










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.2 Buenos Aires abr./jun. 2006
The new genus Multispongia (Porifera) from the Lower Ordovician limestones of the Argentine Precordillera
Marcelo G. Carrera1
1 Centro de Investigaciones Paleobiológicas (CIPAL), Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Avenida Vélez Sársfield 299, 5000 Córdoba, Argentina. mcarrera@com.uncor.edu
Introduction
The sponge fauna of the Early Ordovician San Juan Formation has been the subject of several taxonomic studies (Carrera 1985, 1994, 1996a, 1996b, 2003; Beresi and Rigby, 1993), from which varied analyses focused on paleoecology and paleobiogeography have been carried out (Carrera, 1997, 2000; Carrera and Rigby, 1999, 2004). In spite of these studies, taxonomic analysis of the San Juan Formation sponges is far from complete. Every field work collection provides new specimens for study and new forms to be described. This is the case of the new genus documented in this contribution recovered from the upper levels of the San Juan Formation in the Las Aguaditas creek near Jáchal city, San Juan Province, Argentina (figure 1).
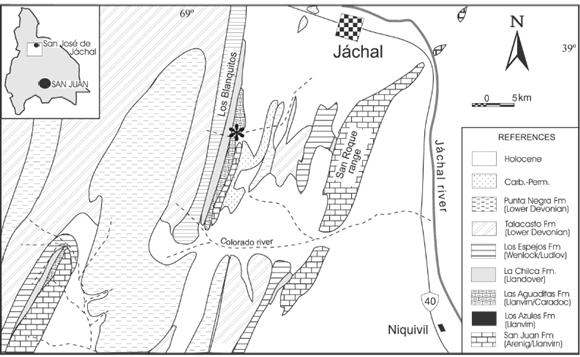
Figure 1. Geologic map showing location of the studied section, Las Aguaditas creek (asterisk) in the Los Blanquitos range / mapa Geológico y ubicación de la localidad estudiada, Quebrada Las Aguaditas (asterisco) en el cordón de Los Blanquitos.
The San Juan Formation in this locality accounts for only 25 meters of wackestones and mudstones, which include an abundant fauna mainly represented by brachiopods, sponges and trilobites. The San Juan outcrops in this section correspond to the upper levels of the unit belonging to the Ahtiella argentina brachiopod Biozone (Benedetto, 2003). These upper levels and the overlying lower part of the Las Aguaditas Formation (figure 2) correspond to the Eoplacognathus suecicus conodont biozone and Paraglossograptus tentaculatus graptolite Biozone (Brussa, 1996), which indicate a Darriwilian age.

Figure 2. Stratigraphic column and location of the sponge levels (asterisk). SJ: San Juan Formation, LA: Las Aguaditas Formation / columna estratigráfica y ubicación de los niveles con esponjas (asterisco).
This interval was included by Carrera (1997) in the "root-tuft" biofacies, the broken stalks of presumably hexactinellid sponges are clearly the dominant forms. Some such tufts were found lying directly on the limestone beds or attached in living positions. The specimen documented here came from a limestone bed five meters below the contact with the overlying black shales and it is slightly older than the root-tuft association which is found in the overlying four meters.
The complete single specimen described here is housed at the Center of Paleobiological Research (CIPAL), Universidad Nacional de C órdoba, under the repository name CEGH-UNC.
Systematic paleontology
Class DEMOSPONGEA Sollas, 1875
Order LITHISTIDA Schmidt, 1870
Suborder ORCHOCLADINA Rauff, 1895
Family ANTHASPIDELLIDAE Miller, 1889
Genus Multispongia new genus
Type species. Multispongia aspera n. sp.
Diagnosis. Compound lamellar-shaped sponge with several coalescent interconnected spongocoels, forming a thick, bulbous laminated body. Spongocoels are hollow cylinders 7 mm across in maximum diameter and 30 to 40 mm long, reaching almost to the base of the sponge. Vertical and horizontal radiating canal systems occur in each oscular pit. Vertical and radial canals interconnect with adjacent canals of neighbouring spongocoels. Radial canals are clustered around the spongocoel margin. Spicular structure, typically of the family, series of horizontally disposed monoclonid dendroclones (0.4-0.7 mm long) united by vertically arranged trabs made by monaxons in a ladder-like appearance. Skeleton opened texture with thick trabs and broadly separated monoclonids. External surface rough marked by deep grooves and emergence of the main skeletal structures.
Etymology. Multi (Lt.): numerous, beyond the regular number. Spongia (Lt.): sponge.
Discussion. Very few lower Paleozoic sponges share the main features found in Multispongia n. gen. The occurrence of several interconnected spongocoels is uncommon in lower Paleozoic forms, except for digitate or dichotomously branching forms. Several os cular areas or shallow pits with a cluster of radial canals are more frequently found. Among such forms is Perissocoelia Rigby and Webby (1988), which is a compound mushroom-shaped sponge with several oscular pits marked by well-defined clusters of excurrent canals. This genus has characteristics stalks covered with a differentiated dermal layer. Multispongia n. gen. is different in morphology; each spongocoel has its own base rather than sharing a single one. It lacks stalks and the radial canals are more irregular and ill defined. In addition, the oscular pits in Perissocoelia are considerably smaller than the relatively large spongocoels of this Argentinean sponge.
Perissocoelia sp. described by Rhebergen et al. (2001) looks more similar in external form to Multispongia n. gen. It has a more expanded form and bulbous appearance than the type species Perissocoelia habra Rigby and Webby (1988). However, Perissocoelia sp. possesses a prominent expanded stalk and the previously noted characteristics of the genus that separate it from the Argentinean sponge. The Silurian species Perissocoelia (?) gelasinina Rigby and Chatterton (1989) and Perissocoelia (?) spinosa Rigby and Chatterton (1989) were tentatively placed in the genus because they have small pits and stalked hemispherical to domical forms, but they lack the typical cluster of radial canals around oscular pits, as pointed out by the authors. All these features are clearly different from those in Multispongia n. gen.
Another somewhat similar genus is Multistella Finks (1960) in its general massive form and the occurrence of numerous excurrent canal clusters. However, these canals do not occur in deep pits but are in sharply delineated stellate clusters. Phacellopegma Gerth, 1927, as described by Finks (1960), is a massive hemispherical form with a surface covered with anastomosing deep grooves into which small circular canals open, features that clearly contrast with the interconnected spongocoels of Multispongia n. gen.
Schismospongia syltensis Rhebergen and von Hacht (2000) is a thick encrusting sponge with a basal layer that has a dense, minutely porous surface. The upper surface of this massive sponge has shallow pits to which converge clusters of long horizontal radial canals on the exterior surface. These characteristics contrast with the presence of deep spongocoels, vertical canals and short radial canals found in Multispongia n. gen.
Clusters of radial excurrent canals located in small mounds is a typical feature of the genus Anthaspidella Ulrich and Everett, 1890, as is particularly evident in A. amplia Rigby and Desrochers (1995) and A. lamellata Liu et al. (1997). However, Anthaspidella lacks prominent pits and its radiating excurrent canal system occurs on the gastral surface of a saucer-shaped sponge.
Aulocopium Oswald, 1847 may appear somewhat similar in external form, but it has only a single central spongocoel and different canal and skeletal arrangements.
Multispongia n. gen. possesses a clearly different functional morphology from that of most other early Paleozoic lithistids. Although interconnected, each spongocoel has its own aquiferous system, which is a condition for dividing the sponge into distinct functional units (see Bergquist, 1978; Fry, 1979; Wood et al. 1992, for a review). Living sponges are generally considered individuals rather than colonies (Hartman and Reiswig, 1973; Berquist, 1978). However, they can be considered as clonal organisms if the sponge body can be divided into distinct functional units. Although these units are morphologically indistinct, highly interdependent entities, they represent a form of modularity (Wood et al. 1992; Dewel, 2000). Each module (duplicate or clone) consists of a number of associated inhalant openings that each lead to a layer of choanocytes from which excurrent canals converge toward the osculum. A solitary sponge will therefore posses one osculum, and a clonal form will be multioscular.
Multispongia n. gen. displays all the essential features for being considered a clonal sponge. Each spongocoel is quite deep and clearly differentiated compared with the oscular pits found, for example, in Perissocoelia. In addition, spongocoels attached to the substrate by its own base rather than a single base in which converge a group of spongocoels. Conversely, it is not a gregarious coalescence of individuals, for their canal systems clearly interact and their skeletons are structurally bound.
Archaeocyaths are considered the oldest sponges with some degree of modularity or integration (Wood et al., 1992), but their phylogenetic position is still somewhat uncertain. Among non-calcified poriferans, Multispongia n. gen. represents one of the oldest clonal forms, and may therefore be useful in clarifying the processes that led to the acquisition of a clonal habit in sponges.
Multispongia aspera new species Figures 3.A-G

Figure 3. A-G, Multispongia aspera n. gen and sp. A, General upper view of the holotype (CEGH-UNC 21868) / vista dorsal general del holotipo. B, Detailed view of the holotype, showing adjacent oscular pits with coalescent radial canals / detalle del holotipo mostrando los ósculos adjacentes con canales coalescentes. C, Lateral view of the holotype, showing coalescent and slightly annulated spongocoels / vista lateral del holotipo mostrando esponjocelos coalescentes y ligeramente anulados. D, Longitudinal thin section of the same specimen with two interconnected spongocoels / corte delgado longitudinal de dos esponjocelos conectados. E, Ventral surface showing the radial canal system at the base of the spongocoel / superficie ventral mostrando el sistema de canales radiales en la base del esponjocelo. F, Detailed view of the spicular net in thin section, showing vertical trabs mainly composed by monaxons and connected by monoclonid dendroclones / detalle de la estructura espicular en corte delgado mostrando las columnas verticales compuestas principalmente por monaxonas y conectadas por dendroclonas monoclonidas. G, Longitudinal section across one spongocoel margin, showing divergence of trabs from the central cavity (left) to the exterior (right) / sección longitudinal del margen del esponjocelo mostrando la divergencia de las columnas desde la cavidad central (izquierda) hacia el exterior (derecha). (figures / figuras A-E: scale bars / escala gráfica 1 cm, figures / figuras F-G: scale bars / escala gráfica 1 mm).
Etymology. Aspera (Lt.): rough, referring to the coarse appearance of the external surface.
Material and occurrence. A single complete specimen, the holotype (CEGH-UNC 21868) is from the upper levels of the San Juan Formation (Darriwilian) of the Las Aguaditas section, San Juan Province.
Diagnosis. As for the genus
Description. Compound lamellar-shaped sponge with several laterally adjacent spongocoels in a thick and bulbous body. Thirteen open oscula were counted in a single specimen and range from 4 to 7 mm in diameter. Radiating external grooves emerge from the oscular margins and represent a cluster of radial canals. This feature and the gross morphology of the skeletal elements (vertical trabs) give a rough appearance to the external surface. Base of each oscular pit marked by cluster of excurrent canals. In lateral view the interconnected spongocoels outline a thick (3- 4 cm.), and irregular lamellar structure with a bulbous appearance at the oscular side. Some spongocoels appear slightly annulated, locally, in the external lateral view (figure 3.C).
Oscular margins range from 4 to 7 mm in maximum diameter, and are located on slightly elevated mounds. Circular spongocoels have moderately irregular walls, and are 4 to 7 mm in diameter near the oscular margin. They are deep, 30 to 40 mm tall, beginning almost at the base of the sponge, where diameters diminish to 3 or 4 mm.
Vertical axial canals are the largest openings of the sponge (1 to 1.5 mm). Some vertical canals terminate at the base of the spongocoel, while others begin at their base or at their margins and later bend outward at angles ranging from 30° to 40° degrees. Vertical canals are cross connected by radial horizontal canals that are 0.8 to 1 mm in diameter. These canals are short and not well defined, beginning at the spongocoel margin and ending at the exterior surface of the sponge or interconnecting with adjacent spongocels. Horizontal radial canals mark the upper surface and occur in the skeletal interior as stacked arcuate radiating canals.
Spicular network is of anthaspidellid type, with vertical trabs composed by fused monaxons (figures 3.F-G) united by monoclonid dendroclones in a ladder- like structure. Vertical trabs radiate upward and outward from a radiante near the base of the sponge. The surface of pinnation occurs at the gastral margin. Trabs are parallel to the vertical canals: straight at the base of the sponge, but curve outward from the gastral margin, and meet the dermal surface at low angles.
Trabs are irregular and complex and somewhat opened structures where vertical coring oxeas and styles, 1-1.7 mm long, are main elements and are fused with dendroclone tips (figure 3.F). Trabs are generally thick and 0.3-0.4 mm in diameter, with exceptionally thicker trabs of 0.5 to 0.6 mm occurring near the gastral surface or at points of divergence in the surface of pinnation.
Monoclonid dendroclones long and smooth, 0.4- 0.7 mm in total length. Shafts 0.2-0.5 mm long with simple and moderately expanded brachyomes and cladomes. They are irregularly spaced in the ladderlike structure, vertical distance between adjacent monoclonids ranges from 1 to 3 mm. As a whole the skeleton looks open textured with thick trabs and widely separated dendroclones.
Less frequent skeletal elements are X- shaped and Y-shaped dendroclones. Four-rayed, X-shaped, forms have moderately short shafts, 0.4-0.5 mm long, from which radiate four moderately curved rays, 0.3- 0.4 mm long. These spicules occur largely as bridging spicules at margins of canals.
Y-shaped dendroclones are approximately the same size, with shafts 0.4-0.5 mm long and proportionally long rays 0.2-0.3 mm. They commonly occur between trabs as part of the ladder-like series of dendroclones, but more frequently occur near where trabs bifurcate. Articulating clad tips are approximately 0.10 mm wide.
Skeletal structure between adjacent spongocoels is complex, with trabs limited in their growth by adjacent sets of trabs. However, neither interference nor contacts with each other were observed. Excurrent canals interconnect in a unique vertical system in the central part of the wall.
Discussion. Distinction between species of the genus, if discovered, should be based on minor skeletal features, such as trab arrangements, kind and proportions of secondary spicules or size and arrangements of canals. General distribution and form of spongocoels could also be taken into account for species differentiation.
Acknowledgements
The author wishes to thank CONICET (PEI 6551) and FONCYT grants to R. Astini (PICT 2002, 07-11741) for financial support. Appreciation is extended to J. Keith Rigby and J. Botting for their constructive reviews.
References
Benedetto, J.L. 2003. Brachiopods. In: J.L. Benedetto (ed.), Ordovician Fossils of Argentina, Secretaría de Ciencia y Técnica, Universidad Nacional de Córdoba, pp. 187-271.
Beresi, S. and Rigby, J.K. 1993. The Lower Ordovician sponges of San Juan, Argentina. Brigham Young University Geology Studies 39: 1-63.
Bergquist, P.R. 1978. Sponges. University of California Press, 268 p.
Brussa, E. 1996. Las Graptofaunas ordovícicas de las Formación Las Aguaditas, Precordillera de San Juan, Argentina. Parte I: Familias Thamnograptidae, Dichograptidae, Abrograptidae y Glossograptidae. Ameghiniana 33: 421-434.
Carrera, M.G. 1985 Descripción de algunos poríferos de la Formación San Juan (Ordovícico) Precordillera de San Juan. Reunión de Comunicaciones Paleontológicas (San Juan), Acta, pp. 41-42.
Carrera, M.G. 1994. An Ordovician sponge fauna from the San Juan Formation, Precordillera basin, western Argentina. Neues Jahrbüch für Geologie und Paläontologie (Abhandlungen) 191: 201- 220.
Carrera, M.G. 1996a. Ordovician Megamorinid Demosponges from San Juan Formation, Precordillera, Western Argentina. Geobios 29: 643-650.
Carrera, M.G. 1996b. Nuevos poríferos de la Formación San Juan (Ordovícico), Precordillera Argentina. Ameghiniana 33: 335- 342.
Carrera, M.G. 1997. Significado paleoambiental de los poríferos y Briozoos de la Formación San Juan (Ordovícico), Precordillera Argentina. Ameghiniana 34: 179-199.
Carrera, M.G. 2000. Sponge-epizoan interactions in the Early Ordovician limestones of the Argentine Precordillera. Palaios 15:261-272.
Carrera, M.G. 2003. Sponges and Bryozoans. In: J.L. Benedetto (ed.), Ordovician Fossils of Argentina, Secretaría de Ciencia y Técnica, Universidad Nacional de Córdoba, pp. 155-186.
Carrera, M.G. and Rigby, J.K. 1999. Biogeography of the Ordovician sponges. Journal of Paleontology 73: 26-37.
Carrera, M.G. and Rigby, J.K. 2004. Sponges. In: B.D. Webby, M. L. Droser, F. Paris and I.G. Percival (eds.), The great Ordovician biodiversification event, Columbia University Press, New York, pp. 102-111.
Dewel, R.A. 2000. Colonial origin for Eumetazoa: Major morphological transitions and the origin of bilaterian complexity. Journal of Morphology 243: 35-74.
Finks, R. 1960. Late Paleozoic sponge faunas from the Texas region. The siliceous sponges. American Museum of Natural History, Bulletin 120: 1-160.
Fry, W. 1979. Taxonomy, the individual and the sponge. In: G. Larwood and B.R. Rosen (eds.), Biology and Systematics of colonial organisms. Academic Press, London, pp. 49-80.
Gerth, H. 1927. Die Spongien aus dem Perm von Timor. Jahrbuch Mijnwesen Nederland-Oest Indien, verhandlung 1926: 99-132.ç
Hartman, W.D. and Reiswig, H. 1973. The individuality of sponges. In: R.S. Boardman, R.S. Cheetham and W. Oliver Jr. (eds.), Animal colonies, developments and function through time. Dowden, Hutchinson and Ross, Pennsylvania, pp. 567-584.
Liu, B.-L., Rigby, J.K., Jiang, Y.-W. and Zhu, Z.-D. 1997. Lower Ordovician lithistid sponges from the eastern Yangtze Gorge area, Hubei, China. Journal of Paleontology 71:194-207.
Miller, S.A. 1889. Class Porifera. North American Geology and Paleontology. Cinncinati, (published by the author), pp. 152- 167.
Oswald, F. 1847. Über die Petrifacten von Sadewitz. Üebersicht Arbeit Veränderungen Schlesischer Gesellschaft Vaterlandische Cultur im Jahr 1846, Breslau, pp. 56-65.
Rauff, H. 1895. Palaeospongiologie. Palaeontographica 43: 223-272. Rhebergen, F.R. and von Hacht, U. 2000. Schismospongia syltensis gen. n. sp. n. (Porifera), ein neuer Geschiebeschwamm aus Plio-/Pleistozänen Kaolinsand Deposits from the Island of Sylt. Archiv für Geschiebekunde 2: 797-804.
Rhebergen, F., Eggink, R., Koops, T. and Rhebergen, B. 2001. Ordovicische zwerfsteensponzen. Staringia 9, Nederlandse Geologische Vereniging, Groondboor and Hamer 52, 114 pp.
Rigby, J.K. and Chatterton, B.D. 1989. Middle Silurian Ludlovian and Wenlockian sponges from Baillie-Hamilton and Cornwallis islands, Artic Canada. Geological Survey of Canada, Bulletin 391: 1-69.
Rigby, J.K. and Desrochers, A. 1995. Lower and Middle Ordovician lithistid demosponges from the Mingan Islands, Gulf of St. Lawrence, Quebec, Canada. Paleontological Society, Memoir 41: 1-35.
Rigby, J.K. and Webby, B.D. 1988. Late Ordovician sponges from the Malongulli Formation of central New South Wales. Palaeontographica Americana 56:1-147.
Schmidt, O. 1870. Grundzüge einer Spongien-fauna des atlantischen Gebietes. Leipzig: 1-88.
Sollas, W.J. 1875. Sponges. Encyclopaedia Britannica, 9th edition, 451 pp.
Ulrich, E.O. and Everett, E.O. 1890. Description of Lower Silurian sponges. Bulletin of the Geological Survey of Illinois 8: 253-282.
Wood, R., Zhuravlev, A. and Debrene, F. 1992. Functional biology and Ecology of Archaeocyatha. Palaios 7: 131-156.
Recibido: 21 de octubre de 2004.
Aceptado: 11 de mayo de 2005.

