










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.1 Buenos Aires ene./mar. 2006
Primer hallazgo de mamíferos en la Formación Mariño (Mioceno) en Divisadero Largo (Mendoza, Argentina)
Esperanza Cerdeño 1 , Bernardo González Riga 1 y Osvaldo Bordonaro 1
1 Departamento de Geología y Paleontología, IANIGLA-CRICYTCONICET, Avda. Ruiz Leal s/n, Parque Gral. San Martín, 5500 Mendoza, Argentina. espe@lab.cricyt.edu.ar
Resumen. Se dan a conocer los primeros restos de mamíferos hallados en sedimentitas de la Formación Mariño, aflorantes en el área de Divisadero Largo, provincia de Mendoza, Argentina. Los niveles cenozoicos de esta zona son conocidos por la asociación faunística de la Formación Divisadero Largo, la cual ha permitido establecer la Edad-mamífero Divisaderense (Eoceno superior-Oligoceno inferior). No obstante, hasta el presente hallazgo no se habían registrado fósiles en las sedimentitas de la Formación Mariño suprayacente asignada al Mioceno. Esta formación aflora también en otras localidades situadas al sudoeste de la ciudad de Mendoza (Cacheuta, Potrerillos), donde recientemente se han encontrado algunos restos vegetales e icnitas. El hallazgo del ejemplar estudiado en este trabajo es de gran importancia, tanto por ser el primer dato fehaciente de mamíferos fósiles provenientes de esta formación como por constituir probablemente un taxón nuevo para Argentina, dentro del grupo de los notoungulados Mesotheriidae Mesotheriinae. El ejemplar consta de un resto maxilar derecho con el M3, el M2 casi completo, un fragmento del M1, el P4 y un fragmento del P3. Estos dientes pertenecen a un individuo adulto y se caracterizan por su pequeño tamaño, significativamente menor que el de los géneros miocenos conocidos, Eutypotherium , Typotheriopsis y Pseudotypotherium . Presenta semejanzas de talla con Microtypotherium de Bolivia y las especies descriptas recientemente de la Formación Chucal de Chile.
Abstract. First discovery of mammals from Mariño Formation (Miocen) in Divisadero Largo (Mendoza, Argentina) . This paper deals with the first mammal remains found in the Mariño Formation outcropping in the Divisadero Largo area, Mendoza, Argentina. The Cenozoic levels in this zone are well known by the faunal assemblage of the Divisadero Largo Formation, which allowed the establishment of the Divisaderan Land Mammal Age (Late Eocene-Early Oligocene), but until now no vertebrate fossils had been described from the overlying Mariño Formation (Miocene). This formation also outcrops in the localities of Potrerillos and Cacheuta, Southwest Mendoza city, where some paleobotanical remains and ichnites have been recently found. The specimen studied in this paper is the first accurate datum of fossil mammals from the Mariño Formation, and it could probably constitutes a new taxon for Argentina within the notoungulates Mesotheriidae Mesotheriinae. The specimen consists of a maxillary fragment with right M3 and, separated from the maxillary, the almost complete M2, a labial fragment of M1, the P4 and a P3 fragment. These teeth belong to an adult individual and are characterized by their small size, clearly lesser than that of known Miocene genera Eutypotherium , Typotheriopsis , and Pseudotypotherium . This material presents size similarities with the genus Microtypotherium from Bolivia and the very recently described species from the Chucal Formation (Chile).
Key words. Mesotheriidae, Notoungulata; Mariño Formation; Miocene; Mendoza; Argentina.
Palabras clave. Mesotheriidae; Notoungulata; Formación Mariño, Mioceno; Mendoza, Argentina.
Introducción
El área de Divisadero Largo, situada unos 8 km al oeste de la ciudad de Mendoza (figura 1), en el centro oeste de la República Argentina, es bien conocida desde el punto de vista paleontológico tanto por sus niveles triásicos (formaciones Cacheuta y Río Blanco) como por sus formaciones cenozoicas. A estas últimas corresponde la asociación faunística procedente de la Formación Divisadero Largo (López, 2002; Minoprio, 1947; Patterson, 1952; Rusconi, 1946a, 1946b; Simpson et al ., 1962). Esta asociación de vertebrados está integrada por ocho especies de mamíferos, un ave corredora y varios reptiles. Su peculiaridad dio lugar a la definición de la Edad-mamífero Divisaderense (Pascual et al ., 1965), considerada actualmente como Eoceno superior-Oligoceno inferior (López, 2002).

Figura 1. Mapa de ubicación (arriba) y perfil geológico (abajo) del área de Divisadero Largo, Mendoza, Argentina. Nota: algunos autores (Kokogian y Mancilla, 1989; Barton y Rodríguez, 1989) incluyen la Formación Papagayos en la Formación Barrancas / location map (upper) and geological section (bellow) of the Divisadero Largo area, Mendoza, Argentina. Note: some authors (Kokogian and Mancilla, 1989; Barton and Rodríguez, 1989) include the Papagayos Formation within the Barrancas Formation.
En contraste con el registro de la Formación Divisadero Largo, en la misma zona existen extensos afloramientos cenozoicos de la Formación Mariño (Biondi, 1936; Rolleri y Criado Roque, 1970) en los que no se habían descripto restos fósiles. Esta formación, asignada primero al Oligoceno-Mioceno inferior (Yrigoyen, 1993) y posteriormente al Mioceno medio (Irigoyen, 1997), también aflora en el área de Cacheuta y Potrerillos, cerca de los perfiles tipo del Triásico de la cuenca Cuyana.
Hasta el momento, el registro paleontológico de la Formación Mariño es relativamente escaso y fragmentario. En niveles aflorantes al sur del cerro Cacheuta, Rusconi (1949) colectó bivalvos de agua dulce, asignándolos a Corbicula elchaensis Rusconi; estos restos son actualmente objeto de revisión. Por su parte, Sepúlveda (1999) describió microfloras halladas en la zona de Salagasta que le permitieron asignar a la formación una edad Oligoceno tardío-Mioceno temprano. Por otro lado, recientes trabajos de prospección en la zona de Potrerillos han proporcionado algunos restos de vegetales (raíces, fragmentos de troncos y una probable semilla) e icnitas indeterminadas atribuibles con dudas a un ave (Zavattieri et al ., 2001).
El único registro de mamíferos procedente de la Formación Mariño se atribuye a un informe inédito de Rolleri (1950) referido por Yrigoyen (1993), quien menciona el hallazgo efectuado al sudeste del Cerro Cacheuta, asignado al Proterotheriidae Licaphrium cf. floweri Ameghino. Según este último autor, los fósiles provienen de los "Estratos de Mariño" (miembro superior de la formación) y pueden ser referidos a la Edadmamífero Santacrucense (Mioceno temprano). Sin embargo, dichos restos no fueron descriptos ni inventariados y no han podido ser localizados. No se hallan depositados en el Museo de Ciencias Naturales y Antropológicas "J.C. Moyano" de Mendoza (obs. pers.) ni en las colecciones del Museo de La Plata o del Museo Argentino de Ciencias Naturales de Buenos Aires (G. López, com. pers.). Además, Soria (2001), en su revisión de los Proterotheriidae argentinos, no cita ningún resto de " Licaphrium floweri " Ameghino, 1887 (= Anisolophus floweri Ameghino, 1887) fuera de la provincia de Santa Cruz. De igual manera, las revisiones de Pascual y de la Fuente (1993), Pascual y Odreman Rivas (1973) y Rolleri y Fernández Garrasino (1979) tampoco citan restos de mamíferos en la Formación Mariño. El informe de Rolleri (1950) no lo pudimos localizar, pero sí una copia de la Hoja geológica 23c-Mendoza de 1956, inédita (biblioteca del Servicio Geológico y Minero Argentino de Mendoza), en la que sólo aparece una breve mención del hallazgo "de restos de mamíferos en las Tobas Grises inferiores y en los Estratos de Mariño (en la zona de Yagüel, Cacheuta)" (p. 79), sin ninguna precisión sobre ellos. Por esta razón, la presencia de L . Floweri en la Formación Mariño puede considerarse dudosa hasta disponer de nuevas evidencias.
En este contexto, el presente trabajo comprende la descripción sistemática del primer hallazgo fehaciente de mamíferos procedente de la Formación Mariño. Su descripción taxonómica junto con una procedencia estratigráfica precisa reviste interés sistemático, bioestratigráfico y paleobiogeográfico.
El espécimen se ha determinado como un mesotérido Mesotheriinae. La Familia Mesotheriidae comprende dos subfamilias: Trachytheriinae y Mesotheriinae. La primera comprende representantes más primitivos, con dentición más completa que los mesoterinos y generalmente sin diastemas, primer incisivo superior con esmalte labial, caninos presentes o ausentes, P1 y P2 reducidos, P3 y P4 no molariformes, molares con tres lóbulos que se pierden con el desgaste (Reguero, 1999). La Subfamilia Mesotheriinae incluye formas con cráneo relativamente más bajo y ancho, de aspecto fuertemente rodentiforme, con gran desarrollo de los I1; su fórmula dentaria es 1023/2013 (Francis, 1965; Pascual, 1966). Los traquiterinos se han registrado en Argentina en el Eoceno Superior-Oligoceno Superior (Edades-mamífero Divisaderense y Deseadense), mientras que los mesoterinos se conocen desde el Mioceno Inferior (Colhuehuapense?) al Pleistoceno (Ensenadense). Los mesoterinos del Mioceno Inferior-Medio son poco conocidos, lo que confiere más interés al hallazgo realizado en la Formación Mariño.
Contexto geológico y cronoestratigráfico
La Formación Mariño (Biondi, 1936, nom . subst . Rolleri y Criado Roque, 1970) constituye una secuencia clástica continental que alcanza 1.900 m de potencia. Aflora extensamente en diversas localidades del sector norte de la provincia de Mendoza, como en Divisadero Largo (figura 1), en las cerrilladas pedemontanas de la Precordillera, y al sudoeste del Cerro Cacheuta, en el valle de Poterillos. También está expuesta en el faldeo oriental de la Sierra de las Higueras, en el área de Salagasta (Dessanti, 1942; Sepúlveda, 1999).
En esta formación se reconocen tres miembros: Conglomerados Violáceos en la base, Areniscas Entrecruzadas o Areniscas Inestratificadas en su sección media y Estratos de Mariño o Serie del Higueral en su sección superior (Chiotti, 1946). Tal como describe Yrigoyen (1993), los Conglomerados Violáceos (15-50 m) comprenden bancos conglomerádicos y arenosos que se apoyan mediante discordancia erosiva y levemente angular sobre otras formaciones. Se le superpone el miembro Areniscas Entrecruzadas (120-250 m), caracterizado por la presencia de bancos arenosos castaños con estratificación entrecruzada. En menor proporción, aparecen pelitas moradas y algunos niveles conglomerádicos. Finalmente, la sección superior de la formación, conocida como Estratos de Mariño (hasta 1.550 m de potencia), muestra una sucesión alternante de arcilitas y areniscas rojizas y castañas, con algunos niveles de conglomerados poligénicos grises.
Desde el punto de vista paleoambiental, los miembros inferior y superior de la Formación Mariño corresponden a depósitos fluviales y aluviales depositados bajo condiciones áridas y semiáridas, mientras que el miembro medio es asignado a depósitos predominantemente eólicos (Irigoyen, 1997).
La edad de la Formación Mariño ha sido objeto de diversas interpretaciones. A partir de los datos de Chiotti (1946) y Minoprio (1947), Groeber (1951) asignó los Estratos de Mariño al Oligoceno, mientras que otros autores los consideraron miocenos (Bracaccini, 1950; Rolleri, 1950). Basados en consideraciones de geología regional, diversos autores han postulado la correlación entre la Formación Mariño y la Formación Agua de la Piedra (Rolleri y Criado Roque, 1968; Gorroño et al ., 1979; Yrigoyen, 1993), aflorante en el Departamento de Malargüe, sur de Mendoza, de probable edad Oligoceno-Mioceno temprano.
Por otro lado, los datos palinológicos de Sepúlveda (1999) para la Formación Mariño en el área de Salagasta, al norte de la provincia de Mendoza, sugieren una edad Oligoceno tardío-Mioceno temprano para el miembro superior de la formación ("Estratos de Mariño").
Por su parte, Irigoyen (1997) efectuó dataciones paleomagnéticas y radioisotópicas que permiten estimar para la Formación Mariño una edad Mioceno medio, entre 15,7 y 12,2 millones de años. Los resultados de estas dataciones (Irigoyen et al ., 2000) permiten referirla a la Edad-mamífero Friasense sensu lato . No obstante, Flynn y Swisher (1995) y Flynn et al . (2002) advierten que la definición del Friasense es controvertida y su uso tradicional abarcaría las Edades-mamífero Friasense sensu stricto , Colloncurense, Laventense y Mayoense, es decir que el Friasense estaría restringido a un periodo mucho más corto entre las edades Santacrucense y Colloncurense (figura 2). Según la estimación de Irigoyen (1997), la Formación Mariño podría corresponder desde el final del Friasense al Laventense.

Figura 2. Esquema cronoestratigráfico de las Edades-mamífero del Mioceno sudamericano (modificado de Reguero, 1999 y Flynn et al ., 2001) / chronostratigraphical scheme of the South American Miocene Land Mammal Ages (modified from Reguero, 1999 and Flynn et al. , 2001).
Estratigrafía y niveles fosilíferos
En el área de Divisadero Largo (figura 1), la Formación Mariño muestra una estructura homoclinal que buza 36-42° hacia el noroeste (310°).
Esta formación se halla limitada al oeste por una falla inversa de orientación meridional que se extiende al sur de la Estancia San Isidro (Barton y Rodríguez, 1989). Hacia el este, una falla inversa de bajo ángulo y orientación noreste-suroeste eleva la Formación Potrerillos (Triásico Medio) sobre la Formación Mariño, atravesando perpendicularmente las quebradas de Divisadero Largo y Papagayos. Esta falla inversa fue inicialmente reconocida por Fossa Mancini (1937) y Rolleri (1950) y luego estudiada por Borrello et al . (1972).
En la quebrada Divisadero Largo, la Formación Mariño se apoya discordantemente sobre la Formación Divisadero Largo (figura 1). En esta localidad, las areniscas medias y finas gris amarillentas de la sección superior de la Formación Divisadero Largo buzan alrededor de 49° hacia el noroeste (312°). Son sobrepuestas mediante una discordancia erosiva y levemente angular por un conglomerado poligénico que marca la base de la Formación Mariño. El miembro inferior (Conglomerados Violáceos) constituye una secuencia clástica granodecreciente de 42,1 m de potencia. Se inicia con conglomerados grises clastosoportados que se intercalan paulatinamente con facies arenosas y pelíticas rojizas y moradas (figura 3). Al igual que en otras secciones descriptas (Irigoyen, 1997), la arquitectura y estructura interna de estas facies indican un origen fluvial predominante. Las facies conglomerádicas y arenosas, de base erosiva y arreglo interno granodecreciente, corresponden a depósitos de canales fluviales de alta energía y escasa agradación vertical, probablemente de tipo entrelazado, mientras que los niveles pelíticos rojizos y violáceos corresponden a rellenos de canal y delgados depósitos de llanura de inundación. En esta sección del perfil no aparecen niveles con evaporitas, en contraste con las facies que describe Irigoyen (1997) en la zona de Poterillos.

Figura 3. Columna estratigráfica de la Formación Mariño en Divisadero Largo, Mendoza, con indicación de los niveles fosilíferos / stratigraphic column of the Mariño Formation in Divisadero Largo, Mendoza, showing fossiliferous levels.
En Divisadero Largo, el techo de los Conglomerados Violáceos puede situarse en el último nivel pelítico rojizo que suprayace a bancos de areniscas conglomerádicas grises y violáceas (figura 3). A partir de allí, se inician las Areniscas Entrecruzadas, caracterizadas por bancos de areniscas castaño-amarillentas con delgadas intercalaciones pelíticas rojizas. Los fósiles de mamíferos han sido hallados en estos niveles basales del miembro medio. Los restos estudiados (nivel fosilífero 1 de la figura 3) provienen de facies arenosas de textura homogénea y estructura maciza (Sm, según nomenclatura de Miall, 1996). Estas facies se asocian con areniscas de estructura tabular (Sh) y areniscas con estratificación entrecruzada planar (Sp) de alto ángulo orientadas en diferentes direcciones. Otros fósiles de mamíferos, restos vertebrales aún no estudiados, provienen del nivel fosilífero 2 (figura 3), de facies arenosas medias que forman bancos lenticulares con clastos pelíticos (depósitos de lag ) en la base. En líneas generales, estas asociaciones de facies son similares a las que describe Irigoyen (1997) en el Toscal de Mariño (sudeste de Potrerillos), las cuales asigna a depósitos de dunas e interdunas eólicas asociadas con sistemas fluviales efímeros.
Microscópicamente, las rocas del nivel fosilífero 1 de la Formación Mariño son arenitas líticas-arcósicas ricas en piroxenos y escaso cemento calcítico. Muestran una fábrica relativamente abierta y clastos subredondeados. La estructura inorgánica del tejido óseo de los restos de mamíferos está formada por fluorapatita (colofana sensu lato ), la cual es incolora con polarizadores paralelos e isótropa y levemente birrefringente con polarizadores cruzados. En cavidades de este tejido se reconoce la presencia de clastos minerales procedentes de la roca portadora (figura 4). En contraste, las arenitas fosilíferas de la Formación Divisadero Largo exhiben un color pardo rojizo debido a la presencia de óxidos de hierro (hematita, limonita). Además, se caracterizan por la abundancia de calcita, la cual ha precipitado tanto entre los granos minerales como dentro de las cavidades vasculares y fracturas de los huesos y dientes de mamíferos (permineralización calcítica). Estas observaciones confirman diferencias mineralógicas entre las rocas fosilíferas de ambas formaciones, estrechamente vinculadas a diferentes procesos fosildiagenéticos que han sufrido los restos de mamíferos.

Figura 4. Microfotografía de un hueso de mamífero procedente de la Formación Mariño (nivel fosilífero 1) tomada mediante un microscopio petrográfico (X 5). La estructura inorgánica del hueso, formada por colofana (fluorapatita), es incolora con polarizadores paralelos. Nótese el relleno de una cavidad ósea por clastos minerales subredondeados de la arenisca portadora de los fósiles / microphotograh of mammal bone from Mariño Formation (fossiliferous level 1) taken by petrographic microscope (X 5). Inorganic bone structure, formed by collophane (fluorapatite), is colorless in plane polarized-light. Note the presence of subrounded mineral clasts infiltrated from the sandstone bone bed.
Estratigrafía de la sección inferior de la Formación Mariño en Divisadero Largo
Espesor
Descripción litológica, color según Rock-Color Chart (Goddard et al ., 1980) y nivel fosilífero de vertebrados (NF)
Base : Formación Divisadero Largo
Discordancia erosiva
Miembro inferior : conglomerados violáceos
0,3 m
Conglomerado clasto-soportado con matriz arenosa gris (5Y 6/1). Los clastos, principalmente vulcanitas, son redondeados y tienen entre 5 y 20 cm. Muestran inbricación hacia el sudoeste (220°).
1,0 m
Areniscas sabulíticas y areniscas medias grises (5Y 6/1).
2,1 m
Conglomerados clasto- y matriz- soportados intercalados con areniscas gruesas grises (5GY 6/1). Los conglomerados forman cuerpos lenticulares (~0,3-0,5 m) de base irregular. Los clastos, de composición vulcanítica y en menor proporción clástica arenítica, tienen 5 cm de diámetro promedio.
7,2 m
Areniscas gruesas y medias grises (5GY 6/1) con estratificación paralela. Hacia el techo aparece un banco de 0,4 m con estratificación entrecruzada planar.
0,3 m
Limoarcilitas pardo rojizas (5YR 3/4).
18,5 m
Areniscas medias gris amarillentas (10YR 6/2) que alternan con niveles arenosos gruesos de base irregular y estructura paralela.
0,3 m
Limoarcilitas pardo rojizas (5YR 3/4) con evidencias de bioturbación. Escasos intraclastos vulcaníticos (~5 mm de diámetro).
4,6 m
Areniscas medias grises (5Y 6/1) con estructura maciza. Hacia el techo aparece estratificación paralela.
6,8 m
Areniscas gruesas grises (5Y 4/1), con intercalaciones de limoarcilitas rojizas (10R 4/2) y moradas (5R 3/4) de 10-20 cm de potencia que gradan a areniscas conglomerádicas con clastos pelíticos y estratificación paralela.
1,0 m
Areniscas finas y limoarcilitas rojizas (10R 4/6), con evidencias de bioturbación.
Miembro medio : areniscas entrecruzadas
12,0 m
Areniscas medias y finas castaño amarillentas (10YR 6/2). Forman bancos (~0,3 m) con base irregular y estratificación paralela.
26,2 m
Areniscas medias grises (10YR 6/2) con estratificación entrecruzada planar de alto ángulo. Forman sets de 0,8 m de potencia que orientan su estratificación en distintas direcciones.
0,4 m
Limoarcilitas y areniscas finas rojo anaranjadas (5YR 5/6) con estructuras heterolíticas de pequeña escala.
11,7 m
Areniscas medias castaño grisáceas (10YR 4/2) de textura homogénea. Exhibe estratificación maciza en los bancos inferiores y estratificación paralela hacia el techo. NF 1: del banco arenoso basal proceden restos de mamíferos notoungulados Mesotheriidae.
3,0 m
Areniscas finas gris amarillentas en bancos de 10 cm intercaladas con limoarcilitas rojizas (5YR 4/4).
7,2 m
Areniscas medias castaño amarillentas (10YR 6/2) con estratificación entrecruzada planar y estratificación paralela.
1,3 m
Limoarcilitas y areniscas finas rojizas (10R 5/4) que forman delgados bancos granodecrecientes. Evidencias de bioturbación y grietas de desecación.
7,9 m
Areniscas medias y gruesas castaño amarillentas (10YR 5/4) con grietas de desecación.
3,4 m
Areniscas medias castaño amarilentas (10YR 5/4).
0,10 m
Limoarcilita pardo rojiza (10YR 4/2) de espesor variable y geometría lenticular.
2 m
Areniscas medias castaño amarillentas (10YR 6/2) en bancos lenticulares y granodecrecientes, que se alternan con areniscas finas. En la base del estrato arenoso aparecen clastos pelíticos grisáceos de pequeño tamaño (~2 cm). NF 2: restos de mamíferos en la base del estrato arenoso, asociados a los clastos pelíticos.
5 m
Areniscas medias castaño amarillentas (10 YR 6/2), con base ondulada irregular. Cuerpos multiepisódicos granodecrecientes de 1-2 m. Se intercalan con limolitas y areniscas finas.
Techo : hacia arriba, continúan las secuencias arenosas del Miembro Areniscas Entrecruzadas de la Formación Mariño.
Paleontología sistemática
Orden Notoungulata Roth, 1903
Familia Mesotheriidae Alston, 1876
Subfamilia Mesotheriinae Simpson, 1945
Mesotheriinae indet.
Figura 5
Material estudiado. El ejemplar estudiado, procedente de la Formación Mariño de Divisadero Largo, está depositado en la Colección de Paleontología del Museo de Ciencias Naturales y Antropológicas "Juan Cornelio Moyano" de Mendoza (MCNAM). Consiste en una serie yugal superior derecha incompleta (MCNAM- PV 3648) de un individuo adulto, cuyos elementos dentarios se encontraron fragmentados y sueltos; sólo el tercer molar (M3) se mantenía en un fragmento de maxilar y, junto a éste, se encontró la mayor parte del M2, un fragmento del M1 (ectolofo), el P4 y un fragmento del P3 (figura 5.A-F).
Descripción. La principal característica del material estudiado es la pequeña talla de los dientes, todos ellos con grado de desgaste avanzado.

Figura 5. Dentición superior (MCNAM-PV 3648) de Mesotheriinae indet. de la Formación Mariño, Divisadero Largo, Mendoza / upper dentition (MCNAM-PV 3648) of Mesotheriinae indet. from the Mariño Formation, Divisadero Largo area, Mendoza. A, P3, vista oclusal / occlusal view ; B, P4, vista oclusal / occlusal view ; C, P4, vista labial / labial view ; D, M2, vista oclusal / occlusal view ; E, M3, vista oclusal / occlusal view ; F, M3, vista labial (sin parastilo, tal como fue hallado) / labial view (incomplete molar, as found) ; G, M3 de " Trachytherus "? mendocensis de la Formación Divisadero Largo, vista oclusal / M3 of " Trachytherus "? mendocensis from the Divisadero Largo Formation, occlusal view. Escala gráfica: 0,5 cm / scale bar: 0.5 cm.
Los premolares, P3 y P4 (figura 5.A-B), tienen contorno claramente triangular, con el ángulo postero-labial bastante recto. No presentan surco lingual. Labialmente, hay un suave surco que marca el parastilo. La cara posterior carece de esmalte. Ésta es más recta en P3 que en P4. Se conservan restos de cemento sobre las paredes de esmalte (figura 5.C).
Los molares presentan el parastilo suavemente marcado. El pliegue del paracono está más marcado en el M2 y el ectolofo es más ondulado que en M1. Lingualmente, los lóbulos anterior y posterior del M2 convergen, pero no llegan a encerrar el lóbulo medio, que es casi tan largo como el anterior (figura 5.D). El M3 es similar al M2, pero con el parastilo un poco más proyectado anteriormente (rasgo que puede estar sobreestimado porque falta parte de la cara anterior; figura 5.E). En la cara posterior del M3, hay una concavidad media marcada. El lóbulo medio es bastante largo, casi como en M2. En ambos molares, hay restos de cemento rellenando los surcos linguales entre los lóbulos. El ectolofo del M3 está expuesto en su totalidad, con una longitud de 26 mm (en el centro) y una fuerte curvatura (figura 5.F).
Discusión y conclusiones
La atribución de los restos hallados a la Subfamilia Mesotheriinae se basa en la semejanza morfológica general con los mesoterinos miocenos conocidos, si bien es cierto que muchos de los caracteres que separan las dos subfamilias de mesotéridos (craneales y de dentición anterior) no son observables en el material estudiado. Por otra parte, si se comparan los dientes de la Formación Mariño con los correspondientes a Trachytherus spegazzinianus (Reguero, 1999), se ven relativamente distintos. El hecho de que se trate de un mesoterino y no de un traquiterino es más acorde con la edad miocena de los sedimentos portadores.
Con respecto al traquiterino conocido de Divisadero Largo, " Trachytherus ?" mendocensis (Simpson y Minoprio, 1949; Simpson et al ., 1962; Reguero, 1999), su holotipo (MCNAM-antes MHNM – PV 2494) está actualmente perdido. Reguero (1994, 1999) considera que la especie corresponde a un género nuevo, sin que aún haya sido descripto formalmente. Las figuras publicadas del holotipo muestran un P3 bastante triangular, como los P3-P4 estudiados; los autores sólo publicaron las dimensiones del M1. Además de este espécimen, hay un M3 (MLP 45-VII-10-2) que presenta bastante similitud con el de la Formación Mariño, si bien se observan algunas diferencias. El parastilo es más corto y el pliegue labial del paracono es más convexo, con un surco vertical anterior más marcado; asimismo, el metacono también queda más señalado por un fino surco labial posterior; la concavidad de la cara posterior es menos aguda que en el nuevo ejemplar (figura 5.G). Aunque el M3 de " T ." mendocensis es un fragmento corto, labialmente parece que la curvatura del ectolofo sería algo menos acusada. Las dimensiones son muy semejantes (tabla 1).
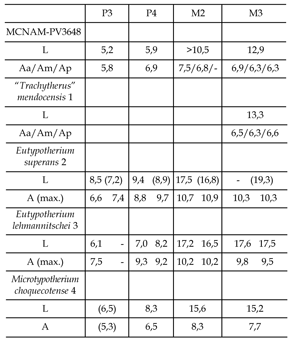
Tabla 1. Dimensiones comparadas de la dentición del mesotérido MCNAM PV-3648 de la Formación Mariño, Divisadero Largo (Mendoza). L, longitud máxima (mesio-distal); Aa, Am y Ap, ancho anterior, medio y posterior a nivel de cada lóbulo / tooth dimensions of the mesotheriid MCNAM PV-3648 from the Mariño Formation, Divisadero Largo (Mendoza). L, maximal length (mesio-distal length); Aa, Am y Ap, anterior, medial, and posterior width. 1, MLP 45-VII-10- 2, Divisaderense, Divisadero Largo (datos pers.) / Divisaderan, Divisadero Largo (pers. data) ; 2, MLP 26-VI-15-1, Friasense, Chubut, calco del holotipo (datos pers.) / Friasian, Chubut, cast of holotype (pers. data) ; 3, MLP 12-1701, "Colloncurense", Chubut, holotipo (datos pers.) / "Colloncuran", Chubut, holotype (pers. data) ; 4, Choquecota, Bolivia (Villarroel, 1974) / Choquecota, Bolivia (Villarroel, 1974) . MLP, Museo de La Plata / La Plata Museum.
Comparado con otros mesotéridos mesoterinos del Mioceno de Argentina, las dimensiones de nuestro espécimen son claramente menores que las de las especies de Eutypotherium del Friasense de Patagonia (tabla 1). El P3 es más pequeño y el P4 es similar o incluso más pequeño que el P3 de E. lehmannitschei . Los P3-P4 de la Formación Mariño son más anchos que largos tal como ocurre en el P4 de E . lehmannitschei , pero no en E. superans . El M3 es más pequeño que en las dos especies de Eutypotherium .
Los taxones más recientes, Typotheriopsis y Pseudotypotherium , son formas de talla más grande (Cerdeño y Montalvo, 2001).
Otros mesoterinos del Mioceno Medio sudamericano proceden de Bolivia y Chile. Villarroel (1974) describió Microtypotherium choquecotense como un nuevo género y especie del Altiplano Boliviano, caracterizado por ser el mesoterino más pequeño conocido. Los dientes encontrados ahora en la Formación Mariño son aún menores (tabla 1). Existen también algunas diferencias morfológicas respecto al material boliviano. Los premolares de Microtypotherium son de contorno más cuadrangular que los de la Formación Mariño y la pared antero-lingual es más convexa. El M2 tiene el ectolofo menos ondulado. El M3 parece presentar un lóbulo medio menos desarrollado que el M3 estudiado. Los niveles sedimentarios con Microtypotherium , Formación Choquecota, fueron correlacionados con los de edad Friasense de Argentina (Villarroel, 1974).
Respecto a los mesoterinos del Mioceno de Chile, Flynn et al . (2002) han descripto un conjunto faunístico de la Formación Chucal caracterizado por la abundancia de restos de mesoterinos. Los autores no describen en ese trabajo dichos restos, pero señalan la presencia de tres taxones (A, B y C), indicando que el taxón A es similar a Microtypotherium . También indican que el taxón B tiene un tamaño ligeramente menor y destacan el hecho de que el P4 es más ancho que largo como un carácter único entre los mesoterinos. Sin embargo, como se ha dicho antes, esto también ocurre en nuestro espécimen y en el P4 de Eutypotherium lehmannitschei . El taxón C es incluso más pequeño que los anteriores, pero tiene caracteres derivados como los P3-P4 cuadrangulares y con surco lingual. Este último carácter está bien desarrollado en Pseudotypotherium y Mesotherium y un surco más corto y menos marcado puede aparecer en Eutypotherium y Plesiotypotherium (Cerdeño y Montalvo, 2001). A partir de los datos radioisotópicos, Flynn et al . (2002) sugieren para la Formación Chucal una edad Mioceno Medio, entre 14 y 17,5 millones de años, pudiendo corresponder a la Edad-mamífero Friasense ( s.l .). Por su parte, Charrier et al . (2002) apoyan una edad santacrucense y Bond y García (2002) sugieren una edad máxima Colloncurense a partir de los restos estudiados de toxodóntidos procedentes de esa formación. Recientemente, Croft et al . (2004) confirman una edad santacrucense.
Considerando las diferencias establecidas, creemos que el ejemplar hallado en la Formación Mariño puede corresponder a un nuevo taxón, quizá relacionado con una de las especies de Chile. Teniendo en cuenta que el estudio detallado de estas formas chilenas (Croft et al ., 2004) ha sido publicado muy recientemente, cuando este manuscrito ya estaba enviado, la comparación en detalle con estos pequeños mesoterinos excede la finalidad de este trabajo. Dicha comparación permitirá establecer inferencias más concluyentes sobre la sistemática y la posición filogenética del taxón de Mendoza.
Asimismo, la semejanza global morfológica y métrica con respecto al traquiterino de la Formación Divisadero Largo, dado el hecho de que los ejemplares proceden de niveles distintos pero en una misma área, confiere un gran interés y plantea incógnitas sobre la verdadera relación entre ambos taxones y la peculiaridad del mesotérido divisaderense (G. López, com. pers.). Es necesario seguir buscando nuevos restos en ambas formaciones, que aporten datos precisos sobre la procedencia estratigráfica de " Trachytherus " mendocensis y proporcionen más caracteres morfológicos comparativos.
En cualquier caso, el material estudiado viene a aumentar el registro de la Subfamilia Mesotheriinae en el Mioceno Medio de Argentina, ampliando su distribución geográfica conocida fuera de Patagonia.
Agradecimientos
Expresamos nuestro especial agradecimiento a A. Acosta (Dirección de Recursos Naturales Renovables, Mendoza) por su dedicación y habilidad para encontrar fósiles durante sus trabajos de prospección en la Reserva de Divisadero Largo. A S. Roig, C. Montalvo y M. Reguero por la revisión crítica del manuscrito original, así como a los árbitros por sus valiosas sugerencias. A M. Galliski y F. Márquez Zavalía por su apoyo y toma de fotografías en microscopía petrográfica. Este trabajo ha sido parcialmente subsidiado por el CONICET y ha recibido apoyo de un proyecto de la ANPCyT.
Bibliografía
Barton, M. y Rodríguez, E. 1989. Geología del piedemonte de la Sierra de Uspallata al oeste de la ciudad de Mendoza. En: Detección y control de la desertificación, 2° Curso Latinoamericano de Desertificación (Mendoza) Actas : 154-159. [ Links ]
Biondi, J.L. 1936. [ Exploraciones geológicas en las inmediaciones del Cerro Cacheuta. Yacimientos Petrolíferos Fiscales, Buenos Aires. Inédito.]. [ Links ]
Bond, M. y García, M. 2002. Nuevos restos de toxodonte (Mammalia, Notoungulata) en estratos de la Formación Chucal, Mioceno, Altiplano de Arica, norte de Chile. Revista Geológica de Chile 29: 81-91. [ Links ]
Borello, A.V., Scanavino, R., Guichon, M. y Méndez, E. 1972. Mapa Tectónico de Mendoza. 5° Congreso Geológico Argentino (Córdoba), Actas 5: 3-9. [ Links ]
Bracaccini, I.O., 1950. Investigaciones tectónicas en la precordillera sanjuanina. Boletín de Informaciones Petroleras, 301 pp. [ Links ]
Cerdeño, E. y Montalvo, C.I., 2001. Los Mesotheriinae (Mesotheriidae, Notoungulata) del Mioceno superior de La Pampa, Argentina. Revista Española de Paleontología 16: 63-75. [ Links ]
Criado Roque, P. 1950. El Terciario del sur de la provincia de Mendoza. Revista de la Asociación Geológica Argentina 4: 233-255. [ Links ]
Croft, D.A., Flynn, J.J. y Wyss, A.R., 2004. Notoungulata and Litopterna of the early Miocene Chucal Fauna, Northern Chile. Fieldiana 50: 1-52. [ Links ]
Charrier, R., Chávez, A.N., Elgueta, S., Hérail, G., Flynn, J.J., Croft, D., Wyss, A.R. y García, M. 2002. Rapid tectonic and paleogeographic evolution: the Chucal Anticline, Altiplano of Arica, Northern Chile. IRD Collection. En: International Symposium of Andean Geodynamics , Toulouse, 5: 137-140. [ Links ]
Chiotti, O.V. 1946. [ Estratigrafía y tectónica del oeste de la ciudad de Mendoza y Las Heras. Tesis Doctoral, Universidad Nacional de Córdoba, Argentina, 2 v., 55 fot., 6 esq. map. Inédita.]. [ Links ]
Dessanti, R.N. 1942. [ Informe geológico de la zona del Cerro de la Cal, Río de las Peñas y Borbollón. Yacimentos Petrolíferos Fiscales, Buenos Aires. Inédito.]. [ Links ]
Flynn, J.J., Croft, D.A., Charrier, R., Hérail, G. and Wyss, A.R. 2002. The first Cenozoic mammal fauna from the Chilean Altiplano. Journal of Vertebrate Paleontology 22: 200-206. [ Links ]
Flynn, J.J. y Swisher, C.C.I. 1995. Cenozoic South American Land Mammal Ages: correlation to global geochronologies. En: W.A. Berggren, D.V. Kent, M.-P. Aubry y J. Hardenbol (eds.), Geochronology, time scales and global stratigraphic correlation. SEPM Soc. Sediment. Geol., Special Publication , 317-333 pp. [ Links ]
Francis, J.C., 1965. Los géneros de la subfamilia Mesotheriinae (Typotheria, Notoungulata) de la República Argentina. Boletín del Laboratorio de Paleontología de Vertebrados (Montevideo) 1: 1-31. [ Links ]
Goddard, E.N., Trask, P.D., De Ford, R.K., Rove, O.N., Singewald, J.T. y Overbeck, R.M. 1980. The Rock-Color Chart. Geological Society of America , Boulder.
Gorroño, R., Pascual, R. y Pombo, R.,1979. Hallazgo de mamíferos eógenos en el Sur de Mendoza. Su implicancia en la datación de los "Rodados Lustrosos" y del primer episodio orogénico del Terciario de la región. 7° Congreso Geológico Argentino (Neuquén, 1978), Actas 2: 125-136.
Groeber, P. 1951. La Alta Cordillera entre las latitudes 34° y 29° 30'. Instituto de Investigaciones de las Ciencias Naturales. Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Revista (Ciencias Geológicas) 1: 1-352.
Irigoyen, M.V. 1997. [ Magnetic polarity stratigraphy and geochronological constraints on the sequence of thrusting in the Principal and Frontal Cordillera and the Precordillera of the Argentine Central Andes (33° S latitude). Ph D. Thesis, Carleton University, Ottawa, Canadá, 392 pp. Inédita.].
Irigoyen, M.V., Buchan, K.L. y Brown, R.L. 2000. Magnetostratigraphy of Neogene Andean foreland-basin strata, lat 33°S, Mendoza Province, Argentina. Geological Society of America, Bulletin 112: 803-816.
Kokogián, D.A. y Mancilla, O.H.,1989. Análisis estratigráfico secuencial de la Cuenca Cuyana. En: G. Chebly y L. Spalletti (eds.), Cuencas Sedimentarias Argentinas . Facultad de Ciencias Naturales de La Universidad Nacional de Tucumán. Correlación geológica Serie 6: 169-201.
López, G. 2002. Redescripción de Ethegotherium carettei (Notoungulata, Hegetotheriidae) de la Formación Divisadero Largo de la Provincia de Mendoza, Argentina. Ameghiniana 39: 295-306.
Miall, A.D., 1996. The Geology of Fluvial Deposits. Springer-Verlag Berlin Heidelberg, New York, 583 pp.
Minoprio, J.L. 1947. Fósiles de la Formación del Divisadero Largo. Anales de la Sociedad Científica Argentina 146: 365-378.
Munsell, 1994. Soil Color Charts. Macbeth Division of Kollmorgan Instruments Corporation, New York.
Pascual, R. 1966. Paleontografía bonaerense. IV. Vertebrata. A.V. Borrello (ed.), La Plata, 202 pp., 101 láms.
Pascual, R. y De la Fuente, M.S. 1993. Vertebrados fósiles cenozoicos. En: V. Ramos (ed.), 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza, 1993). Geología y recursos naturales de Mendoza. Asociación Geológica Argentina e Instituto Argentino de Petróleo, Buenos Aires, pp. 357-363.
Pascual, R. y Odreman Rivas, O. 1973. Las unidades estratigráficas del Terciario portadoras de mamíferos. Su distribución y sus relaciones con los acontecimientos diastróficos. 5° Congreso Geológico Argentino (Villa Carlos Paz, 1972) Actas 3: 293-338.
Pascual, R., Ortega, E.J., Gondar, D. y Tonni, E.P. 1965. Las edades del Cenozoico mamalífero de la Argentina, con especial atención a aquéllos del territorio bonaerense. Anales de la Comisión de Investigación Científica de Buenos Aires 6: 165-193.
Patterson, B. 1952. Un nuevo y extraordinario marsupial Deseadiano. Museo Municipal de Ciencias Naturales y Tradicional de Mar del Plata, Revista, 1: 39-44.
Reguero, M.A. 1994. Consideraciones sobre el status sistemático de Trachytherus ? mendocensis Simpson y Minoprio, 1949 (Notoungulata, Mesotheriidae) de la Formación Divisadero Largo (Eoceno Superior?) de la provincia de Mendoza, Argentina. 6° Congreso Argentino de Paleontología y Bioestratigrafía (Trelew, Chubut), Resúmenes : p. 41.
Reguero, M.A. 1999. [ El problema de las relaciones sistemáticas y filogenéticas de los Typotheria y Hegetotheria (Mammalia, Notoungulata): análisis de los taxones de Patagonia de la Edad-mamífero Deseadense (Oligoceno). Tesis Doctoral. Universidad Nacional de Buenos Aires, Facultad de Ciencias Exactas y Naturales, 350 pp., 60 láms. Inédita.].
Rolleri, E.O. 1950. [ Informe Geológico de la Hoja 23c, Mendoza, Dirección General de Yacimientos Petrolíferos Fiscales. Inédito.].
Rolleri, E.O. y Criado Roque, P. 1970. Geología de la provincia de Mendoza, 4° Jornadas Geológicas Argentinas (Mendoza, 1969), pp. 1-60.
Rolleri, E.O. y Fernández Garrasino, C.A. 1979. Comarca septentrional de Mendoza, Segundo Simposio de Geología Regional Argentina. Academia Nacional de Ciencias (Córdoba), pp. 771-810.
Rusconi, C. 1946a. Ave y reptil oligocenos de Mendoza. Boletín Paleontológico de Buenos Aires 21: 3.
Rusconi, C. 1946b. Nuevo mamífero fósil de Mendoza. Boletín Paleontólogico de Buenos Aires 20: 1-2.
Rusconi, C. 1949. Los moluscos miocénicos de Cacheuta, Mendoza. Revista del Museo de Historia Natural 3: 237-239.
Sepúlveda, E. 1999. Datos palinológicos de la Formación Mariño en la localidad de Salagasta, Mendoza, Argentina. En: C. Náñez (ed.), Simposio "Paleógeno de América del Sur". IGCP Project 301 South American Regional Committee on Paleogene Stratigraphy. Subsecretaría de minería de la Nación, Servicio Geológico Minero Argentino, Anales , Buenos Aires 1996, pp. 75-78.
Simpson, G.G. y Minoprio, J.L. 1949. A new adianthine Litoptern and associated mammals from a Deseadan faunule in Mendoza, Argentina. American Museum Novitates 1434: 1-27.
Simpson, G.G., Minoprio, J.L. y Patterson, B. 1962. The mammalian fauna of the Divisadero Largo Formation, Mendoza, Argentina. Bulletin Museum of Comparative Zoology 127: 139-293.
Soria, M.F. 2001. Los Proterotheriidae (Litopterna, Mammalia), sistemática, origen y filogenia. Monografías del Museo Argentino de Ciencias Naturales , Buenos Aires 1: 167 pp.
Villarroel, C., 1974. Un Mésothériné nouveau (Notoungulata, Mammalia) dans le Miocène supérieur de Bolivie. Comptes Rendus de l´Académie de Sciences de Paris 279: 551-554.
Yrigoyen, M.R. 1993. Los depósitos sinorogénicos terciarios. En: V.A. Ramos (ed.), Geología y recursos naturales de Mendoza. 12° Congreso Geológico Argentino y 2° Congreso de Explotación de Hidrocarburos. Asociación Geológica Argentina e Instituto Argentino de Petróleo, Buenos Aires, pp. 123-148.
Zavattieri, A.M., Cerdeño, E., Bottero, R. y Londero, S. 2001. [ Estudio, prospección y propuesta de rescate y protección de yacimientos paleontológicos en el área de Potrerillos y zonas aledañas, Ministerio de Ambiente y Obras Públicas de Mendoza , Mendoza. Inédito.].
Recibido: 11 de noviembre de 2004.
Aceptado: 14 de julio de 2005.

