










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.1 Buenos Aires ene./mar. 2006
Bioestratigrafía de conodontes y graptolitos silúricos en la sierra de Talacasto, Precordillera de San Juan, Argentina
Guillermo L. Albanesi 1 , Gladys Ortega 1 y Mario A. Hünicken
1 CONICET. Museo de Paleontología, Universidad Nacional de Córdoba, Avenida Vélez Sarsfield 299, Casilla de Correo 1598, 5000 Córdoba. Argentina. galbanes@com.uncor.edu
Resumen. Esta contribución tiene como propósito el estudio taxonómico, bioestratigráfico y paleoambiental de una nueva colección de conodontes y graptolitos silúricos de las formaciones La Chilca y Los Espejos, expuestas en la sección de la quebrada Ancha, sierra de Talacasto, precordillera central de San Juan. Las especies de graptolitos recuperadas de la parte alta de la Formación La Chilca representan a la Serie Llandovery superior (Piso Telychiano). Los conodontes procedentes de coquinas carbonáticas con Saetograptus argentinus (Cuerda), de la parte media-superior de la Formación Los Espejos, permiten determinar la Zona de Kockelella variabilis variabilis del Ludlow inferior (Gorstiano). La secuencia silúrica suprayace en paraconcordancia a la Formación San Juan, cuyas calizas cuspidales produjeron conodontes de la Zona de Lenodus variabilis (Arenig superior - Llanvirn inferior, Ordovícico Medio). La sección estudiada se extiende hasta las primeras lutitas del Devónico Inferior que suprayacen en forma paraconcordante a la secuencia silúrica. La asociación de conodontes registrada en la Formación Los Espejos sugiere una correspondencia con el evento oceánico Linde, entre episodios estables consecutivos. Los conodontes aislados de esta unidad presentan un "CAI" 3, el cual corresponde a paleotemperaturas de soterramiento de entre 110 y 200° C.
Abstract. Conodont-graptolite biostratigraphy of the silurian at Talacasto Range, San Juan Precordillera, Argentina . Present contribution aims to taxonomic, biostratigraphic, and paleoenvironmental studies of new conodont and graptolite collections from the Silurian La Chilca and Los Espejos formations, which crop out in the Ancha Creek, at Talacasto Range, Central Precordillera of San Juan Province. Graptolite species recovered from the upper part of the La Chilca Formation represent the upper Llandovery Series (Telychian Stage). Conodonts from carbonate coquinas with Saetograptus argentinus (Cuerda), of the middle-upper Los Espejos Formation, allow us to determine the Kockelella variabilis variabilis Zone from the lower Ludlow (Gorstian). The Silurian sequence paraconformably overlies the San Juan Formation, whose upper limestones yielded conodonts from the Lenodus variabilis Zone (uppermost Arenig - lowermost Llanvirn, Middle Ordovician). The studied section develops up to the Lower Devonian shales that paraconformably overlies the Silurian sequence. The conodont association recorded in the Los Espejos Formation suggests a correspondence with the Linde Event, which occurred between two consecutive stable episodes. Isolated conodonts from this unit show a CAI 3, which indicates overburden paleotemperatures of 110-200° C.
Key words. Conodonts; Graptolites; Systematics; Biostratigraphy; Events; Silurian; Precordillera; Argentina.
Palabras clave. Conodontes; Graptolitos; Sistemática; Bioestratigrafía; Eventos; Silúrico; Precordillera; Argentina.
Introducción
Las primeras referencias de rocas silúricas en la Precordillera corresponden a los trabajos de Stelzner (1873) y Kayser (1876) realizados en el área de Jáchal, al norte de la provincia de San Juan. Graptolitos de edad silúrica fueron originalmente hallados en las secciones de la Loma de Los Piojos y el cerro del Fuerte, al sur de la ciudad de Jáchal (Stappenbeck, 1910). Especímenes de Monograptus aff. Priodon (Bronn) y Pristiograptus aff. frequens (Jaekel), fueron originalmente referidos al Devónico por Stappenbeck (1910) y luego al Silúrico por Clarke (1912). La sucesión del cerro del Fuerte fue examinada por Keidel (1921) quien corroboró la edad silúrica de estos depósitos. Posteriormente, Turner (1960) clasificó los ejemplares de M. aff. priodon de la colección de Stappenbeck como Monograptus aff. vomerinus (Nicholson), indicando una edad wenlockiana para los estratos portadores.
Numerosos trabajos sobre la cuenca silúrica de la Precordillera se realizaron desde el siglo pasado, pudiendo citarse los de Heim (1948); Furque y Cuerda (1979); Cuerda (1985); Baldis et al. (1982); Benedetto et al . (1986, 1992a); Baldis y Peralta (1999); Peralta (1990, 1993, 2003); Sánchez et al . (1991, 1993); Astini y Piovano (1992); Astini y Maretto (1996); entre otros.
Los depósitos silúricos de la Precordillera corresponden a facies predominantemente clásticas de colores verdosos y ambientes de plataforma, con abundante valvifauna en sus términos superiores, y conodontes y graptolitos localizados en niveles restringidos. Estudios estratigráficos de la cuenca silúrica indican que la misma se habría desarrollado en un ambiente de antepaís ( e.g ., Benedetto et al ., 1986; Astini y Maretto, 1996).
Los depósitos silúricos se registran en la precordillera oriental (Ortiz y Zambrano, 1981) y central (Baldis y Chebli, 1969), en la provincia de San Juan, separados por la dorsal de Villicum-Zonda ( sensu Padula et al ., 1967). Unidades de probada edad silúrica no han sido reconocidas hasta el presente en la precordillera occidental (Baldis et al ., 1982), impidiendo efectuar un análisis adecuado de estratos de la referida edad para esta parte de la cuenca.
El Grupo Tucunuco (Cuerda, 1965), compuesto por las formaciones La Chilca y Los Espejos, presenta extensos afloramientos desde el área de Jáchal, al norte, hasta la sierra de Talacasto por el sur, en la precordillera central. Entre los perfiles clásicos de estudio de este grupo pueden citarse las localidades de Loma de Los Piojos, al suroeste de Jáchal, quebrada de Las Aguaditas, cerro del Fuerte, cerro La Chilca y sierra de Talacasto. La Formación Tambolar (Heim, 1952) está presente en la sierra del Tontal, siendo su perfil clásico el de la cuesta del Tambolar, sobre el río San Juan. Para algunos autores la Formación Tambolar es equivalente al Grupo Tucunuco ( e.g ., Peralta, 1990; Peralta y Carter, 1990), pero para otros se correlaciona sólo con la Formación Los Espejos ( e.g ., Benedetto et al ., 1992a; Astini y Maretto, 1996).
Estas tres unidades se inician con delgados conglomerados de base con rodados de chert y oolitas ferruginosas (Peralta et al ., 1986; Astini, 1992) que implican la presencia de hiatos erosivos aún dentro de la sedimentación silúrica (Astini y Maretto, 1996). Los mismos caracterizan la fase inicial de cortejos transgresivos y son sucedidos por pelitas que indicarían el momento de máxima inundación en la cuenca silúrica (Astini y Maretto, 1996). Las unidades que integran el Grupo Tucunuco (Cuerda, 1965) han sido objeto de numerosas investigaciones. Estas formaciones contienen valvifaunas, microfósiles, graptolitos y palinomorfos descriptos en diversos trabajos, entre los que pueden citarse los de Cuerda (1969; 1971); Cuerda et al . (1982, 1988); Hünicken (1975); Hünicken y Sarmiento (1988); Volkheimer et al . (1980); Pöthe de Baldis (1981); Kerlleñevich y Cuerda (1986); Peralta (1986a, 1986b); Aceñolaza y Peralta (1986); Waisfeld et al . (1988); Benedetto (1995); Benedetto et al . (1992b); Vannier et al. (1995); Rubinstein (1992a, 1992b, 1995); Lehnert (1995); Rickards et al. (1996); Peralta (1998) y Rubinstein y Brussa (1999). De acuerdo con el contenido de fósiles se ha establecido que la depositación del Grupo Tucunuco se inicia en el Hirnantiano (Ordovícico tardío), observable en la sierra de Talacasto (Cuerda et al ., 1988), y culmina en el Lockoviano (Devónico temprano) en una de las secciones expuestas en el cerro del Fuerte (Benedetto et al ., 1992b; Rubinstein, 1995).
En la precordillera oriental se localizan la Formación Rinconada (Amos, 1954) en el faldeo oriental de la sierra Chica de Zonda y cerro Pedernal (Peralta, 1993) y las formaciones Don Braulio (Baldis et al ., 1982) (parte superior) y Rinconada (= Mogotes Negros) en el flanco oriental de la sierra de Villicum (Peralta, 1984, 1993; Peralta y Carter, 1990). En la porción cuspidal de la Formación Don Braulio se registraron graptolitos referidos a la Zona de Atavograptus atavus por Peralta (1986a, 1993), del Llandovery temprano. La Formación Rinconada, una mélange sedimentaria de gran potencia (3750 m sensu Peralta, 1993) contiene restos de biseriados y monográptidos de edad llandoveriana a ludloviana. Los primeros registros de monográptidos de esta unidad fueron mencionados por Keidel (1938), permitiendo referirla al Silúrico. Hallazgos posteriores, realizados por Cuerda (1981) y Peralta (1986b), ampliaron el conocimiento sobre la graptofauna de esta unidad.
El presente trabajo describe asociaciones de conodontes y graptolitos coleccionadas en las formaciones La Chilca y Los Espejos, en el perfil de la quebrada Ancha, sierra de Talacasto (figura 1). El hallazgo de estos taxones brinda precisiones sobre la composición faunística y edad de estas formaciones, así como sobre la estratigrafía de eventos.

Figura 1. Mapa de ubicación y geología del área de estudio (modificado de Baldis et al ., 1984) / location map and geology of the study area (modified from Baldis et al ., 1984) .
Marco geológico del área de estudio
El Grupo Tucunuco fue definido en el área del cerro La Chilca, aproximadamente 14 km al oeste de la estación Tucunuco (Cuerda, 1965). La presencia de Saetograptus argentinus (Cuerda) y Monograptus uncinatus notouncinatus (Cuerda) en la porción media de la Formación Los Espejos permitió inferir una edad ludloviana temprana para esta unidad (Cuerda, 1969, 1971). El hallazgo de biseriados y monográptidos en la parte inferior de la Formación La Chilca aflorante en el área de Los Baños de Talacasto, permite referir la base de esta unidad al Llandovery temprano (Cuerda et al ., 1982) y posteriormente al Ashgill tardío (Hirnantiano) (Cuerda et al ., 1988). La presencia de Monograptus priodon (Bronn) en la parte superior de la citada unidad, unos 3 km al sur de Los Baños de Talacasto, indica una edad llandoveriana tardía a wenlockiana temprana para los niveles cuspidales (Kerlleñevich, 1986; Kerlleñevich y Cuerda, 1986). El hallazgo de los braquiópodos Dedzetina ? Silurica Benedetto y Leangella ( Leangella ) mutabilis Herrera y Benedetto por encima de la secuencia pelítica basal de la Formación Los Espejos, en la quebrada Poblete (Benedetto et al ., 1992b), permitió inferir una edad wenlockiana para este tramo de la secuencia.
Baldis et al . (1984) y Maretto (en Astini y Maretto, 1996) efectuaron un estudio del Grupo Tucunuco en la sierra de Talacasto, describiendo los perfiles de las cabeceras de la quebrada Ancha, tributaria de la quebrada de Talacasto, y los de la quebrada de Salto Macho, ubicada algunos kilómetros al sur del área anterior.
Baldis et al . (1984) dividieron la Formación La Chilca en dos miembros, uno inferior (Miembro Salto Macho) de composición predominantemente pelítica, y otro superior, con mayor contenido de areniscas y cuarcitas (Miembro de Cuarcitas Azules). La depositación se inicia con un conglomerado de base de poco espesor, compuesto por rodados de chert en una matriz areniscosa (Rolleri, 1947; Marchese, 1972) que apoya discordantemente sobre las calizas de la Formación San Juan (Arenig-Llanvirn temprano). Por encima se desarrolla una secuencia de lutitas negras y grises oscuras, con abundante contenido de graptolitos. Esta facies graptolítica de la base de la Formación La Chilca fue originalmente referida al Ordovícico Superior-Silúrico por Ramos et al . (1978). La presencia de Persculptograptus persculptus (Salter) en la base de estas pelitas indica la biozona del mismo nombre de edad hirnantiana tardía en el área de Los Baños de Talacasto (Cuerda et al ., 1988). La graptofauna que se desarrolla inmediatamente por encima del nivel con P . persculptus fue referida a las zonas de Parakidograptus akuminatus y Atavograptus atavus (Llandovery temprano), si bien los fósiles guías de ambas biozonas no fueron hallados (Cuerda et al ., 1988). Recientemente, Lenz et al . (2003) describen una fauna de retiolítidos y diplográptidos procedente de los niveles pelíticos inferiores de la Formación La Chilca que los autores refieren al Telychiano temprano a medio.
La sedimentación continúa con una sucesión de areniscas y limolitas, consideradas como tempestitas distales y fangolitas bioturbadas de depósitos de plataforma bajo somerización progresiva (Astini y Maretto, 1996). Las trazas fósiles son abundantes en los niveles de areniscas, localizándose la facies de Chondrites en la parte inferior de esta secuencia (Baldis et al ., 1984; Aceñolaza y Peralta, 1986). El registro de M. priodon efectuado por Kerlleñevich (1986) y Kerlleñevich y Cuerda (1986) al sur de Los Baños de Talacasto, se ubicaría en limolitas de esta parte de la sección.
El tramo superior de la Formación La Chilca, en el perfil considerado, corresponde a areniscas cuarzosas amalgamadas de ambiente somero de cara de playa ( sensu Astini y Maretto, 1996). La sucesión es fundamentalmente psamítica, con niveles delgados de fangolitas y coquinas intercaladas en algunas partes de la sucesión. La presencia de Zoophycus sp. Fue registrada en esta parte de la unidad (Baldis et al ., 1984; Aceñolaza y Peralta, 1986).
La Formación Los Espejos expuesta en la quebrada Ancha se inicia con un conglomerado de base transgresivo con oolitas ferruginosas que no supera los 50 cm de espesor, sucedido por pelitas verdes y moradas, sin fósiles, de plataforma externa, que coincidirían con el intervalo de máxima inundación de la cuenca (Astini y Maretto, 1996). Por encima se desarrollan tempestitas distales y fangolitas de tiempo normal (Astini y Maretto, 1996) con trazas de la icnofacies de Nereites (Baldis y Aceñolaza, 1978; Baldis et al ., 1984; Aceñolaza y Peralta, 1986).
La sucesión incorpora areniscas medias a gruesas, progresivamente más abundantes hacia el techo, que indican la somerización de la secuencia. Los niveles de coquinas con una rica valvifauna (braquiópodos, trilobites, tentaculites, gastrópodos, ostrácodos y equinodermos) se localizan en los tramos medio a superior de la secuencia. Baldis et al . (1984) registraron un nivel con M . uncinatus notouncinatus y otro, por encima, con S. argentinus en la parte media de la Formación Los Espejos, en el perfil de la quebrada Ancha, indicando una edad ludloviana para los niveles portadores. En los tramos medio a superior de la formación se ha identificado la icnofacies de Cruziana (Aceñolaza y Peralta, 1986; Astini y Maretto, 1996).
La parte cuspidal de la formación en el perfil de la quebrada Ancha presenta una alternancia de pelitas verdes y areniscas deformadas ( boudines y estructuras en almohadilla) de edad pridoliana, generadas presumiblemente por actividad sísmica (Astini y Mareto, 1996). Estos estratos son cubiertos en paraconcordancia por las lutitas verdes de la Formación Talacasto.
Descripción estratigráfica del perfil estudiado en la quebrada Ancha
La sucesión estratigráfica analizada involucra las formaciones La Chilca y Los Espejos (figura 2). La misma se apoya en discordancia sobre las calizas de la Formación San Juan (Arenig-Llanvirn) y es cubierta en paraconcordancia por las pelitas verdes de la Formación Talacasto (Devónico Inferior). El espesor total de la secuencia silúrica es de 219,4 m, correspondiendo 29,1 m a la Formación La Chilca y 190,3 m a la Formación Los Espejos. El contacto entre las dos unidades silúricas es paraconcordante, mediando entre ambas un delgado conglomerado que corresponde a la base de la Formación Los Espejos.

Figura 2. Columna estratigráfica de la sección silúrica de la quebrada Ancha, Precordillera Central de la provincia de San Juan / stratigraphic column of the Silurian section at Ancha Creek, Central Precordillera of San Juan Province.
Formación San Juan
El tramo cuspidal de esta formación está constituido por calizas margosas nodulares con abundante valvifauna (braquiópodos, trilobites, artejos de crinoideos, gastrópodos y microfósiles). Con el objeto de verificar la edad previamente asignada al techo de la Formación San Juan para el área de Talacasto, se procesaron dos muestras de calizas nodulares procedentes del tramo superior de la unidad; i.e ., a un metro del techo (3300 g) y del mismo nivel cuspidal (2000 g). Ambas muestras produjeron escasos elementos ( ca . 10 especímenes por kg) correspondientes a una asociación de conodontes compuesta por Drepanoistodus basiovalis (Sergeeva), Histiodella sinuosa (Graves y Ellison), Juanognathus serpaglii Stouge, Oistodus lanceolatus Pander, Parapaltodus simplicissimus Stouge, Periodon aculeatus Hadding, Rossodus barnesi Albanesi, Scolopodus striatus Pander, Semiacontiodus potrerillensis Albanesi. Esta asociación de especies, en particular por la presencia de H. sinuosa , se restringe a la Zona de Lenodus variabilis , del Arenig superior alto-Llanvirn inferior bajo, según la definición de Albanesi et al . (1998) para el esquema de biozonación del Sistema Ordovícico de la Precordillera. Esta determinación bioestratigráfica es equivalente a registros de otras secciones de la Precordillera donde se observa el contacto erosivo entre la caliza San Juan y los depósitos silicoclásticos silúricos; por ejemplo, en Rinconada (Lehnert, 1995), a diferencia de la documentación previa dada para Talacasto, donde el techo de la Formación San Juan se atribuyó a la Zona de Pygodus serra (Lozano y Hünicken, 1990).
Formación La Chilca
Esta unidad se apoya sobre la anterior a través de un contacto paraconcordante con un conglomerado basal de clastos de chert y en menor proporción de caliza, dispersos en una matriz areniscosa que hacia el tope se torna en una arenisca con clastos dispersos y alto contenido de hierro. Espesor: 0,5 m.
Continúa una alternancia de pelitas negras y grisverdosas. Las lutitas negras, con marcada fisilidad y abundante contenido de graptolitos, están intensamente trituradas en algunos tramos de la secuencia. Estas pelitas tienen un elevado contenido de sulfuro de hierro, siendo característico el reemplazo de los rabdosomas por pirita. Los biseriados son frecuentes en estos estratos, pero su preservación es deficiente debido a la trituración y meteorización de la roca. Las zonas de P . persculptus , P. acuminatus y A. atavus fueron reconocidas en esta parte de la secuencia aflorante en Los Baños de Talacasto, pocos kilómetros al noreste del perfil aquí estudiado (Cuerda et al ., 1988). Algunos metros por encima del conglomerado basal se detectaron especímenes referibles a Talacastograptus leanzai , restos de monográptidos y conodontes coniformes, conservados como moldes en planos de estratificación de pelitas, probablemente del género Panderodus . Espesor: 8,20 metros.
Sigue una sucesión de areniscas finas a medias y limolitas grises a gris verdosas. Esta secuencia se inicia con un banco de arenisca de 30 cm de potencia, gris oscuro en corte fresco, cubierto por una alteración parda oscura. Los restantes estratos psamíticos presentan espesores de 10-15 cm. Algunos bancos de areniscas contienen restos abundantes de Chondrites sp. Las limolitas presentan laminación paralela preservando esporádicamente restos de graptolitos. La muestra G contiene fragmentos de monográptidos (? Stimulograptus sp.) y biseriados de gran porte referibles al género Diplograptus . En la muestra F se coleccionaron restos de M. cf. priodon y unos 2 metros por encima (muestra E), rabdosomas incompletos de M . cf. priodon , y P . aff. nudus . Este último fósil sugiere una edad llandoveriana tardía (Telychiano) para los niveles portadores. Espesor: 11,40 metros.
La formación culmina con areniscas cuarzosas que intercalan fangolitas grises subordinadas. Dentro de las areniscas se observan algunas coquinas delgadas, lenticulares, formadas por valvas de braquiópodos y bancos de intraclastos pelíticos. Las areniscas finas suelen contener restos fragmentarios y escasos de monográptidos. Espesor: 9 metros.
Formación Los Espejos
Esta formación sobreyace a la anterior en contacto paraconcordante, con un conglomerado basal con rodados de areniscas y chert en una matriz arenosa con alteración ferruginosa. Espesor: 0,30 metros. La parter inferior de la unidad esta representada por una sucesión monótona de pelitas verdes y moradas bioturbadas, sin registros de macrofósiles. Espesor: 85 metros.
A esta sucesión le sobreyace una alternancia de areniscas finas a medias y limolitas verde grisáceas que intercalan algunos bancos de areniscas gruesas. Entre los puntos 4 y 5 del perfil comienzan a aparecer niveles de areniscas finas. Por encima del punto 5 se observan estructuras HCS en las areniscas. A partir del punto 7 se reconocen ondulitas simétricas en el tope de algunas capas de areniscas, a veces surcadas por trazas.
En el tramo superior de la sección, las areniscas con cemento cabonático suelen contener niveles de coquinas en sus bases (muestras A, AB, B, C, D). El grado de preservación de los fósiles de estas coquinas es, en general, muy bueno, identificándose braquiópodos (predominantemente chonétidos), gastrópodos, tentaculites, nautiloideos, restos de euriptéridos, trilobites, ostrácodos, conodontes y graptolitos. Se han observado algunos niveles con trazas de la icnofacies de Cruziana . La coquina del punto 16 (muestra AB), localizada en una calcarenita, contiene abundantes rabdosomas de S . argentinus , que se tornan escasos hacia los niveles de calcarenitas que rodean a la coquina. Se trata de ejemplares maduros, bien conservados, con escasa orientación, donde la ausencia de extremos proximales, sículas y formas juveniles coincide con un proceso de selección por corrientes. Las coquinas constituyen niveles irregulares y presentan frecuentemente intraclastos de pelitas pardooscuras. Las calcarenitas tienen estructura laminar planar fina. El tramo final de la Formación Los Espejos (puntos 16 a 17) contiene una sucesión de limolitas gris verdosas con alteración parda y bancos de areniscas finas grises de 5 a 10 cm de espesor, con indicios de deformación sinsedimentaria (boudinage), que termina en un nivel de arenisca de 30 cm de potencia, con icnitas. Espesor: 105 m.
A partir del punto 17, y en contacto paraconcordante sobre la Formación Los Espejos, se superponen limolitas verdosas bioturbadas correspondientes a la Formación Tambolar del Devónico temprano.
Bioestratigrafía y eventos
Conodontes. Tres trabajos previos al presente se refieren a conodontes de edad silúrica en la geología argentina. Hünicken (1975) documenta por primera vez conodontes wenlockianos, procedentes de una muestra de la parte superior del afloramiento de la Formación La Chilca en la localidad de Loma de Los Piojos, departamento Jáchal de la provincia de San Juan. El estado de conservación fragmentario de los especímenes ilustrados no permite asignaciones específicas precisas, y la edad atribuida al nivel de procedencia no ha sido aún verificada. En relación a conodontes del Silúrico inferior de la Precordillera de San Juan, Lehnert et al . (1999) publican una asociación de especies procedente de la parte inferior de la Formación La Chilca, expuesta en el flanco occidental del cerro del Fuerte, unos 20 km al sureste de la ciudad de Jáchal. Esta colección incluye las especies Dapsilodus obliquicostatus (Branson y Mehl), Distomodus cf. kentuckyensis Branson y Branson, Ozarkodina oldhamensis (Rexroad), Distomodus ? sp. Y Panderodus sp., y los autores la asignan a la Zona de Distomodus kentuckyensis , que representa el intervalo Rhuddaniano medio a Aeroniano de la Serie Llandovery. Hünicken y Sarmiento (1988) registran conodontes de la parte media de la Formación Los Espejos, en la sección de la quebrada Ancha, sierra de Talacasto, en la precordillera central de San Juan, probablemente de estratos correlativos a los que produjeron la colección del presente estudio, considerando la posición del nivel muestreado y las especies descriptas por los autores citados; i.e ., Distomodus dubius (Rhodes), Hindeodella excavata (Branson y Mehl), Ligonodina excavata (Branson y Mehl), Panderodus unicostatus (Branson y Mehl), Panderodus sp. A Hünicken, Pelekysgnathus dubius Jeppsson, Pseudooneotodus beckmanni (Bischoff y Sannemann) (véase sinonimia en paleontología sistemática). Hünicken y Sarmiento (1988) atribuyen el intervalo de aparición de la asociación de conodontes referida a la Zona de Polygnathoides siluricus y parte basal de la Zona de Pedavis latialata , según los esquemas de Jeppsson (1974) para Escandinavia y de Walliser (1964) para los Alpes Cárnicos de Austria, que refieren a una edad ludloviana media alta a tardía baja. Sin embargo, ninguna de las especies referidas, ni la combinación de la distribución vertical de las especies asociadas restringen concretamente a la edad indicada, si bien también podrían registrarse en ese intervalo del Silúrico tardío.
La colección de conodontes que aquí se describe se recuperó del residuo insoluble de cinco muestras de coquinas carbonáticas (18290 g) de la Formación Los Espejos (figura 2), las cuales se desagregaron mediante métodos de laboratorio convencionales (ácido acético al 10%, véase Stone, 1987). Se rescataron 983 elementos (figura 3), que se encuentran fragmentados en su mayor proporción, y se observaron escasos especímenes conservados como moldes, o con su material original, en planos de estratificación de pelitas o areniscas finas. Los conodontes aislados presentan la coloración marrón propia del índice de alteración del color ("CAI") 3, que corresponde a paleotemperaturas de soterramiento de entre 110 y 200° C (Epstein et al ., 1977).

Figura 3. Distribución de especies de conodontes en las muestras de la Formación Los Espejos / conodont species distribution in samples of the Los Espejos Formation.
En el presente estudio se describen las especies Kockelella variabilis variabilis Walliser, Ozarkodina excavata excavata (Branson y Mehl), Ozarkodina cf. Confluens (Branson y Mehl), Panderodus unicostatus (Branson y Mehl), Oulodus siluricus (Branson y Mehl), Coryssognathus dubius (Rhodes) y Pseudooneotodus beckmanni (Bischoff y Sannemann), las cuales fueron mencionadas y parcialmente ilustradas por Albanesi (en Peralta et al ., 2003). Este conjunto de taxones es característico de la Serie Ludlow en la localidad tipo y otras secciones de ambientes de plataforma de Gales (Aldridge, 1975), que comprende cuatro biozonas (Aldridge y Schönlaub, 1989) en un esquema basado en la biozonación originalmente propuesta por Walliser (1964) para Europa central; sin embargo, los autores reconocieron que el esquema propuesto para la localidad tipo no es de aplicación general. En un trabajo reciente, Corradini y Serpagli (1998) revisaron el esquema de biozonación de conodontes del Sistema Silúrico sobre la base de nuevos datos de secciones en Cerdeña, Italia, proponiendo nuevas unidades y reintroduciendo biozonas previamente consideradas. De acuerdo con estos autores, el esquema de Cerdeña se adapta a otras partes del mundo así como también a la sección de Cellon en los Alpes Cárnicos de Austria, donde se estableció el primer esquema de biozonación formal para el Silúrico (Walliser, 1964); asimismo, proponen reemplazar la carta de biozonación global de conodontes que apareció en el boletín de la Subcomisión Internacional de Estratigrafía del Silúrico ( Silurian Times n° 3, Nowlan, 1995). El esquema de Corradini y Serpagli (1998) comprende ocho zonas de conodontes para la Serie Ludlow, la cual está dividida en los pisos Gorstiano (inferior) y Ludfordiano (superior). A su vez, el Gorstiano incluye las zonas de Kockelella crassa , K . Variabilis variabilis , Ozarkodina excavata hamata , y la parte inferior de Ancoradella ploeckensis .
El registro de Kockelella v. variabilis en asociación con las otras especies recuperadas de los niveles medios a superiores de la Formación Los Espejos, permite referir el intervalo muestreado a la Zona de K. v. varabilis reintroducida por Corradini y Serpagli (1998). Esta atribución podría confirmarse por la ausencia de O. e. hamata (cuya primera aparición define la base de la biozona siguiente), en una colección importante de conodontes como la proporcionada por los estratos investigados de esta formación. En consecuencia, la parte media a superior de la Formación Los Espejos en la sección de la quebrada Ancha, en el área de Talacasto, es referida por conodontes guías a la segunda biozona del Gorstiano en el Ludlow inferior. En el área tipo Ludlow, este intervalo cronoestratigráfico está representado por la Formación Bringewood que produjo conodontes de las especies que aquí se describen (Aldridge, 1975; Aldridge y Smith, 1985).
El intervalo productivo de conodontes de la Formación Los Espejos aparentemente representa el Evento Linde Primo-Secundo, definido por Jeppsson y Aldridge (2000), compartiendo las mismas especies que se registran aquí. Los eventos y episodios oceánicos descriptos para el Silúrico resultan del modelo predictivo basado en la alternancia de estados del sistema océano-atmósfera fundamentado por Jeppsson (1990), y posteriormente sintetizado y explicado para el Sistema Silúrico por Jeppsson (1998). Los eventos corresponden a breves intervalos perturbados entre episodios estables de distinta naturaleza ( i.e ., primo y secundo), en los cuales se producen extinciones o cambios bióticos significativos. De acuerdo con la escala de severidad de eventos silúricos (Jeppsson, 1998), el Evento Linde alcanza una severidad media. Durante este evento se produce una aparición creciente de elementos coniformes, especialmente del género Panderodus , una fuerte caída en la frecuencia y diversidad de elementos con plataformas, el enanismo de taxones y/o un cambio a poblaciones dominadas por estadios juveniles (Jeppsson y Aldridge, 2000). Todas estas características se observan en la colección de conodontes de la Formación Los Espejos. Cabe destacar el usual carácter monoespecífico de los distintos grupos asociados a los conodontes procedentes de las coquinas carbonáticas depositadas en ambientes de off-shore proximal; en particular, los graptolitos representados por Saetograptus argentinus , si bien dada la condición endémica de este grupo, no es posible efectuar comparaciones de tamaño en relación a otras formas afines. Jeppsson (1990) ha interpretado que el enanismo de los individuos puede resultar de un ajuste a la productividad planctónica baja, probablemente influenciada por las estaciones climáticas, y por el cese en el suministro de nutrientes desde el upwelling de aguas profundas. Incidentalmente, la fauna de conodontes de la Formación Los Espejos se caracteriza por presentar, en general, elementos adultos de tamaño pequeño y las formas juveniles son frecuentes. Probablemente esta particularidad, así como la peculiar fauna monoespecífica de graptolitos, se pueda explicar por las condiciones oceánicas eventuales del modelo de Jeppsson, y por el efecto de las fluctuaciones del nivel del mar, en un período que no se caracteriza por un definido provincialismo faunístico ( cf ., discusión Loydell vs. Jeppsson y Aldridge, 2000).
Graptolitos. El hallazgo de Persculptograptus persculptus (Lapworth) en asociación con Normalograptus angustus (Perner), N . cf. medius (Törnquist), N. Normalis (Lapworth) y Metaclimacograptus robustus Cuerda, Rickards y Cingolani, en la base de la Formación La Chilca, en el área de Los Baños de Talacasto, indica la presencia de estratos del Ashgill más tardío (Hirnantiano) (Cuerda et al ., 1988). Por encima de estos niveles se localiza una graptofauna compuesta por N . angustus , N . cf. medius , N. normalis , M . Robustus y Climacograptus rectangularis (McCoy), referida a la Zona de P. acuminatus de la base del Llandovery (Rhuddaniano) (Cuerda et al ., 1988). Esta asociación es sucedida por fauna de la Zona de A. atavus con Lagarograptus praeacinaces Cuerda, Rickards y Cingolani, Talacastograptus leanzai Cuerda, Rickards y Cingolani, N. angustus y N. normalis (Rhuddaniano) (Cuerda et al ., 1988).
Una fauna compuesta por Metaclimacograptus cf. asejradi Legrand y Pseudoplegmatograptus reticulatus (Boucek y Münch) ha sido recientemente localizada a ca. 1,30 m de la base de la Formación La Chilca (Lenz et al ., 2003). La asociación correspondería, de acuerdo a estos autores, a las zonas de Spirograptus turriculatus (Telychiano temprano) o Streptograptus crispus (Telychiano medio).
Los restos de ? Stimulograptus localizados en la muestra G del perfil medido, por encima de la fauna de atavus , sugieren una edad aeroniana tardía (posiblemente zonas de S. sedgwickii y S. halli ) para los estratos portadores, en términos de la escala británica, si bien estos datos deben corroborarse con colecciones más completas.
Unos 3 m por encima de los estratos con ? Stimulograptus , en la muestra F del perfil, se localizaron restos de monográptidos comparables con M. . priodon ( cf . Bronn), una forma ya descrita para el tramo medio superior de la Formación La Chilca (Kerlleñevich, 1986; Kerlleñevich y Cuerda, 1986). El rango de este graptolito se extiende entre las zonas de S. turriculatus y Cyrtograptus centrifugus (Llandovery tardío-Wenlock temprano) de Europa ( e.g ., Elles y Wood, 1911; Bjerreskov, 1975), China (Mu et al ., 1974) y Canadá (Lenz, 1982; Lenz y Kozlowska-Dawidziuk, 2001; Zhang, 1994).
La muestra E, hacia el tope de la sucesión de areniscas finas a medias y fangolitas verdosas contiene el primer registro de Pristiograptus aff. Nudus (Lapworth). Esta forma se extiende desde la Zona de S. turriculatus hasta la Zona de Monoclimacis crenulata en Gran Bretaña y Dinamarca (Elles y Wood, 1911; Rickards, 1970; Rickards et al ., 1977; Bjerreskov, 1975), indicando una edad llandoveriana alta (Telychiano) para los niveles medio-altos de la Formación La Chilca, inmediatamente por debajo del tramo cuspidal con areniscas amalgamadas.
En la Formación Los Espejos se registran rabdosomas de S. argentinus en una coquina con abundante valvifauna y conodontes (muestra AB) a ca . 176 m de la base de la formación. La especie indica una edad ludloviana temprana para este tramo de la sección (Cuerda, 1969). M. u. notouncinatus , citado por Baldis et al . (1984) para el área de la quebrada Ancha, no fue localizado en el perfil estudiado.
Paleontología sistemática
Conodontes
En la taxonomía de conodontes se aplica el sistema de notación convencional propuesto por Sweet (1981, 1988), con las posiciones espaciales M, S y P, desde el extremo anterior al posterior del aparato multielemental, y las subposiciones de transición en simetría correspondientes. Recientemente, Purnell et al. (2000) propusieron un esquema de nomenclatura para aparatos complejos que integran elementos coniformes, ramiformes y pectiniformes, que respondería a la posición anatómica de los elementos en la cavidad oral del animal. No obstante, la arquitectura de los aparatos de distintos géneros es, en general, poco conocida; por lo cual, mantenemos esquemas de orientación y notación convencionales, siguiendo los autores que describieron las especies determinadas. En particular, para el género Panderodus , con elementos exclusivamente coniformes, adoptamos la nomenclatura propuesta por Sansom et al . (1994). La clasificación sistemática, que incluye a los conodontes como vertebrados primitivos ( e.g ., Donoghue et al ., 2000), mantiene relaciones de niveles supragenéricos de asignación aún dudosa. Recientemente, Sweet y Donoghue (2001) efectuaron una revisión de las categorías formuladas por modelos anteriores, la cual se sigue en el presente trabajo. La distribución geográfica de los taxones descriptos se revela a través de las citas de la sinonimia. La colección de conodontes estudiada se encuentra depositada en el Museo de Paleontología de la Universidad Nacional de Córdoba, Argentina, bajo la sigla de repositorio CORD-MP.
Clase Conodonta Pander, 1856
Orden Ozarkodinida Dzik, 1976
Familia Kockelellidae Klapper, 1981
Género Kockelella Walliser, 1957
Especie tipo. Kockelella variabilis Walliser, 1957.
Kockelella variabilis variabilis Walliser, 1957
Figuras 6.K-U
Kockelella variabilis Walliser, 1957, p. 35, pl. 1, figs. 3-10.
Kockelella variabilis Walliser, 1957, Simpson y Talent, 1995, p. 137-138, pl. 6, figs. 8-17 ( cum syn .).
Kockelella variabilis variabilis Walliser, 1957, Serpagli y Corradini, 1998, p. 288-293, pl. 5, figs. 1.13, pl. 6, figs. 1-9, pl. 6, figs. 1-7 ( cum syn .).
Observaciones. Kockelella variabilis variabilis integra seis morfotipos en su aparato multielemental; i.e ., M, Sa, Sb, Sc, Pa, Pb. Un estudio exhaustivo sobre este taxón, incluyendo la sinonimia completa hasta 1998, fue efectuado por Serpagli y Corradini. El material recuperado en el presente estudio incluye todos los morfotipos de la especie, cuya morfología se ajusta precisamente a la descripción brindada por los autores citados.
Procedencia. Formación Los Espejos, muestras A, AB, B, C, D.
Material y repositorio. 139 elementos, CORD-MP 10048-10049, 10196-10202, 10556-1694.
Familia Spathognathodontidae Hass, 1959
Género Ozarkodina Branson y Mehl, 1933
Especie tipo. Ozarkodina typica Branson y Mehl, 1933 (= sinónimo junior de Hindiodella confluens Branson y Mehl, 1933).
Ozarkodina excavata excavata (Branson y Mehl, 1933)
Figuras 5.M-Y
Prioniodus excavatus Branson y Mehl, 1933, p. 45, pl. 3, figs. 7, 8.
Hindeodella excavata excavata (Branson y Mehl, 1933), sensu Jeppsson, 1969, Jeppsson, 1974, p. 25-31, pl. 4, figs. 1-17 ( cum syn .).
Hindeodella excavata (Branson y Mehl, 1933), sensu Jeppsson, 1969, Hünicken y Sarmiento, 1988, p. 226, lám. 1, figs. 1, 2, 6; lám. 2, figs. 3-5, 10, 11.
Ligonodina excavata (Branson y Mehl, 1933), Hünicken y Sarmiento, 1988, p. 227, lám. 1, figs. 3-5, 8; lám. 2, figs. 1, 2, 6-9.
Ozarkodina excavata excavata (Branson y Mehl, 1933), Barca et al ., 1992, p. 132, pl. 10, figs. 3-5.
Ozarkodina excavata (Branson y Mehl, 1933), Sarmiento et al ., 1994, p. 516, pl. 2, figs. 10, 12, 13, 15.
Ozarkodina excavata excavata (Branson y Mehl, 1933), Simpson y Talent, 1995, p. 147-153, pl. 8, figs. 16-25, pl. 9, figs. 1-24 ( cum syn .).
Ozarkodina excavata (Branson y Mehl, 1933), Männik y Malkowski, 1998, p. 148, pl. 1, figs. 3, 4.
Ozarkodina excavata excavata (Branson y Mehl, 1933), Serpagli et al ., 1998, p. 110, pl. 1.2.1, figs. 4, 5; pl. 1.2.2, fig. 1.
Ozarkodina excavata excavata (Branson y Mehl, 1933), Ferretti et al., 1998, p. 160, pl. 2.2.1, fig. 1.
Ozarkodina excavata (Branson y Mehl, 1933), Benfrika, 1998, p. 315, pl. 1, fig. 7.
Ozarkodina excavata (Branson y Mehl, 1933), Viira, 1998, p. 300, pl. 1, figs. 1-4.
Ozarkodina excavata (Branson y Mehl, 1933), Sarmiento et al ., 1999, p. 761-762, fig. 2, 1, 2, 4-10, 12, 13, 15, 17.
Observaciones. Los distintos morfotipos que integran el aparato multielemental del género Ozarkodina fueron descriptos por Jeppsson (1971). Las variaciones intraespecíficas de O . excavata fueron reconocidas por Walliser (1964) y posteriormente discutidas por Jeppsson (1974). La presente colección incluye todos los morfotipos que integran el aparato de la subespecie O. e. excavata (véase revisión por Simpson y Talent, 1995); i.e ., M, Sa, Sb, Sc, Pa y Pb, ilustrados en diferentes vistas en la figura 5. El complejo intraespecífico de O . excavata tiene una amplia distribución paleogeográfica y su rango estratigráfico se extiende a través de la mayor parte del Silúrico y el Devónico Inferior (véase sinonimia).
Procedencia. Formación Los Espejos, muestras A, AB, B, C, D.
Material y repositorio. 483 elementos, CORD-MP 10055, 10177-10187, 10695-11177.
Ozarkodina cf. confluens (Branson y Mehl, 1933)
Figuras 6.A-D
cf. Hindeodella confluens Branson y Mehl, 1933, p. 45, pl. 3, figs. 22, 23.
cf. Hindeodella confluens Branson y Mehl, 1933; Jeppsson, 1974, p. 31-35, pl. 5 (pars figs. 1-5).
Observaciones. La especie O . confluens se identifica por el característico morfotipo Pa, con dentículos fusionados hacia la base y algunos dentículos del proceso anterior más altos que la cúspide en el estadio adulto. Los especímenes recuperados de la Formación Los Espejos presentan una morfología semejante a los ilustrados para estadios ontogenéticos tempranos de este morfotipo, en los cuales la altura de los dentículos anteriores aún no supera la altura de la cúspide ( cf ., Jeppsson, 1974, pl. 5, figs. 1-5). Por otra parte, el ángulo ántero-basal del elemento ilustrado es obtuso, a diferencia de los típicos elementos de la especie, en los cuales es ligeramente agudo. La ausencia de elementos adultos que confirmen la morfología del elemento Pa, diagnóstico para la especie, no permite verificar la asignación específica.
Procedencia. Formación Los Espejos, muestras C, D.
Material y repositorio. 91 elementos, CORD-MP 10054, 10189-10191, 10459-10555.
Orden Panderodontida Sweet, 1988
Familia Panderodontidae Lindström, 1970
Género Panderodus Ethington, 1959
Especie tipo. Paltodus unicostatus Branson y Mehl, 1933.
Panderodus unicostatus (Branson y Mehl, 1933)
Figuras 5.A-F
Paltodus unicostatus Branson y Mehl, 1933, p. 42, pl. 3, fig. 3.
Panderodus unicostatus (Branson y Mehl), Hünicken y Sarmiento, 1988, p. 227, lám. 3, figs. 11-13a-b.
Panderodus unicostatus (Branson y Mehl), Zhang y Barnes, 2002, pl. 32-33, fig. 15.1-15.24 ( cum syn .).
Observaciones. Desde la primera descripción de elementos de esta especie por Branson y Mehl (1933), la misma se ha registrado en numerosas localidades del mundo (véase sinonimia detallada en Zhang y Barnes, 2002), verificándose la ubicuidad de este taxón euritópico y de extenso rango estratigráfico, a través de la mayor parte del Silúrico. Una primera reconstrucción del aparato multielemental de la especie fue efectuada por Smith et al. (1987) a partir de restos de una asociación natural procedente del Silúrico inferior del estado de Wisconsin, América del Norte. Los autores interpretaron que el aparato consiste en dos mitades opuestas que operan bilateralmente y comprenden siete morfotipos distintos. Dzik y Drygant (1986) efectuaron una nueva reconstrucción a partir de un agregado natural de conodontes ( cluster ), si bien incompleto, hallado en rocas del Llandovery de Podolia, Ucrania, interpretando una disposición de elementos análoga a la de los chaetognathos actuales. Los especímenes que se ilustran de la Formación Los Espejos son todos los morfotipos reconocidos en nuestra colección, los cuales se denominan siguiendo el esquema nomenclatural proporcionado por Sansom et al . (1994). Estos autores reconstruyeron el aparato multielemental del género Panderodus a partir de evidencias proporcionadas por los tres tipos de conservación que presentan los conodontes; i.e ., como asociaciones naturales en planos de estratificación, como agregados ( clusters ) y como elementos aislados. De acuerdo con estos autores, el género posee un aparato nonimembrado incluyendo siete morfotipos distintos (arcuatiforme, qa; graciliforme asimétrico, qg; graciliforme subsimétrico, qg; truncatiforme, qt; aequaliforme, ae; falciforme, pf; tortiforme, pt), dos de los cuales se repiten en la arquitectura del aparato, compuesta por dos grupos de elementos y un elemento simétrico medio, con un total de diecisiete elementos.
Procedencia. Formación Los Espejos, muestras A, AB, C, D.
Material y repositorio. 20 elementos, CORD-MP 10057, 10169-10176, 10203-10213.
Orden Prioniodinida Sweet, 1988
Familia Prioniodinidae Bassler, 1925
Género Oulodus Branson y Mehl, 1933
Especie tipo. Oulodus mediocris Branson y Mehl, 1933 [sinónimo junior de Oulodus serratus (Stauffer, 1930)].
Oulodus siluricus (Branson y Mehl, 1933)
Figuras 6.E-J
Ligonodina silurica Branson y Mehl, 1933, p. 48-49, pl. 3, figs. 18-20.
Oulodus siluricus (Branson y Mehl, 1933), Serpagli et al ., 1998, p. 30, pl. 1.2.1, figs. 1-3.
Oulodus siluricus (Branson y Mehl, 1933), Ferretti et al ., 1998, p. 164, pl. 2.2.2, fig. 6.
Oulodus ? siluricus siluricus (Branson y Mehl, 1933), Männik y Malkowski, 1998, p. 150, pl. 2, figs. 16, 17, 20, 21.
Observaciones. Una revisión de la sinonimia a nivel genérico y la reconstrucción del aparato multielemental de Oulodus fueron efectuadas por Sweet y Schönlaub (1975) y Mawson (1986), incorporando elementos en las posiciones M, Sa, Sb, Sc, Pa, Pb. En el presente trabajo se identifican e ilustran los morfotipos alate Sa, digyrate breviforme Sb y digyrate extensiforme: prioniodiforme Pa y oulodiforme Pb, los cuales permiten una determinación precisa a nivel específico, correspondiendo a las descripciones e ilustraciones citadas en la sinonimia.
Procedencia. Formación Los Espejos, muestras A, AB, B, C, D.
Material y repositorio. 199 elementos, CORD-MP 10052-10053, 10194-10197, 10214-10407.
Orden Prioniodontida Dzik, 1976
Familia Distomodontidae Klapper, 1981
Género Coryssognathus Link y Druce, 1972
Especie tipo. Cordylodus ? dubius Rhodes, 1953.
Coryssognathus dubius (Rhodes, 1953)
Figuras 5.G-J
Cordylodus ? dubius Rhodes, 1953, p. 299, pl. 23, figs. 221-223.
Distomodus dubius (Rhodes, 1953) sensu Jeppsson, 1972, Hünicken y Sarmiento, 1988, p. 226, lám. 3, figs. 1, 3 y 5.
Pelekisgnathus dubius Jeppsson, 1972, Hünicken y Sarmiento, 1988, p. 228, lám 3, figs. 4a-b, 6-9.
Coryssognathus dubius (Rhodes, 1953), Miller y Aldridge, 1993, p. 241-255, pls. 1-4., figs. 1-6 ( cum syn .).
Coryssognathus dubius (Rhodes, 1953), Serpagli et al ., 1997, p. 240-241, pl. 1, figs. 1-15 ( cum syn .).
Observaciones. El aparato multielemental de esta especie fue reconstruido por Miller y Aldridge (1993) a partir de material procedente de la Formación Upper Whitcliffe del Ludloow de Gales. Estos autores describieron siete morfotipos distintos; i.e ., Pa, Pb, Pc, Sa/Sb, Sb, Sc, M, y reconocieron 16 elementos constituyendo el aparato multielemental de un individuo. Las vinculaciones sistemáticas de este taxón fueron consideradas particularmente por los autores citados y por Serpagli et al . (1997) sobre una colección procedente del Silúrico del sur de Cerdeña. En el presente estudio identificamos los morfotipos M, Pa y Pc, con una baja frecuencia de aparición en la Formación Los Espejos.
Procedencia. Formación Los Espejos, muestra C.
Material y repositorio. 6 elementos, CORD-MP 10056-10057, 10175-10176, 10408-10409.
Orden Protopanderodontida Sweet, 1988
Familia ProtopanderodontidaE Lindström, 1970
Género Pseudooneotodus Drygant, 1974
Especie tipo. Oneotodus ? beckmanni Bischoff y Sannemann, 1958 (= sinónimo senior de Coryssognathus dentatus Link y Druce, 1972).
Pseudooneotodus beckmanni (Bischoff y Sannemann, 1958)
Figuras 5.K, L
Oneotodus ? beckmanni Bischoff y Sannemann, 1958, p. 98, pl. 15, figs. 22-25.
Pseudooneotodus beckmanni (Bischoff y Sannemann, 1958), Hünicken y Sarmiento, 1988, p. 228, lám. 1, figs. 7a-b; lám. 3, figs. 10a-b.
Pseudooneotodus beckmanni (Bischoff y Sannemann, 1958), Zhang y Barnes, 2002, p. 33-34, figs. 17.21, 17.22, 17.25, 17.26 ( cum syn .).
Observaciones. La constitución multielemental de Pseudooneotodus incluye morfotipos ligeramente diferenciados; así, Sweet (1988) distinguió dos formas diferenciadas por la altura de la corona ( cf ., Bischoff, 1986). La especie P. beckmanni desarrolla solo un dentículo con forma de nodo apical y se identifica de otras cogenéricas asumiendo que la frecuencia de las distintas morfologías es independiente, como lo observa Jeppsson (1997) en su discusión sobre distintas formas registradas en el Silúrico de Götland. Los elementos ilustrados de nuestra colección representan los dos morfotipos reconocidos por Sweet (1988), uno con corona achatada y otro con corona acuminada, con flancos lisos y ligeramente ondulados en forma concéntrica.
Procedencia. Formación Los Espejos, muestras A, AB, C, D.
Material y repositorio. 48 elementos, CORD-MP 10050, 10176, 10410-10458.
Graptolitos
Los graptolitos que se describen a continuación están depositados en el Museo de Paleontología de la Universidad Nacional de Córdoba, Argentina, identificados con la sigla de repositorio CORD-PZ.
Familia Monograptidae Lapworth, 1873
Género Pristiograptus Jaekel, 1889
Especie tipo. Graptolithus dubius Suess, 1851.
Pristiograptus aff. nudus (Lapworth 1880)
Figuras 4.A-D, G
aff. Monograptus hisingeri Carruthers var. nudus var. nov. Lapworth, 1880, 156, Pl. 4: 7 a-c.
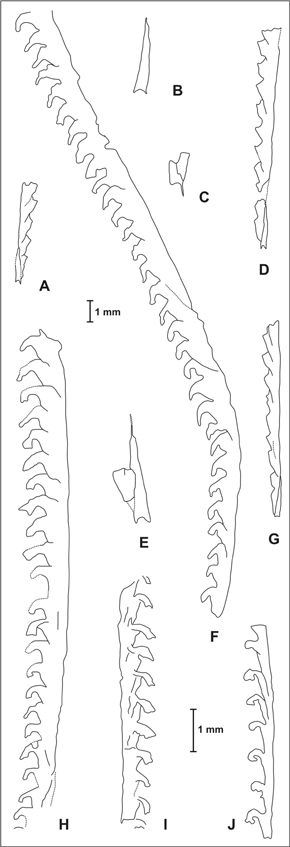
Figura 4. A-D, G, Pristiograptus aff. nudus (Lapworth, 1880), Formación La Chilca, muestra E. A, parte proximal de rabdosoma con la sícula y las cinco primeras tecas / proximal part of rhabdosome with sicula and first five thecae , CORD-PZ 23940, x 6; B, sícula completa / complete sicula , CORD-PZ 23940, x 12; C, parte apertural de sícula e inicio de la primera teca pocos milímetros por encima de la apertura sicular / apertural part of the sicula and beginning of the first theca few milimeters above the sicular aperture , CORDPZ 23940, x 12; D, fragmento de colonia madura / fragment of mature colony, CORD-PZ 23940, x 6; G, parte proximal de rabdosoma / proximal part of the rhabdosome , CORD-PZ 23940, x 6. E, F, H, Monograptus cf. priodon (Bronn, 1835), Formación La Chilca, muestras E, F. E, sícula con primera teca incompleta / sicula with first theca incomplete , CORD-PZ 23934-A, x 12; F, parte media y distal de colonia madura / middle and distal part of mature colony , CORD-PZ 23939, x 6; H, fragmento de rabdosoma maduro / fragment of mature rhabdosome , CORD-PZ 23937-A, x 6. I, J, ? Stimulograptus sp. Formación La Chilca, muestra G. I, fragmento de colonia / fragment of colony , CORD-PZ 23929, x 6; J, fragmento de rabdosoma, probablemente proximal / probably proximal rhabdosome fragment , CORD-PZ 23952-A, x 6.

Figura 5. A-F, Panderodus unicostatus (Branson y Mehl, 1933). A, elemento tortiforme (pt), vista lateral / tortiform element (pt), lateral view , CORD-MP 10169, x 240; B, elemento falciforme (pf), vista lateral del flanco con surco / falciform element (pf), lateral view of the furrowed face , CORD-MP 10170, x 200; C, elemento falciforme (pf), vista lateral del flanco sin surco / falciform element (pf), lateral view of the unfurrowed face , CORD-MP 10171, x 180; D, elemento graciliforme asimétrico (qg),vista lateral del flanco con surco / asymmetrical graciliform element (qg), lateral view of the furrowed face, CORD-MP 10172, x 220; E, elemento graciliforme asimétrico (qg), vista lateral del flanco sin surco / asymmetrical graciliform element (qg), lateral view of the unfurrowed face , CORD-MP 10051, x 180; F, elemento truncatiforme (qt), vista lateral del flanco con surco / truncatiform element (qt), lateral view of the furrowed face , CORD-MP 10173, x 160. Todos los elementos proceden de la muestra C / all elements from sample C . G-J, Coryssognathus dubius (Rhodes, 1953). G, elemento M, vista póstero-lateral, muestra C / Pb element, postero-lateral view, sample C, CORD-MP 10174, x 320; H, elemento Pb, vista póstero-lateral, muestra C / Pc element, postero-lateral view, sample C, CORD-MP 10056, x 220; I, elemento Pa, vista lateral, muestra A / Pa element, lateral view, sample A, CORD-MP 10175, x 200; J, elemento Pa, vista lateral, muestra C / Pa element, lateral view, sample C, CORD-MP 10057, x 180. K, L, Pseudooneotodus beckmanni (Bischoff y Sannemann, 1958). K, vista superior, muestra AB / upper view, sample B, CORD-MP 10050, x 260; L, vista lateral-inferior, muestra AB / lower-lateral view, sample B, CORD-MP 10176, x 320. M-Y, Ozarkodina excavata excavata (Branson y Mehl, 1933). M, elemento Pa izquierdo, vista lateral interna / sinistral Pa element, inner-lateral view , CORD-MP 10177, x 180; N, elemento Pa derecho, vista lateral interna / sinistral Pa element, inner-lateral view , CORD-MP 10178, x 150; O, elemento Pa derecho, vista lateral externa / dextral Pa element, outer-lateral view , CORD-MP 10055, x 150; P, elemento Pa izquierdo, vista lateral externa / sinistral Pa element, outer-lateral view , CORD-MP 10179, x 130; Q, elemento Sa, vista posterior / Sa element, posterior view, CORD-MP 10180, x 220; R, elemento Pb derecho, vista lateral externa / dextral Pb element, outer-lateral view, CORD-MP 10181, x 200; S, elemento Pb izquierdo, vista lateral interna / sinistral Pb element, inner-lateral view , CORD-MP 10182, x 160; T, elemento M derecho, vista lateral externa / destral M element, outer-lateral view , CORD-MP 10183, x 260; U, elemento M izquierdo, vista lateral interna / sinistral M element, inner-lateral view , CORD-MP 10184, x 180; V, elemento Sb izquierdo, vista lateral externa / sinistral Sb element, outer-lateral view , CORD-MP 10185, x 180; W, elemento Sc derecho, vista lateral interna / dextral Sc element, outer-lateral view , CORD-MP 10186, x 130; X, elemento Sc derecho, vista lateral externa / dextral Sc element, outer-lateral view , CORD-MP 10187, x 120; Y, elemento Sc izquierdo, vista lateral interna / sinistral Sc element, inner lateral view , CORD-MP 10188, x 220. Todos los elementos proceden de la muestra AB / all element from sample AB . Todas las ilustraciones de los especímenes son microfotografías MEB / all illustration of specimens are SEM microphotographs.

Figura 6. A-D. Ozarkodina cf. confluens (Branson y Mehl, 1933). A, elemento Sa, vista posterior, muestra D / Sa element, posterior view, sample D , CORD-MP 10189, x 260; B, elemento Pb, vista lateral interna, muestra D / Pb element, inner-lateral view, sample D , CORD-MP 10190, x 200; C, elemento Pa, vista lateral interna, muestra D / Pa element, inner-lateral view, sample D , CORD-MP 10054, x 200; D, elemento Sb, vista superior del flanco interno, muestra D / Sb element, upper view of the inner face, sample D , CORD-MP 10191, x 160. E-J, Oulodus siluricus (Branson y Mehl, 1933). E, elemento Sa, vista posterior, muestra B / Sa element, posterior view, sample B , CORD-MP 10192, x 130; F, elemento Sa, vista anterior, muestra B / Sa element, anterior view, sample B , CORD-MP 10193, x 110; G, elemento Pb izquierdo, vista anterior, muestra B / sinistral Pb element, anterior view, sample B , CORD-MP 10194, x 120; H, elemento Pb izquierdo, vista posterior, muestra B / sinistral Pb element, posterior view, sample B , CORD-MP 10052, x 120; I, elemento Pa derecho, vista anterior, muestra B / dextral Pa element, anterior view, sample B , CORD-MP 10053, x120; J, elemento Sb izquierdo, vista posterior, muestra B / s inistral Sb element, posterior view, sample B, CORD-MP 10195, x 94. K-U, Kockelella variabilis variabilis Walliser, 1957. K, elemento M derecho, vista lateral interna, muestra B / dextral M element, inner lateral view, sample AB , CORD-MP 10196, x 180; L, elemento Sa, vista posterior, muestra AB / Sa element, posterior view, sample B , CORD-MP 10197, x 120; M, elemento Pb derecho, vista lateral interna, muestra AB / dextral Pb element, inner lateral view, sample AB , CORD-MP 10198, x 130; N, elemento Sc derecho, vista lateral interna, muestra AB / destral Sc element, inner lateral view, sample AB , CORD-MP 10199, x 130; O, elemento Sc izquierdo, vista lateral interna, muestra AB / sinistral Sc element, inner-lateral view, sample AB , CORD-MP 10200, x 130; P, elemento Sb, vista posterior, muestra B / Sb element, posterior view, sample AB , CORD-MP 10201, x 150; Q, elemento Pa, vista lateral externa (espécimen juvenil), muestra AB / Pa element, outer-lateral view (juvenil specimen), sample AB , x 360; R, elemento Pa, vista lateral interna (espécimen adulto), muestra AB / Pa element, inner lateral view (adult specimen), sample B , CORD-MP 10202, x 160; S, elemento Pa, vista superior (espécimen adulto), muestra AB / Pa element, upper view (adult specimen), sample AB , CORD-MP 10048, x160; T, elemento Pa, vista superior (espécimen juvenil), muestra AB / Pa element, upper view (juvenil specimen), sample AB , CORD-MP 10049, x180; U, elemento Pa, vista inferior (espécimen adulto), muestra AB / Pa element, lower view (adult specimen), sample AB , CORD-MP 10050, x 130. Todas las ilustraciones de los especímenes son microfotografías MEB / all illustration of specimens are SEM microphotographs.
Descripción. Rabdosoma recto, con la sícula ligeramente inclinada en algunos ejemplares. Los fragmentos más largos superan los 10 mm de longitud; su ancho proximal es de 0,4-0,5 mm a nivel de la apertura de la t1, aumentando progresivamente hasta 1 mm en la novena o décima teca. La sícula es delgada, con una longitud de 1,6-1,7 mm; si bien, en uno de los especímenes (figura 4.D) es de 1,9 mm, aunque esto podría deberse a cierto grado de deformación diagenética sufrida por la colonia. La apertura sicular, de 0,2-0,28 mm de ancho, presenta una virgela corta. El ápice sicular se ubica por encima de la apertura de la t1, sin lograr la apertura de la segunda teca en ninguno de los fósiles estudiados. La primera teca de 1 mm de largo, surge próxima a la apertura sicular, 0,12-0,24 mm por encima de la misma. Las tecas son tubos simples, estando frecuentemente presente una suave flexura ventral. Las aperturas son normales al eje de la teca. Se han medido 10 tecas en 10 mm en la parte proximal del rabdosoma, con una inclinación de 15° - 30° y un traslapamiento de la mitad de su longitud o algo menor.
Observaciones. Los especímenes coleccionados son escasos e incompletos. Los mismos recuerdan a P . Nudus en la forma recta del rabdosoma, la forma y espaciamiento tecal, la longitud de la t1 y el hecho de que el ápice sicular se ubique por debajo de la apertura de la segunda teca. Sin embargo, la longitud sicular es mayor en el material argentino (1-1,23 mm sensu Bjerreskov, 1975) y la inclinación tecal inferior (40° de acuerdo a Bjerreskov, 1975). Otra forma semejante es P . variabilis (Perner), la cual se diferencia de P. Nudus principalmente por presentar un ancho distal del rabdosoma inferior. El hecho de no contar con rabdosomas completos o con fragmentos distales no permite efectuar una comparación con la citada especie.
P . nudus se extiende a través de las zonas de Spirograptus turriculatus y Monoclimacis crenulata del Llandovery tardío (Telychiano) de Europa ( e.g ., Rickards, 1970; Rickards et al ., 1977; Bjerreskov, 1975). De acuerdo a Bjerreskov (1975), los especímenes de P . nudus de la Zona de grienstoniensis son más robustos que los de la Zona de turriculatus , pudiendo ser éste el horizonte al que pertenezcan los ejemplares argentinos, considerando sus dimensiones. Sin embargo, es necesario contar con una colección más amplia para establecer la verdadera identidad y edad del material estudiado. En la puna salteña, Argentina, Isaacson et al . (1976) mencionan la presencia de monográptidos del grupo de P. nudus en asociación con faunas de braquiópodos.
Procedencia. Formación La Chilca, muestra E.
Material y repositorio. Partes proximales de rabdosomas y sículas conservados como una película carbonosa fragmentaria en superficies de pelitas, CORD-PZ 23940, 23941, 23942, 23943, 23944, 23945.
Género Monograptus Geinitz, 1852
Especie tipo. Lomatoceras priodon Bronn, 1835.
Monograptus cf. priodon (Bronn, 1835)
Figuras 4.E, F, H
cf. Lomatoceras priodon Bronn, 1835, 55, 56, Pl. 1: 13.
Descripción. Rabdosoma robusto y largo, ligeramente flexuoso. Uno de los fragmentos supera los 35 mm de largo; su ancho fluctúa entre 1,2 mm en la parte más proximal y 2,25 mm en la distal. Las tecas son curvadas, con forma de gancho, más grandes y más rectas hacia distal, donde la eversión es más marcada. El traslapamiento varía a través de la longitud del rabdosoma, siendo inferior en la porción más proximal (9,5 tecas en 10 mm), aumentando en la parte media (10 tecas en 10 mm) y disminuyendo nuevamente hacia distal (9 - 9,5 tecas en 10 mm).
Observaciones. La ausencia de rabdosomas completos y la preservación deficiente de los mismos nos llevan a comparar nuestro material con M. priodon , especie con la que comparte la forma de las tecas y dimensiones del rabdosoma. Las diferencias con el citado taxón corresponden a la longitud sicular que es superior en el material descripto (1,5 mm en M. Priodon de acuerdo a Bjereskov, 1975) y el traslapamiento tecal, que es más apretado en la parte proximal del rabdosoma (13 tecas en 10 mm sensu Bjereskov, 1975) mientras que en los ejemplares estudiados es de 9,5 tecas en 10 mm.
M. priodon ha sido localizado en las zonas de S. Turriculatus y C. centrifugus (Llandovery tardío-Wenlock temprano) de Europa ( e.g ., Elles y Wood, 1911; Urbaneck, 1958; Schauer, 1971; Bjerreskov, 1975; Loydell et al ., 1998; Štorch et al ., 2002; Loydell et al ., 2003), China (Mu et al ., 1974) y Canadá (Lenz, 1982; Lenz y Kozlowska-Dawidziuk, 2001; Zhang, 1994). En Argentina esta especie fue originalmente reconocida por Stappenbeck (1910) en el cerro del Fuerte y posteriormente por Kerlleñevich (1986) y Kerlleñevich y Cuerda (1986) en la parte superior de la Formación La Chilca, unos 3 km al sur de Los Baños de Talacasto.
Procedencia. Formación La Chilca, muestras E y F.
Material y repositorio. Rabdosomas incompletos y una sícula, posiblemente referible a los mismos especímenes, conservados como una película carbonosa fragmentaria sobre pelitas, CORD-PZ 23934 A-B, 23935 A-B, 23936 A-B, 23937, 23938, 23939.
Género Stimulograptus Pribyl y Štorch, 1983
Especie tipo. Graptolithus halli Barrande, 1850.
? Stimulograptus sp.
Figuras 4.I, J
Descripción. El fragmento más delgado de rabdosoma tiene un ancho de 1,2 mm y una longitud de 10 mm. Las tecas son largas, con escaso traslapamiento y marcada retroversión apertural. Se cuentan 7,5 tecas en 10 mm. El fragmento más robusto, posiblemente una porción media o distal del rabdosoma, presenta una longitud superior a los 11 mm, con un ancho proximal de 1,25 mm y distal de 1,6 mm. Las tecas son evertidas, pero las aperturas no están tan retrovertidas como en el fragmento más delgado. El traslapamiento es mayor que en el otro rabdosoma, observándose 8,5 tecas en 10 mm.
Observaciones. La forma de las tecas evertidas y con retroversión en su parte apertural recuerda al género Stimulograptus Pribyl y Štorch (1983), pero la ausencia de espinas aperturales, características de este taxón, ligado a la escasez y deficiente preservación del material, no permiten efectuar una diagnosis a nivel genérico. Algunas medidas como el espaciamiento tecal y el ancho proximal del rabdosoma y la forma de las tecas recuerdan a especies como S . sedgwickii (Portlock) y S. halli (Barrande). Las espinas aperturales tecales en S . sedgwickii son normalmente escasas o inexistentes en la parte proximal del rabdosoma, volviéndose conspicuas hacia distal (Elles y Wood, 1911). Posiblemente a esto obedezca la ausencia de las mismas en los especímenes de la Formación La Chilca o a un problema de preservación. S. sedgwickii y S. halli son fósiles guías de las dos biozonas finales del Aeroniano británico. S . aff. halli fue citado por Brussa (en Toro, 1995) para la Formación La Chilca, sierra de Talacasto.
Procedencia y distribución. Formación La Chilca, muestra G.
Material y repositorio. Dos fragmentos de rabdosomas, conservados como una película carbonosa, CORD-PZ 23929, 23952.
Agradecimientos
Los autores agradecen el apoyo del CONICET, a través de cuyo subsidio PIP 0933/98, el desarrollo de este trabajo fue posible. Agradecemos a A. Andrada Barone del LAMENOA, Tucumán, quien asistió los estudios sobre microscopía electrónica de barrido. Nuestro reconocimiento a F.J. Zeballo por su colaboración en la preparación de las ilustraciones y en las tareas de laboratorio de micropaleontología. La lectura crítica del manuscrito original fue efectuada por los árbitros G.N. Sarmiento y E.D. Brussa.
Bibliografía
Aceñolaza, F.G. y Peralta, S.H. 1986. Lophoctenium comosum Reinhold Ritcher (traza fósil) en la Formación Los Espejos (Silúrico), Precordillera de San Juan. Primeras Jornadas sobre Geología de Precordillera, Asociación Geológica Argentina, Serie A, Monografías y Reuniones 2: 145-150. [ Links ]
Albanesi, G.L., Hünicken, M.A. y Barnes, C.R. 1998. Bioestratigrafía de conodontes de las secuencias ordovícicas del cerro Potrerillo, Precordillera Central de San Juan, R. Argentina. Academia Nacional de Ciencias (Córdoba), Actas 12: 7-72. [ Links ]
Aldridge, R.J. 1975. The stratigraphic distribution of conodonts in the British Silurian. Journal of the Geological Society , London, 131: 607-618. [ Links ]
Aldridge, R.J. y Schönlaub, H.P. 1989. Conodonts. En: C.H. Holland and M.G. Bassett (eds.), A global standard for the Silurian System. National Museum of Wales, Geological Series 9: 274-279. [ Links ]
Aldridge, R.J. y Smith, M.P. 1985. Lower Palaeozoic succession of the Welsh Borderland. Fourth European Conodont Symposium (ECOS IV) Field Excursion B Guidebook , 1-39. [ Links ]
Amos, A.J. 1954. Estructura de las formaciones paleozoicas de La Rinconada, pie oriental de la sierra Chica de Zonda, San Juan. Revista de la Asociación Geológica Argentina 9: 5-38. [ Links ]
Astini, R.A. 1992. Descripción y génesis de los bancos de oolitas ferruginosas en la base del Silúrico de la Precordillera argentina. Estudios Geológicos 48: 297-303. [ Links ]
Astini, R.A. y Maretto, H.M. 1996. Análisis estratigráfico del Silúrico de la Precordillera Central de San Juan y consideraciones sobre la evolución de la cuenca. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos, Actas 1: 351-368. [ Links ]
Astini, R.A. y Piovano, E.L. 1992. Facies de plataforma terrígena del Silúrico de la Precordillera sanjuanina. Revista de la Asociación Geológica Argentina 47: 99-110. [ Links ]
Baldis, B.A. y Aceñolaza, F.G. 1978. Trazas fósiles del Silúrico de la Precordillera. Acta Geológica Lilloana, Suplemento 14: 101-104. [ Links ]
Baldis, B.A. y Chebli, G. 1969. Estructura profunda del área central de la Precordillera sanjuanina. Cuartas Jornadas Geológicas Argentinas 1: 47-66. [ Links ]
Baldis, B.A. y Peralta, S.H. 1999. Silúrico y Devónico de la Precordillera de Cuyo y Bloque de San Rafael. En: R. Caminos, (ed.), Geología Argentina, Servicio Geológico Minero Argentino, Instituto de Geología y Recursos Minerales, Anales 29: 215-238. [ Links ]
Baldis, B.A., Beresi, M.S., Bordonaro, O. y Vaca, A. 1982. Síntesis evolutiva de la Precordillera argentina. 5° Congreso Latinoamericano de Geología (Buenos Aires) 4: 399-445. [ Links ]
Baldis, B.A., Peralta, S.H. y Uliarte, E. 1984. Geología de la quebrada Ancha y sus alrededores en área de Talacasto, Precordillera sanjuanina. 9° Congreso Geológico Argentino (Bariloche) 4: 233-245. [ Links ]
Barca, S., Ferretti, A., Massa, P. y Serpagli, E. 1992. The Hercynian Arburese tectonic unit of SW Sardinia. New stratigraphic and structural data. Rivista Italiana Paleontologia Stratigrafia 98: 119-136. [ Links ]
Barrande, J. 1850. Graptolites de Bohême . Prague: 1-74. [ Links ]
Bassler, R.S. 1925. Classification and stratigraphic use of the conodonts. Geological Society of America Bulletin 36: 218-220. [ Links ]
Benedetto, J.L. 1995. Braquiópodos del Silúrico temprano (Llandoveriano) Malvinocáfrico, Formación La Chilca, Precordillera argentina. Geobios 28: 425-457. [ Links ]
Benedetto, J.L., Ortega, G., Brussa, E.D. y Toro, B.A. 1986. Estratigrafía y fauna de la secuencia neo-ordovícica y llandoveriana del río Escondido, flanco occidental del cerro del Fuerte, provincia de San Juan. Primeras Jornadas sobre Geología de Precordillera, Asociación Geológica Argentina, Serie A, Monografías y Reuniones 2: 128-133. [ Links ]
Benedetto, J.L., Racheboeuf, P.R., Herrera, Z.A., Brussa, E.D. y Toro, B.A. 1992b. Brachiopodes et biostratigraphie de la Formation de Los Espejos, Siluro-Dévonien de la Précordillère (NW Argentine). Geobios 25: 599-637. [ Links ]
Benedetto, J.L., Sánchez, T.M. y Brussa, E.D. 1992a. Las cuencas silúricas de América Latina. En: J.C. Gutiérrez Marco, J. Saavedra y I. Rábano (eds.), Paleozoico Inferior de Ibero América , Universidad de Extremadura, 119-148. [ Links ]
Benfrika, M. 1998. Some upper Silurian - middle Devonian conodonts from the northern part of Western Meseta of Morocco: systematic and palaeogeographical relationships. Bolletino della Società Paleontologica Italiana 37: 311-319. [ Links ]
Bischoff, G.C.O. 1986. Early and Middle Silurian conodonts from midwestern New South Wales. Courier Forschungsinstitut Senckenberg 89: 1-137. [ Links ]
Bischoff, G.C.O. y Sannemann, D. 1958. Unterdevonische conodonten aus dem Frankenwald. Hessisches landesamt für bodenforschung, Notizblatt 86: 87-110. [ Links ]
Bjerreskov, M. 1975. Llandoverian and Wenlockian graptolites from Bornholm. Fossils and Strata 8: 1-93. [ Links ]
Branson, B.E. y Mehl, M.G. 1933. Conodont studies: University of Missouri Studies , 8, parts 1-4: 1-343. [ Links ]
Bronn, H.G. 1835. Lethaea geognostica . Volume 1. E. Schweizerbart, Stuttgart, 1-768. [ Links ]
Clarke, J.N. 1912. El Devoniano de la Argentina occidental. Anales del Ministerio de Agricultura, Sección Geología, Mineralogía y Minería 8: 3-15. [ Links ]
Corradini, C. y Serpagli, E. 1998. A Silurian conodont biozonation from late Llandovery to end Pridoli in Sardinia (Italy). Bolletino della Società Paleontologica Italiana 37: 255-273. [ Links ]
Cuerda, A.J. 1965. Monograptus leintwardinensis var. Incipiens Word en el Silúrico de la Precordillera. Ameghiniana 4: 171-177. [ Links ]
Cuerda, A.J. 1969. Sobre las graptofaunas del Silúrico de San Juan, Argentina. Ameghiniana 6: 223-235. [ Links ]
Cuerda, A.J. 1971. Monograpten des Unter-Ludlow aus der Vorkordillere von San Juan, Argentinien. Geologisches Jahrbuch 89: 391-406. [ Links ]
Cuerda, A.J. 1981. Graptolitos del Silúrico Inferior de la Formación Rinconada, Precordillera de San Juan. Ameghiniana 18: 241-247. [ Links ]
Cuerda, A.J. 1985. Estratigrafía y bioestratigrafía del Silúrico de San Juan (Argentina) basada en sus faunas de graptolitos. Ameghiniana 22: 233-241. [ Links ]
Cuerda, A.J., Furque, G. y Uliarte, E. 1982. Graptolitos de la base del Silúrico de Talacasto, Precordillera de San Juan. Ameghiniana 19: 239-252. [ Links ]
Cuerda, A.J., Rickards, R.B. y Cingolani, C. 1988. A new Ordovician-Silurian boundary section in San Juan Province, Argentina, and its definite graptolite fauna. Journal of the Geological Society, London, 145: 749-757. [ Links ]
Donoghue, P.C.J., Forey, P. y Aldridge, R. 2000. Conodont affinity and chordate phylogeny. Biological Review 75: 191-251. [ Links ]
Drygant, D.M. 1974. New Middle Ordovician conodonts from the North-Western Volhynia. Paleontologicheskii Sbornik 11: 54-58 (en ucraniano). [ Links ]
Dzik, J. 1976. Remarks on the evolution of Ordovician conodonts. Acta Palaeontologica Polonica 21: 395-455. [ Links ]
Dzik, J. y Drygant, D. 1986. The apparatus of panderodontid conodonts. Lethaia 19: 133-141. [ Links ]
Elles, G.L. y Wood, E.M.R. 1901-1918. A monograph of British graptolites. Palaeontographical Society of London, Monograph , pp. 1-539. [ Links ]
Epstein, A.G., Epstein, J.B. y Harris, L.D. 1977. Conodont color alteration - an index to organic metamorphism. United States Geological Survey, Professional Paper 995: 1-27. [ Links ]
Ethington, R.L. 1959. Conodonts of the Ordovician Galena Formation. Journal of Paleontology 33: 257-292. [ Links ]
Ferretti, A., Corradini, C. y Serpagli, E. 1998. Wenlock-Ludlow conodonts from Perd´e Fogu (Fluminimaggiore). Giornale di Geologia, serie 3° 60: 156-167. [ Links ]
Furque, G. y Cuerda, A.J. 1979. Precordillera de La Rioja, San Juan y Mendoza. Geología Regional Argentina, Academia Nacional de Ciencias (Córdoba) 1: 455-522. [ Links ]
Geinitz, H.B. 1852. Die Vertsteinerungen der Grauwacken-Formation (Die graptolithen). Verlag Wilhelm Engelmann, Leipzig, 1-58 pp. [ Links ]
Hass, W.H. 1959. Conodonts from the Chappel Limestone of Texas. U.S. Geological Survey Professional Paper 294J: 365-400. [ Links ]
Heim, A. 1948. Observaciones tectónicas en La Rinconada, Precordillera de San Juan. Boletín Dirección de Minería y Geología , Buenos Aires, 64: 5-38. [ Links ]
Heim, A. 1952. Estudios tectónicos de la Precordillera de San Juan: Los ríos San Juan, Jáchal y Huaco. Revista de la Asociación Geológica Argentina 7: 11-70. [ Links ]
Hünicken, M.A. 1975. Sobre el hallazgo de conodontes en el Silúrico de Lomas de los Piojos, dpto. Jáchal, pcia. San Juan. 1° Congreso Argentino de Paleontología y Bioestratigrafía (Tucumán), Actas 1: 283-291. [ Links ]
Hünicken, M.A. y Sarmiento, G.N. 1988. Conodontes Ludlovianos de la Formación Los Espejos, Talacasto, provincia de San Juan, R. Argentina. 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 3: 225-233. [ Links ]
Isaacson, P.E., Antelo, B. y Boicot, A.J. 1976. Implications of a Llandovery (Early Silurian) brachiopod fauna from Salta Province. Journal of Paleontology 50: 1103-1112. [ Links ]
Jaekel, O. 1889. Ueber das Alter des sogennanten Graptolithengesteins. Zeitschrift der Deutsches Geologischen Gesellschaft 41: 653-716. [ Links ]
Jeppsson, L. 1969. Notes on some Upper Silurian multielement conodonts. Geologiska Föreningers Stockholm Förhandlingar 91: 12-24 . [ Links ]
Jeppsson, L. 1971. Element arrangement in conodont apparatuses of Hindeodella type and in similar forms. Lethaia 4: 101-123. [ Links ]
Jeppsson, L. 1972. Some Silurian conodont apparatuses and possible conodont dimorphism. Geologica et Palaeontologica 6: 51-69. [ Links ]
Jeppsson, L. 1974 [1975]. Aspects of late Silurian conodonts. Fossils and Strata 6: 1-79. [ Links ]
Jeppsson, L. 1990. An oceanic model for lithological and faunal changes, tested on the Silurian record. Journal of the Geological Society , London, 147: 663-674. [ Links ]
Jeppsson, L. 1997. A new latest Telychian, Sheinwoodian and Early Homerian (Early Silurian) standard conodont zonation. Transactions of the Royal Society of Edinburgh: Earth Sciences 88: 91-114. [ Links ]
Jeppsson, L. 1998. Silurian oceanic events: summary of general characteristics. En: E. Landing and M.E. Johnson (eds.), Silurian cycles: linkages of dynamic stratigraphy with atmospheric, oceanic and tectonic changes. James Hall Centenial Volume, New York State Museum Bulletin 491: 239-257. [ Links ]
Jeppsson, L. y Aldridge, R.J. 2000. Ludlow (late Silurian) oceanic episodes and events. Journal of the Geological Society , London, 157: 1137-1148. [ Links ]
Kayser, E. 1876. Ueber Primordiale und Untersilurische Fossilien aus der Argentinischen Republik. Cassel. Beiträge zur Geologie und Palaeontologie der Argentinischen Republik , 2-Palaeontologischer Theil, 1 Abteilung, Palaeontograph. Suppl. 3: 1-33. [ Links ]
Keidel, J. 1921. Observaciones geológicas en la Precordillera de San Juan-Mendoza. Anales del Ministerio de Agricultura, Sección Geología, Mineralogía y Minería 15: 7-102. [ Links ]
Keidel, J. 1938. Uber die "Gondwaniden" Argentiniens. Geologische Rundschau 30: 148-249. [ Links ]
Kerlleñevich, S.C. 1986. Localidades con Monograptus priodon (Graptolithina) en la Formación La Chilca, Precordillera de San Juan. Primeras Jornadas sobre Geología de Precordillera, San Juan , Asociación Geológica Argentina, Serie A, Monografías y Reuniones 2: 139-144. [ Links ]
Kerlleñevich, S.C. y Cuerda, A.J. 1986. Monograptus priodon (Bronn) (Graptolithina) en la Formación La Chilca, Precordillera de San Juan, Argentina. Ameghiniana 23: 119-126. [ Links ]
Klapper, G. 1981. Systematic descriptions: Families Distomodontidae, Kockelellidae. En: R.A. Robison (ed)., Treatise on Invertebrate Paleontology , Pt. W, Suppl. 2, Conodonta, Geological Society of America and University of Kansas, 1-202. [ Links ]
Lapworth, C. 1873. On an improved classification of the Rhabdophora. Geological Magazine 10: 500-504, 555-560. [ Links ]
Lapworth. C. 1880. On new British graptolites. Annals Magazine Natural History 5: 149-177. [ Links ]
Lehnert, O. 1995. Geodynamic processes in the Ordovician of the Argentine Precordillera: New biostratigraphic contraints. En: J. Cooper, M.L. Droser y S.C. Finney (eds.), 7 th International Symposium on the Ordovician System. The Pacific section Society for Sedimentary Geology (SEPM), Fullerton, Book 77: 75-79. [ Links ]
Lehnert, O., Bergström, S.M., Benedetto, J.L. y Vaccari, N.M. 1999. First record of Lower Silurian conodonts from South America: biostratigraphic and palaeobiogeographic implications of Llandovery conodonts in the Precordillera of Argentina. Geological Magazine 136: 119-131. [ Links ]
Lenz, A.C. 1982. Llandoverian graptolites of the Northern Canadian Cordillera: Petalograptus, Cephalograptus, Rhaphidograptus, Dimorphograptus, Retiolitidea, Monograptidea. Life Sciences Contribution, Royal Ontario Museum 130: 1-154. [ Links ]
Lenz, A.C., Cuerda, A.J. y Peralta, S.H. 2003. Graptolites from the Upper Llandovery of the Talacasto area, Central Precordillera, San Juan, Argentina. Journal of Paleontology 77: 1198-1202. [ Links ]
Lenz, A.C. y Kozlowska-Dawidziuk, A. 2001. Upper Wenlock (Silurian) graptolites of Arctic Canada: pre-extinction, lundgreni Biozone fauna. Palaeontographica Canadiana 20: 1-61. [ Links ]
Lindström, M. 1970. A suprageneric taxonomy of the conodonts. Lethaia 3: 427-445. [ Links ]
Link, A.G. y Druce, E.C. 1972. Ludlowian and Gedinnian conodont stratigraphy of the Yass basin, New South Wales. Bulletin Bureau of Mineral Resources , Australia, 134: 1-136. [ Links ]
Loydell, D.K. vs Jeppsson, L. y Aldridge, R.J. 2000. Discussion on Ludlow (late Silurian) oceanic episodes and events. Journal of the Geological Society , London, 157: 1137-1148. [ Links ]
Loydell, D.K., Kaljo, D. y Mannik, P. 1998. Integrated biostratigraphy of the lower Silurian of the Ohesaare core, Saaremaa, Estonia. Geological Magazine 135: 769-783. [ Links ]
Loydell, D.K., Männik, P. y Nestor, V. 2003. Integrated biostratigraphy of the Lower Silurian of the Aizpute-41 core, Latvia. Geological Magazine 140: 205-229. [ Links ]
Lozano, B. y Hünicken, M.A. 1990. Conodonts and biostratigraphy of the San Juan Formation (Arenigian-Llanvirnian) in the Quebrada de Talacasto, Ullum Department, San Juan Province, Argentina. En: M.A. Hünicken (ed.), 1 Latin American Conodont Symposium, LACON I, Academia Nacional de Ciencias (Córdoba), abstracts, pp. 101-102. [ Links ]
Männik, P. y Malkowski, K. 1998. Silurian conodonts from the Golpad Core, Poland. En: H. Szaniawski (ed.), Proceedings of the Sixth European Conodont Symposium (ECOS VI). Palaeontologica Polonica 58: 141-151. [ Links ]
Marchese, H.G. 1972. Sedimentología de la Formación San Juan (Ordovícico) y del conglomerado basal de la Formación Los Espejos (Silúrico?) en la quebrada de Talacasto, provincia de San Juan, República Argentina. Revista de la Asociación Geológica Argentina 27: 215-222. [ Links ]
Mawson, R. 1986. Early Devonian (Lochkovian) conodont faunas from Windellama, New South Wales. Geologica et Palaeontologica 20: 39-71. [ Links ]
Miller, C.G. y Aldridge, R.J. 1993. The taxonomy and apparatus structure of the Silurian distomodontid conodont Coryssognathus Link and Druce, 1972. Journal of Micropalaeontology 12: 241-255. [ Links ]
Mu, En-zhi et al . 1974. A handbook of stratigraphy and palaeontology of southwest China. Science Press , Nanjing, pp. 1-454. [ Links ]
Nowlan, G. 1995. Left hand column for correlation charts. Silurian Times 3: 7-8. [ Links ]
Ortiz, A. y Zambrano, I.J. 1981. La provincia geológica Precordillera Oriental. 8° Congreso Geológico Argentino (San Luis) 3: 59-74. [ Links ]
Padula, E.L., Rolleri, E.O., Mingramm, A.R.G., Criado Roqué, P., Flores, M.A. y Baldis, B.A. 1967. Devonian of Argentina. International Symposium on the Devonian System , Proceedings , Calgary 2: 165-199. [ Links ]
Pander, C.H. 1856. Monographie der fossilen fische des silurischen systems der russisch-baltischen gouvernements. Buchdrukerei der kaiserlichen Akademie der Wissenschaften, Saint Petersburg pp. 1-91. [ Links ]
Peralta, S.H. 1984. Ludlowiano en la Precordillera Oriental sanjuanina. 9° Congreso Geológico Argentino (Bariloche), 4: 296-304. [ Links ]
Peralta, S.H. 1986a. Graptolitos del Llandoveriano inferior clástico del pie oriental de la sierra de Villicum, Precordillera Oriental de San Juan. Primeras Jornadas sobre Geología de Precordillera, Asociación Geológica Argentina, Serie A, Monografías y Reuniones 2: 134-138. [ Links ]
Peralta, S.H. 1986b. Graptofauna del Llanvirniano Inferior (Ordovícico) y Ludlowiano Inferior (Silúrico), en la Formación Rinconada, Precordillera Oriental de San Juan. 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), 1: 59-66. [ Links ]
Peralta, S.H. 1990. Silúrico de la Precordillera de San Juan, Argentina. 11° Congreso Geológico Argentino (San Juan), Relatorio , pp. 48-64. [ Links ]
Peralta, S.H. 1993. Estratigrafía y consideraciones paleoambientales de los depósitos marino-clásticos eopaleozoicos de la Precordillera Oriental de San Juan. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza) 1: 128-137. [ Links ]
Peralta, S.H. 1998. Storm-dominated shelf sequences and related graptolite faunas in the Silurian of the Talacasto section, San Juan Precordillera, Argentina. En: J.C. Gutiérrez-Marco y I. Rábano (eds.), 6° International Graptolite Conference and 1998 Field Meeting of the IUGS, Subcommission on Silurian Stratigraphy . Temas Geológico-Mineros ITGE, Madrid, 23: 115-118. [ Links ]
Peralta, S.H. 2003. Silurian of the San Juan Precordillera, western Argentina: stratigraphic framework. En: G. Ortega y G.F. Aceñolaza (eds.), Proceedings of the 7th International Graptolite Conference and Field Meeting of the International Subcommission on Silurian Stratigraphy , INSUGEO, Serie Correlación Geológica, 18: 151-155. [ Links ]
Peralta, S.H. y Carter, C.H. 1990. Facies de plataforma e icnofacies asociadas de la Formación Tambolar (Silúrico) en su localidad tipo, Precordillera Central sanjuanina. 3° Reunión Argentina de Sedimentología , Actas : pp. 339-344. [ Links ]
Peralta, S.H., Uliarte, E.R. y Lanzilotta, I. 1986. Estudio de los depósitos sedimentarios ferruginosos del Silúrico de la Precordillera de San Juan. Primeras Jornadas sobre Geología de la Precordillera, Asociación Geológica Argentina, Serie A, Monografías y Reuniones, Actas 2: 301-306. [ Links ]
Peralta, S.H., Albanesi, G.L. y Ortega, G. 2003. Talacasto, La Invernada, and Jáchal River sections, Precordillera of San Juan Province. Ordovician and Silurian of the Precordillera, San Juan, Argentina. Field trip guide, INSUGEO, Miscelánea 10: 81-112. [ Links ]
Pöthe de Baldis, E.D. 1981. Paleomicroplancton y miosporas del Ludloviano inferior de la Formación Los Espejos en el perfil de Los Azulejitos, en la provincia de San Juan, República Argentina. Revista Española de Micropaleontología 13: 231-265. [ Links ]
Pribyl, A. y Štorch, P. 1983. Monograptus ( Stimulograptus ) subgen. n. (Graptolites) from the Lower Silurian of Bohemia. Vestnik Ustredního ústavu geologického 58: 221-224.
Purnell, M.A.P., Donoghue, P.C.J. and Aldridge, R.J. 2000. Orientation and anatomical notation in conodonts. Journal of Paleontology 74: 113-122. [ Links ]
Ramos, V.A., Malanca, S. y Martel, M.H. 1978. Estratos con graptolitos suprayacentes a la Formación San Juan en la quebrada de Talacasto. Reunión Nacional Paleozoico Argentino (Tucumán): pp. 115-116. [ Links ]
Rickards, R.B. 1970. The Llandovery (Silurian) graptolites of the Howgill Fells, Northern England. Palaeontographical Society of London, Monograph, pp. 1-108. [ Links ]
Rickards, R.B., Brussa, E.D., Toro, B.A. y Ortega, G. 1996. Ordovician and Silurian graptolite assemblages from Cerro del Fuerte, San Juan Province, Argentina. Geological Journal 31: 101-122. [ Links ]
Rickards, R.B., Hutt, J.E. y Berry, W.B.N. 1977. Evolution of the Silurian and Devonian graptoloids. Bulletin of the British Museum (Natural History), Geology 28: 1-120. [ Links ]
Rhodes, F.H.T. 1953. Some British Lower Paleozoic conodont faunas. Philosophical Transactions Royal Society London Series B 237: 261-334. [ Links ]
Rolleri, E.O. 1947. [ Estudio geológico de la quebrada de Talacasto y zonas adyacentes, provincia de San Juan . Universidad Nacional y Museo de La Plata. Tesis Doctoral, inédita.]. [ Links ]
Rubinstein, C. 1992a. Palinología del Silúrico Superior (Formación Los Espejos) de la quebrada de Las Aguaditas, Precordillera de San Juan, Argentina. Ameghiniana 29: 231-248. [ Links ]
Rubinstein, C. 1992b. Palinología del Silúrico Superior (Formación Los Espejos) de Loma de Los Piojos, Precordillera de San Juan, Argentina. Ameghiniana 29: 287-303. [ Links ]
Rubinstein, C. 1995. Acritarchs from Upper Silurian of Argentina: their relationships with Gondwana. Journal of South American Earth Sciences 8: 103-115. [ Links ]
Rubinstein, C. y Brussa, E.D. 1999. A palynomorph and graptolite biostratigraphy of the Central Precordillera Silurian Basin, Argentina. Bolletino della Società Italiana 38: 257-266. [ Links ]
Sánchez, T.M., Benedetto, J.L. y Astini, R.A. 1993. Eventos de recambio faunístico en secuencias depositacionales del Ordovícico tardío-Devónico temprano de la Precordillera de San Juan, Argentina. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza) 2: 281-288. [ Links ]
Sánchez, T.M., Waisfeld, B.G. y Benedetto, J.L. 1991. Lithofacies, taphonomy, and brachiopod assemblages in the Silurian of western Argentina. A review of Malvinokafric Realm communities. Journal of South American Earth Sciences 4: 307-329. [ Links ]
Sansom, I.J., Armstrong, H.A. y Smith, P. 1994. The apparatus architecture of Panderodus and its implications for coniform conodont classification. Palaeontology 37: 781-799. [ Links ]
Sarmiento, G.N., Méndez-Bedia, I., Aramburu, C., Arbizu, M., y Truyols J. 1994. Early Silurian conodonts from the Cantabrian Zone, NW Spain. Geobios 27: 507-522. [ Links ]
Sarmiento, G.N., Piçarra, J.M., Rebelo, J.A., Robardet, M., Gutiérrez Marco, J.C., Storch, P. y Rábano, I. 1999. Le Silurien du synclinorium de Moncorvo (NE du Portugal): biostratigraphie et importance paléogéographique. Geobios 32: 749-767. [ Links ]
Schauer, M. 1971. Biostratigraphie und taxonomie der Graptolithen des tieferen Silurs unter besonderer Berücksichtigung der tektonischen deformation. Freiberger ForschHft. C, 273, Paläontologie, 1-185. [ Links ]
Serpagli, E. y Corradini, C. 1998. Taxonomy and evolution of Kockelella (Conodonta) from the Silurian of Sardinia (Italy). Bollettino della Società Paleontologica Italiana 37: 275-298. [ Links ]
Serpagli, E., Corradini, C. y Olivieri, R. 1997. Occurrence and range of the Silurian conodont Coryssognathus dubius (Rhodes, 1953) in Southern Sardinia. Bollettino della Società Paleontologica Italiana 35: 239-243. [ Links ]
Serpagli, E., Corradini, C. y Ferretti, A. 1998. Conodonts from a Ludlow-Pridoli section near the Silus village. Giornale di Geologia, ser. 3° 60: 104-111. [ Links ]
Simpson, A.J. y Talent, J.A. 1995. Silurian conodonts from the headwaters of the Indi (upper Murria) and Buchanan rivers, southeastern Australia, and their implications. En: R. Mawson and J.A. Talent (eds.), Contributions to the First Australian Conodont Symposium (AUSCOS I) held in Sydney, Australia, Courier Forschungsinstitut Senckenberg 182: 79-215. [ Links ]
Smith, M.P., Briggs, D.E.G. and Aldridge, R.J. 1987. A conodont animal from the Lower Silurian of Wisconsin, Palaeobiology of conodonts. En: R.J. Aldridge (ed.), Palaeobiology of Conodonts, British Micropalaeontology Society Series. Ellis Horwood , Chichester, pp. 1-180. [ Links ]
Stappenbeck, R. 1910. La Precordillera de San Juan y Mendoza. Anales del Ministerio de Agricultura, Sección Geología, Minería y Mineralogía 4: 1-187. [ Links ]
Stauffer, C.R. 1930. Conodonts from the Decorah shale. Journal of Paleontology 4: 121-128. [ Links ]
Stelzner, A. 1873. Mitteinlungen an Professor H.B. Geinitz. Ubre seine Reise durch die Argentinischen Provinzen San Juan und Mendoza und die Cordillere zwischen dem 31° und 32° S. Neus Jahrbuch für Mineralogie, Geologie und Palaeontologie, pp. 726-744. [ Links ]
Stone, J., 1987. Review of investigative techniques used in the study of conodonts. En: R.L. Austin (ed.), Conodonts: Investigative Techniques and Applications, Ellis Horwood Ltd. , Chichester, pp. 17-34. [ Links ]
torch, P., Serpagli, E. y Barca, S. 2002. Silurian graptolites of the spiralis and lapworthi biozones (upper Telychian, Llandovery) in the Sulcis area, SW Sardinia. Bollettino della Società Paleontologica Italiana 41: 97-107. [ Links ]
Suess, E. 1851. Ueber böhmische Graptolithen. Naturwissenschaft Abhandlung von W. Haidinger 4: 89-134. [ Links ]
Sweet, W.C. 1981. Macromorphology of elements and apparatuses. En: R.A. Robison (ed.), Treatise on Invertebrate Paleontology, Pt. W, Suppl. 2, Conodonta, Geological Society of America and University of Kansas, pp. 1-202. [ Links ]
Sweet, W.C. 1988. The Conodonta: morphology, taxonomy, paleoecology, and evolutionary history of a long-extinct animal phylum. Clarendon Press , Oxford, pp. 1-211. [ Links ]
Sweet, W.C. y Donoghue, P.C.J. 2001. Conodonts: past, present, future. Journal of Paleontology 75: 1174-1184. [ Links ]
Sweet, W.C. y Schönlaub, H.P. 1975. Conodonts of the genus Oulodus Branson and Mehl, 1933. Geologica et Palaeontologica 9: 41-59. [ Links ]
Toro, B.A. 1995. Primer hallazgo del Silúrico (Llandoveriano) en la Cordillera Oriental, Provincia de Jujuy, Argentina. Ameghiniana 32: 375-384. [ Links ]
Turner, J.C.M. 1960. Faunas graptolíticas de América del Sur. Revista de la Asociación Geológica Argentina 14: 5-180. [ Links ]
Urbaneck, A.1958. Monograptidae from erratic boulders of Poland. Palaeontologica Polonica 9: 1-105. [ Links ]
Vannier, J.M.C., Racheboeuf, P.R. y Benedetto, J.L. 1995. Silurian-early Devonian ostracodes from South America (Argentina, Bolivia): preliminary investigations. Journal of Paleontology 752-772. [ Links ]
Viira, V. 1998. Late Silurian conodont biostratigraphy in the northern East Baltic. Bolletino della Società Paleontológica Italiana 37: 299-310. [ Links ]
Volkheimer, W., Pothe de Baldis, E.D. y Baldis, B.A. 1980. Quitinozoos de la base del Silúrico de la sierra de Villicum (provincia de San Juan, Argentina). Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 2: 121-135. [ Links ]
Waisfeld, B.G., Toro, B.A. y Brussa, E.D. 1988. Trilobites silúricos de la Formación Los Espejos, sector occidental del cerro del Fuerte, Precordillera de San Juan, Argentina. Ameghiniana 25: 305-320. [ Links ]
Walliser, O. 1957. Conodonten aus dem oberen Gotlandium Deutschlands und der Karnischen Alpen. Notizblatt del Hessischen Landesamtes für Bodenforschung zu Wiesbaden 85: 28-52. [ Links ]
Walliser, O. 1964. Conodonten des Silurs. Abhandlungen des Hessischen Landesamtes für Bodenforschung zu Wiesbaden 41: 1-106. [ Links ]
Zhang, S. y Barnes, C.R. 2002. A new Llandovery (early Silurian) conodont biozonation and conodonts from the Becscie, Merrimack, and Gun River formations, Anticosti Island, Québec. Journal of Paleontology 76: 1-46. [ Links ]
Zhang, Yuan-dong. 1994. Wenlock isolated graptolites from Cornwallis Island, Arctic Canada. En: Chen Xu, Erdtmann, B-D y Ni Yu-nan, Graptolite Research Today, Nanjing University Press , pp. 113-127. [ Links ]
Recibido: 12 de febrero de 2005.
Aceptado: 26 de mayo de 2005.

