










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.2 Buenos Aires abr./jun. 2006
Palinología del Holoceno del Canal del Medio, estuario de Bahía Blanca, Buenos Aires, Argentina
C. Marcela Borel1,2 y Eduardo A. Gómez3,4
1 Departamento de Geología, Universidad Nacional del Sur. San Juan 670, 8000 Bahía Blanca, Argentina.
2 Instituto Argentino de Oceanografía. Florida 4500, 8000 Bahía Blanca, Argentina. maborel@criba.edu.ar
3 Consejo Nacional de Investigaciones Científicas y Técnicas, Instituto Argentino de Oceanografía. Florida 4500, 8000 Bahía Blanca, Argentina. gmgomez@criba.edu.ar
4 Universidad Tecnológica Nacional, Facultad Regional Bahía Blanca, 11 de Abril 461, 8000 Bahía Blanca, Argentina.
Abstract. Palynology from the Holocene of Canal del Medio, Bahía Blanca estuary, Buenos Aires, Argentina. Holocene palynological samples from the Canal del Medio (38º 49' S; 62º 17' W), Bahía Blanca inner estuary, south of Buenos Aires province, contain pollen, spores, remains of algae (Dinoflagellata, Chlorophyta and Cyanophyta) and acritarchs. The pollen spectra are dominated by Chenopodiineae and Poaceae, along with Asteraceae and Ephedra. The dinophycean algae are represented by cysts of estuarine dinoflagellates of the order Gonyaulacales (Operculodinium centrocarpum (Deflandre and Cookson) Wall, Spiniferites bulloideus (Deflandre and Cookson) Sarjeant, S. sp. cf. S. pachydermus (Rossignol) Reid, S. bentorii (Rossignol) Wall and Dale and S. ramosus (Ehrenberg) Loeblich and Loeblich). The chlorophycean algae consist of cenobia of Pediastrum boryanum (Turpin) Meneghini and colonies of Botryococcus braunii Kützing. The assemblages also contain sheets of Gloeotrichia sp. The acritarchs are represented by Micrhystridium sp., Cymatiosphaera sp., Halodinium minus Bujak, Acritarch sp. F, Acritarch sp. G and Acritarch sp. H. Three palynological zones are distinguished in the sequence. The lowermost zone, dated in 3,500 14C yr B.P., represents an ancient tidal plain in close relation with a channel of turbid water and variable energy. The intermediate zone suggests better conditions for the development of autotrophic dinoflagellates, in a restricted estuarine environment. For the top zone, the low frequencies of dinocysts and acritarchs indicate similar conditions to those currently present in the inner Bahía Blanca estuary. Along the profile, the pollen reflects the development in the surrounding areas of halophytic communities, with the presence of xerophytic woodland. The characteristics of these deposits indicate that ca. 3,500 14C yr B.P., the mean sea level was located at a similar position than the current one.
Resumen. Las muestras palinológicas de sedimentos holocenos provenientes del perfil Canal del Medio (38º 49' S; 62º 17' O), estuario interno de Bahía Blanca, sur de la provincia de Buenos Aires, contienen polen, esporas, remanentes de algas (Dinoflagellata, Chlorophyta y Cyanophyta) y acritarcos. Los espectros polínicos están dominados por Chenopodiineae y Poaceae, con Asteraceae y Ephedra. Las algas dinofíceas están representadas por quistes de dinoflagelados estuáricos del orden Gonyaulacales (Operculodinium centrocarpum (Deflandre y Cookson) Wall, Spiniferites bulloideus (Deflandre y Cookson) Sarjeant, S. sp. cf. S. pachydermus (Rossignol) Reid, S. bentorii (Rossignol) Wall y Dale y S. ramosus (Ehrenberg) Loeblich y Loeblich). Las algas clorofíceas consisten en cenobios de Pediastrum boryanum (Turpin) Meneghini y colonias de Botryococcus braunii Kützing. Las asociaciones también contienen vainas de Gloeotrichia sp. Los acritarcos están representados por Micrhystridium sp., Cymatiosphaera sp., Halodinium minus Bujak, Acritarco sp. F, Acritarco sp. G y Acritarco sp. H. En la secuencia se distinguen tres zonas palinológicas. La zona inferior, datada en 3.500 14C años A.P., representa una antigua planicie de marea, en estrecha relación con un canal de aguas turbias y energía variable. La zona intermedia sugiere mejores condiciones para el desarrollo de dinoflagelados autotróficos, en un ambiente estuárico restringido. Para la zona superior, las bajas frecuencias de dinoquistes y acritarcos indican condiciones similares a las que actualmente se presentan en el estuario interno de Bahía Blanca. A lo largo del perfil, el polen refleja el desarrollo, en las áreas circundantes, de comunidades halofíticas con la presencia del bosque xerofítico. Las características de estos depósitos indican que a los ca. 3.500 14C años A.P. el nivel medio del mar se encontraba en una posición similar a la actual.
Key words. Palynology; Algae; Acritarchs; Pollen; Holocene; Buenos Aires; Argentina.
Palabras clave. Palinología; Algas; Acritarcos; Polen; Holoceno; Buenos Aires; Argentina.
Introducción
El análisis palinológico de secuencias holocenas costeras permite registrar, además del polen, remanentes de algas (cigósporas, quistes, vainas, cenobios), huevos de resistencia de invertebrados, así como también palinomorfos de afinidad biológica desconocida (acritarcos). Estos palinomorfos acuáticos, no polínicos, representan a las comunidades fito y zooplanctónicas, o bentónicas, que se desarrollaron en los cuerpos de agua, permitiendo inferir los parámetros físicos y químicos del agua y las variaciones ambientales locales (Borel et al., 2003).
Entre los remanentes de origen algal, se incluyen los quistes de dinoflagelados o dinoquistes, el grupo de palinomorfos no polínico más conocido y utilizado en las reconstrucciones paleoambientales del Cuaternario. Asociados a los dinoquistes, aparecen otros microfósiles ácido-resistentes: son los acritarcos y otros palinomorfos acuáticos. Los acritarcos representan un enigmático grupo artificial de microorganismos fósiles, unicelulares, eucariotas, de pared orgánica, morfológicamente variados, de origen polifilogenético asociados consistentemente con sedimentos ácueos, esencialmente marinos (Martin, 1993). Los acritarcos del Cuaternario son escasamente conocidos, sin embargo es altamente probable que muchos de ellos representen esporas de algas. Ante la ausencia de estudios morfológicos detallados, que permitan vincular estas probables esporas con poblaciones vivientes de algas, de una manera conservadora se los clasifica como acritarcos.
En este trabajo se analiza palinológicamente una secuencia holocena aflorante en el Canal del Medio, estuario interno de Bahía Blanca, datada en su base en ca. 3.500 14C años A.P. Además, se realiza el estudio sistemático y taxonómico de los quistes de dinoflagelados y acritarcos, utilizándose su potencial para la caracterización paleoambiental.
Características del área de estudio
El estuario de Bahía Blanca, con una superficie aproximada de 3.000 km2, está integrado por una densa red de canales de diversas dimensiones, orientados en dirección NO-SE y separados por islas y extensas planicies de marea (figura 1). El canal principal tiene una forma elongada con una longitud total de 68 km y un ancho que varía desde 200 m, en la cabecera, a 4 km en su boca. La profundidad media es de 10 m, aunque en la zona exterior se han obtenido valores del orden de los 22 m. El aporte de agua dulce lo proporciona principalmente el río Sauce Chico, en la cabecera, y el arroyo Napostá Grande, en las cercanías de Ingeniero White (Piccolo y Perillo, 1990).
El estuario se divide, sobre la base de la distribución de la salinidad y la temperatura, en dos sectores. El sector interno es de mezcla parcial, con estratificación marcada en las áreas asociadas a la desembocadura de los arroyos, pero con una fuerte tendencia a ser verticalmente homogéneo en condiciones de poca precipitación. Las salinidades varían entre 15 ‰ durante los períodos lluviosos, a 30 ‰ en los períodos secos. En el sector externo, las salinidades son similares a las de las aguas de plataforma (34 ‰), con una distribución vertical homogénea. La temperatura decrece levemente desde la cabecera a la desembocadura, con una distribución vertical homogénea a lo largo de todo el estuario (Piccolo et al., 1987). Las temperaturas oscilan entre 6,8 ºC y 9,5 ºC en invierno, y entre 21,5 ºC y 25,7 ºC durante el verano, con un patrón de ascenso y descenso suave (Marcovecchio, 2000).
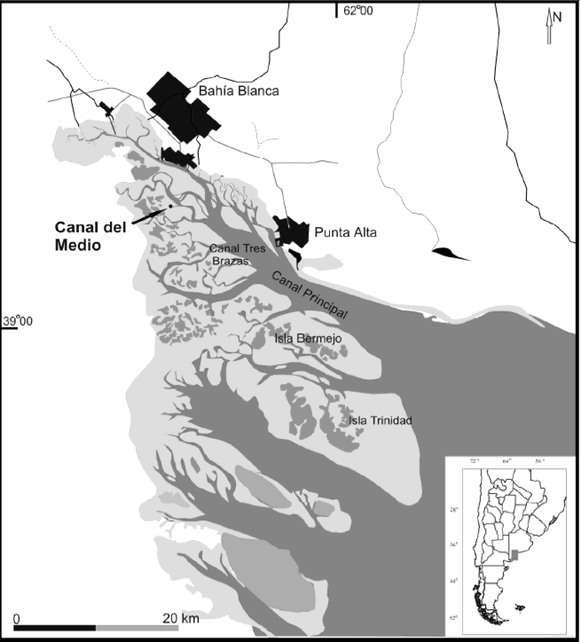
Figura 1. Mapa de ubicación / location map.
Actualmente, el estuario de Bahía Blanca tiene un régimen mesomareal semidiurno con un rango máximo de marea que aumenta desde los 2,4 m en la boca del estuario hasta más de 4 m en su cabecera. Este aumento del rango de marea es causado por la geometría del estuario, y puede incluso duplicarse por la influencia del viento (Perillo y Piccolo, 1991). Las corrientes de marea dentro de los canales del estuario son reversibles, alcanzando velocidades máximas promedio en el Canal Principal de 1,20 y 1,05 m/s, para condiciones de reflujo y flujo respectivamente (Aliotta et al., 1991).
El clima del área de Bahía Blanca es subhúmedo - seco, mesotermal, con nulo o pequeño exceso de agua y de transición hacia los climas típicamente áridos (Burgos y Vidal, 1951). La costa del estuario, hasta la cota de 10 m, está ocupada por la estepa halófita, integrada por arbustos, subarbustos y hierbas halófitas, que llegan a confundirse con las especies propias de la estepa pampeana y del bosque xerofítico (Verettoni y Aramayo, 1976). El espartillar, formado por Spartina alterniflora Loisel, poácea que crece sumergida en el agua de mar, es la vegetación de las planicies de marea en su parte media y superior. En la parte inferior no se desarrolla vegetación, en cambio está habitada por una gran variedad de organismos, entre ellos cangrejos, cirripedios, poliquetos, gasterópodos y bivalvos.
En un estudio de la composición y abundancia del fitoplancton de la parte más interna del estuario, sobre un período de cinco años, Gayoso (1988) muestra la ausencia de una estación con predominancia de dinoflagelados. Este fenómeno es atribuido a la turbulencia del agua, provocada por acción de las mareas y del viento, que permitiría solamente a las diatomeas muy silicificadas (con grandes espinas) mantenerse dentro de la capa eufótica.
La sección estudiada se encuentra expuesta en la barranca de una de las islas del Canal del Medio (38º 49' S; 62º 17' O), correspondiendo al sector erosivo de un meandro de marea de la porción interna del estuario de Bahía Blanca. Este canal exhibe profundidades medias del orden de los 2 m, no presenta descarga de agua dulce, la salinidad es de 30 ‰, las velocidades de corriente máximas, tanto durante la creciente como durante la bajante, están en el orden de los 50 cm/s y la concentración de sedimento en suspensión varía durante el ciclo de marea entre los 210 y los 65 mg/l, siendo unos 20 mg/l mayor en el fondo que en la superficie del canal (Cuadrado et al., 2000).
A lo largo de toda la sección, los sedimentos presentan una estratificación laminar con alternancia cíclica ininterrumpida de limos arcillosos y limos arenosos. Esta alternancia es indicativa de oscilaciones de la energía en el ambiente depositacional, que en este caso particular se atribuye a la acción cíclica de las mareas. Las capas arenosas se depositan durante los períodos en que las corrientes son más veloces, mientras que los materiales limo - arcillosos se depositan principalmente durante los períodos de estoa de pleamar y bajamar. Sin embargo, los materiales limo - arcillosos no pueden permanecer depositados en aquellos sectores que durante el ciclo de marea presentan altas velocidades de corrientes, como ocurre en los canales. Es así que los depósitos de este tipo de materiales sólo pueden permanecer sin ser totalmente resuspendidos en aquellos lugares que presentan una moderada a baja energía ambiental, circunstancia que dentro del ambiente estuarial sólo se da en la zona intermareal y submareal de muy poca profundidad (Cuadrado et al., 2004).
Materiales y métodos
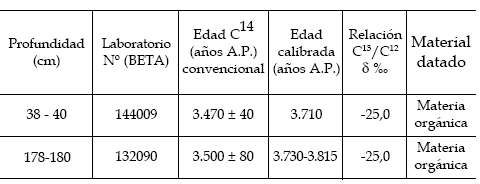
Las muestras de sedimento, obtenidas a intervalos variables entre 10 y 15 cm, provienen de una sección de 2,00 m de longitud, aflorante en la barranca erosiva del Canal del Medio. Se realizaron dos dataciones radiocarbónicas (tabla 1).
Tabla 1. Dataciones radiocarbónicas del perfil Canal del Medio / radiocarbon dates of the Canal del Medio profile.
Para la extracción físico-química de los palinomorfos se procesaron entre 2 y 20 gramos de sedimento de cada muestra, empleando las técnicas de rutina (Gray, 1965; Faegri e Iversen, 1989). Previo al tratamiento se agregaron cinco tabletas con esporas de Lycopodium (taxón foráneo al área de estudio). Se utilizó una solución de pirofosfato de sodio (Na4P2O7) para deflocular las arcillas, HCl 10% en frío para eliminar carbonatos, ZnCl2 (d = 2,2 g/ml) para separar la fracción inorgánica y HF concentrado para eliminar silicatos. No se realizó acetólisis ni se emplearon ácidos calientes para preservar los dinoquistes. Los residuos orgánicos fueron filtrados con malla de nylon de 10 µm y luego teñidos con safranina-o. Con el material retenido se realizaron 1 ó 2 preparados por muestra (designados como a, b), los cuales se encuentran archivados en la Palinoteca del Laboratorio de Palinología de la Universidad Nacional del Sur (sigla LPUNS-CM, números 1 a 24). Los ejemplares ilustrados están caracterizados por el número y letra de preparado, y las coordenadas correspondientes a la escala Vernier del microscopio Nikon Eclipse 600 del Laboratorio de Palinología de la Universidad Nacional del Sur. Las fotomicrografías fueron tomadas con una cámara digital Nikon Coolpix 950.
El agregado de esporas foráneas permitió estimar los valores representativos de la suma palinológica para cada muestra (Bianchi y D'Antoni, 1986). Para la realización de los diagramas de frecuencias relativas (figuras 2 y 3) se utilizó el Programa TILIA 2.0.b.4 (Grimm, 1991-93). El cálculo de porcentajes de cada uno de los taxa de polen fue calculado sobre la suma polínica. El cálculo de porcentajes de algas y acritarcos se realizó utilizando la suma total de palinomorfos reconocidos, incluyendo polen y esporas de plantas terrestres y acuáticas.
Figura 2. Diagrama palinológico, en porcentaje, de polen, algas y acritarcos (frecuencias de algas y acritarcos calculadas sobre la suma total de palinomorfos) del perfil Canal del Medio. Incluye dataciones radiocarbónicas (14C años A.P.) y zonas palinológicas / palynological diagram of pollen, algae and acritarchs at Canal del Medio in percentages (frequencies of algae and acritarchs over total palynomorph sum). Including radiocarbon-dated horizons (14C yr B.P.) and palynological zones.
Figura 3. Diagrama, en porcentaje, de polen (frecuencias calculadas sobre la suma polínica total) del perfil Canal del Medio. Incluye dataciones radiocarbónicas (14C años A.P.) y zonas palinológicas / diagram of pollen at Canal del Medio, in percentages (frequencies over total pollen sum). Including radiocarbon-dated horizons (14C yr B.P.) and palynological zones.
Resultados
Las asociaciones palinológicas recobradas están integradas por polen, esporas de briofitas, esporas de hongos, remanentes de algas y acritarcos. Se identificaron 25 tipos de polen, tres tipos de esporas de briofitas, cinco tipos de esporas de hongos, seis tipos de quistes de dinoflagelados, dos tipos de clorofíceas, un tipo de cianofíceas y seis tipos de acritarcos.
La asociación de quistes de dinoflagelados contiene poca diversidad y está integrada únicamente por gonyaulacoideos: Operculodinium centrocarpum (Deflandre y Cookson) Wall, Operculodinium centrocarpum (Deflandre y Cookson) Wall-de procesos cortos, Spiniferites bulloideus (Deflandre y Cookson) Sarjeant, Spiniferites sp. cf. S. pachydermus (Rossignol) Reid, Spiniferites bentorii (Rossignol) Wall y Dale y Spiniferites ramosus (Ehrenberg) Loeblich y Loeblich. En el 88 % de las muestras el taxón dominante es Operculodinium centrocarpum.
Las Chlorophyta de agua dulce están representadas por cenobios de Hydrodictyaceae (Pediastrum boryanum (Turpin) Meneghini) y agregados coloniales de Botryococcus braunii Kützing. Entre las Cyanophyta se reconocieron vainas de Rivulariaceae (Gloeotrichia sp.).
Los acritarcos están representados por Micrhystridium sp., Cymatiosphaera sp., Halodinium minus Bujak, Acritarco sp. F, Acritarco sp. G y Acritarco sp. H (ver Apéndice). La denominación de los distintos tipos de acritarcos es informal y corresponde a Borel (2003).
Los resultados de los recuentos de polen, esporas, remanentes de algas, quistes de dinoflagelados y acritarcos se presentan en diagramas de porcentajes (figuras 2 y 3). Sobre la base del Cluster Analysis se determinaron tres zonas palinológicas y dos subzonas.
Zona CM-I (190-35 cm): se reconocieron dos subzonas:
Subzona CM-Ia (190-105 cm): está caracterizada por altos valores de polen (hasta 85% sobre el total de palinomorfos). El espectro polínico está dominado por Chenopodiineae (47-79%), junto con Poaceae (9- 30%), Asteraceae (< 15%) y elementos del bosque xerofítico (<15%). Entre estos últimos las mayores proporciones corresponden a Ephedra (<10%). En proporciones menores están presentes otras hierbas y arbustos (<5%) y las plantas acuáticas (<3%). Las algas clorofíceas están representadas por Botryococcus braunii (<4%) y Pediastrum boryanum (<1%). Los dinoquistes y acritarcos representan entre 15 y 25% del total de palinomorfos. Predominan los acritarcos (Micrhystridium sp., Acritarco sp. F, Acritarco sp. H y Cymatiosphaera sp.) que varían entre 10 y 24%. Los dinoquistes, representados por Operculodinium centrocarpum, Spiniferites sp. cf. S. pachydermus, S. bulloideus y S. bentorii se registran en mínimas proporciones (<5,5%).
Subzona CM-Ib (105-35 cm): los valores de polen varían entre 60 y 90% sobre el total de palinomorfos. El polen está dominado por Chenopodiineae (45-70 %) y Poaceae (20-40%), acompañados por Asteraceae (<11%) y elementos del bosque xerofítico (<7%). En proporciones menores están presentes otras hierbas y arbustos (<5%) y las plantas acuáticas (<5%). Las algas clorofíceas están presentes en bajas proporciones: Botryococcus braunii (<3%) y Pediastrum boryanum (<1 %), presentándose también Gloeotrichia sp. (<2%). Los dinoquistes varían entre 1,5 y 18,5%. Los taxa presentes son Operculodinium centrocarpum, O. centrocar pum de procesos cortos, Spiniferites bulloideus, S. sp. cf. S. pachydermus, S. bentorii y S. ramosus. Los acritarcos varían entre 5,5 y 38%, registrándose además de los taxones mencionados Halodinium minus y Acritarco sp. G.
Zona CM-II (35 - 25 cm): el polen alcanza hasta el 75% sobre el total de palinomorfos y no presenta variaciones significativas con respecto a la zona anterior. Los acritarcos presentan frecuencias entre 6,5 y 11%, representados por Micrhystridium sp., Acritarco sp. F y Cymatiosphaera sp. Los dinoquistes alcanzan los mayores valores de toda la sección (14-22,5%).
Zona CM-III (25-0 cm): el espectro polínico no varía significativamente, estando dominado por Chenopodiineae (60-70%) y Poaceae (16-21,5%), junto con Asteraceae (<7%) y elementos del bosque xerofítico (<11%). En proporciones menores están presentes otras hierbas y arbustos (<5%) y las plantas acuáticas (1-6%). Las algas clorofíceas y cianofíceas están presentes en bajas proporciones (1,5-6,5%). Los dinoquistes presentan bajos valores (0,5-4%), mientras que los acritarcos varían entre 23 y 33,5%. Entre los acritarcos los mayores valores corresponden a Micrhystridium sp. (10-20%) y Acritarco sp. F (9-13%); también están presentes Cymatiosphaera sp. (1-3,2%) y Acritarco sp. G (1-3,6%).
Consideraciones ecológicas
La distribución actual de los dinoflagelados en los océanos depende fundamentalmente de la temperatura, la salinidad y la distancia a la costa, esta última relacionada probablemente con el suministro de nutrientes (Dale, 1976, 1996; Wall et al., 1977; Harland, 1983; Edwards y Andrle, 1992). Otros factores que controlan su crecimiento son: luz, influencia de las mareas, polución y predación por otros organismos (Taylor, 1987). A una escala global ocurren en bandas latitudinales amplias, formando asociaciones de latitudes bajas, medias y altas (Mudie y Harland, 1996).
La mayoría de las especies de dinoquistes cuaternarios pertenecen a los órdenes Peridiniales, Gymnodiniales y Gonyaulacales. Los dinoflagelados del orden Gonyaulacales son favorecidos por aguas estratificadas superficialmente templadas, en contraste con los peridinioideos que están asociados con áreas de upwelling donde las aguas son más frías y ricas en nutrientes.
Operculodinium centrocarpum tiene una distribución cosmopolita, desde ambientes costeros a oceánicos profundos y una gran tolerancia a un amplio rango de temperatura y salinidad, encontrándoselo en climas árticos a tropicales (Edwards y Andrle, 1992; Marret y Scourse, 2002). En varios estudios (Harland, 1973; Mudie, 1980; De Vernal et al., 1989, 1992; Matthiessen, 1995; Rochon et al., 1999; Ellegaard, 2000) se describen morfotipos con procesos cortos que han sido relacionados a ambientes de baja salinidad. Los morfotipos con procesos de poca longitud y además gruesos, serían característicos de salinidades bajas en ambientes restringidos (Ellegaard, 2000).
De acuerdo a Marret y Scourse (2002), Spiniferites ramosus y Operculodinium centrocarpum, son abundantes en los sedimentos de las áreas hidrográficamente estratificadas, sugiriendo que condiciones de aguas mezcladas serían menos favorables para estas especies. En cambio, las asociaciones de sedimentos intermareales provenientes de ambientes de aguas mezcladas (región 6 de Reid, 1975) están dominadas por especies de Spiniferites, en particular S. bentorii y S. bulloideus con Lingulodinium machaerophorum (Deflandre y Cookson) Wall. Wall et al. (1977) indican centros de distribución de S. bulloideus en estuarios no estratificados. Edwards y Andrle (1992) sostienen que S. bulloideus se encuentra en asociaciones estuarinas, neríticas y océanicas pero con una tolerancia climática de templado fría a tropical. Según Rochon et al. (1999), la producción de algunas especies de dinoquistes tales como Spiniferites bulloideus y S. ramosus está principalmente limitada por la intensidad de la luz.
En los sedimentos actuales del Canal Tres Brazas, porción interna del estuario de Bahía Blanca, se registra para la zona intermareal a submareal somera una asociación de O. centrocarpum, Spiniferites membranaceus y Spiniferites sp., no superando los dinoquistes proporciones del 5% (Grill y Guerstein, 1995).
En cuanto a los acritarcos, si bien se desconoce su afinidad biológica, pueden inferirse sus preferencias ecológicas a partir de su asociación con otros microfósiles. Micrhystridium y Cymatiosphaera fueron registrados en las muestras de los sedimentos actuales, inter a submareales, del Canal Tres Brazas, estuario interno de Bahía Blanca (Grill y Guerstein, 1995). En ese mismo trabajo, las proporciones de los acritarcos, sobre el total de palinomorfos, varían entre 5,5 y 9%. Los Acritarcos sp. F también están presentes en los sedimentos de las planicies de marea actuales del estuario interno de Bahía Blanca, asociados con dinoquistes estuáricos (revisión realizada por C.M.B. sobre las muestras de Grill y Guerstein, op. cit.). Posiblemente, estos acritarcos representan esporas de algas que habitan en ambientes mareales restringidos. Cabe destacar que, algunas especies de Pyramimonas, algas de agua dulce o ambientes mareales, forman esporas bénticas pequeñas con una morfología similar a estos acritarcos (forma esferoidal y con pared gruesa ornamentada o no). Estas algas prasinofíceas flageladas forman parte del fitoplancton actual del estuario de Bahía Blanca (Gayoso, 1988). Algunos autores (Mudie, 1992; Head, 1993a) otorgan al morfogé nero Cymatiosphaera una afinidad con las Prasinophyceae.
Todos los registros de Halodinium provienen de depósitos marinos o estuáricos, sugiriéndose que su presencia indica una influencia fluvial fuerte y asociaciones con ambientes transicionales (De Vernal et al., 1989, Mudie, 1992; Head, 1993a). Su afinidad biológica es incierta, sin embargo De Vernal et al. (1989) sugieren una afinidad con los rizópodos.
Discusión
Para todo el perfil, el polen evidencia el desarrollo, en las áreas circundantes, de comunidades halófitas, con la presencia del bosque xerofítico. El polen de plantas acuáticas y las algas clorofíceas y cianofíceas responden a la presencia en el área de cuerpos continentales de agua dulce a salobres.
A través de toda la secuencia, las asociaciones de dinoquistes tienen baja diversidad, similar composición y son comparables a las asociaciones de los sedimentos actuales del Canal Tres Brazas (Grill y Guerstein, 1995). No se registraron quistes de dinoflagelados heterotróficos, sino solamente especies autotróficas, tolerantes a rangos amplios de temperatura y salinidad.
La Subzona CM-Ia, datada en 3.500 ± 80 14C años A.P., presenta proporciones de dinoquistes, acritarcos y polen similares a las muestras actuales (Grill y Guerstein, 1995), las que exhiben valores muy altos de polen (hasta 85%) y valores bajos de dinoquistes y acritarcos. Esta subzona representaría un ambiente de planicies de mareas a submareal somero, con una alta turbidez, originada por la importante energía de las corrientes de marea.
La Subzona CM-Ib, datada en 3.470 ± 40 14C años A.P. en su parte superior, presenta proporciones de dinoquistes mayores que la subzona CM-Ia. Sin embargo, esas frecuencias son muy fluctuantes, sugiriendo la alternancia de períodos más y menos favorables para la productividad de los dinoflagelados autotróficos. Los períodos más favorables para la proliferación de estos organismos se vincularían con aguas más claras, con mayor penetración de la luz.
En la Zona CM-II, el espectro palinológico presenta las mayores proporciones de dinoquistes reflejando condiciones más favorables para el desarrollo de estos organismos, posiblemente de menor turbidez, aunque se habrían mantenido las condiciones estuáricas restringidas.
Para la Zona CM-III, las bajas frecuencias de dinoquistes y acritarcos indican condiciones de alta turbulencia y de salinidad variable, similares a las que actualmente se desarrollan en las franjas mareales y submareales de la porción interna del estuario de Bahía Blanca (Grill y Guerstein, 1995).
Mediante las edades radiocarbónicas de la secuencia y con la curva de calibración dada por Stuiver et al. (1998), es posible obtener las edades calibradas correspondientes. Así, sin tener en cuenta el desvío estándar de las dataciones por razones de simplicidad, se obtiene que las edades 14C convencionales de la Zona CM-I, al ser calibradas, corresponderían a 3.730 - 3.815 14C años cal. A.P. y 3.710 14C años cal. A.P. respectivamente (tabla 1). Además, si bien no se cuenta con dataciones para las Zonas CM-II y CM-III, la ausencia de cambios texturales y palinológicos marcados, a lo largo de la sección estudiada, indica que los sedimentos que las componen se depositaron bajo condiciones ambientales similares y durante un intervalo de tiempo relativamente corto que se centraría en los ca. 3.500 14C años A.P.
Según Isla (1989, 1998), Parker y Violante (1982), Cavallotto et al. (1995, 2004) y Violante y Parker (2004), después del máximo transgresivo del Holoceno, el nivel del mar habría sufrido una caída prácticamente gradual hasta alcanzar el nivel del mar actual. Sin embargo, los datos palinológicos y sedimentológicos de este estudio permiten afirmar que a los ca. 3.500 14C años A.P. el nivel medio del mar se situaba en una posición similar a la actual. Si bien no resulta adecuado comparar localidades que presentan diferencias geológicas y ambientales, tal afirmación particularmente contradice la curva de variación relativa del nivel medio del mar dada por Cavallotto et al. (2004) para el Río de la Plata. En esa curva se indica que, para los ca. 3.500 años A.P., el nivel del mar habría estado unos 5 m por encima del nivel actual. Sin embargo entre los 4.130 ± 90 años A.P. y los 2.990 ± 40 años A.P., tal curva fue construida sobre la base de dos dataciones con edades 14C de 3.530 ± 35 años A.P. y 3.470 ± 233 años A.P. obtenidas sobre conchillas y hueso respectivamente, por lo que su grado de representatividad en lo referente a la ubicación relativa del nivel medio del mar es más incierto que aquel dado por depósitos intermareales de fango como los de la sección aquí estudiada.
La altura relativa del nivel del mar representada por la sección aquí analizada no concuerda con aquella determinada por Farinati (1985) sobre Tagelus plebeius Lightfoot en posición de vida, con una edad 14C de 3.373±205 años A.P. en la desembocadura del Arroyo Napostá Grande de Bahía Blanca. Asimismo, la secuencia 4 estudiada por Grill y Quattrocchio (1996), que se ubica en el mismo sector pero a una altura de unos 5 m sobre el nivel medio del mar actual, contiene conchillas articuladas de Tagelus plebeius datadas en 3.560±100 14C años A.P. Para esa secuencia 4, se registra una alta frecuencia de dinoquistes, seguida por una disminución en los valores del microplancton indicando el final de la influencia marina en el perfil. Se debe destacar que estas dataciones posiblemente presenten un importante y variable, aunque desconocido, envejecimiento aparente o efecto de reservorio (ER) propio de los organismos que obtienen el carbono del agua de mar. El carbono presente en los fangos orgánicos de depositación intermareal es de origen principalmente atmosférico debido a que los vegetales, como Spartina, y las diatomeas bentónicas que viven allí, debido al alto grado de turbidez del agua, fotosintetizan principalmente al quedar al descubierto durante la bajamar. Por lo tanto, es probable que los sedimentos analizados en este estudio posean un escaso o incluso nulo ER. En Argentina, el valor del ER Holoceno sólo ha sido estimado en el Canal de Beagle, Tierra del Fuego, en 630 años por Albero et al. (1987) y en 540 años por Rabassa et al. (1986, 1992). Si bien es desconocido el valor de este envejecimiento aparente para el Holoceno en el resto de las costas argentinas, se estima que el mismo podría llegar a alcanzar un máximo del orden de los 600 años de acuerdo al contenido de 14C determinado por Broecker y Olson (1961) para la corriente de Malvinas. Sin embargo, la comparación del contenido de 14C de muestras modernas en equilibrio con reservorios marinos y continentales (Cordero et al., 2003), indica que las diferencias en las edades 14C cubren en realidad un rango que va desde los 80 años hasta los 1.120 años para diversas localidades de las costas patagónicas. Tal disparidad de valores es atribuida al grado de influencia de factores locales como los aportes de aguas continentales, la presencia de carbonatos fósiles en la cuenca, la circulación oceánica, la profundidad de la columna de agua y el grado de mezcla del agua con el CO2 atmosférico. Si bien tales valores no permiten evaluar el ER en el pasado, ellos evidencian un muy alto grado de incertidumbre sobre la representatividad de aquellas edades 14C obtenidas en organismos que obtienen su carbono del agua de mar.
Cabe mencionar que a profundidades similares a las de la sección aquí analizada (sector intermareal), dataciones efectuadas sobre los flancos erosivos de otros canales de marea arrojaron edades más jóvenes. En una barranca del canal Tres Brazas, las dataciones radiocarbónicas efectuadas sobre conchillas de Heleobia australis D'Orbigny (localizadas en el nivel de bajamar), y sobre Tagelus plebeius (cercanos al nivel de tormenta), dieron edades de 1.560 y 1.160 14C años A.P. respectivamente (Aliotta et al., 2000). En el perfil ubicado en el margen de erosión emergido de un canal tributario del Canal Tres Brazas (38° 07' S; 62° 45' O), conchillas de Heleobia australis, provenientes de un nivel ubicado a la altura del nivel medio del mar actual, arrojan una edad de 1.020±30 14C años A.P. (Martínez, 2002). Si bien estas últimas dataciones fueron realizadas sobre organismos que obtienen el carbono principalmente del medio acuático, y por lo tanto es posible que presenten un importante ER, las mismas sugieren fuertemente que el nivel medio del mar se habría mantenido en una posición similar a la actual a los ca. 1.000 a 1.500 14C años A.P.
Sin embargo, estas determinaciones no significan necesariamente que desde hace aproximadamente unos 3.400 años A.P el nivel medio del mar se haya mantenido en forma constante con una altura similar a la actual hasta el presente. Gómez et al. (2005b) presentan evidencias sedimentológicas y micropaleontológicas sobre la ocurrencia de oscilaciones del nivel medio del mar por debajo del actual, las que habrían ocurrido con magnitud dispar alrededor de las edades 14C de 6.300 años A.P., 4.200 años A.P. y 2.400 años A.P. y que se relacionarían con importantes cambios climáticos de escala global. De acuerdo a Gómez (2004) y Gómez et al. (2005a), tales oscilaciones tendrían su origen en valores extremos mínimos y en la gran variabilidad de la irradiancia solar que ocurren con una periodicidad de 2.300 años. Por cuanto, la sección aquí analizada representaría entonces la altura que habría alcanzado el nivel medio del mar entre dos oscilaciones negativas durante una fase en que la irradiancia solar presentaba valores relativamente altos y una baja variabilidad.
Conclusiones
Las características sedimentológicas y los datos palinológicos indican que en el Canal del Medio a los ca. 3.500 14C años A.P. (Zona CM-I) se desarrolló un ambiente intermareal a submareal somero, con alta turbidez, relacionada a una importante energía de las corrientes de marea. Esta sección de sedimentos limo- arcillo-arenosos se habría depositado con un nivel del mar similar al actual, durante el hemiciclo regresivo de la última ingresión marina. Este estudio permite afirmar que a los ca. 3.500 14C años A.P. el nivel medio del mar se situaba en una posición similar a la actual.
Los datos palinológicos y sedimentológicos de las Zonas CM-II y CM-III evidencian que se mantuvieron, durante su depositación, las condiciones de un ambiente mareal a submareal, en una porción interna restringida del estuario de Bahía Blanca.
Apéndice
Palinología sistemática
Division DINOFLAGELLATA (Bütschli)
Fensome et al. 1993
Clase DINOPHYCEAE Pascher 1914
Orden GONYAULACALES Taylor 1980
Suborden GONYAULACINAEAE (autonym)
Familia GONYAULACACEAE Lindemann 1928
Subfamilia CRIBOPERIDINIOIDEAE Fensome et al. 1993
Género Operculodinium (Wall 1967) Matsuoka et al. 1997
Especie tipo. Operculodinium centrocarpum (Deflandre y Cookson) sensu Wall 1967.
Operculodinium centrocarpum (Deflandre y Cookson) sensu Wall 1967 Figuras 4.M-O
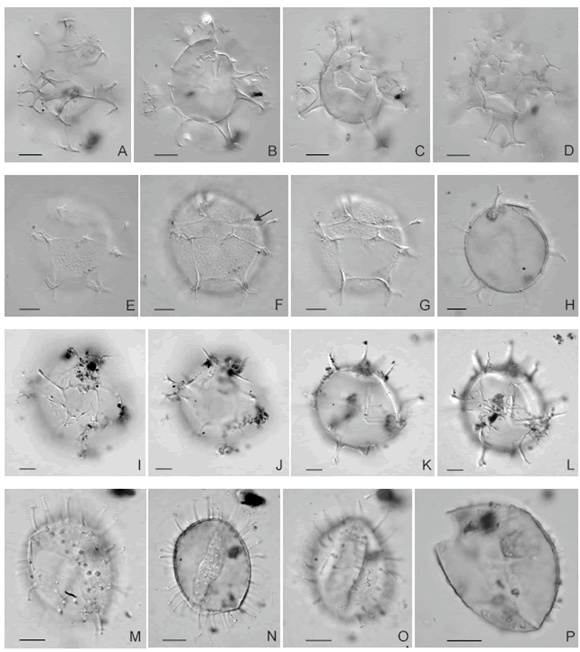
Figura 4. A-D, Spiniferites bulloideus (Deflandre y Cookson) Sarjeant, LPUNS CM 15b: 50,5/106, mostrando sucesivamente focos más bajos desde la superficie superior hacia la superficie inferior / showing successively lower foci from upper through lower surface. E-H, Spiniferites sp. cf. S. pachydermus (Rossignol) Reid, LPUNS CM 03a: 44,2/93,7, mostrando sucesivamente focos más bajos desde la superficie superior hacia el foco intemedio / showing successively lower foci from upper through intermediate focus. F, la flecha señala la ubicación del arqueopilo / arrow shows position archeopyle. I-L, Spiniferites bentorii (Rossignol) Wall y Dale, LPUNS CM 17b: 61,6/99,5, superficie ventral mostrando sucesivamente focos más bajos desde la superficie superior hacia la superficie inferior / ventral surface, showing successively lower foci from upper through lower surface. M-O, Operculodinium centrocarpum (Deflandre y Cookson) sensu Wall, LPUNS CM 13b: 46,7/103, M, foco alto / high focus, N, foco intermedio / intermediate focus, O, foco bajo / low focus. P, Operculodinium centrocarpum (Deflandre y Cookson) sensu Wall - de procesos cortos/ short processes, LPUNS CM 13b: 44,9/107,3, vista general / general view. Todas las fotomicrografías fueron tomadas con contraste de interferencia / All photomicrographs were taken using interference contrast. Escala gráfica / scale bar = 10 µm.
Principal material estudiado. LPUNS-CM 15b: 28,4/100; 15b: 33/110; 13b: 51,2/99,9; 13b: 46,7/103.
Dimensiones. Diámetro del cuerpo central: 35 - 37 x 39 - 41 µm; longitud de los procesos: 7-11 µm (6 ejemplares medidos).
Observaciones. La pared es microgranular; los procesos son delgados, de terminaciones bífidas.
Nombre de la teca. Es el quiste de Protoceratium reticulatum (Claparéde y Lachmann) Bütschli.
Comentarios. En la descripción original Operculodinium centrocarpum (como Hystrichosphaeridium centrocarpum) es una especie del Mioceno con procesos de 13 - 18 µm de longitud, sólidos, mostrando fibrillas radiales en el punto de inserción y ápices ensanchados con pequeñas espinas curvadas. Wall (1967) describió especímenes del Pleistoceno del Mar Caribe que asignó a la especie centrocarpum y transfirió al género Operculodinium. Wall y Dale (1968) incubaron exitosamente quistes actuales que dentificaron como idénticos a los dinoquistes pleistocenos que Wall (1967) había referido a O. centrocarpum. Las tecas móviles incubadas fueron identificadas como Protoceratium reticulatum. Head (1996) y Matsuoka et al. (1997) sostienen que los dinoquistes cuaternarios que son atribuidos a O. centrocarpum en la literatura son más pequeños, están menos ornamentados y tienen procesos menos robustos que el material original del Mioceno. Por eso, proponen referir a los especímenes cuaternarios como quistes de la especie móvil Protoceratium reticulatum. Además, los quistes de Protoceratium reticulatum muestran una gran variación en la morfología de sus procesos (Head, 1993b).
Operculodinium centrocarpum (Deflandre y Cookson) sensu Wall 1967 -de procesos cortos Figura 4.P
Principal material estudiado. LPUNS-CM 13b: 46/106,7; 13b: 44/107,3.
Dimensiones. Diámetro del cuerpo central: 36 x 40 µm; longitud de los procesos: 2 - 3 µm (2 ejemplares medidos).
Observaciones. Los procesos son de reducida longitud y gruesos. En el mismo especimen, algunos son truncados y otros acuminados.
Nombre de la teca. Los estudios de desenquistamiento realizados por Ellegaard et al. (1994) en Dinamarca, confirmaron la afinidad de quistes de procesos reducidos con Protoceratium reticulatum.
Otros registros. Quistes con procesos cortos y gruesos han sido denominados O. centrocarpum var. truncatum (Mudie, 1980) y como P. reticulatum con procesos anchos de longitud reducida por Matthiessen (1995).
Subfamilia INCIERTA
Género Spiniferites Mantel, emend. Sarjeant 1970
Especie tipo. Spiniferites ramosus (Ehrenberg) Loeblich y Loeblich 1966.
Spiniferites bulloideus (Deflandre y Cookson) Sarjeant 1970 Figuras 4.A-D
Principal material estudiado. LPUNS-CM 15b: 50,5/106; 15b: 3,1/105; 13d: 57,6/109.
Dimensiones. Diámetro del cuerpo central: 30-36 x 29-31 µm; longitud procesos antapicales: 14-16 µm; longitud de los otros procesos: 11-13 µm. Altura de las crestas uniendo los procesos: 3-4 µm. Espesor de la pared menor de 1 µm. Ancho del cíngulo: 8 µm (5 ejemplares medidos).
Observaciones. En los especímenes recobrados, generalmente, la forma del cuerpo central es levemente ovoidal. Los procesos son sólo gonales; huecos, cerrados distalmente y trifurcados con terminaciones bífidas. No se observan procesos intergonales. Los procesos antapicales son más anchos y de mayor longitud que los restantes procesos.
Comparaciones. Los ejemplares de Canal del Medio son muy semejantes a los ilustrados por Reid, 1974 (lám. 2, figs 17-19).
Nombre de la teca. Acorde a Wall y Dale (1968) es un quiste de Gonyaulax scrippsae Kofoid.
Spiniferites sp. cf. S. pachydermus (Rossignol) Reid, 1974 Figuras 4.E-H
Principal material estudiado. LPUNS-CM 03a: 44,2/93,7; 15b: 31,6/111; 05b: 40/104,4.
Descripción. Quiste escolocorado, de cuerpo central ovoidal con ápices redondeados. Los procesos son trifurcados, unidos por una membrana baja. En el endofragma se observan columelas o fibras en posición radial, el perifragma es más delgado y granular. Las membranas que conectan los procesos son claramente granulares.
Dimensiones. Diámetro del cuerpo central 39 - 45 µm x 34 - 41 µm; pared: endofragma de 1,5 µm de espesor y perifragma más delgado, de 1 µm de espesor; longitud de los procesos: 11 - 13 µm (3 ejemplares medidos).
Observaciones. Los ejemplares muestran el tipo de pared típica de S. pachydermus, sin embargo las dimensiones son menores y los procesos más cortos y delgados. Además en la descripción de Reid (1974) no se indica que las membranas suturales sean granulares.
Spiniferites bentorii (Rossignol) Wall y Dale 1970 Figuras 4.I-L
Principal material estudiado. LPUNS-CM 02b: 31/109; 17b: 61,6/99,5.
Dimensiones. Diámetro del cuerpo central 50 - 55 µm x 44 - 48 µm; pared de 1,5 µm de espesor; longitud de los procesos: 10 - 15 µm (3 ejemplares medidos).
Grupo ACRITARCHA Evitt, 1963
Género HALODINIUM Bujak 1984
Especie tipo. Halodinium major Bujak 1984 Halodinium minus Bujak 1984 Figuras 5.A-C
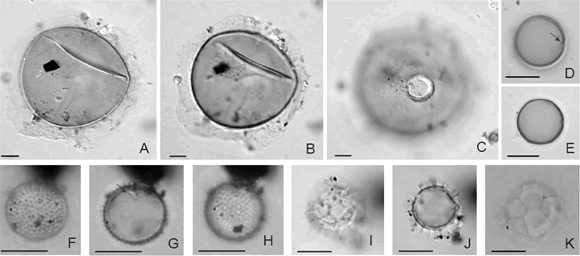
Figura 5. A-C, Halodinium minus Bujak, LPUNS CM 11b: 61,6/99,5; A, foco alto / high focus; B, foco intermedio / intermediate focus; C, foco bajo mostrando piloma con opérculo in situ / low focus showing pylome with operculum in situ. D-E, Acritarco / Acritarch sp. F, LPUNS CM 15b: 29,7/110, D, foco alto; la flecha señala una pequeña apertura / high focus; the arrow shows a small slit; E, foco intermedio / intermediate focus. F-H, Micrhystridium sp. F, LPUNS CM 07b: 20,5/94,8, F, foco alto / high focus; G, foco intermedio; la flecha señala una pequeña apertura / intermediate focus; the arrow shows a small slit. H, foco bajo / low focus. I-K, Cymatiosphaera sp., I-J, LPUNS CM 22a: 31,7/111,3, I, foco alto / high focus; J, foco intermedio / intermediate focus; K, LPUNS CM 23a: 23,4/118,2, foco alto / high focus. Todas las fotomicrografías fueron tomadas con contraste de interferencia / all photomicrographs were taken using interference contrast. Escala gráfica / scale bar = 10 µm.
Principal material estudiado. LPUNS-CM 11b: 61,6/99,5.
Dimensiones. Diámetro del cuerpo central (excluyendo la membrana): 56 x 63 µm; altura máxima de la membrana: 11 - 12 µm; diámetro piloma: 10 -13 µm (3 ejemplares medidos)
Observaciones. El piloma central es subcircular y está rodeado con un anulus engrosado bien definido. El opérculo es discernible en uno de los ejemplares.
Otros registros. Halodinium minus ha sido registrado en sedimentos recientes y del Holoceno inferior en la Bahía de Hudson y en el estuario del río St. Lawrence (Mudie y Harland, 1996). Halodinium spp. fue registrado en sedimentos actuales de estuarios del Mar de Beaufort y de las Bahías de Baffin.
Género Micrhystridiun (Deflandre) emend. Sarjeant y Stancliffe 1994
Especie tipo. Micrhystridium inconspicuum (Deflandre) Deflandre emend. Deflandre y Sarjeant 1970.
Micrhystridium sp. Figuras 5.F-H
Principal material estudiado. LPUNS-CM 15b: 29/96; 15b: 21,6/94,5; 01a: 36,1/96,6.
Dimensiones. Diámetro: 9-19 µm; pared: 0,5-1 µm de espesor; espinas: 1 - 2 µm de longitud (15 ejemplares medidos).
Observaciones. En algunos especímenes se observan pequeñas hendiduras en forma de rasgaduras rectas. Género CYMATIOSPHAERA (Wetzel) Deflandre 1954
Especie tipo. Cymatiosphaera radiata (Wetzel) emend. Sarjeant 1985.
Cymatiosphaera sp. Figuras 5.I-K
Principal material estudiado. LPUNS-CM 05b: 49,3/95,9; 13b: 48,2/105,8; 15b: 17/94,5; 04a: 53,9/110,1.
Dimensiones. Diámetro 12 - 16 µm de diámetro, lúmenes 3 - 6 µm de diámetro; crestas 2 - 3 µm de altura; pared 1 µm de espesor (9 ejemplares medidos).
Acritarco sp. F Figuras 5.D-E
Principal material estudiado. LPUNS-CM 15b: 29,7/110; 15b: 29,7/110; 04a: 57/99,7.
Descripción. Palinomorfos esferoidales, de pared gruesa y psilada. Presentan una ruptura en forma de rasgadura desarrollada variablemente; y en algunos ejemplares de forma sigmoide.
Dimensiones. Diámetro 12 - 24 µm; espesor de la pared 1 - 1,5 µm (20 ejemplares medidos).
Acritarco sp. G
Principal material estudiado. LPUNS-CM 01a: 57,8/100,1; 01a: 13,7/121,3.
Descripción. Palinomorfos esferoidales, de pared gruesa, escabrada a verrucada. En algunos especímenes se observan rasgaduras simples muy poco desarrolladas.
Dimensiones. Diámetro 15 - 23 µm; espesor de la pared 1 µm (5 ejemplares medidos).
Acritarco sp. H
Principal material estudiado. LPUNS-CM 24a: 45,6/110,8; 24a: 40,5/98,4; 24a: 95,5/23,1; 17a: 16,5/104,2.
Descripción. Palinomorfos elipsoidales, de pared gruesa y escabrada, con una ruptura en forma de rasgadura recta y bien desarrollada que abre en dos mitades el cuerpo central.
Dimensiones. Diámetro 22 - 29 µm; espesor de la pared 1 - 1,5 µm (7 ejemplares medidos).
Agradecimientos
Los autores desean agradecer a G. Daners y a un revisor anónimo, quienes con sus correcciones y sugerencias enriquecieron este trabajo. C.M.B agradece especialmente a G.R. Guerstein por su constante apoyo, tanto en el trabajo de campo como en el estudio de las muestras y a A.R. Prieto quien contribuyó con sus va- liosos aportes en las distintas etapas de este trabajo. Este trabajo constituye una contribución a los proyectos financiados por la Agencia de Promoción Científica y Tecnológica (BID 1201/OC-AR PICT 07-09659; BID 1201/OC-AR PICT 07-14652 y BID 1201/OCAR PICT 07-14653).
Bibliografía
Albero, M.C., Angiolini, F. y Piana, L. 1987. Holocene 14C reservoir effect at Beagle Channel (Tierra del Fuego, Argentina Republic). Quaternary of South America and Antarctic Peninsula 5: 59-71.
Aliotta, S., Lizasoain, G.O. y Lizasoain, W.O. 1991. Sedimentología y paleoambientes cuaternarios en el sector interno del estuario de Bahía Blanca, Argentina. Anales del Instituto de Ciencias del Mar y Limnología de la Universidad Autónoma de México 18: 99-107.
Aliotta, S., Lizasoain, G. y Ginsberg, S. 2000. Diferenciación geofísica del fondo marino y su relación geológica, canal Tres Brazas, estuario de Bahía Blanca (Argentina). 2° Congreso Latinoamericano de Sedimentología y 8° Reunión Argentina de Sedimentología (Mar del Plata), Resúmenes: 27-28.
Bianchi, M. y D'Antoni, H. 1986. Depositación del polen actual en los alrededores de Sierra de los Padres (Provincia de Buenos Aires). 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas: 16-27.
Borel, C.M. 2003. [Palinología (con énfasis en algas y acritarcos) del Holoceno del sud-sudeste bonaerense. Tesis Doctoral, Universidad Nacional del Sur, Bahía Blanca, 113 pp. Inédito.].
Borel, C.M., Guerstein, G.R. y Prieto, A.R. 2003. Palinomorfos acuáticos (algas y acritarcos) del Holoceno de la laguna Hinojales (Buenos Aires, Argentina): interpretación paleoecológica. Ameghiniana 40: 531-544.
Broecker, W.S. y Olson, E.A. 1961. Lamont radiocarbon measurements VIII, Radiocarbon 3: 176-204.
Bujak, J.P. 1984. Cenozoic dinoflagellate cysts and acritarchs from the Bering Sea and northern North Pacific, D.S.D.P. Leg 19. Micropaleontology 30: 180-212.
Burgos, J. y Vidal, A. 1951. Los climas de la República Argentina, según la nueva clasificación de Thornthwaite. Meteoros 1: 3-32.
Cavallotto, J.L., Parker, G. y Violante, R.A. 1995. Relative sea level changes in the Río de la Plata during the Holocene. 2° Annual Meeting of International Geoscience Programme 367: Late Quaternary coastal records of rapid change: application to present and future conditions (Antofagasta), Abstracts: 19-20.
Cavallotto, J.L., Violante, R.A. y Parker, G. 2004. Sea-level fluctuations during the last 8600 years in the de la Plata river (Argentina). Quaternary International 114: 155-165.
Cordero, R.R, Panarello, H., Lanzelotti, S. y Favier Dubois, C.M. 2003. Radiocarbon Age Offsets between Living Organisms from the Marine and Continental Reservoir in Coastal Localities of Patagonia (Argentina). Radiocarbon 45: 9-15.
Cuadrado, D.G., Ginsberg, S.S y Gómez, E.A. 2000. Características físicas de un canal interior en el Estuario de Bahía Blanca (Buenos Aires). En: 3° Jornadas Nacionales de Geografía Física, Santa Fé, Actas: 113-120.
Cuadrado, D.G., Ginsberg, S.S y Gómez, E.A. 2004. Geomorfología. En: M.C. Píccolo y M.S. Hoffmeyer (eds.), Ecosistema del Estuario de Bahía Blanca, Instituto Argentino de Oceanografía, Bahía Blanca, pp. 29-38.
Dale, B. 1976. Cyst formation, sedimentation and preservation: factors affecting dinoflagellate assemblages in recent sediments from Trondheimsfjord, Norway. Review of Palaeobotany and Palynology 22: 39-60.
Dale, B. 1996. Dinoflagellate cysts ecology: modeling and geological applications. En: J. Jansonius y D.C. McGregor (eds.), Palynology: principles and applications, American Association of Stratigraphic Palynologists Foundation, pp. 1249-1275.
Deflandre, G. 1954. Systématique des Hystrichosphaeridés: sur l`acception du genre Cymatiosphaera O Wetzel. Compte Rendu Sommaire des Séances de la Société Géologique de France 12: 257-258.
Deflandre, G. y Sarjeant, W.A.S. 1970. Nouvel examen de quelques holotypes de Dinoflagellés fósiles et d`Acritarches. Cahiers de Micropaléontologie 2: 1-10.
De Vernal, A., Goyette, C. y Rodrigues, C.G. 1989. Contribution palynostratigraphique (dinoquistes, pollen et spores) á la mer de Champlain: coupe de Saint-Césaire Quebec. Canadian Journal of Earth Sciences 26: 2450-2464.
De Vernal, A., Londeix, L., Mudie, P.J., Harland, R., Morzadec- Kerfourn, M.T., Turon, J.l. y Wrenn, J.H. 1992. Quaternary organic- walled dinoflagellate cysts of the North Atlantic ocean and adjacent seas: ecostratigraphy and biostratigraphy. En: M.J. Head y J.H. Wrenn (eds.), Neogene and Quaternary dinoflagellate cysts and acritarchs, American Association of Stratigraphic Palynologists Foundation, pp. 289-328.
Edwards, L.E. y Andrle, A.S. 1992. Distribution of selected dinoflagellate cysts in modern marine sediments. En: M.J. Head y J.H. Wrenn (eds.), Neogene and Quaternary dinoflagellate cysts and acritarchs, American Association of Stratigraphic Palynologists Foundation, 69-87 pp.
Ellegaard, M. 2000. Variations in dinoflagellate cyst morphology under conditions of changing salinity during the last 2000 years in the Limfjord, Denmark. Review of Palaeobotany and Palynology 109: 65-81.
Ellegaard, M., Christensen, N.F. y Moestrup, O. 1994. Dinoflagellate cysts from Recent Danish marine sediments. European Journal of Phycology 29: 183-194.
Evitt, W.R. 1963. A discussion and proposals concerning fossil dinoflagellates, hystrichospheres, and acritarchs. National Academy of Sciences, Washington, Proceedings 49: 158-164.
Faegri, K. e Iversen, J. 1989. Textbook of Pollen Analysis. John Wiley and Sons, London, 328 pp.
Farinati, E.A. 1985. Radiocarbon dating of Holocene marine deposits, Bahía Blanca area, Buenos Aires Province, Argentina. Quaternary of South America and Antarctic Peninsula, 3: 197-206.
Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S., Wharton, D.I. y Williams, G.L. 1993. A classification of fossil and living dinoflagellates. Micropaleontology Press Special Paper 7: 351 pp.
Gayoso, A.M. 1988. Variación estacional del fitoplancton del estuario de Bahía Blanca. Gayana Botánica 45: 241-247.
Gómez, E.A., 2004. Sea Level Oscillations Related to Past Global Climatic Changes. Inter-American Institute for Global Change Research News Letter 36: 11-13.
Gómez, E.A., Martínez, D.E. y Cusminsky, G.C. 2005a. A cycle in solar irradiance as trigger of abrupt Holocene global climatic changes and negative sea level oscillations. 3° Joint Meeting of International Geoscience Programme 490: Holocene environmental catastrophes in South America: from the lowlands to the Andes (Córdoba), Abstracts: 25-26.
Gómez, E.A., Martínez, D.E., Borel, C.M., Guerstein, G.R. y Cusminsky, G.C. 2005b. Submarine evidence of Holocene sea-level fluctuations in the Bahía Blanca Estuary, Argentina. Journal of South American Earth Sciences.
Gray, J. 1965. Palynological techniques. En K. Bernard y D. Raup (eds.), Handbook of Paleontological techniques, W.H. Freeman, San Francisco, pp. 470-587.
Grimm, E. 1991-93. Tilia Software. Illinois State Museum. Research and Collection Center. Springfield, Illinois.
Grill, S. y Guerstein, G.R. 1995. Estudio palinológico de sedimentos superficiales en el estuario de Bahía Blanca, provincia de Buenos Aires, Argentina. Polen 7: 41-49.
Grill, S.C. y Quattrocchio, M.E. 1996. Fluctuaciones eustáticas durante el Holoceno a partir del registro de paleomicroplancton; arroyo Napostá Grande, sur de la provincia de Buenos Aires. Ameghiniana 33: 435-442.
Harland, R. 1973. Quaternary (Flandrian) dinoflagellate cysts from the Grand Banks of Newfoundland, Canada. Review of Palaeobotany and Palynology 16: 229-242.
Harland, R. 1983. Distribution maps of recent dinoflagellate cysts in bottom sediments from the North Atlantic Ocean and adjacent seas. Palaeontology 26: 321-387.
Head, M. J. 1993a. Dinoflagellates, sporomorphs, and other palynomorphs from the Upper Pliocene St. Erth Beds of Cornwall, Southwestern England. Journal of Paleontology 67: 62 pp.
Head, M. J. 1993b. A forum on Neogene and Quaternary dinoflagellate cysts. Palynology 17: 201-239.
Head, M.J. 1996. Modern dinoflagellate cysts and their biological affinities. En: J. Jansonius y D.C. McGregor (eds.), Palynology: principles and applications, American Association of Stratigraphic Palynologists Foundation 3: 1197-1248.
Isla, F.I. 1989. Holocene sea-level fluctuation in the Southern Hemisphere. Quaternary Science Review 8: 359-368.
Isla, F.I. 1998. Holocene coastal evolution in Buenos Aires Province, Argentina. Quaternary of South America and Antarctic Peninsula 11: 297-321.
Lindemann, E. 1928. Abteilung Peridineae (Dinoflagellatae). En: A. Engler y K. Prantl (eds.), Die Natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren Arten insbesondere den Nutzpflanzen. Zweite Stark vermehrte und verbesserte Auflage herausgegeben von A. Engler. 2 Band, Leipzig, Wilhelm Engelmann, pp. 3-104.
Loeblich, A.R.JR. y Loeblich, A.R.III. 1966. Index to the genera, subgenera, and sections of the Pyrrhophyta. Studies in Tropical Oceanography 3: 94
Marcovecchio, J. 2000. [Estudio de la calidad del agua en la ría de Bahía Blanca (2° etapa). Informe Final, Instituto Argentino de Oceanografía, Bahía Blanca, 53pp. Inédito.].
Marret, F. y Scourse, J. 2002. Control of modern dinoflagellate cyst distribution in the Irish and Celtic seas by seasonal stratification dynamics. Marine Micropaleontology 47: 101-116.
Martin, F. 1993. Acritarchs: a review. Biological Review 68: 475-538.
Martínez, D.E. 2002. [Microfauna (Ostracoda, Crustacea) del Cuaternario de las cercanías de Bahía Blanca, República Argentina. Tesis Doctoral, Universidad Nacional del Sur, Bahía Blanca, 378 pp. Inédito.].
Matthiessen, J. 1995. Distribution patterns of dinoflagellate cysts and other organic walled microfossils in recent Norwegian- Greenland Sea sediments. Marine Micropaleontology 24: 307-334.
Matsuoka, K., McMinn, A. y Wrenn, J.H. 1997. Restudy of the holotype of Operculodinium centrocarpum (Deflandre and Cookson) Wall (Dinophyceae) from the Miocene of Australia, and the taxonomy of related species. Palynology 21: 19-33.
Mudie, P.J. 1980. [Palynology of later Quaternary marine sediments. Tesis doctoral, Department of Geology, Dalhousie University, Halifax, 628 pp. Inédito.].
Mudie, P.J. 1992. Circum-arctic Quaternary and neogene marine palynofloras: paleoecology and statistical analysis. En: M.J. Head y J.H. Wrenn (eds.), Neogene and Quaternary dinoflagellate cysts and acritarchs, American Association of Stratigraphic Palynologists Foundation, pp. 347-390.
Mudie, P.J. y Harland, R. 1996. Chapter 21. Aquatic Quaternary. En: J. Jansonius y D.C. McGregor (eds.), Palynology: principles and applications, American Association of Stratigraphic Palynologists Foundation, pp. 843-877.
Parker, G. y Violante, R.A. 1982. Geología del frente de costa y plataforma interior entre Pinamar y Mar de Ajó. Acta Oceanográfica Argentina 3: 57-91.
Pascher, A. 1914. Uber Flagellaten und Algen. Deutsche Botanische Gessellschaft 32: 136-160.
Perillo, G.M.E. y Piccolo, M.C. 1991. Tidal response in the Bahía Blanca estuary. Journal of Coastal Research 7: 437-449.
Piccolo, M.C. y Perillo, G.M. 1990. Physical characteristics of the Bahía Blanca Estuary (Argentina). Estuarine, Coastal and Shelf Science 34: 303-347.
Piccolo, M.C., Perillo, G.M.E. y Arango, J.M. 1987. Hidrografía del estuario de Bahía Blanca, Argentina. Geofísica 26: 75-89.
Rabassa, J., Heusser, C.J. y Stuckenrath, R. 1986. New data on Holocene sea transgression in the Beagle Channel, Tierra del Fuego, Argentina. Quaternary of South America and Antarctic Peninsula 4: 291-309.
Rabassa, J., Bujalesky, G., Meglioli, A., Coronato, A., Gordillo, S., Roig, C. y Salemme, M. 1992. The Quaternary of Tierra del Fuego, Argentina: the status of our knowledge. Sveriges Geologiska Undersokning 81: 249-256.
Reid, P.C. 1974. Gonyaulacacean dinoflagellate cysts from the British Isles. Nova Hedwigia 25: 579-637.
Reid, P.C., 1975. A regional sub-divisionof dinoflagellate cysts around the British Isles. New Phytology 75: 589-603.
Rochon, A., De Vernal, A., Turon, J.L., Matthiessen, J. y Head, M.J. 1999. Distribution of recent dinoflagellate cysts in surface sediments from the North Atlantic ocean and adjacent seas in relation to sea-surface parameters. American Association of Stratigraphic Palynologists Contributions Series 35: 152 pp.
Sarjeant, W.A.S. 1970. The genus Spiniferites Mantell, 1850 (Dinophyceae). Grana 10: 74-78.
Sarjeant, W.A.S. 1985. A restudy of some dinoflagellate cysts holotypes in the University of Kiel Collections: 6. Late Cretaceous dinoflagellate cysts and other palynomorphs in the O. Wetzel Collection. Meyniana 37: 129-185.
Sarjeant, W.A.S. y Stancliffe, R.P.W. 1994. The Micrhystridium and Veryhachium complexes (Acritarcha: Acantomorphitae and Poligonomorphitae): a taxonomic reconsideration. Micropaleontology 40: 1-77.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen, K.A., Kromer, B., McCormac, G., van der Plicht, J. y Spurk, M. 1998. INTCAL98 radiocarbon age calibration, 24,000 cal BP. Radiocarbon 40: 1041-1084.
Taylor, F.J.R. 1980. On dinoflagellate evolution. Biosystems 13: 65- 108.
Taylor, F.J.R. 1987. Ecology of dinoflagellates. En: F.J.R. Taylor (ed.), The biology of dinoflagellates, Botanical Monographs 21: 399-502.
Verettoni, H.N. y Aramayo, E. 1976. Comunidades vegetales de la región de Bahía Blanca. Harris Ed., Bahía Blanca, 175 pp.
Violante, R.A. y Parker, G. 2004. The post-last glacial maximum transgression in the de la Plata River and adjacent inner continental shelf, Argentina. Quaternary International 114: 167-181.
Wall, D. 1967. Fossil microplankton in deep-sea cores from the Caribbean Sea. Palaeontology 10: 95-123.
Wall, D. y Dale, B. 1968. Modern dinoflagellate cysts and evolution of the Peridiniales. Micropaleontology 14: 265-304.
Wall, D. y Dale, B. 1970. Living hystrichosphaerid dinoflagellate spores from Bermuda and Puerto Rico. Micropaleontology 16: 47-58.
Wall, D., Dale, B., Lohmann, G.P. y Smith, W.K. 1977. The environmental and climatic distribution of dinoflagellate cysts in modern marine sediments from regions in the North and South Atlantic ocean and adyacent seas. Marine Micropaleontology 2: 121-200. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 9 de diciembre de 2004.
Aceptado: 1 de setiembre de 2005.

