










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.2 Buenos Aires abr./jun. 2006
Análisis tafonómico e inferencia del grado de mezcla temporal y espacial de la macrofauna del Miembro Pilmatué de la Formacion Agrio, Cretácico Inferior de cuenca Neuquina, Argentina
Darío G. Lazo1
1 Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Ciudad Universitaria, 1428 Buenos Aires, Argentina, CONICET, <dlazo@gl.fcen.uba.ar>
Abstract. Taphonomy, time-averaging and lateral transport of the macrofossils of the Pilmatué Member of the Agrio formation, lower cretaceous, neuquén basin, argentina. A taphonomic and sedimentary facies analysis of the Pilmatué Member of the Agrio Formation in central Neuquén basin is presented to determine the time-averaging and degree of lateral transport of its macrofauna. The Pilmatué Member is characterized by abundant fossil concentrations dominated by benthic organisms such as corals, serpulids, bivalves and gastropods. Stratigraphic occurrence of benthic elements was accurately dated by a refined ammonite zonation of the Agrio Formation. Three sections were studied between Chos Malal and Zapala (Neuquén) situated on a north-south line of 44.2 km. Five litofacies and nine taphofacies were defined. The depositonal system is interpreted as a storm influenced homoclinal shoreface to shelf environment. Taphonomic analysis showed that most macrofaunistic associations are autochthonous or slightly parautochthonous with no significant lateral transport. Census, within-habitat time-averaged and environmentally condensed assemblages were identified. Time scales of fossil accumulation span from days-years up to 0.5 My. Nearshore assemblages are less time-averaged than shelf assemblages as in modern settings.
Resumen. Se describen e interpretan las tafofacies del Miembro Inferior o Pilmatué de la Formación Agrio en el sector central de la cuenca Neuquina para determinar el grado de mezcla temporal y lateral que presentan sus macrofósiles. A su vez se presenta una breve descripción e interpretación de las litofacies presentes en la misma unidad. El Miembro Pilmatué se caracteriza por abundantes concentraciones fósiles dominadas por organismos bentónicos dentro de los que se destacan corales, serpúlidos, bivalvos y gastrópodos. Los mismos se registran junto a amonites, lo que permite ubicarlos estratigráficamente en forma muy precisa. Tres perfiles ubicados entre Chos Malal y Zapala (Neuquén) fueron analizados con sumo detalle. Los mismos abarcan una distancia de 44,2 km. Se definieron cinco litofacies que permitieron interpretar al Miembro Pilmatué como depositado en un ambiente de shoreface-plataforma influido por tormentas sin quiebres de pendiente hacia el offshore. Las tafofacies analizadas son autóctonas o ligeramente parautóctonas y su grado de transporte lateral fue nulo o bajo en dirección al offshore. La mezcla temporal es variable ya que se han registrado tafofacies de censo con mínima mezcla temporal (días-años) y tafofacies promediadas dentro del hábitat y condensadas ambientalmente con un promedio temporal mayor, desde años hasta un máximo de 0,5 Ma. Al igual que ocurre en ambientes actuales el grado de mezcla temporal aumenta en dirección al offshore.
Key words. Valanginian-Hauterivian; Storm influenced shoreface-shelf. Macrofauna; Taphonomy.
Palabras clave. Valanginiano-Hauteriviano; Shoreface-plataforma influenciada por tormentas; Macrofauna. Tafonomía.
Introducción
En las últimas dos décadas la tafonomía ha experimentado un crecimiento significativo no sólo en el número de contribuciones sino en sus alcances y objetivos (Behrensmeyer et al., 2000). Dentro de ellos se destaca la interpretación paleoambiental de secuencias con abundante contenido fosilífero. Combinando datos tafonómicos, sedimentológicos y estratigráficos es posible subdividir a los ambientes actuales y antiguos en tafofacies o facies tafonómicas (Speyer y Brett, 1986). Las tafofacies son tanto indicadoras paleoecológicas (e.g., fidelidad espacial) como paleoambientales (e.g., tasa de sedimentación) y los análisis de tafofacies han sido aplicados exitosamente tanto en ambientes actuales como en el registro geológico (e.g., Fürsich y Flessa, 1991). En nuestro país, los estudios combinados de litofacies y tafofacies en ambientes marinos son relativamente pocos, pero están demostrando ser muy útiles en el refinamiento de interpretaciones paleoambientales (Del Río et al., 2001; Palma y Lanés, 2001). En particular, las rocas del Cretácico Inferior de la cuenca Neuquina son una oportunidad ideal para realizar estudios tafonómicos y paleoecológicos. Sus afloramientos, potentes y extensos lateralmente, son portadores de una gran cantidad de fósiles comparable a las más importantes secuencias cretácicas del mundo.
El presente trabajo tiene como objetivo describir e interpretar las litofacies y tafofacies del Miembro Inferior o Pilmatué de la Formación Agrio en el sector central de la cuenca Neuquina para inferir el grado de mezcla temporal y lateral que presenta su macrofauna en los distintos paleoambientes de dicha unidad. Esta información será luego utilizada en otro trabajo para la definición de biofacies o paleocomunidades con significado paleoambiental.
Zona de estudio
La cuenca Neuquina es una cuenca de retroarco ubicada principalmente en las provincias de Mendoza y Neuquén. Su desarrollo abarcó el lapso Triásico Superior-Oligoceno (figura 1.A). Desde el Triásico tardío en algunos sectores y desde el Pliensbaquiano en la mayor parte de la cuenca ocurrió una conexión con el océano Pacífico que duró hasta el Barremiano (Legarreta y Gulisano, 1989). Sin embargo, la forma de golfo, la presencia de un arco de islas en el margen oeste y cambios relativos del nivel del mar habrían generado aislamientos de la cuenca al menos en forma temporaria. Durante el Valanginiano-Barremiano ocurrió la depositación de la Formación Agrio. Sus afloramientos aparecen a lo largo de casi toda la cuenca aunque con importantes variaciones en espesor, litología y contenido fosilífero (Marchese, 1971; Leanza et al., 1977). En su localidad tipo se la puede dividir en tres miembros: Inferior o Pilmatué, Medio o Avilé y Superior o Agua de la Mula. Tanto el Miembro Inferior como el Superior están compuestos por pelitas, areniscas y coquinas de origen marino, mientras que el Miembro Medio es un cinturón delgado de areniscas de origen continental. Se apoya en concordancia sobre la Formación Chachao en Mendoza y sobre la Formación Mulichinco en Neuquén. Su techo es cubierto por el Miembro Chorreado o por el Miembro Troncoso Inferior de la Formación Huitrín (Leanza, 2003).

Figura 1. Ubicación del área de estudio. A, ubicación de la cuenca Neuquina en el margen centro-oeste de Argentina. B, mapa de Neuquén mostrando la ubicación de los tres perfiles analizados: 1, Agua de la Mula; 2, Salado Sur; 3, Bajada del Agrio. Nótese que los perfiles se encuentran ubicados sobre una transecta N-S de 44,2 km de largo / location of the study area. A, location map of the Neuquén Basin in west-central Argentina. B, fossil localities: 1, Agua de la Mula; 2, Salado Sur; 3, Bajada del Agrio. Note that the three localities are on a 44.2 km north-south transect.
Los perfiles seleccionados para este estudio se encuentran dentro de la faja plegada y corrida del Agrio en el centro-norte de Neuquén (figura 1.B). El perfil Agua de la Mula está ubicado a 90 km al sur de Chos Malal en el sector noroeste de la cordillera del Salado (cerro Mula). El perfil Salado Sur está ubicado a 16,3 km al sur de Agua de la Mula en el sector sudoeste de la misma cordillera (cerro Salado). Las capas son continuación lateral de las aflorantes en Agua de la Mula y presentan un rumbo e inclinación semejantes. El perfil Bajada del Agrio está ubicado a 27,9 km al sur de Salado Sur en un paraje denominado Bajada Vieja sobre la margen derecha del río Agrio. En los tres perfiles la base de la Formación Agrio está dada por la aparición de la primeras pelitas negras sobre calizas grisáceas con Aetostreon (= "Exogyra") tomadas aquí como techo de la Formación Mulichinco. En los tres perfiles se registraron las mismas biozonas de amonites aunque hacia Bajada del Agrio se registra una disminución del espesor (figura 2).

Figura 2. Perfiles litológicos del Miembro Pilmatué de la Formación Agrio mostrando la biozonación de amonites y la distribución de tafofacies / lithological columns of the Pilmatué Member of the Agrio Formation showing the ammonite zonation and the distribution of taphofacies.
Criterios y metodología
La clasificación de litofacies y tafofacies se elaboró a partir de observaciones de campo. El control estratigráfico se basó en la biozonación de amonites propuesta por Aguirre-Urreta y Rawson (1997, 2003). A partir de la misma se pudo establecer que cada biozona abarcaría unos 0,5 Ma en promedio. Esto marca un límite superior para la mezcla temporal de fósiles ya que en la zona de estudio no hay condensación bioestratigráfica.
Las litofacies se definieron en función de litología, textura, estructuras sedimentarias, geometría, contactos, empaquetamiento, color y espesor. En primera instancia se subdivieron en tres grandes grupos: pelitas, areniscas y concentraciones fósiles. Luego se subdividieron sobre la base de las estructuras sedimentarias y fueron interpretadas en términos de los procesos deposicionales. Finalmente se agruparon en asociaciones para interpretarlas en función de distintos subambientes deposicionales. El modelo paleoambiental utilizado corresponde a un ambiente marino dominado por oleaje caracterizado por una zonación de subambientes pendiente abajo y en dirección al offshore (figura 3). Las tempestitas se clasificaron en proximales, proximales amalgamadas y distales según los esquemas planteados por Aigner y Reineck (1982), Dott y Burgeois (1982) y Cheel y Leckie (1993). Dentro de las tempestitas distales se distinguió un tipo particular denominado tempestita fangosa (mud layer) definida por Brett (1983).

Figura 3. Perfil esquemático de un ambiente de shoreface-plataforma mostrando la ubicación de elementos morfológicos, icnofacies y tempestitas. Modelo basado en Hampson y Storms (2003). Datos de bases de ola de buen tiempo y de tormenta tomados de Howard y Reineck (1981) / schematic shoreface-shelf profile showing main morphological elements, ichnofacies, and storm beds. Model from Hampson and Storms (2003). Fairweather wave base and storm wave base from Howard and Reineck (1981).
Las tafofacies fueron definidas sobre la macrofauna del Miembro Pilmatué para evaluar su grado de mezcla temporal y espacial. En Agua de la Mula y Bajada del Agrio se realizaron 48 muestreos aleatorios de 45 individuos cada uno en distintos niveles del perfil. En cada individuo seleccionado se relevaron, cuando fue posible, los siguientes rasgos: 1, orientación (convexo hacia arriba/convexo hacia abajo/concordante/vertical/posición de vida/indeterminada); 2, articulación (articulado/desarticulado/ bivalvo en mariposa/indeterminada); 3, fragmentación (entero/fragmento); 4, modificación de bordes (sin alteración/roto/redondeado); 5, corrasión (externa/interna, sin alteración/alteración menor/ alteración mayor); 6, bioerosión (externa/interna, presente/ausente); 7, tipo de bioerosión (tipo cliónido/Gastrochaenolites isp./indeterminado); 8, incrustamiento (externo/ interno, presente/ ausente) y 9, organismo incrustante (Amphidonte (Ceratostreon) sp./Parsimonia antiquata (Sowerby)/ Plicatula sp. indet./ briozoario indeterminado/Demospongea indeterminada/ indeterminado). Con los datos obtenidos se calcularon porcentajes de cada rasgo y el grado de modificación tafonómica se determinó de la siguiente manera: 0-25%, leve o bajo; 25-50%, intermedio; 50- 75%, alto y 75-100%, muy alto.
Las identificaciones taxonómicas y los hábitos de vida se tomaron de Lazo (2004). Los rasgos tafonómicos utilizados están basados en Brett y Baird (1986) y Kidwell (1991) y la clasificación según el grado de time-averaging en Kidwell y Bosence (1991). Un espécimen se consideró bioerosionado y/o incrustado si la bioerosión y/o el incrustamiento cubrían más del 2% de la superficie de la conchilla. Los bivalvos fueron los fósiles mayormente relevados debido a su mayor abundancia relativa, aunque algunos niveles estaban dominados por serpúlidos o corales.
Análisis paleoambiental
En el cuadro 1 se presenta una breve descripción e interpretación de las litofacies (figura 4.A, B,C,D). Las mismas fueron agrupadas en tres asociaciones de facies. La asociación A está compuesta por las litofacies Fl y Fm acompañadas de concreciones carbonáticas, Thalassinoides isp. concrecionados, niveles con trazas de la icnofacies de Glossifungites e intercalaciones de concentraciones delgadas de bioclastos (figura 4.E). Comprende intervalos de hasta 25 m de espesor. Es la más común de las asociaciones descriptas en la zona de estudio. Estos depósitos resultan principalmente de la decantación de materiales finos por debajo del nivel de olas de buen tiempo en la zona de transición y en la plataforma media-externa. Las intercalaciones delgadas de conchillas pueden interpretarse como tempestitas distales y fangosas.
Cuadro 1. Descripción e interpretación de las litofacies identificadas en el Miembro Pilmatué de la Formación Agrio en la zona de estudio / lithofacies of the Pilmatué Member of the Agrio Formation in the study area: description and interpretation.
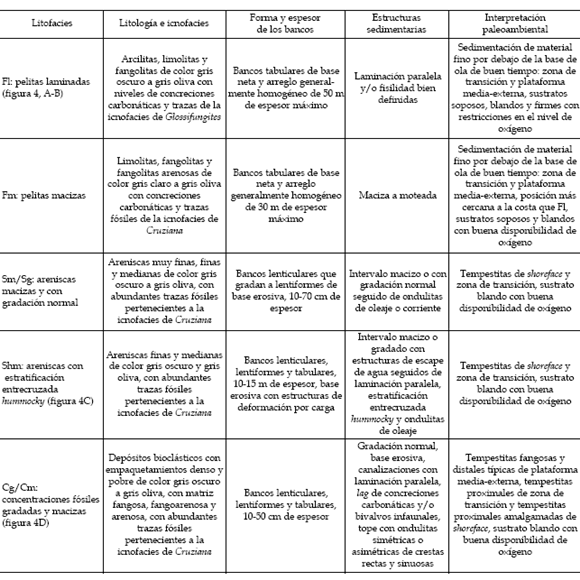

Figura 4. Fotos de campo de las litofacies del Miembro Pilmatué de la Formación Agrio. A, litofacies Fl, corte tranversal. B, litofacies Fl/Fm, corte transversal mostrando trazas fósiles (cf. Teichichnus isp.) de la icnofacies de Glossifungites bajo un contacto erosivo. C, litofacies Shm, corte transversal de un banco lenticular. D, litofacies Cg/Cm, corte transversal. E, asociación de litofacies A. F, asociaciones de litofacies B y C. Fotos A, C-E de Agua de la Mula; B y F de Bajada del Agrio. Escalas gráficas en centímetros; tapa de lente de 5 cm de diámetro; martillo de aprox. 30 cm de largo / Field pictures of lithofacies of the Pilmatué Member of the Agrio Formation. A, lithofacies Fl, transversal section. B, lithofacies Fl/Fm, transversal section showing trace fossils (cf. Teichichnus isp.) belonging to Glossifungites ichnofacies below an erosive contact. C, lithofacies Shm, lenticular bed in transversal section. D, lithofacies Cg/Cm, transversal section. E, association of lithofacies A. F, association of lithofacies B and C. A, C-E from Agua de la Mula; B and F from Bajada del Agrio. Graphic scales in centimeters; lens cap has 5 cm in diameter; hammer is approx. 30 cm-long.
La asociación B está compuesta de una alternancia de pelitas (Fl/Fm), areniscas medianas a finas o muy finas (litofacies Sg/Sm y Shm) y concentraciones fósiles formando una estratificación heterolítica de tipo lenticular que grada a ondulosa (figura 4.F). Comprende intervalos de hasta 30 m de potencia. Las areniscas y las concentraciones pasan de una geometría lenticular de menos de 15 cm de espesor a una geometría lentiforme de hasta 40 cm de espesor. Esta asociación se ubica sobre la asociación A. Estos depósitos reflejan la alternancia de depositación mecánica y decantación por lo cual pueden ubicarse dentro de la zona de transición. En los períodos de buen tiempo se producía la decantación de fango mientras que en períodos de tormenta se generaban las capas de areniscas y concentraciones intercaladas interpretadas como tempestitas proximales. La tendencia estratocreciente de los intervalos heterolíticos indica progradación de la zona de transición.
La asociación C está compuesta de cuerpos arenosos de geometría tabular de hasta 30 m de espesor (figura 4.F) formados por la amalgamación de bancos de areniscas (Sg/Sm y Shm) y concentraciones fósiles (Cg/Cm). Las capas individuales poseen una geometría lentiforme y hasta 80 cm de espesor. Los bancos formados por capas amalgamadas son fácilmente correlacionables entre las tres secciones analizadas. Estos depósitos se ubican sobre las asociaciones A o B. La presencia de bancos amalgamados indica la acción de procesos de tormenta en un espacio de acomodación reducido y aquí son interpretados como depósitos de shoreface, justo por encima de la base de ola de buen tiempo.
Tafofacies
P1: pavimentos de bivalvos y amonites
Descripción. Corresponde a pavimentos de bivalvos y amonites inmersos en las litofacies Fl y Fm en zona de transición y plataforma media-externa. No hay estructuras sedimentarias asociadas. Además pueden encontrarse restos post-craneales desarticulados de reptiles marinos (Lazo y Cichowolski, 2003). Las especies más abundantes son: Cucullaea (N.) gabrielis Leymerie, Steinmanella pehuenmapuensis Leanza y Steinmanella transitoria (Steinmann) (figura 5.A). Además pueden aparecer en forma esporádica Pinna (P.) robinaldina d'Orbigny, Sphaera koeneni (Behrendsen), Disparilia sp. indet. y Ptychomya koeneni Behrendsen. Es común encontrar pavimentos monoespecíficos de Modiolus cf. M. subsimplex d'Orbginy de conchilla muy delgada y tamaño pequeño (largo máximo = 63 mm).

Figura 5. Fotos de campo de las tafofacies del Miembro Pilmatué de la Formación Agrio. A, tafofacies P1, corte transversal mostrando dos ejemplares desarticulados de Cucullaea (Noramya) gabrielis en posición vertical.A, taphofacies P1, transversal section showing two disarticulated specimens of C. (N.) gabrielis in vertical orientation.
Los rasgos tafonómicos principales son: muy alta articulación y niveles muy bajos de fragmentación, modificación de bordes, corrasión y bioerosión. Las conchillas poseen un nivel intermedio de incrustación externa y un nivel bajo de incrustación interna. Las orientaciones predominantes fueron valvas convexas hacia arriba (36%) y verticales (48%); se registraron además algunas conchillas en posición de vida (4%). Los incrustantes observados fueron Amphidonte (C.) sp. (77,8%) y Parsimonia antiquata (22,2%). Algunos ejemplares de ostras incrustantes se encontraban articulados. Los moldes internos de Modiolus cf. M. subsimplex pueden tener trazas de tipo criptococlear. El relleno de las conchillas es pelítico y concordante con la roca hospedante.
Los amonites se registran con el plano de enroscamiento paralelo a la estratificaci ón. Tienen baja corrasión externa, baja fragmentación, baja bioerosión externa y no presentan modificación de bordes. Presentan incrustaciones externas de Amphidonte (C.) sp., Plicatula sp. indet. o Parsimonia antiquata. La cámara habitación siempre se rellena. El fragmócono eventualmente aparece relleno. Es interesante destacar que no se han encontrado nautílidos en esta tafofacies.
Interpretación. Dada la excelente preservación de los bivalvos, a veces en posición de vida, se supone que las conchillas no han tenido un transporte significativo y por lo tanto se habrían preservado en su hábitat original. Los pavimentos de amonites son el resultado de la caída o lluvia de estos organismos desde la columna de agua hacia un fondo fangoso de plataforma. Luego de la muerte caían al fondo y se acomodaban en forma paralela a la estratificación.
La preservación buena de la mayoría de los fósiles indica también un enterramiento relativamente rápido, sin embargo la presencia de incrustantes en la cara interna de algunas valvas indica que antes del enterramiento final ocurrió desarticulación y exposición de conchillas sobre el sustrato. La orientación en posiciones inestables se puede explicar también a través de un enterramiento rápido. Este tipo de concentraciones ha sido interpretado en la literatura como depósitos de flujos de baja energía que no alcanzan a desplazar significativamente a los organismos, más bien tienden a lavar la matriz que los rodea (winnowed concentrations), concentrándolos o simplemente cubriéndolos con una capa de fango. Este tipo de procesos pueden darse en la última fase de un flujo de relajación de tormenta hacia la plataforma mediaexterna (tempestita fangosa) o como producto de una tormenta de baja intensidad en la zona de transición.
En el caso de los pavimentos monoespecíficos de bivalvos pequeños podría pensarse que algún factor ambiental estaba controlando su desarrollo (e.g., nivel de oxígeno o salinidad). Esta tafofacies puede ser interpretada como una asociación promediada dentro del hábitat o condensada ambientalmente en el caso de los pavimentos monoespecíficos. Ambos tipos reflejan la superposición de distintas generaciones de organismos sin o con un cambio ambiental respectivamente y su mezcla temporal es mayor a la de una asociación de censo. La ausencia de nautílidos sugeriría que sus conchillas tendrían un comportamiento post-mortem diferente o un hábito de vida distinto al de los amonites (véase Lazo et al., 2005).
P2: niveles de concreciones
Descripción. Se compone de niveles de concreciones carbonáticas en las litofacies Fl y Fm que incluyen amonites, bivalvos, crustáceos, huesos y fragmentos de madera. Los niveles son continuos lateralmente a nivel afloramiento (centenas de metros). Los fósiles pueden tener distintas improntas tafonómicas. Los bivalvos que se han registrado en concreciones son los siguientes: Cucullaea. (N.) gabrielis, Disparilia sp. indet., Modiolus cf. M. subsimplex, Pinna (P.) robinaldina, Panopea gurgitis (Brongniart), Ptychomya koeneni, Sphaera koeneni, Steinmanella pehuenmapuensis y Steinmanella transitoria. Ocasionalmente las concreciones pueden presentar incrustaciones externas de Amphidonte (C.) sp. y Parsimonia antiquata y perforaciones del tipo Gastrochaenolites isp. Los bivalvos pueden provenir de las tafofacies P1 o R1, los amonites y huesos de P1 y los fragmentos de troncos (sólo en Bajada del Agrio) se interpretan que han llegado por balsaje y posteriormente se han hundido hacia un fondo fangoso. Aparecen en zona de transición y plataforma media-externa.
Interpretación. Las concreciones tienen un origen diagenético temprano o sinsedimentario y relativamente cercano a la interfase agua-sedimento. Alojan fósiles con una variada impronta tafonómica adquirida previamente a la formación de la concreción. A partir de la forma de las concreciones es posible distinguir dos tipos particulares. Existen concreciones con valvas desarticuladas y fragmentadas cuya forma se puede relacionar con la generación de hoyos de erosión en el fango o pot casts, que se conectan corriente abajo con pequeños canales o gutter casts. Estas estructuras se forman por erosión del fondo submarino y son moldeadas por el movimiento de valvas en su interior (Aigner et al., 1978). Por otro lado hay concreciones que poseen restos de crustáceos que coinciden en su forma externa con partes de la traza Thalassinoides isp. (figura 5.B). Esto sugiere que una porción de dichas excavaciones puede concrecionarse durante la diagénesis temprana preservando restos, muchas veces completos, del organismo constructor (Aguire-Urreta, 2003). Esta tafofacies puede ser interpretada como una asociación promediada dentro del hábitat ya que posee modificaciones tafonómicas significativas pero no hay evidencias de mezcla entre ambientes y/o bioestratigráfica.

Figura 5. B, tafofacies P2, corte transversal mostrando tres niveles de trazas concrecionadas pertenecientes a Thalassinoides isp.B, taphofacies P2, transversal section showing three concretionary levels of Thalassinoides isp.
R1: Moluscos retrabajados
Descripción. Se compone de cefalópodos, gastrópodos y bivalvos retrabajados que se registran desde el shoreface hasta la plataforma. Los cefalópodos corresponden a conchillas y moldes internos de amonites y nautílidos. Se trata en general de fragmentos de fragmóconos cuyas cámaras pueden estar vacías o rellenas por material sedimentario concordante con la matriz de la roca hospedante. Los restos presentan orientación variable del plano de enroscamiento con respecto a la estratificación, pudiendo ser concordan te, oblicua o perpendicular (figura 5.C). Es relativamente común que los moldes y conchillas posean incrustaciones por Amphidonte (C.) sp. y Parsimonia antiquata en ambos flancos. Usualmente los incrustantes están desarticulados y fragmentados. Los gastrópodos pertenecen a las siguientes especies: Pleurotomaria gerthi Weaver, Tylostoma jaworskii Weaver y Naticoidea gen. et sp. indet., interpretados como epifaunales herbívoros, y Protohemichenopus neuquensis Camacho, interpretado como infaunal suspensívoro. Las modificaciones tafonómicas incluyen la fragmentación del ápice, la rotura del borde de la última vuelta con la pérdida de digitaciones espiniformes, el aplastamiento lateral y la incrustación externa por Amphidonte (C.) sp. y Parsimonia antiquata. El relleno de las conchillas es de arenisca o coquina y es siempre concordante con la roca hospedante, aunque pueden aparecer conchillas vacías en tempestitas fangosas. Los bivalvos son epifaunales, semiinfaunales e infaunales. Las especies involucradas son: Aetostreon sp., Amphidonte (Ceratostreon) sp., Aphrodina (A.) quintucoensis (Weaver), Astartidae gen. et. sp. indet., Cucullaea (Noramya) gabrielis, Disparilia sp. indet., Eriphyla argentina Burckhardt, Gervillaria alatior (Imlay), Gervillella aviculoides (Sowerby), Isognomon (I.) ricordeanus (d'Orbigny), Isognomon (I.) lotenoensis (Weaver), Limidae gen. et sp. indet., Mimachlamys robinaldina (d'Orbigny), Modiolus cf. M. subsimplex, Myoconcha transatlantica Burckhardt, Mytilus sp. indet., Panopea gurgitis, Pholadomya gigantea (Sowerby), Pinna (P.) robinaldina, Pterotrigonia sp. indet., Ptychomya koeneni, Sphaera koeneni, Steinmanella pehuenmapuensis, Steinmanella transitoria y Trigonia carinata Agassiz. Los ejemplares relevados presentaron desarticulación muy alta, fragmentación baja y modificación de bordes muy alta. Tanto la incrustación como la bioerosión externa son intermedias. El resto de los índices son bajos. La orientación de las conchillas es variable entre convexo hacia arriba, convexo hacia abajo y verticales (figura 5.D). En el caso en que exista relleno, éste es siempre concordante con la matriz de la roca hospedante.

Figura 5. C, tafofacies R1, vista en planta mostrando un molde interno de un nautílido en posición vertical.C, taphofacies R1, plan view of articulated specimens of S. transitoria and C. (N.) gabrielis, both out of life position.

Figura 5.D, tafofacies R1, vista en planta mostrando a la izquierda un ejemplar articulado de Steinmanella transitoria y a la derecha uno de Cucullaea (N.) gabrielis, ambos están articulados y fuera de su posición de vida.D, taphofacies R1, plan view of an internal mould of a nautiloid oriented vertically.
Interpretación. La muy alta desarticulación, la presencia de cámaras rellenas y los niveles intermedios de incrustación y bioerosión externas indica que las conchillas estuvieron expuestas un cierto tiempo en la zona tafonómicamente activa (TAZ), sin embargo niveles de fragmentación y corrasión bajos indican que la energía del ambiente y el transporte lateral no fueron significativos. Los moldes internos fragmentados e incrustados permiten inferir una historia tafonómica compleja con al menos un ciclo de enterramiento- exhumación en un proceso denominado reelaboración. Esto quiere decir que las conchillas se rellenaron con material sedimentario, se enterraron y cementaron. Luego, los moldes internos incluyendo restos de tabiques y muralla fueron exhumados al menos una vez por algún evento erosivo el cual los expuso sobre la interfase agua-sedimento siendo susceptibles de ser modificados. El enterramiento no fue muy profundo ya que los moldes pudieron ser erosionados. El hecho de que la mayoría posea una matriz concordante con la de la roca hospedante implica que la reelaboración se produjo dentro del mismo ambiente sedimentario.
Esta tafofacies puede ser interpretada como una asociación promediada dentro del hábitat ya que posee modificaciones tafonómicas significativas, pero no hay evidencias de mezcla entre ambientes y/o bioestratigráfica. Los moldes internos reelaborados con matriz concordante son típicos de dichas asociaciones (Kidwell, 1998).
R2: serpúlidos coloniales retrabajados
Descripción. Se compone de serpúlidos coloniales de la especie Sarcinella occidentalis (Leanza y Castellaro), retrabajados y fuera de su posición de vida. La preservación más común es en fragmentos subcilíndricos de 10 cm de longitud máxima y 2-3 cm de diámetro. Cada fragmento es un manojo de tubos delgados cementados entre sí. En Bajada del Agrio, dentro de la subzona de Neocomites sp., se ha encontrado un fragmento más grande (70 x 55 x 15 cm) con forma de arbusto. Estos serpúlidos poseen un registro escaso en shoreface y zona de transición.
Interpretación. A pesar de ser colonial, S. occidentalis nunca alcanzó el tamaño de los agregados de serpúlidos (véase tafofacies Gr) sino que habría formado pequeños arbustos aislados sobre el fondo. Este hallazgo concuerda con el hecho de que los serpúlidos coloniales actuales tampoco forman estructuras grandes (Ten Hove com. pers., 2003). Esta tafofacies puede ser interpretada como una asociación promediada dentro del hábitat ya que posee modificaciones tafonómicas significativas sin evidencias de mezcla entre ambientes y/o bioestratigráfica.
R3: corales retrabajados
Descripción. Se compone de corales coloniales de las especies Columastrea antiqua (Gerth) y Placocoenia neuquensis Gerth. Se restringe a facies de shoreface únicamente en el perfil Bajada del Agrio. C. antiqua corresponde a colonias ramosas que nacen de colonias incrustantes que a su vez se cementan sobre conchillas articuladas y desarticuladas de bivalvos. Las ramas pueden alcanzar unos 15-20 cm de altura y un diámetro máximo de 4,5 cm. P. neuquensis corresponde a colonias globosas de hasta 17 cm de diámetro y 13 cm de altura. Las formas ramosas son mucho más abundantes que las globosas, las cuales se encuentran sólo ocasionalmente. Todas las ramas relevadas estaban fragmentadas, aunque el grado de corrasión es bajo. La gran mayoría de ellas (97,1%) se encontraba en posición concordante y ninguna en posición de vida (figura 5.E). El grado de incrustación es intermedio mientras que la bioerosión externa es muy alta. Los incrustantes más abundantes corresponden a Amphidonte (C.) sp., seguidos por igual proporción de Parsimonia antiquata y Demospongea indeterminada. La bioerosión corresponde a Gastrochaenolites isp. sin lineamientos internos.

Figura 5. E, tafofacies R3, vista en planta mostrando una alta concentración de corales ramosos pertenecientes a Columastrea antiqua, las ramas están fragmentadas y en posición concordante. E, taphofacies R3, plan view showing a high concentration of fragmented and concordant ramose corals of C. antiqua.
Interpretación. Dado que las colonias no se han encontrado en posición de vida se puede inferir que las mismas han sufrido retrabajo; sin embargo, dada la baja corrasión y que no existe homogeneización en la abundancia de las colonias entre perfiles, se puede inferir que el grado de transporte ha sido bajo. De esta forma las colonias serían parautóctonas. No obstante, el tiempo de exposición sobre el sustrato parece haber sido significativo ya que presentan incrustación y bioerosión. Es probable que los incrustantes y los perforantes hayan infestado a los corales luego de la muerte de los mismos, dado que no se han observado modificaciones en el crecimiento del coral en respuesta a la presencia de cementantes o perforantes ni lineamientos internos en las perforaciones (Morton, 1990). En arrecifes actuales los corales exhiben bioerosión de bivalvos y esponjas luego de tres años de exposición sobre la interfase (Chazzotes et al., 1995) y lo mismo se puede suponer para la tafofacies analizada. La restricción paleogeográfica de los corales está indicando que las condiciones para el asentamiento y el crecimiento de las colonias fue mejor en Bajada del Agrio, probablemente por una disminución en la tasa de sedimentación. Esta tafofacies puede ser interpretada como una asociación promediada dentro del hábitat ya que posee modificaciones tafonómicas significativas, pero sin evidencias de mezcla entre ambientes y/o bioestratigráfica.
R4: vertebrados retrabajados
Descripción. Se compone de restos de peces picnodontiformes (Osteichthyes, Actinopterygii, Neop terygii) y reptiles marinos correspondientes a plesiosaurios elasmosáuridos e ictiosaurios (Lazo y Cichowolski, 2003). Se trata de fragmentos de piezas dentarias, vértebras y otros fragmentos post-craneales desarticulados, fragmentados, corrasionados y muy aislados. Muchos de ellos presentan incrustaciones de Amphidonte (C.) sp. Aparece en shoreface y zona de transición.
Interpretación. Las carcazas de estos vertebrados llegaron al fondo donde sufrieron distintos procesos tafonómicos que llevaron a la desarticulación, fragmentación y corrasión externa de los huesos. Luego fueron removilizados por el oleaje de buen tiempo y de tormentas y fueron finalmente depositados en tempestitas. Esta tafofacies puede ser interpretada como una asociación promediada dentro del hábitat ya que posee modificaciones tafonómicas significativas, pero no hay evidencias de mezcla entre ambientes y/o bioestratigráfica.
Dc: Detrito conchil
Descripción. Se compone de conchillas, valvas y otros bioclastos triturados, de 0,2 a 4 cm de eje mayor (figura 5.F). La fracción menor a 0,2 cm se toma como parte de la matriz de la roca. El empaquetamiento, la selección de tamaño, la modificación de bordes y la orientación son muy variables. A veces se puede llegar a distinguir entre fragmentos recristalizados de aquellos que no lo están. Dentro de los que no recristalizan se encuentran las valvas de ostras, isognomónidos y pectínidos. Aparece en shoreface y zona de transición.

Figura 5. F, tafofacies Dc, corte transversal.F, taphofacies Dc, transvsersal section.
Interpretación. El detrito conchil probablemente deriva de la acción del oleaje de buen tiempo sobre bioclastos expuestos, los cuales pueden luego ser transportados hacia la plataforma por flujos de relajación de tormentas. En cuanto a la mezcla temporal, esta tafofacies puede ser interpretada como una asociación promediada dentro del hábitat o condensada ambientalmente ya que puede tener bioclastos que deriven de cualquiera de las tafofacies descriptas.
Vi: bivalvos en posición de vida
Descripción. Se compone de bivalvos infaunales y/o semi-infaunales en posición de vida (figura 5.G). Si bien ésta no es la única orientación posible, el porcentaje de bivalvos en posición de vida debe ser comparable o mayor al de las otras orientaciones. Los bivalvos registrados en posición de vida fueron Panopea gurgitis de hábito infaunal profundo; Cucullaea (N.) gabrielis de hábito infaunal somero y Pinna (P.) robinaldina de hábito endobisado. La mayoría de los ejemplares se encuentran articulados y enteros. Los niveles de corrosión y modificación de bordes son intermedios, mientras que los de incrustación y bioerosión externas son bajos. Ocasionalmente se registra la especie Cucullaea (N.) gabrielis articulada con cemento carbonático en su interior. Aparece en shoreface (figuras 2 y 6).
Figura 5. G, tafofacies Vi, vista en planta mostrando tres ejemplares de Cucullaea (N.) gabrielis en posición de vida en el tope de un banco de arenisca. G, taphofacies Vi, plan view of three specimens of C. (N.) gabrielis in life position at the top of a sandstone.

Figura 5. H, tafofacies Gr, corte transversal mostrando un agregado de Parsimonia antiquata. Fotos A, E, G y H de Bajada del Agrio; B-D y F de Agua de la Mula. Escalas gráficas en centímetros; tapa de lente de 5 cm de diámetro / Field pictures of taphofacies of the Pilmatué Member of the Agrio Formation. H, taphofacies Gr, transversal section of gregarious P. antiquata. A, E, G and H from Bajada del Agrio; B-D and F from Agua de la Mula. Graphic scales in centimeters; lens cap has 5 cm in diameter.
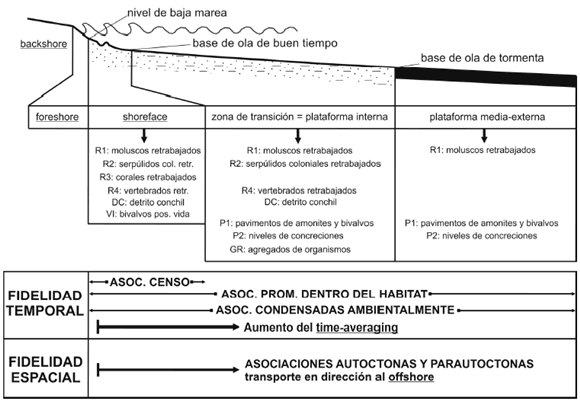
Figura 6. Perfil esquemático de un ambiente de shoreface-plataforma mostrando la distribución de las tafofacies y la fidelidad espacial y temporal de la macrofauna del Miembro Pilmatué de la Formación Agrio / Schematic shoreface-shelf profile showing the distribution of taphofacies and the degree of temporal and spatial mixing of the macrofauna of the Pilmatué Member of the Agrio Formation.
Interpretación. La presencia de bivalvos en posición de vida implica que se han sepultado rápidamente y han quedado por debajo de la TAZ, protegidos de los posibles efectos de bioturbadores y eventos erosivos. Los bivalvos articulados vacíos también indican un sepultamiento rápido dado que no alcanzaron a rellenarse con sedimento y es probable además que se hayan sepultado mientras estaban vivos. La presencia en un mismo horizonte de bivalvos cavadores someros y profundos en posición de vida implica la mezcla de dos escalones. Este tipo de mezcla ha sido descripta para las trazas fósiles, donde se habla de una capa histórica que contiene la superposición de escalones profundos sobre someros (Bromley, 1990). En este caso hay superposición de escalones de bivalvos lo que probablemente se produjo por efecto de la sedimentación progresiva y del ascenso en el tiempo de los cavadores profundos mezclándose con los someros. La presencia de corrasión y bordes modificados no indica en este caso retrabajo de los organismos, sino la disolución diagenética de la conchilla que deja a veces un aspecto alterado a los moldes internos y compuestos. Dado sus atributos esta tafofacies puede ser interpretada como una asociación de censo que refleja una mezcla temporal mínima de días- años en comparación con las asociaciones promediadas dentro del hábitat. En general son producto de un sepultamiento rápido y/o mortalidad en masa (Kidwell y Bosence, 1991).
Gr: agregados de organismos
Descripción. Se compone de agregados monoespecíficos de ostras y serpúlidos en sustratos fangosos (figura 5.H). Los taxones involucrados son Amphidonte (C.) sp. y Parsimonia antiquata. En ambos casos se trata de formas cementantes que ocurren bajo un hábito gregario. En forma muy dispersa pueden encontrarse amonites y bivalvos. Las modificaciones tafonómicas son: alta fragmentación, alta desarticulación, moderada incrustación y orientación variable para las ostras. En el caso de los serpúlidos los tubos poseen orientación concordante. Las conchillas y tubos pueden contener un relleno pelítico o pueden estar vacíos. Los rasgos sedimentológicos más característicos son los siguientes: empaquetamiento denso, mala selección (detrito conchil mezclado con bivalvos enteros), escasa matriz pelítica y base neta o erosiva. Los agregados están inmersos en litofacies Fl y Fm y su mayor expresión es en Bajada del Agrio donde llegan a tener 30 m de extensión lateral y 2,1 m de espesor. Aparece en zona de transición y reviste especial importancia en la subzona de Neocomites sp., hacia el tope de la subzona de Holcoptychites agrioensis y en la subzona de Olcostephanus (O.) laticosta.
Interpretación. El modo de ocurrencia de los agregados se asemeja a las bioconstrucciones de ostras y serpúlidos actuales y probablemente estuvieron sujetas a controles ambientales similares. Algunas especies de ostras y serpúlidos pueden, frente a un estímulo ambiental físico o químico, alcanzar el hábito gregario y formar bioconstrucciones e incluso arrecifes en bahías, estuarios, lagunas costeras, fiordos y plataformas (Stenzel, 1971;Ten Hove y van den Hurk, 1993). Los factores que controlan a las bioconstrucciones actuales son: tipo de sustrato, modo de fijación, nutrientes, tasa de sedimentación, temperatura, enfermedades, interacciones con otros organismos y salinidad (Norris, 1953; Puffer y Emerson, 1953). Según los rasgos tafonómicos se puede inferir que los organismos eran retrabajados frecuentemente. Las tormentas pueden haber ido rompiendo los agregados aportando fragmentos al sistema para ser transportados y/o modificados tafonómicamente por ejemplo por incrustantes. Para el crecimiento pudieron haber utilizado sustratos firmes o conchillas y tubos retrabajados, como ocurre en el caso de los corales coloniales. La decantación probablemente limitó el crecimiento de los agregados dado que el sedimento aumenta la mortalidad de individuos juveniles y adultos e inhibe la colonización por nuevas larvas. Cambios bruscos en la química del agua también pueden haber afectado su crecimiento. En base a los modelos actuales se podría pensar que en el momento en que los agregados crecían, la salinidad era distinta a la marina normal. Sin embargo la presencia de organismos típicamente marinos como los amonites indica lo contrario. La ausencia de macrobioerosión, aunque se trata de una evidencia negativa, también podría estar indicando un ambiente estresado. La monoespecificidad concuerda con un modelo de ambiente estresado. Dado que esta tafofacies posee mezcla temporal es probable que no se pueda resolver en forma delicada los cambios temporales y/o espaciales en la salinidad. En realidad lo que queda registrado es un promedio temporal de organismos que pudieron haber vivido bajo diferentes salinidades. Sería interesante testear los cambios de salinidad estudiando microfósiles, palinomorfos o isótopos lo cual está fuera del alcance de este trabajo. Las diferencias paleogeográficas en cuanto al desarrollo de los agregados son significativas. En Bajada del Agrio las condiciones para el gregarismo habrían sido óptimas, lo cual probablemente esté relacionado con un menor aporte clástico en comparación con el perfil Agua de la Mula el cual posee un mayor predominio de areniscas a lo largo de todo el perfil y está directamente relacionado con la zona de aporte (Marchese, 1971). Por otro lado tanto en Salado Sur como en Bajada del Agrio se puede ver que los agregados tienen su eje mayor ubicado en dirección este-oeste, en forma perpendicular a la paleocosta y a la corriente de deriva litoral, lo que también ocurre en los arrecifes actuales. Esta tafofacies puede ser tomada como una asociación condensada ambientalmente ya que involucra la superposición de sucesivas generaciones de ostras y serpúlidos mediando cambios en algún parámetro ambiental, posiblemente tasa de sedimentación y/o salinidad. Los agregados habitaban un ambiente estresado y de energía intermedia a alta.
Discusión
En cuanto al análisis paleoambiental realizado se infiere que, en la zona de estudio, el ambiente más cercano a la costa fue un shoreface arenoso caracterizado por un sustrato blando a bioclástico, energía intermedia a alta, nutrientes en suspensión, baja turbidez, 2-9 m de profundidad, salinidad marina normal y oxigenación normal. Estuvo influido por olas de buen tiempo y periódicamente por olas de tormentas. La sedimentación fue de tipo episódica donde se depositaron tempestitas que erosionaban depósitos previos y formaban bancos de tempestitas proximales amalgamadas. Este ambiente estuvo habitado por una fauna bentónica muy diversa dominada por bivalvos a los que se suman gastrópodos, serpúlidos coloniales, crustáceos y equinoideos. En Bajada del Agrio aparecen corales coloniales establecidos sobre conchillas retrabajadas, los cuales no alcanzaron a generar una barrera ambiental significativa sino que habrían formado un jardín de unos 15 cm de altura sobre el fondo.
La zona de transición estaba caracterizada por un sustrato fangoso producto de la decantación de material fino durante el buen tiempo, mientras que durante las tormentas llegaban arenas y bioclastos desde el shoreface depositándose como tempestitas proximales. La decantación de material fino no fue constante ya que la presencia ubicua de horizontes de concreciones carbonáticas indica que hubo pausas en la sedimentación. La consistencia del sustrato fangoso varió entre blando y firme. La zona de transición no estuvo tan ampliamente colonizada como el shoreface. Un bajo nivel de oxígeno o una consistencia soposa pudieron haber inhibido el asentamiento de organismos bentónicos. La presencia de abundante materia orgánica también apunta a fondos con bajo nivel de oxígeno. Sin embargo, estos fondos eran colonizados periódicamente, cuando las condiciones lo permitían, dado que se registra una variedad de asociaciones que pueden llegar a tener una alta densidad de organismos. Los agregados de ostras y serpúlidos se establecieron en la zona de transición, pero hacia el sector sur de la zona de estudio. En base a modelos actuales estos agregados se habrían desarrollado en un ambiente inestable relacionado probablemente con cambios en salinidad y/o tasa de sedimentación.
En la plataforma media-externa predominaba la decantación de material fino. A este sector de la plataforma sólo llegaban flujos de relajación distales muchas veces cargados de fango los cuales se depositaban como tempestitas distales y fangosas. Aquí disminuye aún más la presencia de organismos bentónicos. Es probable que la consistencia soposa del fondo o un bajo nivel de oxígeno inhibiera la colonización del fondo.
La distribución de tafofacies (figura 6) muestra que parte de la macrofauna no ha sufrido transporte lateral significativo preservándose en el lugar donde habitaba mientras que otra parte fue ligeramente desplazada en dirección al offshore. El transporte ocurrió a través de flujos de relajación que erosionaban organismos en el shoreface y zona de transición y los depositaban en zona de transición y plataforma media- externa. Incluso a pesar de haber sufrido retrabajo, los niveles de corales coloniales y los agregados de ostras y serpúlidos aún se ubican en un solo ambiente y con restricciones paleogeográficas importantes. Esto implica que las tormentas no tenían energía suficiente como para mezclar organismos de una localidad con otra. Estos resultados coinciden con datos obtenidos en estudios de tormentas actuales. Estos estudios demuestran que las tormentas no producen una mezcla lateral significativa de conchillas. Por lo tanto la distribución de conchillas muertas coincide con la distribución de la comunidad de la cual derivan (Miller et al., 1992).
El grado de mezcla temporal en promedio aumenta hacia el offshore (figura 6). En el shoreface se registra una mezcla de asociaciones con distinto timeaveraging reflejando procesos de sedimentación y erosión que se combinan en forma compleja. En cambio, hacia el offshore disminuye el poder erosivo de las tormentas y ocurre un descenso en la tasa de sedimentación. Esto genera acumulaciones fosilíferas más delgadas y simples que las del shoreface aunque con un mayor tiempo involucrado en su génesis. La retroalimentación tafonómica, el efecto de la bioturbación u otros factores biológicos pueden generar fábricas caóticas incluso en el offshore. Este resultado concuerda con estudios realizados sobre asociaciones de muerte en ambientes modernos (Kidwell, 1998). Debido a que existe mezcla temporal en las concentraciones analizadas cualquier estudio paleontológico o geológico que intente hilar muy fino en el tiempo puede errar sus conclusiones. Sin embargo, existen asociaciones de censo en el shoreface que dan una idea de la estructura de las comunidades del Miembro Pilmatué. Se las reconoce en el campo por presentar organismos en posición de vida, bajos índices tafonómicos y menores índices de diversidad y equitatividad que las promediadas ambientalmente.
Conclusiones
El Miembro Pilmatué en la zona de estudio fue depositado en un ambiente de shoreface-plataforma influido por tormentas sin presentar quiebres de pendiente hacia el offshore.
En los depósitos de shoreface predominan las trazas de sustrato blando de la icnofacies de Cruziana, mientras que hacia la plataforma se registran tanto trazas de sustrato blando en tempestitas como de sustrato firme de la icnofacies de Glossifungites en los depósitos pelíticos.
Hacia el sur de la zona de estudio, el Miembro Pilmatué presenta menor espesor, mayor amalgamación de capas, menor participación de clásticos y mayor expresión de agregados de ostras y serpúlidos. Todo señala que el perfil Bajada del Agrio se encontraba en una ubicación más proximal que los perfiles Agua de la Mula y Salado Sur.
El análisis tafofacial demostró que la mayoría de las asociaciones fósiles son autóctonas o ligeramente parautóctonas y que el grado de transporte lateral fue bajo y en dirección al offshore.
La mezcla temporal de las asociaciones fósiles es variable ya que se han registrado asociaciones de censo con mínima mezcla temporal (días-años) y asociaciones promediadas dentro del hábitat y condensadas ambientalmente con un promedio temporal mayor, desde años hasta 0,5 Ma. No se han registrado asociaciones condensadas bioestratigráficamente. La mezcla temporal aumenta en dirección al offshore.
Las concentraciones fósiles del shoreface tuvieron un origen sedimentológico complejo por repetidos procesos de erosión y sedimentación por tormentas aunque su time-averaging en promedio es bajo. Hacia la zona de transición y plataforma media-externa las concentraciones tuvieron un origen mixto biogénicosedimentológico y poseen en promedio mayor timeaveraging que las concentraciones del shoreface.
Agradecimientos
El autor desea agradecer a M.B. Aguirre-Urreta, A. Concheyro, P. Rawson, M. Cichowolski, D. Rodríguez, S. Lanés, R. Palma, G. Mángano y L. Buatois por su constante colaboración y estímulo. R. Sanci, C. Rodríguez y G. Iovine ayudaron en distintas tareas de campo. El financiamiento de esta investigación provino de una beca de formación de postgrado del CONICET y un subsidio para estudiantes de postgrado de la International Association of Sedmentologists ambos otorgados al autor, y los subsidios CONICET PIP 5960, PICT 14143 (ANPCyT) y UBACyT X084 a cargo de Aguirre-Urreta.
Bibliografía
Aguirre-Urreta, M.B. 2003. Early Cretaceous decapod Crustacea from the Neuquén Basin, west-central Argentina. Contributions to Zoology 72: 79-81.
Aguirre-Urreta, M.B. y Rawson, P.F. 1997. The ammonite sequence in the Agrio Formation (Lower Cretaceous), Neuquén Basin, Argentina. Geological Magazine 134: 449- 458.
Aguirre-Urreta, M.B. y Rawson, P.F. 2003. Lower Cretaceous ammonites from the Neuquén Basin, Argentina: the Hauterivian genus Holcoptychites. Cretaceous Research 24: 589-613
Aigner, T. y Reineck, H-E. 1982. Proximality trends in modern storm sands from the Helgoland Bight (North sea) and their implications for basin analysis. Senckenbergiana maritima 14: 183-215.
Aigner, T., Hagdorn, H. y Mundlos, R. 1978. Biohermal, biostromal and storm generated coquinas in the Upper Muschelkalk. Neues Jahrbuch für Geologie und Paläontologie Abhanlungen 157: 42-52.
Behrensmeyer, A.K., Kidwell, S.M. y Gastaldo, R.A. 2000. Taphonomy and paleobiology. Paleobiology 26: 103-147.
Brett, C.E. 1983. Sedimentology, facies and depositional environments of the Rochester Shale (Silurian; Wenlockian) in western New York and Ontario. Journal of Sedimentary Petrology 53: 947-971.
Brett, C.E. y Baird, G.C. 1986. Comparative Taphonomy: A key to paleoenvironmental interpretation based on fossil preservation. Palaios 1 : 207-227.
Bromley, R.G. 1990. Trace fossils. Biology and Taphonomy. Special Topics in Palaeontology 3, 280 pp.
Chazzotes, V., Le Campion-Alsumard, T. y Peyrot-Clausade, M. 1995. Bioerosion rates on coral reefs: interaction between macroborers, microborers and grazers (Moorea, French Polynesia). Palaeogeography, Palaeoclimatology, Palaeoecology 113: 189-198.
Cheel, R.J. y Leckie, D.A. 1993. Hummocky cross-stratification. En: V.P. Wright (ed.), Sedimentology Review 1, Blackwell Scientific Publications, Oxford, pp. 103-122.
Del Río, C.J., Martínez, S.A. y Scasso, R.A. 2001. Nature and origin of spectacular marine miocene shell beds of northeastern Patagonia (Argentina): paleoecological and bathymetric significance. Palaios 16: 3-25.
Dott, R.H.Jr. y Burgeois, J. 1982. Hummocky stratification: significance of its variable bedding sequences. Geological Society of America Bulletin 93: 663-680.
Fürsich, F.T. y Flessa, K.W. 1991. Ecology, taphonomy, and paleoecology of Recent and Pleistocene molluscan fauna of Bahía La Choya, northern Gulf of California. Zitteliana 18: 1-180.
Hampson, G.J. y Storms, J.E.A. 2003. Geomorphological and sequence stratigraphic variability in wave-dominated, shoreface-shelf parasequences. Sedimentology 50: 667-701.
Howard, J.D. y Reineck, H-E. 1981. Depositional facies of high-energy beach-to-offshore sequence: comparison with low-energy sequence. American Association of Petroleum Geologists Bulletin 65: 807-830.
Kidwell, S.M. 1991. The stratigraphy of shell concentrations. En: P.A. Allison y D.E.G. Briggs (eds.), Taphonomy: Releasing the data locked in the fossil record, Volume 9 of Topics in Geobiology, Plenum Press, New York, pp. 211- 290.
Kidwell, S.M. 1998. Time-averaging in the marine fossil record: overview of strategies and uncertainties. Geobios 30: 977-995.
Kidwell, S.M. y Bosence, D.W.J. 1991. Taphonomy and timeaveraging of marine shelly faunas. En: P.A. Allison y D.E.G. Briggs (eds.), Taphonomy: Releasing the data locked in the fossil record, Volume 9 of Topics in Geobiology, Plenum Press, New York, pp. 115-209.
Lazo, D.G. 2004. [Análisis de concentraciones fósiles del Cretácico Inferior de cuenca Neuquina. Tesis doctoral, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 337 pp., 7 perfiles. Inédito.].
Lazo, D.G. y Cichowolski, M. 2003. First plesiosaur remains from the Lower Cretaceous of the Neuquén basin, Argentina. Journal of Paleontology 77: 784-789.
Lazo, D.G., Cichowolski, M., Rodríguez, D.L. y Aguirre- Urreta, M.B. 2005. Lithofacies, palaeocology and palaeoenvironments of the Agrio Formation, Lower Cretaceous of the Neuquén Basin, Argentina. En: G.D. Veiga, L.A. Spalletti, J. Howell y E. Schwarz (eds.), The Neuquén Basin: A case study in sequence stratigraphy and basin dynamics, Geological Society of London, Special Publication 252: 295-315.
Leanza, H.A. 2003. Las sedimentitas huitrinianas y rayosianas (Cretácico Inferior) en el ámbito central y meridional de la cuenca Neuquina, Argentina. Serie Contribuciones Técnicas, Geología 2, Servicio Geológico Minero Argentino, 31 pp.
Leanza, H.A., Marchese, H.G. y Riggi, J.C. 1977. Estratigrafía del Grupo Mendoza con especial referencia a la Formación Vaca Muerta entre los paralelos 35° y 40° L.S., cuenca Neuquina-Mendocina. Revista de la Asociación Geológica Argentina 32: 190-208.
Legarreta, L. y Gulisano, C.A. 1989. Análisis estratigráfico secuencial de la cuenca Neuquina (Triásico Superior- Terciario Inferior). En: G.A. Chebli y L.A. Spalletti (eds.), Cuencas Sedimentarias Argentinas, Instituto Superior de Correlación Geológica (INSUGEO), Serie Correlación Geológica 6, pp. 221-243.
Marchese, H.G. 1971. Litoestratigrafía y variaciones faciales de las sedimentitas mesozoicas de la cuenca Neuquina, provincia de Neuquén, República Argentina. Revista de la Asociación Geológica Argentina 26: 343-410.
Miller, A.I., Llewellyn, G., Parsons, K.M., Cummins, H., Boardman, M.R., Greenstein, B.J. y Jacobs, D.K. 1992. Effect of hurricane Hugo on molluscan skeletal distributions, Salt River Bay, St. Croix, U.S. Virgin Islands. Geology 20: 23-26.
Morton, B. 1990. Corals and their bivalve borers-The evolution of a symbiosis. En: B. Morton (ed.), The Bivalvia, Proceedings of a memorial symposium in honour Sir Charles Maurice Yonge (1899-1986), Edinburgh, 1986, Hong Kong Univrsity Press, Hong Kong, pp. 11-46.
Norris, R.M. 1953. Buried oyster reefs in some Texas bays. Journal of Paleontology 27: 569-576.
Palma, R.M. y Lanés, S. 2001. Shell bed stacking patterns in the Chachao Formation (Early Valanginian) in Malargüe Area, Mendoza province, Neuquén Basin-Argentina. Carbonates and Evaporites 16: 168-180.
Puffer, E.L. y Emerson, W.K. 1953. The molluscan community of the oyster-reef biotope on the central Texas coast.Journal of Paleontology 27: 537-544.
Speyer, S.E. y Brett, C.E. 1986. Trilobite taphonomy and Middle Devonian taphofacies. Palaios 1: 312-327.
Stenzel, H. 1971. Oysters. En: R.C. Moore y C. Teichert (eds.), Treatise on Invertebrate Paleontology. Part N. Mollusca 6.Bivalvia Volume 3, University of Kansas and Geological Society of America, Lawrence, pp. N953-N1224.
Ten Hove, H.A. y van den Hurk, P. 1993. A review of recent and fossil serpulid "reefs"; actuopalaeontology and the "Upper Malm" serpulid limestones in NW Germany. Geologie en Mijnbouw 72: 23-67. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 6 de octubre de 2004.
Aceptado: 7 de julio de 2005.

