










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.2 Buenos Aires abr./jun. 2006
Variabilidad de la icnofacies de Glossifungites en el contacto entre las Formaciones Sarmiento (Eoceno medio - Mioceno temprano) y Chenque (Mioceno temprano) en el Golfo San Jorge, Chubut, Argentina
Noelia B. Carmona1, Juan José Ponce1, María Gabriela Mángano2 y Luis A. Buatois2
1 Laboratorio de Geología Andina. Centro Austral de Investigaciones Científicas. CONICET. B. Houssay 200, 9410 Ushuaia, Tierra del Fuego, Argentina. carmonanb@yahoo.com.ar
2 Department of Geological Sciences. University of Saskatchewan. 114 Science Place, Saskatoon, SK S7N 5E2, Canadá
Abstract. Variability of the Glossifungites ichnofacies at the boundary between the Sarmiento Formation (middle Eocene- early Miocene) and Chenque Formation (early Miocene) in San Jorge Gulf, Chubut, Argentina . This study focuses on the ichnologic and stratigraphic characteristics of the boundary surface between the Sarmiento (middle Eocene- early Miocene) and Chenque Formations (early Miocene). The analyzed outcrops are located near Comodoro Rivadavia city, Chubut Province, Patagonia, Argentina. The studied boundary represents a co-planar surface of lowstand erosion and transgressive erosion that produced exhumation of deposits belonging to the Sarmiento Formation, allowing the development of the Glossifungites ichnofacies. This ichnofacies is characteristic of stiff and dewatered sediments (firmgrounds) and comprises a particular suite of substrate-controlled trace fossils. In the studied outcrops (Astra, bahía Solano and Infiernillo), this boundary surface was colonized by decapod crustaceans that constructed galleries (Thalassinoides isp.) and bivalves that constructed dwelling structures (Gastrochaenolites ornatus). Although in all these localities the ichnotaxonomic composition does not show significant variations, important paleoecological differences between the trace fossil suites preserved in each outcrop can be inferred. Recent assemblages that colonize firmground surfaces seem to be controlled by three main parameters: sediment texture, substrate firmness and bathymetry. In the Miocene examples, the observed variability of the Glossifungites ichnofacies was probably controlled by the firmness of the surface exhumed during the Leonense transgression. Analysis of the ichnologic content, truncation of trace fossils, and the relationship between biogenic structures suggests a complex history for this erosive surface, involving successive events of ravinement erosion and benthic colonization.
Resumen. Este estudio está centrado en las características icnológicas y estratigráficas del contacto entre la Formación Sarmiento (Eoceno medio - Mioceno temprano) y la Formación Chenque (Mioceno temprano). Los afloramientos analizados se localizan en las cercanías de la ciudad de Comodoro Rivadavia, provincia del Chubut, Patagonia, Argentina. El contacto estudiado representa una superficie co-planar de erosión de nivel de mar bajo y erosión transgresiva que produjo la exhumación de los depósitos pertenecientes a la Formación Sarmiento, permitiendo el desarrollo de la icnofacies de Glossifungites. Esta icnofacies es característica de sedimentos compactados y deshidratados (sustratos firmes) y comprende una suite de trazas fósiles particulares que se desarrollan en ambientes con control del sustrato. En los afloramientos analizados (Astra, bahía Solano e Infiernillo), este contacto fue colonizado por crustáceos decápodos constructores de galerías (Thalassinoides isp.) y bivalvos productores de estructuras de habitación (Gastrochaenolites ornatus). A pesar que la composición taxonómica no presenta variaciones significativas, se pueden inferir importantes diferencias de carácter paleoecológico entre las asociaciones de trazas fósiles preservadas en cada uno de los afloramientos. Las asociaciones modernas de organismos que colonizan superficies firmes parecen estar controladas por tres parámetros principales: la textura del sedimento, la firmeza del sustrato y la batimetría. En las sucesiones miocenas, la variabilidad hallada en los diferentes ejemplos de la icnofacies de Glossifungites estaría controlada probablemente por la firmeza de la superficie exhumada durante la transgresión Leonense. El análisis del contenido icnológico, el truncamiento de las estructuras y la relación existente entre las trazas fósiles sugiere una compleja historia para esta superficie, con sucesivos eventos de ravinamiento erosivo y colonización bentónica.
Key words. Trace fossils; Glossifungites ichnofacies; Chenque Formation; Miocene; Patagonia; Transgression; Firmground.
Palabras clave. Trazas fósiles; Icnofacies de Glossifungites; Formación Chenque; Mioceno; Patagonia; Transgresión; Sustrato firme.
Introducción
En los últimos años la icnología ha experimentado un importante desarrollo. En parte, este hecho se ha debido al creciente reconocimiento de las trazas fósiles como herramientas de gran utilidad en la identificación y entendimiento de superficies con importancia estratigráfica (MacEachern et al., 1992; Pemberton et al., 1992a, 1992b, 1992c, 1997; Gingras et al., 2001; Gibert y Robles, 2005). En tal sentido, el desarrollo de estudios en icnofacies sustrato-controladas permitió un importante avance en los análisis estratigráficos secuenciales para el reconocimiento de superficies aloestratigráficas.
Actualmente se reconocen tres icnofacies sustrato controladas: la icnofacies de Trypanites (subdividida posteriormente en las asociaciones de Entobia y Gnatichnus) (Bromley y Asgaard, 1993; Gibert et al., 1998), la cual se desarrolla en sustratos litificados o cementados (hardgrounds); la icnofacies de Teredolites, que se desarrolla en sustratos de madera (woodgrounds) y la icnofacies de Glossifungites, desarrollada en sustratos firmes y compactados, no litificados (firmgrounds). La icnofacies de Glossifungites fue definida por Seilacher (1967) y posteriormente revisada por Frey y Seilacher (1980) y Pemberton y Frey (1985). Esta icnofacies se caracteriza por la presencia de estructuras biogénicas con morfologías variadas, siendo comunes las excavaciones ramificadas y las estructuras verticales, cilíndricas, en forma de gota o U, con límites bien definidos y netos, presentando en algunos casos marcas de rasguñaduras (scratch marks) (Frey y Pemberton, 1984, 1985; Pemberton et al., 1992a, 1992c; MacEachern et al., 1992; Uchman et al., 2000; Gingras et al., 2001; Buatois et al., 2002). Generalmente el relleno de las estructuras biogénicas es pasivo, con una textura diferente a la de la roca hospedante y similar a la de los estratos suprayacentes. Las trazas fósiles que caracterizan esta icnofacies pertenecen en su mayoría a organismos suspensívoros. También son comunes las estructuras con desarrollo de spreiten protrusivos, formados en respuesta al crecimiento de los organismos productores (e.g. Diplocraterion) (MacEachern et al., 1992). La icnodiversidad de esta icnofacies es generalmente baja, aunque la abundancia de especimenes de cada icnotaxón puede ser elevada.
El reconocimiento de la icnofacies de Glossifungites en el registro geológico es de suma importancia debido a sus implicancias en estratigrafía genética. En este trabajo se analiza la superficie de contacto entre las Formaciones Sarmiento (Eoceno medio - Mioceno temprano) y Chenque (Mioceno temprano) en tres localidades de los alrededores de la ciudad de Comodoro Rivadavia, provincia de Chubut, Argentina. Las características del límite entre ambas formaciones fueron siempre controversiales ya que algunos autores postularon un límite transicional mientras que otros reconocieron una superficie de discontinuidad entre ambas sucesiones (Feruglio, 1949; Camacho y Fernández, 1956; Camacho, 1974; Andreis et al., 1975; Andreis, 1977; Expósito, 1977; Bellosi, 1987, 1995).
En las tres localidades analizadas se encuentra bien expuesto el contacto entre las Formaciones Sarmiento y Chenque. Este contacto está representado esencialmente por una superficie de discontinuidad, con el desarrollo de la icnofacies de Glossifungites en el tope de los estratos de la Formación Sarmiento. Sin embargo, en las diferentes localidades se encontraron variaciones en las abundancias relativas de las estructuras biogénicas y en las relaciones espaciales que las mismas presentan, sugiriendo una compleja historia de interacción entre procesos físicos y biogénicos. Por otra parte, un análisis detallado de esta superficie palimpséstica revela aún más complejidades, estando la suite de sustrato firme posiblemente representada por más de un evento erosivo y subsiguiente colonización. El objetivo principal de este trabajo se centró en el estudio de detalle de la superficie de contacto entre las Formaciones Sarmiento y Chenque en las distintas localidades analizadas, a fin de evaluar las diferencias en la icnofauna en función de posibles controles físicos y biológicos. Asimismo, en este trabajo se analizan posibles analogías con estudios de la icnofacies de Glossifungites en ambientes modernos, evaluando las limitaciones de una perspectiva estrictamente actualista, donde la variante temporal a escala geológica no entra en juego.
Área de estudio y marco geológico
Las localidades analizadas se encuentran en las cercanías de la ciudad de Comodoro Rivadavia (figura 1). De norte a sur, la primera localidad se ubica al oeste de bahía Solano, la segunda en Astra, y la tercera, denominada Infiernillo, sobre la costa atlántica, al este del Cerro Chenque.
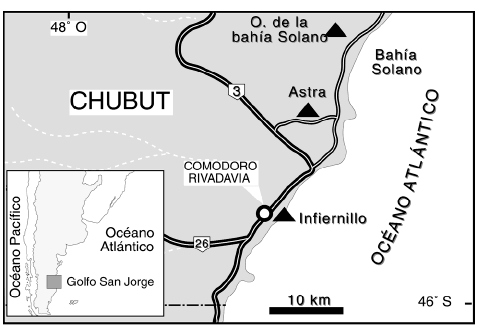
Figura 1. Mapa de ubicación del área estudiada. Las tres localidades analizadas se encuentran indicadas por triángulos negros (Oeste de la bahía Solano, Astra e Infiernillo) / map showing the studied area. The three analyzed localities are indicated by black triangles (west of Solano bay, Astra and Infiernillo).
En todas las localidades estudiadas se identificaron los estratos tobáceos pertenecientes a la Formación Sarmiento, sobre los cuales se apoyan las capas arenosas y areno-pelíticas de la Formación Chenque (figura 2). La Formación Sarmiento incluye principalmente tobas masivas depositadas en ambientes continentales durante el Eoceno medio y el Mioceno temprano (Bellosi, 1987, 1995, Bellosi y Genise, 2004). A partir del Mioceno temprano, la sedimentación en la cuenca San Jorge estuvo dominada por procesos marinos vinculados al desarrollo de dos transgresiones de extensión regional (Bellosi, 1987, 1995). Du rante la primera de estas transgresiones, denominada Leonense, tuvo lugar la depositación de las Secuencias I y II de la Formación Chenque (Bellosi, 1987). Estas secuencias se encuentran integradas por depósitos de pelitas tobáceas y areniscas finas glauconíticas, dispuestas en arreglos estrato-grano creciente, depositadas mayormente en ambientes marinos abiertos. Durante la segunda transgresión (transgresión Superpatagoniense) se produjo la acumulación de otras tres secuencias, las cuales se caracterizan por presentar un mayor contenido arenoso y el dominio de procesos mareales en ambientes restringidos (Bellosi, 1987). En este trabajo nos referiremos a las fases iniciales de la primera transgresión (Leonense), que se inicia con la superficie erosiva que separa las Formaciones Sarmiento y Chenque.
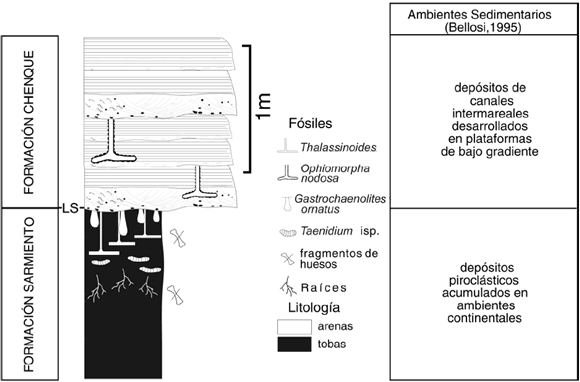
Figura 2. Sección esquemática mostrando el contacto entre las Formaciones Sarmiento y Chenque en las localidades estudiadas. En el gráfico se resumen las principales características litológicas, icnológicas y ambientes depositacionales (estos últimos basados en Bellosi, 1995) / schematic section showing the contact between the Sarmiento and Chenque Formations. The figure includes the main lithologic and ichnologic characteristics, as well as the interpreted environments (based on Bellosi, 1995).
Contenido icnológico y características sedimentológicas
En todas las localidades analizadas se identificó la icnofacies de Glossifungites con una compleja estructura interna. La icnodiversidad es similar en todas las localidades. Básicamente se identificaron dos estructuras biogénicas principales. La primera corresponde a galerías tridimensionales, asignadas a Thalassinoides isp. Estas estructuras son atribuidas a la actividad de crustáceos decápodos y se caracterizan por la presencia de componentes horizontales y verticales, con una clara sección cilíndrica, un diámetro variable entre 2-4,5 cm y ramificaciones que forman en general ángulos rectos. Los especimenes presentan límites netos, sin pared revestida, y se encuentran rellenos de modo pasivo principalmente por arenas gruesas a finas, provenientes de los depósitos suprayacentes. La abundancia relativa de estas estructuras es notablemente variable en las distintas localidades, siendo muy conspicuas en Astra y muy pobremente representadas al oeste de bahía Solano.
La otra estructura biogénica que caracteriza al límite entre las Formaciones Sarmiento y Chenque corresponde a Gastrochaenolites ornatus. Estas estructuras corresponden a habitaciones o domicilios de bivalvos suspensívoros y tienen forma de botella o gota, con diámetros basales que varían entre 0,2 a 2,5 cm y longitudes máximas de 9 cm. Un análisis de detalle revela que muchas de estas estructuras se en cuentran ornamentadas con bioglifos, producto del movimiento de las valvas del organismo constructor contra las paredes de la excavación. Si bien el icnogénero Gastrochaenolites fue primariamente definido para perforaciones desarrolladas en sustratos duros (hardgrounds) y no para sustratos firmes como los de este estudio, la morfología fina de estos especimenes es indistinguible de aquella presente en ejemplares comúnmente descriptos para sustratos duros. Por tal motivo, y considerando que la clasificación icnotaxonómica es morfo-etológica, resultaría artificial asignar el material analizado a un nuevo icnogénero. Tal cual fue señalado por Ekdale y Bromley (2001) con respecto a Gastrochaenolites oelandicus, la distinción entre excavaciones y perforaciones es en algunos casos difícil de establecer y a su vez resultaría confuso otorgar distinta asignación icnotaxonómica a estructuras que presentan la misma morfología y que parecen resultar esencialmente del mismo comportamiento.
Si bien la icnodiversidad observada regionalmente a lo largo de la superficie es similar, se registraron notables diferencias en cuanto a la abundancia relativa y a las relaciones espaciales de las estructuras biogénicas. Dichas diferencias son evaluadas en función de los parámetros que actuaron al momento de la colonización (variable ecológica) y del filtro tafonómico que representa colonizaciones sucesivas sobreimpuestas en un mismo depósito (variable temporal). Es artificial tratar de determinar un índice de bioturbación para la capa que subyace a la superficie de contacto. Un análisis de icnofábricas revela una compleja historia que resultó en una superposición de estructuras correspondientes a sucesivas icnocenoses, produciendo una icnofábrica compleja, situación en la cual la utilidad de índices de bioturbación es algo limitada (cf. Ekdale y Bromley, 1991).
En la localidad ubicada al Oeste de bahía Solano, la superficie de erosión se encuentra completamente retrabajada por Gastrochaenolites ornatus (figura 3), reflejando la actividad de bivalvos de diferentes tamaños (figura 3.A). En general, los límites de estas estructuras están realzados por la precipitación de yeso, lo cual permite una mejor identificación y visualización de los detalles morfológicos de estas trazas fósiles de elite (figuras 3.B, C y E). En general, pueden distinguirse dos estilos preservacionales de Gastrochaenolites ornatus: aquellos que presentan en su interior moldes internos de los bivalvos productores (pertenecientes a la familia Pholadidae) (figura 3.D) y aquellos que se encuentran rellenos en su totalidad por material arenoso. Los ejemplares de Gastrochaenolites ornatus poseen cuellos alargados y no exhiben evidencias de haber sufrido erosión significativa. En forma aislada también se identificaron estructuras asignables a galerías de crustáceos decápodos (figura 3.A). Si bien la arquitectura de estas estructuras no se encuentra bien preservada, por similitud en morfología y tamaño las mismas son asignadas a Thalassinoides isp. Cabe destacar que Bellosi (1987) mencionó la presencia de una superficie colonizada por bivalvos en la zona de Pico Salamanca, cercana a la localidad del Oeste de bahía Solano.
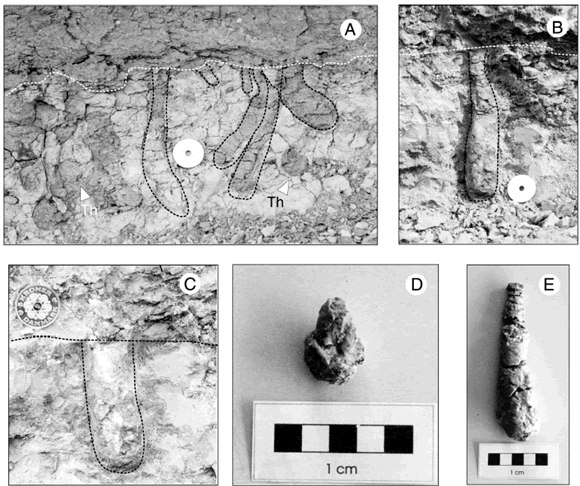
Figura 3. Icnofacies de Glossifungites en el Oeste de la bahía Solano. A, Desarrollo de las estructuras de habitación de bivalvos en la superficie que limita ambas formaciones. B y C, Ejemplares excelentemente preservados por el precipitado de yeso. En A, B y C, diámetro de la moneda de 28,4 mm. D y E, Ejemplares colectados en el campo. En D, se observa preservado el molde interno del bivalvo constructor de la excavación / Glossifungites ichnofacies at west of Solano bay. A, Bivalve dwelling structures developed on the boundary surface between the formations. B and C, Specimens superbly preserved due to a gypsum precipitate. In A, B and C, coin diameter is 28.4 mm. D and E. Collected specimens. The internal mould of the bivalve is preserved in D.
En la localidad de Astra, la icnofacies de Glossifungites tiene características bastante diferentes a las registradas en la localidad anterior (figuras 4.A-D). La superficie de contacto y el depósito infrayacente están dominados por Thalassinoides isp. La abundancia de estas estructuras biogénicas es muy alta por lo que en un primer relevamiento en esta localidad, no se identificaron otras trazas fósiles, y se consideró que la asociación era monoespecífica (Carmona et al., 2004). En un trabajo de campo posterior se reconocieron escasas estructuras de habitación de bivalvos (Gastrochaenolites ornatus, figuras 4.C y D).

Figura 4. Icnofacies de Glossifungites en la localidad de Astra. A, Vista de la superficie de contacto entre las Formaciones Sarmiento y Chenque (indicada por una línea de puntos). B, C y D, Estructuras biogénicas que caracterizan esta localidad (Th = Thalassinoides isp., Gs = Gastrochaenolites ornatus). En B, moneda de 23 mm de diámetro. En C y D, tapa de la lente de 55 mm de diámetro. En todos los casos el límite entre las formaciones se encuentra indicado por una línea de puntos / Glossifungites ichnofacies in Astra. A, View of the boundary surface between the Sarmiento and Chenque Formations (indicated by a dash line). B, C and D, Characteristic ichnofossils from this locality (Th = Thalassinoides isp., Gs = Gastrochaenolites ornatus). In B, coin diameter is 23 mm; in C and D, lens cap diameter is 55 mm.
Finalmente, en la localidad denominada Infiernillo la icnofacies de Glossifungites presenta tanto estructuras de habitación de bivalvos correspondientes a Gastrochaenolites ornatus (figuras 5.A-D), como galerías asignadas a Thalassinoides isp., si bien estas últimas no son muy abundantes. Numerosas estructuras de bivalvos están preservadas parcialmente, siendo identificables en algunos casos, sólo las bases de las mismas. Curiosamente, este tipo de preservación también ha sido observada por Doménech et al. (2001) para una plataforma rocosa del Mioceno medio. En dichos depósitos, los especimenes de Gastrochaenolites se caracterizan por tener preservadas sólo sus bases, debido a que la intensa bioerosión junto a la acción física del oleaje habrían producido el debilitamiento de los sectores superiores de la plataforma rocosa. En los depósitos estudiados en el presente trabajo, la preservación de las bases de las estructuras de bivalvos responde básicamente a la erosión, la cual cumplió un importante papel durante la transgresión, no sólo al inicio de la misma, sino en los sucesivos pulsos erosivos que modificaron la arquitectura y estructuras biogénicas de esta superficie. Al igual que en la localidad al oeste de bahía Solano, se detectaron dos tipos de preservaciones en la población de Gastrochaenolites ornatus. Por un lado, se preservaron las estructuras con los moldes internos de los bivalvos productores (figura 5.D), mientras que otros ejemplares sólo mostraron relleno arenoso (figura 5.C). De este modo, la icnofacies de Glossifungites en esta localidad revela un complejo interjuego entre los eventos erosivos que dieron lugar a esta superficie y las sucesivas colonizaciones. El material de Gastrochaenolites ornatus colectado en esta localidad se encuentra en la Colección de Paleontología de Invertebrados del Centro Austral de Investigaciones Científicas (CADIC PI 39-45).

Figura 5. Icnofacies de Glossifungites en el Infiernillo. A, Vista general del afloramiento. B, Thalassinoides isp. (moneda de escala de 17 mm de diámetro). C, Vista basal de ejemplares de Gastrochaenolites ornatus (CADIC PI 39). Notese las diferencias de tamaño de las estructuras biogénicas. D, Vista lateral de las estructuras de bivalvos en las que se destacan los moldes internos de los organismos productores. En C y D diámetro de la moneda 18,2 mm / Glossifungites ichnofacies in Infiernillo. A, General view of the outcrop. The boundary between the Sarmiento and Chenque Formations is clearly visible in this locality. B, Thalassinoides isp. (coin is 17 mm in diameter). C. Basal view of Gastrochaenolites ornatus (CADIC PI 39). Note differences in size between the biogenic structures. D, Lateral view of the bivalve structures, with preservation of the internal moulds of the bivalves. In C and D, coin is 18.2 mm in diameter.
Las características sedimentológicas de las sucesiones clásticas analizadas son similares en cuanto a facies y geometría de los cuerpos en las tres localidades. Los depósitos de la Formación Sarmiento están integrados por niveles tobáceos masivos dispuestos en bancos tabulares, que localmente contienen trazas de raíces y asociaciones monoespecíficas de estructuras meniscadas asignadas a Taenidium isp. Esto pone de relieve la existencia de una icnofauna previa, representando a la icnofacies de Scoyenia. Las estructuras biogénicas de la icnofacies de Glossifungites se encuentran hacia el techo de esta formación. Sobre la Formación Sarmiento y en relación de discordancia (superficie co-planar), se acumularon los depósitos basales de la Formación Chenque, integrados por areniscas medianas a finas con delgados niveles pelíticos intercalados y abundante contenido bioclástico. Internamente estos bancos presentan estructuras de tipo tidal bundle, relacionadas a procesos mareales, y ondulitas simétricas en el techo, generadas por corrientes oscilatorias. En los depósitos de canales mareales presentes en la Formación Chenque se disponen sistemas de galerías atribuibles a Ophiomorpha nodosa, que indican el establecimiento de una icnofauna de sustratos blandos con posterioridad al desarrollo de la superficie de omisión.
Análogos modernos e implicancias paleoecológicas
La variabilidad en el contenido de estructuras bio génicas en las diferentes localidades no es totalmente explicable en función de una historia tafonómica, sino que posiblemente refleje un control a nivel paleoecológico. Pemberton y Frey (1985) estudiaron un ejemplo moderno de la icnofacies de Glossifungites en la costa de Georgia. Estos autores observaron que las asociaciones de organismos se encontraban distribuidas de acuerdo a ciertos controles ambientales como la energía del oleaje, la firmeza del sustrato y el tiempo de exposición subaérea. Recientemente Gingras et al. (2001) estudiaron el desarrollo de una icnofacies de Glossifungites actual en la bahía de Willapa. Estos autores determinaron que las diferencias encontradas en las asociaciones de estructuras biogénicas respondían a una serie de parámetros que incluyen la batimetría, la textura del sedimento, la firmeza del sustrato y la presencia o ausencia de una delgada capa de sedimento blando sobre la superficie firme. El análisis de estos parámetros les permitió determinar tres asociaciones de estructuras biogénicas en la bahía de Willapa. La zona intermareal superior está caracterizada por el dominio de la asociación de Polydora (organismos vermiformes que construyen estructuras similares a Diplocraterion y Arenicolites). La zona intermareal media a inferior está dominada por la asociación de Petricola, bivalvos que construyen estructuras similares a Gastrochaenolites en el registro fósil. Esta asociación se caracteriza además por encontrarse en sustratos de mayor firmeza. Finalmente, la tercera asociación, dominada por estructuras del crustáceo decápodo Upogebia, se encuentra en los sectores submareales a intermareales medios, en sustratos de menor firmeza que la asociación anterior, y preferentemente en sedimentos finos. Las estructuras que construyen estos crustáceos han sido comparadas por Gingras et al. (2001) con Thalassinoides. Sin embargo muchas de las morfologías ilustradas en dicho trabajo, muestran formas en "Y" propias del icnogénero Psilonichnus (ver figura 4.28 en Bromley, 1996).
En analogía con estudios modernos, las variaciones en las abundancias relativas encontradas en las sucesiones miocenas del Golfo San Jorge, responderí an a parámetros paleoecológicos que habrían condicionado el establecimiento y la colonización de la fauna marina durante la transgresión Leonense en las distintas localidades estudiadas.
Oeste de la bahía Solano
La superficie transgresiva en este afloramiento fue profusamente colonizada por bivalvos; siendo las estructuras de Gastrochaenolites ornatus muy abundantes y constituyendo las trazas fósiles dominantes de la icnofacies de Glossifungites en esta localidad (figuras 3 y 6.A). Localmente se registraron secciones aisladas cilíndricas de estructuras horizontales, con relleno pasivo asignadas a Thalassinoides isp. (figura 3.A). La abundancia de estructuras de habitación de bivalvos y la escasa presencia de estructuras de crustáceos sugiere la existencia de un factor limitante al momento de la colonización de la fauna bentónica. Esto estaría en acuerdo con el estudio realizado por Pemberton y Frey (1985) y Gingras et al. (2001), quienes notaron que los bivalvos prefieren sustratos relativamente más firmes que las faunas de crustáceos. La exhumación del sustrato habría dejado disponibles en superficie, niveles estratales relativamente profundos y más compactos. La escasez de estructuras de crustáceos y la gran variabilidad de tamaños en Gastrochaenolites ornatus (figura 3.A) sugieren que, si bien existió tiempo disponible para la colonización, los crustáceos no habrían encontrado condiciones óptimas para habitar este sustrato, siendo el grado de compactación del sedimento el posible factor limitante.
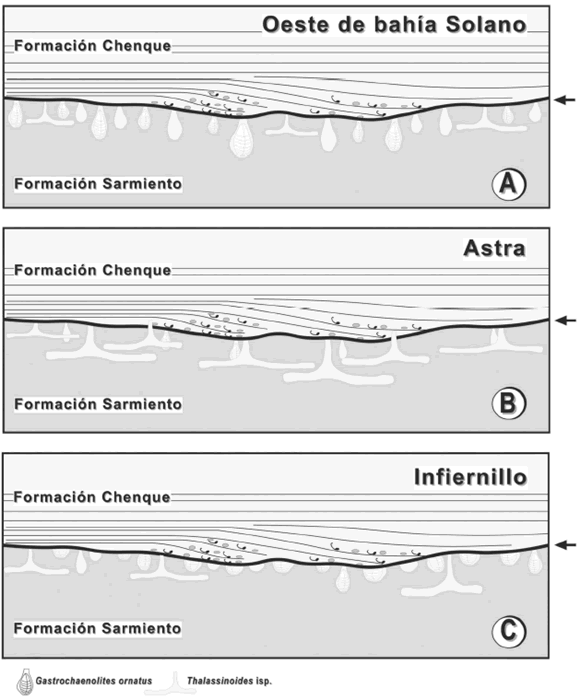
Figura 6. Íconos de las asociaciones icnofosilíferas en A, Oeste de la bahía Solano, B, Astra y C, Infiernillo. Las flechas indican el contacto entre la Formación Sarmiento y la Formación Chenque / icons of the trace fossil associations in A, West of Solano Bay-, B, Astra and C, Infiernillo. Arrows indicate the boundary between Sarmiento and Chenque Formations.
Astra
En esta localidad la icnofacies de Glossifungites está dominada por Thalassinoides isp. (figura 4). En forma subordinada hay ejemplares de Gastrochaenolites ornatus, los cuales en la mayoría de los casos fueron obliterados por las galerías de crustáceos decápodos (figura 6.B). Esta relación de corte indica que la colonización por parte de crustáceos decápodos se produjo con posterioridad al establecimiento de los bivalvos. La abundancia original de las estructuras de bivalvos no se pudo evaluar con certeza. Una posibilidad es considerar que este patrón responde a causas ecológicas. En este contexto, la colonización de bivalvos fue incipiente e interrumpida por la colonización de crustáceos, cuya actividad excluyó a los bivalvos. Otra interpretación alternativa sería la de un "efecto tafonómico", en la cual la profusa colonización de estos niveles por parte de los crustáceos decápodos haya obliterado significativamente cualquier estructura biogénica previa, produciendo una disminución aparente en la abundancia de las estructuras de habitación de los bivalvos. Si el extenso desarrollo de galerías de Thalassinoides responde a causas ecológicas (e.g. no es un efecto tafonómico), esto podría sugerir que el sustrato exhumado y disponible para ser colonizado no fue muy firme en esta localidad. La baja abundancia relativa de Gastrochaenolites ornatus (en comparación con las otras localidades estudiadas) favorecería esta interpretación.
Infiernillo
La asociación de trazas fósiles en esta localidad se caracteriza por Gastrochaenolites ornatus (figuras 5 y 6.C) y en menor proporción por Thalassinoides isp. Asimismo, cabe destacar que la superficie analizada está completamente retrabajada por trazas de bivalvos que exhiben truncamiento erosivo, preservándose en algunos casos, sólo las bases de los ejemplares de Gastrochaenolites ornatus (figura 5.C). Esta condición sugiere que la historia de colonización y erosiónsedimentación fue compleja, indicando que la superficie transgresiva estuvo afectada por sucesivos eventos erosivos durante el inicio de la transgresión Leonense. Por otra parte, la gran abundancia y el amplio rango de tamaños de Gastrochaenolites ornatus permite inferir que el sustrato firme, representado por la superficie exhumada de la Formación Sarmiento, estuvo disponible suficiente tiempo para la extensa colonización de los bivalvos.
Discusión
Además de la firmeza del sustrato, otros parámetros pudieron jugar un rol en relación con las variaciones encontradas en la icnofacies de Glossifungites. En el caso del control batimétrico, al comparar con los análogos modernos, se detectó que, si bien los bivalvos tienden a localizarse mayormente en ambientes más someros que los crustáceos decápodos, existe cierta superposición entre los hábitats preferidos por ambos organismos. Por esta razón se infiere que este parámetro no es un factor de primer orden para explicar las variaciones encontradas en el golfo San Jorge. Asimismo, los estudios realizados no revelan ningún patrón de distribución de estructuras biogénicas de tipo proximal-distal que ponga de manifiesto un control batimétrico. La textura del sedimento tampoco parece haber tenido un rol determinante en la composición de las asociaciones registradas, debido a que la composición de las capas en las que se desarrolló las icnofacies de Glossifungites en las diferentes localidades es básicamente la misma (los bancos tobáceos de la Formación Sarmiento). Entre los controles biológicos, el tipo trófico no parece haber jugado un rol de importancia. Si bien los crustáceos decápodos tienen una plasticidad mayor en cuanto a las estrategias de alimentación que los bivalvos aquí analizados, la presencia de rellenos claramente pasivos en ambas estructuras biogénicas permite inferir que las mismas fueron construidas por organismos suspensívoros, razón por la cual se descarta que la distribución de las trazas fósiles haya sido controlada por el tipo de alimentación. De este modo, se considera que la firmeza del sustrato, posiblemente regulada por el efecto local del descenso del nivel del mar y la dinámica de la transgresión, sería el parámetro que determinó en mayor medida las diferencias entre las asociaciones de Glossifungites en las localidades estudiadas.
El entendimiento de la icnofacies de Glossifungites se ha visto enriquecido recientemente por la dimensión ecológica de los análogos modernos (e.g. Pemberton y Frey, 1985; Gingras et al., 2001). Esta perspectiva "instantánea" requiere, sin embargo, de la adición de la variable temporal en el desarrollo de estas asociaciones en el registro estratigráfico. La relación local entre la erosión y la sedimentación, y el tiempo de disponibilidad del sustrato exhumado, previo al evento de sedimentación subsiguiente, modelan las características particulares de cada asociación incluidas en la icnofacies de Glossifungites. El análisis del desarrollo de estas asociaciones nos permite comprender la complejidad de los eventos que actuaron durante la transgresión Leonense. Es evidente que los eventos erosivos ocurridos durante esta transgresión borraron parte del registro icnológico, de manera tal que la reconstrucción de la paleocomunidad colonizadora a partir de las trazas fósiles, puede no corresponder a la verdadera comunidad endobentónica pionera. Existe una relación entre el factor temporal y la sucesión de eventos físicos propios de la transgresión que debe ser tomada en cuenta. También existe una relación entre el factor temporal y el desarrollo intrínseco de una comunidad bentónica, que sólo puede ser inferido al analizar las relaciones de corte entre las estructuras biogénicas, siendo las trazas más profundas (e.g. Thalassinoides isp. en Astra) las que presentan mejor grado de preservación y definición, mientras que las estructuras más someras se encuentran en general retrabajadas por los bioturbadores posteriores. La incorporación de estas diferentes escalas de análisis temporal a los estudios de las asociaciones de trazas fósiles nos permite obtener un mayor entendimiento de los parámetros paleoecológicos que regularon el desarrollo de las comunidades bentónicas, así como la complejidad de la historia tafonómica que registra la superficie analizada.
Historia tafonómica e implicancias aloestratigráficas
El reconocimiento de la icnofacies de Glossifungites en las localidades estudiadas, así como el análisis de su complejidad interna tiene un importante significado estratigráfico secuencial y es fundamental para el entendimiento de la relación entre las Formaciones Sarmiento y Chenque.
En ambientes silicoclásticos, los ejemplos de la icnofacies de Glossifungites se encuentran principalmente relacionados a exhumaciones erosivas del sustrato, que corresponden a discontinuidades erosivas, comúnmente producidas durante variaciones relativas del nivel del mar (MacEachern et al., 1992). El desarrollo de esta asociación de trazas fósiles implica que hubo un período de enterramiento, compactación y posterior exhumación que afectó arealmente a gran parte de la cuenca. De este modo, la presencia de la icnofacies de Glossifungites se puede relacionar con cambios relativos del nivel del mar que afectaron a los ambientes costeros de la cuenca San Jorge durante el Mioceno temprano.
La superficie erosiva que caracteriza al contacto entre las Formaciones Sarmiento y Chenque corresponde a una superficie co-planar, en la cual se encuentran amalgamadas la superficie erosiva de nivel de mar bajo (Surface of Lowstand Erosion) y la superficie erosiva de transgresión (Surface of Transgressive Erosion). Sumado a esto, la icnofauna de sustrato firme constituye en sí misma una icnofábrica compuesta. La presencia de una amplia gama de tamaños en los ejemplares de Gastrochaenolites ornatus y las variaciones en cuanto al tipo de relleno sugiere que la superficie estuvo disponible para la colonización durante un lapso considerable y que probablemente más de un evento de bioturbación estuvo involucrado a lo largo de su historia. A su vez, es notable el emplazamiento a distintas profundidades de los ejemplares de Thalassinoides en las tres localidades analizadas. La figura 7 presenta un modelo esquemático que, basado en la evidencia icnológica, permite reconstruir la sucesión de eventos que tuvieron lugar a partir del Mioceno temprano en la cuenca del Golfo San Jorge. En una primera etapa, se produjo la erosión de los depósitos continentales de la Formación Sarmiento (figura 7.A), los cuales presentan una icnofauna previa de estructuras meniscadas asignadas a Taenidium isp. y trazas de raíces. Esta superficie erosiva fue generada durante una fase de nivel de mar bajo (LST) y subsecuentemente sirvió de sustrato para la transgresión Leonense (figura 7.B). Esta transgresión está representada por una superficie de ravinamiento sobre la cual se encuentran depósitos arenosos con intraclastos tobáceos pertenecientes a la Formación Sarmiento, así como fragmentos de conchillas de invertebrados marinos de la Formación Chenque. Dentro de este contexto transgresivo y antes de la depositación de los niveles arenosos de la Formación Chenque, tuvo lugar una primera fase de colonización del sustrato (figura 7.C). Basado en analogías con estudios modernos, un parámetro determinante a nivel ecológico fue el grado de firmeza del sustrato que posiblemente condicionó la abundancia relativa de los distintos colonizadores. De este modo, las variaciones en abundancia de Thalassinoides y Gastrochaenolites ornatus entre las distintas localidades analizadas estarían reguladas posiblemente por parámetros ecológicos (ver Análogos modernos e implicancias paleoecológicas), siendo las condiciones óptimas para la colonización de Thalassinoides en Astra y menos propicias en Infiernillo y Oeste de bahía Solano (estas dos últimas localidades con mayor abundancia de Gastrochaenolites ornatus). Subsecuentemente durante la fase transgresiva, la superficie habría sido nuevamente retrabajada, siendo re-exhumada y quedando nuevamente disponible para la colonización de organismos capaces de excavar en un sustrato firme (figura 7.D). En algunas localidades (e.g. Infiernillo) los ejemplares de Gastrochaenolites ornatus están decapitados y sólo están preservadas las bases de los ejemplares, posiblemente reflejando el grado de exposición a la erosión por oleaje durante la transgresión. Por el contrario, la preservación de especimenes de Gastrochaenolites ornatus con cuellos alargados en el oeste de bahía Solano revela escasa erosión y probablemente emplazamiento en zonas más protegidas de la costa. Esta compleja arquitectura registra las sucesivas suites de omisión. La etapa final en la evolución del sistema muestra el desarro llo de depósitos marinos someros que forman parte de los niveles basales de la Formación Chenque, con presencia de galerías de crustáceos decápodos atribuidas a Ophiomorpha nodosa, que indican el establecimiento de una icnofauna característica de sustratos blandos desarrollada con posterioridad a la superficie de omisión.

Figura 7. Representación esquemática de los eventos ocurridos durante la transgresión Leonense en el Mioceno temprano. A, Desarrollo de una superficie erosiva de descenso del nivel del mar en el techo de la Formación Sarmiento (Cortejo sedimentario de nivel del mar bajo). B, Inicio del evento transgresivo. C, Colonización inicial de la superficie de ravinamiento. D, Fase de colonización subsecuente y depositación de la Formación Chenque. Ophiomorpha nodosa registra colonización de un sustrato blando en depósitos de canales mareales / idealized representation of the Leonense transgression during the early Miocene. A, Development of a lowstand surface of erosion at the top of the Sarmiento Formation (Lowstand Systems Tract). B, Initial transgressive event. C, Initial colonization of the ravinement surface. D, Subsequent colonization and deposition of the Chenque Formation. Ophiomorpha nodosa records colonization of a softground substrate in the tidal channel deposits.
Conclusiones
La identificación y el análisis de diversos parámetros que regulan la composición taxonómica de las icnofacies sustratos-controladas, junto con la inclusión de la variable temporal, permiten comprender desde un punto de vista paleoecológico y tafonómico las variaciones registradas en las sucesiones fósiles. En este estudio se infiere que la firmeza del sustrato fue un parámetro de primer orden a nivel ecológico que reguló las diferencias en las asociaciones de trazas fósiles en las localidades analizadas. Otros parámetros como la batimetría y la textura del sustrato, parecen no haber ejercido un control significativo sobre la composición de las suites atribuidas a la icnofacies de Glossifungites. Sin embargo, a estos resultados se debe incorporar la variable temporal, analizando las relaciones de corte entre las estructuras biogénicas, las diferencias en el grado de preservación de las mismas, la evaluación del tiempo que el sustrato estuvo disponible para su colonización, así como la superposición de eventos que dieron lugar a la asociación encontrada en cada uno de los afloramientos. De este modo, se puede concluir que la historia transgresiva fue compleja, con sucesivos eventos de erosión por ravinamiento y múltiples eventos de colonización de la fauna marina bentónica.
Agradecimientos
Agradecemos muy especialmente a E. Musacchio y J.C. Sciutto por sus comentarios y ayuda en el campo. También agradecemos a Eduardo Bellosi, Richard Bromley y Marcelo Reguero por sus comentarios y discusiones y a Jeremías González por su colaboración durante el trabajo de campo. Agradecemos las sugerencias aportadas por los árbitros Eduardo Olivero y Jordi de Gibert. Este estudio constituye parte del trabajo de Tesis doctoral de la Beca interna de Formación de Postgrado del CONICET de Noelia Carmona y fue financiado parcialmente por una beca otorgada por la International Association of Sedimentologists (IAS). Asimismo, durante la fase final de este estudio hemos contado con una NSERC -Discovery Grant (311727-05)- otorgada a Gabriela Mángano.
Bibliografía
Andreis, R.R. 1977. Geología del área de Cañadón Hondo, Dpto. Escalante, Provincia del Chubut, República Argentina. Revista del Museo de la Plata 4: 77-102.
Andreis, R.R., Mazzoni, M. y Spalletti, L. 1975. Estudio estratigráfico y paleoambiental de las sedimentitas terciarias entre Pico Salamanca y bahía Bustamante, provincia del Chubut. Revista de la Asociación Geológica Argentina 30: 85-103.
Bellosi, E. 1987. [Litoestratigrafía y Sedimentación del "Patagoniano" en la Cuenca del Golfo San Jorge, Terciario de Chubut y Santa Cruz. Universidad de Buenos Aires. Tesis Doctoral. 252 pp. Inédita.].
Bellosi, E. 1995. Paleogeografía y cambios ambientales de la Patagonia central durante el Terciario medio. Boletín de Informaciones Petroleras (B.I.P.). Tercera época. Año 11, 44: 50- 83.
Bellosi, E. y Genise, J. 2004. Insect trace fossils from paleosols of the Sarmiento Formation (Middle Eocene-Lower Miocene) at Gran Barranca (Chubut province). 1º International Congress on Ichnology. Fieldtrip Guidebook. Chapter 1: 15-29.
Bromley, R.G. 1996. Trace Fossils. Biology, taphonomy and applications. Chapman & Hall, Londres. 361 pp.
Bromley, R.G. y Asgaard, U. 1993. Two bioerosion icnofacies produced by early and late burial associated with sea-level change. Geologische Rundschau 82: 276-280.
Buatois, L.A., Mángano, M.G. y Aceñolaza, F.G. 2002. Trazas fósiles. Señales de comportamiento en el registro estratigráfico. Edición Especial Museo Egidio Feruglio N° 2. 382 p.
Camacho, H.H. 1974. Bioestratigrafía de las Formaciones marinas del Eoceno y Oligoceno de la Patagonia. Anales de la Academia de Ciencias Exactas, Físicas y Naturales 26: 39-57.
Camacho, H.H. y Fernández, J.A. 1956. La transgresión patagoniense en la costa atlántica entre Comodoro Rivadavia y el curso inferior del río Chubut. Revista de la Asociación Geológica Argentina 12: 23-55.
Carmona, N.B., Ponce, J.J. Mángano, M.G. y Buatois, L.A. 2004. Variability of the Glossifungites ichnofacies at the Lower Miocene boundary between the Sarmiento and Chenque Formations, Patagonia, Argentina. 1º Ichnological Congress on Trace Fossils (Trelew, 2004), Abstract Book: 23.
Domènech, R., Gibert, J.M. de y Martinell, J. 2001. Ichnological features of a marine transgression: Middle Miocene rocky shores of Tarragona, Spain. Geobios 34: 99-107.
Ekdale, A.A. y Bromley, R.G. 1991. Análisis of composite ichnofabrics: An example in uppermost Cretaceous chalk of Denmark. Palaios 6: 232-249.
Ekdale, A.A. y Bromley, R.G. 2001. Bioerosional innovation for living in carbonate hardgrounds in the Early Ordovician of Sweden. Lethaia 34: 1-12.
Expósito, E.S. 1977. [Estratigrafía del Terciario marino de Astra, provincia de Chubut. Trabajo Final de Licenciatura FCEN. Universidad de Buenos Aires. 70 pp. Inédita.].
Feruglio, E. 1949. Descripción Geológica de la Patagonia. Yacimientos Petrolíferos Fiscales. 1-3. Buenos Aires.
Frey, R.W. y Seilacher, A.1980. Uniformity in marine invertebrate ichnology. Lethaia 13: 183-207.
Frey, R.W. y Pemberton, S.G. 1984. Trace fossils Facies Models. En: R.G. Walker (ed.), Facies Models. Geoscience Canada Reprints Series, pp. 189-207.
Frey, R.W. y Pemberton, S.G. 1985. Biogenic structures in outcrops and cores. I. Approaches to ichnology. Bulletin of Canadian Petroleum Geology 33: 72-115.
Gibert J.M. de y Robles J.M. 2005. Firmground ichnofacies recording high-frequency marine flooding events (Langhian transgression, Vallès-Penedès Basin, Spain). Geologica Acta 3: 215-225.
Gibert, J.M. de, Martinell, J. y Domènech, R. 1998. Entobia ichnofacies in fossil rocky shores, Lower Pliocene, Northwestern Mediterranean. Palaios 13: 476-487.
Gingras, M.K., Pemberton, S.G. y Saunders, T. 2001. Bathymetry, sediment texture, and substrate cohesiveness; their impact on modern Glossifungites trace assemblages at Willapa Bay, Washington. Palaeogeography, Palaeoclimatology, Palaeoecology 169: 1-21.
MacEachern, J.A., Raychaudhuri, I. y Pemberton, S.G. 1992. Stratigraphic applications of the Glossifungites ichnofacies; delineating discontinuities in the rock record. En: S.G. Pemberton (ed.), Applications of ichnology to petroleum exploration; a core workshop, Society of Economic Paleontologists and Mineralogists, Core Workshop N° 16: 169-198.
Pemberton, S.G. y Frey, R.W. 1985. The Glossifungites ichnofacies: modern examples from the Georgia Coast, USA. En: H.A. Curran (ed.), Biogenic structures: their use in interpreting depositional environments. Society of Economic Paleontologists and Mineralogists. Special Publication 35: 237-259.
Pemberton, S.G., Van Wagoner, J.C. y Wach, G.D. 1992a. Ichnofacies of wave-dominated shoreline. En: S.G. Pemberton (ed.), Applications of ichnology to petroleum exploration - A core workshop. Society of Economic Paleontologists and Mineralogists, Core Workshop 17: 339-382.
Pemberton, S.G., MacEachern, J.A. y Frey, R.W. 1992b. Trace fossils facies models: environmental and allostratigraphic significance. En: R.G. Walker and N.P. James (eds.), Facies models and sea level changes. Geological Society of Canada, pp. 47-72.
Pemberton, S.G., Frey, R.W., Ranger, M.J. y MacEachern, J.A. 1992c. The conceptual framework of ichnology. En: S.G. Pemberton (ed.), Applications of ichnology to petroleum exploration - A core workshop. Society of Economic Paleontologists and Mineralogists, Core Workshop 17: 1-32.
Pemberton, S.G., MacEachern, J. y Buatois, L.A. 1997. Criterios icnológicos en el reconocimiento e interpretación de límites estatales claves. Boletín de la Sociedad Venezolana de Geólogos 22: 7- 32.
Seilacher, A. 1967. Bathymetry of trace fossils. Marine Geology 5: 413-428.
Uchman, A., Bubniak, I. y Bubniak, A. 2000. The Glossifungites ichnofacies in the area of its nomenclatural archetype, Aviv, Ukraine. Ichnos 7: 183-189. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 11 de agosto de 2004.
Aceptado: 1 de setiembre de 2005.

