










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.2 Buenos Aires abr./jun. 2006
Significado paleobiológico de agrupamentos (coloniais/gregários) de Conularia quichua Ulrich, 1890 (Cnidaria), Formação Ponta Grossa, Devoniano (Pragiano-Emsiano), Bacia do Paraná, Brasil
Sabrina C. Rodrigues1, Juliana M. Leme1 e Marcello G. Simões1
1Depto. Zoologia, IB, UNESP, CP 510, Rubião Júnior, 18618-000, Botucatu, SP, Brasil. scoelho@ibb.unesp.br; leme@ibb.unesp.br; btsimoes@ibb.unesp.br
Abstract. Paleobiological significance of clusters (clonal/gregarious) of Conularia quichua Ulrich, 1890 (cnidaria), Ponta Grossa Formation, Devonian (Gragian-Emsian), Paraná Basin, Brazil . The "Conularia beds" of the Ponta Grossa Formation (Devonian) of the Paraná Basin, southern Brazil, yield well-preserved specimens of Conularia quichua Ulrich and Paraconularia africana Sharpe. Many of these are preserved in life orientation. Also, one of the C. quichua specimens has five faces instead of four, providing additional evidence of a cnidarian affinity for conulariids. Conulariids occur in the Jaguariaíva Member (or Sequence B, transgressive system tract) containing several obrution deposits beneath marine flooding surfaces. Taphonomic data obtained from these beds show conclusively that both C. quichua and P. africana were epibenthic, sessile invertebrates originally oriented with their long axis perpendicular to the bottom and with their aperture opening upward. Of the 136 C. quichua specimens examined here, 125 occur isolated. Eleven of the C. quichua specimens collectively occur in five discrete clusters consisting of two or three specimens. All of the clustered specimens are fully inflated (exhibiting a rectangular transverse cross section) or slightly compressed longitudinally. In all of these specimens the apex is missing, and thus the problem of whether the clusters were clonal colonies or formed through preferential larval settlement cannot be resolved conclusively. However, in the single cluster consisting of three specimens, the specimens are oriented perpendicular to bedding, and thus they do not converge adapically. The three specimens are in contact with each other along the upper portion of their median region. These and the lack of any evidence of a sheet of budding stolons, suggest that this cluster was formed by preferential larval settlement.
Resumo. Camadas ricas em conulários da Formação Ponta Grossa (Devoniano) da Bacia do Paraná, sul do Brasil, contêm espécimes de Conularia quichua Ulrich e Paraconularia africana Sharpe bem preservados, muitos em posição de vida. Adicionalmente, um exemplar de C. quichua apresenta cinco faces verdadeiras (ao invés de quatro), constituindo evidência adicional para a afinidade entre conulários e cnidários. Os conulários em rochas do Membro Jaguariaíva (ou Seqüência B, um trato de sistemas trangressivos) que encerra diversos depósitos de sufocamento abaixo de superfícies transgressivas. Dados tafonômicos obtidos dos depósitos de sufocamento mostram que C. quichua e P. africana foram invertebrados sésseis de epifauna originamente orientados com seu eixo maior perpendicular ao fundo e com a região da abertura voltada para cima. Dos 136 espécimes de C. quichua examinados, 125 ocorrem isolados. 11 espécimes de C. quichua formam cinco agrupamentos, consistindo de dois ou três espécimes. Todos os espécimes estão inflados (exibindo seção transversal retangular) ou ligeiramente comprimidos longitudinalmente. Em todos os espécimes o ápice está faltando e, desse modo, o problema se, os agrupamentos são colônias clonais ou formados pelo assentamento preferencial de larvas, não pode ser solucionado. Entretanto, três espécimes de um único agrupamento, todos orientados perpendicularmente ao plano de acamamento, não convergem adapicalmente. Contato entre os espécimes se dá apenas ao longo da porção média/superior das tecas. Essas características, juntamente com a ausência de estolão laminar de brotamento, sugerem que, pelo menos esse agrupamento, foi formado por assentamento preferencial das larvas.
Key words. Conularia quichua; Paleoecology; Devonian; Ponta Grossa Formation; Paraná Basin.
Palavras chaves. Conularia quichua; Paleoecologia; Devoniano; Formação Ponta Grossa; Bacia do Paraná.
Introdução
Conulários são cnidários (mas veja Termier e Termier, 1949, 1953; Kozlowski, 1968; Oliver, 1984; Steul, 1984; Mortin, 1985; Feldmann e Babcock, 1986; Babcock e Feldmann, 1986a; Babcock, 1991, para uma interpretação distinta das afinidades desse grupo), extintos, bentônicos, sésseis de epifauna (Bischoff, 1978; Harland e Pickerill, 1987; Van Iten, 1987, 1991a, 1991b, 1992a, 1992b; Jerre, 1994; McKinney et al., 1995; Simões et al., 2000; Leme, 2002; Rodrigues, 2002; Rodrigues et al., 2003; Marques e Collins, 2004; Leme et al., 2004). Eles ocorrem em uma ampla variedade de fácies sedimentares, especialmente de águas rasas, plataformais, em ambientes siliciclásticos ou carbonáticos (McMenamin, 1987; Jerre, 1994; Van Iten, 1991b; Van Iten et al., 1996). Possuem teca fosfática, piramidal, alongada, flexível e, na maioria das vezes, com quatro faces (Babcock, 1991; Van Iten, 1991a; Leme, 2002; Leme et al., 2004). Esses invertebrados distribuem-se do Neocambriano (Hughes et al., 2000) ao Triássico (McMenamin, 1987), embora recentemente Ivantsov e Fedonkin (2002) e Van Iten et al. (2005) tenham sugerido possíveis ocorrências de formas aparentadas no Neoproterozóico. Foram, contudo, mais diversificados no Ordoviciano, incluindo, pelo menos, 12 dos 43 gêneros conhecidos para o grupo (Leme et al., 2003). Conulários são invertebrados tipicamente encontrados como formas isoladas, sugerindo hábito de vida solitário. Entretanto, diversos autores (tabela 1) documentaram a ocorrência de espécimes formando agrupamentos (clusters), indicando que algumas espécies podem apresentar também hábito gregário ou colonial. Em todos os agrupamentos descritos na literatura, no entanto, os espécimes estão preservados horizontais ou inclinados, em relação ao plano de acamamento e, na maioria das vezes, ocorrem achatados lateralmente (tabela 1). Com exceção de Paraconularia tenuis, que provêm da Escócia, as demais ocorrências documentadas na literatura são da América do Norte (tabela 1).
Tabela 1. Lista das ocorrências documentadas de formas agrupadas de conulários, incluindo os agrupamentos da Formação Ponta Grossa (Slater, 1907; Ruedemann, 1925; Sinclair, 1940; Babcock e Feldmann, 1986a, 1986b; Van Iten e Cox, 1992; Van Iten et al., 1996). O modo de preservação dos agrupamentos foi recuperado a partir das ilustrações, quando possível / list of documented occurrences of clustered conularids, including the Ponta Grossa Formation clusters (Slater, 1907; Ruedemann, 1925; Sinclair, 1940; Babcock and Feldmann, 1986a, 1986b; Van Iten and Cox, 1992; Van Iten et al., 1996). Preservation characteristics were obtained from the illustrations, when possible.

No presente artigo, são descritos agrupamentos (clusters) de Conularia quichua Ulrich in Steinmann e Döderlein, 1890 e seu possível significado paleobiológico é comentado. Tais ocorrências são provenientes das camadas de conulários da Formação Ponta Grossa ("Conularia beds", Simões et al., 2000; Rodrigues et al., 2003), as quais materializam depósitos de sufocamento (obrution deposits), onde os conulários estão preservados in situ, isso é, em posição de vida (Petri e Fúlfaro, 1983; Simões et al., 2000; Rodrigues et al., 2003).
Material e métodos
Geologia da área
O material aqui estudado provém de ocorrências da área de afloramentos da Formação Ponta Grossa, Devoniano (Pragiano-Emsiano, Grahn et al., 2000, mas veja Bergamaschi e Pereira, 2001 para uma interpretação diferente da idade da Formação Ponta Grossa), na Sub-bacia Apucarana, município de Jaguariaíva (figura 1). Na região de Jaguariaíva, PR, estão bem preservadas as rochas do chamado Membro Jaguariaíva (Lange e Petri, 1967) ou da Seqüência B (Bergamaschi, 1999; Bergamaschi e Pereira, 2001), referente ao ciclo de sedimentação caracterizado por depósitos marinhos rasos de face praial inferior e costa afora, respectivamente, shoreface e offshore. Os sedimentitos estudados localizam-se no corte da estrada de ferro Jaguariaíva-Arapoti, desde o km 2,2 (altitude de 860 m) até o km 6,6 (altitude de 960 m), conforme informações em Lange e Petri (1967). As coletas foram realizadas ao longo de toda a seção, sendo que os conulários ocorrem, preferencialmente, nos intervalos de 29 a 32 metros, a 39 metros e, de 44 a 48 metros do contato basal com a Formação Furnas (figura 2) (Simões et al., 2000; Rodrigues et al., 2003; Leme et al., 2004). Os pontos de coleta estão, respectivamente, situados próximos ao km 3,8 (24º14'05"S / 49º42'54"W) e ao km 4,5 (24º14'68"S / 49º43'19"W) da estrada de ferro.

Figura 1. Mapa de localização das ocorrências fossilíferas estudadas no Município de Jaguariaíva, Estado do Paraná, PR, Formação Ponta Grossa / location map of the studied fossil concentrations from Jaguariaíva County, Paraná State, Ponta Grossa Formation.

Figura 2. Seção colunar da Formação Ponta Grossa em Jaguariaíva, PR, modificada de Bergamaschi (1999) / columnar section of the Ponta Grossa Formation, Jaguariaíva, modified from Bergamaschi (1999).
Nos locais acima, predominam os depósitos de sufocamento (Simões et al., 2000; Rodrigues et al., 2003), os quais são progressivamente sucedidos por folhelho cinza escuro, laminado e siltitos e arenitos interlaminados, intensamente bioturbados (Bergamaschi e Pereira, 2001). Os folhelhos escuros correspondem a superfícies de inundação marinha, enquanto os siltitos e arenitos apresentam evidências da ação de ondas (Bergamaschi, 1999; Bergamaschi e Pereira, 2001).
Coleta e preparação dos blocos orientados de rochas
As amostras foram coletadas na forma de blocos de rocha, seguindo metodologia sugerida por Simões e Kowalewski (1998a, 1998b), posteriormente ampliada e modificada por Simões e Ghilardi (2000). Após uma detalhada inspeção visual, os siltitos da parede do afloramento, que continham conulários, foram demarcados e sua posição estratigráfica, bem como as coordenadas geográficas das ocorrências, anotadas. O trabalho envolveu a extração dos fósseis contidos em blocos de rochas, com volume aproximado de 0,002 m3 (veja Simões e Ghilardi, 2000). Uma vez extraídos do afloramento, os 97 blocos retirados (contendo um total de 114 espécimes) foram numerados e preparados de acordo com as técnicas usualmente utilizadas na preparação de macroinvertebrados fósseis (Feldmann et al., 1989). Finalmente, após as diversas etapas de preparação em laboratório, as feições sedimentares e tafonômicas presentes nos blocos de rocha foram descritas (veja Rodrigues, 2002; Rodrigues et al., 2003). Todo material coletado está depositado na Coleção Científica do Laboratório de Paleozoologia Evolutiva do Instituto de Biociências, UNESP, campus de Botucatu, Estado de São Paulo, sob a sigla DZP. Em adição, as Coleções Científicas da Universidade de São Paulo, da Universidade de Guarulhos, Estado de São Paulo e da Universidade Estadual de Ponta Grossa, Estado do Paraná, foram examinadas, as quais forneceram um total de 22 espécimes de conulários.
Modo de preservação
Os conulários da Formação Ponta Grossa estão preservados segundo três classes tafonômicas, reconhecidas por Simões et al. (2000) e Rodrigues et al. (2003). Esses autores identificaram tais classes a partir de evidências tafonômicas, sedimentológicas e estratigráficas. Abaixo, segue um resumo, fundamentado nesses autores.
Segundo Simões et al. (2000) e Rodrigues et al. (2003), a classe tafonômica 1 é representada por conulários isolados ou agrupados, verticalmente orientados (90º a 70º), em relação ao plano de acamamento. As tecas dos conulários apresentam-se infladas, com a região da abertura voltada para cima, preenchidas por material muito fino. Essas podem apresentar algum achatamento longitudinal, que, na maioria das vezes, é mais acentuado na região da abertura da teca. Com exceção dos espécimes que ocorrem agrupados, todos os espécimes isolados ocorrem completos, isso é, com as três regiões da teca (região da abertura, mediana e basal) preservadas. Os espécimes dessa classe tafonômica estão preservados tipicamente em siltitos maciços ou incipientemente laminados, às vezes, com icnofósseis isolados, discretos (índice de bioturbação 2, Miller e Smail, 1997), estando presentes acima ou abaixo dos estratos onde os espécimes horizontalmente preservados ocorrem.
Já a classe tafonômica 2 é representada por conulários isolados ou agrupados, inclinados (60º a 20º), em relação ao plano de acamamento (Simões et al., 2000; Rodrigues et al., 2003). Na maioria dos casos, os conulários apresentam tecas completas, cuja região da abertura dos espécimes está orientada para o topo, em relação ao plano de acamamento. Estão preservados inflados, podendo apresentar algum achatamento longitudinal ou lateral. Os conulários estão presentes em siltitos com icnofósseis isolados (e.g., Zoophycus, índice de bioturbação 2) ou zonas de generalizada bioturbação (índice de bioturbação 3, Miller e Smail, 1997).
Finalmente, a classe tafonômica 3 é representada por conulários isolados e horizontalmente orientados (20º ou menos), em relação ao plano de acamamento. Inclui tanto conulários completos e inflados, como incompletos e achatados (Simões et al., 2000; Rodrigues et al., 2003). Podem estar preservados em siltitos maciços ou intensamente bioturbados (índice de bioturbação 3 ou 4, Miller e Smail, 1997), com marcas onduladas ou, ainda, associados a estratificação cruzada hummocky ou a pavimentos métricos de conchas de braquiópodes articulados [e.g., Australospirifer heringi (Kayser)] (Simões et al., 2000; Rodrigues et al., 2003).
Especialmente os conulários representantes das classes tafonômicas 1 e 2, são encontrados em depósitos gerados abaixo do nível de base de onda de tempestade. Por tratar-se de invertebrado séssil de epifauna (Harland e Pickerill, 1987; Simões et al., 2000; Leme, 2002; Rodrigues, 2002; Rodrigues et al., 2003; Leme et al., 2004), sua preservação in situ implica, necessariamente, na ausência de correntes ou fluxos junto ao substrato. Aumento brusco nas taxas de sedimentação, por decantação de finos, em áreas mais distais, durante eventos de tempestades é o principal mecanismo gerador dos chamados depósitos de sufocamento (Miller et al., 1988; Brett e Seilacher, 1991; Brett, 1995; Brett et al., 1997). Por outro lado, representantes da classe tafonômica 3, cujas tecas ocorrem preservadas junto a estruturas sedimentares do tipo estratificação cruzada hummocky indicam que os conulários foram retirados de sua posição de vida ou dos sedimentos onde estavam preservados, sofrendo curto transporte e rápido soterramento ou sendo redepositados, acima do nível de base de ondas de tempestades (Rodrigues et al., 2003).
Resultados
Os conulários da Formação Ponta Grossa, Estado do Paraná, ocorrem como formas isoladas ou formando agrupamentos (clusters) de dois ou três espécimes de Conularia quichua, segundo as recentes revisões taxonômicas de Leme (2002), Siviero (2002) e Leme et al. (2004). De um modo geral, os conulários dos agrupamentos aqui descritos estão preservados como molde externo de uma ou mais faces ou, ainda, mantendo restos da teca, representados por uma película escura, onde estão preservadas as principais feições morfológicas externas da teca. Essas ocorrem orientadas com a região da abertura voltada para cima, infladas, podendo apresentar achatamento lateral ou longitudinal (veja Simões et al., 2003, para as formas de achatamento). O material que preenche a cavidade do corpo é muito fino, contendo, às vezes, bioclastos representados por conchas isoladas de bivalves. Embora, tipicamente, a base dos espécimes esteja ausente em todos os agrupamentos, é notável que os conulários convirjam para um centro comum, com a exceção da amostra DZP-3686, cujos espécimes tocam-se apenas na porção superior da região mediana, havendo um espaço entre os espécimes na porção inferior da mesma.
De um total de 136 espécimes estudados, apenas 11 conulários formam 5 agrupamentos, os quais estão preservados segundo as classes tafonômicas 1 (amostras DZP-3081, DZP-3632 e DZP-3686) e 2 (amostra DZP-3505 e DZP-3633) (Rodrigues et al., 2003).
O agrupamento representado pela amostra DZP- 3081 é constituído por dois conulários, com tecas incompletas, ambas representadas apenas pela região mediana da teca. As dimensões da teca são de 1,82 cm e de 2,19 cm, respectivamente, sendo o primeiro representado apenas por molde externo de uma face verdadeira, isso é, delimitada por duas arestas adjacentes. Desse modo, não é possível saber quantas faces verdadeiras formaram a teca desse conulário. Já o espécime maior preserva restos da teca e é constituído de cinco faces verdadeiras. Tal exemplar apresenta- se inflado na matriz, porém sua teca está achatada longitudinalmente, com dobras na porção superior da região mediana. Os dois espécimes de conulários estão preservados verticalmente, em relação ao plano de acamamento e paralelos entre si, tocando-se pelas faces ao longo de toda a sua extensão (figuras 3.A, 3.B).

Figura 3. Conularia quichua. A-B, DZP-3081. A, Vista lateral da região mediana de agrupamento constituído por, pelo menos, dois exemplares (setas brancas) / lateral view of the cluster median region with, at least, two specimens (white arrows);

Figura 3. Conularia quichua.B, Detalhe, em seção (vista inferior), do exemplar com cinco faces verdadeiras (detalhe em branco) / cross section detail (lower view) of a five sided specimen (white detail)
Outro agrupamento formado por dois conulários é representado pela amostra DZP-3632. Os espécimes apresentam tecas incompletas, ambas representadas apenas pela região mediana, com dimensões de 5,12 cm e de 4,43 cm. Os conulários mantêm restos da teca preservada e constituída de quatro faces verdadeiras. As tecas dos espécimes estão totalmente infladas, isso é, sem achatamento visível. Assim como observado no agrupamento anteriormente descrito, os dois espécimes de conulários estão preservados verticais, em relação ao plano de acamamento e paralelos entre si, tocando-se pelas faces ao longo de toda a sua extensão.
A amostra DZP-3686 corresponde a um agrupamento de tr ês conulários, todos com teca incompleta, novamente representadas apenas pela região mediana. Os três espécimes apresentam dimensão igual a 3,15 cm cada, os quais mantêm restos da teca preservados e são constituídos de quatro faces verdadeiras. Embora inflados na matriz, os conulários apresen tam-se achatados longitudinalmente, com dobras ao longo de toda teca. Os espécimes estão verticalmente orientados, em relação ao plano de acamamento e preservados paralelos entre si, cujas faces dos diferentes conulários tocam-se na porção superior da região mediana das tecas. Entretanto, ocorre um espaço entre os indivíduos desse agrupamento, preenchido pela rocha matriz, que aumenta adapicalmente, chegando a 3,5mm na porção inferior da região preservada da teca (figuras 3.C, E-G; 4).




Figura 3. C, Vista lateral da região mediana de dois espécimes (DZP-3686) / lateral view of the median region of two specimens (DZP-3686); E-G, DZP- 3686; E, Vista superior, em seção, de agrupamento formado por três espécimes (setas brancas) / cluster with three (white arrows), upper view (cross section)F, Vista superior, em seção, de agrupamento mostrando dois exemplares, o terceiro foi retirado (setas brancas), cujas faces se tocam na região superior da teca (vide também figura 4) / cluster (white arrows) showing two specimens (the third one is missing), upper view (cross section). Note that the faces are touching each other in the superior portion of the median region (see also figure 4); G, Vista inferior, em seção, de agrupamento mostrando dois exemplares, o terceiro foi retirado (setas brancas), cujas faces não se tocam na porção inferior da teca (vide também figura 4) / the same as F, lower view. Note the matrix between the specimens, indicating that the tecae are not in close contact (see also figure 4). Escala gráfica: 1 cm / graphic scale: 1 cm.
O agrupamento correspondente à amostra DZP- 3633 apresenta dois conulários, dentre os quais um apresenta teca com regiões basal, mediana e da abertura preservadas, ainda que a primeira e a última estejam incompletas. A teca do outro espécime corresponde às regiões basal e mediana. Os dois espécimes mostram dimensões de 5,47 cm e de 3,30 cm, respectivamente, sendo que o maior apresenta restos da teca preservados, constituído por quatro faces verdadeiras. Entretanto, o menor (DZP-3633b) representa o molde externo de uma face. Ocorrem inflados na matriz sedimentar, porém achatados longitudinalmente. Os dois espécimes desse agrupamento estão preservados inclinados, em relação ao plano de acamamento, com o mesmo ângulo de inclinação (40º) e tocamse pelas faces, ao longo de toda a teca, não havendo, portanto, espaço preenchido por rocha matriz entre as tecas dos indivíduos. Tal agrupamento é distinto de todos os demais aqui descritos, pois os conulários estão associados a restos de braquiópodes inarticulados (Orbiculoidea) e moldes de moluscos bivalves (Nuculacea), na mesma amostra de mão. Embora os conulários desse agrupamento ocorram associados a outros invertebrados, nenhuma evidência de fixação da teca foi identificada, uma vez que, conforme já mencionado, a base não está preservada (figura 3.D). Finalmente, a amostra DZP-3505 reúne dois espécimes, sendo um deles (DZP-3505a) mais completo, com teca representada pelas três regiões e o segundo espécime (DZP-3505b) apresenta as regiões basal e parte da mediana da teca preservadas. As dimensões das tecas dos conulários são de 10,96 cm e de 4,46 cm de comprimento, respectivamente, ambas apresentam restos da teca preservados na forma de uma película escura e são representadas por quatro faces verdadeiras, cada. Embora achatados lateralmente, os conulários ocorrem inflados na matriz sedimentar. Estão preservados inclinados, em relação ao plano de acamamento, porém com diferentes graus de inclinação, sendo o ângulo de inclinação do espécime maior (DZP-3505a) igual a 50º e, do menor (DZP- 3505b), igual a 20º.

Figura 3. D, Vista lateral de agrupamento com dois exemplares (DZP-3633), (setas brancas) / cluster (DZP-3633) with two specimens (white arrows), lateral view
Discussão
Modo de vida
Fundamentados em sua morfologia funcional e/ou inferências filogenéticas, conulários já foram interpretados como invertebrados planctônicos, pseudoplanctônicos ou bentônicos (Babcock, 1991; Van Iten, 1991a, 1991b). Estudos tafonômicos realizados para as ocorrências da Formação Ponta Grossa, Estado do Paraná (Simões et al., 2000; Rodrigues, 2002; Rodrigues et al., 2003), bem como dados de literatura (Harland e Pickerril, 1987), indicam que os conulários foram fixados ao substrato pela base (abertura voltada para cima). Tais observações tafonômicas estão de acordo com as interpretações de Sinclair (1948), Finks (1955), Petri e Fúlfaro (1983), Babcock et al. (1987a), Van Iten (1991a, 1991b) e Van Iten e Cox (1992). Vários autores (e.g., Ruedemann, 1898; Slater, 1907; Kozlowski, 1968; Bischoff, 1978; Hemish, 1986) sugeriram evidências de fixação por disco basal, projeções do tipo "raízes", além de outros sistemas de ancoragem (Harland e Pickerill, 1987).
Babcock (1991) e Babcock e Feldmann (1986a, 1986b), por sua vez, propuseram uma nova interpretação para a anatomia da região basal da teca dos conulários, ou seja, que a terminação basal dos conulários seja afilada, associada a um pedúnculo flexível, talvez quitinoso. Para esses autores, as evidências apresentadas por Kozlowski (1968) (e.g., projeções tipo "raízes") estão fundamentadas em exemplares que não pertenceriam aos conulários, embora outros autores apresentassem caracteres diagnósticos de Conulatae, para os mesmos exemplares (Van Iten e Cox, 1992). No entanto, os exemplares que suportam essa reconstrução anatômica (Babcock e Feldmann, 1986b, figs. 21.1, 24.2 e 32.5) apresentam sua porção basal pobremente preservada e a hipótese do pedúnculo ser artefato de preparação ou restos vegetais associados não é descartada (Van Iten e Cox, 1992). Embora o ponto de fixação no substrato não tenha sido observado no material examinado, uma vez que a região basal não está preservada, as orientações vertical e inclinada (abertura voltada para cima), em relação ao plano de acamamento, são dificilmente conciliadas com um modo de vida plactônico (veja também Harland e Pickerill, 1987). Visto que todos os espécimes dos agrupamentos examinados (verticais ou inclinados) estão orientados com a abertura para cima, torna-se improvável ser esse um fato acidental (Simões et al., 2000; Rodrigues et al., 2003).
Significado paleobiológico
Conulários são geralmente referidos como formas solitárias, embora agrupamentos de conulários tenham sido ilustrados na literatura por Hall (1876, 1879), Slater (1907), Ruedemann (1925), Sinclair (1940), Babcock e Feldmann (1986a, 1986b), Van Iten e Cox (1992) e Rodrigues et al. (2001). Os agrupamentos apresentados na literatura são monoespecíficos (tabela 1), com os espécimes preservados paralelos ou inclinados, em relação ao plano de acamamento, na maioria das vezes, ocorrem achatados lateralmente. Os indivíduos convergem para um centro comum, em direção à sua região basal, distribuindo-se radialmente (Van Iten e Cox, 1992). As únicas evidências de agrupamentos de espécimes inflados, com bases preservadas são descritas para Paraconularia crustula, Pensilvaniano da América do Norte (Van Iten e Cox, 1992). Esse agrupamento apresenta uma lâmina de material extremamente fino, bem próximo das bases dos exemplares, cuja constituição é apatítica, semelhante a do material da teca dos conulários, representando, possivelmente, o tecido de conexão entre os espécimes (Van Iten e Cox, 1992). Conforme advertem os autores, "o material da teca na extremidade basal dos dois conulários parece ser contínuo com a lâmina apatítica, entretanto, a pobre preservação dessas áreas torna difícil testar essa hipótese conclusivamente" (Van Iten e Cox, 1992, p. 423).
De modo similar às ocorrências da literatura, os agrupamentos de conulários da Formação Ponta Grossa são monoespecíficos. Conularia quichua é o elemento predominante da fauna de conulários da Formação Ponta Grossa (Leme, 2002; Leme et al., 2004), até então observado como formas solitárias. Outra feição importante desses agrupamentos é o fato de os espécimes estarem preservados inflados e verticalmente orientados, em relação ao plano de acamamento, ou inclinados. Formas isoladas de Conularia quichua de mesma condição tafonômica (classes tafonômicas 1 e 2) foram interpretadas como ocorrências in situ, preservadas em depósitos de sufocamento (Simões et al., 1999, 2000; Rodrigues, 2002; Rodrigues et al., 2003).
Os exemplos disponíveis na literatura (tabela 1), bem como a maioria dos espécimes aqui descritos mostra, tipicamente, que as tecas tocam-se lateralmente e convergem para um centro comum, com exceção do exemplar DZP-3686 (veja discussão a seguir). Nos exemplares da Formação Ponta Grossa, embora inflados e preservados in situ, com teca praticamente completa, a base está faltando. Conforme discussão em Simões et al. (2003), a perda dessa estrutura deve-se a diagênese e ao intemperismo, sugerindo que essa porção da teca era menos mineralizada. De fato, Van Iten e Cox (1992) descrevem material de constituição apatítica semelhante a teca dos conulários, próximo das bases dos espécimes agrupados. É importante notar que tal feição (perda da base) não é exclusiva dos espécimes preservados em agrupamentos, mas uma característica do registro tafonômico do grupo. O problema é que a ausência da base dificulta o entendimento da natureza desse agrupamento. Em outras palavras, não é possível saber, por exemplo, se os indivíduos foram conectados pela base, o que refletiria um comportamento colonial (Van Iten e Cox, 1992) ou se são representantes de assentamentos de duas ou mais larvas de conulários próximas entre si, refletindo um comportamento gregário (Babcock, 1991). Essa questão é muito importante porque colônias são formadas por clones provenientes de reprodução assexuada, interligados por tecidos, apresentando mesma fisiologia, morfologia ou anatomia. Já o comportamento gregário resulta do assentamento de larvas, por estímulo químico do substrato ou pela presença de outros indivíduos da mesma espécie ou de espécies diferentes e, portanto, envolve reprodução sexuada (Werner, 1979). Infelizmente, o material examinado não pode elucidar a questão da natureza dos agrupamentos de conulários, ou seja, se refletem um comportamento gregário ou colonial. No entanto, a mostra DZP-3686, mesmo com espécimes com bases ausentes, os conulários tocam-se apenas na porção superior da região mediana, havendo um espaço entre os espécimes na porção inferior da mesma (figura 4). Dessa forma, embora preservados lado a lado, provavelmente, em vida, esses espécimes não convergiram para um centro comum, sendo essa uma evidência mais favorável a um comportamento gregário, do que colonial (figura 4).
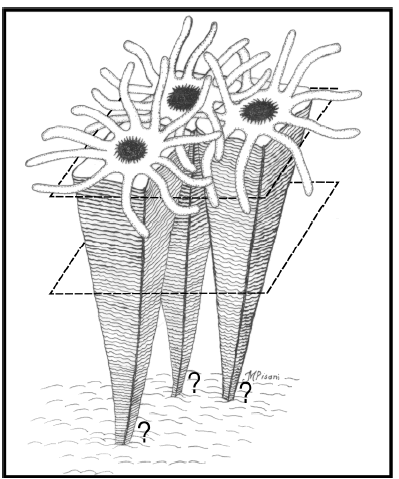
Figura 4. Possível comportamento gregário de conulários inferido a partir dos exemplares preservados no agrupamento DZP-3686 veja também figuras 3.E e 3.G. Note que há dúvida também quanto ao modo de fixação dos conulários junto ao fundo (explicação no texto) / possible conulariid gregarious behavior inferred from the specimens in the DZP-3686 cluster (see also figures 3.E e 3.G). The doubt regarding their attachment in the sea floor is also expressed (see the text for further explanation).
No contexto acima, é digno de nota também a ocorrência verificada no agrupamento (DZP-3081), onde um dos exemplares (DZP-3081a, figura 3.B) exibe cinco faces verdadeiras, ou seja, delimitadas por duas arestas adjacentes, uma diferença marcante entre C. quichua e as demais espécies de Conularia (Leme, 2002; Leme et al., 2004). A ocorrência de tecas não apresentando quatro faces tem sido observada em outros gêneros de conulários, incluindo Conularina triangulata (Raymond) (três faces), Paraconularia missouriensis (Swallow) e Vendoconularia triradiata Ivantsov e Fedonkin (ambas com seis faces) (Babcock et al., 1987b; Ivantsov e Fedonkin, 2002; Van Iten et al., 2005), mas não em Conularia. Portanto, o espécime de C. quichua (DZP-3081b) é o único na literatura com teca exibindo cinco faces (figura 3.B). Babcock et al. (1987b) interpretaram a ocorrência dos espécimes com três e seis faces como resultado de fenômenos teratológicos. No entanto, a presença de um espécime com cinco faces, em um mesmo agrupamento, permite especular sobre a possibilidade dessa condição ter origem relacionada à limitação de espaço no agrupamento e não apenas como resultado de fenômenos teratológicos. A existência de espécimes de conulários que apresentam três, seis ou, agora cinco faces, pode ser relevante nas discussões sobre as afinidades filogenéticas do grupo (Van Iten, 1992a). Em análises anteriores sobre o problema da afinidade dos conulários (e.g., Kiderlen, 1937; Werner, 1966, 1967; Babcock e Feldmann, 1986c; Van Iten, 1992a; Jerre, 1994; Bergström, 1995), a simetria da teca é tratada como um caráter morfológico diagnóstico. De fato, Kiderlen (1937), com base nas ilustrações de Wiman (1895), foi o primeiro a interpretar a simetria radial dos conulários, propondo que esses foram cnidários cifozoários, um grupo que geralmente é caracterizado pela presença de simetria tetrâmera bem desenvolvida (Brusca e Brusca, 1990; Gershwin, 1999). Desde Kiderlen (1937), muitos autores (Knight, 1937; Moore e Harrington, 1956a, 1956b; Werner, 1966, 1967, 1969, 1973; Bischoff, 1978; Van Iten, 1991a, 1991b, 1992a, 1992b; Van Iten e Cox, 1992; Van Iten et al., 1996, 2000; Jerre 1993, 1994; McKinney et al., 1995; Hughes et al., 2000) obtiveram novas evidências que reforçaram, conclusivamente, a idéia de que conulários são afins aos Cnidaria (Van Iten, 1991a, 1991b, 1992a, 1992b; Hughes et al., 2000), como, por exemplo, a comparação entre a morfologia e a microestrutura das feições internas dos conulários (septos e carenas) e estruturas internas dos cifozoários (septos gástricos), que apresentam similaridades em número, arranjo, tamanho e forma, sendo consideradas estruturas homólogas (Jerre, 1994; McKinney et al., 1995; Van Iten, 1991a, 1991b, 1992a, 1992b; Van Iten e Cox, 1992; Van Iten et al., 1996, 2000). No entanto, deve ser comentado que, recentes análises filogenéticas, apoiadas em dados paleontológicos, morfológicos e moleculares indicaram que Conulatae é grupo irmão de Stauromedusae e Scyphozoa é um grupo parafilético (Collins et al., 2000; Marques e Collins, 2004). No entanto, a plasticidade observada na simetria dos conulários não exclui uma afinidade com Scyphozoa, uma vez que, embora a maioria dos cifozoários apresente simetria tetrâmera por definição, algumas espécies (e.g., Aurelia aurita, A. labiata, Chrysaora fuscescens, Pelagia colorata e Phacellophora camtschatica), freqüentemente, não exibem simetria tetrarradial (Gershwin, 1999). Indivíduos aberrantes podem mostrar simetria variando de birradial a heptarradial (Gershwin, 1999). Adicionalmente, anormalidades similares também ocorrem em hidrozoários (Agassiz e Woodworth, 1896; Hargitt, 1901).
As causas da variação na simetria em Cnidaria são ainda pouco entendidas, podendo estar relacionadas a fatores genéticos, ambientais, ou mesmo por uma combinação entre eles (Gershwin, 1999). De qualquer forma, a plasticidade na simetria em Cnidaria representa uma estratégia favorável em condições adversas, pois apresentam habilidade de produzirem fenótipos alternativos, contribuindo para o aumento da diversificação entre as linhagens (West-Eberhard, 1986, 1989; Gershwin, 1999). Finalmente, conulários são similares aos cifozoários, por apresentarem mesmo padrão de simetria, normalmente tetraradial, mas podendo variar entre trirradial, pentarradial e hexaradial. No entanto, a ocorrência de conulários aberrantes não constitui uma evidência independente de uma relação de parentesco somente entre conulários e cifozoários, uma vez que esse padrão também ocorre em espécies de hidrozoários (Agassiz e Woodworth, 1896; Hargitt, 1901). De qualquer maneira, esse fenômeno, acrescenta novas evidências em relação à afinidade entre conulários e cnidários.
Comentários finais
Conularia quichua é o conulário mais comum e abundante na Formação Ponta Grossa, da Bacia do Paraná (Leme, 2002; Leme et al., 2004). Até o presente, eram conhecidas apenas ocorrências de espécimes isolados, preservados em posição de vida ou não. No presente estudo, são descritas ocorrências de agrupamentos in situ, com as tecas tocando-se pelas faces e, na maioria das vezes, convergindo para um centro comum, em direção à base. Os espécimes ocorrem todos inflados, verticalmente orientados ou inclinados e abertura voltada para cima, em relação ao plano de acamamento, os quais correspondem às classes tafonômicas 1 e 2 (Rodrigues et al., 2003).
Esses dados são importantes do ponto de vista paleobiológico, porque mostram que conulários, especialmente C. quichua, podem apresentar um modo de vida tanto solitário, como agrupado. Entretanto, a natureza desse agrupamento, isso é, se refletem um comportamento clonal ou gregário, não pode ser esclarecida para todos os agrupamentos aqui estudados, uma vez que, assim como ocorre nos exemplos da literatura internacional, a região basal desses não está preservada. Por outro lado, o fato de alguns conulários apresentarem um espaço maior na porção inferior da teca (em direção à base) (figura 4) parece ser uma evidência mais plausível com um comportamento gregário do que colonial. Adicionalmente, a ocorrência de um agrupamento de conulário com cinco faces verdadeiras (figura 3.B) acrescenta novas evidências em favor de afinidades desse grupo com os cnidários.
Agradecimentos
Os autores agradecem os pesquisadores M.J. Garcia (UNG), R. C.T. Cassab (CPRM), L.E. Anelli (IG/USP) e E. Bosetti (UEPG), pelo acesso às coleções científicas de conulário, utilizados para comparações, Heyo Van Iten e um revisor anônimo pelas suas construtivas sugestões à versão final do trabalho. Os autores agradecem também os alunos do GSA/IG-USP, R.P. Ghilardi, A.M.F. Sales, L.H.C. Mello, F.F. Torello e o H.A. Rodrigues (IBB/UNESP) pelo auxílio em campo e a L.B. Simões, pelo trabalho fotográfico. Suporte financeiro foi oferecido pela FAPESP e CNPq.
Referências
Agassiz, A. e Woodworth, W.M. 1896. Some variations in genus Eucope. Bulletin of Museum of Comparative Zoology at Harvard College 30: 121-150.
Babcock, L.E. 1991. The enigma of conulariid affinities. In: A.M. Simonetta e S.C. Morris (eds.), The early evolution of Metazoa and the significance of problematic fossil taxa, Cambridge University Press, pp. 113-143.
Babcock, L.E. e Feldmann, R.M. 1986a. Devonian and Mississipian conulariids of North America. Part A. General discription and Conularia. Annals of Carnegie Museum 55: 349-410.
Babcock, L.E. e Feldmann, R.M. 1986b. Devonian and Mississipian conulariids of North America. Part B. Paraconularia, Reticulaconularia, new genus and organisms rejected from Conulariida. Annals of Carnegie Museum 55: 411- 479.
Babcock, L.E. e Feldmann, R.M. 1986c. The phyllum Conulariida. In: A. Hoffmann e M.H. Nitecki (eds.), Problematic fossil taxa, Oxford University Press, Oxford, pp. 135-147.
Babcock, L.E., Feldmann, R.M., Wilson, M.T. e Suárez-Riglos, M. 1987a. Devonian conulariids of Bolivia. National Geographic Research 3: 210-231.
Babcock, L.E., Feldmann, R.M. e Wilson, M.T. 1987b. Teratology and pathology of some Paleozoic conulariids. Lethaia 20: 93- 105.
Bergamaschi, S. 1999. [Análise estratigráfica do Siluro-Devoniano (Formação Furnas e Ponta Grossa) da sub-bacia de Apucarana, Bacia do Paraná, Brasil. Programa de Pós-graduação em Geologia Sedimentar, Universidade de São Paulo, Tese de Doutoramento, 167 pp. Inédito].
Bergamaschi, S. e Pereira, E. 2001. Caracterização de Seqüências Deposicionais de 3a ordem para o Siluro-Devoniano na Sub- Bacia de Apucarana, Bacia do Paraná, Brasil. In: J.H.G. Melo e G.J.S. Terra (eds.), Correlações de Seqüências Paleozóicas Sul- Americanas, Ciência Técnica Petróleo, Seção Exploração de Petróleo 20: 63.
Bergström, J. 1995. Conulariid affinities: a discution. Geologiska Foreningens i Stockholm Forhandlingar 117: 245.
Bischoff, G.C.O. 1978. Internal structures of conulariid tests and Circonulariina n. suborder (Cnidaria, Scyphozoa). Senckenbergiana Lethaea 59: 275-327.
Brett, C.E. 1995. Sequence, stratigraphy, biostratigraphy, and taphonomy in shallow marine environments. Palaios 10: 597-616.
Brett, C.E. e Seilacher, A. 1991. Fossil Lagerstätten: a taphonomic consequence of event sedimentation. In: G. Einsele, W. Ricken e A. Seilacher (eds.), Cycles and events in stratigraphy. New York, Berlin, Heidelberg, Springer Verlag, pp. 283-297.
Brett, C.E., Baird, G.C. e Speyer, S.E. 1997. Fossil Lagerstätten: stratigraphic record of paleontological and taphonomic events. In: C.E. Brett and G.C. Baird (eds.), Paleontological Events: stratigraphic, ecological, and evolutionary implications. New York, Columbia University Press, pp. 3-40.
Brusca, R.C. e Brusca, G. 1990. Invertebrates. Sunderland, Mass, Sinauer Associates, 992 pp.
Collins, A.G., Marques A.C. e Simões, M.G. 2000. The phylogenetic placement of Conulatae within Cnidaria. Geological Society of America, Abstracts with Programs 32: A-443.
Feldmann, R.M., Chapman, R.E. e Hannibal, J.T. 1989. Paleotechniques. The Paleontological Society, 4, Special Publications. 358 pp.
Feldmann, R.M. e Babcock, L.E. 1986. Exceptionally preserved conulariids from Ohio- reinterpretation of their anatomy. National Geographic Research 2: 464-472.
Finks, R.M. 1955. Conularia in a sponge from the west Texas Permian. Journal of Paleontology 29: 831-836.
Gershwin, L. 1999. Clonal and population variation in jellyfish symmetry. Journal of the Marine Biological Association of United Kingdom 79: 993-1000.
Grahn, Y., Pereira, E. e Bergamaschi, S. 2000. Silurian and Lower Devonian chitinozoan biostratigraphy of the Paraná Basin in Brazil and Paraguay. Palynology 24: 147-176.
Hall, J. 1876. Illustrations of Devonian Fossils: Gasteropoda, Pteropoda, Cephalopoda, Crustacea and Corals of the Upper Helderberg, Hamilton and Chemung Groups. Albany, 7 pp.
Hall, J. 1879. Geological survey of New York. Paleontology. Containing descriptions of the gasteropoda, pteropoda and cephalopoda of the Upper Helderberg, Hamilton, Portage and Chemung Groups. C.V. Benthuysen and Sons. Albany, 492 pp.
Hargitt, C.W. 1901. Variation among Hydromedusae. Biological Bulletin, Marine Biological Laboratory, Woods Hole 2: 221-251.
Harland, T.L. e Pickerill, R.K. 1987. Epizoic Schizocrania sp. from the Ordovician Trenton Group of Quebec, with comments on mode of life of conulariids. Journal of Paleontology 61: 844-849.
Hemish, L.A. 1986. Pennsylvanian conulariids from Okfuskee County, Oklahoma. Oklahoma Geology Notes 46: 202.
Hughes, N.C., Gunderson, G.O. e Weedon, M.J. 2000. Late Cambrian conulariids from Wisconsin and Minnesota. Journal of Paleontology 74: 828-838.
Ivantsov, A.Y. e Fedonkin, M.A. 2002. Conulariid-like fossil from the Vendian of Russia: a metazoan clade across the Proterozoic/Palaeozoic boundary. Palaeontology 45: 1219-1229.
Jerre, F. 1993. Conulariid microfossils from the Silurian Lower Visby Beds of Gotland, Sweden. Palaeontology 36: 403-424.
Jerre, F. 1994. Anatomy and phylogenetic significance of Eoconularia loculata, a conulariid from the Silurian of Gotland. Lethaia 27: 97-109.
Kiderlen, H. 1937. Die Conularien. Über Bau and Leben der ersten Scyphozoa. Neues Jahrbuch für Mineralogie, Beilage-Band 77: 113-169.
Knight, J.B. 1937. Conchopeltis Walcott, an Ordovician genus of the Conulariida. Journal of Paleontology 11: 186-188.
Kozlowski, R. 1968. Nouvelles observations sur les conulaires. Acta Palaeontologica Polonica 13: 497-535.
Lange, F.W. e Petri, S. 1967. The Devonian of the Paraná Basin. In: J.J. Bigarella (ed.), Problems in Brazilian Devonian Geology, Boletim Paranaense de Geociências 21-22: 5-55.
Leme, J.M. 2002. [Revisão sistemática dos Conulatae Collins et al. 2000, Formação Ponta Grossa, Devoniano (?Lochkoviano- Frasniano), Bacia do Paraná, Brasil: implicações paleobiogeográficas e comentários sobre as relações filogenéticas entre os Conulatae. Programa de Pós-graduação em Geologia Sedimentar, Universidade de São Paulo, Dissertação de Mestrado, 100 pp. Inédito].
Leme, J.M., Heredia, S., Rodrigues, S.C., Simões, M. G., Aceñolaza, G.F. e Milana, J.P. 2003. Teresconularia gen. nov. from the Lower Ordovician of the Cordillera Oriental of Salta, (NW Argentina): the Oldest Conulariid (Cnidaria) from South America. Revista Española de Micropaleontologia 35: 265-273.
Leme, J.M., Rodrigues, S.C., Simões, M.G. e Van Iten, H. 2004. Sistemática dos conulários (Cnidaria) da Formação Ponta Grossa (Devoniano), do Estado do Paraná, Bacia do Paraná, Brasil. Revista Brasileira de Paleontologia 7: 213-222.
Marques, A.C. e Collins, A.G. 2004. Cladistic analysis of Medusozoa and cnidarian evolution. Invertebrate Biology 123: 23-42.
McKinney, F.K., Devolvé, J.J. e Sobieraj, J. 1995. Conularia sp. from the Pyrénées: further support for scyphozoan affinities of the Conularida. Lethaia 28: 229-236.
McMenamin, M.A.S. 1987. The fate of Ediacaran Fauna, the nature of conulariids, and the basal paleozoic predator revolution. Geological Society of America, Abstracts with Programs 19: 29.
Miller, K.B., Brett, C.E. e Parsons, K.M. 1988. The paleocologic significance of storm generated disturbance within a Middle Devonian muddy empiric sea. Palaios 3: 35-52.
Miller, M.F. e Smail, S.E. 1997. A semiquantitative field method for evaluating bioturbation on bedding planes. Palaios 12: 391- 396.
Moore, R.C. e Harrington, H.J. 1956a. Scyphozoa. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part F, Coelenterata. Geological Society of America and University of Kansas Press, F27-F38.
Moore, R.C. e Harrington, H.J. 1956b. Conulata. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part F, Coelenterata. Geological Society of America and University of Kansas Press, F54-F66.
Mortin, J. 1985. The shell structure and zoological affinities of conulariids. Palaeontological Association Annual Conference Abstracts, (Wales) 12-13.
Oliver, W.A.Jr 1984. Conchopeltis: its affinities and significance. Paleontographica Americana 54: 141-147.
Petri, S. e Fúlfaro, V.J. 1983. Geologia do Brasil: Fanerozóico. EDUSP, São Paulo, SP, 631 pp.
Rodrigues, S.C. 2002. [Tafonomia comparada dos Conulatae Collins et al. 2000, Formação Ponta Grossa, Devoniano (?Lochkoviano- Frasniano), Bacia do Paraná: implicações paleoeautocológicas e paleoambientais. Programa de Pós graduação em Geologia Sedimentar, Universidade de São Paulo, Dissertação de Mestrado, 100 pp. Inédito].
Rodrigues, S.C., Leme, J.M. e Simões, M.G. 2001. Conularia cf. quichua Ulrich: a solitary or clustered marine invertebrate? In: Reunião Anual da Sociedade Brasileira de Paleontologia, Paleo- 2001/SP (Rio Claro, 2001), Boletim de Resumos p. 12.
Rodrigues, S.C., Leme, J.M. e Simões, M.G. 2003. Tafonomia comparada dos Conulatae (Cnidaria), Formação Ponta Grossa (Devoniano), Bacia do Paraná, Estado do Paraná, Brasil. Revista Brasileira de Geociências 33: 381-390.
Ruedemann, R. 1898. The discovery of sessile Conularia. New York State Geology 15: 701-720.
Ruedemann, R. 1925. Some Silurian (Ontarian) Faunas of New York. New York State Museum Bulletin 265: 5-84.
Simões, M.G. e Ghilardi, R.P. 2000. Protocolo tafonômico/paleautoecológico como ferramenta nas análises paleossinecológicas: exemplos de aplicação em concentrações fossilíferas do Paleozóico da Bacia do Paraná, Brasil. Pesquisas em Geociências 27: 3-13.
Simões, M.G. e Kowalewski, M. 1998a. Shell beds as paleoecological puzzles: a case study from the Upper Permian on the Paraná Basin, Brazil. Facies 38: 175-196.
Simões, M.G. e Kowalewski, M. 1998b. Genetic complexity and geobiological implications of "simple" shell beds: an example from the Upper Permian of the Paraná Basin, Brazil. Journal of African Earth Sciences 27: 179-180.
Simões, M.G., Mello, L.H.C., Leme, J.M., Rodrigues, S.C. e Marques, A.C. 1999. Devonian conulariid taphonomy and their paleoecological implications. Geological Society of America, Abstracts with Programs: A-468.
Simões, M.G., Mello, L.H.C., Rodrigues, S.C, Leme, J.M. e Marques, A.C. 2000. Conulariid taphonomy as a tool in paleoenvironmental analysis. Revista Brasileira de Geociências 30: 757-762.
Simões, M.G., Rodrigues, S.C., Leme, J.M. e Van Iten, H. 2003. Some Middle Paleozoic conulariids (Cnidaria) as possible examples of taphonomic artifacts. Journal of Taphonomy 1: 165-186.
Sinclair, G.W. 1940. A discussion of the genus Metaconularia with descriptions of new species. Transactions of the Royal Society of Canada 34: 101-121.
Sinclair, G.W. 1948. [The biology of the Conularia. Ph.D. thesis, McGill University, Montreal, Quebec, 328 pp. Inédito].
Siviero, F.N. 2002. [Revisão sistemática das conulárias brasileiras. Programa de Pós-graduação em Geologia, Universidade Federal do Rio de Janeiro, Dissertação de Mestrado, 80 pp. Inédito].
Slater, I.L. 1907. A monograph of British Conulariae. Palaeontographical Society of London, 41 pp.
Steinmann, G. e Döderlein L. 1890. Elemente der Paläontologie. Verlag von Wilhelm Engelmann, Leipzig, 848 pp.
Steul, H. 1984. Die systematische Stellung der Conularien. Giessener Geologische Schriften 37: 117 pp.
Termier, H. e Termier, G. 1949. Position systématique et biologie des conulaires. Revue Scientifique 86: 711-722.
Termier, H. e Termier, G. 1953. Les Conularides. In: J. Piveteau (ed.), Traité de Paléontologie, Onychophores, Arthropodes, Echinodermes, Stomocordes. Masson and Cie., Paris, 3, pp. 1006- 1013.
Van Iten, H. 1987. The mode of life of the Conulariida and its implications for conulariid affinities. Geological Society of America, Abstracts with Programs 19: 876.
Van Iten, H. 1991a. Evolutionary affinities of conulariids. In: A.M. Simonetta e S.C. Morris (eds.), The early evolution of Metazoa and the significance of problematic fossil taxa, Cambridge University Press, pp. 145-155.
Van Iten, H. 1991b. Anatomy, patterns of occurrence, and nature of the conulariid schott. Palaeontology 34: 939-954.
Van Iten, H. 1992a. Morphology and phylogenetic significance of the corners and midlines of the conulariid test. Palaeontology 35: 335-358.
Van Iten, H. 1992b. Microstructure and growth of the conulariid test: implications for conulariid affinities. Palaeontology 35: 359-372.
Van Iten, H. e Cox, R.S. 1992. Evidence of clonal budding in a radial cluster of Paraconularia crustula (White) (Pennsylvanian: ?Cnidaria). Lethaia 25: 421-426.
Van Iten, H., Fitzke, J.A. e Cox, R.S. 1996. Problematical fossil cnidarians from the Upper Ordovician of the North-Central USA. Palaeontology 39: 1037-1064.
Van Iten, H., Zhu, Z.K e Zhu, M.Y. 2000. Anatomy and systematic of the Devonian conulariids Changshaconus Zhu, 1985 and Reticulaconularia Babcock and Feldmann, 1986. Acta Palaeontologica Sinica 39: 466-475.
Van Iten, H., Leme, J.M., Rodrigues, S.C. e Simões, M.G. 2005. Reinterpretation of a conulariid-like fossil from the Vendian of Russia. Palaeontology.
Werner, B. 1966. Stephanoscyphus (Scyphozoa Coronatae) und seine direkte Abstammung von den fossilen Conulata. Helgoländer Wissenschaftliche Meeresuntersuchungen 13: 17-21.
Werner, B. 1967. Stephanoscyphus Allman (Scyphozoa Coronatae), ein rezenter Vertreter der Conulata? Paläontologische Zeitschrift 41: 137-153.
Werner, B. 1969. Neue Beitraege zur Evolution der Scyphozoa und Cnidaria. 1º Simpósio Internacional de Zoofilogenia. University of Salamanca, Salamanca, pp. 223-244.
Werner, B. 1973. New investigations on systematics and evolution of the class Scyphozoa and the phylum Cnidaria. Seto Marine Biology Labaratory Publicattions 20: 35-61.
Werner, B. 1979. Coloniality in the Scyphozoa: Cnidaria. In: G. Larwood e B.R. Rosen (eds.), Biology and Systematics of colonial organisms, Academic Press, London and New York, pp. 81- 103.
West-Eberhard, M.J. 1986. Alternative adaptions, speciation, and phylogeny (a review). Proceedings of the National Academy of Sciences of the United States of America 83: 1388-1392.
West-Eberhard, M.J. 1989. Phenotypic plasticity and the origins of diversity. Annual Review of Ecology and Systematics 20: 249-278.
Wiman, C. 1895. Palaeontologische Notizen 1-2. Bulletin of the Geological Institution of Upsala 3: 1-9. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 6 de octubre de 2004.
Aceptado: 25 de enero de 2005.

