










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.2 Buenos Aires abr./jun. 2006
Molluscan assemblages from the marine Holocene of Uruguay: composition, geochronology, and paleoenvironmental signals
Sergio Martínez1, Alejandra Rojas1, Martín Ubilla1, Mariano Verde1, Daniel Perea1 and Graciela Piñeiro1
1 Departamento de Evolución de Cuencas Facultad de Ciencias, Iguá 4225, 11400 Montevideo, Uruguay. smart@fcien.edu.uy
Abstract. Dates of marine Holocene molluscs from Uruguay (Villa Soriano Formation) range from 6849 to 1858 cal. years BP. There are no significative gaps among the different ages obtained, with the exception of the interval 2276-2830 calculated years BP. Two kinds of concentrations are distinguished: autochthonous, for which a low energy depositional environment is inferred, and parautochthonous, deposited in foreshore and nearshore high-energy environments. Results of different types of multivariate analyses on presence-absence and abundance data show that only three localities are clearly differentiated from the others. The discrimination of two of them (Arazatí, Las Cañas) is explained by the presence of only one or two species, but the third (Punta Rasa) is the only one where characteristic species of rocky or hard substrates are undoubtedly represented . Analyses of salinity and temperature ranges of the various mollusc assemblages suggest a displacement of the Rio de la Plata estuary about 300 km westwards along the Uruguayan coast from its present position, and the presence of warmer waters than those found today at the same latitude. With the exception of the currently northwards displaced Anomalocardia brasiliana (Gmelin), Marshallora nigrocincta (C.B. Adams), Nioche subrostrata (Lamarck), Bulla striata Bruguìere and Miralda sp., all species are present today at the Uruguayan coast.
Resumen. Asociaciones de moluscos del holoceno marino de Uruguay: composición, geocronología e indicadores paleoambientales. Las asociaciones de moluscos marinos holocenos contenidos en la Formación Villa Soriano (Uruguay) presentan un rango máximo de 6849-1858 años cal. AP. No existen discontinuidades significativas entre las diversas edades obtenidas, con la excepción del intervalo 2276-2830 años calculados AP. Se distinguen dos tipos de concentraciones: autóctonas, propias de un ambiente de baja energía, y parautóctonas, depositadas en ambientes de alta energía del foreshore y nearshore. Los resultados de diferentes técnicas de análisis multivariado sobre datos de presencia-ausencia y abundancia de especies muestran que solamente tres se distinguen claramente de las demás localidades. La separación de dos de ellas (Arazatí, Las Cañas) se explica porque presentan una o dos especies solamente, en tanto que la tercera (Punta Rasa) es la única donde están claramente representados elementos propios de fondos duros. Sobre la base de las asociaciones estudiadas, se infiere un desplazamiento del frente estuárico del Río de la Plata de aproximadamente 300 km sobre la costa uruguaya, y una temperatura del agua mayor que la actual para la latitud considerada. Con la excepción de Anomalocardia brasiliana (Gmelin), Marshallora nigrocincta (C.B. Adams), Nioche subrostrata (Lamarck), Bulla striata Bruguìere y Miralda sp., cuyo límite de distribución meridional presente se encuentra más al norte, todas las especies viven hoy día en la costa de Uruguay.
Key words. Uruguay; Holocene; Paleoecology; Shell-beds. Mollusca; Bivalvia; Gastropoda.
Palabras clave. Uruguay; Holoceno; Peleoecología; Concentraciones fosilíferas; Mollusca; Bivalvia; Gastropoda.
Introduction
Quaternary shelly marine deposits are widespread along the Atlantic coast of South America (Clappertton, 1993; Aguirre and Whatley, 1995; Suguio, 1999; Pastorino, 2000; Martínez et al., 2001, and references therein). Although poorly known, the geographic location of the Uruguayan deposits in the Río de la Plata area constitute an interesting case for studying fluctuations in biological and environmental parameters, including faunal distributions, temperature and salinity.
The earliest published papers on Quaternary marine fossiliferous deposits from Uruguay are those by d'Orbigny (1842) and Darwin (1846). However, the first reference belongs to Larrañaga, who recognised shell-beds in 1819, but whose observations were not published until 1894. Since then, several studies have been carried out on those deposits, focalizing on their geology, paleontology and cartography, but many questions about the marine Quaternary of Uruguay remain unanswered. In this paper we focus on widespread Holocene molluscan associations, their radiocarbon ages, and the information that these associations imply about temperature and salinity, using a uniform sampling strategy and a taxonomic approach.
Geological setting and study area
The deposits bearing the mollucs presently known as Villa Soriano Formation have been cited with different names along the years. Some of the terms arose from Argentine stratigraphy (e.g. "Querandinense", "Platense", or Querandina Formation), or are local expressions ("Arcillas grises de Vizcaíno", Vizcaíno Formation, Villa Soriano Formation). A summary of the nomenclature of Uruguayan Quaternary is presented in figure 1.
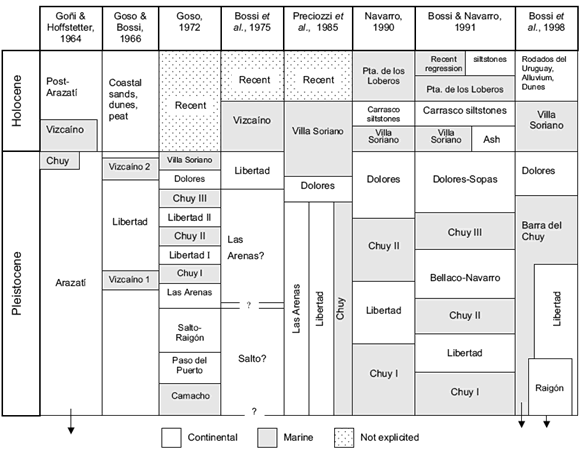
Figure 1. Units and ages proposed for the Uruguayan Quaternary (from Martínez et al., 2001) / unidades y edades propuestas para el Cuaternario de Uruguay (tomado de Martínez et al., 2001).
The highly fossiliferous Villa Soriano Formation crops out patchily along a narrow strip parallel to the present coast line, spreading from the mouth of the Negro river to the Merín lagoon margins. According to most authors (e.g. Preciozzi et al., 1985), this unit comprises claystones, medium sandstones and conglomerates of up to 9 meters of thickness. However, due to its wide lithologic definition the best criteria to define the litostratigraphic unit are the abundant shell content and the stratigraphic relationships, taking into consideration that the Villa Soriano Formation is not overlaid by any unit (Martínez and Ubilla, 2004).
The published radiocarbon ages appear either in abstracts or papers lacking information about taphonomic/ diagenetic conditions or proper taxonomic identification (e.g. Cortelezzi and Lerman, 1971, Bracco et al., 1994, García et al., 2002).
The Villa Soriano Formation has often been correlated with units from Argentina (e.g. "Platense", "Querandinense", Las Escobas Formation) (Goñi and Hoffstetter, 1964; Aguirre and Whatley, 1995). Forti- Esteves (1974) and Martínez (1990) correlated it with the Patos Group of Rio Grande do Sul, Brazil.
It has been extensively discussed whether these sediments have been originated in one or more transgressive events (e.g. Figueiras, 1961, 1962; Goso and Bossi, 1966; Sprechmann, 1978; Preciozzi et al., 1985), but no conclusive data has been presented yet.
Molluscan content has been mentioned by several authors, such as Ihering (1907) Teisseire (1928), De Mata (1947) Figueiras (1961, 1962, 1967), but mostly limited to species identification. Sprechmann (1978) intended to reconstruct past depositional environments on the basis of foraminifers and molluscs, but some information about the latter group was compiled from very heterogeneous data (of sampling effort or taxonomic criteria), and Pleistocene and Holocene deposits still had not been discriminated, and were treated as contemporaneous.
The area of study comprises the Uruguayan coastline in the zones where the shell-bearing deposits of Villa Soriano Formation outcrop. Localities where collection took place are shown in figure 2.

Figure 2. Localities mentioned in the text / localidades mencionadas en el texto.
Taphonomy and depositional environment
According to our own observations, two kinds of shell-beds have been found: 1) autochthonous assemblages, and 2) parautochthonous shell concentrations.
Autochthonous associations (La Caballada, Arazatí and Playa Pascual, see figure 3) are defined by the predominance of articulated valves in life position found in frequently bioturbated mudstones, which are very rich in organic content. Shells are unsorted by size. A low energy regime is clear, and the depositional environment is interpreted as coastal lagoons.

Figure 3. Stratigraphic sections of the localities studied. Samples were taken from levels shown by the arrows. c = clay, s = silt, fs = fine sand, ms = medium sand, cs = coarse sand, p = pebble. 0 = local base of stratigraphic section / secciones estratigráficas de las localidades estudiadas. Las muestras fueron tomadas de los niveles señalados con las flechas. c = arcilla, s = limo, fs = arena fina, ms = arena media, cs = arena gruesa, p = grava. 0 = base local de la sección estratigráfica.
Parautochthonous shell concentrations (the remaining localities) are composed of accumulations of disarticulated, sometimes broken and abraded, mostly chaotically arranged (although sometimes nested) shells, in a coarse to medium sandy matrix, showing parallel or cross-bedded stratification. They are interpreted as littoral bars deposited in foreshore to near shore high-energy environments, strongly influenced by waves and currents.
Material and methods
After making taphonomic and stratigraphic observations, bulk sample units of about 3 dm3 of the mollusc bearing levels were collected at the localities shown in figure 2. Stratigraphic sections for each locality studied are shown in figure 3.
Data on the present distribution, temperature and salinity tolerance of the molluscs (Appendix 1) were taken from Figueiras and Sicardi (1968-1974, 1979, 1980), Sprechmann (1978), Layerle and Scarabino (1984), Di Persia and Olazarri (1986), Mansur et al. (1991), Rios (1994), and Scarabino (2003, 2004). Regarding salinity, species were classified into two categories: estuarine and marine (no limnic species were found). The first one comprises species considered true estuarine, that is, those that only live in brackish water. Since this was the delimiting criterion, the remaining species were considered to be marine, regardless their tolerance to salinity changes. To be euryhaline implies a potential attribute, not a true environmental restriction. In relation to temperature, species were grouped according to their origin: tropical- subtropical, those that are shared with northern warmer areas (Brazilian Province); cold, those shared with the southern colder areas (Magallanic Province); and endemic, those present only in the Argentinean Province.
Specimens were counted (right and left valves of bivalves were counted separately, taking into account the maximum number), and relative abundance of species was expressed as (i) percentages (see appendix 1), and (ii) intervals: 1 (>50%), 2 (50%- 25%), 3 (25%-5%), 4 (<5%).
Similarity-dissimilarity in mollusc species content among the localities (presence-absence) was asserted using both Dice association Coefficient and Raup and Crick (1979) Index. (Dice Coefficient= 2a/(2a+b+c), where "a" is the number of taxa shared by the samples; "b" and "c" are the taxa present in one sample and absent from the other; RC Index= the probability that kerl will be less than or equal to kobs, being kerl theoretically possible values of k generated by Monte Carlo simulations; kobs the number of taxa actually shared by assemblages, and k the number of taxa common to both assemblages). Although Dice Coefficient provides a minimum distortion in binary data (Archer and Maples, 1987; Maples and Archer, 1988), the Raup and Crick Index was also calculated because it uses a randomization of Monte Carlo procedure. Non Metric Multidimensional Scaling, minimum spawning tree, and Q and R modes (UPGMA) Cluster Analysis were carried out. Correspondence analyses were performed for abundance data. It is an ordination method that uses eigenanalysis of the Chi-squared distances between all data, very appropiated for counted data. (Kent and Coker, 1994; Hammer et al., 2001). PAST 1.34 (Hammer et al., 2001) was the computer program used.
Mineralogical alteration of the shells used for the radiocarbon dating was evaluated by analysing their mineralogical composition using X-ray diffraction. Four samples showed some degree of mineralogical alteration (La Caballada, San Luis, Chuy, Saglia 2B), however, their results do not disagree with the range of ages obtained by the other samples.
Radiocarbon and X-ray diffraction analyses were performed at the Laboratorio de Tritio y Radiocarbono (LATYR) at the Museo de La Plata (Argentina). Ages were calibrated using the program CALIB 4.4.1 (Stuiver and Reimer, 1993), with the Stuiver et al. (1998) calibration dataset, assumed 15+ -2 per mil delta 13C, and 82+ -46 delta R.
Samples used for dating in La Caballada, Arazati, and Playa Pascual were articulated valves found in life position (autochthonous). but the others were isolated valves recovered from parautochthonous assemblages that presumably represent time-averaged accumulations (single or multiple events). Flessa et al. (1993), Flessa and Kowalewski (1994), Wehmiller et al. (1995), Schellmann and Radtke (1997), and Kowalewski et al. (1998), have demonstrated time averaging from hundreds (usually) up to three thousands years, for recent and near shore Quaternary shell-beds. Estimations of the exact extent of this time averaging are strongly dependent on several variables for each shell-bed. In our case, for example, and due to reworking (although alteration may be playing a role), we found in Villa Soriano a stratigraphical lower sample with a radiocarbon age around 300 years younger than the upper sample. Although it gives some uncertainty, both are within the range of ages presented here.
Specimens are deposited in the collection of invertebrate fossils, Facultad de Ciencias, Montevideo, Uruguay (FCDPI).
Results and discussion
Age
Radiocarbon ages and related data are shown in table 1. According to our results, mollusc-bearing Holocene deposits of Uruguay would have a maximum range of 6849-1858 cal. years B.P. These results are similar to the radiocarbon ages obtained for the Argentinean (Buenos Aires province) Las Escobas Formation, with ranges from 7600 to 1400 14C yr B.P. (Aguirre and Whatley, 1995; Aguirre and Urrutia, 2002). However, Rojas (2002) has reported in a brief communication an age of 9830 +/- 90 14C yr B.P. for a Uruguayan locality (Sauce creek, near Nueva Palmira city) a datum that must be confirmed.
Table 1. Radiocarbon and calculated ages for molluscs and additional data / edades radiocarbónicas y calculadas de los moluscos y datos complementarios.

As is shown in figure 4, age-ranges make a rather compact group, with a pair of exceptions, that would be Los Rodríguez and Punta Rasa. The range of the former does not overlap with others in general, but just for a brief period of time (ca. 100 years), and it can be safely included into the group. On the other hand, Punta Rasa is clearly disconnected from the other ages, being the youngest of all. According to our data, if the age hiatus between Punta Rasa and the other lo calities is consequence of a lowering of sea level and represents a true absence of a marine record, it would be the only relevant oscillation registered.

Figure 4. Age ranges (bars) for the studied localities / rangos de edades (barras) de las localidades estudiadas.
Paleoecology
Remarks on faunal composition. Recorded taxa are listed by locality in Appendix 1. With the exceptions of Anomalocardia brasiliana (Gmelin), Marshallora nigrocincta (C.B. Adams), Nioche subrostrata (Lamarck), Bulla striata Bruguìere, and Miralda sp., all species are present today along the Uruguayan coast. The geographic distribution of Anomalocardia brasiliana and Bulla striata ranges from the Caribbean region to Rio Grande do Sul (Brazil), Marshallora nigrocincta from North Carolina (USA) to Santa Catarina (Brazil), and Nioche subrostrata from Florida (USA) to São Paulo (Brazil). The genus Miralda is present from Florida to northern Brazil (Rios, 1994).
Some taxa are recorded for the Uruguayan Quaternary for the first time: Nuculana janeiroensis E. A. Smith, Crepidula plana Say, Miralda sp., and a representative of the Family Ellobidae, not determined yet.
Results of Cluster analysis and NMDS (figure 5) show that three localities are discriminated from the others: Punta Rasa in all cases, and Arazatí and Las Cañas in some (both in dendrograms applying Dice coefficient, and Arazatí in the result of cluster analysis applying Raup and Crick coefficient). The two latter differ from the other localities in being monospecific or almost [Las Cañas: Erodona mactroides Daudin; Arazatí: Ostrea equestris Say, and Ennucula puelcha (d'Orbigny)], but Punta Rasa is the richest in number of species. It includes some species that are not shared with any other site. These are: Glycymeris longior (G. B. Sowerby I), Brachidontes darwinianus (d´Orbigny), Plicatula gibbosa Lamarck, Crassinella maldonadoensis (Pilsbry), Amiantis purpuratus (Lamarck), Lottia subrugosa (d´Orbigny), Diodora patagonica (d´Orbigny), Tegula patagonica (d´Orbigny), Calliostoma jucundum (Gould), Nodiottorina lineolata (d'Orbigny), Seila adamsi (H.C. Lea), Urosalpinx cala (Pilsbry), Hanetia haneti (Petit de la Saussaye), Siphonaria lessoni (Blainville), Ellobidae indet. It is suggestive the high proportion of species that inhabit rocky or hard substrates (B. darwinianus, P. gibbosa, L. subrugosa, D. patagonica, H. haneti, U. cala, S. lessoni, see Appendix 1), the maximum among the samples studied.

Figure 5. A, Two-way cluster analysis, Dice coefficient. B, NMDS with superimposed minimum spawning tree, Dice coefficient. C, Twoway cluster analysis, Raup-Crick coefficient. D, NMDS with superimposed minimum spawning tree, Raup-Crick coefficient. Codes of localities and species as in Appendix 1 / A, Análisis de agrupamiento de dos vías, coeficiente de Dice. B, NMDS con árbol de distancias mínimas sobreimpuesto, coeficiente de Dice. C, Análisis de agrupamiento de dos vías, coeficiente de Raup y Crick. D, NMDS con árbol de distancias mínimas sobreimpuesto, coeficiente de Raup y Crick. Las codificaciones de las localidades son las mismas del Apéndice 1.
When analyzing abundance (figure 6, Correspondence analysis), Arazatí and Punta Rasa are also differentiated from the rest of the localities, but Las Cañas is grouped with others (especially Punta Pereira 1 and 2) where Erodona mactroides largely predominates. Arazatí is distinguished either by the presence of Ennucula puelcha, when taking into account abundance intervals, or Ostrea equestris, when using percentages. Considering abundance intervals, Punta Rasa is related with Mytilus edulis Linné, and following percentages with Glycymeris longior, Brachidontes darwinianus, Plicatula gibbosa, Crassinella maldonadoensis, Amiantis purpuratus, Lottia subrugosa, Diodora patagonica, Tegula patagonica, Seila adamsi, Urosalpinx cala, Siphonaria lessoni, Ellobidae indet., Corbula lyoni Pilsbry, Corbula sp. and Tawera gayi (Hupé). This species list is very similar to the resultant of the presence-absence data, with the addition of the two species of Corbula and T. gayi, and the elimina tion of Calliostoma jucundum, Nodilittorina lineolata and Hanetia haneti.

Figure 6. A, Correspondence analysis, abundance of species expressed as intervals (see text). Only the species close to Punta Rasa and Arazatí are included, since the others are too crowded in the remaining localities. B, Correspondence analysis, abundance of species expressed as percentages. Only the species close to Punta Rasa, Arazatí and Las Cañas are included, since the others are too crowded in the remaining localities. Codes of localities and species as in Appendix 1 / A, Análisis de Correspondencia, la abundancia de las especies tomada en intervalos (ver texto). Solamente las especies cercanas a Punta Rasa y Arazatí se han incluido, ya que las demás forman un grupo muy compacto junto con las restantes localidades. B, Análisis de Correspondencia, la abundancia de las especies tomada como porcentajes. Solamente las especies cercanas a Punta Rasa, Arazatí y Las Cañas se han incluido, ya que las demás forman un grupo muy compacto junto con las restantes localidades.
Salinity
Species were classified according to their salinity range (see Appendix 1), and the relative frequency of salinity groups was calculated for each sample. This information is summarized in figure 7, altogether with the current distribution of mollusc salinity ranges.

Figure 7. Proportion (by locality) of molluscan species according to their salinity range / proporciones por localidad de las especies de moluscos según su rango de salinidad.
We then propose that the marine front during the Holocene was further to the northwest than at present times (about 300 km farther) (figure 8), beyond the limits proposed by Sprechmann (1978). This conclusion must be interpreted as a general trend throughout the estimated age range of the deposits (taking into account the shell-beds time averaging).

Figure 8. Holocene and Present Uruguayan coastal area environments according to mollusc content / ambientes del Holoceno y Actual de la costa Uruguaya de acuerdo a sus moluscos.
Temperature
Species were also classified according to their thermal ranges (see Appendix 1), and the relative frequencies of these classes compared with the modern ones, according to the data of Sicardi (1967). The results (figure 9) indicate that for the time interval considered, a greater proportion of warm-water species and a smaller proportion of cold-water species characterized the fossil mollusc assemblages. Furthermore, there are in the Holocene assemblages five warm water species that do not live today along the Uruguayan coast, having their southern distribution limit displaced northwards. Stronger south ward-flowing warm Brazil currents could be postulated as a mechanism to explain warmer temperatures in the Quaternary (Martínez, 1990; Martínez et al., 2001). This would be reinforced by the shift of the estuarine and marine fronts of the Río de la Plata towards the West, given that today the decrease in salinity caused by the discharge of fresh water from the Rio de la Plata is a significant barrier for many subtropical species (Scarabino, 1977; Piola et al., 2000).

Figure 9. Proportion (by locality) of molluscan species according to their thermal range / proporciones por localidad de las especies de moluscos según su rango térmico.
Conclusions
The age of Holocene shell-beds from Uruguay range from 6849-1858 cal. years B.P. and indicate almost continuously a higher sea level than that present today.
The only possible gap could be between 2276 and 2830 cal. years B.P. (oldest and youngest calculated ages of Punta Rasa and Punta Pereira).
The salinity front of the Rio de la Plata estuary was displaced in the Uruguayan coast about 300 km westwards during the Holocene transgression. For the time interval considered, temperature was warmer than at present.
Nearly all the recorded species are present today along the Uruguayan coast.
Exceptions to the condition stated above are Anomalocardia brasiliana, Marshallora nigrocincta, Nioche subrostrata, Bulla striata and Miralda sp., whose present geographical ranges do not reach as far south as Uruguay.
Appendix 1. Recorded species by locality (percentages) and classification according to temperature and salinity ranges / especies registradas en cada localidad (abundancia expresada en porcentajes), y clasificación de acuerdo a sus rangos térmicos y de salinidad. * = less than 0.1 / menor que 0,1.
C: Las Cañas, VS1: Villa Soriano 1, VS2: Villa Soriano 2, P1: Punta Pereira 1, P2: Punta Pereira 2, Ca: La Caballada, A: Arazatí, Pa: Playa Pascual, F: La Floresta, R: Punta Rasa, C: Arroyo Chuy, SL: Arroyo San Luis, S1: Saglia 1, S2A: Saglia 2A, S2B: Saglia 2B, LR: Los Rodríguez.
TS: tropical-subtropical, AR: endemic to Argentinean Province, CO: cold, MA: marine, ES: estuarine. See Material and Methods for details. TS: tropical-subtropical, AR: endémica de la Provincia Argentina, CO: fría, MA: marina, ES: estuarina. Ver Material y Métodos por detalles.


Acknowledgments
This research was supported by Project 1023, CONICYTFondo "Prof. Clemente Estable". Claudia del Río improved earlier versions of the manuscript. Fabrizio Scarabino helped with some taxonomic identifications and ecologic data. The late Alfredo Figueiras transmitted generously his vast experience about Uruguayan molluscs. Richard Madden and Anita Eisenberg kindly gave assistance with the language.
References
Aguirre, M.L. and Urrutia, M.I. 2002. Morphological variability of Littoridina australis (d' Orbigny, 1835) (Hydrobiidae) in the Bonaerensian marine Holocene (Argentina). Palaeogeography, Palaeoclimatology, Palaeoecology 183: 1-23.
Aguirre, M.L. and Whatley, R.C. 1995. Late Quaternary marginal marine deposits and palaeonvironments from northeastern Buenos Aires Province, Argentina: a review. Quaternary Science Reviews 14: 223-254.
Archer, A.W. and Maples, C.G. 1987. Monte Carlo simulation of selected binomial similarity coefficients (I): effect of number of variables. Palaios 2: 609-617.
Bossi, J.C. and Navarro, R. 1991. Geología del Uruguay, 2. Departamento de Publicaciones, Universidad de la República. Montevideo, 966 pp.
Bossi, J.C., Ferrando, L.A., Fernández, N., Elizalde, G., Morales, H., Ledesma, J.J., Carballo, E., Medina, E., Ford, I. and Montaña, J.R. 1975. Carta Geológica del Uruguay. Escala 1:1.000.000. Dirección de Suelos y Fertilizantes, MAP. Montevideo, 32 pp.
Bossi, J., Ferrando, L., Montaña, J., Campal, N., Morales, H., Gancio, F., Schipilov, A., Piñeyro, D. and Sprechmann, P. 1998. Carta Geológica del Uruguay. Escala 1:500.000. CD-ROM Version 1.01, Montevideo.
Bracco, R., Navarro, R. and Ures, C. 1994. Primer intento cronológico de una ingresión holocénica en Carrasco (Montevideo, Uruguay). 1º Jornada de Paleontología del Uruguay (Montevideo), Resúmenes: 33-35.
Clappertton, C. 1993. Quaternary geology and geomorphology of South America. Elsevier. New York, 779 pp.
Cortelezzi, C.R. and Lerman, J.C. 1971. Estudio de las formaciones marinas de la costa atlántica de la Provincia de Buenos Aires. LEMIT 1: 77-83.
Darwin, C. 1846. Geological observations on South America, being the third part of the geology of the voyage of the Beagle, under the command of capt. Fitzroy, R.N. during the years 1832-1836. Smith, Elder and Co. London, 279 pp .
De Mata, O. 1947. La formación Holocena en el Departamento de Montevideo. Author´s Edition. Montevideo, 37 pp.
Di Persia, D.H. and Olazarri, J. 1986. Zoobenthos of the Uruguay system. In: B.R. Davies and K.F. Walker (eds.), The Ecology of River Systems, Junk Publishers, The Netherlands, pp. 623-629.
d' Orbigny, A. 1842. Voyage dans l' Amerique Meridionale (le Brésil, la République Orientale de l' Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivie, la République du Pérou, executé pendant les anné 1826-1833, 3, Geologie. P. Bertrand. Paris, 290 pp.
Figueiras, A. 1961. Contribución al conocimiento de la malacofauna holocena del Uruguay. Comunicaciones de la Sociedad Malacológica del Uruguay 1: 15-21.
Figueiras, A. 1962. Sobre nuevos hallazgos de moluscos subfósiles de la Transgresión Querandina. Comunicaciones de la Sociedad Malacológica del Uruguay 1: 53-68.
Figueiras, A. 1967. Contribución al conocimiento de los moluscos del Holoceno uruguayo. Comunicaciones de la Sociedad Malacológica del Uruguay 2: 61-76.
Figueiras, A. and Sicardi, O. E. 1968-1974. Catálogo de los moluscos marinos del Uruguay. Comunicaciones de la Sociedad Malacológica del Uruguay. 1968a. 2: 233-241. 1968b. 2: 255-273. 1969. 2: 355-376. 1970. 2: 407-421. 1970. 3: 15-22. 1970. 3: 25-34. 1972. 3: 101-127. 1973. 3: 169-186. 1973. 3: 259-286. 1974. 3: 323- 352.
Figueiras, A. and Sicardi, O.E. 1979. Catálogo de los moluscos marinos del Uruguay. Parte X. Revisión actualizada de los moluscos marinos del Uruguay con descripción de las especies agregadas. Sección I-Polyplacophora-Scaphopoda- Bivalvia. Comunicaciones de la Sociedad Malacológica del Uruguay 5: 107-157.
Figueiras, A. and Sicardi, O. E. 1980. Catálogo de los moluscos marinos del Uruguay. Parte X. Revisión actualizada de los moluscos marinos del Uruguay con descripción de las especies agregadas. Sección II-Gastropoda y Cephalopoda. Comunicaciones de la Sociedad Malacológica del Uruguay 5: 179- 272.
Flessa, K.W. and Kowalewski, M. 1994. Shell survival and time averaging in near shore and shelf environments: estimates from the radiocarbon literature. Lethaia 27: 153-165.
Flessa, K.W., Cutler, A.H. and Meldahl, K. H. 1993. Time and taphonomy: quantitative estimates of time-averaging and stratigraphic disorder in a shallow marine habitat. Paleobiology 19: 266-286.
Forti-Esteves, I.E. 1974. Bioestratigrafia e Paleoecologia (Mollusca) do Quaternário da Planicie costeira do Rio Grande do Sul (Brasil). 28º Congresso Brasileiro de Geologia (Porto Alegre), Anais: 133-149.
García, F., Castiñeira, C., Scharf, B. and Sprechmann, P. 2002. The relationship between trophic state and sea level variation in the Rocha Lagoon, Uruguay. Neues Jahrbuch für Geologie und Paläontologie, Mh. 2002: 27-47.
Goñi, J.C. and Hoffstetter, R. 1964. Uruguay. Lexique Stratigraphique. International, 5 Amérique. Latine, 9a. CNRS. Paris, 200 pp.
Goso, H. 1972. Cuaternario. Programa de Estudio y Levantamiento de Suelos, MGA. Montevideo, 12 pp.
Goso, H. and Bossi, J.C. 1966. Cenozoico. In: J.C. Bossi (ed.), Geología del Uruguay, Departamento de Publicaciones, Universidad de la República, Montevideo, pp. 259-305.
Hammer, Ø., Harper, D.A.T. and Ryan, P.D. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 4: 9 pp. http://palaeoelectronica.org/2001_1/past/issue1_01.htm
Ihering, H. von. 1907. Les Mollusques fossiles du Tertiaire et du Cretacé Supérieur de l' Argentine. Anales del Museo Nacional de Buenos Aires 3: 611 pp.
Kent, M. and Coker, P. 1994. Vegetation Description and Analysis, John Wiley and Sons, New York, 363 pp.
Kowalewski, M., Goodfriend, G.A. and Flessa, K.W. 1998. Highresolution estimates of temporal mixing within shell beds: the evils and virtues of time-averaging. Paleobiology 24: 287-304.
Larrañaga, D.A. 1894. Memoria geológica sobre la formación del Río de la Plata, deducida de sus conchas fósiles, escrita por los años 1819. Anales del Museo Nacional de Montevideo 1: 3-12.
Layerle, C. and Scarabino, V. 1984. Moluscos del frente marítimo uruguayo entre 9 y 78 m de profundidad: Análisis biocenológico. Contribuciones Departamento de Oceanografía (FHC) 1: 1-17.
Mansur, M.C.D., Schulz, C., da Silva, M.G.O. and de Campos- Velho, N.M.R. 1991. Moluscos Bivalves límnicos da estação ecológica do Taim e áreas adjacentes, Rio Grande do Sul, Brasil. Iheringia (Zoologia) 71: 43-58.
Maples, C.G. and Archer, A.W. 1988. Monte Carlo simulation of selected binomial similarity coefficients (II): effect of sparse data. Palaios 3: 95-103.
Martínez, S. 1990. Taphonomy and Paleoecology of Holocene mollusks from the western margin of the Merin Lagoon (Villa Soriano Fm., Uruguay). Quaternary of South America and Antarctic Peninsula 7: 121-135.
Martínez, S. and Ubilla, M. 2004. El Cuaternario de Uruguay. In: G, Veroslavsky, M. Ubilla and S. Martínez (eds.), Cuencas sedimentarias de Uruguay. Cenozoico, EUDECI, Montevideo, pp. 195-228.
Martínez, S, Ubilla, M., Verde, M., Perea, D., Rojas, A., Guérèquiz, R. and Piñeiro, G. 2001. Paleoecology and Geochronology of Uruguayan Coastal Marine Pleistocene Deposits. Quaternary Research 55: 246-254.
Navarro, R. 1990. A brief review on the knowledge on the Quaternary of the Southern and Southeastern Coast of Uruguay. Quaternary of South America and Antarctic Peninsula 7: 107-117.
Pastorino, G. 2000. Asociaciones de moluscos de las terrazas cuaternarias de Río Negro y Chubut, Argentina. Ameghiniana 37: 131-156.
Piola, A.R, Campos, E.J.D., Möller Jr., O.O., Charo, M. and Martínez, C. 2000. The subtropical shelf front off eastern South America. Journal of Geophysical Research 105: 6565-6578.
Preciozzi, F., Spoturno, J., Heinzen, W. and Rossi, P. 1985. Memoria explicativa de la Carta Geológica del Uruguay a la escala 1:500.000. DINAMIGE. Montevideo, 90 pp.
Raup, D. and Crick, R.E. 1979. Measurement of faunal similarity in paleontology. Journal of Paleontology 53: 1213-1227.
Rios, E. de C. 1994. Seashells of Brazil. Editora Fundação Universidade do Rio Grande. Rio Grande, 368 pp.
Rojas, A. 2002. Tafonomía y cronología de nuevos depósitos fosilíferos del Holoceno marino de Uruguay. 2º Jornadas uruguayas del Cenozoico (Montevideo), Actas: 80-82.
Scarabino V. 1977. Moluscos del Golfo San Matías (Provincia de Río Negro, República Argentina). Inventario y claves para su identificación. Comunicaciones de la Sociedad Malacológica del Uruguay 4: 177-285.
Scarabino, F. 2003. Lista sistemática de los Bivlavia marinos y estuarinos vivientes de Uruguay. Comunicaciones de la Sociedad Malacológica del Uruguay 8: 229-259.
Scarabino, F. 2004. Lista sistemática de los Gastropoda marinos y estuarinos vivientes de Uruguay. Comunicaciones de la Sociedad Malacológica del Uruguay 8: 305-346.
Schellmann, G. and Radtke, U. 1997. Electron Spin Resonance (ESR) techniques applied to mollusc shells from South America (Chile, Argentina) and implications for palaeo sealevel curve. Quaternary Science Reviews 16: 465-475.
Sicardi, O.E. 1967. La influencia de las corrientes marinas sobre la malacofauna uruguaya. Comunicaciones de la Sociedad Malacológica del Uruguay 2: 49-60.
Sprechmann, P. 1978. The paleoecology and paleogeography of the Uruguayan coastal area during the Neogene and Quaternary. Zitteliana 4: 3-72.
Stuiver, M. and Reimer, P. J. 1993. Extended 14C database and revised CALIB radiocarbon calibration program. Radiocarbon 35: 215-230.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen, K.A., Kromer, B., McCormac, F.G., v. d. Plicht, J. and Spurk, M. 1998. INTCAL98 Radiocarbon age calibration 24,000 - 0 cal BP. Radiocarbon 40: 1041-1083.
Suguio, K. 1999. Recent progress in Quaternary geology of Brazil. Episodes 22: 217-220.
Teisseire, A. 1928. Contribución al estudio de la geología y paleontología de la República Oriental del Uruguay. Región de Colonia. Anales de la Universidad 37: 319-469.
Wehmiller, J.F., York, L.L. and Bart, M.L. 1995. Amino acid racemiztion geochronology of reworked Quaternary mollusks on U.S. Atlantic coast beaches: implications for chronostratigraphy, taphonomy, and coastal sediment transport. Marine Geology 124: 303-337. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 15 de noviembre de 2004
Aceptado: 26 de agosto de 2005

