










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.43 no.1 Buenos Aires Jan./Mar. 2006
Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans
Sergio F. Vizcaíno 1 , M. Susana Bargo 1 and Guillermo H. Cassini 2
1 Departamento Científico Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n, 1900 La Plata, Argentina. CONICET-CIC. vizcaino@museo.fcnym.unlp.edu.ar, msbargo@museo.fcnym.unlp.edu.ar
2 Departamento de Ciencias Básicas, Universidad Nacional de Luján, Rutas 5 y 7, 6700 Luján, Argentina. ghnanca@yahoo.com.ar
Abstract. The Xenarthra includes the most intriguing mammals from the Cenozoic of South America: the glyptodonts (Cingulata) and the ground sloths (Tardigrada). Their masticatory apparatuses are diverse and peculiar, with a strongly reduced, hypselodont dentition that lacks enamel and displays different degrees of lobation. The goal of this study is to investigate the relationship between dental occlusal surface area (OSA) and diet, and other physiological factors in fossil xenarthrans. Over one hundred and fifty specimens, including living herbivorous epitherians and both extinct and living xenarthrans, were measured and photographed, and their OSA estimated and plotted against body mass. For most fossil xenarthrans OSA is smaller than expected for extant herbivorous mammals of equivalent body size. Within xenarthrans, cingulates show the highest OSA values, suggesting more extensive oral food processing than in tardigrades. Among ground sloths, mylodontids have extremely low OSA values, suggesting low efficiency in oral food processing that was probably compensated by high fermentation in the digestive tract, or lower metabolic requirements, or a combination of both adaptations. On the other hand, Megatherium americanum has an OSA expected for, or even higher than that of, a mammal of its size, which indicates higher oral food processing, lower fermentation capacity, and/or higher metabolic requirements.
Resumen. Área de la superficie oclusal dentaria en relación con la masa corporal, hábitos alimenticios y otros rasgos biológicos en Xenartros fósiles. El grupo de los Xenarthra incluye los mamíferos más intrigantes del Cenozoico de América del Sur: los gliptodontes (Cingulata) y los perezosos terrestres (Tardigrada). Sus aparatos masticatorios son diversos y peculiares, con una dentición fuertemente reducida, hipselodonte, que carece de esmalte y presenta distintos grados de lobulación. El objetivo de este trabajo es investigar la relación que existe entre el área de la superficie oclusal dentaria (OSA), la dieta y otros factores fisiológicos en xenartros fósiles. Se midieron y fotografiaron más de ciento cincuenta especímenes, que incluyen epiterios herbívoros actuales y xenartros vivientes y fósiles; se estimaron sus OSAs y graficaron contra la masa corporal. Para la mayoría de los xenartros fósiles la OSA es menor que la esperada para mamíferos herbívoros vivientes de tamaño corporal equivalente. Dentro de los xenartros, los cingulados arrojan los valores mayores de OSA, lo que sugiere un procesamiento del alimento en la cavidad oral más extensivo que en los tardigrados. Entre los perezosos terrestres, los milodóntidos presentan valores de OSA extremadamente bajos, lo que indica una baja eficiencia en el procesamiento oral del alimento, que podría compensarse con una alta fermentación en el tracto digestivo o con requerimientos metabólicos menores, o una combinación de ambas adaptaciones. Por otra parte, Megatherium americanum posee la OSA esperada para un mamífero de tamaño equivalente, o inclusive mayor, lo que sugiere un mayor procesamiento oral del alimento, menor capacidad de fermentación y/o mayores requerimientos metabólicos.
Key words. Mammalia; Xenarthra; Dental Occlusal Surface Area; Body mass; Diet; Physiology.
Palabras clave. Mammalia; Xenarthra; Área de la Superficie Oclusal Dentaria; Masa corporal; Dieta; Fisiología.
Introduction
The Order Xenarthra represents one of the four major clades of placental mammals (Murphy et al ., 2001; Madsen et al ., 2001; Delsuc et al ., 2003). The other three, Afroteria, Euarchontoglires and Laurasitheria, were previously believed to be a single clade, Epitheria (McKenna, 1975). Within xenathrans, glyptodonts and ground sloths are, without doubt, the most intriguing South American Cenozoic mammals. The former (Suborder Cingulata) are armoured animals related to the living armadillos while the latter (Suborder Tardigrada) are related to the living tree sloths. The masticatory apparatuses of cingulates and tardigrades are truly diverse and peculiar. The dentition is strongly reduced: armadillos and glyptodonts usually have no incisors or canines and nine or ten cheek teeth in each quadrant, while sloths usually have four or five. All living xenarthrans, and possibly all extinct ones, lack enamel in the adult, deciduous dentitions except Dasypus Linné and the cuspal pattern observed in other mammals. Teeth are always hypselodont ( i.e ., high-crowned and evergrowing) and, although they can be lobate, are usually simple and separated by short spaces (figure 1).

Figure 1. Upper left tooth series of different fossil xenarthrans / series dentarias izquierdas superiores de distintos xenartros fósiles. A, Eutatus seguini (Dasypodidae), B, Holmesina occidentalis (Pampatheriidae), C, Panochthus tuberculatus (Glyptodontidae), D, Glossotherium robustum (Mylodontidae), E, Megatherium americanum (Megatheriidae). Scale bar = 5 cm / Escala = 5 cm.
Morpho-functional analyses of the masticatory apparatus of fossil xenarthrans, including biomechanical and morpho-geometric methods, have been performed recently as a basis for the interpretation of their dietary adaptations. These studies include dasypodid armadillos (Vizcaíno and Fariña, 1997; Vizcaíno and Bargo, 1998; Vizcaíno and De Iuliis, 2003), pampatheriid armadillos (Vizcaíno et al ., 1998; De Iuliis et al ., 2000), glyptodonts (Fariña, 1985,1988; Fariña and Vizcaíno, 2001) and ground sloths (Bargo, 2001a and 2001b).
At the same time, much research has focused on relationships between aspects of mammalian dental morphology ( i.e ., size, shape and wear) and trophic characterizations. Understanding these relationships allows inferences on feeding adaptations from the teeth of fossil forms (Ungar and Williamson, 2000). Most of this research has been carried out in extant and extinct herbivorous ungulates (Fortelius, 1985; Janis, 1988, 1990, 1995; Spencer, 1995; Fortelius and Solounias, 2000; Pérez-Barbería and Gordon, 1998, 2001; Williams and Kay, 2001; Mendoza et al ., 2002) and primates (see Kay and Ungar, 1997 and references therein).
Although the xenarthran dental peculiarities mentioned above reflect some phylogenetic constraints (Vizcaíno and De Iuliis, 2003) that should be considered (see Discussion), it is possible to quantify and compare some of these variables within xenarthrans, using the “form-function correlation approach" noted by Radinsky (1987). This approach assumes that a close relation exists between form and function, so that the latter can be predicted from the former. Such investigations have benefited greatly when combined with knowledge of closely related taxa. In the absence of suitable homologies, the tendency has been to argue for function based on analogy, usually biological. Moreover, when biological analogues are not available, mechanical ones have often been used (see Plotnick and Baumiller, 2000, for an extensive discussion on this matter).
Janis (1995) demonstrated that some craniodental variables allow discrimination among ungulates of grazing, browsing and mixed feeding habits (but see, Pérez-Barbería and Gordon, 2001 for an alternative interpretation with a different methodological approach). Several authors (Janis, 1990, 1995; Janis and Constable, 1993; Mendoza et al ., 2002) have noted that the cheek-teeth occlusal surface area (OSA) is larger in grazers than in browsers among ungulates and kangaroos. For instance, Janis (1988, 1995) stated that monogastric ungulates, such as perissodactyls, have longer molarized premolar rows than ruminant artiodactyls. Janis and Constable (1993) and Janis (1995) proposed that these differences are due to different alimentary strategies in relation with differences in the physiology of digestion. Horses in comparison with cows (ruminants) spend more time chewing and they also chew more when they eat food with a high fiber content.
The purpose of this paper is to investigate the relationships among cheek-teeth ( i.e ., molariforms) occlusal surface area, body mass, inferred diet, and other biological factors in different fossil xenarthrans. These results provide insightful information that, together with the above-mentioned studies, will allow the development of reasonable interpretations of their dietary behaviour.
Material and methods
For the purpose of this paper, more than one hundred and fifty specimens from collections housed in different museums were measured (see Appendix 1). The data base includes forty-seven species of living, mostly herbivorous, epitherian mammals of very different sizes of the orders Rodentia, Hyracoidea, Tubulidentata, Proboscidea, Artiodactyla, and Perissodactyla, and twenty-four species of Xenarthra, including ten fossil and four living Cingulata and seven fossil and three living Tardigrada.
The fossil tardigrades and cingulates analysed include most of the Pleistocene species that have been previously studied (see above). As these were specialized forms of great size (from approximately 800 to 5000 kg; Fariña et al ., 1998; Bargo et al ., 2000), several older species (Miocene and Pliocene in age) with lower body masses were included to widen the sample. The sloth taxa include Glossotherium robustum (Owen), Lestodon armatus Gervais, Mylodon darwini Owen, and Scelidotherium leptocephalum Owen (Mylodontidae), Hapalops sp. Ameghino and Megatherium americanum Cuvier (Megatherioidea), and Eucholoeops sp. Ameghino (Megalonychidae). The cingulates include Propalaeohoplophorus australis Ameghino, Propalaeohoplophorus incisivus Ameghino, Asterostemma depressa Ameghino, Plohophorus sp. Ameghino, Panochthus tuberculatus Burmeister, Glyptodon sp. Owen (Glyptodontidae), Holmesina occidentalis (Hoffstetter) (Pampatheriidae) and Eutatus seguini Gervais (Dasypodidae).
Usually, cheek-teeth OSA has been measured indirectly by multiplying the width and length of molars and premolars in some ungulates ( e.g ., Janis, 1988, 1995; Pérez-Barbería and Gordon, 1998, 2001), which have nearly quadrangular teeth. This simple method does not consider infolding of the enamel, fossae, cusps and lophs. Hence it would be not suitable for many xenarthrans such as the glyptodonts and mylodontids with their lobated molariforms and the bilophodont molariforms of megatheres which require a different technical approach. Since the relationships between OSA and the three-dimensional surface of the tooth are unknown for different tooth types (Pérez-Barbería and Gordon, 1998), for this paper we will approach this problem considering OSA as the two-dimensional projection of a three-dimensional structure. Digital photos of the occlusal surfaces of the upper cheek tooth rows were taken, including molars and premolars in epitherians and molariforms in xenarthrans. The choice of the upper cheek teeth for this analysis was due to limitations in the availability of material, especially among fossil specimens. The OSA of teeth depends on the individual history or age ( i.e ., old individuals have a greater OSA than juveniles, Pérez-Barbería and Gordon, 1998). In order to avoid these differences, all specimens included in this analysis were adults. Therefore, adults with highly worn teeth ( i.e ., little or no enamel left on the occlusal surface) and juveniles ( i.e. , animals in which the last molar is not erupted) were excluded. These considerations are irrelevant for xenarthrans, since they lack enamel and have ever-growing teeth, so that OSA does not vary much with age in adults.
The outlines of the teeth were digitized in palatal view using WinDIG 2.5 software ( http://www.unige.ch- /cpb/windig.html ). For each individual a set of xi , yi coordinate pairs describing the surface contour of each tooth was collected, and the area enclosed by these points was calculated by Simpson's numerical integer approximation. This method is frequently used in continental aquatic ecology to calculate the surface area of irregular lakes or lagoons (Dangavs, 1995). OSA is the sum of the area of each tooth in the series.
The OSA values obtained were plotted against body mass. For living species, when body mass was not available in museum records it was estimated from 3 to 5 cranial and/or postcranial allometric equations for each specimen, depending on the completeness of the material, and excluding dental measurements (Janis, 1990; Biknevicius et al ., 1993). The values obtained were reasonably within the range for adult specimens reported in the literature (Redford and Eisenberg, 1989; Nowak, 1991). Given the strikingly peculiar design of the skulls, jaws, teeth and limbs of fossil xenarthrans, and the absence of proper analogues among living members of the clade, body mass estimations should be approached cautiously (see Vizcaíno and De Iuliis, 2003, for an extensive discussion on the phylogenetic constraints of the group). In this study, body masses were obtained from the best estimators taken from Fariña et al . (1998) and Bargo et al . (2000). In these articles the mass of fossil xenarthrans were estimated from regression equations adjusted in modern ungulates with craniodental and postcranial variables, and the averages obtained were contrasted with estimates obtained from scale and geometric models. In the few cases for which the available skeletal elements were insufficiently complete, geometric similarity with a phylogenetically close relative with an appropriate estimation was assumed.
The two variables (OSA and body mass) were log-transformed to convert the relationship of these variables from an exponential function (Y=aWb) to a straight linear function (LogY=log a + b logW) and reduce heteroscedasticity ( i.e ., the dispersion associated with high values) of the data (see Peters, 1983).
Regression lines, with log OSA taken as dependent variable and log body mass as independent variable, were calculated by least squares method independently for all extant mammals, living epitherians, and living and extinct xenarthrans. Additionally, they were also calculated for various subgroups within epitherians and xenarthrans, as follows:
1. Rodents (South American hystricomorphs), 8 species.
2. Perissodactyls plus hyracoids (Families Equidae, Tapiridae, Rhinocerotidae and Procaviidae), 17 species.
3. Artiodactyls (Families Antilocapridae, Cervidae, Bovidae, Giraffidae, Camelidae and Hippopotamidae), 18 species.
4. Ruminant artiodactyls (Families Antilocapridae, Cervidae, Bovidae), 12 species.
5. Cingulates (Families Dasypodidae, Glyptodontidae and Pampatheriidae), 14 species.
6. Dasypodids, 5 species.
7. Glyptodontids, 8 species.
8. Tardigrades (Families Bradypodidae, Megatheriidae and Mylodontidae), 10 species.
Perissodactyls and hyracoids are lumped together, following Janis (1990), based on the convergence of dental morphology and digestive physiology in the two groups. All regression slopes were tested for isometry ( i.e ., if the calculated slope differs significantly from the expected value of 0.67) and for the expected scaling of metabolic requirements with mass ( i.e ., 0.75, as described by Kleiber´s law) by Student's t statistic (two-tailed test). Following Bell (1989), differences among groups, that is Epitherians vs. Xenarthrans and within xenarthran taxa, were studied by means of analyses of covariance (ANCOVA), using the log-transformed body mass as a covariable. The distributions around the regression line of different physiological or taxonomic groups were studied by means of a residual analysis. The residuals of each group were compared by means of the Mann-Whitney U test, which is the most powerful (or sensitive) non-parametric alternative to the t-test for independent samples. All statistical analyses were performed using the program Statistica (StatSoft, 1996).
Results
Appendix 1 lists the body mass and the OSA for each specimen studied, and Appendix 2 lists the statistics of both features for each species. Note that when compared with epitherians of similar body size (for instance living xenarthrans against hyracoids and rodents, or mylodontid fossil sloths against rhinos, hippos and elephants), xenarthrans have smaller OSA. Also, it can be seen that within the largest xenarthrans, ground sloths have smaller OSA values than the cingulates.
ANCOVA results show that the three pairs of comparisons (epitherians vs xenarthrans, perissodactyls plus hyracoids vs ruminants, and cingulates vs tardigrades) do not differ significantly (p = 0.05) in their adjusted values for slopes (table 1). In the first case, the OSA values of xenarthrans are significantly smaller than in epitherians of similar body mass (p < 0.0001); in the second, the ruminants have significantly smaller OSA values than perissodactyls of similar body mass (p < 0.0001); and in the last comparison, the OSA values of tardigrades are significantly smaller than in cingulates of similar body mass (p < 0.001).
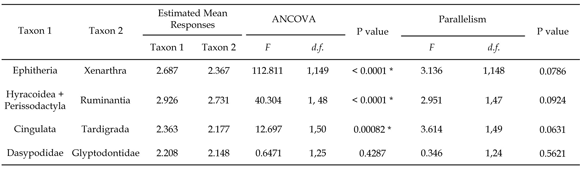
Table 1. Results of ANCOVA for comparisons between taxa listed of log-transformed data of occlusal surface area, using the log-transformed body mass as covariable. Assumptions violating test of parallelism are also presented. Significant differences between groups are indicated by an asterisk / análisis de covarianza de la comparación entre los taxones listados de los datos de área de superficie oclusal transformados a logaritmo usando la masa corporal transformada a logaritmo como covariable. También se presentan los resultados del test de paralelismo. Las diferencias significativas entre los grupos están indicadas por un asterisco.
Figure 2 shows the relationships between OSA and body mass for living mammals. Specimens belonging to different orders and/or families are identified by different symbols. This regression clearly shows that living xenarthrans that fall below the adjusted line have smaller OSA values than those expected for mammals of their size. In addition, it is clear how the different groups of taxa tend to cluster above or below the line: most rodents, hyracoids, proboscideans, perissodactyls (especially horses) fall above the regression line, while most artiodactyls (especially ruminants) fall below the regression line.
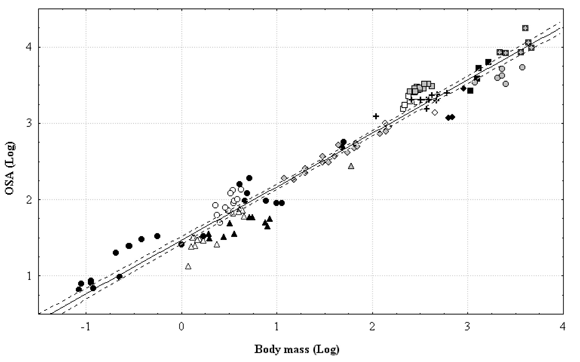
Figure 2. Regression of OSA against body mass for living mammals (n = 125). Symbols: open triangles = Dasypodidae; black triangles = Bradypodidae; gray triangle = Tubulidentata; black circles = Caviomorpha; open circles = Hyracoidea; open squares = Tapiridae; gray squares = Equidae; black squares = Rhinocerotidae; gray diamonds = Cervidae; open diamonds = Bovidae; black diamonds = Giraffidae; gray circle = Hippopotamidae; crosses = Camelidae; gray cross within square = Elephantidae. Dashed lines above and below the regression line = 95 % confidence interval / Regresión de las áreas de las superficies oclusales dentarias (OSA) contra la masa corporal en mamíferos vivientes (n = 125). Símbolos: triángulos en blanco = Dasypodidae; triángulos negros = Bradypodidae; triángulos grises = Tubulidentata; círculos negros = Caviomorpha; círculos en blanco = Hyracoidea; cuadrados en blanco = Tapiridae; cuadrados grises = Equidae; cuadrados negros = Rhinocerotidae; rombos grises = Cervidae; rombos en blanco = Bovidae; rombos negros = Giraffidae; círculos grises = Hippopotamidae; cruces = Camelidae; cruces grises dentro de un cuadrado = Elephantidae. Líneas punteadas por encima y por debajo de la línea de regresión = intervalo de confianza del 95%.
Figure 3 compares independent regressions of epitherians and all xenarthrans (including fossils). Both regression lines seem to be parallel and the slopes do not differ significantly (p=0.05; see table 1). The taxa distributions of epitherians around the regression line do not show marked differences with the distribution observed for all living mammals regression in figure 2. The regression line of epitherians lies above that of xenarthrans. Among the latter, there are two taxa from different clades that are outliers: the pampatheriid cingulate Holmesina occidentalis and the giant ground sloth Megatherium americanum . Both lie well above the xenarthran regression line, falling on the line of epitherians. It is also remarkable that the mylodontid ground sloths fall well below the regression line for xenarthrans.

Figure 3 . Regression of OSA against body mass of Xenarthra (n = 53) compared with Epitheria (n = 99). Symbols: circles = Epitheria; black triangles = Xenarthra; E = Epitheria regression line; X = Xenarthra regression line. Dashed lines above and below the regression line = 95 % confidence interval. The black triangles contained by the ellipse correspond to the mylodontid ground sloths. a. case of the Pampatheriidae Holmesina . b. two cases of Megatherium ./ regresión de las áreas de las superficies oclusales dentarias (OSA) contra la masa corporal de Xenarthra (n = 53) comparado con Epitheria (n = 99). Símbolos: círculos = Epitheria; triángulos negros = Xenarthra; E = línea de regresión de Epitheria; X = línea de regresión de Xenarthra. Líneas punteadas por encima y por debajo de la línea de regresión = intervalo de confianza del 95 %. Los triángulos negros dentro de la elipse corresponden a los milodóntidos. a. caso del Pampatheriidae Holmesina . b. dos casos de Megatherium .
Figure 4 includes independent regressions for cingulates and tardigrades. The regression of cingulates lies above that of tardigrades, and has a higher, although not statistically different (p = 0.05), slope (table 1). The tardigrade regression clearly shows a differential distribution of some groups, which parallels that of figure 3. Mylodontids fall below the line, while M. americanum lies far above the regression line, being an outlier due to a great value of OSA (and not to a measuring error). When regression parameters are recalculated without M. americanum , the values adjusted for the slope become smaller ( i.e ., 0.52), compared to the value provided in table 2 for tardigrades (see below). The living sloths overlap both regression lines and do not show any particular distribution. In contrast, the cingulates are distributed tightly around the regression line, with the exception of H. occidentalis , which lies well above the line.

Figure 4. Regression of OSA against body mass of xenarthrans (n = 53). Symbols: C = Cingulata; T = Tardigrada; black triangles = dasypodid cingulates; gray triangle = pampatheriid cingulate; open triangles = glyptodontid cingulates; open circles = living tardigrades; gray circle = Eucholoeops ; open squares = Hapalops ; black squares = Megatherium ; black circles = mylodontids. Dashed lines above and below the regression line = 95% confidence interval / regresión de las áreas de las superficies oclusales dentarias (OSA) contra la masa corporal de Xenarthra (n = 53). Símbolos: C = Cingulata; T = Tardigrada; triángulos negros = cingulados dasipódidos; triángulos grises = cingulados pampateridos; triángulos en blanco = cingulados gliptodóntidos; círculos en blanco = tardigrados vivientes; círculos grises = Eucholoeops ; cuadrados en blanco = Hapalops ; cuadrados negros = Megatherium ; círculos negros = mylodontids. Líneas punteadas por encima y por debajo de la línea de regresión = intervalo de confianza del 95%.
Table 2. Results of simple linear regression for each group. The more common parameters, R-square value and estimators of regression coefficients in bold. Differences between the values adjusted for the slopes and that expected for isometry (2/3) were not significant in all groups at p-level of 0.05 except for that tagged by an asterisk (p = 0.01). Differences between the values adjusted for the slopes and that expected for Kleiber´s law (0.75) were not significant only in groups tagged by an asterisk (p = 0.01) or two asterisks (p = 0.05). Residual analysis results are present when a pair of comparisons was statistically significant (p < 0.01). Labels : E = Epitheria, X = Xenarthra, P = Perissodactyla, A = Artiodactyla, C = Cingulata, T = Tardigrada, Eq = Equidae, Tp = Tapiridae / resultados de las regresiones lineares simples de cada grupo. Los parámetros más comunes, valor R-cuadrado y los estimadores de los coeficientes de regresión están marcados en negrita. Las diferencias entre los valores ajustados para las pendientes y aquellos esperados para isometría (2/3) fueron no significativos para todos los grupos (p = 0.05), excepto por aquél marcado con un asterisco (p = 0.01). Las diferencias entre los valores ajustados para las pendientes y aquel esperado para la ley de Kleiber (0.75) fueron no significativos solamente en los grupos marcados con un asterisco (p = 0.01) o dos asteriscos (p = 0.05). Los resultados del análisis de residuos fueron presentados cuando el par de comparaciones fue estadísticamente significativo (p < 0.01). Leyendas: E = Epitheria, X = Xenarthra, P = Perissodactyla, A = Artiodactyla, C = Cingulata, T = Tardigrada, Eq = Equidae, Tp = Tapiridae.
The parameters for the regression lines in figures 2, 3 and 4, and some subgroups, are summarised in table 2. For all cases the R2 values obtained are greater than 0.9, indicating a high correlation between OSA and body mass. According to the t-test, all slopes do not differ significantly from the expected value for isometric relationship ( i.e ., 0.67 for a log relation between area and volume) in each case considered. This indicates that they would be parallel, in correspondence with parallelism test results. In all groups except for the perissodactyls plus hyracoids, dasypodids and glyptodontids (table 2), the t-tests allow us to conclude with 99% confidence, that the values adjusted for the slopes differ from that expected from Kleiber´s law ( i.e ., a 0.75 slope). Only in the three mentioned groups are the values adjusted for the slopes not significantly different from the two null hypothesis models ( i.e ., 0.67 and 0.75). In dasypodids and glyptodontids values for the two models for the slope are included in a wide confidence interval due to the small sample available. The graphic distributions of different groups around the regression line are statistically significant and supported by the residual analyses. In the regression line for living mammals, epitherians have greater OSA values than living xenathrans; in the epitherian regression line perissodactyls have greater OSA values than artiodactyls; for the xenarthran regression line (including living and fossil taxa) the residual analyses, as well as the ANCOVA, indicates that the cingulates have greater OSA values than tardigrades; and for the perissodactyls plus hyracoid regression line, residual analyses shows that equids have greater OSA values than tapirs.
Discussion
When structures differ in size but have the same proportion, they are said to be isometric, and their areas scale with mass to the power 2/3 (or 0.67). In biology isometry usually exists only for limited size ranges within single species, for instance insects and arachnids (Prange, 1977) and lizards (McMahon and Bonner, 1983). For larger ranges, proportions change with size and that change is described as allometric. For our general sample of mammals, which includes a large range of body sizes, OSA increases with mass by the power 0.70, which, according with the Student's t-test results (see Table 2 to review the two null hypothesis), suggests isometry.
This is particularly evident in the slope of Epitheria (0.66). Within ungulates, the artiodactyls (especially the ruminant artiodactyls) are distributed below the regression line with lower OSA values, while the monogastric forms lie above the regression line, which indicates that their OSA values are larger than expected for their body size. This is in accordance with Janis' (1988, 1995) conclusion that the monogastric ungulates, such as the perissodactyls, have longer and more molarized premolar rows than ruminants; and the molars of the former are also bigger and more quadrangular than in the latter. Following Janis and Constable (1993) and Janis (1995), this difference is due to different alimentary strategies reflecting differences in the physiology of digestion. Horses chew their food more than ruminants during the first stages of ingestion and, in contrast to the latter, chew more when the proportion of fiber in the food increases. Within the perissodactyls, the equids, which are mainly grazers and live in open habitats, have greater OSAs than tapirs, which are browsers and live in more closed habitats. Similarly, within artiodactyls, mixed feeding or grazing bovids and camelids have larger OSAs than browsing cervids and giraffids. This suggests that in ungulates chewing area is also related to the nutritional value of food: browsers consume succulent leaves while grazers ingest forage that is of lower quality and highly abrasive for teeth. Additionally, within ruminants Texera (1974) observed that subadult individuals of the cervid Hippocamelus Leuckart spent more time chewing than adults, a behaviour attributed to the smaller OSA of subadults due to unerupted third molars. A remarkable case is the hippopotamus. Its teeth are unusually small for such a large grazing mammal, a condition probably related to its low metabolic rate (Owen-Smith, 1988). The preceding examples indicate that OSA may thus be correlated with the capacity to process food in the oral cavity and, indirectly, with some aspects of feeding physiology and metabolism.
Xenarthrans have less OSA available for triturating food than epitherians of similar sizes. This fact may be related to the low basal metabolic rates characteristic of living xenarthrans, which fall between 40 and 60 percent of the rates expected from mass in Kleiber's (1932) relation for placental mammals (McNab, 1985). This implies that xenarthrans have less energetic requirements than epitherians and, therefore, for a specific type of food, require lower intakes than epitherians of similar body masses.
Within xenarthrans, cingulates have the highest OSA values, suggesting a greater oral cavity food processing than in tardigrades. It is interesting to note that large herbivorous cingulates, such as pampatheres and glyptodonts, have lobated teeth. Feeding behaviour in these forms was postulated as ranging from browsing to grazing (Vizcaíno et al ., 1998; De Iuliis et al ., 2000; Fariña and Vizcaíno, 2001). Tooth lobation is more complex in glyptodonts than in pampatheres, producing more extensive shearing edges for food processing. Moreover, a glyptodont tooth is highly hypselodont and bears a longitudinal osteodentine (compact dentine) crest and, in each lobe, a transverse crest. This morphology, coupled with other morphometric features of their skulls and jaws, led Fariña and Vizcaíno (2001) to place them near the group of grazers in the graphs presented by Janis (1990). In the case of pampatheres, tooth lobation is less complex, but OSA is much larger. It seems probable that, in some way, a reduction in one feature is compensated for by an increase in the other. Holmesina occidentalis , with its high OSA, also lacks the hard osteodentine crest, indicating another aspect to consider with respect to compensation. In general, pampatheres seem to have higher OSAs and fewer ridges and less shearing surfaces. Although there is no direct evidence of the rate of metabolism of pampatheres and glyptodonts, indirect evidence, such as the pattern of geographic distribution in North America, indicates that their northern limit were restricted by the combination of low metabolism and high thermal conductance (McNab, 1985). Also, there may be a relationship between the OSA and stomach morphology that may help to explain why cingulates fall above the regression line compared to tardigrades. Modern armadillos have a simple sac-like stomach (Grassé, 1955) compared to living sloths with a chambered stomach (Britton, 1941) so presumably do not have the same level of digestive efficiency. Cingulates like glyptodonts and pampatheres probably also had a simple sac-like stomach that lacked chambers and therefore would need a more complex dental apparatus, i.e., lobate teeth and greater OSA to better process vegetation than sloths. Among sloths, the extremely low OSA values for mylodontids might reflect poor food oral processing abilities. If this were the case these ground sloths, in order to maintain diets similar to those of the ungulates of equal body masses, would have been expected to compensate for the low efficiency in food processing with high fermentation capability in the digestive tract, and/or lower metabolic requirements. The living tree sloths, Bradypus Linné and Choloepus Illiger, have an extremely large four-chambered stomach and so presumably this is also true for all fossil sloths. Food passage through the digestive tract is very slow (up to one week) and, consequently, the digestion and absorption rhythm is also extremely slow (Britton, 1941). Moreover, the generally slow nature of tree sloths, due to the poor skeletal musculature, also implies lower metabolic requirements than in other mammals (Scholander, 1955; McNab, 1985). Naples (1989) assumed that Glossotherium also had a slow rate of passage of food through the gut, which might have enabled this sloth to obtain sufficient nutrition from high fiber foods. If the presumed chambered stomach of mylodontids was a functional equivalent to the chambered stomach of ruminants, then in both groups digestion in the stomach is an important component that allows a more efficient extraction of nutrients and permits a smaller OSA. Consequently, it could be argued that the smaller OSA in mylodontids indicates increased digestive efficiency and a longer transit time in the gut that permitted a more efficient absorption of nutrients. As a consequence there may not be as strong selection for the oral mechanical processing of vegetation. Small OSA may also in part be a reflection of the low metabolism, as occurs in the hippopotamus, which has a complex three chambered stomach, relatively small teeth, a restricted rate of food intake and a relatively low metabolic rate (Owen-Smith, 1988); and individuals have been known to survive for many weeks in a mud wallow without food, water, or shade (Nowak, 1991).
Surprisingly, Megatherium americanum has an expected, or even higher, OSA value for a mammal of its size, and a much larger OSA value than expected compared with mylodontids, which may represent a high specialization for the former. Following Janis (1995), if a parallel with ungulates is assumed, mylodontids might be inferred as foregut fermenters, while M. americanum would have been a hindgut fermenter, which in turn reflect lower and higher quality nutritional diets, respectively (Alexander, 1996). However, the absence of a caecum in living xenarthrans might indicate that M. americanum was not a hindgut fermenter. In this case the great difference in OSA values between M. americanum and mylodontids might suggest that the former had less developed chambers (in number and/or size) than the latter. While stomach anatomy, except in a few exceptional cases, cannot be compared among fossil taxa, it is clear that M. americanum was better suited for food processing in the oral cavity than were mylodontids. This is in accordance with other morphological and biomechanical evidence discussed extensively by Bargo (2001a). The teeth of M. americanum are extremely hypselodont and bilophodont, with the sagittal section of each loph being triangular and sharp-edged (figure 1). This occlusal morphology indicates that the way OSA has been calculated in this study does not reflect the sum of the total area present on the anterior and posterior surfaces of the two lophs.
From a functional point of view, the morphology described above suggests that the teeth were well adapted for strong, predominantly orthal movements and were used mainly for cutting rather than crushing and grinding (Bargo, 2001a). We do not know yet how much one function is enhanced over the other, or how much one complements the other.
The evidence provided by Bargo (2001a) indicates that M. americanum could have fed on moderate to soft tough food, probably browsing (including fruits) in open habitats, and the possibility that it was capable of processing flesh cannot be excluded (Fariña, 1996; Fariña and Blanco, 1996). Fruits and flesh are food items nutritionally richer than most grasses or leaves; richer diets require smaller fermentation chambers (Alexander, 1996) and strict carnivores do not have chambers at all. Frugivory and carnivory imply that flesh and fruit eaters presumably do not require the same degree of mastication, and then the larger OSA in Megatherium may have been an indirect way of enlarging the cutting area, since cutting does not result in the breakage of cell walls and the release of nutrients as efficiently as crushing and grinding does. Additionally, OSA is assumed to be directly correlated with the quantity of food trapped between the upper and lower tooth rows. Hence, in species with strong jaw muscles which crush their food between sets of interlocking cheek-teeth, as is the case in Megatherium , a large OSA is functionally important (Pérez-Barbería and Gordon, 1998 and references therein), increasing the amount of food fragments cut or ground per chewing cycle.
The great OSA of M. americanum would also suggest a basal metabolic rate similar to that of an extant herbivorous mammal of equal body size. If M. Americanum had a digestive tract as efficient for processing cellulose as mylodonts, it would have been capable of maintaining a higher basal metabolism compared to mylodonts. The ground sloths, in general, would have had low metabolic rates, due to their large body masses (McNab, 1985; Naples, 1989), probably lower than those ungulates of equivalent size. The skeletal musculature, in contrast to living sloths, was well developed, given their totally terrestrial locomotory habits, and some mylodontids ( i.e ., G. robustum and S. leptocephalum ) were well adapted for digging burrows (Bargo et al ., 2000; Vizcaíno et al ., 2001). This adaptation would probably indicate an even lower basal metabolism for these sloths compared with that for Megatherium americanum . McNab (1979, 1985) found that in armadillos low metabolic rates correlate with burrowing habits, probably as a mean to reduce heat storage during this activity, which would be even more important for the large burrowing ground sloths.
Finally, it is worth remembering that the absence in xenarthrans of many dental specializations characteristic of advanced epitherians is most probably due to biomechanical (Mendoza et al ., 2002) and phylogenetic constraints (Vizcaíno and De Iuliis, 2003). Several authors (Winge, 1941; Hirschfeld and Webb, 1968; Patterson and Pascual, 1972; Hirschfeld, 1976; Webb, 1985; Naples, 1987) have proposed that specializations for insectivory among early xenarthrans imposed a severe constraint in the subsequent adaptations to different diets among the various clades. From their earliest records, xenarthrans are known to have possessed a homodont, hypselodont dentition that lacked enamel, which would be a singularly derived condition for epitherians. Given the early loss of enamel and the typical tribosphenic molar, we should not expect xenarthrans to have evolved morphological responses convergent on those of many epitherians. Specific biomechanical and morphofunctional analyses provide more accurate reconstructions of these unusual extinct creatures than straightforward, traditional comparisons with modern closely allied taxa or supposed analogs, which alone can lead to misleading conclusions (Vizcaíno and De Iuliis, 2003).
Acknowledgments
The authors would like to thank F. Momo, P. Palmqvist, G. De Iuliis and G. McDonald for their comments and valuable input on the manuscript. To J. Cuisin (Laboratoire Zoologie Mammifères et Oiseaux) and F. Renourt (Laboratoire d'Anatomie Comparée) of the Muséum National d'Histoire Naturelle, Paris, A. Kramarz of the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia", Buenos Aires, and M. Merino of the Mammalogy Collection of Museo de La Plata, Argentina, for allowing access to specimens in their care. This paper is a contribution to the projects Agencia Nacional de Promoción Científica y Tecnológica PICT 07-06348, Universidad Nacional de La Plata N-336.
Appendix 1. (Table3). Acronyms and list of the material used with their catalogue numbers, the estimated body masses in kg and the oclussal surface area (OSA) in mm2 of each specimen. / Acrónimos y lista de materiales utilizados con sus números de catálogo, masas corporales estimadas en Kg y superficies oclusales dentarias (OSA) en mm 2 de cada espécimen .

Appendix 2. (Table 4). Figures (means and standard deviations) of dental occlusal surface area (OSA) in mm2 and estimated body masses (in kg) for each species studied / valores (promedios y desvíos standard) de las áreas de las superficies occlusales dentarias (OSA) en mm2 y masas corporales estimadas (en kg) para cada especie estudiada.

References
Alexander, R.McN. 1996. Optima for animals. Princeton University Press, New Jersey, 169 pp. [ Links ]
Bargo, M.S. 2001a. The ground sloth Megatherium americanum : skull shape, bite forces, and diet. In: S.F. Vizcaíno, R.A. Fariña and C. Janis (eds.), Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica , Special Issue 46: 41-60. [ Links ]
Bargo, M.S. 2001b. [ El aparato masticatorio de los perezosos terrestres (Xenarthra, Tardigrada) del Pleistoceno de la Argentina. Morfometría y biomecánica. Doctoral Thesis, Universidad Nacional de La Plata, La Plata, Argentina 400 pp., Unpublished]. [ Links ]
Bargo, M.S., Vizcaíno, S.F., Archuby, F.M. and Blanco, R.E. 2000. Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20: 601-610. [ Links ]
Bell, G. 1989. A comparative method. American Naturalist 133: 553-571. [ Links ]
Biknevicius, A.R., McFarlane, D.A. and MacPhee, R.D. 1993. Body size in Amblyrhiza inundata (Rodentia: Caviomorpha), an extinct megafaunal rodent from the Anguilla Bank, West Indies: Estimates and implications. American Museum Novitates 3079: 1-25. [ Links ]
Britton, W.S. 1941. Form and function in the sloth. The Quarterly Review of Biology 16: 190-207. [ Links ]
Dangavs, N.V. 1995. Morfometría de cuerpos lénticos. In: E.C. Lopretto y G. Tell (eds.), Ecosistemas de aguas continentales. Metodologías para su estudio . Ediciones Sur, La Plata, pp. 27-45. [ Links ]
De Iuliis, G., Bargo, M.S. and Vizcaíno, S.F. 2000. Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution. Journal of Vertebrate Paleontology 20: 743- 754. [ Links ]
Delsuc, F., Vizcaíno, S.F. and Douzery, E. 2004. Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. Evolutionary Biology 4: 1-13. [ Links ]
Fariña, R.A. 1985. Some functional aspects of mastication in Glyptodontidae (Mammalia). Fortschritte der Zoologie 30: 277-280. [ Links ]
Fariña, R.A. 1988. Observaciones adicionales sobre la biomecánica masticatoria en Glyptodontidae (Mammalia, Edentata). Boletín de la Sociedad Zoológica, Montevideo 4: 5-9. [ Links ]
Fariña, R.A. 1996. Trophic relationships among Lujanian mammals. Evolutionary Theory 11: 125-134. [ Links ]
Fariña, R.A. and Blanco, R.E. 1996. Megatherium , the stabber. Proceedings of the Royal Society B 263: 1725-1729. [ Links ]
Fariña, R.A. and Vizcaíno, S.F. 2001. Carved teeth and strange jaws: How glyptodonts masticated. In: S.F. Vizcaíno, R.A. Fariña and C. Janis (eds.), Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica, Special Issue 46: 87-102. [ Links ]
Fariña, R.A, Vizcaíno, S.F. and Bargo, M.S. 1998. Body size estimations in Lujanian (Late Pleistocene-Early Holocene of South America) mammal megafauna. Mastozoología Neotropical 5: 87-108. [ Links ]
Fortelius, M. 1985. Ungulate cheek teeth: developmental, functional, and evolutionary interrelations. Acta Zoologica Fennica 180: 1-76. [ Links ]
Fortelius, M. and Solounias, N. 2000. Functional characterization of ungulate molars using abrasion-attrition wear gradient: a new method for reconstructing paleodiets. American Museum Novitates 3301: 1-36. [ Links ]
Grassé, P. 1955. Traite de Zoologie. Mammiferes. Les Ordres: Anatomie, Éthologie, Systematique. 17: 1174-2300. [ Links ]
Hirschfeld, S.E. 1976. A new fossil anteater (Edentata, Mammalia) from Colombia, S.A., and evolution of the Vermilingua. Journal of Paleontology 50: 419-432. [ Links ]
Hirschfeld, S.E. and Webb, S.D. 1968. Plio-Pleistocene megalonychid sloths of North America. Bulletin of the Florida State Museum 12: 213-296. [ Links ]
Janis, C.M. 1988. An estimation of tooth volume and hypsodonty indices in Ungulate mammals, and the correlation of these factors with dietary preference. In: D.E. Russell, J.P. Santoro and D. Sigogneau-Russell (eds.), Teeth revisited: Proceedings of the VII International Symposium on Dental Morphology. Mémoirs du Muséum national d'Histoire naturelle (série C) 53: 367-387. [ Links ]
Janis, C.M. 1990. Correlation of cranial and dental variables with dietary preferences: a comparison of macropodoid and ungulate mammals. Memoirs of the Queensland Museum 28: 349-366. [ Links ]
Janis, C.M.. 1995. Correlations between craniodental morphology and feeding behavior in ungulates: reciprocal illumination between living and fossil taxa. In: J. Thomason (ed.), Functional Morphology in Vertebrate Palaeontology , Cambridge University Press, pp. 76-98. [ Links ]
Janis, C.M. and Constable, E. 1993. Can ungulate craniodental features determine digestive physiology? Journal of Vertebrate Paleontology 13: 43A. [ Links ]
Kay, R.F. and Ungar, P.S. 1997. Dental evidence for diet in some Miocene Catarrhines with comments of the effects of phylogeny on the interpretation of adaptation. In: D.R. Begun, C. Ward, and M. Rose (eds.), Function, Phylogeny and Fossils: Miocene Hominoid Evolution and Adaptation s, Plenum Press, New York, pp. 131-151. [ Links ]
Kleiber, M. 1932. Body size and metabolism. Hilgardia 6: 315-353. [ Links ]
Madsen, O., Scalli, M., Douady, C.J., Kao, D.J., DeBry, R.W., Adkins, R., Amrine, H.M., Stanhope, M.J., de Jong, W.W. and Springer, M.S. 2001. Parallel adaptative radiations in two major clades of placental mammals. Nature 409: 610-614. [ Links ]
McKenna, M.C. 1975. Toward a phylogenetic classification of the Mammalia. In: W.P. Luckett and P.S. Szalay (eds.), Phylogeny of the Primates , Plenum Publishing Corporation, New York, pp. 21-46. [ Links ]
McMahon, T.A and Bonner, J.T. 1983. On Size and Life. Scientific American Books. W. H. Freeman, New York, 255 pp. [ Links ]
McNab, B.K. 1979. The influence of body size on the energetic and distribution of fossorial and burrowing mammals. Ecology 60: 1010-1021. [ Links ]
McNab, B.K. 1985. Energetics, population biology, and distribution of xenarthrans, living and extinct. In: G.G. Montgomery (ed.), Evolution and ecology of armadillos, sloths and vermilinguas , Smithsonian Institution Press, Washington and London, pp. 219-232. [ Links ]
Mendoza, M., Janis, C.M. and Palmqvist, P. 2002. Characterizing complex craniodental patterns related to feeding behaviour in ungulates: a multivariate approach. Journal of Zoology 258: 223-246. [ Links ]
Murphy, W.J, Eizirik, E, Johnson, W.E., Zhang, Y.P., Ryderk, O.A. and O'Brien, S.J. 2001. Molecular phylogenetics and the origins of placental mammals. Nature 409: 614-618 [ Links ]
Naples, V.L. 1987. Reconstruction of cranial morphology and analysis of function in Nothrotheriops shastensis. Contributions in Science , Los Angeles County Museum of Natural History 389: 1-21. [ Links ]
Naples, V.L. 1989. The feeding mechanism in the Pleistocene ground sloth, Glossotherium. Contributions in Science , Los Angeles County Museum of Natural History 415: 1-23. [ Links ]
Nowak, R.M. 1991. Walker's Mammals of the World. 5th Edition. The Johns Hopkins University Press, Baltimore, 1629 pp. [ Links ]
Owen-Smith, R.N. 1998. Megaherbivores. The influence of very large body size on ecology . Cambridge University Press, Cambridge, 369 pp. [ Links ]
Patterson, B. and Pascual, R. 1972. The fossil mammal fauna of South America. In: A. Keast, F. Erk, and B. Glass (eds.), Evolution, Mammals, and Southern Continents . State University of New York Press, Albany, New York, pp. 247-309. [ Links ]
Pérez-Barbería, F. J. and Gordon, I.J. 1998. Factors affecting food comminution during chewing in ruminants: a review. Biological Journal of the Linnean Society 63: 233-256. [ Links ]
Pérez-Barbería, F.J. and Gordon, I.J. 2001. Relationships between oral morphology and feeding style in the Ungulata: a phylogenetically controlled evaluation. Proceedings of the Royal Society London 268: 1021-1030. [ Links ]
Peters, R.H. 1983. The ecological implications of body size . Cambridge University Press, Cambridge, 324 pp. [ Links ]
Plotnick, R.E. and Baumiller, T.K. 2000. Invention by evolution: functional analysis in paleobiology. In: D.H. Erwin, and S.L. Wing (eds.), Deep Time. Paleobiology's Perspective. Supplement to Paleobiology 26: 305-323. [ Links ]
Prange, H.D. 1977. The scaling and mechanics of arthropods exoskeletons. In: T.J. Pedley, (ed.), S cale effects in animal locomotion, Academic Press, New York, pp. 169- 181. [ Links ]
Radinsdky, L.B. 1987. The evolution of vertebrate design. University of Chicago Press, Chicago, 188 pp. [ Links ]
Redford, K.H. and Eisenberg, J.F. 1989. Mammals of the Neotropics. The Southern Cone . Vol. 2. Chile, Argentina, Uruguay, Paraguay. The University of Chicago Press, 430 pp. [ Links ]
Scholander, P.F. 1955. Evolution of climatic adaptations in homeotherms. Evolution 9: 15-26. [ Links ]
Spencer, L.M. 1995. Morphological correlates of dietary resource partitioning in the African Bovidae. Journal of Mammalogy 76: 448-471. [ Links ]
StatSoft, Inc. 1996. STATISTICA for Windows [ Computer program manual]. Tulsa, OK: StatSoft, Inc., 2300 East 14th Street, Tulsa, OK 74104. ( http://www.statsoftinc.com ). [ Links ]
Texera, W. 1974. Algunos aspectos de la biología del huemul ( Hippocamelus bisulcus ) (Mammalia: Artiodactyla, Cervidae) en cautividad. Anales del Instituto de la Patagonia 5: 155-188. [ Links ]
Ungar, P. and Williamson, M. 2000. Exploring the effects of tooth wear on functional morphology: a preliminary study using dental topographic analysis. Paleontologia Electronica 3: 1-18. [ Links ]
Vizcaíno, S.F. and Bargo, M.S. 1998. The masticatory apparatus of the armadillo Eutatus (Mammalia, Cingulata) and some allied genera. Paleobiology and evolution. Paleobiology 24: 371-383. [ Links ]
Vizcaíno, S.F. and De Iuliis, G. 2003. Evidence for advanced carnivory in fossil armadillos (Mammalia: Xenarthra: Dasypodidae). Paleobiology 29: 123-138. [ Links ]
Vizcaíno, S.F. and Fariña, R.A. 1997. Diet and locomotion of the armadillo Peltephilus : a new view. Lethaia 30: 79-86. [ Links ]
Vizcaíno, S.F., De Iullis, G. and Bargo, M.S. 1998. Skull shape, masticatory apparatus, and diet of Vassallia and Holmesina (Mammalia: Xenarthra: Pampatheriidae). When anatomy constrains destiny. Journal of Mammalian Evolution 5: 293-321. [ Links ]
Vizcaíno, S. F., Zárate, M.A. Bargo, M.S. and Dondas, A. 2001. Pleistocene burrows in the Mar del Plata area (Buenos Aires Province, Argentina) and their probable builders. In: S.F. Vizcaíno, R.A. Fariña, and C. Janis, (eds.), Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica , Special Issue 46: 157-169. [ Links ]
Webb, S.D. 1985. The interrelationships of tree sloths and ground sloths. In: G.G. Montgomery (ed.), Evolution and ecology of armadillos, sloths and vermilinguas , Smithsonian Institution Press, Washington, pp. 105-112. [ Links ]
Williams, S.H. and Kay, R.F. 2001. A comparative test of adaptive explanations for hypsodonty in ungulates and rodents. Journal of Mammalian Evolution 8: 207-229. [ Links ]
WinDIG 2.5. Free Data Digitizer , Copyrigth 1994-1996, D. Lovy. ( http://www.unige.ch/cpb/windig.html ) [ Links ]
Winge, H. 1941. Edentates (Edentata). In: S. Jensen, R. Spärck and H. Volsoe (eds.), The Interrelationships of the Mammalia Genera , Reitzels Forlag, Copenhagen, pp. 319-341. [ Links ]
Recibido: 20 de abril de 2004.
Aceptado: 17 de febrero de 2005.

