










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.1 Buenos Aires ene./mar. 2006
Ostrácodos recientes de la llanura pampeana, Buenos Aires, Argentina: ecología e implicancias paleolimnológicas
Cecilia Laprida1
1 Laboratorio de Micropaleontología, Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Intendente Güiraldes 2160, Ciudad Universitaria, 1428 Ciudad de Buenos Aires, Argentina. chechu@gl.fcen.uba.ar
Resumen. Veinte especies de ostrácodos no marinos fueron recolectadas de la Pampa bonaerense. Simultáneamente se midieron parámetros ambientales y propiedades químicas de las aguas con el objetivo de determinar qué condiciones ambientales prefieren las distintas especies y qué factores determinan su distribución. Los estudios sugieren que la mayor parte de las especies prefieren una calidad de agua particular en términos de salinidad e iones dominantes. Los ostrácodos que habitan ambientes estuarinos viven exclusivamente en aguas cloruradas sódicas, de salinidades oligo-polihalinas con amplias variaciones diarias de salinidad. Los ostrácodos que habitan bañados y charcas semipermanentes viven casi exclusivamente en aguas oligohalinas HCO3-/Na+ dominadas. Por último, los ostrácodos que habitan en las lagunas permanentes mayoritariamente oligohalinas pueden vivir en más de un tipo de agua, sean éstas Cl-/Na+ o HCO3 -/Na+, que son los estadíos entre los cuales oscilan las lagunas pampásicas en función del balance entre la evaporación y la precipitación. Adicionalmente, son considerados otros factores tales como los modos reproductivos, la disponibilidad del alimento y las presiones de predación. Estos resultados autorizan el uso de las asociaciones fósiles de ostrácodos no marinos de la pampa bonaerense como indicadores de las condiciones paleoclimáticas del Cuaternario tardío.
Abstract. Recent ostracodes from the pampas, Buenos Aires, Argentina: ecology and paleolimnological implications . Twenty non-marine ostracod species were collected from the pampas of Buenos Aires Province. Water parameters were analyzed in order to determine what environmental conditions each species prefers and which factor affects their occurrence. Studies suggest that most species prefer waters containing particular salinity and solute types. Marginal estuarine ostracods can only survive in oligo-polihaline Cl-/Na+ dominated waters with dayly fluctuating salinities. Freshwater ostracods living in temporary ponds can only survive in oligohaline HCO3-/Na+ dominated waters. Finally, ostracods inhabiting permanent lakes can live in more than one solute type (Cl-/Na+ or HCO3-/Na+), but these solute type variations must be gradual, related with the balance between evaporation and precipitation. Reproductive modes, trophic level and predation are also taking into account. These conclusions encourage the use of fossil non marine ostracods from the Pampas of Buenos Aires as valuable tools to reconstruct late quaternary paleoclimatic conditions.
Key words. Recent Ostracoda; Distribution; Ecology; Pampa Deprimida Plains.;Argentina.
Palabras clave. Ostrácodos recientes; Distribución; Ecología; Pampa Deprimida, Argentina.
Introducción
Conocidos desde el Devónico Superior, los ostrácodos no marinos son frecuentes en sedimentos cuaternarios de origen continental (Swain, 1990). Poseen un caparazón calcáreo bivalvo de pequeño tamaño que presenta un alto potencial de fosilización. Son excelentes indicadores paleoecológicos, sensibles a las variaciones de los parámetros físico-químicos del medio. El estudio de asociaciones fósiles permite la reconstrucción de los paleoambientes continentales y, dado que éstos son climáticamente controlados, los ambientes reconstruidos pueden ser interpretados en términos paleoclimáticos (Forester,1987, 1991; De Deckker y Forester, 1988; Carbonel et al., 1988; Forester y Smith, 1992; Curry,1999). Sin embargo, la utilización de los ostrácodos como indicadores paleoambientales requiere de una base de datos que relacione la distribución de las especies actuales con datos físicos, químicos y climáticos propios de los hábitats que ocupan, de modo de extrapolar dichos datos a las asociaciones fósiles.
Las investigaciones sobre ostrácodos actuales de ambientes continentales son relativamente escasas en la Argentina. Si bien existen algunos trabajos en los cuales se describen y/o citan especies actuales (Daday, 1902; Mèhes, 1914; Behm, 1934; Ferguson, 1967; Ramírez, 1967, entre otros), las investigaciones más metódicas se efectuaron sobre fauna cuaternaria (Whatley y Cusminsky, 2000; Cusminsky y Whatley, 1996; Zabert y Herbst, 1986; Bertels y Martínez, 1997; Ferrero, 1996; Laprida et al ., 2004). Existen trabajos más recientes que consideran simultáneamente fauna actual y ciertos parámetros ambientales (Schwalb et al ., 2002; César et al ., 2004). Sin embargo, no ha habido en la Pampa Deprimida una recolección sistemática de datos en un área lo suficientemente extensa de modo de examinar la relación entre la distribución de las especies y los principales parámetros ambientales.
La gran diversidad de ambientes acuáticos de la Pampa Deprimida brinda una excelente oportunidad para el estudio de especies con requerimientos ecológicos diferentes. En este trabajo se muestrearon diversos cuerpos de agua con el objetivo de determinar la ocurrencia de las especies en los distintos hábitats y los factores que la determinan. Con la presente contribución se intenta obtener datos que permitan el uso de los ostrácodos de ambientes no-marinos en la interpretación de paleoambientes cuaternarios de la Pampa Deprimida, una de las áreas de mayor interés para el desarrollo de estudios paleoclimáticos (Iriondo, 1994) y donde la escasez de información limita las potencialidades de reconstrucción paleoclimática. Se intenta básicamente determinar: 1) qué especies habitan los distintos ambientes; 2) qué condiciones ambientales prefieren las diversas especies; 3) cuáles son los principales factores que determinan su distribución y 4) qué especies pueden ser utilizadas como potenciales indicadoras en estudios paleoclimáticos.
Materiales y métodos
Se muestrearon 15 cuerpos de agua incluyendo lagunas permanentes, charcas, bañados, arroyos, canales y humedales en la Pampa Deprimida, Buenos Aires, entre enero de 1994 y julio de 1995, y en enero de 2002 (figura 1). Consideramos que, en el caso de los hábitats permanentes, la presencia de una especie no es resultado únicamente de los factores ambientales medidos en el momento del muestreo, sino de su variabilidad en el corto y mediano plazo. Por eso, de acuerdo con Mezquita et al . (1999), los sitios fueron muestreados una única vez, aunque algunos fueron visitados en más de una oportunidad con el objeto de analizar su variabilidad. Este único muestreo restringe las posibilidades de hallazgo de especies poco numerosas en el momento de muestreo por poseer sus poblaciones un carácter estacional, lo que resulta una limitación del método. En algunas estaciones, la temperatura, el pH y la concentración de oxígeno disuelto del agua (DO) fueron medidos in situ . Los análisis químicos (total de sólidos disueltos -TDS- e iones principales) fueron efectuados por el Laboratorio de Análisis de Trazas (INQUIMAE, FCEN, UBA) y el Limnological Research Center (LRC), Universidad de Minnesota, EE.UU. Cuando estas mediciones no pudieron ser efectuadas, se consideraron los datos disponibles en investigaciones previas. En los cuadros 1 y 2 se consignan los datos obtenidos y/o recopilados para cada sitio. Para la caracterización hidroquímica de las aguas se efectuaron diagramas de Maucha y se siguió la terminología propuestas por Dangavs (1988).

Figura 1. Mapa de ubicación mostrando las localidades estudiadas en la llanura pampeana bonaerense / map showing the location of the samples examined from Pampa plain of Buenos Aires.

Cuadro 1. Datos hidroquímicos y ambientales obtenidos de los sitios muestreados y recopilados de ambientes no marinos de carácter permanente de la Pampa bonaerense. Los datos medidos o recopilados incluyen: temperatura (T), oxígeno disuelto (DO), salinidad (TDS), pH, materia orgánica (MO) e iones mayoritarios. TDS, DO, MO e iones mayoritarios expresados en mg/L, salvo indicación específica / hydrochemical and environmental data collected during sampling and compiled from permanent pampa environments. Water parameters measured include temperature (T), dissolved oxygen (DO), salinity (TDS), pH, organic matter (MO) and major solutes. TDS, DO, MO and solute composition reported in mg/L save specific indication.
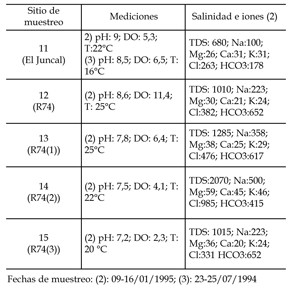
Cuadro 2. Datos hidroquímicos y ambientales obtenidos de los sitios muestreados de ambientes semipermanentes de la Pampa bonaerense. Los datos medidos incluyen: temperatura (T), oxígeno disuelto (DO), salinidad (TDS), pH, materia orgánica (MO) e iones mayoritarios. TDS, DO, MO e iones mayoritarios expresados en mg/L, salvo indicación específica / hydrochemical and environmental data collected during sampling from semipermanent pampa environments. Water parameters measured include temperature (T), dissolved oxygen (DO), salinity (TDS), major solute composition and pH. DO, TDS and solute composition are reported in mg/L.
Los muestreos fueron efectuados desde la costa mediante una red con copo colector de 1 dm3 (30 cm de diámetro; malla de 58 µm). Se muestrearon los primeros centímetros de la interfase agua/sedimento así como la vegetación superficial de acuerdo a lo propuesto por Moguilevsky y Whatley (1995). El material fue narcotizado con etanol e inmediatamente fijado con formaldehído 4% con tetraborato de sodio 2%. Las lagunas de Chascomús y del Monte fueron muestreadas en enero de 2002 mediante un sacatestigo tipo gravity core provisto por el LCR. Una vez en el laboratorio las muestras fueron lavadas con agua corriente sobre tamiz de 63 µm. En el caso de las muestras tomadas mediante sacatestigo, se procesaron aproximadamente 5 cm3 de las 3 muestras superiores (equidistancia del submuestreo 1,5 cm).
Dada la imposibilidad de estandarizar los tamaños de las muestras, los datos de abundancia presentados son de índole cualitativa. Se consideraron exclusivamente ejemplares adultos vivos al momento del muestreo, completos y con las partes blandas en perfecto estado. Dado que este trabajo tiene principalmente una aplicación paleolimnológica, la determinación específica se basó en la morfología de las partes duras, las únicas que superan la barrera tafonómica. Los ejemplares estudiados se hallan depositados en el Repositorio del Laboratorio de Micropaleontología, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, bajo los números FCEN- LM N° 2900 al 2919. En el apartado sistemático, cuando se hace mención al material estudiado, se indica el número de valvas y caparazones ilustrados y/o depositados en dicho repositorio y no al total de individuos recuperados, cuya abundancia relativa en cada sitio de muestreo se indica en el cuadro 3.

Cuadro 3. Distribución y abundancia de las especies de ostrácodos en los sitios relevados. XXX= muy abundante; XX= abundante; X= escaso / distribution and abundance of the Ostracoda by sample. XXX= very abundant; XX= abundant; X= scarce.
Área de estudio
El área de estudio se encuentra en el sector oriental de la Pampa Deprimida (figura 1), incluyendo la cuenca inferior del río Salado y el litoral de la bahía de Samborombón. Fuera de esta área, la laguna de Monte está ubicada en el extremo suroccidental de la Pampa Deprimida.
La Pampa Deprimida presenta clima húmedo a subhúmedo, mesotermal, con un gradiente de sequedad creciente hacia el oeste. Las temperaturas medias anuales oscilan entre 13°C y 16°C, aumentando en sentido sur-norte. Para enero, el mes más cálido, la temperatura media varía entre 21°C y 23°C, y para julio, el mes más frío, entre 7°C y 9°C. Las precipitaciones varían entre 850 mm anuales en el sur, a 1000 mm en el norte y disminuyen de este a oeste (Sala, 1975). Los inviernos son húmedos con excesos hídricos y anegamientos frecuentes como consecuencia de la baja evapotranspiración. Estos excesos disminuyen hacia el oeste y suroeste (Vega et al ., 1995). En los veranos, si bien la precipitación suele ser mayor, hay un pronunciado déficit de humedad del suelo como consecuencia de la mayor evapotranspiración y, en ciertos períodos, como consecuencia de las sequías. Esta alternancia de períodos húmedos y secos se refleja rápidamente en los recursos hídricos (Sala, 1975).
Desde el punto de vista fisiográfico, el sector oriental de la Pampa Deprimida se caracteriza por ser una zona baja, de relieve poco pronunciado. La red de drenaje no está claramente definida y existe una marcada desproporción entre la abundancia de cuerpos lénticos y la escasez de cursos fluviales. En este contexto, la presencia y el comportamiento de las aguas superficiales son el resultado de la interacción entre factores climáticos, morfológicos e hidrogeológicos (Sala, 1975; Miretzky, 2001).
Sitios de muestreo
Los sitios de muestreo se indican con números en la figura 1. Para los muestreos se seleccionaron ambientes lóticos y lénticos, permanentes (cuadro 1) y temporarios (cuadro 2).
Ambientes lóticos permanentes
Sitio n° 1. Punta Rasa . Se encuentra en el extremo sur de la bahía de Samborombón y constituye el límite austral del Río de la Plata. Es un ambiente estuarino de transición entre las aguas del río y el océano Atlántico. El rasgo más conspicuo es la porción distal de una espiga de barrera detrás de la cual se define una planicie de mareas surcada por canales activos. La salinidad y el pH presentan grandes variaciones dependiendo de los factores meteorológicos y de la fuerza relativa de la corriente del río y las mareas. Las muestras se extrajeron de la ría de Ajó ubicada al oeste de la espiga.
Sitio n° 2. Canal 15. Es un canal antrópico construido para prevenir inundaciones en la cuenca inferior del río Salado, pero hay serias dudas acerca de su operatividad como controlador del nivel de las aguas (Conzono et al ., 2001). Fluye desde el curso inferior del río Salado hacia la bahía de Samborombón con dirección oeste noroeste-este sureste. Fue ampliado y profundizado, por lo cual lleva agua no sólo en períodos de crecidas, sino también bajo condiciones de flujo normal. Su régimen hidrológico es sumamente variable y sufre la influencia de las mareas provenientes de la bahía. Esto provoca grandes variaciones en los valores de conductividad, así como en el transporte de materia particulada y disuelta (Miretzky et al ., 1998).
Sitio n° 3. Arroyo Chico. Es un curso de agua meandriforme, de ancho fluctuante entre 5 y 15 m. Pertenece a la cuenca hídrica de los arroyos del sudeste de la provincia de Buenos Aires y forma parte de la misma cuenca que la laguna Mar Chiquita, donde desemboca. En el sitio de muestreo, el lecho está constituido por abundantes restos de arenas conchíferas retrabajadas, asociadas al último evento ingresivo del Holoceno. Carece de vegetación macroscópica y lleva abundante sedimento en suspensión.
Ambientes lénticos permanentes
Sitio n° 4. Laguna Salada Grande. Constituye el área receptora del Complejo Lagunar Salada Grande (Dangavs, 1988), un grupo de lagunas costeras interconectadas de manera semipermanente que mantuvieron conexión marina hasta principios del siglo pasado (Vega et al ., 1995). Su profundidad media es de 0,63 m y la profundidad máxima es de 1,33 m. Presenta una superficie de 47 km2. El fondo es de arena limosa, pero en ciertos sectores el lecho está constituido por abundantes restos de conchillas marinas retrabajadas.
Sitio n° 5. Laguna Las Chilcas. Pertenece al complejo lagunar Salada Grande. Presenta una superficie de 17 km2. El fondo está constituido por arcillas limosas a limos arcillosos de origen marino (Dangavs, 1988). Si bien entre la Salada Grande y Las Chilcas no existe conexión superficial permanente, sí se conectan en épocas de inundaciones.
Sitio n° 6. R56 . Es una pequeña laguna innominada originada a partir de la inundación de una tosquera en desuso, 13 km al norte de General Madariaga. Presenta unos 4 km2 de superficie y una profundidad máxima cercana a 1,50 m. Carece de vegetación macroscópica y se desarrolla sobre sedimentos cuaternarios continentales asignables al Pampeano.
Sitio n° 7. Laguna Los Horcones. Es una cuenca de carácter endorreico. El espejo tiene una superficie de 30 km2, con una profundidad máxima aproximada de 1,50 m. Carece de vegetación subacuática aunque posee escasos juncales que se localizan en los lugares más bajos de su ribera, donde desembocan algunos arroyos de poco caudal. Su reducida superficie la expone a las sequías prolongadas.
Sitio n° 8. Laguna de Chascomús. Forma parte del sistema de lagunas encadenadas de Chascomús, originado sobre un cauce fluvial preexistente pero en cuya génesis también intervinieron el retroceso de las pendientes y la acción deflacionaria del viento (Miretzky, 2001). La laguna de Chascomús (30 km2) posee las características de un sistema eutrófico y alcalino. En épocas de crecientes, funciona como colectora de los excesos pluviales. Hay enriquecimiento salino de las aguas debido a la concentración por evaporación y el afloramiento de aguas subterráneas (Miretzky et al ., 1998).
Sitio n° 9. Laguna del Monte. Forma parte del sistema endorreico de lagunas encadenadas del oeste. Se localiza en el área medanosa del distrito fitogeográfico Surpampeano. Desde el punto de vista zoogeográfico, marca la transición de la región Pampeana a las regiones Central o Subandina. Presenta un área de 174,5 km2 y una profundidad media de 4,8 m, aunque la profundidad máxima alcanza los 7,7 m (Van Eerden y Ledesma, 1994). La vegetación está restringida a pequeños sectores costeros y consiste básicamente en juncales.
Ambientes lénticos temporarios y semipermanentes
Estos ambientes corresponden a charcas y bañados sin cuenca definida, sin sedimento propio y sin exhondación posterior, con vegetación emergente y flotante sumamente abundante. Estos microlimnótopos no exceden las 103 m2. Representan zonas anegables de perímetro irregular con agua semipermanente asociada principalmente al régimen pluvial y al aporte freático.
Sitio n° 10. M'Chabot. Está situada en el extremo sur del delta de Paraná, en el sector conocido como Bajo Delta (Vega et al ., 1995), límite norte de la Pampa Deprimida. Esta región posee una gran cantidad de riachos y arroyos, con islas formadas por acumulación de sedimentos fluviales (Vega et al ., op . cit .). El sitio muestreado es una pequeña depresión alimentada por desborde embalsada por vegetación. Presenta una superficie aproximada de 250 m2 y 40 cm de profundidad. Situado en el interior de la isla definida por los arroyos Espera y Gélvez, se encuentra sujeto a aislamiento y reconexión periódicos con el arroyo Espera. Posee abundante vegetación marginal, sumergida y flotante.
Sitio n° 11. El Juncal. Es una charca de lluvia originada en una pequeña depresión deflacionaria de unos 200 m2 de superficie y 50 cm de profundidad. Se trata de una depresión cerrada, que depende del régimen de lluvias pero que recibe aguas de desborde del complejo lagunar Salada Grande. Presenta una profusa vegetación flotante y arraigada, aunque en otros sectores el junco es la vegetación dominante.
Localidades 12 a 15 - R74, R74(1), R74(2) y R74(3). Serie de pequeños bañados ubicados a lo largo de la ruta 74 entre Pinamar y General Madariaga. Son alimentados principalmente por agua de lluvia y el escurrimiento superficial. En el caso de R74(2), recibe además aguas de desborde de la laguna Los Horcones. Se trata de un conjunto de pequeños cuerpos de agua de contornos poco definidos, con abundantes fanerógamas sumergidas y flotantes, con coberturas del 80 al 100%. Suelen ser de profundidad muy reducida [menores a 10 cm en R74(2)], pero R74(3) alcanza 1,50 m de profundidad.
Sistemática
Phylum Crustacea Pennant, 1777
Clase Ostracoda Latreille, 1806
Orden Podocopida G. W. Müller, 1894
Suborden Podocopa Sars, 1866
Superfamilia Cypridoidea Baird, 1845
Familia Cyprididae Baird, 1845
Género Chlamydotheca Saussure, 1858
Especie tipo. Cypris ( Chlamydotheca ) azteca Saussure en Moore y Pitrat, 1961 (pág. 214, fig. 150.1).
Chlamydotheca iheringi (Sars, 1901)
Figuras 2.A, 4.A-C

Figura 2. Ostrácodos no marinos de la pampa bonaerense. Vex: Vista lateral externa; Vi: valva izquierda; Vd: valva derecha; Ca: caparazón; Vin: vista interna; Vld: vista lateral derecha; Vliz: vista lateral izquierda; la: largo; al: alto; He: hembra; Ma: macho / non marine ostracods from the pampas from Buenos Aires Province. Ex: external view; Lv: left valve; Rv: right valve; Ca: carapace; In: internal view, Rlv: right lateral view; Llv: left lateral view; Do: dorsal view; l: length; h: height; Fe: female; Ma: male. A, Chlamydotheca iheringi (Sars), Vex / Ex , Vd / Rv , M'Chabot, la / l : 3,24 mm, al / h : 1,88 mm. B, Chlamydotheca incisa (Claus), Vex / Ex , Vi / Lv , Fe / He , El Juncal, la / e : 2,39mm; al / h : 1,32mm. C, Chlamydotheca incisa (Claus), Vin / In , Vd / Rv , El Juncal, la / l, al / h : 1,34 mm. D, Heterocypris similis (Wierzejski), Ca / Ca , Vliz / Llv , Las Chilcas, la / l : 1,22 mm, al / h : 0,69 mm; E, Heterocypris similis (Wierzejski), Ca / Ca , Vld / Rlv , Las Chilcas, la / l : 1,19 mm, al / h : 0,67 mm; F, Heterocypris incongruens (Ramdohr), Ca / Ca , Vliz / Llv , Las Chilcas, la / l : 1,14, al / h : 0,61 mm; G, Cypridopsis vidua (O.F. Müller), Vex / Ex , Vd / Rv , El Juncal, la / l : 0,48 mm; al / h : 0,28 mm; H, Cypridopsis vidua (O.F. Müller), Vex / Ex , Vi / Lv , M'Chabot, la / l : 0,55 mm, al / h : 0,35mm; I, Potamocypris sp., Ca / Ca , Vld / Rlv , R74 / R74 , la / l : 0,45 mm, al / h : 0,22 mm; J. Candona sp. Vex / Ex , Vi / Lv , R74 / R74 , la / l : 0,76 mm, al / h : 0,41 mm; K, Candona sp. aff. C. annae Méhes, Vex / Ex , Vd / Rv , R74 / R74 , la / l : 0,56 mm, al / h : 0,30mm; L, Candona sp. aff. C. pedropalensis Mèhes, Vex / Ex , Vd / Rv , R74(3) / R74(3) , la / l : 0,55 mm, al / h : 0,25 mm; LL, Candonopsis brasiliensis Sars, Vex / Ex , Vi / Lv , M'Chabot, la / l : 0,69 mm, al / h : 0,33 mm; M, Ilyocypris ramirezi Cusminsky y Whatley, Ca / Ca , Vliz / Llv , Arroyo Chico, la / l : 0,86 mm, al / h : 0,47mm; N, Vestanula pagliolii (Pinto y Kotzian), Ca / Ca , Vld / Rlv , R74 / R74 , la / l : 0,56 mm, al / h : 0,28 mm.
1901. Cypris iheringi Sars: 25-26; lám. 4, figs. 1-4.
1905. Eucypris ( Chlamydotheca ) iheringi (Sars). Daday: 248.
1934. Chlamydotheca iheringi G.O. Sars. Behm: 82-83.

Figura 3. Ostrácodos no marinos de la Pampa bonaerense. Para la explicación de las abreviaturas véase la leyenda de la figura 2 / non marine ostracods from the pampas from Buenos Aires Province. See figure 2 for explanation. A, Darwinula stevensoni (Brady y Robertson), Vex / Ex , Vi / Lv , M'Chabot, la / l : 0,58 mm, al / h : 0,22 mm; B, Cyprideis multidentata Hartmann, Ca / Ca , Vliz / Llv , He / Fe , Punta Rasa, la / l : 1,14 mm, al / h : 0,61; C, Cyprideis multidentata Hartmann, Vex / Ex , Vd / Rv , Ma / Ma , Punta Rasa, la / l : 1,26 mm, al / h : 0,65 mm; D, Cyprideis salebrosa Van den Bold, Vex / Ex , Vd / Rv , Ma / Ma , Arroyo Chico, la / l : 1,21 mm, al / h : 0,63 mm; E, Cyprideis salebrosa Van den Bold, Vex / Ex , Vi / Lv , He / Fe , Arroyo Chico, la / l : 1,04 mm, al / h : 0,65 mm; F, Perissocytheridea sp., Vex / Ex , Vd / Rv , Punta Rasa, la / l : 0,39 mm, al / h : 0,20 mm; G, Cytherura dimorphica Bertels y Martínez, Vex / Ex , Vi / Lv , Punta Rasa, la / l : 0,41 mm, al / h : 0,20 mm; H, Limnocythere sp., Ca / Ca , Vliz / Llv , Ma / Ma , Laguna del Monte, la / l : 0,54 mm, al / h : 0,26 mm; I, Limnocythere sp., Vex / Ex , Vd/ Rv , He / Fe , Laguna del Monte, la / l : 0,61 mm, al / h : 0,29 mm; J, Limnocythere sp. aff. L. Staplini Gutentag y Benson, Vex / Ex , Vd / Rv , Ma / Ma , Arroyo Chico, la / l : 0,51 mm, al / h : 0,25 mm; K, Limnocythere sp. aff. L. Staplini Gutentag y Benson, Ca / Ca , Vliz / Llv , Ma / Ma , Arroyo Chico, la / l : 0,47 mm, al / h : 0,22 mm; L, Limnocythere sp. aff. L. Staplini Gutentag y Benson, Vex / Ex , Vd / Rv , He / Fe , Arroyo Chico, la / l : 0,59 mm, al / h : 0,29 mm; M, Cytheridella ilosvayi Daday, Vex / Ex , Vi / Lv , He / Fe , M'Chabot, la / l : 0,65 mm, al / h : 0,41 mm.
Repositorio. FCEN-LM 2900-Dos hembras adultas y un ejemplar macho.
Procedencia. M'Chabot (sitio n°10).
Observaciones. Escasos ejemplares fueron recuperados de ambientes temporarios, de escasa profundidad, vegetados, hipohalinos, del Bajo Delta.
Distribución geográfica. Ampliamente distribuido en Brasil, Paraguay y Argentina (Martens y Behen, 1994), asociado exclusivamente a ambientes no-marinos, temporarios y permanentes (Kotzian, 1974).
Chlamydotheca incisa (Claus, 1893)
Figuras 2.B-C, 4.D-G

Figura 4. Esquemas en cámara clara mostrando la morfología externa e interna de los ostrácodos no marinos de la Pampa bonarense. No a escala. Para la explicación de las abreviaturas véase la leyenda de la figura 2 / camera lucida drawings showing external and internal morphology of non marine ostracodes of the pampas from Buenos Aires Province. Drawings not to scale. See figure 2 for explanation . A-C, Chlamydotheca iheringi (Sars); A, Vex / Ex , Vi / Lv ; B, Vex / Ex , Vd / Rv , He / Fe ; C, Vin / In , Vi / Lv , He / Fe ; D-G, Chlamydotheca incisa (Claus); D, Vex / Ex , Vd / Rv , He / Fe ; E, Vin / In , Vd / Rv , He / Fe ; F, Vex / Ex , Vi / Lv , He / Fe ; G, Vin / In , Vi / Lv , He / Fe ; H-K, Heterocypris similis (Wierzejski); H, Vex / Ex , Vd / Rv , He / Fe ; I, Vin / In , Vi / Lv ; J, Vex / Ex , Vd / Rv , Ma / Ma ; K, Vin / In , Vd / Rv , Ma / Ma ; L-O, Heterocypris incongruens (Rahmdor); L, Vex, Vd. Ex, Rv; M, Vin, Vd, In, Rv; N, Vex, Vi. Ex, Lv; O, Vin / In , Vi / Lv ; P-R, Cypridopsis vidua (O. F.Müller); P, Vex / Ex , Vd / Rv ; Q, Vex / Ex , Vi / Lv ; R, Vin / In , Vi / Lv .
1893. Pachycypris incisa Claus: 205; lám. 8, figs. 7-15.
Repositorio. FCEN-LM 2901-Dos ejemplares.
Procedencia. El Juncal (sitio n° 11) y R7(2) (sitio n° 14).
Observaciones. Ambientes oligohalinos temporarios de escasa profundidad y vegetados. Alcalinidad media a fuerte; pH=7,5-9; DO= 4,1- 6,5 mg/L; TDS= 680-2070 mg/L; aguas cloruradas sódicas oligobicarbonatadas y bicarbonatadas sódicas hemicloruradas oligomagnésicas.
Distribución geográfica. Extensamente distribuida por América del Sur, habiéndose registrado en Argentina, Bolivia, Brasil, Chile, Paraguay, Perú y Uruguay (Martens y Behen, 1994). También fue hallada en sedimentos cuaternarios (Bertels y Martínez, 1990) y ambientes actuales (Ramírez, 1967; César et al ., 2004) de la provincia de Buenos Aires.
Género Heterocypris Claus, 1893
Especie tipo. Cypris incongruens Rahmdor en Moore y Pitrat, 1961 (pág. 221, fig. 155.5).
Heterocypris similis (Wierzejski en Ramírez, 1967)
Figuras 2.D-E, 4.H-K
1967. Cyprinotus similis Wierzejski. Ramírez: 29-30; lám. 5, figs. 27-32.
Repositorio. FCEN-LM 2902-Cuatro ejemplares femeninos y cinco masculinos.
Procedencia. Arroyo Chico (sitio n°3), Salada Grande (sitio n°4), Las Chilcas (sitio n°5), Chascomús (sitio n°8) y R74(2) (sitio n°14).
Observaciones. Esta especie fue hallada en la costa de lagunas permanentes, en ambientes lóticos y en charcas temporarias de salinidad hipohalina mesopoikilohalina positiva a oligohalina; aguas fuertemente alcalinas; pH=7,5-10,1; DO=4,1-14,5 mg/L; TDS=478-3540 mg/L; aguas cloruradas o hemicloruradas sódicas o hemisódicas hemibicarbonatadas oligo a hipomagnésicas. En Chascomús y R74(2) se la halló asociada a aguas hemibicarbonatadas o bicarbonatadas sódicas cloruradas u oligocloruradas.
Distribución geográfica. Se encuentra en ambientes no marinos de Argentina y Brasil (Martens y Behen, 1994). Según Moguilevsky y Whatley (1995), las especies de este género habitan charcas temporarias y lagunas permanentes.
Heterocypris incongruens (Ramdohr, 1808)
Figuras 2.F, 4.L-O
1808. Cypris incongruens Rahmdor: 86; lám. 3, figs. 1-12, 15-16 y 19-20.
Repositorio. FCEN-LM N°2903-Dos ejemplares hembra.
Procedencia. Salada Grande (sitio n°4), Las Chilcas (sitio n°5) y Los Horcones (sitio n°7).
Observaciones. La especie fue hallada en ambientes costeros de lagunas oligohalinas de fuerte alcalinidad; pH=8-12,8; DO=9,2-14,5 mg/L; TDS=2395-4435 mg/L; aguas cloruradas a hemicloruradas sódicas o hemisódicas hemibicarbonatadas hipo a oligomagnésicas.
Distribución geográfica. Es una especie de distribución cosmopolita; en América del Sur se la registró en Argentina, Brasil, Uruguay y Venezuela (Kotzian, 1974; Purper y Würdig-Maciel, 1974; Martens y Behen, 1994). Es frecuente en sedimentos no marinos del Cuaternario de la provincia de Buenos Aires (Ferrero, 1996) y en ambientes actuales del norte de la Patagonia (Schwalb et al ., 2002).
Género Cypridopsis Brady, 1867
Especie tipo. Cypris vidua O.F. Müller en Moore y Pitrat, 1961 (pág. 230, fig. 164.1).
Cypridopsis vidua (O.F. Müller, 1776)
Figuras 2.G-H, 4.P-R, 5.A

Figura 5. Esquemas en cámara clara mostrando la morfología externa e interna de los ostrácodos no marinos de la Pampa bonarense. No a escala. Para la explicación de las abreviaturas véase la leyenda de la figura 2 / camera lucida drawings showing external and internal morphology of non marine ostracodes of the pampas from Buenos Aires Province. Drawings not to scale. See figure 2 for explanation. A, Cypridopsis vidua (O.F. Müller), Vin / In , Vd / Rv ; B-C, Potamocypris sp.; B, Vex / Ex , Vi / Lv ; C, Vin / In, Vi / Lv; D-G, Candona sp.; D, Vex / Ex , Vd / Rv ; E, Vin / In , Vd / Rv ; F, Vex / Ex , Vi / Lv ; G, Vin / In , Vi / Lv ; H-J, Candona sp. aff. C. anae (Mèhes); H, Vex / Ex , Vd / Rv ; I, Vex / Ex, Vi / Lv; J, Vin / In , Vd / Rv ; K-L, Candona sp. aff. C. pedropalensis (Mèhes); K, Vex / Ex , Vd / Rv ; L, Vin / In , Vd / Rv ; M-P, Candonopsis brasiliensis Sars; M, Vex / Ex , Vi / Lv ; N, Vex / Ex , Vd / Rv ; O, Vin / In , Vi / Lv ; P, Vin / In , Vd / Rv ; Q-S, Vestanula pagliolii (Pinto y Kotzian); Q, Vex / Ex , Vd / Rv ; R, Vex / Ex , Vi / Lv ; S, Vin / In , Vi / Lv ; T-U, Cyprideis multidentata Hartmann; T, Vex / Ex, Vi / Lv, He / Fe; U, Vex / Ex , Vd / Rv , Ma / Ma .
1776. Cypris vidua O.F. Müller: 199.
1962. Cypridopsis vidua (O.F. Müller). Gutentag y Benson: 26-28; fig. 6.
Repositorio. FCEN-LM 2904-Seis ejemplares.
Procedencia. Arroyo Chico (sitio n°3), Salada Grande (sitio n°4), Las Chilcas (sitio n°5), R56 (sitio n°6), M'Chabot (sitio n°10), El Juncal (sitio n°11), R74 (sitio n°12), R74(1) (sitio n°13) y R74(2) (sitio n°14).
Observaciones. Es una de las especies con mayor distribución en el área. Se la halló en ambientes hipohalinos y oligohalinos lóticos, lénticos, permanentes y temporarios; pH=7,5-10,1; DO=14,5-4,1 mg/L; TDS=1010-3540 mg/L; aguas fuertemente alcalinas, bicarbonatadas sódicas y cloruradas a hemicloruradas sódicas.
Distribución geográfica. Especie de distribución cosmopolita; en Sudamérica se la encuentra en ambientes actuales de Argentina, Brasil, Paraguay, Uruguay y Venezuela (Martens y Behen, 1994; César et al ., 2004); en Argentina fue recuperada de ambientes cuaternarios desde el Chaco hasta la Patagonia (Zabert, 1981; Zabert y Herbst, 1986; Cusminsky, 1995).
Género Potamocypris Brady, 1870
Especie tipo. Bairdia fulva Brady en Moore y Pitrat, 1961 (pág. 230, fig. 165.4).
Potamocypris sp.
Figuras 2.I, 5.B-C
Repositorio. FCEN-LM N° 2905-Seis ejemplares.
Procedencia. Arroyo Chico (sitio n°3), R74 (sitio n°12) y R74(1) (sitio n°13).
Observaciones. Escasos ejemplares fueron hallados en ambientes lénticos y lóticos con aguas oligohalinas fuertemente alcalinas; pH=7,8-9,1; DO=6,4-11,4 mg/L; TDS=1010-1850 mg/L; aguas bicarbonatadas sódicas cloruradas a hemicloruradas hipo a oligopotásicas u oligomagnésicas y cloruradas hemisódicas hemibicarbonatadas oligomagnésicas.
Familia Candonidae Kaufmann, 1900
Género Candona Baird, 1845
Especie tipo. Cypris reptans Baird en Moore y Pitrat, 1961 (pág. 232, fig. 167.2).
Candona sp.
Figuras 2.J, 5.D-G
Repositorio. FCEN-LM 2906-Dos ejemplares.
Procedencia. R74 (sitio n° 12).
Observaciones. Escasos ejemplares fueron hallados viviendo en un ambiente temporario de escasa profundidad, sin drenaje, con aguas oligohalinas fuertemente alcalinas; pH=8,6; DO=11,4 mg/L; TDS=1010 mg/L; aguas bicarbonatadas sódicas hemicloruradas oligomagnésicas.
Candona sp. aff. C. annae Mèhes, 1914
Figuras 2.K, 5.H-J
aff. 1914. Candona annae Mèhes: 653-658; figs. 9 a-c; figs. 11 a-c.
Repositorio. FCEN-LM N°2907-Tres ejemplares.
Procedencia. R74 (sitio n°12) y R74(1) (sitio n°13).
Observaciones. Un reducido número de individuos fue hallado en ambientes semipermanentes de aguas oligohalinas fuertemente alcalinas; pH=7,8-8,6; DO=6,4-11,4 mg/L; TDS=1010-1285 mg/L; aguas bicarbonatadas sódicas cloruradas a hemicloruradas oligomagnésicas u oligopotásicas.
Candona sp. aff. C. pedropalensis Mèhes, 1914
Figuras 2.L, 5.K-L
aff. 1914. Candona pedropalensis Mèhes: 651-653; figs. 7a y b.
Repositorio. FCEN-LM N°2908-Tres ejemplares.
Procedencia. R74(3) (sitio n°15).
Observaciones. Escasos ejemplares fueron hallados en un ambiente temporario oligohalino fuertemente alcalino con abundante vegetación; pH=7,2; DO=2,3 mg/L; TDS=1015 mg/L; agua bicarbonatada sódica oligoclorurada oligomagnésica.
Familia Eucandonidae Swain, 1961
Género Candonopsis Vávra, 1891
Especie tipo. Candona kingsleyi Brady y Robertson en Moore y Pitrat, 1961 (pág. 235, fig. 171.2).
Candonopsis brasiliensis Sars, 1901
Figura 2.LL, 5.M-P
1901. Candonopsis brasiliensis Sars: 45; lám. 8, figs. 21-22.
1981. Candona elliptica Furtos. Zabert: 79; lám. 1, fig. 2; lám. 3, figs. 6a-c.
Repositorio. FCEN-LM N°2909-Cuatro ejemplares adultos.
Procedencia. M'Chabot (sitio n°10) y R74(3) (sitio n°15).
Observaciones. La especie se halló exclusivamente asociada a ambientes temporarios hipo-oligohalinos de alcalinidad fuerte; pH=7,5; DO= 2,3 mg/L; TDS=1015 mg/L; de aguas bicarbonatadas sódicas hemicloruradas oligomagnésicas.
Distribución geográfica. Citada para ambientes actuales del Brasil (Würdig, 1983), es relativamente frecuente en sedimentos cuaternarios del noreste argentino (Zabert, 1981; Zabert y Herbst, 1986), la provincia de Buenos Aires (Bertels y Martínez, 1990) y el norte de la Patagonia (Cusminsky y Whatley, 1996; Whatley y Cusminsky, 2000). Según Moguilevsky y Whatley (1995), este género es común en microambientes acuosos retenidos en hojas y en fondos barrosos.
Familia Ilycyprididae Kaufmann, 1900
Género Ilyocypris Brady y Norman, 1889
Especie tipo. Cypris gibba Rahmdor en Moore y Pitrat, 1961 (pág. 239, fig. 176.1).
Ilyocypris ramirezi Cusminsky y Whatley, 1996
Figura 2.M
1996. Ilyocypris ramirezi Cusminsky y Whatley: 148; lám. 1, figs. 11-14.
Repositorio. FCEN-LM N°2910-Cinco ejemplares.
Procedencia. Arroyo Chico (sitio n°3), Salada Grande (sitio n°4), Las Chilcas (sitio n°5) y R56 (sitio n°6).
Observaciones. La especie fue hallada en ambientes costeros de lagunas permanentes y cuerpos lóticos, asociada a aguas oligohalinas de alcalinidad muy fuerte; pH=8-10,1; DO=8,4-14,5 mg/L; TDS= 745-3540 mg/L; aguas cloruradas a hemicloruradas sódicas hemibicarbonatadas oligo a hipomagnésicas y bicarbonatadas sódicas oligocloruradas hipopotásicas.
Distribución geográfica. La especie fue descripta para sedimentos lacustres cuaternarios del norte patagónico, pero había sido previamente registrada en ambientes actuales de Buenos Aires (Ramírez, 1967; Moguilevsky y Whatley, 1995) y el norte patagónico (Schwalb et al. , 2002), como así en sedimentos cuaternarios de Buenos Aires (Bertels y Martínez, 1990) y el Chaco (Zabert, 1981).
Superfamilia Darwinuloidea Brady y Norman, 1889
Familia Darwinulidae Brady y Norman, 1889
Género Vestanula Rossetti y Martens, 1998
Especie tipo. Vestanula boteai Rossetti y Martens, 1998.
Vestanula pagliolii (Pinto y Kotzian, 1961)
Figuras 2.N, 5.Q-S
1961. Darwinula pagliolii Pinto y Kotzian: 9-10; lám. 1, figs. 1-5.
Repositorio. FCEN-LM 2911-Dos ejemplares hembras.
Procedencia. R74 (sitio n°12).
Observaciones. Se encontraron escasos ejemplares en un ambiente temporario de escasa profundidad y agua oligohalina fuertemente alcalina; pH=8,6; DO=11,4 mg/L; TDS=1010 mg/L; agua bicarbonatada sódica hemiclorurada oligomagnésica.
Distribución geográfica. Originalmente descripta en Brasil (Pinto y Kotzian, 1961), es considerada una especie cosmopolita (Martens et al ., 1997).
Género Darwinula Brady y Robertson en Jones, 1885
Especie tipo. Polycheles stevensoni Brady y Robertson en Moore y Pitrat, 1961 (pág. 254, fig. 183.2a-c).
Darwinula stevensoni (Brady y Robertson 1870)
Figura 3.A
1870. Polycheles stevensoni Brady y Robertson en Moore y Pitrat,
1961: 254; lám. 183, figs. 2a-c.
Repositorio. FCEN-LM N°2912-Un ejemplar hembra.
Procedencia. M'Chabot (sitio n°10).
Observaciones. Sólo dos ejemplares adultos de esta especie fueron hallados en un ambiente temporario de aguas hipohalinas, con abundante vegetación flotante y sumergida y una profundidad aproximada de 30 cm.
Distribución geográfica. Descripta originalmente para Inglaterra (Brady y Robertson, 1870), D. Stevensoni es una especie cosmopolita euritérmica (Martens et al ., 1997). Martens y Tudorance (1991) hallaron esta especie viviendo en ambientes de escasa profundidad (< 2 m) con un rango de pH entre 8,8-9,1. La tolerancia a la salinidad es amplia: entre 800 mg/L a 2-3 g/L, aunque hay datos que establecen que fue recuperada en aguas de salinidades cercanas a los 15 g/L (Külköylüoglu y Vinyard, 2000). En Europa se conoce fósil desde el Mioceno (Martens et al ., 1997), habiéndose registrado en sedimentitas oligocenas de la Patagonia (Bertels-Psotka y Cusminsky, 1999).
Superfamilia Cytheroidea Baird, 1850
Familia Cytherideidae Sars, 1925
Género Cyprideis Jones, 1857
Especie tipo. Candona torosa Jones en Moore y Pitrat, 1961 (pág. 276, figs. 201.3 y 207.1).
Cyprideis multidentata Hartmann, 1955
Figuras 3.B-C, 5.T-U, 6.A

Figura 6. Esquemas en cámara clara mostrando la morfología externa e interna de los ostrácodos no marinos de la Pampa bonarense. No a escala. Para la explicación de las abreviaturas véase la leyenda de la figura 2 / camera lucida drawings showing external and internal morphology of non marine ostracodes of the pampas from Buenos Aires Province. Drawings not to scale. See figure 2 for explanation. A, Cyprideis multidentata Hartmann, Vin / In , Vd / Rv , Ma / Ma ; B-D, Cyprideis salebrosa Van den Bold; B, Vex / Ex , Vi / Lv , Ma / Ma ; C, Vex / Ex , Vd / Rv , He / Fe ; D, Vin / In , Vi / Lv , Ma / Ma ; E-F, Perissocytheridea sp.; E, Vex / Ex , Vd / Rv ; F, Vin / In , Vi / Lv ; G-I, Cytherura dimorphica Bertels y Martínez; G, Vex / Ex , Vd / Rv ; H, Vin / In , Vd / Rv ; I, Vin / In , Vi / Lv ; J-N, Limnocythere sp.; J, Vex / Ex , Vi / Lv , Ma / Ma ; K, Vin / In , Vd / Rv , Ma / Ma ; L, Vex / Ex , Vd / Rv , He / Fe ; M, Vin / In , Vd / Rv , He / Fe ; N, Vin / In , Vd / Rv , He / Fe ; O-R, Limnocythere sp. aff. L. staplini Gutentag y Benson; O, Vex / Ex , Vd / Rv, Ma / Ma ; P, Vex / Ex , Vi / Lv , Ma / Ma ; Q, Vin / In, Vd / Rv , Ma / Ma ; R, Vex / Ex , Vd / Rv , He / Fe ; S-U, Cytheridella ilosvayi Daday; S, Vex / Ex , Vd / Rv , He / Fe ; T, Vin / In , Vd / Rv , He / Fe ; U, Ca / Ca , Vdo / Do , He / Fe .
1955. Cyprideis multidentata Hartmann: 119, 121-123; figs. 1-6.
Repositorio. FCEN-LM N°2913-10 caparazones machos y 10 caparazones hembras.
Procedencia. Punta Rasa (sitio n° 1) y Canal 15 (sitio n° 2).
Observaciones. Fue hallada en ambientes estuáricos de salinidades oligohalinas mesopoikilohalinas positivas hasta polihalinas fuertemente alcalinas; TDS= 3,1 y 34 g/L (López, 1995); aguas cloruradas a hemicloruradas sódicas hemibicarbonatadas oligo a hipomagnésicas, moderadamente básicas (Conzono et al ., 2001).
Distribución geográfica. Originalmente descripta en Brasil (Hartmann, 1955), se halló en los canales que unen una laguna marginal con el mar (Pinto y Ornellas, 1965). Es muy abundante en los sedimentos cuaternarios de la provincia de Buenos Aires (Aguirre y Whatley, 1995; Bertels y Martínez, 1997).
Cyprideis salebrosa Van den Bold, 1963
Figuras 3.D-E, 6.B-D
1963. Cyprideis salebrosa Van den Bold: 377-378; lám. 7, figs. 9a-d; lám. 11, figs. 1a-c.
Repositorio. FCEN-LM 2914-Cinco caparazones machos y seis hembras adultos.
Procedencia. Canal 15 (sitio n° 2), Arroyo Chico (sitio n°3), Salada Grande (sitio n°4), Las Chilcas (sitio n°5), R56 (sitio n°6) y Chascomús (sitio n°8).
Observaciones. En lagunas permanentes y cuerpos lóticos hipohalinos a oligohalinos de alcalinidad fuerte a muy fuerte (entre 285-771 mg/L HCO-3); pH=8-10,1; DO=8,4-14,5 mg/L; TDS=478-3540 mg/L; agua clorurada a hemiclorurada sódica hemibicarbonatada oligo a hipomagnésica. También se la halló asociada a aguas bicarbonatadas a hemibicarbonatadas sódicas oligocloruradas hipocálcicas o hipopotásicas.
Distribución geográfica. Descripta originalmente para sedimentos pliocenos de Trinidad (Van den Bold, 1963), en la actualidad está bien representada en América desde Argentina hasta Venezuela (Martens y Behen, 1994). Es muy frecuente en ambientes actuales (Ramírez, 1967) y sedimentos cuaternarios de la provincia de Buenos Aires (Bertels y Martínez, 1997) y Entre Ríos (Zabert y Herbst, 1986).
Familia Cytheridae Baird, 1850
Género Perissocytheridea Stephenson, 1938
Especie tipo. Cytheridea matsoni Stephenson en Moore y Pitrat, 1961 (pág. 280, fig. 206.2 i-j).
Perissocytheridea sp.
Figuras 3.F, 6.E-F
Repositorio. FCEN-LM 2915-Dos caparazones y dos valvas.
Procedencia. Punta Rasa (sitio n°1) y Canal 15 (sitio n°2).
Observaciones. Esta especie se halló exclusivamente en ambientes estuarinos. Los datos disponibles en la bibliografía indican un rango de salinidades entre 3,1 y 34 g/L, aguas fuertemente alcalinas, moderadamente básicas, cloruradas a hemicloruradas sódicas hemibicarbonatadas oligo a hipomagnésicas (Conzono et al ., 2001).
Familia Cytheruridae G.W. Müller, 1894
Género Cytherura Sars, 1866
Especie tipo. Cythere gibba O.F. Müller en Moore y Pitrat, 1961 (pág. 292, figs. 219.2 y 220.4).
Cytherura dimorphica Bertels y Martínez, 1997
Figuras 3.G, 6.G-I
1997. Cytherura dimorphica Bertels y Martínez: 43-44 ; lám. 3, figs. 2-5; lám. 6, fig. 10.
Repositorio. FCEN-LM 2916-Dos caparazones y dos valvas.
Procedencia. Punta Rasa (sitio n°1) y Canal 15 (sitio n°2).
Observaciones. Esta especie se halló exclusivamente en ambientes estuarinos. Los datos disponibles en la bibliografía indican un TDS=3,1-34 g/L, aguas fuertemente alcalinas, moderadamente básicas, cloruradas a hemicloruradas sódicas hemibicarbonatadas oligo a hipomagnésicas (Conzono et al ., 2001).
Distribución geográfica. Ambientes actuales y sedimentos cuaternarios de la provincia de Buenos Aires (Bertels y Martínez, 1997). Martínez (en prensa) la halló en el estuario de Bahía Blanca, sur de la provincia de Buenos Aires.
Familia Limnocytheridae Klie, 1938
Género Limnocythere Brady, 1868
Especie tipo. Cythere inopinata Baird en Moore y Pitrat, 1961 (pág. 309, fig. 235.2).
Limnocythere sp.
Figuras 3.H-I, 6.J-N
Repositorio. FCEN-LM 2917-Dos caparazones hembra y dos caparazones machos.
Procedencia. Laguna del Monte (sitio n°9).
Observaciones. Fue hallada en una laguna permanente de aguas mesohalinas en el límite entre la Pampa Deprimida y la Pampa Arenosa, el sector más árido del área analizada. En este ambiente en el que la evaporación excede a la precipitación, las aguas son mesohalinas (TDS= 8933 mg/L), de fuerte alcalinidad, fuertemente básicas (pH= 9,05), muy poco oxigenadas (DO= 2 mg/L), hemi a cloruradas sódicas hemibicarbonatadas hipomagnésicas.
Limnocythere sp. aff. L . Staplini Gutentag y Benson, 1962
Figuras 3.J-L, 6.O-R
aff. 1962. Limnocythere staplini Gutentag y Benson: 51-53; fig. 15; lám. 1, figs. 1-3.
Repositorio. FCEN-LM 2918-Cinco caparazones machos y cuatro hembras.
Procedencia. Salada Grande (sitio n°4), Los Horcones (sitio n°5), R56 (sitio n°6), Arroyo Chico (sitio n°3) y Chascomús (sitio n°8).
Observaciones. Fue hallada en lagunas permanentes y ambientes lóticos de aguas oligohalinas a hipohalinas fuertemente alcalinas; pH=8-12,8; DO=8,4-12,4 mg/L; TDS=478-4435 mg/L; aguas cloruradas a hemicloruradas sódicas a hemisódicas hemibicarbonatadas oligomagnésicas. En R56 y en Chascomús se la halló asociada a condiciones bicarbonatadas a hemibicarbonatadas sódicas oligocloruradas hipopotásicas o hipocálcicas.
Distribución geográfica. Esta especie es abundante en sedimentos cuaternarios del sur de la provincia de Buenos Aires (Bertels y Martínez, 1990, 1997; Ferrero, 1996).
Género Cytheridella Daday, 1905
Especie tipo. Cytheridella ilosvayi Daday, 1905.
Cytheridella ilosvayi Daday, 1905
Figuras 3.M, 6.S-U
1905. Cytheridella ilosvayi Daday: 263-267; lám. 17, figs. 15-18; lám. 18, figs. 1-11.
Repositorio. FCEN-LM 2919-Dos caparazones hembra y una caparazón macho.
Procedencia. M'Chabot (sitio n°10), El Juncal (sitio n°11), R74, R74(1), R74(2) y R74(3) (sitios n°12-15).
Observaciones. Exclusivamente en ambientes temporarios y semipermanentes de aguas oligohalinas de alcalinidad media a fuerte (178-650 mg HCO-3/L); pH=7,5-9; DO=2,3-11,4 mg/L; TDS=680-2070 mg/L. Sin embargo, es tolerante a variaciones en la composición de las aguas, ya que se la halló asociada tanto a aguas bicarbonatadas sódicas cloruradas a hemicloruradas oligomagnésicas u oligopotásicas como a cloruradas sódicas oligobicarbonatadas hipopotásicas.
Distribución geográfica. Está extensamente distribuida por toda América del Sur, desde Chile hasta Venezuela (Martens y Behen, 1994). En Argentina, es frecuente en sedimentos cuaternarios estuáricos de la provincia de Buenos Aires y continentales de Entre Ríos (Bertels y Martínez, 1990; Zabert y Herbst, 1986).
Discusión
En la provincia de Buenos Aires existe una enorme diversidad de categorías de aguas lénticas. Los espejos de agua son permanentes o temporarios, someros y de carácter eutrófico. Carecen de estratificación química o térmica (Dangavs, 1998). En estos ambientes acuáticos pampeanos, la salinidad, la composición iónica y la estabilidad determinan los patrones de distribución de las poblaciones de ostrácodos. Dado que estos factores son, en última instancia, climáticamente controlados, el conocimiento de cómo estos factores regulan dicha distribución tiene importantes connotaciones para la interpretación paleolimnológica del registro fósil cuaternario.
Composición iónica de las aguas pampeanas
La composición iónica de las aguas superficiales de la llanura pampeana es muy variable. Uno de los principales factores que la determina es la naturaleza geológica de las cuencas sobre las cuales se desarrollan. La presencia de depósitos de origen marino favorece la formación de aguas Cl-/Na+ dominadas mientras que los de origen continental dan origen a aguas enriquecidas en HCO 3 -(Delorme, 1969; Dangavs, 1998). A partir de la composición determinada por estas condiciones iniciales, los mecanismos de fraccionamiento iónico determinarán la composición final (Eugster y Jones, 1979), siendo el balance entre las precipitaciones y la evaporación el principal proceso responsable del fraccionamiento en la llanura pampeana. Esto resulta de particular importancia ya que la composición iónica es uno de los factores que determina la distribución de los ostrácodos (Forester y Brouwers, 1985).
En la mayoría de las lagunas pampeanas, la composición varía entre clorurada sódica y bicarbonatada sódica (Dangavs, 1998). En nuestro estudio, la mayor parte de las lagunas y cursos permanentes resultaron ser clorurados sódicos, como consecuencia de su relación directa con la bahía de Samborombón (Punta Rasa, Canal 15), de su origen (lagunas costeras relícticas tales como Salada Grade, Las Chilcas), de los sedimentos cuaternarios marinos sobre las cuales se desarrollan (arroyo Chico) o bien como consecuencia de su evolución natural en ambientes en los cuales la evaporación excede las precipitaciones (laguna del Monte). Las únicas excepciones las constituyen la laguna de Chascomús y R56, ambas hemibicarbonatadas sódicas. Los datos publicados previamente describen a Chascomús como un cuerpo de agua clorurado sódico (Ringuelet, 1962; Miretzki, 2001), pero lluvias intensas pueden provocar modificaciones no sólo en términos de salinidad sino también de la composición iónica, pudiendo pasar de una condición clorurada a una bicarbonatada como consecuencia del aporte de aguas meteóricas.En efecto, el muestreo de Chascomús (15/01/2002) se efectuó luego de un invierno particularmente lluvioso, con algunas lluvias intensas concentradas en pocos días ( i.e. en la semana del 19/10/2001 al 25/10/2001 llovieron en la estación de Chascomús 130 mm según estadísticas Ministerio de Economía y Producción). Una situación diferente se verifica en R56, ya que este cuerpo de agua se desarrolla sobre sedimentos continentales pampeanos, evidenciándose sobre la costa un banco de tosca, lo que explicaría su condición bicarbonatada.
En contraposición al carácter dominantemente clorurado de los cuerpos permanentes, la mayor parte de los cuerpos no permanentes son bicarbonatados sódicos. Dado que suelen ubicarse sobre suaves depresiones y no sobre cubetas, su condición hidrológica está controlada principalmente por la presencia de sedimentos no marinos (generalmente asignables al Pampeano s.l .) y el aporte freático. Existen dos casos que se apartan de esta generalización. La estación R74(2) presenta aguas cloruradas sódicas, pero por su ubicación y morfología se puede inferir que recibe aguas de desborde de la laguna Los Horcones. La otra excepción la constituye El Juncal, desarrollado sobre el extremo occidental de la cuenca de drenaje del Complejo Lagunar Salada Grande, presentando por lo tanto identidad hidrológica con dicha cuenca.
Salinidad de las aguas pampeanas y diversidad
En los ambientes no marinos, la salinidad tiene una gran influencia en el número de especies, mientras que la estabilidad temporal y la composición iónica de las aguas determinan las especies presentes (Forester, 1983; Smith, 1993). El número de especies suele ser bajo en los ambientes hipohalinos, aumentando con el incremento de la salinidad, pero descendiendo abruptamente más allá del punto de precipitación de la calcita (TDS=3 g/L) (De Deckker y Forester, 1988). Si analizamos el número de especies de ostrácodos en los sitios relevados en función del TDS (cuadros 3 y 4), veremos que estas tendencias se verifican en los ambientes de carácter permanente de la Pampa Deprimida. El mayor número de especies se registró en los ambientes oligohalinos con un TDS cercano a los 2000 mg/L (Salada Grande, arroyo Chico), disminuyendo a medida que la salinidad aumenta o disminuye. Cabe destacar que el bajo número de especies hallado en los ambientes meso e hipohalinos (laguna de Monte y Chascomús, respectivamente) puede ser el resultado del arte de muestreo utilizado (un gravity core ). Dadas las diferencias metodológicas de extracción, estos datos no pueden ser comparados con el resto de las muestras, obtenidas mediante barrido de red. Sin embargo son, al menos, comparables entre sí: en los ambientes hipohalinos se identificaron más especies que en los ambientes mesohalinos.
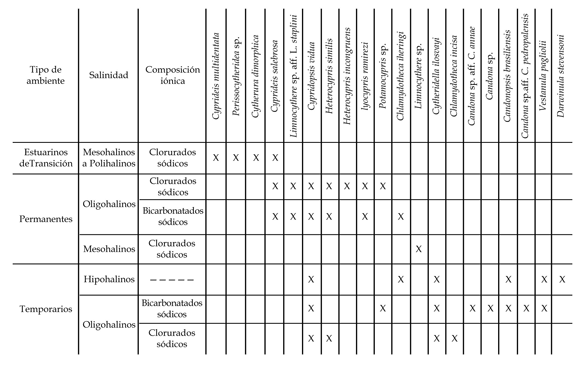
Cuadro 4. Distribución de los ostrácodos en los ambientes actuales de la pampa bonaerense / distribution of the Ostracoda in modern environments of the Pampas from Buenos Aires Province.
Distribución de los ostrácodos en los ambientes pampeanos
Dentro del amplio espectro de ambientes de la Pampa bonaerense, los distintos taxones de ostrácodos pueden ser agrupados de acuerdo a su preferencia por el tipo de agua y la estabilidad del ambiente (cuadro 4).
El primer grupo incluye especies que habitan exclusivamente ambientes estuáricos en conexión directa con la bahía de Samborombón (Punta Rasa y Canal 15). En estos ambientes hallamos a Cyprideis multidentata , Perissocytheridea sp. y Cytherura dimorphica . Estas especies se desarrollan exclusivamente en aguas cloruradas sódicas con variaciones diarias muy marcadas de salinidad y pH como consecuencia de la influencia de las mareas provenientes del estuario platense. Según Forester y Brouwers (1985), muchas especies estuarinas requieren fluctuaciones periódicas en las concentraciones de Cl- y Na+, por lo cual la distribución de ostrácodos estuáricos no es una simple respuesta a la salinidad, sino a variaciones de muy alta frecuencia en las concentraciones iónicas, lo que explica la ausencia de estas especies en otro tipo de ambientes clorurados sódicos de salinidad semejante.
Otro grupo de especies se encuentra asociado a lagunas permanentes y arroyos oligohalinos, sean sus aguas cloruradas, hemicloruradas o bicarbonatadas sódicas. En estos ambientes hallamos a Cyprideis salebrosa , Ilyocypris ramirezi , Limnocythere sp. aff. L. staplini , Heterocypris incongruens y Heterocypris similis . La ausencia de estas especies en cuerpos temporarios y semipermanentes parece implicar que la estabilidad temporal constituye un factor limitante para su desarrollo. Esta asociación incluye especies moderadamente eurihalinas, capaces de sobrevivir a fluctuaciones lentas y graduales de la salinidad y de la composición iónica, frecuentes en una región sometida a ciclos periódicos de inundaciones y sequías como lo es la pampa bonaerense, donde las lagunas varían gradualmente de hipohalinas bicarbonatadas (en momentos de mayor precipitación relativa) a oligomesohalinas cloruradas (en momentos de menor precipitación relativa) (Laprida et al ., 2004). Según Forester y Brouwers (1985), la variación tolerada por estas especies capaces de vivir en más de un tipo de agua es limitada, lo que explica la ausencia de estas especies en los ambientes estuarinos Cl-/Na+ dominados.
Cabe destacar que si bien Heterocypris incongruens es considerada una especie cosmopolita que se encuentra en todo tipo de agua y bajo todo régimen soportando salinidades de hasta 20 g/L (Külköylüoglu y Vinyrad, 2000), en este estudio se la halló exclusivamente asociada a cuerpos permanentes clorurados sódicos de aguas oligohalinas. Por otra parte, si bien en estudios previos Cyprideis salebrosa fue hallada en lagunas litorales con conexión marina y abundante vegetación (Würdig, 1983), en la pampa bonaerense se la halló exclusivamente en lagunas permanentes sin conexión marina.
En términos generales, la fauna de ambientes temporarios es altamente especializada e incapaz de sobrevivir en ambientes permanentes (Martens y de Moor, 1995). La gran mayoría de las especies registradas en la pampa bonaerense están limitadas a un solo tipo de agua (clorurada sódica o bicarbonatada sódica) indicando que la composición iónica constituye un factor determinante en su distribución. Una notable excepción la constituye Cytheridella ilosvayi . Las especies que caracterizan los ambientes temporarios hipohalinos u oligohalinos bicarbonatados sódicos son Candona sp. aff. C. annae , Chlamydotheca iherengi y Candonopsis brasiliensis . De hallazgo más esporádico resultaron Vestanula pagliolii , Candona sp. aff. C . pedropalensis y Candona sp.
Las asociaciones que caracterizan a los ambientes temporarios oligohalinos clorurados sódicos merecen un párrafo aparte. Como fuera mencionado previamente, estos ambientes son bañados de lluvia que reciben aguas de desborde de lagunas permanentes. Como consecuencia de ello, las asociaciones son una mezcla de especies cosmopolitas fuertemente eurioicas presentes tanto en ambientes permanentes como temporarios ( Cypridopsis vidua ); especies típicas de ambientes no permanentes clorurados sódicos ( Chlamydotheca incisa ); especies eurihalinas típicas de ambientes no permanentes ( Cytheridella ilosvayi ) pero también especies eurioicas típicas de ambientes permanentes ( Heterocypris similis ) transportadas con las aguas de desborde. Las biocenosis de este tipo de ambientes ponen de manifiesto la necesidad de considerar a la asociación y no a taxones individuales como indicadores de tipo de hábitat.
Por último, cabe destacar que Cypridopsis vidua fue hallada en todo tipo de ambientes. Se trata de una especie tolerante a amplias variaciones de pH, temperatura y salinidad, bajo todo régimen de estabilidad por lo que no puede ser utilizada como indicador de salinidad o calidad de agua.
Modos reproductivos, estrategias de dispersión y hábitos
Además de la preferencia por un tipo particular de agua o régimen temporal, otros aspectos tales como los modos reproductivos, las estrategias de dispersión y los hábitos son determinantes en los patrones de distribución de ostrácodos continentales (Horne et al ., 1998). En los ambientes permanentes de la pampa bonaerense la gran mayoría de las especies posee reproducción sexual. Es sabido que la partenogénesis es común en ambientes ecológicamente estables (en términos de previsibilidad) y que la reproducción sexual tiene lugar con el incremento de la inestabilidad ambiental (Horne et al ., 1998; Horne y Martens, 1999). La gran cantidad de especies sexuales en los ambientes permanentes de la pampa bonaerense podría ser consecuencia de las marcadas variaciones en el balance precipitación/evaporación en diversas escalas temporales, desde anuales a interdecádicas, que caracterizan esta región (Deschamps et al ., 2003; Laprida et al ., 2004). Por su parte, en los ambientes temporarios se registran tanto especies partenogenéticas como sexuales. Si bien las especies asexuales pueden colonizar más fácilmente los cuerpos de agua no permanentes (Horne et al ., 1998; Horne y Martens, 1999), la partenogénesis no es un factor que asegure el éxito en estos ambientes. Aunque no pudo verificarse la presencia de machos en los Candonidae, la gran mayoría se reproduce sexualmente (Horne y Martens, 1999), por lo que podríamos establecer que la sexualidad es el principal modo de reproducción en estos ambientes no permanentes, ya que es la estrategia más exitosa para sobrevivir a las variaciones drásticas de la salinidad que tienen lugar estacionalmente (Schwalb et al ., 2002).
Adicionalmente, algunas especies pueden tener ciertos atributos que resultan preadaptaciones significativas para colonizar los hábitats sometidos a desecación periódica. La existencia de cámaras de incubación, común en la mayoría de los darwinulidos, frecuente en los cytheroideos y presente en algunos cypridoideos, permite soportar períodos de sequía asegurando la recolonización cuando las condiciones son de nuevo propicias. De acuerdo a nuestras observaciones, hembras de Chlamydotheca incisa , Cytheridella ilosvayi , Darwinula stevensoni y Vestanula pagliolii , exclusivas de ambientes no permanentes, retienen los primeros estadíos juveniles dentro del caparazón. Estos "cuidados maternales" aumentan la probabilidad de supervivencia en ambientes inestables y representan una preadaptación para la dispersión pasiva por el viento (Horne et al ., 1998).
Por último, existe una clara relación entre la distribución y el hábito de las diferentes especies. Si bien existen especies de hábitos semiterrestres (Horne et al ., 2004), la gran mayoría de las especies de ostrácodos no marinos nadan cortas distancias sobre la interfase agua/sedimento y/o habitan dentro del sedimento (Henderson, 1995). La capacidad de natación está determinada por la presencia de sedas natatorias en las antenas, siendo la antena 2 el principal apéndice locomotor (Van Morkhoven, 1959), de modo que la presencia de sedas natatorias en la antena 2 puede ser considerada como una evidencia de capacidad de natación. Las especies infaunales estrictas habitan exclusivamente los primeros cm del sedimento y son incapaces de nadar, mientras que las especies bentónicas vagabundas habitan el sedimento pero son capaces de nadar cortas distancias por encima de la interfase.
En la pampa bonaerense, la totalidad de las especies que habitan los ambientes estuarinos son infaunales estrictas como consecuencia de la dinámica del medio, sometido periódicamente al flujo y reflujo de las mareas y la exposición subaérea. En estos ambientes el sedimento arenoso o areno-limoso favorece la circulación intersticial, el intercambio de nutrientes y por lo tanto el desarrollo de la infauna (Carbonel, 1980).
En los ambientes no marinos permanentes, lóticos o lénticos, el 66% de las especies presentes son bentónicas vagabundas ( Cypridopsis , Heterocypris , Ilyocypris , Potamocypris y Chlamydotheca ), mientras que en los ambientes oligohalinos no permanentes y temporarios, el 75% de las especies son infaunales estrictas ( Cytheridella , Candona , Candonopsis , Vestanula y Darwinula ). Esto es consecuencia de que, en sedimentos de grano fino, la distribución de las especies también es influida por la disponibilidad de alimento y la predación (Benzie, 1989). La mayor parte de los ambientes permanentes de la pampa poseen aguas turbias como consecuencia de una elevada cantidad de masa fitoplanctónica que determina estados eutróficos a hipertróficos (Quirós et al ., 2002). En ello, la oferta alimentaria sigue los ciclos de la productividad primaria asociada al desarrollo del fitoplancton (Gabellone et al ., 2001). Esta productividad primaria se exporta al medio bentónico, depositándose en la interfase y decreciendo abruptamente con la profundidad (Laprida et al ., 2004), de modo que las especies capaces de explotar la interfase estarán mejor posicionadas al momento de utilizar el recurso alimenticio. En contraposición, los ambientes no permanentes suelen poseer aguas claras y escasa masa fitoplanctónica. Al ser zonas anegables sin sedimentos propios, el fondo constituye una mezcla de materiales húmicos. Por ello, es probable que en estos ambientes las especies exploten intersticialmente la materia orgánica lábil, favoreciéndose la presencia de taxones infaunales estrictos.
Este no es el único factor que influye en los hábitos dominantes. Si bien no existen muchos datos publicados sobre la influencia de la predación sobre la distribución de los ostrácodos, se cree que la presencia de peces y otros predadores (por ejemplo, larvas de insectos y moluscos) puede tener un considerable impacto (Henderson, 1995; Benzie, 1989). Por ello, la turbidez y la presencia de vegetación también pueden influir en la distribución (Ruiz et al ., 2000; Bronshtein, 1947; Argollo y Mourguiart, 2000). Los ambientes límnicos permanentes relevados presentan aguas turbias y abundante vegetación arraigada en sus orillas, proveyendo refugio y protección frente a los potenciales predadores y favoreciendo la presencia de bentos vagabundo (Carbonel, 1980). Por el contrario, los ambientes no permanentes suelen poseer aguas claras y presentan vegetación flotante, por lo que las especies de ambientes temporarios están más expuestas a la predación (Martens y de Moor, 1995).
Conclusiones
Los principales factores que determinan la distribución de los ostrácodos en la llanura pampeana son: la naturaleza química de las aguas, la estabilidad temporal del ambiente, los modos reproductivos, la disponibilidad del alimento y las presiones de predación. Dado que algunos de estos factores son climáticamente controlados, la distribución de las especies tiene una directa aplicación en estudios paleolimnológicos.
Se reconocieron tres tipos de ambientes: estuáricos, lagunas y arroyos permanentes, y bañados no permanentes. Estos ambientes son portadores de una asociación particular de especies de ostrácodos que puede ser considerada como indicadores ambientales en la pampa bonaerense.
Cyprideis multidentata , Perissocytheridea sp. Y Cytherura dimorphica son indicadoras de aguas oligomesohalinas a polihalinas, Cl-/Na+ dominadas, sometidas a fuertes variaciones diarias de la salinidad, características propias de los ambientes estuáricos. En estos ambientes dominan las especies infaunales estrictas.
Cyprideis salebrosa , Limnocythere sp. aff. L. staplini , Heterocypris incongruens , Ilyocypris ramirezi y Heterocypris similis son indicadoras de ambientes oligohalinos, Cl-/Na+ o HCO3-/Na+ dominados, eutróficos a hipertróficos, tales como lagunas y arroyos permanentes en los cuales suelen registrarse variaciones graduales de la salinidad y composición. En estos ambientes dominan las especies bentónicas vagabundas.
Candona sp. aff. C . annae , Chlamydotheca iherengi , Candonopsis brasiliensis , Vestanula pagliolii , Candona sp. aff. C. pedropalensis y Candona sp. son indicadoras de ambientes no permanentes o temporarios de aguas hipohalinas u oligohalinas bicarbonatadas sódicas. En estos ambientes dominan las especies infaunales estrictas.
Cytheridella ilosvayi indica aguas temporarias oligohalinas, pero no puede ser utilizada para caracterizar tipo de agua.
Cypridopsis vidua no puede ser utilizada como un indicador del tipo de ambiente ni calidad de agua.
Chlamydotheca incisa caracteriza cuerpos temporarios de aguas oligohalinas de carácter clorurado sódico. Su asociación con Cytheridella ilosvayi y otras especies eurioicas y cosmopolitas tales como Cypridopsis vidua y Heterocypris similis indica la influencia de aguas de desborde de cuerpos permanentes.
Más datos deben ser recopilados para generar una plataforma conceptual más amplia sobre la cual puedan apoyarse las interpretaciones basadas en asociaciones fósiles. Las reconstrucciones que consideran exclusivamente la ausencia/presencia de determinadas especies marcadoras pueden ser erróneas. Por ello, deben efectuarse estudios que incluyan aspectos tales como diversidad, riqueza y dinámica poblacional de las asociaciones de ostrácodos pampásicos, los que deben ser considerados en las interpretaciones paleolimnológicas tendientes a revelar la evolución paleoclimática de la pampa bonaerense.
Agradecimientos
Este trabajo fue financiado por los proyectos "Micropaleontología (foraminíferos, ostrácodos y radiolarios) del Mesozoico y Cenozoico de la República Argentina y mares adyacentes" (Proyecto ANCyT-CONICET PIP 4091) y "Climatologia-Sinóptica de la Circulación Atmosférica en el Hemisferio Sur y su Relación con la Variabilidad Climática y Paleoclimática sobre América del Sur" (Proyecto UBACyT X002). Mi reconocimiento a R.H. Compagnucci por alentarme a realizar este estudio; a Blas Valero-Garcés (Instituto Pirenaico de Ecología, España) y a M.J. Orgeira (INGEODAV) del proyecto CAHCILA por proveer las muestras correspondientes a Laguna del Monte y Chascomús. Mi agradecimiento al invalorable aporte de los revisores R. Whatley y G. Cusminsky, quienes contribuyeron notablemente al mejoramiento del manuscrito.
Bibliografía
Aguirre, M.L. y Whatley, R.C. 1995. Late Quaternary marginal marine deposits and palaeoenvironments from northeastern Buenos Aires Province, Argentina: a review. Quaternary Science Reviews 14: 223-254. [ Links ]
Argollo, J. y Mourguiart, Ph. 2000. Late Quaternary climate history of the Bolivian Altiplano. Quaternary International 72: 37-51. [ Links ]
Baird, W. 1845. Arrangement of the British Entomostraca, with a list of species, particularly noticing those which have as yet been discovered within the bounds of the club. Transactions of the Berkwickshire naturalist´ club 2: 145-158. [ Links ]
Behm, V. 1934. Über sudamerikanische Ostrakoden des Zoologischen Staatinstitutes und Museums in Hamburg. Zoologischen Anzeiger 108: 74-85. [ Links ]
Benzie, J.A.H. 1989. The distribution and habitat preference of ostracods (Crustacea: Ostracoda) in a coastal sand-dune lake, loch of Strahbeg, northeast Scotland. Freshwater Biology 22: 309-321. [ Links ]
Bertels, A. y Martínez, D.E. 1990. Quaternary ostracodes of continental and transitional littoral-shallow marine environments. Courier Forschunginstitut Senckenberg 123: 141-159. [ Links ]
Bertels, A. y Martínez, D. 1997. Ostrácodos holocenos de la desembocadura del Arroyo Napostá Grande, sur de la provincia de Buenos Aires, Argentina. Revista Española de Micropaleontología 29: 29-69. [ Links ]
Bertels-Psotka, A. y Cusmisky, G.C. 1999. Nuevas especies de ostrácodos de la Formación Ñirihuau (Oligoceno) en su área tipo (alrededores de San Carlos de Bariloche), provincia de Río Negro, República Argentina. Ameghiniana 36: 71-81. [ Links ]
Brady, G.S. 1867. A synopsis of the recent British Ostracoda. The intellectual Observer 12: 110-130. [ Links ]
Brady, G.S. 1868. A monograph of the recent British Ostracoda. Transactions of the Linnean Society of London 26: 353-495. [ Links ]
Brady, G.S. 1870. Notes on Entomostraca taken chiefly in the Northumber land and Durham districts. Natural History Society of Northum land an Durham, Transactions 3: 361-373. [ Links ]
Brady, G.S. y Norman, A.M. 1889. A monograph of marine and non-marine fresh-water Ostracoda of the North Atlantic and of Northwestern Europe. Section 1. Podocopa: Royal Dublin Society Sciences Transactions, ser. 2, v. 4, p. 63-270, pl. 8-23. [ Links ]
Brady, G.S. y Robertson, D. 1870. The Ostracoda and Foraminifera of tidal rivers. Annals and Magazine of Natural History , serie 4: 1-33, 307-309. [ Links ]
Bronshtein, Z.S. 1947. Fresh-water Ostracoda. Fauna of the USSR. Volume 2, N° 1, Crustacean. RussianTranslation Series 64. Academy of Sciences of the USSR Publishers, Moscow. A. A. Balkema, Ámsterdam (reprinted 1988), 445 pp. [ Links ]
Carbonel, P. 1980. Les ostracodes et leur intêret dans la définition des écosystèmes estuariens et de plateforme continentale. Essais d'application a des domaines anciens. Mémoires de l'Institut de Géologie du Bassin d'Aquitaine 11, 350 pp. [ Links ]
Carbonel, P., Colin, J.-P., Danielopol, D.L., Löffler, H. y Neustrueva, I. 1988. Palaeoecology of limnic ostracods: a review of some major topics. Palaeogeography, Palaeoclimatology, Palaeoecology 62: 413-461. [ Links ]
César, I.I., Armendáriz, L.C. y Damborenea, M.C. 2004. Ostrácodos (Crustacea) de la Isla Martín García, Río de la Plata, Argentina. Natura Neotropicalis 32 (2001): 147-151. [ Links ]
Claus, C. 1893. Beitrage zur kenntniss der süsswasser-Ostracoden. I. Über den Korper und Gliedmassenbau der Cypriden, nebst Bemerkungen über einzelne organe derselben. Arbeiten aus Zoologische Institut der Universitat Wien zoologische 10: 147-216. [ Links ]
Conzono, V., Miretzky, P. y Fernández Cirelli, A. 2001. The impact of man-made hydrology on the lower stream bed of the Salado River drainage basin (Argentina). Environmental Geology 40: 968-972. [ Links ]
Curry, B.B. 1999. An environmental tolerance index for ostracodes as indicators of physical and chemical factors in aquatic habitats. Palaeogeography, Palaeoclimatology, Palaeoecology 148: 51-63. [ Links ]
Cusminsky, G.C. 1995. Ostrácodos lacustres cuaternarios en la transecta Tronador-Laguna Cari-Laufquen, Patagonia Norte, Argentina. 6° Congreso Argentino de Paleontología y Bioestratigrafía (Trelew, 1994), Actas : 99-105. [ Links ]
Cusminsky, G.C. y Whatley, R. 1996. Quaternary non-marine ostracodes from lake beds in northern Patagonia. Revista Española de Paleontología 11: 143-154. [ Links ]
Daday, E. 1902. Mikroskopische Süswasserthiere aus Patagonien, gesammelt von Dr Filippo Silvestri. Természetrajzi fützetek 25: 201-310. [ Links ]
Daday, E. 1905. Untersuchungen über die Süsswasser. Mikrofauna Paraguayans. Zoologica 18: 1-374. [ Links ]
Dangavs, N. 1988. Geología, sedimentología y limnología del Complejo Lagunar Salada Grande, partidos de General Madariaga y General Lavalle, provincia de Buenos Aires, República Argentina. Informe CIC, 143 pp. [ Links ]
Dangavs, N. 1998. Los ambientes lénticos de la pampasia bonaerense. En: A. Fernández Cirelli (ed.), Agua problemática regional , Eudeba, pp. 145-152. [ Links ]
De Deckker P. y Forester, R.M. 1988. The use of ostracods to reconstruct continental palaeoenvironmental records. En: P. De Deckker, J.-P. Colin y J.-P. Peypouquet (eds.), Ostracoda in the Earth Sciences , Elsevier, pp. 175-200. [ Links ]
Delorme, L.D. 1969. Ostracodes as Quaternary indicators. Canadian Journal of Earth Sciences 6: 1471-1476. [ Links ]
Deschamps, J.R., Otero, O. y Tonni, E.P. 2003. Cambio climático en la pampa bonaerense: las precipitaciones de los siglos 18 al 20. Documentos de Trabajo N° 109. Departamento de Investigación. Universidad de Belgrano, 18 pp. [ Links ]
Eugster, H.P. y Jones, B.F. 1979. Behaviour of major solutes during closed-basin brine evolution. American Journal of Science 279: 609-631. [ Links ]
Ferguson, E. Jr. 1967. Three new species of freshwater otracods (Crustacea) from Argentina. Notulae Naturae of the Academy of Natural Sciences of Philadelphia 405: 1-7. [ Links ]
Fernández, C.A y Romero, E.J. 1984. Palynology of Quaternary Sediments of Lake Chascomús, Northeastern Buenos Aires Province, Argentina. Quaternary of South America and Antarctic Peninsula 2: 201-221. [ Links ]
Ferrero, L. 1996. Paleoecología de ostrácodos holocenos del estuario del río Quequén Grande (provincia de Buenos Aires). Ameghiniana 32: 209-222. [ Links ]
Forester, R.M. 1983. Relationship of two lacustrine ostracod species to solute composition and salinity: implications for paleohydrochemistry. Geology 11: 435-438. [ Links ]
Forester, R.M. 1987. Late Quaternary paleoclimate records from lacustrine ostracodes. En: W.F. Ruddiman y H.E. Wright Jr. (eds.), North America and Adjacent Oceans During the Last Deglaciation. The Geology of North America K-3. GeologicalSociety of America, Boulder, CO, pp. 261-276. [ Links ]
Forester, R. M. 1991. Ostracode assemblages from springs in the western United States: Implications for paleohydrology. Memoirs of the Entomological Society of Canada 155: 181-201. [ Links ]
Forester, R.M. y Brouwers, E.M. 1985. Hydrochemical parameters governing the occurrence of estuarine and marginal estuarine ostracodes: an example from south-central Alaska. Journal of Paleontology 59: 344-369. [ Links ]
Forester, R.M. y Smith, A.J. 1992. Microfossils as Indicators of Paleohydrology and Paleoclimate. Paleohydrogeological Methods and Their Applications. Proceedings of NEA Workshop , Paris, pp. 39-57. [ Links ]
Gabellone, N.A, Solari, L.C. y Claps, M.C. 2001. Planktonic and physico-chemical dynamics of a markedly fluctuating backwater pond associated with a lowland river (Salado River, Buenos Aires, Argentina). Lakes & Reservoirs: Research and Management 6: 133-142. [ Links ]
Gutentag, E.D. y Benson, R.H. 1962. Neogene (Plio-Pleistocene) fresh water ostracodes from the Central High Plains. State Geological Survey of Kansas, Bulletin 157: 1-60. [ Links ]
Hartmann, G. 1955. Neue marine ostracoden der Familie Cytheridae. Zoologischer Anzeiger 154: 109-157. [ Links ]
Henderson, P.A. 1995. Freshwater Ostracoda. Synopses of the British Fauna 41. The Linean Society & The Estuarine and Coastal Association. Universal Books Services, 228 pp. [ Links ]
Horne, D.J. y Martens, K. 1999. Geographical parthenogenesis in European non-marine ostracods: post-glacial invasion or Holocene stability?. Hydrobiologie 391: 1-7. [ Links ]
Horne, D.J., Baltanás, A. y Paris, G. 1998. Geographical distribution of reproductive modes in living non-marine ostracods. En: K. Martens (ed.), Sex and Parthenogenesis, Backuys Publishers, pp. 77-100. [ Links ]
Horne, D.J., Smith, R.J., Whittaker, J.E. y Murray, J.W. 2004. The first British record and a new species of the superfamily Terrestricytheroidea (Crustacea, Ostracoda): morphology, ontogeny, lifestyle and phylogeny. Journal of the Linnean Society 142: 253-288. [ Links ]
Iriondo, M. 1994. Los climas cuaternarios de la región pampeana. Comunicaciones del Museo Provincial de Ciencias Naturales Florentino Ameghino (nueva serie) 4: 46 pp. [ Links ]
Jones, T.R. 1857. A monograph of Tertiary Entomostraca of England. Monographs of the Palaeontological Society 9: 1-68. [ Links ]
Jones, T.R. 1885. On the Ostracoda form Purberk Formation with notes on the Wealden species. Quarterly Journal of Geological Society of London 41: 311-353. [ Links ]
Kotzian, S.C.B. 1974. New fresh-water ostracodes of the genus Chlamydotheca from Brazil. Ecology, Geographic distribution and stratigraphical position. Anales Academia Brasileira de Ciencias 46: 423-467. [ Links ]
Külköylüoglu, O. y Vinyard, G.L. 2000. Distribution and ecology of freshwater Ostracoda (Crustacea) from springs of Nevada, Idaho, and Oregon: preliminary study. Western North American Naturalist 60: 291-303. [ Links ]
Laprida, C., Gómez, A., Rosemblat, A. y Castañeda, E. 2004. Cambios ambientales en la Laguna de Chascomús mediante el análisis de ostrácodos y datos históricos: estudio preliminar. 22° Reunión Científica de la Asociación Argentina de Geofísicos y Geodestas (Buenos Aires, 2004) Resúmenes : 85-86. [ Links ]
López, R.A. 1995. [ Dinámica costera e impacto ambiental generado por la urbanización en el Municipio de la Costa. Informe de Beca de Perfeccionamiento, Universidad de Buenos Aires, 123 pp. Inédito.]. [ Links ]
Martens, K. y Behen, F. 1994. A checklist of the Recent non-marine ostracods (Crustacea, Ostracoda) from the Inland waters of South America and adjacent islands. Travaux scientifiques du Musée National d'Historie Naturelle de Luxembourg 22: 84 pp. [ Links ]
Martens, K. y de Moor, F. 1995. The fate of the Rhino Ridge pool at Thomas Barnes Nature Reserve: a cautionary tale for nature conservationists. South African Journal of Science 91: 385-387. [ Links ]
Martens, K. y Tudorance, C. 1991. Seassonality and spatial distribution of ostracods of Lake Zwai, Ethiopia (Crustacea, Ostracoda). Freshwater Biology 25: 233-241. [ Links ]
Martens, K., Rossetti, G. y Fuhrmann, R. 1997. Pleistocene and Recent species of the Family Darwinulidae Brady & Norman, 1889 (Crustacea, Ostracoda) in Europe. Hydrobiologia 357: 99-116. [ Links ]
Martínez, E.D. (en prensa). Asociaciones de ostrácodos modernos del estuario de Bahía Blanca, Buenos Aires, Argentina. Ameghiniana . [ Links ]
Mèhes, G. 1914. Süsswasser-ostracoden aus Columbien und Argentinien. Mémoires de la Societé Neuchateloise des Sciences Naturelles 5: 639-663. [ Links ]
Mezquita, F., Tapia, G. y Roca, J.R. 1999. Ostracoda from springs on the eastern Iberian peninsula: ecology, biogeography and palaeolimnological implications. Palaeogeography, Palaeoclimatology, Palaeoecology 148: 65-89. [ Links ]
Miretzky, P.S. 2001. [ Procesos geoquímicos en la cuenca baja del Río Salado, Provincia de Buenos Aires. Tesis doctoral Universidad de Buenos Aires, 383 pp. Inédito.]. [ Links ]
Miretzky, P.S., Herrero, M.A., Galindo, G. y Fernández Cirelli, A. 1998. Caracterización de los recursos hídricos en la zona del sistema de las Encadenadas de Chascomús, provincia de Buenos Aires. Revista de Geología Aplicada a la Ingeniería y al Ambiente 12: 85-100. [ Links ]
Moguilevsky, A. y Whatley, R. 1995. Crustacea Ostracoda. En: E. Lopretto y G. Tell (eds.), Ecosistemas de aguas continentales. Metodologías para su estudio, Eudeba, pp. 973-999. [ Links ]
Moore, R.C. y Pitrat, C.W. (eds.). 1961. Treatise on Invertebrate Paleontology. Part Q, Arthropoda 3, Crustacea, Ostracoda, Kansas University Press, 442 pp. [ Links ]
Müller, O.F. 1776. Zoologiae Danicae Prodromus , Havniae, 282 pp. [ Links ]
Pinto, I.D. y Kotzian, S.C.B. 1961. Novos ostracodes da Familia Darwinulidae e a variaçao das impressoes musculares. Boletim Instituto de Ciencias Naturais 6: 31. [ Links ]
Pinto, I. D y Ornellas, L. Pinto de 1965. A new brackishwater ostracode Cyprideis riograndensis Pinto et Ornellas sp. nov., from Southern Brazil and its ontogenetic carapace development. Universidade Federal do Porto Alegre. Escola de Geologia de Porto Alegre. Publicaçao especial 8: 78 pp. [ Links ]
Purper, I. y Würdig-Maciel, N.L. 1974. Occurrence of Heterocypris incongruens (Ramdohr), 1808-Ostracoda-in Rio Grande do Sul. Discussion on the allied genera: Cyprinotus , Hemicypris , Homocypris and Eucypris . Pesquisas 3: 69-91. [ Links ]
Quirós, R. 1988. Relationships between air temperature, depth, nutrients and chlorophyll in 103 Argentinian lakes. Verhandlungen Internationale Vereinigung für Limnologie 23: 647-658. [ Links ]
Quirós, R., Rennella, A.M., Boveri, M.B., Rosso, J.J. y Sosnovsky, A. 2002. Factores que afectan la estructura y el funcionamiento de las lagunas pampeanas. Ecología Austral 12: 175-185. [ Links ]
Rahmdor, F.A. 1808. Übber die gattung Cypris Müller und drei zur derselben gehorige neue Arten. Gesellschaft Naturforschender Freunde zu Berlin 2: 83-91. [ Links ]
Ramírez, F.C. 1967. Ostrácodos de lagunas de la provincia de Buenos Aires. Revista del Museo de La Plata , nueva serie; 10, Zoología 73: 5-79. [ Links ]
Ringuelet, R.A. 1962. Ecología acuática continental . Eudeba, 138 pp. [ Links ]
Rossetti, G. y Martens, K. 1998. Taxonomic revision of the Recent and Holocene representatives of the Family Darwinulidae (Crustacea, Ostracoda), with a description of three new genera. Biologie 68: 55-110. [ Links ]
Ruiz, F., González-Regalado, M.L., Baceta, J.I. y Muñoz, J.M. 2000. Comparative ecological analysis of the ostracod faunas from low-and high-polluted southwestern Spanish estuaries: a multivariate approach. Marine Micropaleontology 40: 345-376. [ Links ]
Sala, J.M. 1975. Recursos hídricos (especial mención de las aguas subterráneas). En: V. Angelelli et al ., (eds.), Relatorio. Geología de la provincia de Buenos Aires. 6° Congreso Geológico Argentino, Actas : 169-194. [ Links ]
Sars, G.O. 1866. Oversigt af norges marine ostracoder. Forhandlinger I videnskabs-selskabet I Christiania : 130 pp. [ Links ]
Sars, G.O. 1901. Contributions to the knowledge of the freshwater Entomostraca of South America. Part II. Copepoda-Ostracoda. Archives Mathematischen Naturvidensk B14: 1-52. [ Links ]
Saussure, H. de. 1858. Mémoire sur divers crustacées nouveaux des Antilles et du Méxique. Mémoires de la Societé de Physique et d´Histoire Naturelle de Géneve 14: 417-496. [ Links ]
Schwalb, A., Burns, S.J., Cusminsky, G., Kelts, K. y Margraff. V. 2002. Assemblage diversity and isotopic signals of modern ostracodes and host waters from Patagonia, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 187: 323-339. [ Links ]
Smith, A.J. 1993. Lacustrine ostracod diversity and hydrochemistry in lakes of northern Midwest of the United States. En: K.G. McKenzie y P.J. Jones (eds.), Ostracoda in the Earth and Life Sciences , A. A. Balkema, Rotterdam, pp. 493-500. [ Links ]
Stephenson, M.B. 1938. Miocene and Pliocene Ostracoda of the genus Cytheridea from Florida. Journal of Paleontology 12: 127-148. [ Links ]
Swain, F.M. 1990. Fossil non-marine Ostracoda of the United States. Developments in Palaeontology and Stratigraphy 16. Elsevier, 410 pp. [ Links ]
Van Eerden, M.R. y Ledesma, C.W. 1994. The Lagunas Encadenadas del Oeste: Pearls of the Pampas. Ecological Aspects of Flood Control Alternatives for the Lagunas Encadenadas. Ministry of Transport Public, Works and Water Management Directorate Flevoland. The Netherlands, 53 pp. [ Links ]
Van den Bold, W.A. 1963. Upper Miocene and Pliocene Ostracoda of Trinidad. Micropaleontology 9: 361-424. [ Links ]
Van Morkhoven, F.P.C.M. 1959. The morphology, taxonomy and economic use of post-Paleozoic Ostracoda. Part 1: General. Bataafse Internationale Petroleum Maatschappij, 244 pp. [ Links ]
Vávra, W. 1891. Monographie der ostracoden boehems. Archiv der naturwissenschaftlichen landesdurchforschung von Boehmen 8: 1-116. [ Links ]
Vega, M.A.L., Sistac, R. y González, M.A. 1995. Diagnóstico ambiental de la provincia de Buenos Aires. Tomo I. Aspectos básicos. Ministerio de la Producción de la Provincia de Buenos Aires, 245 pp. [ Links ]
Whatley, R.C. y Cusminsky, G.C. 2000. Quaternary lacustrine Ostracoda from northern Patagonia, Argentina: a review. En: E.H. Gierlowski y K. Kelts (eds.), Lake basins through space and time, American Association of Petroleum Geologist, Studies in Geology 46: 581-590. [ Links ]
Würdig, N.L. 1983. Fresh and brackish water ostracodes from the East coast of the State of Rio Grande do Sul, Brazil. En: R.F. Maddocks (ed.), Application of Ostracoda , Elsevier, pp. 591-504. [ Links ]
Zabert, L. 1981. Ostrácodos cuartarios de Taco Pozo (provincia de Chaco, Argentina) con algunas consideraciones paleoecológicas. FACENA 4, 77-87. [ Links ]
Zabert, L. y Herbst, R. 1986. Ostrácodos Pleistocénicos del arroyo Perucho Verna, provincia de Entre Ríos, Argentina. Ameghiniana 23: 213-224. [ Links ]
Recibido: 6 de octubre de 2004.
Aceptado: 14 de julio de 2005.

