica Argentina, Serie A, Monografías y Reuniones 2 (1986): 168-173. [ Links ]
Herbst, R. and Crisafulli, A. 1997. Kaokoxylon zalesshyi (Sahni) Maheshwari (Coniferopsida), en el Pérmico Superior del Cerro Colorado de la Antigua, La Rioja, Argentina. Ameghiniana 34: 447-451. [ Links ]
Kneller, B., Milana, J.P., Buckee, C. and Al Ja´aidi, O. 2004. A depositional record of deglaciation in a paleofjord (Late Carboniferous [Pennsylvanian] of San Juan Province, Argentina): The role of catastrophic sedimentation. Bulletin of the Geological Society of America 116: 348-367. [ Links ]
Kräusel, R. 1956. Hölzer aus dem südlichen Gebiet der Karru-Schichten Südwest-Afrika. Senckenbergiana Lethaia 37: 447-453. [ Links ]
Kräusel, R. and Range, P. 1928. Beiträge zur Kenntnis der Karruformation Deutsch-Südwest Afrikas. Beiträge zur Geologie Erforschung der Deutschen Schutzgebiete 20: 1-54. [ Links ]
Lepekhina, V.G. 1972. Woods of Palaeozoic pycnoxylic gymnosperms with special reference to north Eurasia representatives. Palaeontographica B 138: 44-106. [ Links ]
Mésigos, M. 1953. El Paleozoico superior de Barreal y su continuación austral, Sierra de Barreal, provincia de San Juan. Revista de la Asociación Geológica Argentina 8: 65-109. [ Links ]
Petriella, B. 1982. Medullopitys menendezii n. sp., leño picnoxílico de gimnospermas del Paleozoico Superior de Mendoza, Argentina. Ameghiniana 19: 253-257. [ Links ]
Pujana, R.R. 2003. Nuevos hallazgos de leños con anatomía preservada en el Carbonífero Superior de la provincia de San Juan, Argentina. Revista del Museo Argentino de Ciencias Naturales, nueva serie 5: 201-204. [ Links ]
Richter, H.G., Grosser, D., Heinz, I. and Gasson, P.E. (eds.). 2004. International Association of Wood Anatomists List of microscopic features for softwood identification. Journal of the International Association of Wood Anatomists 25: 1-70. [ Links ]
Vega, J.C. 1995. La flora fósil de la Formación Jejenes (Carbonífero). Implicancias paleoclimáticas y paleogeográficas. Ameghiniana 32: 31-40. [ Links ]
Vega, J.C. and Archangelsky, S. 2001. Austrocalyxaceae, a new pteridosperm family from Gondwana. Palaeontographica B 257: 1-16. [ Links ]
Recibido: 28 de julio de 2004.
Aceptado: 28 de diciembre de 2004.

