










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAmeghiniana
On-line version ISSN 1851-8044
Ameghiniana vol.43 no.2 Buenos Aires Apr./June 2006
Estratigrafía y mamíferos fósiles de la Formación Toro Negro (Neógeno), Departamento Vinchina, noroeste de la provincia de La Rioja, Argentina
Rafael Rodríguez Brizuela1 y Adán Tauber2
1 Museo Argentino de Ciencias Naturales, Av. Ángel Gallardo 470, 1405 Buenos Aires, Argentina. rrbrizuela@macn.gov.ar - rafarodriguezb@csnat.unt.edu.ar
2 Museo de Paleontología. Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Av. Vélez Sarsfield 299, 5000 Córdoba, Argentina.
Abstract. Stratigraphy and fossil mammals from the Toro Negro Formation (Neógene), Vinchina Department, northwestern La Rioja Province, Argentina. The Toro Negro Formation is exposed in the western range of the Sierra de Los Colorados, in the North-West of La Rioja Province, Argentina. The sequence in La Troya creek (type section) is composed by a succession of epiclastic and piroclastic deposits, where thick-bedded sandstones and mudstones with interbedded conglomeradic lenses and tuff levels predominate. The facies analysis suggests a fluvial depositional environment of sand bed braided rivers for the lower Member and high efficiency alluvial fans for the upper Member. The species recorded are: Proeuphractus limpidus Ameghino, Chorobates villosissimus (Rovereto) Reig, Paleuphractus sp. Bordas, Doellotatus inornatus (Rovereto) Castellanos, Lomaphorops corallinus Rovereto, Neuryurus cf. giganteus, cf. Urotherium sp. (Mammalia, Cingulata), Xyophorus aff. bondesioi Scillato Yané, cf. Plesiomegatherium, cf. Proscelidodon sp. (Mammalia, Tardigrada), Abrocoma sp., Protabrocoma sp., Propediolagus sp. (Mammalia, Rodentia), Eoauchenia sp. (Mammalia, Litopterna). This faunistic association suggests: 1) Lower Pliocene age; 2) climate conditions that changed from arid to warm-temperate, or the development of arid areas in a general context of more humid conditions; 3) open areas of herbaceous and shrub vegetation and arboreal formation probably existed; 4) the record of the nothrotherin Xyophorus aff. bondesioi is related to the faunistic and climatic changes during the Middle Miocene; 5) the existence of a zoogeographic connection between the subandean and Pampean regions during the Pliocene.
Resumen. La Formación Toro Negro aflora en el borde occidental de la sierra de Los Colorados, en el Noroeste de la provincia de La Rioja, Argentina. La secuencia en la quebrada de La Troya (sección tipo) está conformada por la alternancia de depósitos epiclásticos y piroclásticos, predominando areniscas y pelitas en potentes bancos estratificados y lentes conglomerádicas intercaladas, y niveles de tobas. El análisis de facies permitió inferir un ambiente de depositación fluvial caracterizado por ríos entrelazados de baja sinuosidad y carga dominantemente arenosa para el miembro inferior y de abanicos aluviales de alta eficiencia de transporte para el miembro superior. Los taxones reconocidos en este trabajo son: Proeuphractus limpidus Ameghino, Chorobates villosissimus (Rovereto) Reig, Paleuphractus sp. Bordas, cf. Doellotatus inornatus (Rovereto) Castellanos, Lomaphorops corallinus Rovereto, Neuryurus cf. giganteus, cf. Urotherium sp. (Mammalia, Cingulata), Xyophorus aff. bondesioi Scillato Yané, cf. Plesiomegatherium, cf. Proscelidodon sp. (Mammalia, Tardigrada), Abrocoma sp., Protabrocoma sp., Propediolagus sp. (Mammalia, Rodentia), Eoauchenia sp. (Mammalia, Litopterna). Este conjunto faunístico sugiere: 1) una Edad Plioceno Inferior; 2) condiciones climáticas que fueron inicialmente áridas y secas, y que cambiaron a templado-cálidas; o bien el desarrollo de subambientes áridos dentro de un contexto general de mayor humedad 3) que existirían áreas abiertas con vegetación herbácea y arbustiva y formaciones arbóreas más o menos cerradas; 4) la presencia del nothrotherino Xyophorus aff. bondesioi estaría relacionada a los cambios climáticos y faunísticos ocurridos a partir del Mioceno Medio; 5) una conexión zoogeográfica de la región Subandina con la región Pampeana para el Plioceno.
Key words. Stratigraphy; Mammals; Toro Negro Formation; Neogene; La Rioja;
Palabras clave. Estratigrafía; Mamíferos; Formación Toro Negro; Neógeno; La Rioja.
Introducción
En la provincia de La Rioja la mayoría de los estudios paleontológicos se refieren a restos procedentes de depósitos triásicos, aún cuando afloran en ella secuencias terciarias continentales de gran espesor. Este es el caso de las formaciones Vinchina y Toro Negro (Turner, 1964), aflorantes en la sierra de Los Colorados y que superan entre ambas los 10.000 m de potencia en la quebrada del Yeso (Ramos, 1970, 1999). Recientemente se han publicado nuevos hallazgos (Tauber, 2005; Georgieff et al., 2004) en el Mioceno de la Rioja, que confirman el potencial fosilífero en el Neógeno de la provincia.
La Formación Vinchina está compuesta principalmente por areniscas y pelitas, que son portadoras de niveles con vegetales, huellas de aves y de mamíferos. Estas trazas fueron asignadas por Frenguelli (1950) al Pérmico, pero más tarde, mediante estudios realizados en la quebrada del Yeso, Bonaparte (1965) las atribuye a mamíferos (Venatoripes riojanus, tardígrado de familia incierta y macrauquénidos) y aves reiformes, charadriformes y probablemente ralliformes, de posible edad miocena. Recientes estudios realizados en esta unidad (Marenssi et al., 2000 y Tripaldi et al., 2001) propusieron modelos de evolución y paleoambientes de depositación.
La suprayacente Formación Toro Negro es portadora de troncos indeterminados y elementos óseos de aves y mamíferos atribuidos al Plioceno por Ramos (1970) sobre la base del hallazgo de restos de Plesiomegatherium sp. Recientemente los mismos fueron revisados y atribuidos a una nueva especie de Pyramiodontherium (De Iullis et al., 2004). Posteriormente se realizaron nuevos hallazgos de vertebrados en la zona de la quebrada de La Troya (Arcucci et al., 1999; Rodríguez Brizuela, 2003 y Rodríguez Brizuela y Tauber, 2003a) y se realizaron los primeros estudios destinados a determinar el ambiente depositacional de la unidad (Rodríguez Brizuela y Tauber, 2003b).
Es necesario destacar que los vertebrados pliocénicos de Argentina más intensamente estudiados, provienen de los depósitos aflorantes en la costa atlántica de la provincia de Buenos Aires (Cione y Tonni, 1995) y, en menor medida, de restos procedentes de la provincia de Catamarca (Marshall y Patterson, 1981). Aportes realizados recientemente en la provincia de Córdoba (Tauber et al., 1997 y Tauber, 2000) han ampliado el conocimiento de la fauna de ese lapso de tiempo. Sin embargo es necesaria la prospección de nuevos yacimientos de vertebrados de otras regiones, lo que permitirá confrontar hipótesis y tener una visión más completa sobre aspectos geológicos, climáticos, ambientales, zoogeográficos, taxonómicos y paleobiológicos.
Para la realización de este trabajo se eligió la zona de la quebrada de La Troya, en el sector central de la sierra de Los Colorados, entre las localidades de Villa San José de Vinchina y Alto Jagüé (figura 1). Allí se encuentra la sección tipo de la Formación Toro Negro, con óptimos afloramientos para el levantamiento de perfiles estratigráficos.

Figura 1. Mapa de ubicación de la sierra de Los Colorados y localización del sitio de estudio. A la derecha, mapa geológico de la zona estudiada / locality map of the Sierra de Los Colorados and localization of the study place. At right, geological map of the study place.
Abreviaturas. NF es el nivel fosilífero de procedencia. En los casos de materiales que no lo posean se utilizó SNF (sin nivel fosilífero). Para los dientes de los roedores es: P4, premolar superior; M1, M2, M3, molares superiores; p4, premolar inferior; m1, m2, m3, molares inferiores. Los materiales se encuentran depositados en el Centro Regional de Transferencia Científica y Tecnológica La Rioja, bajo el acrónimo CRILAR-PZ.
Geología, estratigrafía y edad
La sierra de Los Colorados se dispone como una extensa faja de unos 30 km de ancho por unos 85 km de largo, siguiendo una dirección noreste-sudoeste. Esta unidad geomorfológica separa el valle de Vinchina, al este, del bolsón de Jagüé, al oeste (figura 1). La estructura general es la de un homoclinal de rumbo noreste-suroeste presentando bruscas inflexiones, como en la quebrada del Yeso donde los estratos adquieren un rumbo noroeste-sudeste y hasta norte-sur (Ramos, 1970). Las capas buzan 35° al oeste. En la sierra de Los Colorados aflora una extensa secuencia terciaria continental, correspondiente a las formaciones Vinchina y Toro Negro. Entre ambas superan los 10.000 metros de potencia (Ramos, 1970, 1999), conformando uno de los mayores espesores de terciario aflorante del mundo. La sección superior de la Formación Vinchina fue fechada radimétricamente en los 7,3±1,3 Ma y la toba basal de la Formación Toro Negro en los 4,3±1,0 Ma (Tabutt et al., 1989; véase Ramos, 1999).
La Formación Toro Negro es, litológicamente, muy homogénea. El perfil estratigráfico realizado en la quebrada de La Troya (sección tipo) tiene un espesor de 1.643 m para el miembro inferior de la formación (figura 2). Se caracteriza por la alternancia de sedimentos clásticos y piroclásticos. Los primeros consisten en una alternancia de potentes estratos de areniscas pardas y rojizas con niveles pelíticos verdes y marrones; eventualmente se intercalan cuerpos lentiformes de conglomerados gruesos. Los estratos de arenisca tienen entre 2 y 4 metros promedio de potencia y una geometría tabular. Se trata de areniscas finas, medianas y gruesas de moderada a buena selección y con predominio de matriz ferruginosa. Los niveles pelíticos se desarrollan a lo largo de todo el tramo estratigráfico y se presentan como cuerpos tabulares. Estos sedimentos están finamente laminados, en algunos casos se pudieron observar estructuras de calcos de carga y grietas de desecación en la superficie del estrato. Los cuerpos conglomerádicos son lentiformes y no presentan desarrollo horizontal significativo. Se trata de paraconglomerados medianos a gruesos, polimícticos, de aspecto masivo. Los clastos son subangulosos y están pobremente seleccionados, alcanzando los 15-20 cm de diámetro en su eje mayor. En general la matriz es arenosa y en algunos casos arcillosa.
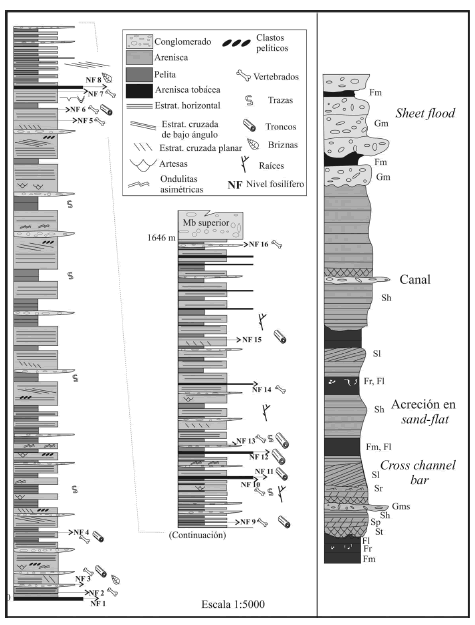
Figura 2. Columna estratigráfica del miembro inferior de la Formación Toro Negro en la quebrada de La Troya (sección tipo) con los niveles fosilíferos identificados y esquema de la secuencia de facies a la derecha / stratigraphic column of the lower Member of the Toro Negro Formation in Quebrada de La Troya (type section) with the fossiliferous levels and sketch of the facies sequence at right.
En la bibliografía se ha mencionado una aparente superficie de erosión que la separa de la infrayacente Formación Vinchina. Esta superficie no se identifica en el perfil, observándose un paso transicional de una unidad a otra. Para realizar el perfil se identificó el depósito tobáceo descrito por Ramos (1970), asumiéndose como la base de la Formación Toro Negro. En total se reconocieron nueve niveles tobáceos a lo largo del perfil. Los cuatro primeros tienen un espesor promedio de 5 metros, mientras que en los restantes el espesor disminuye considerablemente alcanzando los 2 metros de espesor promedio. La ubicación estratigráfica de cada nivel es: toba 1, base; toba 2 a 963 m, toba 3 a 1.193 m, toba 4 a 1.241 m, toba 5 a 1.369 m, toba 6 a 1.512 m, toba 7 a 1.546 m, toba 8 a 1.596 m y toba 9 a 1.611 m. Son de color claro, blanco o gris; en algunos casos presentan mezcla con material epiclástico.
El miembro superior de la Formaci ón Toro Negro comienza en una superficie de erosión que lo separa del miembro inferior. Está conformado por depósitos de conglomerados gruesos masivos pobremente seleccionados que se intercalan con bancos pelíticos oscuros. Los conglomerados son polimícticos, en parte grano sostén y en parte matriz soportado. Predominan clastos subredondeados a subangulosos con tamaños muy variados, desde pocos centímetros hasta 40-60 centímetros de diámetro en su eje mayor. En algunos casos presentan estructuras internas como clastos imbricados. Estos cuerpos conglomerádicos se presentan como cuerpos tabulares, de gran extensión areal y base erosiva.
Litofacies
En la secuencia estudiada se identificaron en forma preliminar las siguientes litofacies detríticas fluviales (tabla 1), la nomenclatura de las litofacies está basada en Miall (1992).
Tabla 1. Código de litofacies utilizado (basado en Miall, 1992) / Used litofacies code (based in Miall, 1992).
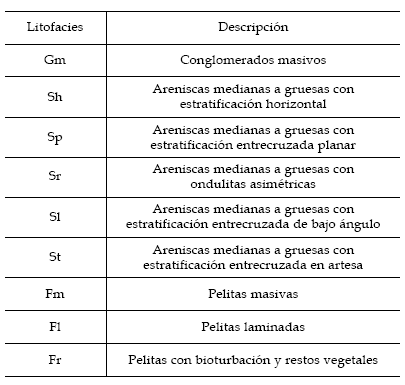
Miembro inferior. La facies psefítica está representada por el tipo Gm, correspondiente a depósitos de conglomerados matrizsoportados masivos. Esta litofacies se presenta en forma de cuerpos lenticulares que pueden alcanzar el metro de espesor y aparecen recurrentemente durante todo el desarrollo del perfil; el límite inferior de estos cuerpos es erosivo y no alcanzan un importante desarrollo horizontal.
Las facies psamíticas son las más abundante en el perfil, y son las que imprimen el aspecto general de la secuencia a gran escala. Los estratos alcanzan los dos metros de espesor, forman cuerpos tabulares muy potentes y de importante desarrollo horizontal. Son facies: Sm, correspondiente a depósitos de areniscas medianas a gruesas masivas; Sh, correspondiente a depósitos de areniscas medianas a gruesas con estratificación horizontal; y Sl, correspondiente a depósitos de areniscas finas a gruesas con estratificación entrecruzada de bajo ángulo. También están representadas las facies: St, correspondiente a depósitos de areniscas medianas a gruesas con estratificación entrecruzada en artesa de pequeña y mediana escala; Sr, correspondiente a depósitos de areniscas medianas a gruesas con ondulitas unidireccionales; y Sp, correspondiente a areniscas finas y medianas con laminación entrecruzada tabular planar.
Las facies pelíticas, al igual que las psamíticas, se presentan en forma de cuerpos tabulares de hasta dos metros de potencia que se intercalan durante todo el perfil. Representan el segundo tipo litológico más abundante, son facies: Fl, correspondiente a depósitos de limos y arcillas finamente laminadas siendo común encontrar asociadas a esta litofacies estructuras de escapes de fluidos (laminación convoluta); Fr, correspondiente a depósitos de limos y arcillas con restos de vegetales y bioturbación; y Fm, correspondiente a depósitos de limos y arcillas masivas.
Miembro superior. Predominan las facies conglomerádicas masivas (Gm), que en algunos casos presentan clastos imbricados. Estos depósitos conforman potentes cuerpos tabulares de gran extensión areal y de base erosiva, que se intercalan con facies finas de pelitas de aspecto masivo (Fm).
Asociación de facies e interpretación paleoambiental
Los elementos arquitecturales del sistema fluvial (nomenclatura basada en Miall, 1992) están representados por: canales (facies Gm, St, Sp y Sr), formas arenosas (facies St predominantemente), barras de gravas (facies Gm), arenas con estratificación paralela (facies Sh), depósitos finos de desbordamiento (facies Fm y Fl) y macroformas con "foreset" (facies Sh y Sl predominantemente).
En el miembro inferior se reconocieron tres asociaciones de facies: la primera corresponde a depósitos de llanura arenosa. Ella está constituida por bancos amalgamados de areniscas medianas y finas masivas o con estratificación horizontal y estratificación entrecruzada de bajo ángulo; la segunda asociación de facies se desarrolla en forma subordinada y corresponde a depósitos de canal. Está conformada por bancos de areniscas medianas y gruesas con estratificación entrecruzada en artesa de pequeña y mediana escala, areniscas medianas y finas con laminación entrecruzada tabular planar y lentes de conglomerados masivos, medianos y gruesos; la tercera asociación está representada por los depósitos finos, pelitas masivas, laminadas y con restos de plantas y bioturbación, correspondiente a facies de planicie de inundación.
Teniendo en cuenta la presencia de depósitos de canal, los potentes bancos de areniscas amalgamadas, la arquitectura fluvial caracterizada por cuerpos tabulares y la ausencia de cuerpos de acreción lateral, se interpreta un ambiente de depositación fluvial caracterizado por una red de canales entrelazados de baja sinuosidad y poca profundidad asociados a extensos complejos de llanura arenosa. La abundancia de formas arenosas con facies St indica que la carga de fondo estaría representada por un predominio de las arenas por sobre las gravas.
La llanura arenosa representa grandes áreas de acumulación de arenas que están emergidas durante el régimen bajo y moderado del río principal. En los canales se desarrollarían barras transversas que pueden alcanzar todo el ancho del canal, son en general barras someras y extendidas. Es probable que las areniscas con estratificación entrecruzada de bajo ángulo se formaran por migración de este tipo de barras. El bajo ángulo de inclinación de las capas reflejaría la poca pendiente de las barras, lo cual es una consecuencia directa de la escasa profundidad de los canales (Smith, 1971). Las areniscas con estratificación entrecruzada en artesa, reflejan la migración de ondas de arenas de crestas sinuosas que pudieron desarrollarse en el piso de los canales. La presencia de depósitos conglomerádicos gruesos, estaría representando una primera fase de mayor energía en la avenida fluvial. Al disminuir la capacidad de transporte del sistema, los materiales gruesos se habrían depositado conformando barras de gravas longitudinales. Estos depósitos están representados por cuerpos tabulares y lentiformes conformados por facies Gm.
Los depósitos finos (Fm, Fl y Fr), proponen la existencia de una extensa planicie de inundación con desarrollo de vegetación. Los restos de plantas identificados corresponden a equisetales y ciperáceas. En los niveles con bioturbación se reconocieron trazas fósiles típicas de la ichnofacies de Scoyenia (Taenidium, Beaconites y Skolithos), pedotúbulos y rizoconcreciones, las cuales se asociarían a paleosuelos. Éstos pudieron desarrollarse en la planicie de inundación o en áreas protegidas de intercanal con niveles freáticos altos (Heward, 1978).
En varios niveles del perfil se observaron clastos pelíticos de diferentes tamaños, los que evidencian la erosión por avenidas posteriores de la planicie de inundación y son indicadores de la alta cohesividad de estos depósitos.
En el miembro superior predominan las facies conglomerádicas masivas (Gm). Estos depósitos conforman potentes cuerpos tabulares que se intercalan con facies finas de pelitas masivas (Fm). Este miembro correspondería a facies medias y distales de abanicos aluviales de alta eficiencia de transporte (abanicos húmedos), con predominio de canales entrelazados y mantiformes. La existencia de estos potentes cuerpos conglomerádicos masivos, de base erosiva y con algunas estructuras internas producidas por corrientes tractivas (clastos imbricados), evidencia el desarrollo de flujos acuosos de alta energía. La geometría tabular y la gran extensión areal de los depósitos de conglomerados sugieren una ausencia de confinamiento en los flujos acuosos, los cuales habrían circulado en forma de arroyada en manto. Por otra parte es común la presencia de clastos de sobre medida que indican la existencia de flujos muy densos y de gran competencia. Corresponderían a depósitos de arroyada en manto de alta energía que localmente presentan encajonamiento (Heward, 1978).
La Formación Toro Negro, en su conjunto (miembros inferior y superior), corresponde a depositación sinorogénica asociada a un frente montañoso tectónicamente activo (frente orogénico andino; Ramos, 1999). El miembro superior representa la sedimentación en abanicos aluviales de alta eficiencia de transporte (abanicos húmedos) que pasan lateralmente a llanuras aluviales con desarrollo de canales entrelazados de baja sinuosidad, representadas por el miembro inferior (figura 3). El arreglo vertical de facies (figura 2) se debe a la progradación del sistema, producto del avance del frente orogénico.

Figura 3. Esquema del ambiente depositacional de la Formación Toro Negro. 1, frente tectónico; 2, abanicos aluviales; 3, llanura aluvial / sketch of the depositional environment of the Toro Negro Formation. 1, tectonic front; 2, alluvial fans; 3, alluvial flat.
Paleontología sistemática
Clase MAMMALIA Linneo, 1758
Superorden XENARTHRA Cope, 1869
Orden CINGULATA Illiger, 1811
Familia DASYPODIDAE Bonaparte, 1838
Subfamilia EUPHRACINAE Pocock, 1924
Tribu EUPHRACTINI Simpson, 1945
Género Proeuphractus Ameghino, 1886
Especie tipo. Proeuphractus limpidus Ameghino, 1886.
Proeuphractus limpidus Ameghino, 1886 Figura 4.A
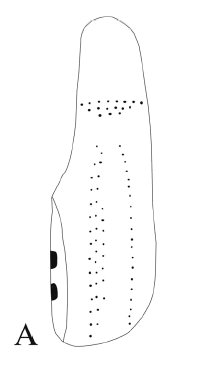
Figura 4. A, Proeuphractus limpidus Ameghino, CRILAR-PZ 251. Placa móvil / movable plate.
Material y procedencia. Dos placas de bandas móviles y tres mitades anteriores de placas de bandas móviles, CRILAR-PZ 251. El material procede del miembro inferior de la Formación Toro Negro, SNF.
Observaciones. Las placas presentan tres figuras longitudinales separadas por dos surcos paralelos y no alcanzan el área de articulación con la placa anterior. Los forámenes pilíferos son numerosos y se ubican en los márgenes laterales y posterior de las placas, lo cual es una característica distintiva de este género. Según Rovereto (1914) éstos nunca se ubican en los surcos. Las diferencias entre Proeuphractus y Paleuphractus se basan principalmente en la ubicación de los forámenes pilíferos y en que la figura central en Proeuphractus no es tan elevada como en Paleuphractus (Cerdeño, 2000). Este taxón fue registrado recientemente en el Mioceno Tardío de La Rioja (Tauber, 2003).
Género Chorobates Reig, 1958
Especie tipo. Chorobates recens Ameghino, 1887.
Chorobates villosissimus (Rovereto) Reig, 1958 Figura 4.B
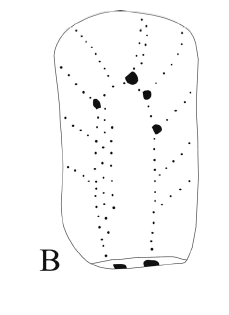
Figura 4.B, Chorobates villosissimus (Rovereto) Reig, CRILAR-PZ 252. Placa móvil / movable plate.
Material y procedencia. Seis placas de bandas móviles y varios fragmentos de placas móviles y fijas sueltas, CRILAR-PZ 252. El material procede del miembro inferior de la Formación Toro Negro, SNF.
Observaciones. La figura central está ligeramente elevada y presenta una cresta central no muy marcada. Esta característica permitió distinguir la especie Chorobates villosissimus de Chorobates recens, en esta última éste rasgo es más notorio (Esteban et al., 2001). Se identifican forámenes pilíferos en los surcos longitudinales y radiales. Se diferencia claramente un número variable de figuritas periféricas, definidas por surcos radiales a partir de la figura central.
Género Paleuphractus Kraglievich, 1934
Especie tipo. Paleuphractus argentinus Moreno y Mercerat, 1891.
Paleuphractus sp. Kraglievich, 1934 Figura 4.C

Figura 4.C, Paleuphractus sp., CRILAR-PZ 253. Placa escapular /scapular plate.
Material y procedencia. Doce placas del escudo escapular aisladas, CRILAR-PZ 253 y ocho fragmentos de placas de bandas móviles, CRILAR-PZ 254. El material procede del miembro inferior de la Formación Toro Negro, SNF.
Observaciones. Las placas del escudo escapular son pequeñas, poligonales cuadradas o subrectangulares. La superficie externa presenta una figura central de forma elíptica y siete figuras periféricas. Los forámenes pilíferos se ubican en las intersecciones de cada surco longitudinal con los surcos radiales. Las placas correspondientes a bandas móviles están rotas transversalmente, conservándose sólo la parte lisa anterior de la placa y un fragmento de la parte posterior en la cual se observa el desarrollo de tres figuras longitudinales separadas por dos surcos. La ubicación de los forámenes pilíferos permitió descartar al género Paraeuphractus en el cual éstos se encuentran casi exclusivamente en el borde posterior de la placa.
Los registros mio-pliocénicos del género Paleuphractus se distribuyen en Argentina hasta los 27° de latitud Sur (Carlini y Scillato Yané, 1996). El registro de Paleuphractus de este trabajo amplía la distribución geográfica del género.
Tribu EUTATINI Bordas, 1933
Género Doellotatus Bordas, 1932
Especie tipo. Doellotatus inornatus (Rovereto) Bordas, 1932.
cf. Doellotatus inornatus (Rovereto) Bordas, 1932 Figura 4.D

Figura 4.D, cf. Doellotatus inornatus (Rovereto) Bordas, CRILAR-PZ 255. Placa móvil / movable plate. Escala 1 cm / scale bar 1 cm.
Material y procedencia. Placa de una banda móvil, CRILAR-PZ 255. El material procede del miembro inferior de la Formación Toro Negro, NF 16. Las coordenadas del punto de extracción son 28° 41' 693 S y 68° 19' 712 O.
Observaciones. La placa presenta una figura central en relieve de forma ovalada que ocupa la mitad posterior de la placa y unas cuatro figuritas periféricas delimitadas por surcos radiales no muy marcados. La placa es de menor tamaño que aquellas de D. chapadmalensis y la figura central más angosta y con más relieve. Esta especie no se encuentra muy difundida en Argentina, solo presenta registros en la Formación Monte Hermoso (Scillato Yané et al., 1995) y en la Formación Cerro Azul del Mioceno Tardío de la provincia de La Pampa (Esteban et al., 2001).
Superfamilia GLYPTODONTOIDEA (Gray, 1869) Simpson, 1931
Familia GLYPTODONTIDAE Gray, 1869
Subfamilia SCLEROCALYPTINAE Trouessart, 1893
Tribu LOMAPHORINI Hoffstetter, 1958
Género Lomaphorops Castellanos 1931-32
Especie tipo. Lomaphorops corallinus (Rovereto) Castellanos 1931-32.
Lomaphorops corallinus (Rovereto) Castellanos 1931-32 Figura 5.A

Figura 5. A, Lomaphorops corallinus (Rovereto) Castellanos, CRILAR-PZ 256. Placas del caparazón dorsal / dorsal plates.
Material y procedencia. Treinta y nueve placas sueltas del caparazón dorsal, CRILAR-PZ 256; tres fragmentos de placas del escudo cefálico, CRILAR-PZ 257; siete placas del borde delantero de un anillo caudal, CRILAR-PZ 258; dos vértebras caudales fragmentadas, CRILAR-PZ 259. El material procede del miembro inferior de la Formación Toro Negro, NF 7. Las coordenadas del punto de extracción son 28º 41' 11" S y 68º 18' 16" O. Todos los materiales corresponden a un mismo individuo.
Observaciones. Las placas correspondientes al caparazón dorsal son poligonales hexagonales, pentagonales o subcuadradas. Presentan una figura central circular o ligeramente ovalada con una sensible depresión en su centro. Se observan numerosas perforaciones y surcos radiales que definen figuras periféricas, en algunos casos éstas no se diferencian y en otros se observan claramente hasta 12 y 14.
La asignación de género y especie del material del presente trabajo se alcanzó siguiendo las descripciones e ilustraciones de restos procedentes del valle de Santa María en la provincia de Catamarca (Castellanos, 1948). Recientemente se identificó este taxón en el Mioceno Tardío de La Rioja (Tauber, 2003) y en el Plioceno de Córdoba (Giannoncelli y Tauber, 1997).
Tribu NEURYURINI Hoffstetter, 1958
Género Urotherium Castellanos, 1926
Especie tipo. Urotherium simplex Castellanos, 1926.
cf. Urotherium sp. Castellanos, 1926 Figura 5.B

Figura 5.B, cf. Urotherium sp., CRILAR-PZ 260. Placa de un anillo caudal / caudal ring plate.
Material y procedencia. Placa de un anillo movible de la región caudal, CRILAR-PZ 260. El material procede del miembro inferior de la Formación Toro Negro, NF 13. Las coordenadas del punto de extracción son 28° 41' 798" S y 68° 19' 337" O.
Observaciones. Placa poligonal hexagonal con la superficie interna plana y rugosa. En la superficie externa se desarrolla una figura central con numerosas puntuaciones que abarca toda la superficie de la placa. No presenta figuras periféricas. Los bordes de la placa son de superficies muy rugosas. El largo es de 32,5 mm; el ancho de 20,0 mm; y el espesor de 10,7 milímetros.
El material es similar a aquél de Plohophorus, diferenciándose por la falta de figuras periféricas. Los registros del género se limitan al Plioceno Superior de la provincia de Buenos Aires.
Género Neuryurus Ameghino, 1889
Especie tipo. Nuryurus rudis (Gervais) Ameghino, 1889.
Neuryurus cf. giganteus Rovereto, 1914 Figura 5.C

Figura 5.C, Neuryurus cf. giganteus Rovereto, CRILAR-PZ 261. Placas del caparazón dorsal / dorsal plates.
Material y procedencia. Dos placas del caparazón dorsal soldadas y un fragmento de una tercera placa soldada a las otras dos, CRILAR- PZ 261. El material procede del miembro inferior de la Formación Toro Negro, NF 9. Las coordenadas del punto de extracción son 28° 41' 71" S y 68° 19' 19" O.
Observaciones. Las placas son gruesas y presentan forma rectangular. Se encuentran unidas por medio de una sutura discernible en la cara interna. La cara externa no presenta figuras definidas y su superficie es muy rugosa y uniformemente perforada por numerosos orificios dándole un aspecto esponjoso. El largo de las placas es de 43,4 mm, el ancho es de 31,8 mm, y el espesor de 28,0 mm.
Las placas estudiadas en este trabajo fueron asignadas tentativamente a Neuryurus cf. giganteus, teniendo en cuenta la descripción y las ilustraciones de Rovereto (1914, lámina XXXI, figura 3), esperando poder hallar más material que permita confirmar esta asignación.
Orden TARDIGRADA Latham y Davies, 1795
Superfamilia MEGATHERIOIDEA Owen, 1843
Familia MEGATHERIIDAE Owen, 1843
Subfamilia NOTHROTHERIINAE Ameghino,1920
Género Xyophorus Ameghino, 1887
Especie tipo. Xyophorus sulcatus Ameghino, 1887.
Xyophorus aff. bondesioi Scillato Yané, 1978 Figura 5.D

Figura 5.D, Xyophorus cf. bondesioi Scillato Yané, CRILAR-PZ 262. Dentario izquierdo en vista lateral / left dentary in lateral view.
Material y procedencia. Fragmento de dentario izquierdo, CRILAR- PZ 262. El material procede del miembro inferior de la Formación Toro Negro, NF 7. Las coordenadas del punto de extracción son 28º 41' 134" S y 68º 18' 255" O.
Observaciones. Está preservada casi la totalidad de la rama horizontal y parte de la rama vertical. Conserva implantados los cuatro dientes (un caniniforme y tres molariformes), de los cuales sólo el segundo molariforme está completo. En éste puede observarse la superficie oclusal, conformada por dos crestas transversales separadas por un valle transversal profundo. Tanto el primer como el segundo molariforme presentan surcos verticales, interno y externo, bien marcados y profundos. El cuarto alvéolo es de sección elíptico rectangular y dentro se conserva parte del tercer molariforme. Su cara lingual es redondeada y la cara labial plana, observándose el desarrollo de un surco vertical poco desarrollado. El orificio de la rama externa del canal dentario se ubica en la base de la raíz de la apófisis coronoides, ocupando una posición más interna y posterior que la del holotipo de Xyophorus bondesioi. La longitud total del dentario es de 63,1 mm, la longitud total de la serie dentaria es de 33,6 mm.
La presencia del primer diente (caniniforme) es característica de estos Nothrotherinos, en las formas más avanzadas se va reduciendo hasta desaparecer en Nothrothrium y Nothrotheriops. Xyophorus presenta marcadas diferencias con otros géneros del Mioceno Tardío o Plioceno Inferior. Se diferencia de Pronothotherium por la longitud de la región sinfisiaria que, en las formas más avanzadas, tiende a alargarse formando un canal longitudinal y por la ausencia en éste último de surcos verticales en el primer molariforme y en la cara interna del segundo. La forma del tercer molariforme que es típica de Xyophorus y de los Nothrotheriinae del Mioceno Tardío-Pleistoceno en general lo diferencia de formas más primitivas como Hapalops y Chasicobradys en los cuales el último molariforme es de sección subelíptica y sin surcos longitudinales.
Es característico de Xyophorus bondesioi la presencia del surco vertical en la cara labial del tercer molariforme, el cual está ausente en las demás especies del género; en el material descripto se observa un leve desarrollo del surco.
En Xyophorus bondesioi el orificio de la rama externa del canal dentario se ubica en la cara externa de la raíz de la apófisis coronoides, a semejanza de los Megatherioidea primitivos en general. La ubicación de dicho orificio en el material aquí descripto se asemeja a la de Chasicobradys, Senetia y Pronothotherium, es decir en la base del margen anterior de la rama ascendente, presumiéndose esta condición como más evolucionada (Scillato Yané et al., 1987).
Xyophorus tiene sus primeros representantes en las unidades estratigráficas de Edad "Santacrucense" (Mioceno Temprano y Medio) de Patagonia austral (Ameghino, 1889) y llegan hasta el "Chasiquense", Mioceno Tardío de la provincia de Buenos Aires (Scillato Yané, 1986), estando también registrado en la Formación Collón Curá, Mioceno Tardío de la provincia de Río Negro (Scillato Yané, 1978).
Subfamilia MEGATHERIINAE Owen, 1842
Género Plesiomegatherium Roth, 1911
Especie tipo. Plesiomegatherium hansmeyeri Roth, 1911.
cf. Plesiomegatherium Roth, 1911 Figura 5.F
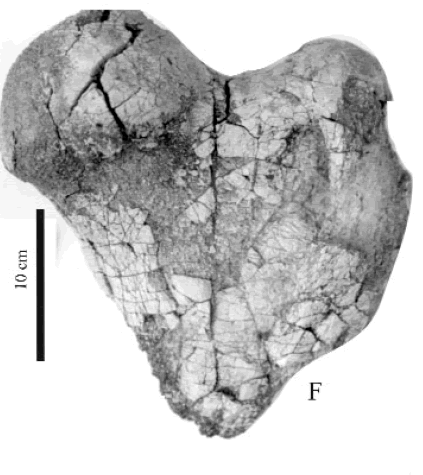
Figura 5.F, cf. Plesiomegatherium sp., CRILAR-PZ 263. Extremo proximal de fémur derecho en vista posterior / proximal extreme of right femur in posterior view.
Material y procedencia. Extremo proximal de fémur derecho, extremo distal de fémur derecho y extremo proximal de una costilla, CRILAR-PZ 263. Estos materiales proceden del miembro inferior de la Formación Toro Negro, NF 13. Las coordenadas del punto de extracción son 28° 41' 693" S y 68° 19' 712" O. Metacarpiano 4 izquierdo, CRILAR-PZ 264, procedente del miembro inferior de la Formación Toro Negro, SNF. Las coordenadas del punto de extracción son 28° 41' 20" S y 68° 18' 13" O.
Observaciones. El fragmento proximal de fémur derecho presenta un cuello estrecho y no muy profundo que separa la cabeza del fémur del trocánter; éste se desarrolla en un plano ligeramente por debajo de la cabeza. En la región de la diáfisis el hueso es aplanado en sentido ántero posterior. La cara posterior conserva la inserción del ligamento ileofemoral.
El metacarpiano se conserva casi completo y presenta evidencias de deformación por compresión lateral. Es un hueso robusto y de contorno surectangular. Está roto en el extremo distal conservando parte de la quilla la cual se inclina hacia la derecha en dirección palmar interna. En el extremo proximal se conservan las articulaciones para los carpianos. Su longitud total es de 168,9 mm; el ancho proximal es de 42,1 mm; el ancho distal es de 26,4 mm; el espesor proximal es de 44,5 mm; y el espesor distal es de 51,3 milímetros.
El material estudiado es demasiado fragmentario como para realizar una determinación genérica precisa. Fue asignado tentativamente como cf. Plesiomegatherium, teniendo en cuenta además que el género ya fue mencionado en la Formación Toro Negro (Ramos, 1970 y Arcucci et al., 1999). Por otro lado, no se descarta la posibilidad que correspondan a Pyramiodontherium (De Iullis et al., 2004).
Familia MYLODONTIIDAE Gill, 1872
Subfamilia SCELIDOTHERIINAE Ameghino, 1889
Género Proscelidodon Bordas, 1935
Especie tipo. Proscelidodon patrius Bordas, 1935.
cf. Proscelidodon sp. Figura 5.G
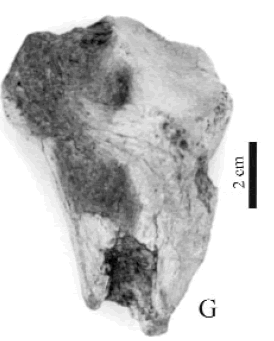
Figura 5.G, cf. Proscelidodon sp., CRILAR-PZ 265. Extremo proximal de fíbula derecha en vista lateral / fibula in lateral view.
Material y procedencia. Extremo proximal de fíbula derecha, CRILAR- PZ 265, fragmento del extremo proximal de la tibia, CRILARPZ 266, fragmento de un tarsal probablemente el primer cuneiforme, CRILAR-PZ 267. El material procede del miembro inferior de la Formación Toro Negro, NF 9. Las coordenadas del punto de extracción son 28° 41' 71" S y 68° 19' 19" O.
Observaciones. El género Proscelidodon surge al desdoblarse el género preexistente Scelidodon, de esta manera fue propuesto Proscelidodon para las especies de Scelidodon provenientes del "Hermosense", "Chapadmalalense" y algunas del "Pampeano inferior" (Bordas, 1935).
Si bien el material es fragmentario, la asignación tentativa de los materiales descriptos en este trabajo se realizó mediante la comparación con restos depositados en el Museo de Paleontología de la Universidad Nacional de Córdoba, siguiendo el criterio de Pascual et al. (1966), quienes dan como rasgo característico de Proscelidodon su talla, un tercio menor que las mayores especies de Scelidotherium y aproximadamente un quinto menor que Scelidodon. Por otro lado, ya se verificó la presencia de Proscelidodon en los valles andinos del norte argentino, en la Formación Chiquimil del Mioceno Superior en la provincia de Catamarca (Powell et al., 1998).
Orden RODENTIA Bowdich, 1821
Suborden HYSTRICOGNATHI Tullberg, 1899
Infraorden CAVIOMORPHA Wood y Patterson (en Wood, 1955)
Superfamilia CHINCHHILLOIDEA Kraglievich, 1940
Familia ABROCOMIDAE Miller y Gidley, 1918
Género Abrocoma Waterhouse, 1837
Especie tipo. Abrocoma benetti Waterhouse, 1837.
Abrocoma sp. Figuras 6.A-6.B
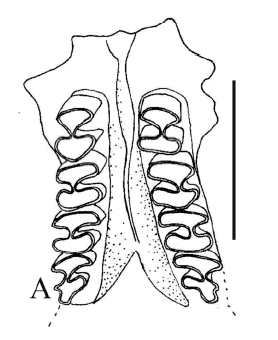
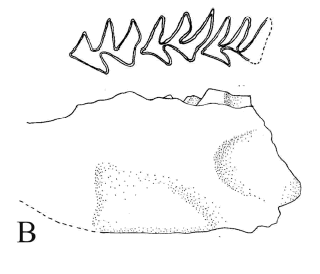
Figura 6. A-B, Abrocoma sp., CRILAR -PZ 272, CRILAR-PZ 268. Maxilar superior con las series P4-M3 izquierda y derecha en vista oclusal, dentario izquierdo con p4-m2 en vista lateral y oclusal / superior maxillary with right and left P4-M3 in occlusal view, left dentary with p4-m2 in lateral and occlusal view.
Material y procedencia. Fragmento de rama mandibular izquierda con p4-m1 y parte del m2, CRILAR-PZ 268, fragmento de rama mandibular izquierda con parte del incisivo y parte del p4, CRILAR- PZ 269, rama mandibular derecha con parte del incisivo y del p4, CRILAR-PZ 270, fragmento de rama mandibular izquierda con m1- m2 y parte del m3, CRILAR-PZ 271, fragmento de cráneo con el maxilar superior con ambas series P4-M3, CRILAR-PZ 272, once vértebras caudales, dos extremos distales de fémur, dos cabezas de fémur aisladas, dos extremos proximal de tibia, numerosas astillas óseas y fragmentos de extremidades, CRILAR-PZ 273. El material procede del miembro inferior de la Formación Toro Negro, SNF. Las coordenadas de los puntos de extracción son 28° 42' 02" S y 68° 18' 26" O; 28° 41' 11" S y 68° 18' 22" O.
Observaciones. Se describe a continuación los fragmentos de ramas mandibulares con las series dentarias por ser los materiales diagnósticos del género. Los molariformes son euhipsodontes, una capa continua de esmalte rodea a todos los dientes, la cual en algunas secciones se encuentra disminuida en su espesor. Los premolares y molares inferiores están compuestos por un repliegue externo y dos internos fuertemente oblicuos hacia adelante quedando cada diente inferior conformado por tres prismas triangulares de ángulos agudos presentando la parte anterior una forma de flecha en vista oclusal.
El fragmento de cráneo conserva las series P4-M3 izquierda y derecha. Estas series son más simplificadas que las inferiores, son bilobuladas por un repliegue interno y uno externo, siendo el interno más profundo y oblicuo hacia atrás. Las series derecha e izquierda son prácticamente paralelas y el diente más complejo es el M3 que presenta una prolongación adicional posterior. El hueso palatal es triangular. Rovereto (1914) postula que "los maxilares superiores de Abrocoma es difícil distinguirlos de los de Plataeomys, Pseudoplataeomys y Pithanotomys". Sin embargo al analizar el material surgen diferencias significativas dadas principalmente por la marcada forma de ocho en la dentición de estos últimos géneros, los molares están constituidos por dos lóbulos elípticos transversales más o menos comprimidos reunidos por un istmo que puede ser perfectamente opuesto como en Pithanotomys o estar ligeramente desplazados como en Pseudoplataeomys. Por otro lado, los lóbulos en algunos casos están cementados, característica que no se aprecia en los abrocómidos y no presentan el desarrollo del prolongamiento adicional en el M3.
Género Protabrocoma Kraglievich, 1927
Especie tipo. Protabrocoma paranensis Kraglievich, 1927.
Protabrocoma sp. Figura 6.C
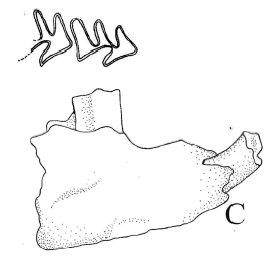
Figura 6.C, Protabrocoma sp., CRILAR-PZ 274. Dentario derecho con el incisivo, p4-m1 en vista lateral y oclusal / right dentary with the incisive and p4-m1 in lateral and occlusal view.
Material y procedencia. Fragmento de rama mandibular izquierda con p4-m1, CRILAR-PZ 274, fragmento de rama mandibular derecha con parte del incisivo, p4 y parte del m1, CRILAR-PZ 275. El material procede del miembro inferior de la Formación Toro Negro, SNF. Las coordenadas del punto de extracción son 28° 41' 17" S y 68° 18' 17" O.
Observaciones. La diferencia morfológica en la dentición entre Abrocoma y Protabrocoma está dada por las aristas de los molares inferiores las cuales son más angulosas en el primer género, además el p4 de Protabrocoma es más simple que el de Abrocoma (Pascual et al., 1966). Powell et al. (1998) citan a Protabrocoma para el Mioceno Superior de la provincia de Catamarca, en la Formación Chiquimil.
Superfamilia CAVIOIDEA Kraglievich, 1930
Familia CAVIIDAE Gray,1821
Subfamilia DOLICHOTINAE Pocock, 1922
Género Propediolagus Ortega Hinojosa, 1963
Especie tipo. Propediolagus marplatensis Ortega Hinojosa, 1963.
Propediolagus sp. Figuras 5.H, 6.D-E

Figura 5.H, Propediolagus sp., CRILAR-PZ 276. Cráneo y dentario izquierdo con el incisivo y la serie p4 - m3 en vista lateral / skull and left dentary with incisive and p4-m3 in lateral view.
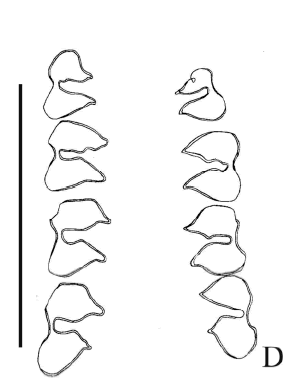

Figura 6.D-E, Propediolagus sp., CRILAR-PZ 276. Series p4-m3 y P4-M3 izquierda y derecha en vista oclusal / left and right p4-m3 and P4-M3 in occlusal view. Escala 1 cm / scale bar 1 cm.
Material y procedencia. La asignación del material se realizó siguiendo las descripciones de Ortega Hinojosa (1963). Cráneo casi completo, la región palatal conserva las series P4- M3 izquierda y derecha, la rama mandibular izquierda conserva el incisivo y la serie p4- m3, la rama mandibular derecha con la serie p4-m3 y el incisivo aislado, CRILAR-PZ 276, fragmento de cintura pélvica derecha, cinco segundas falanges, mitad proximal de una falange primera, húmero izquierdo, mitad distal del húmero derecho, cúbito izquierdo, radio izquierdo, extremo distal de fémur, mitad proximal de la tibia derecha, mitad proximal de la tibia izquierda, cinco mitades distal de metatarsos y una mitad proximal de metatarso, CRILARPZ 277. Estos materiales fueron hallados articulados y proceden del miembro inferior de la Formación Toro Negro, SNF. Las coordenadas del punto de extracción son 28º 41' 06" S y 68º 18' 18" O.
Observaciones. El cráneo es de contorno superior convexo y se aprecian claramente las suturas. Le falta la región anterior con los incisivos y tampoco se conservan los arcos cigomáticos. La ampolla auditiva izquierda es la única que se conserva y presenta el meato auditivo externo bien desarrollado. La morfología dental es simple. Los molariformes superiores e inferiores están compuestos fundamentalmente por dos prismas de sección cordiforme separados por un surco intreprismático lingual y labial. Los molariformes presentan el típico aspecto dolicotino, es decir son simétricos respecto a un imaginario plano interprismático normal a la serie dentaria. Los p4 y el M3 son los únicos elementos que presentan una mayor complejidad por el agregado de prismas accesorios.
La región palatal es de forma triangular. En todos los dientes se desarrolla un surco interno en forma de U que define dos prismas orientados oblicuamente hacia atrás. El surco externo es en general simple encontrándose enfrentado al externo, con la excepción de los M3 en los que se ubica desplazado hacia atrás. Los M3 desarrollan un prisma adicional posterior. Todos los molariformes superiores se encuentran rodeados por una capa continua de esmalte.
La rama mandibular izquierda conserva el incisivo y la serie p4-m3, faltándole la rama ascendente, en tanto que la rama mandibular derecha sólo conserva la serie p4-m3. Los dientes presentan un surco externo bien desarrollado y en forma de U que definen dos prismas. En los p4 se desarrolla un prisma adicional anterior no muy notorio. En las series dentarias inferiores la capa de esmalte es continua pero se adelgaza en las secciones convexas de los molariformes.
Los molariformes inferiores son semejantes a aquellos de Orthomyctera y Pediolagus, la diferencia está marcada por la profundidad del surco externo, la cual es menor que en Pediolagus y mayor que en Orthomyctera. El prisma accesorio anterior que se desarrolla en los p4 es semejante al homólogo de Orthomyctera pero considerablemente menor que aquel de Pediolagus. Es diagnóstico de Propediolagus la capa continua de esmalte que rodea a los molariformes así como su tendencia a adelgazarse en las secciones convexas. Además es característica la tendencia de desplazar el surco interprismático hacia el prisma posterior.
Género Eoauchenia Ameghino, 1887
Especie tipo. Eoauchenia primitiva Ameghino, 1887.
Eoauchenia sp. Figura 5.E
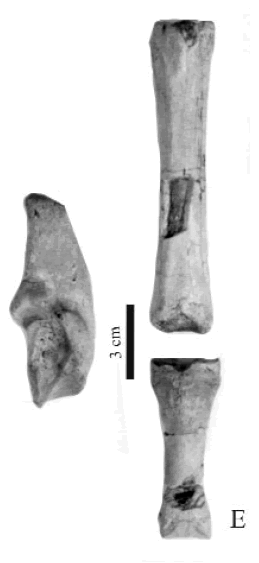
Figura 5. E, Eoauchenia sp., CRILAR-PZ 279. Metatarso III, falange proximal y calcáneo derecho / Metatarsian III, proximal phalange and right calcaneum.
Material y procedencia. Metatarsiano III sin la epífisis distal, falange proximal, calcáneo derecho, un tarsal, epífisis de la tibia, CRILAR-PZ 279. El material procede del miembro inferior de la Formación Toro Negro, NF 7. Las coordenadas del punto de extracción son 28° 41' 174" S y 68° 18' 261" O.
Observaciones. Los metapodianos III son algo más robustos que los de Thoatherium pero más gráciles que en los demás Proterotheriidae. Eoauchenia presenta las falanges proximales III con su longitud total triple al ancho proximal e igual a la mitad de la longitud de los metapodianos. La longitud total del metatarsiano es de 84,2 mm, la longitud total de la falange es de 48,4 mm y la longitud total del calcáneo es de 58,9 milímetros.
En la región andina sólo se ha citado la presencia de proterotheridos del Plioceno en la provincia de Catamarca (Rovereto, 1914; Marshall y Patterson, 1981; Esteban y Nasif, 1999).
Discusión y conclusiones
Edad
Los trabajos sobre bioestratigrafía y geocronología del Terciario del centro oeste del territorio argentino son escasos. En general fueron referidos a localidades clásicas de la paleontología argentina en particular al valle de Santa María y Corral Quemado en la provincia de Catamarca (Marshall y Patterson, 1981).
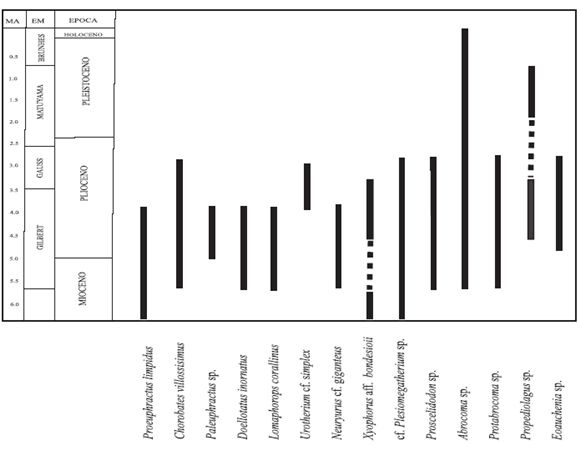
Los taxones registrados en este trabajo y que fueron citados en el Terciario de Catamarca son: los dasipódidos Chorobates villosissimus, Proeuphractus y Paleuphractus, los Gliptodóntidos Lomaphorops corallinus y Urotherium, los Megaterinos Plesiomegatherium, los Scelidoterinos Proscelidodon sp., y entre los roedores se ha citado a la Familia Abrocomidae representada por el género Protabrocoma. A este registro hay que agregar la presencia de Thylatheridium dolgopolae que también fue hallado en la Formación Toro Negro (Arcucci et al., 1999).
Estos materiales presentan en su gran mayoría el problema de la falta de control estratigráfico y geográfico, problema que es común para todo el material proveniente del tradicional piso "Araucanense" del valle de Santa María. De los materiales registrados en la Formación Toro Negro y citados en Catamarca por Marshall y Patterson (1981) Thylatheridium dolgopolae es mencionado como procedente de la unidad XX de la sección de Chiquimil. De esta misma sección proceden los géneros Chorobates y Protabrocoma, ambos ubicados en las unidades XVII y XX. En la sección de Corral Quemado mencionan la presencia de los dasipódidos Paleuphractus, procedente de las unidades XVIIIy XXXII, y Chorobates procedente de la unidad XXIV. Estos niveles son referidos a la Formación Andalhuala (Bossi et al., 1987) de Edad "Huayqueriense- Montehermosense" (Mioceno Superior- Plioceno Inferior). En la Formación Chiquimil asignada al Mioceno Superior, Powell et al. (1998) citan la presencia de Chorobates sp., Paleuphractus sp., Proscelidodon sp. y Protabrocoma sp.
En el "Huayqueriense" (Mioceno Superior) de la provincia de La Pampa están registrados los dasipódidos Proeuphractus, Chorobates villosissimus y Doellotatus inornatus (Esteban et al., 2001), que tienen registros hasta el "Montehermosense". El gliptodóntido Neuryurus giganteus sólo fue registrado hasta el momento en el piso Tunuyanense, Plioceno Inferior de la localidad típica de Huayquerías de Mendoza. Por su parte Urotherium tiene registros en el "Chapadmalalense" de la provincia de Buenos Aires (Pascual et al., 1966).
El notroterino Xyophorus bondesioi fue registrado por primera vez en la Formación Arroyo Chasicó en el suroeste de la provincia de Buenos Aires (Scillato Yané, 1978) asignándole una Edad "Chasiquense" (Plioceno Temprano, actualmente Mioceno Tardío). El cávido Propediolagus tiene su primer registro en las barrancas del suroeste de Mar del Plata (Ortega Hinojosa, 1963), aunque no se especifica la procedencia estratigráfica del material, fue atribuida con dudas al "Chapadmalalense", Plioceno Medio-Superior; sin embargo, Vucetich y Verzi (1995) proponen una Edad Ensenadense para esos mismos restos. El proterotérido Eoauchenia está registrado en la provincia de Buenos Aires, Bond et al. (1995) asignan al "Montehermosense" la especie E. primitiva mencionando Eoauchenia sp. en el "Chapadmalalense".
Las dataciones radimétricas realizadas en la Formación Toro Negro son conflictivas. Recientes edades K/Ar arrojaron valores de 8,6 ± 0,3 Ma y 6,8 ± 0,2 Ma (Ciccioli et al., 2005) para niveles de tobas en la sección superior de la unidad, mientras que Tabutt et al. (1989, véase Ramos, 1999) la ubicaban en los 4,3 ± 1,0 Ma. Teniendo en cuenta lo expresado en los párrafos anteriores respecto de la distribución estratigráfica de los taxones identificados en este trabajo y por superposición de los mismos (tabla 2), se sugiere una edad pliocena temprana para la Formación Toro Negro, en congruencia con las edades radimétricas publicadas por Tabutt et al. (1989). La edad de la Formación Toro Negro es un claro ejemplo de la dicotomía que muchas veces surge al combinar métodos bioestratigráficos y radimétricos. La presencia de Xyophorus en sedimentos de la edad propuesta podría estar evidenciando la subsistencia de una fauna relictual en la Formación Toro Negro relacionada con el importante recambio faunístico como consecuencia de los cambios climáticos-ambientales ocurridos en el continente a partir del Mioceno Medio (Goin et al., 2000 y bibliografía allí citada). Otro trabajo en preparación tiene como objetivo analizar la implicancia de este hallazgo.
Tabla 2. Distribución temporal de los taxones registrados / temporal distribution of the recorded taxa.
Paleoecología
La abundancia de vertebrados hervíboros, numerosos troncos, briznas y trazas de invertebrados evidencian una intensa actividad biológica y el desarrollo de vegetación en las planicies fluviales.
En los valles del centro oeste argentino las condiciones bióticas habrían sido de tipo "chaqueñas" durante el Mio-Plioceno (Scillato Yané, 1975; Kay y Madden, 1997). Los Xenarthra registrados en la Formación Toro Negro sugieren en general la exis tencia de condiciones templado-cálidas, con áreas abiertas de pastizales y formaciones arbóreas más o menos cerradas (Scillato Yané et al., 1995).
El género extinto Xyophorus habría estado relacionado con el actual perezoso arbóreo Bradypus (Scillato Yané, 1978), el cual por su muy imperfecta termorregulación está restringido a ámbitos tropicales y subtropicales. Xyophorus habría requerido por lo menos condiciones templado-cálidas. Los Nothrotheriinae fueron de régimen alimentario folívoro y es probable que hayan sido de hábitos arbóreos o semiarbóreos, la existencia de niveles con troncos fósiles encontrados en la Formación Toro Negro sería congruente con esta hipótesis.
Los proterotéridos en general han sido considerados como equivalentes ecológicos de los équidos y los Artiodactyla (Cervidae y Camelidae). Actualmente se los considera como formas cursoriales aunque no necesariamente de ambientes abiertos (Bond et al., 1995). Dos aspectos referidos a la morfología pedal de esta familia cobran interés en el análisis paleoecológico: el número de dedos funcionales y la gracilidad de las extremidades. Las formas tridáctilas (conservadoras) estarían adaptadas a suelos blandos y húmedos, a diferencia de las formas monodáctilas (e.g. Thoatherium del Mioceno Temprano de Patagonia), adaptadas a suelos más duros y secos. La gracilidad o robustez de los miembros tendría un significado adaptativo similar al de los équidos.
Las extremidades largas y gráciles indican tipos más corredores y ambientes abiertos mientras que los miembros más cortos y robustos se relacionan con ambientes boscosos. Eoauchenia del Plioceno es un proterotérido de morfología pedal conservadora, es tridáctilo y de metápodos largos y gráciles. Habría estado adaptado para la carrera en terrenos blandos y húmedos en áreas abiertas.
Otro aspecto que cobra interés en el análisis paleoecológico es el tipo de crecimiento y la altura de las coronas dentales (véase Mones, 1982; Tauber, 1997 y bibliografía allí citada) en los mamíferos consumidores de vegetales. La fauna reconocida en este trabajo en la Formación Toro Negro está compuesta predominantemente por formas euhipsodontes. Este tipo de adaptación está vinculada con una dieta compuesta por frutos, hojas, tallos, raíces y principalmente por hierbas que contienen elementos abrasivos relacionándose indirectamente con ambientes con vegetación abierta.
Los roedores abrocómidos y cávidos se encuentran en los niveles más bajos. Esta ubicación posiblemente se relacione a condiciones más secas (Vucetich y Verzi, 1995) durante el comienzo de la depositación de la Formación Toro Negro o bien al desarrollo de subambientes dentro de un contexto general más húmedo. Por otro lado, la comparación con registros de la región Pampeana sugiere que Chorobates, Doellotatus, Urotherium, Proscelidodon, Plesiomegatherium, Propediolagus y Eoauchenia indicarían una conexión zoogeográfica de la región Subandina con la región Pampeana, conexión ya propuesta por Scillato Yané (1975) y Scillato Yané et al. (1995).
Agradecimientos
Este trabajo es dedicado por el primer autor (RRB) a su padre (Fernando Rodríguez Brizuela), madre (María Haydeé Brandan) y hermanos. También agradece especialmente a Luis Benedetto por haber motivado en primera instancia la publicación de este trabajo y a Patricia Ciccioli por las numerosas y formativas charlas mantenidas sobre la Formación Toro Negro. Los autores agradecen a José Di Ronco por la invalorable colaboración durante las tareas de campo, sin cuyo apoyo no hubiera sido posible realizar este trabajo. Se agradece la lectura crítica del manuscrito a G. Esteban y los oportunos aportes realizados por los revisores G. Re, A. Carlini y C. Marsicano. Este trabajo se realizó en el Centro Regional de Investigación Científica y Transferencia Tecnólogica de la Rioja (Anillaco), en el Museo de Paleontología de la Facultad de Ciencias Exactas, Físicas y Naturales de la Universidad Nacional de Córdoba y en el Museo de Ciencias Naturales "Bartolomé Mitre" de la ciudad de Córdoba.
Bibliografía
Ameghino, F. 1886. Contribución al conocimiento de los mamíferos fósiles de los terrenos antiguos de Paraná. Boletín de la Academia Nacional de Ciencias (Córdoba), 9: 5-228.
Ameghino, F. 1887. Apuntes preliminares sobre algunos mamíferos extinguidos del yacimiento de Monte Hermoso. Imprenta "El Censor", 20 pp. Buenos Aires.
Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias (Córdoba) 6: 32-1028.
Arcucci, A.B., Tambussi, C. P., Pardiñas, U. F. y Petrulevicius, J. F. 1999. Nuevos restos de vertebrados en el terciario superior del noroeste de La Rioja, Argentina. 14° Jornadas Argentinas de Paleontología de Vertebrados, Ameghiniana 36: 95-96.
Bonaparte, J. 1965. Nuevas icnitas de la Quebrada del Yeso (La Rioja) y consideraciones acerca de la edad de los afloramientos. Acta Geológica Lilloana 7: 5-16.
Bond, M., Cerdeño, E. y López, G. 1995. Los ungulados nativos de América del Sur. En:. M.T. Alberdi, G. Leone y E.P. Tonni (eds.), Evolución biológica y climática de la Región Pampeana durante los últimos cinco millones de años: 259-275. Madrid.
Bordas, A. F. 1932. Proposición de un nuevo género para "Eutatus inornatus". Physis 11: 167-168.
Bordas, A.F. 1935. Observaciones sobre los géneros "Scelidodon" Amegh. y "Proscelidodon" n.g. Physis 11: 484-491.
Bossi, G.E., Ovejero, R. y Strecker, M. 1987. Correlación entre los perfiles del Terciario Superior en la Puerta de Corral Quemado-Hualfin y de Entre Ríos (Chiquimil), provincia de Catamarca, Argentina. 10° Congreso Geológico Argentino, Actas 2: 117-120.
Carlini, A.A. y Scillato Yané, G.J. 1996. Chorobates recens (Xenarthra, Dasypodidae) y un análisis de la filogenia de los Euphractini. Revista del Museo de La Plata (nueva serie), Paleontología 59/60, 9: 225-238.
Castellanos, A. 1948. Nuevos restos de "Lomaphorops corallinus (Rov.)". Publicaciones del Instituto de Fisiografía y Geología, Universidad Nacional del Litoral 34: 5-42.
Castellanos, A. 1926. Sobre un nuevo gliptodóntido chapadmalense. "Urotherium simplex" y las formas afines. Anales del Museo Nacional de Historia Natural de Buenos Aires "Bernardino Rivadavia" 34: 263-278.
Castellanos, A. 1931-32. Nuevos géneros de gliptodontes en relación con su filogenia. Physis 11: 99-100.
Cerdeño, E. 2000. Los mamíferos de la Formación Arroyo Chasicó (Mioceno superior), provincia de Buenos Aires, del Museo "J. C. Moyano" de Mendoza. Ameghiniana 37: 503-508.
Ciccioli, P.L., Limarino, C.O. y Marenssi, S. A. 2005. Nuevas edades radimétricas para la Formación Toro Negro en la Sierra de los Colorados, Sierras Pampeanas Noroccidentales, provincia de La Rioja. Revista de la Asociación Geológica Argentina 60: 251- 254.
Cione, A.L. y Tonni, E.P. 1995. Bioestratigrafía y cronología del Cenozoico superior de la Región Pampeana. En: M.T. Alberdi, G. Leone y E.P. Tonni (eds.), Evolución biológica y climática de la Región Pampeana durante los últimos cinco millones de años, pp. 49-74.
De Iullis, G., Ré, G.H. y Vizcaíno, S.F. 2004. The Toro Negro megatheriinae (Mammalia, Xenarthra): a new species of Pyramiodontherium and a review of Plesiomegatherium. Journal of Vertebrate Paleontology 24: 214-227.
Esteban, G.I. y Nasif, N.L. 1999. Mamíferos fósiles de la Formación Chiquimil (Mioceno tardío), provincia de Catamarca, Argentina. Bioestratigrafía. 15° Jornadas Argentinas de Paleontología de Vertebrados, Ameghiniana Suplemento Resúmenes 36: 11R.
Esteban, G.I., Nasif, N.L. y Montalvo, C.I. 2001. Nuevos registros de Dasypodidae (Xenarthra) del Mioceno tardío de la provincia de La Pampa, Argentina. Revista Española de Paleontología 16: 77-87.
Frenguelli, J. 1950. Ichnites en el Paleozoico Superior del oeste argentino. Revista de la Asociación Geológica Argentina 5: 136-148.
Georgieff, S.M., Herbst, R., Esteban, G.I. y Nasif., N. 2004. Análisis paleoambiental y registro paleontológico de la Formación Desencuentro (Mioceno Superior), Alto de San Nicolás, La Rioja, Argentina. Ameghiniana 41: 45-56.
Giannoncelli, R.M. y Tauber, A.A. 1997. Nueva localidad fosilífera del Plioceno en la provincia de Córdoba, República Argentina. Ameghiniana 34: 119.
Goin, F.J., Montalvo,C.I. y Visconti, G. 2000. Los marsupiales (Mammalia) del Mioceno Superior de la Formación Cerro Azul (provincia de La Pampa, Argentina). Estudios Geológicos 56: 101-126.
Heward, A.P. 1978. Alluvial fan sequence and megasequence models from the Stephanian A and B (La Magdalena, Ciñera- Metallana and Sabero) coalfields, northern Spain. Sedimentology 25: 451-488.
Kay, R.F. y Madden, R.H. 1997. Paleogeography and Paleoecology. En: R. Kay, R. Madden, R. Cifelli y J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press pp. 521-550.
Kraglievich, L. 1927. Nota preliminar sobre nuevos géneros y especies de roedores de la fauna argentina. Physis 8: 591-598.
Kraglievich, L. 1934. La antigüedad pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron. Imprenta "El Siglo Ilustrado", 136 pp. Montevideo.
Marenssi, S., Net, L., Caselli, A., Tripaldi, A. y Limarino, C.O. 2000. Hallazgo e interpretación de discordancias intraformacionales en la Formación Vinchina (Neógeno), quebrada de La Troya, La Rioja, Argentina. Revista de la Asociación Geológica Argentina 55: 414-418.
Marshall, L.G. y Patterson, B. 1981. Geology and geocronlogy of the mammal-bearing Tertiary of the valle de Santa María and río Corral Quemado, Catamarca province, Argentina. Fieldiana Geology, New Series 9: vii + 1-80.
Miall, A.D. 1992. Alluvial deposits. In: R.G. Walker y N.P. James (Eds.), Facies models: response to sea level change. Geological Association of Canada, pp. 119-142
Mones, A 1982. An equivocal nomenclature: What means hypsodonty? Palaontologisches Zeitschrift 56: 107-111.
Ortega Hinojosa, E.J. 1963. Dos nuevos Caviidae de la región de Chapadmalal, datos complementarios a las diagnosis de otros Caviinae poco conocidos. Ameghiniana 3: 21-28.
Pascual, R., Ortega Hinojosa, E., Gondar, D.E. y Tonni, E. 1966. Paleontografía Bonaerense, Fascículo IV, Vertebrata (Ed. Borrello, A.V.). Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, La Plata, 202 pp., CI láminas.
Powell, J., Mulé, P., Duarte, R., Ortiz, P., Sanagua, J. y Muruaga, C. 1998. Mamíferos de la Formación Chiquimil (Mioceno Superior) de la vertiente occidental de la sierra de Hualfin, provincia de Catamarca. 11° Jornadas Argentinas de Paleontología de Vertebrados, Acta Geológica Lilloana 18: 178.
Ramos, V.A. 1970. Estratigrafía y estructura del Terciario en la Sierra de los Colorados (Provincia de la Rioja), República Argentina. Revista de la Asociación Geológica Argentina 25: 359- 382.
Ramos, V.A. 1999. Los depósitos sinorogénicos terciarios de la Región Andina. En: Geología Argentina. Anales del Instituto de Geología y Recursos Minerales de Buenos Aires 29: 651-682.
Reig, O.A. 1958. Notas para una actualización del conocimiento de la fauna de la Formación Chapadmalal. I. Lista faunística preliminar. Acta Geológica Lilloana 11: 241-253
Rodríguez Brizuela, 2003. Nuevos hallazgos de vertebrados fósiles en la Formación Toro Negro (Plioceno de La Rioja) e interpretación paleoambiental. 19° Jornadas Argentinas de Paleontología de Vertebrados, Ameghiniana Suplemento Resúmenes 40: 70R.
Rodríguez Brizuela, R. y Tauber, A. 2003a. Consideraciones paleoecológicas de la Formación Toro Negro (Plioceno, dpto. Vinchina, La Rioja) en base a sus vertebrados. Instituto Miguel Lillo, Serie Monográfica y Didáctica 42: 92.
Rodríguez Brizuela, R. y Tauber, A. 2003b. Análisis preliminar de las litofacies sedimentarias de la Formación Toro Negro (Plioceno), departamento Vinchina, La Rioja. Instituto Miguel Lillo, Serie Monográfica y Didáctica 42: 91.
Roth, S. 1911. Nuevo género de la familia "Megatheridae". Revista del Museo de La Plata 18: 1-21.
Rovereto, C. 1914. Los estratos araucanos y sus fósiles. Anales del Museo Nacional de Historia Natural de Buenos Aires 25: 1-250.
Scillato Yané, G.J. 1975. Nuevo género de Dasypodidae (Edentata, Xenarthra) del Plioceno de Catamarca (Argentina). Algunas consideraciones filogenéticas y zoogeográficas sobre los Euphractini. 1° Congreso Argentino de Paleontología y Bioestratigrafía, Actas 2: 449-461.
Scillato Yané, G.J. 1978. Nuevo Nothrotheriinae (Edentata, Tardigrada) de edad chasiquense (Plioceno Temprano) del sur de la provincia de Buenos Aires (Argentina). Su importancia bioestratigráfica, filogenética y paleobiogeográfica. 7° Congreso Geológico Argentino, Actas 2: 449-457.
Scillato Yané, G.J. 1986. Los Xenarthra fósiles de Argentina (Mammalia, Edentata). En: Simposio "Evolución de vertebrados cenozoicos", 4° Congreso Argentino de Paleontología y Bioestratigrafía, Actas 2: 151-156.
Scillato Yané, G.J., Carlini, A.A. y Vizcaíno, S.F. 1987. Nuevo Nothrotheriinae (Edentata, Tardigrada) de edad chasiquense (Mioceno Tardío) del sur de la provincia de Buenos Aires (Argentina). Ameghiniana 24: 211-215.
Scillato Yané, G.J., Carlini, A.A., Vizcaíno, S.F. y Ortiz Jaureguizar, E.O. 1995. Los Xenartros. En:. M.T. Alberdi, G. Leone y E.P. Tonni (eds.), Evolución biológica y climática de la Región Pampeana durante los últimos cinco millones de años, pp. 183-209. Madrid.
Smith, D.G. 1971. Pseudo-planar stratification produced by very low amplitude sand waves. Journal of Sedimentary Petrology 41: 69-73.
Tabutt, K.D., Naeser, C.W., Jordan, T.E. y Cerveny, P.F. 1989. New fissio-track ages of Mio-Pliocene tuffs in the Sierras Pampeanas and Precordillera of Argentina. Revista de la Asociación Geológica Argentina 44: 408-419.
Tauber, A.A. 1997. Paleoecología de la Formación Santa Cruz (Mioceno Inferior) en el extremo sudeste de la Patagonia. Ameghiniana 34:517-529.
Tauber, A.A. 2000. Las Caleras, provincia de Córdoba, primer yacimiento fosilífero de origen cárstico de Argentina. Revista Española de Paleontología 15: 233-244.
Tauber, A.A. 2005. Mamíferos fósiles y edad de la Formación Salicas (Mioceno tardío) de la sierra de Velasco, La Rioja, Argentina. Ameghiniana 42: 443-460.
Tauber, A.A., Cellenza, L. y Di Ronco, J. 1997. Aspectos tafonómicos del yacimiento fosilífero plioceno de Las Caleras, Departamento de Calamuchita, Córdoba, República Argentina. Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina, Ameghiniana 34: 125-126.
Tripaldi, A., Net, L., Limarino, C., Marenssi, S., Re, G. y Caselli, A. 2001. Paleoambientes sedimentarios y procedencia de la Formación Vinchina, Mioceno, noroeste de la provincia de La Rioja. Revista de la Asociación Geológica Argentina 56: 443-465.
Turner, J.C. 1964. Descripción geológica de la Hoja 15c, Vinchina, provincia de la Rioja. Boletín de la Dirección Nacional de Geología y Minería 100: 1-81.
Vucetich, M.G. y Verzi, D. 1995. Los Roedores Caviomorfos. En: M.T. Alberdi, G. Leone y E.P. Tonni (eds.), Evolución biológica y climática de la Región Pampeana durante los últimos cinco millones de años, pp. 213-225. Madrid.
Waterhouse, G.R. 1837. Mammalia, Part 2, Rodents. En: C. Darwin (ed.), The Zoology of the Voyage of H.M.S. Beagle, Under the Command of Captain Fitzroy, R.N., during the years 1832 to 1836. 97 pp. + 35 láminas. Londres. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 30 de octubre de 2003.
Aceptado: 17 de diciembre de 2004.

