










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.2 Buenos Aires abr./jun. 2006
Cuantificación del grado de consolidación del substrato: Aplicación en análisis icnológicos
Francisco J. Rodríguez-Tovar1 y José Delgado2
1 Departamento de Estratigrafía y Paleontología, Universidad de Granada, 18002 Granada, España, fjrtovar@ugr.es
2 Departamento de Ciencias de la Tierra y del Medio Ambiente, Universidad de Alicante, 03080 Alicante, España
Introducción
La consistencia o grado de consolidación del substrato es un factor limitante mayor para la biota bentónica y, por tanto, para la producción y distribución de bioturbación. Asimismo, son conocidos los efectos de la bioturbación sobre la consistencia del sustrato (Bromley, 1996), dando lugar a cambios en la estabilidad del sedimento (de Deckere, 2003). Usualmente aquélla se caracteriza en términos cualitativos, diferenciándose varias categorías básicas (Goldring, 1995). Términos como substratos sopa (soupground), blandos (softground), firmes (firmground), duros (hardground), o vegetales (woodground), son frecuentemente utilizados, así como otros en referencia a substratos sueltos (looseground), rocosos (rockground), y de conchas (shellground). La distinción entre categorías se fundamenta en criterios como la abundancia y tipo de organismo colonizador o las características de las trazas, aunque dada la variedad de factores implicados la diferenciación es difícil, esto sumado a la existencia de límites transicionales entre sucesivas categorías (Buatois et al., 1997; Mángano et al., 1998).
En este trabajo se presenta una metodología, el test de molinete, barata y fácil de usar, que permite cuantificar la consistencia del substrato a partir del análisis de su resistencia al corte sin drenaje, y que puede ser de gran utilidad en icnología.
Resistencia al corte del terreno. El ensayo de molinete
La manifiesta relación entre el tipo de sedimento y la capacidad de bioturbación (de Deckere, 2003) es difícil de cuantificar (Bromley, 1996) dada la variedad de factores que afectan a la resistencia al corte en sedimentos cohesivos (Paterson, 1997).
La resistencia al corte de un suelo es función de su cohesión (c), ángulo de rozamiento (Ø) y esfuerzos efectivos que soporta (criterio de rotura de Mohr-Coulomb), además de la forma en que el suelo es deformado. Para deformaciones comparativamente rápidas, como la que produce un organismo cuando se desplaza sobre la superficie, el suelo se deforma sin drenaje, es decir, sin tiempo para expulsar el agua de los poros. En tal caso se denomina "resistencia al corte sin drenaje" (Su), numéricamente igual a la cohesión del suelo, independientemente del ángulo de rozamiento (Ø) (Smith y Smith, 1998).
Varios métodos cuantifican la resistencia al corte directamente sobre el substrato (Black y Paterson, 1997), diferenciándose los que no producen erosión sobre la superficie (Briggs y Richardson, 1996), incluido el test de molinete.
El equipo consta de una rueda con varias aspas y un brazo, donde ésta queda alojada (figura 1). El brazo posee una escala graduada (dial) donde se lee la resistencia del sedimento. Esta resistencia viene dada por:
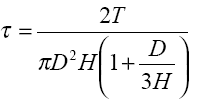
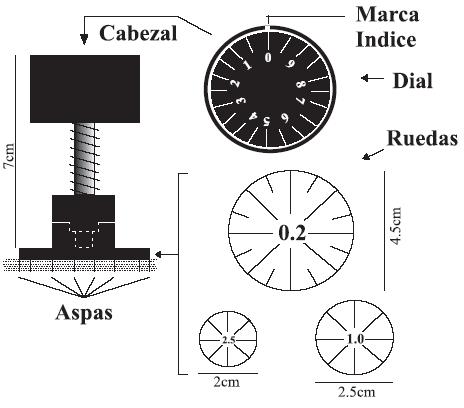
Figura 1. Dibujo esquemático del molinete. Nótese la escala graduada (dial) y los tres tipos de ruedas de aspas con radios de 2.5, 1 y 0.2 / schematic sketch of Torvane-test apparatus. Note the dial showing the index marks on the knob and the three types of vanes with factor ratios of 2.5, 1 and 0.2.
donde D y H son el diámetro de la rueda y la altura de las aspas, respectivamente.
Este test es muy común en geotecnia para determinar la resistencia Su de suelos en la superficie. Para realizarlo se requiere un área despejada, no necesariamente horizontal, de al menos 25 cm2. Existen varias ruedas de aspas para medir resistencias dentro de cierto rango de esfuerzos (figura 1). Se selecciona la rueda más apropiada a la resistencia esperada del substrato (cuanto más grande sea la rueda, más blando se supone será el terreno). El molinete se clava en el terreno hasta que las aspas quedan completamente alojadas en el suelo y, entonces, se aplica un par de fuerzas para rotar las aspas. El par se aplica con suavidad, de manera que el suelo rompa transcurridos entre 5 y 10 segundos (mayor rapidez puede inducir valores anómalamente altos). Una vez roto el suelo se lee el dial y se multiplica la cifra obtenida por la constante característica de la rueda (2,5, 1 ó 0,2 para nuestro equipo), cuantificando así la resistencia del terreno. Dado que el ensayo se realiza en pocos segundos, se obtiene la resistencia sin drenaje (Su).
Algunas ventajas del test de molinete respecto de otros métodos de cuantificación del grado de consolidación son: a) se realiza sobre la superficie, de manera que no afecta la textura o propiedades mecánicas de los sedimentos; b) puede ser repetido tantas veces como sea necesario, sin coste extra alguno; c) es comparativamente barato; d) es fácil de aplicar y e) su generalizada utilización en ingeniería permite la comparación con una importante base de datos.
Esta metodología puede ser aplicada a sedimentos no consolidados, medianamente compactos, desde substratos sopa a firmes, siendo especialmente útil en substratos blandos.
Aplicación
La relación entre actividad bentónica, bioturbación y resistencia al corte ha sido estudiada generalmente en el medio marino. En este trabajo se han seleccionado áreas en ambientes periódicamente inundados pertenecientes a medios continentales (fluvial y lacustre marginal) y marginales marinos (intertidal y estuario), colonizadas por organismos que generan distintos tipos de trazas.
En cada caso se han realizado 20 medidas, utilizando la rueda mayor (factor de escala de 0,2) e incluyendo áreas con y sin bioturbación. Nueve experimentos (IV-XII) no muestran diferencias significativas en los valores de resistencia al corte entre las áreas con y sin bioturbación; puede asumirse que los valores originales no se han modificado por la bioturbación (el experimento X necesita una investigación más detallada). En los experimentos I a III, substratos colonizados por Lumbricus terrestris, teniendo en cuenta además su etología (ver debajo), se asume que los datos originales han sido modificados por la actividad bioturbadora.
Ambientes fluviales periódicamente inundados
Se han realizado cinco experimentos (I-V), en barras laterales de los márgenes del río Genil (Granada, Sur de España), localizados en tres puntos separados aproximadamente 1 km (tabla 1 y figura 2). El área está colonizada por lombrices (Lumbricus terrestris) y gasterópodos (Lymnaea).
Tabla 1. Estadísticos básicos de la resistencia al corte obtenidos con el método del molinete en ambientes periódicamente inundados continentales (fluvial; I-V y lacustre VI, VII) y marginales marinos (intermareal VIII-X y estuario XI, XII). Todos los valores en Pascales / basic statistics of substrate firmness measurements using the Torvane test in periodically inundated nonmarine environments (fluvial; I-V and lacustrine VI, VII) and marginal marine environments (intertidal VIII-X and estuarine XI, XII). All values are given in Pascals.
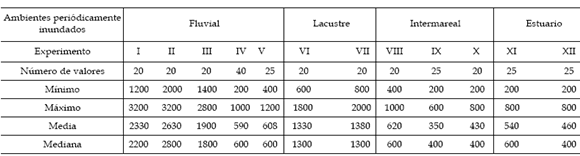
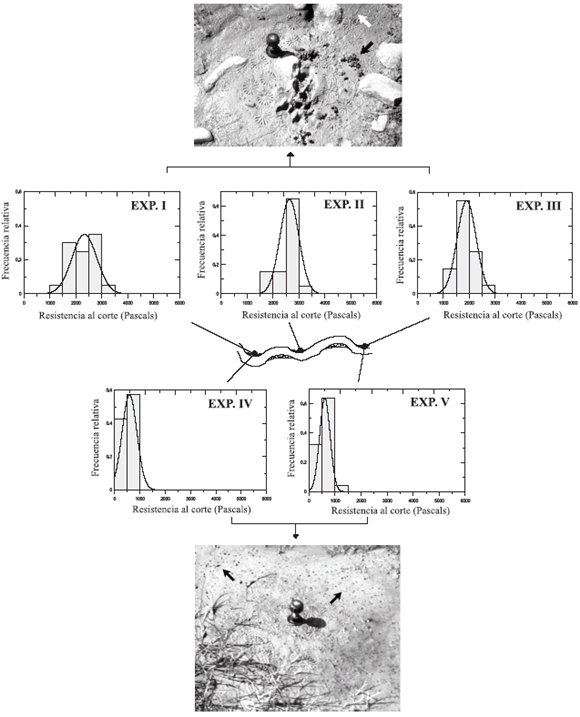
Figura 2. Experimentos en ambientes fluviales periódicamente inundados. Los gráficos superiores (I-III) corresponden a substratos bioturbados por Lumbricus terrestris (la flecha blanca en la fotografía muestra las pistas sobre la superficie y la flecha negra el sedimento expulsado), con valores medios alrededor de 2000 Pa. Los gráficos inferiores (IV y V) corresponden a substratos bioturbados por Lymnaea (las flechas negras muestran numerosos ejemplares en superficie), con valores medios cerca de 600 Pa. Nótese la marca de las aspas sobre el substrato blando / torvane experiments carried out in periodically inundated fluvial settings. Upper frequency graphs (I-III) corresponde to substrates bioturbated by Lumbricus terrestris (white arrow showing trails on the surface, and blach arrow for soil material pushed aside), with similar mean values around 2000 Pa. Lower frequency graphs (IV and V) correspond to substrates bioturbated by Lymnnaea (black arrows showing numerous specimens travelling on the surface), with similar mean values around 600 Pa. Note the star-like blade marks on the soft substrate.
Lumbricus terrestris es una lombriz de tierra, activa en superficie y en el interior del sedimento. Las excavaciones son continuas, tubulares, con aperturas en superficie rodeadas de sedimento expulsado desde el interior. Los factores principales que controlan su distribución son la disponibilidad de nutrientes y la cantidad de agua, así como la compactación del suelo (Buck et al., 2000). Su actividad puede influenciar la estabilidad del suelo (Schrader y Haiquan, 1997) y determinar modificaciones en la resistencia original. Sobre esta base, consideramos que los datos obtenidos corresponden a valores modificados respecto de los originales.
Lymnaea es un caracol pulmonado, común en ambientes de agua dulce, que se desplaza originando pistas ligeramente sinuosas, sobre substratos lodosos.
Los cinco experimentos corresponden a tres áreas de estudio (figura 2); dos (I, IV y III, V) presentan substratos bioturbados por ambos organismos, Lumbricus y Lymnaea, y la tercera (II) bioturbada sólo por Lumbricus. Los valores obtenidos permiten diferenciar dos agrupaciones: el primer grupo (I-III) con valores medios de resistencia alrededor de 2000 Pascales (Pa) (2330, 2630 y 1900 Pa), corresponde a substratos bioturbados por Lumbricus terrestris, y el segundo (IV, V) con valores medios de unos 600 Pa (590 y 608 Pa) a los bioturbados por Lymnaea (figura 2).
Ambientes lacustres periódicamente inundados
Se han analizado dos casos (VI, VII) en substratos periódicamente subaéreos de ambientes lacustres marginales del Pantano de Canales (Granada, Sur de España), con rastros de pisadas de aves. Ambos experimentos muestran valores de resistencia similares (tabla 1), con mínimos de 600 y 800 Pa, máximos de 1800 y 2000 Pa, y medias de 1330 y 1380 Pa.
Ambientes intermareales periódicamente inundados
Tres casos (VIII-X; tabla 1) han sido estudiados en substratos de áreas intermareales de la costa de Huelva (sudoeste de España, zona atlántica), cerca de los pueblos de Rompido y Cartaya, colonizados por gasterópodos (Hydrobia) y cangrejos (Uca tangeri).
Hydrobia es un caracol de aguas salobres, zonas intermareales marinas/estuario, donde origina pistas durante la locomoción y alimentación. Su distribución y abundancia se relacionan con la biomasa de microalgas, corrientes mareales, salinidad, temperatura y parámetros del sedimento como tamaño de grano, resistencia al corte y grado de consolidación (Blanchard et al., 1997).
Las actividades de excavación y alimentación del cangrejo Uca tangeri se relacionan con las características del sedimento, siendo especialmente importante los efectos de la bioturbación en su retrabajo (Wolfrath, 1992).
En el Rompido se ha realizado un experimento sobre substratos bioturbados por Hydrobia (VIII). En Cartaya se han realizado dos experimentos, el primero (IX) en substratos colonizados por Hydrobia, y el segundo (X) por Uca tangeri. Existe bastante similitud entre los experimentos, con valores medios de 620 y 350 Pa para los substratos bioturbados por Hydrobia (VIII y IX), y de 430 Pa para los bioturbados por Uca (tabla 1).
Ambientes de estuario periódicamente inundados
Se han realizado dos experimentos (XI y XII; tabla 1) en un área de estuario del río Piedras (provincia de Huelva), en substratos blandos bioturbados por bivalvos (Scrobicularia; XI) y gasterópodos (Hydrobia; XII). Scrobicularia es un bivalvo infaunal, de substratos blandos, lodosos, que origina excavaciones oblicuas en el interior del sedimento, y trazas en superficie durante la alimentación. Ambos experimentos muestran valores similares (tabla 1), mínimos de 200 Pa, máximos de 800 Pa, y medias de 540 y 460 Pa.
Discusión y perspectivas
El trabajo realizado debe considerarse como una investigación preliminar que demuestra el potencial del test de molinete para cuantificar el grado de consistencia en substratos bioturbados de medios continentales y marinos marginales. Sobre los datos obtenidos, algunas aplicaciones potenciales serían:
1. Discriminación entre y en el interior de las categorías de consistencia.
a) Una determinada categoría (especialmente aplicable a substratos blandos) podría ser caracterizada cuantitativamente y asignada a un rango de valores de resistencia al corte, de manera que se pudiera comparar y discriminar entre categorías. Los experimentos IV a XII permitirían asignar a los substratos blandos unos valores medios entre 350 Pa (IX) y 1380 Pa (VII).
b) Variaciones en el interior de una categoría podrían cuantificarse a partir de diferencias en los valores de resistencia al corte. Los experimentos IV y V (ambientes fluviales) muestran valores similares, alrededor de 600 Pa, frente a los experimentos VI y VII (ambientes lacustres), con valores medios alrededor de 1350 Pa (tabla 1).
2. Relación entre organismos colonizadores y valores de resistencia al corte. Substratos bioturbados por el mismo organismo muestran valores similares, lo que puede deberse a la influencia de la resistencia al corte sobre el organismo o bien a la modificación de la resistencia original por la actividad del organismo. a) Experimentos IV y V, en dos substratos distantes 1 km en el río Genil, bioturbados por Lymnaea, muestran valores medios similares (590 y 608 Pa) (tabla 1 y figura 2). Experimentos en substratos bioturbados por Hydrobia de ambientes marginales marinos poseen valores medios similares, ya sean áreas intermareales (VIII y IX; 620 y 350 Pa) o estuarios (XII; 460 Pa) (tabla 1).
b) Datos similares en substratos bioturbados por Lumbricus terrestris (I-III; entre 1900 y 2630 Pa; tabla 1 y figura 2), podrían relacionarse con modificaciones de los valores originales de la resistencia al corte.
3. Relación entre bioturbación y valores de resistencia al corte, siempre que se pruebe la influencia directa de la consistencia sobre la colonización. La relación entre distribución y abundancia de Hydrobia, valores de resistencia al corte y cohesión del substrato permitirían relacionar sus pistas con valores medios entre 350 y 620 Pa (VIII, IX y XII; tabla 1).
La posible relación con el grado de consistencia podría referirse a un tipo particular de comportamiento. Valores similares de resistencia al corte (entre 350 y 620 Pa) en substratos colonizados por dos diferentes gasterópodos, Lymnaea (IV, V) e Hydrobia (VIII, IX y XII), podrían relacionarse con un patrón de comportamiento parecido (organismos depositívoros de superficie) que generan pistas similares.
Posibilidades en el análisis de icnofacies
La influencia de factores ecológicos, ambientales y tafonómicos en el modelo de icnofacies arquetípicas es un hecho contrastado (Bromley y Asgaard, 1991; Pemberton et al., 1992; de Gibert y Martinell, 1998; Genise et al., 2000), de manera que la inclusión de datos que mejoren la interpretación del medio ecológico y deposicional será de gran utilidad para su caracterización. El test de molinete ofrece una aplicación potencial al aportar información sobre el grado de consistencia en relación con valores de resistencia al corte, en ejemplos actuales de icnofacies, siendo aplicable fundamentalmente a substratos blandos de ambientes continentales y marginales marinos en condiciones subaéreas.
En el medio continental, la disponibilidad de agua y su influencia en la consistencia del substrato es de gran importancia en la caracterización de icnofacies (Buatois y Mángano, 2004), especialmente las registradas en ambientes subaéreos (Buatois y Mángano, 1998; Genise et al., 2000); icnofacies de Scoyenia (ambientes como llanuras de inundación de los sistemas fluviales o depósitos marginales de lagos) y Coprinisphaera (paleosuelos de ecosistemas de comunidades herbáceas, llanuras aluviales, llanuras de inundación desecadas, etc.). Buatois y Mángano (2002), distinguen dos "suites" en las icnofacies de Scoyenia caracterizadas por diferentes estructuras relacionadas con substratos de distinto grado de consistencia; la inclusión de valores de la resistencia al corte podría mejorar esta diferenciación cualitativa.
En medios marginales marinos temporalmente subaéreos se reconocen dos principales icnofacies de substrato blando; icnofacies de Psilonichnus (ambientes transicionales entre marinos y terrestres, ambientes costeros, etc.; Frey y Pemberton, 1987) y de Skolithos (ambientes litorales inferiores a sublitorales; Pemberton et al., 1992). En estos ambientes las características del substrato (principalmente el tamaño de grano y la proporción entre lodo y arena), salinidad, temperatura, turbulencia del agua y grado de oxigenación, constituyen un control mayor en la distribución de los organismos. Datos cuantitativos de la resistencia al corte permitirán evaluar la incidencia real del grado de consistencia.
Conclusiones
Se presenta el test de molinete como un método para caracterizar el grado de consolidación del substrato a partir de la cuantificación de la resistencia al corte, de potencial utilidad en icnología. Su aplicación es especialmente recomendable en substratos blandos, bajo condiciones subaéreas permanentes o temporales. El método posee numerosas ventajas: bajo coste, fácil uso, no distorsiona las propiedades físicas del substrato y puede ser repetido tantas veces como sea necesario.
Su aplicación en ambientes continentales (fluviales y lacustres marginales) y marginales marinos (intermareales y estuarios), sobre substratos bioturbados por distintos organismos con comportamientos diversos que generan diferentes trazas, ofrece posibles líneas de avance futuras: a) caracterización y discriminación entre y en el interior de categorías de consistencia; b) cuantificación de la relación entre grado de consolidación del substrato, organismo colonizador, tipo de traza y/o comportamiento; y c) evaluación de la influencia de la actividad del organismo sobre la resistencia al corte original. El test de molinete se revela de gran utilidad para la caracterización de icnofacies en medios continentales y marginales marinos.
Agradecimientos
Esta investigación ha contado con el apoyo de los Proyectos PB97-0803, BTE2001-3029 (DGICYT), y REN2000-0798 (MCYT), y de los Grupos de Investigación RNM-178 (Junta de Andalucía) y 03/085 (Generalitat Valenciana).
Bibliografía
Black, K.S. y Paterson, D.M. 1997. Measurement of the erosion potential of cohesive, marine sediments: A review of current in situ technology. Journal of Marine Environmental Engineering 4 : 43-84.
Blanchard, G.F., Sauriau, P.G., Cariou-Le Gall, V., Gouleau, D., Garet, M.J. y Olivier, F. 1997. Kinetics of tidal resuspension of microbiota: testing the effects of sediment cohesiveness and bioturbation using flume experiments. Marine Ecology Progress Series 151: 17-25.
Briggs, K.B. y Richardson, M.D. 1996. Variability in in situ shear strength of gassy muds. Geo-Marine Letters 16: 189-195.
Bromley, R.G. 1996. Trace fossils: Biology, taphonomy and applications. Chapman y Hall. London, 361 pp.
Bromley, R.G. y Asgaard, U. 1991. Ichnofacies: A mixture of taphofacies and biofacies. Lethaia 24: 153-163.
Buatois, L.A. y Mángano, M.G. 1998. Trace fossil analysis of lacustrine facies and basins. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 367-382.
Buatois, L.A. y Mángano, M.G. 2002. Trace fossils from Carboniferous floodplain deposits in western Argentina: Implications for ichnofacies models of continental environments. Palaeogeography, Palaeoclimatology, Palaeoecology 183: 71- 86.
Buatois, L.A. y Mángano, M.G. 2004. Animal-substrate interactions in freshwater environments: applications of ichnology in facies and sequence stratigraphic analysis of fluvio-lacustrine successions. En: D. McIlroy (ed.), The Application of Ichnology to Palaeoenvironmental and Stratigraphic Analysis, Geological Society of London, Special Publication 228: 311-335.
Buatois, L.A., Jalfin, G. y Aceñolaza, F.G. 1997. Permian nonmarine invertebrate trace fossils from southern Patagonia, Argentina: Ichnologic signatures of substrate consolidation and colonization sequences. Journal of Paleontology 71: 324-336.
Buck, C., Langmaack, M. y Schrader, S. 2000. Influence of mulch and soil compaction on earthworm cast properties. Applied Soil Ecology 14: 223-229.
de Deckere, E.M.G.T. 2003. [Faunal influence on sediment stability in intertidal mudflats. NIOO Thesis 20, Rijksuniversiteit Groningen, The Netherlands, 112 p. Inédito.].
de Gibert, J.M. y Martinell, J. 1998. El modelo de icnofacies, 30 años después. Revista Española de Paleontología 13: 167-174.
Frey, R.W. y Pemberton, S.G. 1987. The Psilonichnus ichnocoenose, and its relationship to adjacent marine and nonmarine ichnocoenoses along the Georgia Coast. Bulletin of Canadian Petroleum Geology 35: 333-357.
Genise, J.F., Mángano, M.G., Buatois, L.A., Laza, J.H. y Verde, M. 2000. Insect trace fossil associations in paleosols: The Coprinisphaera ichnofacies. Palaios 15: 49-64.
Goldring, R. 1995. Organisms and the substrate: Response and effects. En: D.W.J. Bosence y P.A. Allison (eds.), Marine paleoenvironmental analysis from fossils, Geological Society of London, Special Publication 83: 151-180.
Mángano, M.G., Buatois, L.A., West, R.R. y Maples, C.G. 1998. Contrasting behavioural and feeding strategies recorded by tidal- flat bivalve trace fossils from the Upper Carboniferous of Eastern Kansas. Palaios 13: 335-351.
Paterson, D.M. 1997. Biological mediation of sediment erodibility: ecology and physical dynamics. En: N. Burt, R. Parker y J. Watts (eds.), Cohesive Sediments, John Wiley y Sons Ltd., Chichester: 215-229.
Pemberton, S.G., MacEachern, J.A. y Frey, R.W. 1992. Trace fossil facies models: Environmental and allostratigraphic significance. En: R.G. Walker y N.P. James (eds.), Facies Models: Response to Sea Level Change, Geological Association of Canada: 47-72.
Schrader, S. y Haiquan, Z. 1997. Earthworm casting: stabilization or destabilization of soil structure?. Soil Biology and Biochemistry 29: 469-475.
Smith, G.N. y Smith, I.G.N. 1998. Elements of soil mechanics, 7º ed. Blackwell Science Professional. Oxford, 494 pp.
Wolfrath, B. 1992. Burrowing of the fiddler crab Uca tangeri in the Ria Formosa in Portugal and its influence on sediment structure. Marine Ecology Progress Series 85: 237-243.
Recibido: 21 de febrero de 2005.
Aceptado: 1 de setiembre de 2005.

