










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.4 Buenos Aires sept./dic. 2006
La Familia Rzehakinidae (Foraminifera) en el Cretácico superior-Paleogeno de la cuenca Austral y la plataforma continental atlántica adyacente, Argentina
Andrea Caramés1 y Norberto Malumián1
1Consejo Nacional de Investigaciones Científicas y Técnicas, Servicio Geológico Minero Argentino, Benjamín Juan Lavaisse 1194, 1107 Buenos Aires, Argentina. postmaster@mpgeo1.gov.ar
Abstract. The Family Rzehakinidae (Foraminifera) from the upper Cretaceous-Paleogene of the Austral Basin and the adjacent Atlantic Continental shelf, Argentina. The stratigraphic and paleoenvironmental significance of the Rzehakinidae in the widespread Cretaceous-Paleogene agglutinated foraminifera assemblages of southernmost South America, particularly the Austral Basin, is reviewed. Rzehakina epigona (Rzehak), R. lata Cushman y Jarvis and transitional forms are found in the Bahía Thetis Formation (Upper Campanian-? Lower Maastrichtian); scarce specimens of R. fissistomata (Grzybowski), in the Bahía Thetis Formation, in the Maastrichtian part of the Policarpo Formation and in the Punta Torcida Formation (Lower Eocene). More delicate and small size, evolute and thin walled forms assigned to R. minima Cushman y Renz occur from the upper Campanian up to the Punta Noguera Formation (uppermost Paleocene/lowermost Eocene), becoming widespread and abundant in the "Inoceramus Superior", "Arcillas Fragmentosas" and "Senoniano" (Maastrichtian- Paleocene? subsurface beds). Few specimens of Spirosigmoilinella naibensis Turenko ? are recorded from the López de Bertodano Formation (lower Upper Maastrichtian) in the Antarctic Peninsula, the Río Claro (Upper Paleocene) and Río Bueno (lower Middle Eocene) Formations in Tierra del Fuego Island. Spirosigmoilinella compressa Matsunaga is abundant in Patagonia from the upper Oligocene up to the lower Miocene, and has its oldest record in Tierra del Fuego Island, in the lowest Oligocene, between the last ocurrences of Isthmolithus recurvus Deflandre and Subbotina angiporoides angiporoides (Hornibrook). Silicosigmoilina angusta Turenko is recorded from the Policarpo and the Río Claro Formations. Typically all these genera are mostly restricted to clastic deep settings of the Andean thrusted and folded belt. The genus Miliammina is recorded in an almost monospecific assemblage of shallow water environment from the Cerro Cazador Formation (Upper Campanian).
Resumen. Se revisa el significado estratigráfico y paleoambiental de los Rzehakinidae en las asociaciones de foraminíferos aglutinados diseminadas en el Cretácico-Paleogeno del extremo austral de Sudamérica, particularmente en la cuenca Austral. Rzehakina epigona (Rzehak), R. lata Cushman y Jarvis y formas transicionales, fueron halladas en la Formación Bahía Thetis (Campaniano superior-? Maastrichtiano inferior); y escasos ejemplares de R. fissistomata (Grzybowski) en la Formación Bahía Thetis, en niveles maastrichtianos de la Formación Policarpo y en la Formación Punta Torcida (Eoceno inferior). Formas más delicadas, de menor tamaño, evolutas y de pared más delgada, asignadas a R. minima Cushman y Renz, se registran a partir del Campaniano superior, aumentan su abundancia y distribución en las capas de subsuelo denominadas informalmente como "Inoceramus Superior", "Arcillas Fragmentosas" y "Senoniano" (Maastrichtiano-?Paleoceno); y alcanzan el entorno del límite Paleoceno/Eoceno con escasos registros en la Formación Punta Noguera. Pocos ejemplares de Spirosigmoilinella naibensis Turenko ? se registran en las Formaciones López de Bertodano (Maastrichtiano superior temprano) de la Península Antártica, y Río Claro (Paleoceno superior) y Río Bueno (Eoceno medio temprano) de la isla de Tierra del Fuego. Spirosigmoilinella compressa Matsunaga es abundante en Patagonia desde el Oligoceno superior hasta el Mioceno inferior, y tiene su registro más antiguo en la isla de Tierra del Fuego, en el Oligoceno basal, entre los últimos registros de Isthmolithus recurvus Deflandre y Subbotina angiporoides angiporoides (Hornibrook). Silicosigmoilina angusta Turenko se registra en las Formaciones Policarpo y Río Claro. Todos estos géneros están restringidos a depósitos clásticos profundos de la faja plegada y corrida andina. El género Miliammina se registra en asociaciones casi monospecíficas de ambiente marino poco profundo de la Formación Cerro Cazador (Campaniano superior).
Key words. Rzehakinidae; Upper Cretaceous; Paleogene; Argentina.
Palabras clave. Rzehakinidae; Cretácico superior; Paleogeno; Argentina.
Introducción
Las transgresiones del Cretácico tardío- Paleogeno en la cuenca Austral, incluyendo su porción costa afuera conocida como cuenca Marina Austral, y en la cuenca de Malvinas, depositaron sedimentitas de diversas facies clásticas (figura 1 y cuadro 1). Estas facies, aunque con escasos foraminíferos en asociaciones de aglutinados de muy baja diversidad, son claramente tipificables.
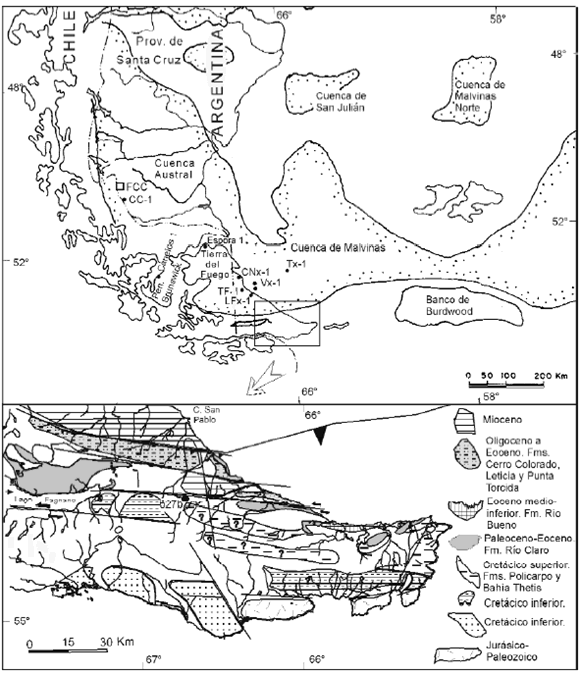
Figura 1. Localidades estudiadas / studied localities.
Cuadro 1. Distribución de los Rzehakinidae en el Cretácico superior y Paleogeno de la cuenca Austral / distribution of the Rzehakinidae in the Upper Cretaceous and Paleocene of the Austral Basin.
Las facies someras de plataforma están representadas en la Formación Cerro Cazador (Campaniano superior) por las asociaciones de Miliammina y de Reticulophragmium, intercaladas entre los niveles con Hoplitoplacenticeras y Maorites (Náñez y Malumián, 2000; Malumián y Náñez, 2002). Una asociación de muy baja diversidad y dominada por formas biseriales se reconoce desde el Santoniano cobrando su mayor extensión en el Campaniano tardío en las capas denominadas informalmente "Inoceramus Superior" y "Arcillas Fragmentosas", y en los términos superiores de la Formación Cabeza de León (Campaniano superior-Maastrichtiano, del subsuelo), resultado de un ciclo regresivo (Malumián y Náñez, 1996, 2002).
En la faja plegada y corrida de la cordillera fueguina, dentro de las facies disóxicas a anóxicas de plataforma y/o talud de las Formaciones Bahía Thetis (Campaniano superior-?Maastrichtiano inferior) y Policarpo (Maastrichtiano-Paleoceno inferior), se desarrollaron conspicuos niveles cuasi monoespecíficos con Rzehakina (cf. Olivero et al., 2003). En facies someras desconectadas del mar abierto, interpretadas como turbiditas clásicas intercaladas con areniscas masivas depositadas al pie de pendientes de abanicos deltaicos, de la Formación Punta Noguera (entorno del límite Paleoceno/Eoceno, cf. Olivero et al., 2002) se encontraron escasos ejemplares de R. minima Cushman y Renz. Las sedimentitas ricas en carbono orgánico de la Formación La Barca (Paleoceno superior), contienen microfaunas de muy baja diversidad (Malumián y Caramés, 2002) dominadas por Spiroplectammina spectabilis (Grzybowski) sin que se hayan reconocido los Rzehakinidae. En las areniscas de ambiente somero de la Formación Río Claro (Paleoceno) sólo se hallaron muy pocos ejemplares de Spirosigmoilinella naibensis Turenko ?, Silicosigmoilina angusta Turenko, S. spectabilis y de aglutinados trocoespirales indeterminables.
En facies profundas por debajo de la línea de compensación de la calcita, asociada a condiciones de corrosión (cf. Malumián y Náñez, 1991), se reconoce Spirosigmoilinella compressa Matsunaga, ampliamente distribuida en Tierra del Fuego. En toda la Patagonia, S. compressa es rara en la Formación San Julián (Oligoceno), y abundante en la base o niveles próximos a la base de la Formación Monte León (Oligoceno cuspidal-Mioceno inferior) y unidades equivalentes (cf. Malumián y Náñez, 2002). En la cuenca del Colorado costa afuera, se reconoce en paleoambientes de plataforma media e interna en el Mioceno inferior-medio inferior del pozo Estrella x-1 (Caramés y Malumián, 2000).
Los representantes de la familia Rzehakinidae, pese a haber sido reconocidos mediante menciones desde antiguo en la región sudamericana austral (Macfadyen, 1933) dentro de algunas de las asociaciones de aglutinados, no fueron objeto de estudios sistemáticos comprensivos. La marcada significación facial y cronológica dentro de la cuenca Austral y la falta de consenso en su clasificación alientan su tratamiento sistemático.
Antecedentes de rzehakinidos en el área austral
Las citas de rzehakinas se concentran mundialmente entre los 45º N y los 15º S, siendo mucho menos frecuentes en altas latitudes (véase figura 1 y cuadros 1 y 2).
Cuadro 2. Distribución estratigráfica mundial de algunos Rzehakinidae./ Worldwide stratigraphic distribution of some Rzehakinidae. (1) Tappan (1957). (2) Turenko (1983). (3) Neagu (1970). (4) Bubík (1995). (5) Hanzliková y Roth (1963). (6) Olszewska (1997). (7) Bak (2004). (8) Bubík y Kamminski (2000). (9) Rzehak (1895). (10) Charnock y Jones (1990). (11) Van den Akker et al. (2000). (12) Kuhnt y Kaminski (1997). (13) Volat et al. (1996). (14) Kuhnt et al. (1998). (15) Krasheninnikov y Pflaumann (1977). (16) Gradstein et al. (1994). (17) Miller et al. (1982). (18) Koutsoukos (2000). (19) Latgé Milward de Azevedo et al. (1987). (20) Wong (1976). (21) Malumián y Náñez (1996). (22) Olivero et al. (2003) y este estudio/ Olivero et al. (2003) and this work. (23) Duque Caro (1972 a,b). (24) Martínez R. (1989). (25) Stainforth (1955) y Weiss (1955). (26) Sigal (1970). (27) Santos y Ramírez (1986). (28) Rögl (1976). (29) Hornibrook et al. (1989). (30) Strong et al. (1995). (31) Grzybowski (1901), *¹ Corresponde a las citas originales de las especies R. occulta, R. inclusa y R. fissistomata, registradas en los estratos de la Formación Ropianka = "Inoceramus Bed", en "Magura Unit", Polonia. Geroch y Koszarski (1988) datan estos estratos con Inoceramus en el lapso Senoniano-límite Paleoceno/Eoceno, pero según estudios posteriores en perforaciones oceánicas profundas los inocerámidos desaparecen hacia la base de la Zona de Abatomphalus mayaroensis del Maastrichtiano tardío y en algunas regiones como en la Península Antártica y Tierra del Fuego, desaparecen más tempranamente hacia la base del Campaniano tardío (cf. Olivero et al., 2003) /*¹ It belongs to the original citation of the species R. occulta, R. inclusa and R. fissistomata, recorded in the beds of the Ropianka Formation = "Inoceramus Bed", in "Magura Unit", Polonia. Geroch y Koszarski (1988) assigned these beds with Inoceramus to the interval Senonian- Paleocene/Eocene boundary, but later studies in deep sea drillings suggested that the inoceramids disappeared toward the base of the Abatomphalus mayaroensis Zone in the Late Maastrichtian and in some regions as Península Antárctica and Tierra del Fuego they disappeared early toward the base of the Late Campanian (cf. Olivero et al., 2003). (32) Gawor-Biedowa (1992). (33) Szczechura y Pozaryska (1974). (34) Bak et al. (1997). (35) Olszewska y Smagowicz (1977). (36) Kaminski et al. (1988). (37) Caramés (1993). (38) Loeblich y Kniker (1950). (39) Natland et al. (1974). (40) Webb (1975). (41) Montanaro-Gallitelli (1955), *2 Bubík y Kaminski (2000) no hallaron ejemplares similares al holotipo y al no conocer otros registros mundiales dudan de la validez y relaciones con las otras especies / *2 Bubík y Kaminski (2000) did not find specimens similar to the holotype, and as they do not know other worlwide records, they doubt about the species validity. (42) Fukuta (1957). (43) Serova et al. (1980). (44) Kaminski y Geroch (1997). (45) Kaminski et al. (1990). (46) Kaminski y Austin (1999). (47) Schröeder-Adams y McNeil (1994). (48) Matsunaga (1955). (49) Hanagata et al. (2001). (50) En la Argentina se conoce a partir de la capas de La Herminita, Tierra del Fuego (Olivero y Malumián, 2005) y los registros más modernos corresponden al Mioceno inferior-Mioceno medio inferior del pozo Estrella x-1, cuenca del Colorado, plataforma continental argentina (Caramés y Malumián, 2000) / In Argentina it is known from the La Herminita beds, Tierra del Fuego (Olivero y Malumián, 2005), and the younger records correspond to the Early Miocene-early Middle Miocene of the Estrella x-1 well, Colorado Basin, Argentine continental shelf (Caramés y Malumián, 2000). (51) Bolli et al. (1994).
En el Banco Burdwood, al sur de las islas Malvinas en medio del talud continental, se tiene la primera mención de rzehakinas del área austral (Macfadyen, 1933).
En la Ant ártida, Huber (1988) registra baja a moderada abundancia de Rzehakina epigona en un paleoambiente de plataforma media a externa, en la parte inferior (unidades informales 3 y 4 de Macellari, 1988) de la Formación López de Bertodano (Maastrichtiano superior temprano, cf. Olivero y Medina, 2000) de las islas Marambio, Vega y James Ross, y la considera una especie diagnóstica de la Zona de Gaudryina healyi en la isla James Ross. La ilustración de Huber (1988) corresponde a un ejemplar mal conservado, con morfología y dimensiones más cercanas a las de Rzehakina minima.
En la porci ón chilena de la cuenca Austral, en el sector oriental, Distrito Springhill, González y colaboradores (1965), mencionan en las Lutitas Arenosas una abundante microfauna que incluye Rzehakina en las fangolitas glauconíticas del Campaniano-Maastrichtiano; Herm (1966), menciona frecuentes rzehakinas junto con Bolivinoides draco dorreeni Finlay y Bolivina incrassata Reuss, en la parte inferior, maastrichtiana, de la Zona Glauconítica, que suprayace a las Lutitas Arenosas. En la región al sur de Última Esperanza, Loeblich y Kniker (1950) ilustran R. minima en lutitas de la Formación Fuentes de los pozos Canelos nº 1 y Espora nº 1, y R. lata Cushman y Jarvis en el primero; Charrier y Lahsen (1968; 1969) asignan al Maastrichtiano tardío a los estratos superiores de la Formación Fuentes por los foraminíferos planctónicos y por B. draco dorreeni, y mencionan dos ejemplares de R. minima en limolitas y arcilitas de la Formación Chorrillo Chico, pero opinan que podrían ser retrabajados. Esta última formación fue recientemente asignada al Paleoceno medio-superior por sus dinoflagelados (Quattrocchio y Sarjeant, 2003) y la edad más antigua ha sido confirmada por la datación de circón detrítico (Hervé et al., 2004). En la región de Última Esperanza, Herm (1966) se refiere a una faúnula casi monoespecífica de Miliammina en afloramientos del Maastrichtiano medio al norte del lago Sarmiento y en el cerro Cazador. Por último, Natland et al. (1974), indican que la última aparicion de Psamminopelta minima, entre otras, definen el techo del piso Riescoiano, que se extiende por toda la cuenca Magallánica y aumenta el espesor hacia el oeste llegando a más de 3000 m en la región de Última Esperanza. El Riescoiano está representado en península Brunswick e isla Dawson por turbidítas de la Formación Rocallosa, asignada por bioestratigrafía al Campaniano superior-Maastrichtiano y cuya edad más antigua fue confirmada por datación de circón (Hervé et al., 2004). Bolivina incrassata gigantea Wicher junto con Bolivinoides draco se encuentran asociados en cuenca Austral con Nephrolithus frequens Gorka nanofósil calcáreo indicador del Maastrichtiano superior (Malumián y Náñez, 1996).
En el talud continental, en el sitio 328 en la cuenca externa de Malvinas, al este de la plataforma de Malvinas, Tjalsma (1976) cita a R. epigona (Rzehak) en una fauna de aglutinados con dominio de uniseriales y triseriales típica de ambientes abisales por debajo de la línea de compensación de carbonato de calcio; si bien la asigna al lapso Cretácico tardío-Eoceno temprano, menciona entre otros aglutinados a Uvigerinammina sp., por cuya distribución cronológica es probable que la asociación sea campaniana.
Antecedentes en la porción argentina de la cuenca Austral
Los Rzehakinidae de los trabajos mencionados a continuación forman parte del material estudiado en el presente trabajo.
En la cuenca Austral santacruceña, en el Campaniano superior de la Formación Cerro Cazador (cf. Malumián y Panza, 2000), Náñez y Malumián (2000) y Malumián y Náñez (2002), reconocen las asociaciones de Miliammina y de Reticulophragmium, y en la última mencionan un ejemplar de Psamminopelta? sp., que bien podría ser un ejemplar de R. minima muy distorsionado.
En el Campaniano inferior de la cuenca Austral costa afuera-cuenca de Malvinas y la Plataforma de Malvinas, Malumián y Náñez (1996), indican Rzehakina epigona. En el Maastrichtiano superior de la cuenca marina Austral, Caramés (1993) reconoce abundantes R. minima y Spiroplectammina spectabilis en la zona de foraminíferos arenáceos de la Formación Cabeza de León.
En Tierra del Fuego, en la Formación Bahía Thetis (Campaniano superior-?Maastrichtiano inferior), Olivero et al. (2003) hallan rzehakinas formando parte de una microfauna escasa a relativamente abundante de foraminíferos aglutinados, y en un asomo de la Formación Río Claro (Paleoceno superior), en la desembocadura del río Turbio en el lago Fagnano, Caramés (1993) menciona muy escasos ejemplares de Spirosigmoilinella naibensis Turenko y Spiroplectammina spectabilis, en una asociación muy pobre de foraminíferos aglutinados. Muestreos posteriores del último afloramiento recobraron escasos ejemplares de Silicosigmoilina angusta. En la sección tipo de la Formación Punta Torcida, Olivero y Malumián (1999), identifican como R. epigona un molde piritizado de un organismo no identificado. En niveles arenosos del Eoceno inferior, en la localidad de punta Gruesa, Jannou (comunicación verbal, junio 2005), halló un ejemplar de R. fissistomata (Grzybowski).
Spirosigmoilinella compressa es sumamente abundante y hasta dominante en las asociaciones residuales que se encuentran por debajo de la línea de compensación de la calcita en las facies profundas del Oligoceno y Mioceno inferior del fore deep de las capas de La Herminita (Olivero y Malumián, en prensa). Los registros más antiguos en la Argentina se hallan acotados en el Oligoceno inferior por debajo de la última aparición de Subbotina angiporoides angiporoides (Hornibrook) y por encima de la de Isthmolithus recurvus Deflandre que de acuerdo con las calibraciones de Berggren et al. (1995), se encuentran entre los 30 y 32 Ma.
Edad de las muestras analizadas
El miembro inferior de la Formación Río Bueno es Eoceno medio bajo (Olivero et al., 2002).
En los pozos de la cuenca Austral costa afuera, se reconocen la Zona de Gaudryina healyi y su suprayacente, la zona de foraminíferos arenáceos, ambas de la Formación Cabeza de León y referidas respectivamente al Campaniano medio-Maastrichtiano inferior y "probable maastrichtiano" (cf. Malumián, 1990).
El tramo 690-740 metros bajo boca de pozo del pozo Vega x-1, con R. minima y Spiroplectammina spectabilis, que suprayace niveles con elementos de la Zona de G. healyi, contiene exclusivamente foraminíferos arenáceos y se considera Maastrichtiano. El nivel 680 m carece de foraminíferos y sólo contiene radiolarios esféricos.
El tramo 840-900 m del pozo Cruz x-1, con abundante R. minima, se considera Maastrichtiano superior porque suprayace niveles con Notoplanulina australis Malumián y Masiuk y Ramulina sp. A de Malumián y Masiuk, característicos de la Zona de G. healyi; pero no se descarta que podría alcanzar el Paleoceno superior, por correlación con el pozo Polux x-1 que exhibe un hiato daniano seguido de facies calcáreas con los planctónicos Acarinina mckannai (White) y ?A. nicoli (Martin) (cf. Caramés, 1993). Indicios de este hiato en el Cruz x-1 son: 1) el cambio en la asociación de foraminíferos aglutinados con predominio de las formas más simples, uniseriales y biseriales, sobre las enroscadas entre 840-850 m, y a la inversa, predominio de enroscadas sobre las uniseriales o biseriales entre 860-900 m, 2) cambios en la proporción de glauconita, frecuente en 840-850 m, y muy escasa a escasa de 860-900 m, 3) brusco aumento de radiolarios en 840-850m, y de S. spectabilis que en 840 m llega a ser la especie dominante.
El tramo 1440-1470 m del pozo Cachimba Norte x-1, con escasos ejemplares de R. minima y Spirosigmoilinella naibensis?, infrayace niveles con foraminíferos del Eoceno inferior (Jannou, comunicación verbal, marzo 2005) y, aunque suprayace niveles con elementos de la Zona de Notoplanulina rakauroana (Coniaciano-Campaniano inferior, cf. Malumián, 1990), la edad más antigua del tramo se restringe al Maastrichtiano por la presencia de S. spectabilis, pudiendo alcanzar el Paleoceno superior por su abundancia y el predominio de radiolarios espumellarios. El nivel 890-900 m del pozo Tiburón x-1, con un solo ejemplar de R. minima, se asigna al Campaniano medio-Maastrichtiano inferior porque infrayace niveles maastrichtianos con Frondicularia rakauroana (Finlay) (850-860 m) y contiene elementos de la Zona de G. healyi.
El tramo 2180-2270 m del pozo Lago Fuego x-1, con R. minima y R. epigona entre la escasa microfauna compuesta casi exclusivamente por foraminíferos aglutinados, corresponde a la unidad conocida informalmente como Arcillas Fragmentosas y se considera Campaniano superior-Maastrichtiano inferior por su posición estratigráfica.
La muestra de inyección de 2960 m del pozo Cancha Carrera, con Miliammina sp., corresponde al Campaniano superior de la Formación Cerro Cazador.
El pozo Tierra del Fuego 1, contiene R. minima, en muestras de inyección de la Zona de Notoplanulina rakauroana (por debajo de 1548-1549 m), junto con Praeglobobulimina kickapoensis (Cole), Marginulinopsis sp. de Malumián y Masiuk, características de la Zona de G. healyi, y la segunda también típica de las Arcillas Fragmentosas, prismas de inocerámidos y radiolarios nassellarios en testigos y muestras de inyección del Campaniano medio-Maastrichtiano inferior (tramo 1503-1548 m), en testigos maastrichtianos con microfauna exclusiva de arenáceos (1468-1473 m), y junto con S. spectabilis en muestras de inyección de difícil datación por la gran cantidad de formas caídas de niveles superiores (entre 1447-1468 m).
La muestra de superficie 627b, Camping-Estancia La Correntina/río Irigoyen con R. minima y abundante S. spectabilis se asigna al Paleogeno inferior (cf. Olivero y Malumián, en prensa).
El tramo 1260-1620 m del Pozo Estrella x-1 con Spirosigmoilinella compressa, se asigna al Mioceno inferior- Mioceno medio inferior (cf. Caramés y Malumián, 2000).
Formación Desdémona, Capas de La Herminita, Capas del Cabo Ladrillero y Capas del Cabo San Pablo, con abundante S. compressa se refieren al Oligoceno y/o Mioceno inferior (cf. Olivero y Malumián, en prensa).
Paleoecología
El género Rzehakina es uno de los pocos constituyentes de las asociaciones de foraminíferos aglutinados de sedimentos de tipo flysch, que se depositaron en ambientes profundos de salinidad normal donde la escasez de fósiles se explica por la paleogeografía y/o paleotectónica de la cuenca. Así las especies del género pueblan paleoambientes profundos como el de los Cárpatos, donde las asociaciones están formadas casi exclusivamente por aglutinados de facies profundas, y según Bak et al. (1997), el alimento es aportado por corrientes turbidíticas distales en ambientes por debajo de la línea de disolución de carbonato de calcio, o como en el Mar del Norte, en un paleoambiente que Charnock y Jones (1990) interpretan como fuertemente relacionado con la sedimentación de turbiditas terrígenas durante tiempos de nivel del mar bajo, en ambientes batial y abisal, con alto Eh y bajo Ph y por lo tanto pobre en carbonatos. Por regla general los aglutinados de aguas profundas tienen distribución cosmopolita, sobre todo en planicies abisales (Kaminski y Geroch, 1997).
En contraste, otros géneros de rzehakinidos se encuentran en ambientes someros, como el género Miliammina que conforma asociaciones cuasi monoespecíficas en el Cretácico superior de cuenca Austral (cf. Herm, 1966; Náñez y Malumián, 2000; Malumián y Náñez, 2002) o como Psamminopelta en ambiente intertidal- deltaico y sublitoral interno y externo de Alaska (cf. Tappan, 1962). En general, los depósitos intertidales salobres tienen pocas especies de arenáceos, comúnmente conformando faunas enanas; mientras que en el ambiente costero y de plataforma, la gran turbulencia, turbidez y la rápida depositación, originan biofacies de aglutinados grandes y robustos, en general muy deformados, acompañados por raros calcáreos y muy raros radiolarios, ambos generalmente representados por moldes piritizados que sugieren rápido enterramiento (Tappan, 1962).
En el talud superior los aglutinados medianos de grano grueso son poco abundantes, en el talud medio los aglutinados son abundantes, grandes y de grano grueso, y en el talud inferior o fondo de cuenca, los aglutinados se caracterizan por ser pequeños, de grano fino y están acompañados por radiolarios. En tanto que la diversidad y equitatividad son altas en el talud superior e inferior, en el talud medio hay amplia fluctuación de los valores con los máximos y los mínimos. En la asociación tipo flysch del Mar del Norte, Rzehakina minima y Recurvoides ex gr. walteri (Grzybowski), tienen un rango paleobatimétrico restringido al fondo de la cuenca (Jones, 1988).
Bak (2004) incluye a Rzehakina en el grupo de los infaunales de ambiente somero a profundo, de hábito detritívoro.
Las rzehakinas por su morfología planoespiral, comprimida, con alta relación superficie a volumen y tamaño pequeño o mediano, predominan en ambientes anóxicos. En general, ante anoxias localizadas son frecuentes ejemplares de tamaños variables; y en anoxias extendidas se producen asociaciones de ejemplares pequeños (Bernhard,1986).
Rzehakina arca (Schröeder-Adams y McNeil), es rara en los paleoambientes de talud inferior y elevación continental del Oligoceno y Mioceno de la cuenca de Beaufort-Mackenzie (Schröeder-Adams y Mc- Neil, 1994).
Rzehakina gradsteini (Kaminski y Geroch), se registra en ambiente batial inferior-abisal de los océanos Atlántico y Tethys (Kaminski y Geroch, 1997).
Charnock y Jones (1990) incluyen a Spirosigmoilinella naibensis Turenko en su morfogrupo C2 (Jones y Charnock,1985), con hábito infaunal y capacidad para extraer los nutrientes concentrados en los primeros 5 cm de sedimento.
En la Formación Bahía Thetis, la asociación litológica fangolita-toba con abundantes diminutos fragmentos de materia orgánica carbonizada junto con la ausencia de bioturbación y relativa abundancia de Rzehakina y radiolarios esféricos, indica fondos disaeróbicos hasta anóxicos ricos en materia orgánica. Los abundantes radiolarios esféricos sugieren aguas superficiales con alta productividad. El contenido exclusivo de foraminíferos aglutinados es típico de ambiente turbidítico, y la relativa diversidad sugiere sedimentación alejada de la zona de canales submarinos donde dominan asociaciones casi monoespecíficas. La ausencia de icnofósiles es coherente con la interpretación de fondos disaeróbicos-anóxicos sugerido por la microfauna (Olivero et al., 2003).
En la Formación Policarpo, con R. fissistomata y Silicosigmoilina angusta, el dominio de sedimentos finos bioturbados sugiere ambientes tranquilos de plataforma y/o talud, la abundancia de aglutinados y escasez y conservación de los calcáreos indica ambientes turbidíticos corrosivos, la abundancia de S. spectabilis considerada infaunal detritívora junto con Gerochammina conversa (Grzybowski) pueden caracterizar ambientes con altos flujos orgánicos, y la marcada dominancia de formas biseriales como S. spectabilis indica condiciones de disoxia (Olivero et al., 2002).
Para la zona de foraminíferos aglutinados del Cruz x-1 se interpreta un ambiente de plataforma externa, con una baja tasa de sedimentación, manifestada por la abundante glauconita, que puede haber favorecido la disolución de foraminíferos calcáreos, alto pH, condiciones de disoxia marcadas por el dominio de S. spectabilis y aguas superficiales con alta productividad sugerida por la abundancia de radiolarios.
Métodología
Para diferenciar a las especies epigona, lata, fissistomata e inclusa (Grzybowski), se aplica el diagrama de Bubík (1999) que relaciona L/B vs. Z. Con L y B como largo y ancho del rectángulo mínimo donde se circunscribe el contorno lateral de la conchilla; resultando L no necesariamente paralelo al eje mediano de la conchilla, y Z como la relación porcentual entre el ancho de la depresión central limitada por las suturas espirales de las dos últimas cámaras y el ancho total de la conchilla, ambos medidos perpendicularmente al eje mediano que pasa por los dos puntos donde se unen la sutura espiral de la última y anteúltima cámaras.
Los caracteres morfológicos visibles por luz reflejada y por trasparencia se dibujaron usando como base imágenes muy pobres de los ejemplares sin metalizar obtenidas con el microscopio electrónico de barrido ambiental Phillps XL30 ESEM.
Sistemática
Los ejemplares ilustrados se depositaron en la colección de Micropaleontología de la Dirección Nacional del Servicio Geológico (DNSG 2234 y 2412 a 2428).
El material investigado de la Argentina comprende (figura 1):
Provincia de Santa Cruz: Formación Cerro Cazador (FCC) muestra de inyección del pozo Cancha Carrera 2960 mbbp.
Provincia de Tierra del Fuego: Formaciones Bahía Thetis (FBT) muestras 393-1 a 4 y 406-1 a 3 (cf. Olivero et al., 2003); Policarpo (FP) muestra 439-1d (cf. Olivero et al., 2002); Río Claro (FRC) muestra F15 (cf. Caramés, 1993) y muestra 238 equivalente a la anterior; Punta Torcida (FPT) muestra PGs-arenisca media de punta Gruesa; Río Bueno (FRB) muestra 342-3 y Punta Noguera (FPN) muestra 369 4c (cf. Olivero et al., 2002) y una muestra sin numerar, estratigráficamente más alta, próxima al punto 363 en Olivero et al. (2002); muestra de afloramiento 627b Camping- Estancia La Correntina/río Irigoyen; muestras de afloramiento de la Formación Desdémona, Capas de La Herminita, Capas del Cabo Ladrillero y Capas del Cabo San Pablo; testigos y muestras de inyección del pozo Tierra del Fuego 1 (TF-1); muestras de inyección de los pozos Lago Fuego x-1 (LF x-1) y Cachimba Norte x-1 (CN x-1).
Cuenca marina Austral: muestras de inyección de los pozos Cruz x-1 (C x-1), Vega x-1 (V x-1) y Tiburón x-1 (T x-1).
Antártida, isla Marambio: Formación López de Bertodano (FLB) LB4-2 correspondiente a la unidad sedimentaria 4, Ma18 y Ma19 de la parte inferior y media respectivamente, de la unidad sedimentaria 5 ambas de Macellari (1988).
Suborden TEXTULARIINA Delange y Hérouard, 1896 Familia RZEHAKINIDAE Cushman, 1933
Género Rzehakina Cushman, 1927, genotipo Silicina epigona Rzehak, 1895
El género Rzehakina fue propuesto para los foraminíferos arenáceos silíceos, con abertura terminal y enroscamiento planoespiral, en los que cada cámara forma media vuelta, resultando una conchilla de sección oval comprimida. Loeblich y Tappan (1987) señalan que usualmente en el género Rzehakina, las cámaras se superponen parcialmente a las de vueltas anteriores, resultando en sección transversal, una conchilla de pared mucho más engrosada en el centro de los lados. En realidad, el engrosamiento de la conchilla por superposición de cámaras, directamente relacionado con la involución de las mismas, común en Rzehakina, engrosa una parte de los lados próxima a la periferia y no exactamente al centro, como puede claramente apreciarse en la ilustración original de la especie tipo R. epigona, y en la descripción de los topotipos de Bubík y Kaminski (2000)... "con una parte fuertemente excavada en el centro de ambos lados". Estos autores al no poder hallar al único ejemplar que Rzehak designó como holotipo, proveniente de Zdounky, Moravia, propusieron una nueva localidad tipo próxima a Zdounky, investigaron la variación morfológica de R. epigona y describieron detalladamente los topotipos incluidos en las margas del Maastrichtiano superior sin designar un neotipo dado que recuperaron escasos ejemplares de pobre preservación que dificultaba visualizar las estructuras internas.
La mayoría de las especies incluidas en Rzehakina fueron descritas a comienzos del siglo XX en secuencias clásticas del entorno del límite Cretácico- Paleogeno del Flysch de los Cárpatos (Grzybowski, 1986, 1901; Friedberg, 1901; Liebus y Schubert, 1903; Jurkiewicz, 1967). Thalmann (1949) fue el primero en ponderar a la especie tipo en las correlaciones interregionales de sedimentos marinos profundos del Cretácico superior-Paleogeno. En los Cárpatos se diferenciaron varias especies como indicadores temporales. Posteriormente se comprobó la amplia distribución mundial del género y se propusieron nuevos taxones, la cantidad de formas morfológicamente cercanas indujo a cuestionar la validez de algunos y puso de manifiesto la necesidad de un criterio normalizador. Surgieron entonces distintos estudios biométricos.
El primer estudio biométrico, el del índice de crecimiento de Hanzliková (en Homola y Hanzliková, 1955), toma en cuenta la relación largo/ancho conchular. La autora, no pudo comprobar el valor taxonómico del índice por la insuficiencia de ejemplares; pero diferenció a los paleocenos con un índice cercano a 1,5 de los maastrichtianos con uno de alrededor de 1,3, en los montes Beskides (límite entre Polonia y Checoslovaquia) y en el Cáucaso.
Scott (1961) consideró el tipo de enroscamiento cameral, las relaciones largo/ancho, y el tamaño del prolóculo de las "rzehakinas" del Santoniano al Paleoceno de Nueva Zelanda. Definió cuatro tipos de generaciones: dos generaciones con enroscamiento en un plano (tipo espiroloculínido), la generación macrosférica alpha y la microsférica gamma con prolóculo mayor y menor a 50 micrones respectivamente, y dos generaciones con rotación de los planos de enroscamiento de las primeras cámaras, con un ángulo de 60-100º entre el plano de enroscamiento de las dos primeras vueltas y el de las últimas, beta, microsférica y delta, megalosférica. Y observó que: a) Las formas con rotación de los planos aparecen hacia el Cretácico terminal, pero la forma delta, megalosférica, es más rara. b) Hacia el Cretácico terminal y el Paleoceno hay una disminución progresiva de la forma microesférica gamma y un aumento de las formas con rotación de los planos de enroscamiento de las primeras cámaras. c) Todos los grupos están en los depósitos del Paleoceno inferior y luego desaparecen. Concluyó que las proporciones de crecimiento de todas las formas están dentro del rango de la variación intraespecífica y las asignó a un grupo simple. No desechó la posibilidad de que las formas beta y delta pertenezcan a un grupo genético independiente, aunque la consideró poco probable basándose en que, donde la forma beta es muy abundante la delta se presenta muy raramente, y en que tanto las formas con enroscamiento en un plano como las que cambian el plano fueron halladas juntas en depósitos del Maastrichtiano y Paleoceno.
Serova (1969) criticó duramente el criterio de Scott (1961) aduciendo que este autor eliminó especies distinguibles del género Rzehakina y desconoció la validez del género Silicosigmoilina Cushman y Church. Bubík y Kaminski (2000) consideraron que beta y especialmente delta encajan bien en la definición de Silicosigmoilina.
A juzgar por las ilustraciones de Scott (1961), coincidimos con Bubík y Kaminski (2000, p. 75), en que por su morfología general el grupo delta encaja en la definición de Silicosigmoilina; pero a diferencia de estos, consideramos que los ejemplares de las figuras 17 y 18, que Scott incluye en su grupo beta, pueden corresponder a Rzehakina, la primera con rotación de los primeros planos y la segunda a una Rzehakina donde se oculta el prolóculo como en las ilustraciones originales de Psamminopelta bowsheri.
Hiltermann (1974), al estudiar por transparencia medio centenar de ejemplares del Cretácico y Terciario inferior del Ecuador, del flysch austríaco y de los Cárpatos, remarcó que, exceptuando las primeras cámaras, el enroscamiento espiral de todas las vueltas se produce en un plano. En su búsqueda de parámetros para diferenciar las especies, definió el ángulo de crecimiento o ángulo sigmoidal, que aunque inútil para el propósito original permitió reconocer que el eje de simetría longitudinal puede ser recto o sigmoideo. Propuso un ordenamiento simple para clasificar tanto material tangible como ilustrado. Entre los varios parámetros definidos, el más útil es el valor de involución Z (véase en "Metodología"). Basándose en las mediciones de sus ejemplares y de un centenar de figuras publicadas, concluyó que R. epigona comprende cinco subespecies: inclusa, epigona, fissistomata, minima y bowsheri, cuyos valores de Z forman una serie continua creciente (0-5, 4 -24, 27-34, 35-56 y 56-76 respectivamente). Consideró que R. epigona var. lata Cushman y Jarvis entra en el rango de variación morfológica de R. epigona; y que como las rzehakinas tienen un grado de involución variable, Psamminopelta es sinónimo de Rzehakina, y bowsheri, la especie tipo de Psamminopelta, es una especie de Rzehakina totalmente evoluta. En su opinión, los rasgos morfológicos que diferencian las subespecies aparecen independientemente del tiempo y se deberían a condiciones ecológicas.
Turenko (1983) reconoce en los Rzehakinidae los mismos enroscamientos que en los miliólidos. Su diagnosis del género Rzehakina corresponde a un caparazón alargado, comprimido lateralmente, con hendiduras simétricas en los lados, y cámaras involutas a evolutas que varían la forma del contorno exterior. De manera que en corte transversal las conchillas involutas tienen forma de lanza o silla de montar; y las evolutas, sección redondeada u oval. Llamativamente, por primera vez mencionó que la abertura puede tener un diente. También, al igual que Hiltermann (1974), observó que las primeras cámaras pueden agregarse en un ángulo distinto de 180º, ya que al enroscamiento espiroloculínido (planoespiral) a veces le antecede un reducido estadio quinqueloculínido (cámaras cada 144º) o sigmoilínido (cada 154º) de dos o tres cámaras.
Bubík (1999), graficó la relación L/B versus Z (véase en "Metodología"), para un grupo de rzehakinas del flysch checoslovaco y confirmó la distinción de grupos morfológicos asignables a especies individuales.
Bubík y Kaminski (2000) aplicaron esta relación a medio centenar de ejemplares de Zdounky, localidad tipo de R. epigona. La mayoría de los ejemplares forman un agrupamiento compacto con valores de Z centrados alrededor del 10% y L/B entre 1,2 y 2. Los valores de Z del grupo de las rzehakinas de Zdounky son menores que los medidos en Checoslovaquia (Bubík 1999), y considerablemente menores que dos posibles interpretaciones de la figura original de Silicina epigona, pero consideran que sus ejemplares son subjetivamente asignables a R. epigona y que las dos interpretaciones extremas de la figura tipo posiblemente sean erróneas porque las suturas no son claramente visibles. También encontraron ejemplares únicos de R. inclusa, R. lata, R. minima y R. fissistomata, y como referencia importante incluyeron los valores obtenidos de las figuras tipo de estas especies (figura 2). Concluyen que: el grupo de R. epigona de Zdounky puede definirse biométricamente por una relación L/B entre 1,2 y 2, y Z entre 4 y 15%, aunque formas transicionales a R. lata alcanzan valores de Z de alrededor del 20%; el grupo de R. lata se diferencia por sus conchillas más anchas y valores de Z entre 20 y 30%; el grupo de R. fissistomata está delimitado por valores de Z entre 40 y 50%, y el de R. minima entre 30 y 60%. Consideran que las especies del género Rzehakina serían: Silicina epigona Rzehak, Spiroloculina fissistomata Grzybowski, S. inclusa Grzybowski, S. occulta Grzybowski, R. epigona var. lata Cushman y Jarvis, R. epigona var. minima Cushman y Renz, R. spiroloculinoides Montanaro-Gallitelli, R. sogabei Fukuta, R. uryuensis Fukuta, R. kakyineica Serova y R. macilenta Turenko; y que R. sakhalinica Turenko probablemente sea sinónima de R. fissistomata.

Figura 2. Gráfico L/B versus Z, con mediciones de los hipotipos de la figura 3, de las figuras tipo de Rzehakina lata, R. fissistomata, R. minima, R. inclusa y R. epigona según Bubík y Kaminski (2000), e indicación de la zona que ocupan la mayoría de los topotipos de R. epigona según Bubík y Kaminski (2000) en el material de Zdounky / L/B -Z diagram with measurements of the hipotypes of the figure 3, of the type figures of R. lata, R. fissistomata, R. minima, R. inclusa and R. epigona after Bubík and Kaminski (2000), and the zone of the measured set from Bubík and Kaminski (2000) with the bulk of the specimen subjectively referable to R. epigona from Zdounky.
Hiltermann (1974) y Bubík y Kaminski (2000), difieren en los valores límites de Z que separan las subespecies y especies, respectivamente. Bubík (1999) al proponer la relación L/B vs. Z, agregó otro parámetro directamente relacionado con la morfología externa de la conchilla. La utilidad de esta relación, se ejemplifica claramente con los ejemplares de R. fissistomata de cuenca Austral (véase figura 2): el ejemplar C, con Z ˜ 35%, según Hiltermann (1974) podría identificarse como R. fissistomata o R. minima; mientras que según Bubík y Kaminski (2000), debería identificarse como R. minima. Como L/B es de alrededor de 1,38, un valor muy próximo al de la figura original de R. fissistomata, se la asigna a la última clasificación. El ejemplar J, con Z ˜ 43%, siguiendo a Bubík y Kaminski (op. cit.) podría identificarse como R. fissistomata o como R. minima; y según Hiltermann (op. cit.), como R. minima. Al incorporar el índice L/B, vemos que se trata de un ejemplar mucho más ancho que la ilustración original de R. minima y sólo un poco más elongado que la original de R. fissistomata. En la cuenca Austral, por medio del diagrama de L/B vs. Z se pueden diferenciar las especies epigona, lata, fissistomata y minima (cuadro 1 y figura 2).
Una diferencia que surge de las figuras originales de Psamminopelta bowsheri, P. subcircularis y R. minima, es que en las dos primeras, la superposición de las primeras cámaras oculta el prolóculo circular, siempre visible en la última.
Los registros mundiales m ás frecuentes del género Rzehakina se circunscriben al Cenomaniano- límite Paleoceno/Eoceno, pero se conocen algunos registros en el Eoceno inferior (cuadro 2). Si las especies gradsteini y arca se incluyen en el género, la distribución se extiende al Mioceno medio.
Rzehakina epigona (Rzehak, 1895) Figuras 3.Oa,b

Figura 3. a y b, vistas laterales del mismo ejemplar con luz reflejada y transmitida / a and b, lateral views of the same specimen under rreflectted and transmitted light. Aa,b, Spirosigmoilinella compressa Matsunaga, pozo Estrella x-1. 1260 bbpm, Mioceno inferior-Mioceno medio inferior, DNSG 2234 . Ba,b, Spirosigmoilinella naibensis Turenko?, muestra 342-3, miembro inferior de la Formación Río Bueno, Eoceno medio superior, DNSG 2412. Ca,b, Rzehakina fissistomata (Grzybowski), muestra PGs-arenisca media de punta Gruesa, Formación Punta Torcida, Eoceno inferior, DNSG 2413. Da,b, Rzehakina minima Cushman y Renz, muestra 627b, Camping-Estancia La Correntina/río Irigoyen, Paleogeno inferior, DNSG 2414. Ea,b, Silicosigmoilina angusta Turenko, muestra 439-1d, Formación Policarpo, Maastrichtiano, DNSG 2415. Fa,b-Ga,b, Silicosigmoilina angusta Turenko, muestra 238, Formación Río Claro, Paleoceno superior, DNSG 2416 y 2417. Ha,b, Rzehakina minima Cushman y Renz, pozo Cruz x-1 850 mbbp, zona de foraminíferos arenáceos, Formación Cabeza de León, Maastrichtiano, DNSG 2418. Ia,b, Rzehakina minima Cushman y Renz, muestra 369-4c, Formación Punta Noguera, entorno del límite Paleoceno-Eoceno, DNSG 2419. Ja,b, Rzehakina fissistomata (Grzybowski), muestra 439-1d, Formación Policarpo, Maastrichtiano, DNSG 2420. Ka,b-La,b, Rzehakina minima Cushman y Renz, pozo Cruz x-1 870 mbbp, zona de foraminíferos arenáceos, Formación Cabeza de León, Maastrichtiano, DNSG 2421 y 2422. Ma,b, Silicosigmoilina? sp., muestra 439-1d, Formación Policarpo, Maastrichtiano, DNSG 2423. Na,b, Spirosigmoilinella naibensis Turenko?, muestra LB42, Formación López de Bertodano, Maastrichtiano superior bajo, DNSG 2424. Oa,b, Rzehakina epigona (Rzehak), muestra 406-1, Formación Bahía Thetis, Campaniano superior-? Maastrichtiano inferior, DNSG. Pa,b, Qa,b, Rzehakina lata Cushman y Jarvis, muestra 406-1, F. Bahía Thetis, Campaniano superior-? Maastrichtiano inferior, DNSG 2426 y 2427. R, Miliammina sp., pozo Cancha Carrera 2960 m, Formación Cerro Cazador, Campaniano superior, DNSG 2428.
1895. Silicina epigona Rzehak, p. 214, lám. 6, fig. 1.
Material. FBT 406-1, 5 ejemplares. LF x-1 2180-2190 mbbp, 1 ejemplar.
Rzehakina lata Cushman y Jarvis, 1928 Figuras 3.Pa,b-Qa,b
1928. Rzehakina epigona var. lata Cushman y Jarvis, p. 93, lám. 13, fig. 11a,b.
Material. FBT 406-1, 21 ejemplares; 406-2, 3 ejemplares.
Formas transicionales entre Rzehakina epigona y Rzehakina lata
Material. FBT 406-1, 4 ejemplares; 406-2, 2 ejemplares.
Rzehakina fissistomata (Grzybowski, 1901) Figuras 3.Ca,b, Ja,b
1901. Spiroloculina fissistomata Grzybowski, p. 261, lám. 7, figs. 22- 24.
Material. FBT 393-2, 2 ejemplares. FP 439-1d, 1 ejemplar. FPT PGsarenisca media de punta Gruesa, 1 ejemplar.
Rzehakina minima Cushman y Renz, 1946 Figuras 3.Da,b, Ia,b, Ha,b, Ka,b-La,b
1946. Rzehakina epigona var. minima Cushman y Renz, p. 24, lám. 3, fig. 5.
1975. Rzehakina epigona (Rzehak), Webb, p. 834, lám. 2, figs. 4-5.
Material. FPN 639-4c, 2 ejemplares. Localidad 627b Correntina/ río Irigoyen, 2 ejemplares dudosos por deformación. TF-1 1447-1589 mbbp, 36 ejemplares (testigos 1468-1469 m, 2; 1515-1519 m, 2; 1539-1543 m, 6; muestras de inyección 1447-1582 m, 30). CN x-1 1440-1470 mbbp, 5 ejemplares (1440-1450 m, 1; 1450-1460 m, 3; 1460-1470 m, 1). Cx-1 840-900 mbbp, 54 ejemplares (840 m, 29; 850 m, 5; 860 m, 2; 870 m, 5; 880 m, 4; 900 m, 7). Vx-1-1 690-740 mbbp , 8 ejemplares (690 m, 2; 710 m, 1; 740 m, 5). Tx-1 890-900 mbbp, 1 ejemplar. LFx-1 2180-2270 mbbp, 17 ejemplares (2190 m, 4; 2200- 2210 m, 2; 2220-2230 m, 7; 2260-2270 m, 4).
Observaciones. La mayoría de los trabajos (cuadro 2) indican la extinción de R. minima en el límite Paleoceno/Eoceno. El único registro en el Eoceno inferior, en la zona P6 y NP11 de los Cárpatos checoslovacos, corresponde a Bubík (1995); otro, por encima del techo de la Formación Sele, Paleoceno del Mar del Norte (Gradstein et al., 1988), no se ha incluido ya que en 1988 no estaba fijado el límite Paleoceno/ Eoceno.
Género Psamminopelta Tappan, 1957, genotipo P. bowsheri Tappan, 1957
El género Psamminopelta según la descripción original del Albiano de Alaska (Tappan, 1957) abarca formas planoespirales, en las que cada cámara forma media vuelta, superponiéndose muy ligeramente en las primeras vueltas, y se diferencia de Rzehakina porque cada cámara forma exactamente media vuelta resultando una conchilla con un eje de simetría longitudinal recto en lugar de sigmoideo. En nuestra opinión, la forma del eje de simetría no es un rasgo útil para diferenciar los géneros. Como puede apreciarse en las ilustraciones de los paratipos de P. subcircularis y P. bowsheri, las dos especies que Tappan (1957) incluyó en su género, el eje de simetría longitudinal de Psamminopelta puede ser recto o sigmoideo (casos extremos de eje longitudinal sigmoideo son los paratipos de P. bowsheri y de P. subcircularis de las figuras 12 y 8 respectivamente). Además, como señaló Hiltermann (1974) al definir el ángulo sigmoidal, en el género Rzehakina, aunque es más frecuente que el eje longitudinal sea sigmoideo, se conocen casos de eje longitudinal recto.
Según Loeblich y Tappan (1987) Psamminopelta se diferencia de Rzehakina por los lados planos y la ausencia de cualquier engrosamiento de la pared, en realidad aumento del espesor, sobre los lados planos; pero los mismos autores al referirse a Rzehakina, señalan que el engrosamiento de la conchilla por superposición de cámaras es solamente "usual". Además, aunque Loeblich y Tappan (1987) indican que Psamminopelta tiene lados planos, se han incluído en el género las especies gradsteini y arca, con engrosamiento o aumento de espesor hacia la periferia de la conchilla por el natural aumento del tamaño del lumen entre cámaras sucesivas.
Otra característica de Psamminopelta, mencionada por Tappan (1957) para diferenciar el género de Spirolocammina, es que como se ve en sección transversal, Psamminopelta es perfectamente planoespiral y carece del suave desarrollo sigmoidal en el arreglo de las cámaras. Aunque no se tiene conocimiento de que esta característica se haya utilizado para diferenciar Psamminopelta de Rzehakina, se aclara su valor parcial ya que según Turenko (1983), Rzehakina abarca formas con un reducido estadio inicial quinqueloculínido o sigmoilínido y formas completamente planoespirales.
Como vemos, la diferenciación de los géneros Psamminopelta y Rzehakina es bastante indefinida provocando disparidad de opiniones y problemas en su discriminación. Hiltermann (1974) considera que Psamminopelta es sinónimo de Rzehakina, y que bowsheri, es una especie de Rzehakina totalmente evoluta. Turenko (1983) incluye a subcircularis en Rzehakina, sin mencionar a bowsheri. Charnock y Jones (1990) observan que la conchilla evoluta de R. minima recuerda a Psamminopelta.
Originalmente han sido referidas al género Psamminopelta las especies bowsheri, subcircularis, gradsteini y arca, y a continuación analizaremos su posición genérica.
Kaminski y Geroch (1997) describen a Psamminopelta gradsteini del Eoceno medio tardío-?Mioceno temprano de los mares del Labrador y Noruega. Señalan que P. gradsteini es morfológicamente muy similar a R. minima, pero diferencian a R. minima por su prolóculo más grande y sus topotipos más anchos y espesos con lados perfectamente aplanados, y a P. gradsteini por su conchilla bicóncava, con la pared del prolóculo y de las primeras vueltas muy delgada por lo que muchos ejemplares tienen el centro roto. Nótese que estos autores, por una parte siguiendo a Loeblich y Tappan (1987), mencionan que Psamminopelta se diferencia de Rzehakina por los lados planos y ausencia de cualquier espesamiento o engrosamiento de la pared de los lados; pero por otra parte, no solo describen a P. gradsteini como bicóncava, sino que hacen referencia a topotipos de R. minima con lados perfectamente planos sin hacer ningún comentario sobre su posición genérica.
Respecto a la distribución de P. gradsteini, Kaminski y Geroch (1997) indican que se reconoce en el Eoceno medio superior-Oligoceno inferior del Mar del Labrador (Kaminski et al., 1989) y en el Eoceno superior-Oligoceno del Mar de Noruega (Kaminski et al., 1990), y que fue identificada como Spirosigmoilinella compressa por Miller et al. (1982) en el Eoceno medio superior-Eoceno superior del Mar de Labrador y por Ostermann y Spiegler (1996) en el Mioceno del Atlántico Norte. Indican que P. gradsteini puede haber sido observada para el Paleoceno de los Cárpatos polacos, ya que la ilustración de Ammodiscus polygyrus (abnormal) (Grzybowski, 1901) recuerda a su especie, pero no pueden asegurar su sinonimia porque el ejemplar no se preservó en la colección. También aclaran que Kaminski en Kaminski et al. (1989) creyó que en el Mar de Labrador esta forma representaba una de las generaciones de S. compressa, pero que en un examen más detallado del material de los mares de Labrador y Noruega-Groenlandia no hallaron ningún ejemplar con enroscamiento inicial espirosigmoilínido.
Psamminopelta arca Schröder-Adams y McNeil (1994), del Oligoceno-Mioceno medio, Canadá ártico, según su descripción original, se diferencia de otras especies del género por la conchilla más espesa, de sección lateral redondeada en forma de llama y fundamentalmente, por sus cámaras tubulares comprimidas por el canto más que por los lados y con un surco longitudinal en el canto. Como señalan Schröeder-Adams y McNeil (1994), P. arca posee la última cámara (en realidad el último par de cámaras) mucho más grande que las anteriores, por lo tanto, según puede apreciarse en las ilustraciones originales, tampoco posee lados planos sino una depresión central amplia y profunda.
Dados estos antecedentes, no sólo existen problemas en la diferenciación de los géneros Rzehakina y Psamminopelta, sino también en la caracterización de Psamminopelta. Parecería que, más que la ausencia de engrosamiento de la conchilla, se restringiría Psamminopelta para aquellas conchillas en las que el engrosamiento hacia la periferia se debe únicamente al natural aumento del tamaño del lumen entre cámaras sucesivas, dejando el engrosamiento por superposición de las cámaras como un carácter exclusivo de Rzehakina. Sin embargo, no todas las rzehakinas engrosan la conchilla por superposición de cámaras, y por consiguiente el engrosamiento por superposición carece de valor para diferenciar los géneros.
Rzehakina minima aparentemente no presenta engrosamiento por involución de las últimas cámaras y el escaso engrosamiento hacia la periferia se debería al aumento del tamaño del lumen de cámaras sucesivas. En la descripción e ilustración original de la especie de Trinidad y en ilustraciones de Kaminski et al. (1988), que corresponden a vistas laterales de dos topotipos, uno con luz reflejada y el otro con transmitida, no se aporta información que permita conocer el engrosamiento de la conchilla y su espesor. La descripción original, sólo señala que la conchilla es muy comprimida, planteando dudas sobre su posición genérica. Sin embargo, que según las mediciones de Rzehakina de Hiltermann (1974) el grado de involución es gradual y variable entre formas totalmente evolutas y totalmente involutas, y que Bubik y Kaminski (2000) encontraron juntos ejemplares asignables a R. minima y a otras especies de Rzehakina en la localidad tipo de R. epigona, son argumentos a favor de mantener a minima en Rzehakina y de que en ésta, el engrosamiento de la conchilla no siempre se debe a la involución de las cámaras y por lo tanto la involución no siempre tendría valor en la diferenciación de los géneros Rzehakina y Psamminopelta.
Tappan (1957) indicó que Psamminopelta posee prolóculo y cámaras tubulares con enroscamiento planoespiral, pero en todas las ilustraciones del genotipo y de P. subcircularis, que corresponden a vistas con luz reflejada, no se aprecia ninguna insinuación del prolóculo sino una cámara tubular que aparentemente se le superpone. El prolóculo oculto podría ser característico de Psamminopelta y de ser así, Psamminopelta tendría dos especies, bowsheri y subcircularis, en tanto que gradsteini y arca se incluirían mejor en Rzehakina; sin embargo no es una característica útil para diferenciar Psamminopelta de Rzehakina, ya que según Turenko (1983), en Rzehakina al enroscamiento planoespiral a veces le antecede un reducido estadio quinqueloculínido o sigmoilínido de dos o tres cámaras y el prolóculo queda oculto, y nuevamente este rasgo (prolóculo oculto) tendría un valor parcial en la diferenciación de ambos géneros.
Es probable que no exista diferencia morfológica que justifique separar a minima y gradsteini. Según señalan Kaminski y Geroch (1997), son morfológicamente muy similares; pero los topotipos de R. minima son más anchos y espesos con lados perfectamente aplanados. Si bien es cierto que tanto el holotipo de R. minima como los dos ejemplares que ilustran Kaminski et al. (1988) tienen una conchilla más ancha que la de P. gradsteini, la mayoría de los ejemplares de la cuenca Austral presentan proporciones más próximas a las de P. gradsteini, sólo unos pocos tienen una relación L/A similar a la de la figura del holotipo de R. minima y otros tienen proporciones intermedias, algunos ejemplares poseen conchilla plana y otros ligeramente cóncava. Además, el concepto de la especie minima de Kaminski no es tan rígido en cuanto al ancho de la conchilla, así en Bubik y Kaminski (2000) expresa que el grupo gamma de Scott (1961) se compara bien con R. minima y especialmente la fig. 16 de Scott con L= 0,48 mm y A= 0,225 mm, corresponde a un ejemplar bastante más elongado que las ilustraciones de R. minima de Trinidad, y en Gradstein et al. (1994) ilustra tres ejemplares elongados con proporciones similares a las de algunas R. minima de la cuenca Austral o las de P. gradsteini. De confirmarse la sinonimia entre gradsteini y minima, habría una ausencia de registros en el Eoceno temprano tardío y medio inferior, pero esta ausencia bien podría deberse a su rareza o conservación.
La discontinuidad del registro fósil entre las especies bowsheri y subcircularis por un lado y gradsteini y arca por otro (véase cuadro 2), y los contrastantes requerimientos paleoambientales, las primeras de ambiente somero y las últimas de ambiente profundo, son otros argumentos para considerar que las dos últimas tendrían mejor ubicación en Rzehakina.
Respecto a la validez del género Psamminopelta, para las especies bowsheri y subcircularis se necesita un examen minucioso del tipo de enroscamiento e involución de las primeras cámaras de los holotipos.
Según antecedentes la distribución estratigráfica del género Psamminopelta es Albiano y Eoceno medio- límite Mioceno medio/superior; en caso de comprobarse la validez de Psamminopelta, en nuestra opinión se restringiría al Albiano.
Género Spirosigmoilinella Matsunaga, 1955, genotipo S. compressa Matsunaga, 1955
El género Spirosigmoilinella, fue propuesto para los foraminíferos arenáceos silíceos, con abertura terminal sin diente en el extremo de un cuello corto, en los que cada cámara forma media vuelta; en las cámaras del estadio temprano el plano de enroscamiento forma una curva sigmoidea suave en sección transversal, las cámaras siguientes se agregan en un único plano de enroscamiento. Según Turenko (1983), el género puede o no tener diente, en las formas macrosféricas las primeras cámaras tienen arreglo sigmoilínido; en las microsféricas, quinqueloculínido. Las cámaras tardías de ambas formas se disponen de manera spiroloculínida.
Se conocen pocas especies del género Spirosigmoilinella, a diferencia del género Rzehakina con varias especies frecuentemente citadas, y exceptuando a la especie tipo, del resto los muy pocos registros son de distribución geográfica restringida. Spirosigmoilinella compressa, se reconoce en forma confiable desde el Eoceno superior, y es muy abundante y frecuente en sedimentitas de ambientes profundos a partir el Oligoceno.
Un conjunto de especies asignadas al género fueron originalmente descritas por Turenko (1983) en Sakhalin, Rusia: S. gorbushensis, Coniaciano-Campaniano; S. manuensis, S. holmskensis y S. naibesis, Campaniano superior-Daniano. La última reconocida además en el Campaniano-Paleoceno del Mar del Norte (Charnock y Jones, 1990). Silicomassilina sinegorica Serova (1966), genotipo de Silicomassilina, del Daniano-Paleoceno de Sakhalin, Rusia, con un prolóculo redondeado seguido por 5-6 cámaras con arreglo quinqueloculínido y luego cámaras en un único plano tipo Spiroloculina (cf. Loeblich y Tappan, 1987), según Turenko (op. cit.) sería otra especie de Spirosigmoilinella. La discontinuidad de registros entre estas especies y S. compressa, plantea la posibilidad de que este conjunto conforme un género diferente.
La distribución estratigráfica mundial de Spirosigmoilinella se restringe al Coniaciano-Paleoceno y Eoceno superior-Mioceno cuspidal (cuadro 2). Este trabajo reconoce el género en el Eoceno medio inferior de la Formación Río Bueno.
Spirosigmoilinella naibensis Turenko, 1983 ? Figuras 3.Ba,b, Na,b
? 1983. Spirosigmoilinella naibensis Turenko, p. 49, lám. 2, figs. 3,4.
Material. FRC 15, 1 ejemplar roto. FRB 342-3, 1 ejemplar. FLB LB42, 1 ejemplar; Ma18, 1 ejemplar; Ma19, 1 ejemplar. CNx-1 1440- 1450 mbbp, 1 ejemplar.
Observaciones. Los ejemplares pueden corresponder a esta especie, pero la gran similitud con S. compressa pone de manifiesto la necesidad de comparar material topotípico. Según Turenko (1983) S. naibensis se diferencia de S. compressa por la abertura triangular redondeada y dentada, y suturas menos marcadas. En nuestro material la pobre conservación impide ver las características de la abertura.
Hasta el momento, los registros seguros más antiguos de S. compressa corresponden al límite Eoceno medio/superior (aunque Charnock y Jones,1990, en su cuadro de distribución, indican su presencia con línea punteada en la parte superior del Eoceno medio del Mar del Norte) y los registros más jóvenes de S. naibensis al Paleoceno superior (cf. Charnock y Jones, 1990). Probablemente la discontinuidad en el registro (cuadro 2) desalentó su comparación e hizo que no se percibiera la similitud entre ambas. En cambio se compararon S. naibensis y Rzehakina minima, ambas concurrentes desde el Campaniano inferior al Paleoceno superior.
El hallazgo de un ejemplar de Spirosigmoilinella en una muestra de afloramiento de la Formación Río Bueno, datada como eocena media inferior (cf. Olivero et al., 2002), de gran similitud con el ejemplar maastrichtiano de la Formación López de Bertodano, con el ejemplar del Paleoceno superior de la Formación Río Claro, y con numerosos ejemplares de S. compressa (incluso comparado con 11 topotipos de la Formación Nanatani), plantea una posible relación entre ambas. Por otra parte, tanto S. compressa como S. naibensis, aunque más frecuentes en ambientes de flysch con típicas asociaciones de aglutinados, también se registran en asociaciones con foraminíferos calcáreos.
Spirosigmoilinella compressa Matsunaga, 1955 Figuras 3.Aa,b
1955. Spirosigmoilinella compressa Matsunaga, p. 50, fig. texto 1.
Material. Formación Desdémona, Capas de La Herminita, Capas del Cabo Ladrillero y Capas del Cabo San Pablo, más de 500 ejemplares. Pozo Estrella x-1, 1260-1620 mbbp, 25 ejemplares. Topotipos de la Formación Nanatani, 11.
Observaciones. Probablemente sean sinónimas de S. compressa, Psamminopelta venezuelana Hedberg (1937) descrita para el Oligoceno-Mioceno medio de la Formación Carapita y Rzehakina advena Bermúdez y Fuenmayor (1966) del Mioceno medio de estratos equivalentes a la Formación Cubagua, ambas de Venezuela.
Género Silicosigmoilina Cushman y Church, 1929, genotipo Silicosigmoilina californica Cushman y Church, 1929
El género Silicosigmoilina, según Loeblich y Tappan (1987), posee un prolóculo seguido por cámaras elongadas y angostas que se agregan con un ángulo algo inferior a 180º, que en sección transversal forma una serie sigmoidea, y abertura circular a oval con un diente simple. Según Turenko (1983), el género tiene un caparazón con aspecto de "ovillo" con enroscamiento sigmoilínido y este tipo de disposición se presenta luego de una cámara macrosférica o de dos o tres cámaras quinqueloculínidas microsféricas.
Se conocen las siguientes especies: S. californica Cushman y Church, Cretácico superior y Paleoceno y S. perplexa Israelesky, Paleoceno, ambas de California; S. ezoensis (Takayanagi), Santoniano-Campaniano superior; S. futabaensis Asano, Senoniano; S. futabaensis tokachiensis Yoshida, S. kushiroensis Yoshida y S. akkeshiensis Yoshida del Maastrichtiano, todas del Japón. Silicosigmoilina compacta Serova, Daniano-Paleoceno de Rusia; S. elegantissima Serova, Paleoceno de Kamchatka. S. seimensis y S. explicata, Campaniano superior- Daniano; S. angusta, Santoniano-Campaniano inferior y S. convexa, Campaniano inferior, todas descritas por Turenko en Sakhalin. Rzehakina sogabei Fukuta del Campaniano de Japón, es considerada una Rzehakina según Bubík y Kaminski (2000), pero Turenko (1983) la incluye en el género Silicosigmoilina. Según Kaiho (1992), S. futabaensis tokachiensis, S. kushiroensis y S. akkeshiensis, son sinónimas de S. californica que es una especie dominante del Maastrichtiano al Eoceno inferior del área de Hokaido central, Japón.
La distribución de Silicosigmoilina se extiende del Santoniano al Paleoceno.
Silicosigmoilina angusta Turenko, 1983 Figuras 3.Ea,b, Fa,b-Ga,b
1983. Silicosigmoilina angusta Turenko, p. 45, lám. 1, figs. 5,6.
Material. FP 439 1d, 1 ejemplar. FRC F15, 1 ejemplar perdido; 238, 2 ejemplares.
Observaciones. Nuestros ejemplares son de pequeñas dimensiones relativas respecto a los tipos del Santoniano-Campaniano inferior de Sakhalin de Turenko (0,26-0,27 mm vs. 0,50-0,56 mm), y aunque también son más pequeños, su tamaño es más próximo al de los escasos ejemplares del Maastrichtiano temprano-Paleoceno temprano de Trinidad de Bolli et al. (1994) (0,33-0,55 mm).
En los pocos ejemplares disponibles se ve claramente que el último par de cámaras se dispone en un ángulo inferior a los 180°. Sin embargo estas características se constatan también en ejemplares juveniles en el abundante material de Spirosigmoilinella compressa, poniendo en duda al género asignado.
Silicosigmoilina? sp. Figuras 3.Ma,b
Material. FP 439-1d, 2 ejemplares.
Observaciones. Los dos ejemplares presentan enroscamiento sigmoilínido, pero el prolóculo visible, hace sospechar que se trate de ejemplares de R. minima muy deformados. Por otra parte, como no se hallaron ejemplares de la última especie en la Formación Policarpo se opta por la presente clasificación.
Género Miliammina Heron-Allen y Earland, 1930, genotipo Miliolina oblonga (Montagu) var. arenacea Chapman, 1916
El género Miliammina, posee conchilla oval elongada con cámaras angostas con arreglo quinqueloculínido, pared relativamente espesa, aglutinada muy fina sobre una base orgánica e insoluble en ácido, y abertura terminal redondeada o semilunada dependiendo del grado de compresión entre las vueltas, que puede estar en un cuello.
Loeblich y Tappan (1987) incluyen a Miliammina en la Familia Rzehakinidae del Suborden Textulariina; en cambio Haynes (1981) y Fahrni et al. (1997) lo consideran un miliólido.
Distribución. Cretácico inferior-Holoceno.
Miliammina sp. Figura 3.Ra
Material. CC 2960 mbbp, 2 ejemplares (1 perdido).
Discusión y conclusiones
En la cuenca Austral, dentro de las asociaciones de foraminíferos aglutinados, aparentemente las microfaunas con Uvigerinammina del Santoniano tardío- Campaniano temprano, son reemplazadas por otras formadas casi exclusivamente por Rzehakina en el Campaniano tardío. A partir de esta edad sucede un relevo de especies.
Las de conchillas más robustas, R. lata y R. epigona, mundialmente están bien representadas en el Cretácico tardío, y permanecen hasta el límite Paleoceno/Eoceno y el Eoceno temprano, respectivamente. En cuenca Austral son dominantes en la Formación Thetis y en las Arcillas Fragmentosas (Campaniano superior-Maastrichtiano inferior) desapareciendo luego drásticamente.
Las conchillas más delicadas de R. minima, reconocidas mundialmente desde el Campaniano y que llegan con un sólo caso al Eoceno inferior (zona P6, Bubík, 1995) en los Cárpatos checoslovacos, tienen en cuenca Austral algunos escasos registros dudosos, por encontrarse en muestras de inyección, desde la Zonas de Notoplanulina rakauroana y Gaudryina healyi (Coniaciano-Campaniano inferior y Campaniano medio-Maastrichtiano inferior); luego aumentan su abundancia en testigos corona llegando a ser una especie característica de la zona de foraminíferos arenáceos (Maastrichtiano-?Paleoceno), y es escasa en la Formación Punta Noguera.
Rzehakina fissistomata, mundialmente reconocida en rocas pre-eocenas, es una forma escasa en cuenca Austral en el Campaniano superior-Maastrichtiano de las Formaciones Bahía Thetis y Policarpo y en el Eoceno inferior de la Formación Punta Torcida.
La Formación Punta Noguera, fue asignada al entorno del límite Paleoceno/Eoceno, por contener foraminíferos bentónicos, tanto formas consideradas de edad no más joven que el Paleoceno, como de los géneros Elphidium y Cribrorotalia, típicamente considerados post-paleocenos. El hallazgo de R. minima, limita su menor edad a la zona P6, cuyo techo ha sido calibrado en 52,3 Ma (cf. Berggren et al., 1995).
La coexistencia de los géneros Rzehakina y Spirosigmoilinella, mencionada en el Mar del Norte por Charnock y Jones (1990) y que sugiere una relación entre ambos, se verificó sólo con un ejemplar de cada género en el pozo CN x-1, pero en material chileno de la Formación Rocallosa, junto con una docena de ejemplares de R. minima se encontraron el doble de ejemplares de S. naibensis?
Dada la semejanza entre Spirosigmoilinella compressa y S. naibensis, y la pobre preservación de nuestro material que impide reconocer la existencia del diente que caracteriza a la segunda especie, se la identifica con dudas en las Formaciones López de Bertodano y Río Claro, bastante más antiguo que los primeros registros mundiales de S. compressa en el límite Eoceno medio/superior. El hallazgo de un ejemplar en el Eoceno medio inferior de la Formación Río Bueno, temporalmente más próximo a los primeros registros de S. compressa, podría indicar una relación filogenética o sinonímica entre ambas especies y se extendería el biocrón de S. naibensis a niveles más jóvenes, o el de S. compressa a niveles más antiguos.
Se identificaron como Silicosigmoilina angusta cuatro ejemplares regularmente conservados, tres de los cuales provienen de la Formación Río Claro donde también se halló el género Spirosigmoilinella. Teniendo en cuenta que entre el abundante material de Spirosigmoilinella compressa del Oligoceno- Mioceno inferior, se hallaron algunos ejemplares, aparentemente coespecíficos, con el último par de cámaras dispuestas en un ángulo inferior a los 180°, no se descarta que los primeros correspondan a formas juveniles del género Spirosigmoilinella.
La fuerte disimilitud entre el ambiente somero habitado por Miliammina y el profundo habitado por Rzehakina, soportaría la sistemática de Haynes (1981) y los estudios moleculares (Fahrni et al., 1997) que consideran al primero perteneciente a los Miliolida.
Agradecimientos
Se agradece a E. Olivero muestras de las Formaciones López de Bertodano y Río Claro, a G. Jannou muestra y ejemplares de Rzehakina de la Formación Punta Torcida, a A. Concheyro muestras de la Formación López de Bertodano, y a D. Boltovskoy y P. Kress las traducciones de bibliografía rusa y alemana, respectivamente. Es un aporte del Proyecto PICT 2002 N°12257- Cenozoico de la Plataforma Continental Argentina y Patagonia.
Bibliografía
Bak, K. 2004. Deep-water agglutinated foraminiferal changes across the Cretaceous/Tertiary and Paleocene/Eocene transitions in the deep flysch environments; eastern Outer Carpathians (Bieszczady Mts., Poland). En: M. Bubík y M.A. Kaminski, (eds.), Proceedings of the Sixth International Workshop on Agglutinated Foraminifera, Grzybowski Foundation Special Publication 8: 1-56. [ Links ]
Bak, K., Bak, M., Geroch, S. y Manacki, M. 1997. Biostratigraphy and Paleoenvironmental analysis of benthic foraminifera and radiolarians in Paleogene variegated shales in the Skole Unit, Polish Flysch Carpathians. Annales Societatis Geologorum Poloniae 67: 135-154. [ Links ]
Berggren, W.A., Kent, D.V., Swisher III, C.C. y Aubry, M.-P. 1995. A revisal Cenozoic geochronology and chronostratigraphy. Society for Sedimentary Geology, Special Publication 54: 129-212. [ Links ]
Bermúdez, P.J. y Fuenmayor, A.N. 1966. Consideraciones sobre los sedimentos del Mioceno medio al Reciente de las costas central y oriental de Venezuela. Segunda parte. Los foraminíferos bentónicos. Boletín de Geología, Venezuela 7: 413-611. [ Links ]
Bernhard, J.M. 1986. Characteristic assemblage and morphologies of benthic foraminifera from anoxic, organic-rich deposits: Jurassic through Holocene. Journal of Foraminiferal Research 16: 207-215. [ Links ]
Bolli, H.M., Beckmann, J-P. y Saunders, J.B. 1994. Benthic foraminiferal biostratigraphy of the south Caribbean region. Cambridge University Press, Cambridge, 408 pp. [ Links ]
Bubík, M. 1995. Cretaceous to Paleogene agglutinated foraminifera of the Bílé Karpaty unit (West Carpathians, Czech Republic). En: M.A. Kaminski, S. Geroch y M.A. Gasinski (eds.), Proceedings of the Fourth International Workshop on Agglutinated Foraminifera, Grzybowski Foundation Special Publication 3: 71-116. [ Links ]
Bubík, M. 1999. Results of taxonomic research on Cretaceous- Paleogene flysch-type agglutinated foraminifera. Geologica Carpatica 50, Special Issue (International Geological Conference "Carpathian Geology 2000", Smolonice, Slovak Republic, 1999): 17-19. [ Links ]
Bubík, M. y Kaminski, M.A. 2000. Remarks on the type locality and current status of the foraminiferal specie Rzehakina epigona (Rzehak, 1895). En: M.B. Hart, M.A. Kaminski M.A. y C.W. Smart (eds.), Proceedings of the Fifth International Workshop on Agglutinated Foraminifera, Grzybowski Foundation Special Publication 7: 71-80. [ Links ]
Caramés, A.A. 1993.[Foraminíferos y bioestratigrafía del Cretácico superior y Terciario inferior del sur de Argentina. Universidad de Buenos Aires. Tesis doctoral, 1: 208 pp., 2: 182 pp. Inédito. [ Links ]].
Caramés, A. y Malumián, N. 2000. Foraminíferos bentónicos del Cenozoico del pozo Estrella x-1, cuenca del Colorado, plataforma continental Argentina. Ameghiniana 37: 387-419. [ Links ]
Chapman, F. 1916. Report on the foraminifera and ostracoda out of marine muds from soundings in the Ross Sea. Reports of Scientific Investigations, British Antarctic Expedition 1907-1909, Geology 2: 53-80. [ Links ]
Charnock, M.A., y Jones, R.W. 1990. Agglutinated foraminifera from the Paleogene of the North Sea. En: C. Hemleben, M.A. Kaminski, W. Kuhnt y D.B. Scott (eds.), Paleoecology, Biostratigraphy, Paleoceanography and Taxonomy of Agglutinated Foraminifera, Kluwer Academic Publishers, pp. 139-244. [ Links ]
Charrier, R. y Lahsen, A. 1968. Contribution a l'étude de la limite Crétacé-Tertiaire de la Province de Magellan, extréme-sud du Chili. Revue de Micropaléontologie 2: 111-120. [ Links ]
Charrier, R. y Lahsen, A. 1969. Stratigraphy of Late Cretaceous- Early Eocene, Seno Skyring-Strait of Magellan Area, Magallanes Province, Chile. American Association Petroleum Geologists Bulletin 53: 568-590. [ Links ]
Cushman, J.A. 1927. An outline of a re-classification of the foraminifera. Contributions from the Cushman Laboratory for Foraminiferal Research 1: 1-105. [ Links ]
Cushman, J.A. 1933. Foraminifera: their classification and economic use. Special Publications Cushman Laboratory for Foraminiferal Research 4: 1-349. [ Links ]
Cushman, J.A. y Church, C. 1929. Some Upper Cretaceous Foraminifera from near Coalinga, California. Proceedings of the California Academy of Sciences, ser. 4, 18: 497-530. [ Links ]
Cushman, J.A. y Jarvis, P.W. 1928. Cretaceous foraminifera from Trinidad. Contributions from the Cushman Laboratory for Foraminiferal Research 4: 85-103. [ Links ]
Cushman, J.A. y Renz, H. H. 1946. The foraminiferal fauna of the Lizard Springs Formation of Trinidad, British West Indies. Cushman Laboratory for Foraminiferal Research, Special Publication 18: 1-48. [ Links ]
Duque Caro, H. 1972a. Ciclos tectónicos y sedimentarios en el Norte de Colombia y sus relaciones con la paleoecología. Boletín Geológico 19: 1-23. [ Links ]
Duque Caro, H. 1972b. Relaciones entre la bioestratigrafía y la cronoestratigrafía en el llamado geosinclinal de Bolívar. Boletín Geológico 19: 25-68. [ Links ]
Fahrni, J.F., Pawlowski, J., Richardson, S., Debenay, J.P. y Zaninetti, L. 1997. Actin suggests Miliammina fusca (Brady) is related to porcellanous rather than to agglutinated foraminifera. Micropaleontology 42: 211-214. [ Links ]
Friedberg, W. 1901. Otwornice warstw inoceramowych okolic Rzeszowa i Debicy . The Foraminifera from the Inoceramus beds near Rzeszow and Debica. Rozprawy Akademii Umiejetnosci w Krakowie, Wydziat Matematyczno-Przyrodniczy, Kraków, ser. 2, 41: 601-668. [ Links ]
Fukuta, O. 1957. Upper Cretaceous foraminifera from the Rumoi coal field, Hokkaido, Japan. Geological Survey Japan, Report 172: 1-17. [ Links ]
Gawor-Biedowa, E. 1992. Campanian and Maastrichtian foraminifera from the Lublin Upland, Eastern Poland. Paleontologia Polonica 52, 187 pp. [ Links ]
Geroch, S. y Koszarski, L .1988. Agglutinated foraminiferal stratigraphy of the Silesian Flysch Trough. En: F.M. Gradstein y F. Rögl (eds.), Second Workshop on Agglutinated Foraminifera, Abhandlungen der Geologischen Bundesanstalt, Band 41: 73-79. [ Links ]
González, P.E. y colaboradores. 1965. La cuenca petrolífera de Magallanes. Minerales, Revista Trimestral del Instituto de Ingenieros de Minas de Chile 20: 43-62. [ Links ]
Gradstein, F.M., Kaminski, M.A. y Berggren, W.A. 1988. Cenozoic foraminiferal biostratigraphy of the Central North Sea. En: F.M. Gradstein y F. Rögl. (eds.), Second Workshop on Agglutinated Foraminifera, Abhandlungen der Geologischen Bundesanstalt 41: 97-108. [ Links ]
Gradstein, F.M., Kaminski, M.A., Berggren, W.A., Kristiansen, I.L. y Díorio, M.A. 1994. Cenozoic biostratigraphy of the North Sea and Labrador shelf. Micropaleontology 40, supplement 1994: 152 pp. [ Links ]
Grzybowski, J. 1896. Otwornice czerwonych itów z Wadowic. Rozprawy Akademii Umiejetnosci w Krakowie, Wydziat Matematyczno-Przyrodniczy, Kraków, ser. 2, 30: 261-308. [ Links ]
Grzybowski, 1901. Otwornice warstw inoceramowych okolicy Gorlic. Rozprawy Akademii Umiejetnosci w Krakowie, Wydziat Matematyczno-Przyrodniczy, Kraków, ser. 3, 41: 219-288. [ Links ]
Hanagata, S., Motoyama, I. y Miwa, M. 2001. Geologic ages of the last occurrence of Spirosigmoilinella compressa and first occurrence of Miliammina echigoensis (benthic foraminifera) and their paleoceanographic implications-response to the latest Miocene-earliest Pliocene sea level change. Journal of the Geological Society of Japan 10: 101-116. [ Links ]
Hanzliková, E. y Roth, Z. 1963. Review of the Cretaceous stratigraphy of the flysch zone in west carpathians. Geologický Sborník 37-81. [ Links ]
Haynes, J.R. 1981. Foraminifera. MacMillan Publishers, Londres. 433 p. [ Links ]
Hedberg, D.H. 1937. Foraminifera of the middle Tertiary Carapita Formation of northeastern Venezuela. Journal of Paleontology 11: 661-697. [ Links ]
Herm, D. 1966. Micropaleontological aspects of the Magallanese Geosyncline, Southernmost Chile, South America. Proceedings of the Second West African Micropaleontological Colloquium (Ibadan June 18th-July 1st 1965): 72-86. [ Links ]
Heron-Allen, E. y Earland, A. 1930. Some new foraminifera from the South Atlantic: III Miliammina, a new siliceous genus. Journal of the Royal Microscopical Society of London, ser. 3, 50: 38- 45. [ Links ]
Hervé, F., Godoy, E., Mpodozis, C. y Fanning, M. 2004. Monitoring magmatism of the Patagonian batholith through the U-PB shrimp dating of detrital zircons in sedimentary units of the Magallanes Basin. Bollettino di Geofisica teorica ed applicata. An International Journal of Earth Sciences. International Symposium on the Geology and Geophysics of the Southernmost Andes, the Scotia Arc and the Antarctic Peninsula. Extended Abstracts of the GeoSur 2004 International Symposium, 22-23 November, Buenos Aires, Argentina, 45 supplement: 113-116
Hiltermann, [ Links ] H.H. 1974. Rzehakina epigona und Unterarten dieser Foraminifere. Paläontologische Zeitschrift 48: 36-56. [ Links ]
Homola, V. y Hanzliková, E. 1955. Biotragrafiké, tektoniké a litologické studie na Tesinsku. Sbornik Üstrední ústav geologický, paleontologiký 21: 317-496. [ Links ]
Hornibrook, N. de B., Brazier, R.C. y Strong, C.P. 1989. Manual of New Zealand Permian to Pleistocene Foraminiferal Bioestratigraphy. New Zealand Geological Survey Paleontological Bulletin 56: 1-175. [ Links ]
Huber, B.T. 1988. Upper Campanian-Paleocene foraminifera from the James Ross Island region, Antarctic Peninsula. Geological Society of America, Memoir 169: 163-252. [ Links ]
Jones, G.D. 1988. A paleoecological model of late Paleocene "Flysh-Type" agglutinated foraminifera using the paleoslope transect approach, Viking Graben, North Sea. En: F.M. Gradstein y F. Rögl (eds.), Second Workshop on Agglutinated Foraminifera, Abhandlungen der Geologischen Bundesanstalt, Band 41: 143-153. [ Links ]
Jones, R.W. y Charnock, M.A. 1985. "Morphogroups" of agglutinating foraminifera. Their life positions and feeding habits and potential applicability in (paleo)ecological studies. Revue de Paléobiologie 4: 311-320. [ Links ]
Jurkiewicz, H. 1967. Foraminifers in the sub-Melinitic Paleogene of the Polish Middle Carpathians. Biuletyn Instytutu Geologicznego 210: 5-116. [ Links ]
Kaiho, K. 1992. A low extinction rate of intermediate-water benthic foraminifera at the Cretaceous/Tertiary boundary. Marine Micropaleontology 18: 229-259. [ Links ]
Kaminski, M.A. y Austin, W.E.N. 1999. Oligocene Deep Water Agglutinated Foraminifers at Site 985, Norwegian Basin, southern Norwegian Sea. En: M.E. Raymo, E. Jansen, P. Blum y T.D. Herbert (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 162: 169-177. [ Links ]
Kaminski, M.A. y Geroch, S. 1997. Psamminopelta gradsteini n. sp., a new species of Paleogene deep-water agglutinated foraminifera from the northern North Atlantic and Polish Outer Carpathians. En: C. Hass y M.A, Kaminski. (eds.), Micropalaeontology & Paleoceanography of the northern North Atlantic, Grzybowski Foundation Special Publication 5: 249-252. [ Links ]
Kaminski, M.A., Gradstein, F.M. y Berggren, W.A. 1989. Paleogene benthic foraminiferal stratigraphy and paleoecology at Site 647, Southern Labrador Sea. En: M.A. Arthur, S. Srivastava, y colaboradores (eds.). Proceedings of the Ocean Drilling Program, Scientific Results, 105: College Station, TX (ocean Drilling Program): 705-730. [ Links ]
Kaminski, M.A., Gradstein, F.M., Goll, R.M. y Grieg, D. 1990. Biostratigraphy and paleoecology of deep-water agglutinated foraminifera at ODP Site 643, Norwegian-Greenland Sea. En: C.
Hemleben, M.A. Kaminski, W. Kuhnt y D.B. Scott (eds.), Paleoecology, Biostratigraphy, Paleoceanography and Taxonomy of Agglutinated Foraminifera, Kluwer Academic Publishers, pp 53-76. [ Links ]
Kaminski, M.A., Gradstein, F.M., Berggren, W.A., Geroch, S. y Beckmann, J.P. 1988. Flysch-type agglutinated foraminiferal assemblages from Trinidad: Taxonomy, stratigraphy and paleobathymetry. En: F.M. Gradstein y F. Rögl (eds.), Second Workshop on Agglutinated Foraminifera, Abhandlungen der Geologischen Bundesanstalt, Band 41: 155-227. [ Links ]
Koutsoukos, E.A.M. 2000. "Flysch-type" foraminiferal assemblages in the Cretaceous of northeastern Brazil. En: M.B. Hart, M.A. Kaminski y C.W. Smart (eds.), Proceedings of the Fifth International Workshop on Agglutinated Foraminifera, Grzybowski Foundation Special Publication 7: 243-260. [ Links ]
Krasheninnikov, V.A. y Pflaumann, U. 1977. Cretaceous agglutinated foraminifera of the Atlantic Ocean off west Africa (Leg. 41, Deep sea Drilling Project). En: Y. Lancelot, E. Seibold, y colaboradores (eds.), Initial Reports of the Deep Sea Drilling Project, Washington (U.S. Government Printing Office) 61: 565-580. [ Links ]
Kuhnt, W. y Kaminski, M.A. 1997. Cenomanian to Lower Eocene deep-water agglutinated foraminifera from the Zumaya section, Northern Spain. Annales Societatis Geologorum Poloniae 67: 257-270. [ Links ]
Kuhnt, W., Moullade, M. y Kaminski, M.A. 1998. Upper Cretaceous, K/T boundary, and Paleocene agglutinated foraminifers from Hole 959D (ODP Leg 159, Côte D'Ivoire-Ghana transform margin). En: J. Mascle, G.P. Lohmann y M. Moullade (eds.), Proceedings of the Ocean Drilling Program, Scientific Results 159: 389-411. [ Links ]
Latgé Milward de Azevedo, R., Gomide, J. y Viviers, M.C. 1987. Geo-História da Bacia de Campos, Brasil: do Albiano ao Maastrichtiano. Revista Brasileira de Geociências 17: 139-146. [ Links ]
Liebus, A. y Schubert, R.J. 1903. Die Foraminiferen der Karpathischen Inoceramenschichten von Gbellan in Ungarn (Puchower Mergel). Jahrbuch der kauserlich-koniglichen geol. Reichsanstalt (Wien) 52: 285-310. [ Links ]
Loeblich, A.R. y Kniker, H.T. 1950.[The foraminiferal genus Rzehakina in Magallanes Province, Chile. Empresa Nacional de Petroleo, ENAP, Chile, 12 pp. Inédito. [ Links ]].
Loeblich, A. y Tappan, H. 1987. Foraminiferal genera and their classification. Van Nostrand Reinhold, New York, 1: 970 pp., 2: 212 pp., 847 lám. [ Links ]
Macfadyen, W.A. 1933. Fossil foraminifera from the Burdwood Bank and their geological significance. Discovery Reports 7: 3- 16. [ Links ]
Macellari, C.E. 1988. Stratigraphy, sedimentology and paleoecology of Upper Cretaceous-Paleocene shelf-deltaic sediments of Seymour Island. Geological Society of America, Memoir 169: 25- 53. [ Links ]
Malumián, N. 1990. Foraminíferos bentónicos del Cretácico de cuenca Austral, Argentina. En: W. Volkheimer (ed.), Bioestratigrafía de los Sistemas Regionales del Jurásico y Cretácico de América del Sur (Mendoza, 1986): 429-495. [ Links ]
Malumián, N. y Caramés, A. 2002. Foraminíferos de sedimentitas ricas en carbono orgánico: Formación La Barca, Paleoceno superior, Tierra del Fuego, República Argentina. Revista de la Asociación Geológica Argentina 37: 219-231. [ Links ]
Malumián, N. y Náñez, C. 1991. Paleogeografía del Terciario medio del cono sur: avance de aguas antárticas. 6° Congreso Geológico Chileno, Resúmenes expandidos: pp. 847-851. [ Links ]
Malumián, N. y Náñez, C. 1996. Microfósiles y nanofósiles calcáreos de la plataforma Continental. En: V.A. Ramos y M.A. Turic (eds.) (Buenos Aires), 13º Congreso Geológico Argentino y 3º Congreso de Exploración de Hidrocarburos, Geología y Recursos Naturales de la Plataforma Continental Argentina, Relatorio 5: 73- 93. [ Links ]
Malumián, N. y Náñez, C. 2002. Los foraminíferos. Su significado geológico y ambiental. En: M.J. Haller (ed.), Geología y Recursos Naturales de Santa Cruz, 15° Congreso Geológico Argentino (El Calafate), Relatorio, Capítulo II-8: 481-493. [ Links ]
Malumián, N. y Panza, J.L. 2000. Hoja Geológica 5172-III Yacimiento de Río Turbio Provincia de Santa Cruz. Programa Nacional de Cartas Geológicas de la república Argentina 1:250.000. Secretaría de Energía y Minería, Servicio Geológico Minero Argentino, Instituto de Geología y Recursos Minerales, Boletín 247: 1-66. [ Links ]
Martínez R., J.I. 1989. Foraminiferal biostratigraphy and paleoenvironments of the Maastrichtian Colon mudstones of northern South America. Micropaleontology 35: 97-113. [ Links ]
Matsunaga, T. 1955. Spirosigmoilinella, a new foraminiferal genus from the Miocene of Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Ser. 18: 49-50. [ Links ]
Miller, K.G., Gradstein, F.M. y Berggren, W.A. 1982. Late Cretaceous to Early Tertiary agglutinated benthic foraminifera in the Labrador sea. Micropaleontology 28: 1-30. [ Links ]
Montanaro-Gallitelli, E. 1955. Foraminiferi cretacei delle marne a fucoidi di Serramazzoni (appennino modenese). Atti e Memorie Accademia di Scienze, Lettere ed Arti Modena, ser. 5, 13: 175-204. [ Links ]
Náñez, C. y Malumián, N. 2000. Foraminíferos aglutinados de la Formación Cerro Cazador. En: N. Malumián y J.L. Panza. Hoja Geológica 5172-III Yacimiento Río Turbio, Provincia de Santa Cruz. Servicio Geológico Minero Argentino, Boletín 247, Apéndice 1: 69- 78. [ Links ]
Natland, M.L., González, P.E., Cañón, A. y Ernst, M. 1974. A system of stages for correlation of Magallanes Basin sediments. Geological Society of America, Memoir 139: 1-126. [ Links ]
Neagu, T. 1970. Micropaleontological and stratigraphical study of the Upper Cretaceous deposits between the upper valleys of the Buzau and Riul Negru rivers (eastern Carpathians). Institut Géologique, Mémoires 12: 7-109. [ Links ]
Olivero, E.B. y Malumián, N. 1999. Eocene stratigraphy of southeastern Tierra del Fuego Island, Argentina. Bulletin of the American Association of Petroleum Geologist 183: 295-313. [ Links ]
Olivero, E.B. y Malumián, N. (en prensa). Mesozoic-Cenozoic stratigraphy of the Fuegian Andes, Argentina. Geologica Acta. [ Links ]
Olivero, E.B. y Medina, F. A. 2000. Patterns of Late Cretaceous ammonite biogeography in southern high latitudes: The Family Kossmaticeratidae in Antarctica. Cretaceous Research 21: 269-279. [ Links ]
Olivero, E.B., Malumián, N. y Palamarczuk, S. 2003. Estratigrafía del Cretácico Superior-Paleoceno del área de Bahía Thetis, Andes fueguinos, Argentina: acontecimientos tectónicos y paleobiológicos. Revista Geológica de Chile 30: 245-263. [ Links ]
Olivero, E.B., Malumián, N., Palamarczuk, S. y Scasso, R.A. 2002. El Cretácico superior-Paleogeno del área del Río Bueno, costa atlántica de la Isla Grande de Tierra del Fuego. Revista de la Asociación Geológica Argentina 57: 199-218. [ Links ]
Olszewska, B. 1997. Foraminiferal boostratigraphy of the Polish Outer Carpathians: a record of basin geohistory. Annales Societatis Geologorum Poloniae 67: 325-337. [ Links ]
Olszewska, B. y Smagowicz, M. 1977. Comparison of biostratigraphic subdivisions of the Upper Cretaceous and Paleogene of the Dukla Unit on the basis of planktonic foraminifers and nannoplankton. Przeglad Geologiczny 7: 359-363 (en polaco con resumen en inglés). [ Links ]
Ostermann, L.E. y Spiegler, D. 1996. Agglutinated benthic foraminiferal biostratigraphy of the Sites 909 and 913, northern North Atlantic. En: J. Thiede, A.M. Myrhe y colaboradores (eds.). Proceedings of the Ocean Drilling Program, Scientific Results 151: 169-185. [ Links ]
Quattrocchio, M. E. y Sarjeant, W.A.S. 2003. Dinoflagellates from the Chorrillo Chico Formation (Paleocene) of the southern Chile. Ameghiniana 40: 129-153. [ Links ]
Rögl, F., 1976. Late Cretaceous to Pleistocene foraminifera from the southeast Pacific Basin, DSDP Leg 35. En: C.D. Hollister, C. Craddock y colaboradores (eds.). Initial Reports of the Deep Sea Drilling Project, Washington (U.S. Government Printing Office) 35: 539-555. [ Links ]
Rzehak, A. 1895. Über einige merkwurdige Foraminiferen aus dem Österreichischen Tertiär. Annalen Naturhistorisches Hofmuseum 10: 213-230. [ Links ]
Santos N., M. y Ramírez, H.F. 1986. La Formación Apagua, una nueva unidad eocénica en la Coordillera Occidental ecuatoriana. 4° Congreso Ecuatoriano de Geología, Minas y Petróleo, (Quito) Memoria 1: 179-189. [ Links ]
Schröder-Adams, C.J. y McNeil, D.H. 1994. New paleoenvironmentally important species of agglutinated foraminifera from the Oligocene and Miocene of the Beaufort Sea, Artic Canada. Journal of Foraminiferal Research 24: 179-190. [ Links ]
Scott, G.H. 1961. Contribution to the knowledge of Rzehakina Cushman (Foraminifera) in New Zealand. New Zealand Journal of Geology and Geophysics 4: 3-43. [ Links ]
Serova, M. Ya. 1969. Comparison of characteristics of Rzehakinidae in the Carpathian region and Pacific province, Annales de la Société Géologique de Pologne 39: 225-240. [ Links ]
Serova, M. Ya., Danilesko, L. A., Kolyada, A. A. y Petrina, N. M. 1980. Raschlenenie vyerkhnemelovykh i paleogenovykh otlozhenii Koryakskogo Nagor'ya. Izvyestiya Akademii Nauk SSSR, Seriya Géeologicheskaya 10: 73-83. [ Links ]
Sigal, J. 1970. Quelques acquisitions recentes concernant la chronostratigraphie des formations sedimentaires de l'Equater. Revista Española de Micropaleontología 1: 205-236. [ Links ]
Stainforth, R.M. 1955. Ages of Tertiary formations in northwest Perú. American Association Petroleum Geologists Bulletin 39: 2068-2077. [ Links ]
Strong, C.P., Hollis, C.J. y Wilson, G.J. 1995. Foraminiferal, radiolarian, and dinoflagelate biostratigraphy of the Late Cretaceous to Middle Eocene pelagic sediments (Muzzle Group), Mead Stream, Marlborough, New Zealand. New Zealand Journal of Geology and Geophysics 38: 171-212. [ Links ]
Szczechura, J. y Pozaryska, K. 1974. Foraminiferida from the Paleocene of Polish Carpathians (Babica Clays). Paleontologia Polonica 31: 142 pp. [ Links ]
Tappan, H. 1957. New Cretaceous Index Foraminifera from Northern Alaska. Bulletin United States National Museum 215: 201-222. [ Links ]
Tappan, H. 1962. Foraminifera from the Arctic Slope of Alaska. Part 3, Cretaceous Foraminifera. Geological Survey Professional Paper 236-C: 91-209. [ Links ]
Thalmann, H.E. 1949. Regional-stratigraphische Verbreitung der Foraminiferen-Gattung Rzehakina Cushman, 1927. Eclogae Geologicae Helvetiae 42: 506-507. [ Links ]
Tjalsma, R.C. 1976. Mesozoic foraminifera from the South Atlantic, DSDP Leg 36. En: P.F. Barker, I.W.D. Dalziel y colaboradores (eds.), Initial Reports of the Deep Sea Drilling Project, Washington (U. S. Government Printing Office) 36: 493-517. [ Links ]
Turenko, T.V. 1983. New foraminiferal species of the Family Rzehakinidae from the Upper Cretaceous of Sakhalin. Paleontologicheskiy Zhurnal 3: 13-21. [ Links ]
van den Akker, T.J. H.A., Kaminski, M.A., Gradstein, F.M. y Wood, J. 2000. Campanian to Palaeocene biostratigraphy and palaeoenvironments in the Foula Sub-basin, west of the Shetland Island, UK. Journal of Micropaleontology 19, 23-43. [ Links ]
Volat, J-L., Hugo, B. y Bignoumba-Ilogue, J. 1996. Foraminifères arénacès du Crétacé supérieur du Gabon. Bulletin des Centres de Recherches Exploration-Production Elf Aquitaine 20: 229-275. [ Links ]
Webb, P.N. 1975. Paleocene foraminifera from DSDP Site 283, South Tasman Basin. En: J.P. Kennett, R.E. Houtz, et al. (eds.), Initial Reports of the Deep Sea Drilling Project, Washington (U.S. Government Printing Office) 29: 833-843. [ Links ]
Weiss, L. 1955. Planktonic index foraminifera of northwestern Perú. Micropaleontology 1: 301-319. [ Links ]
Wong, T.E. 1976. Tertiary stratigraphy and micropaleontology of the Guiana Basin. Geologie en Mijnbouw Dienst Suriname, Mefddeling 25: 13-109. [ Links ]
Recibido: 3 de agosto de 2004.
Aceptado: 6 de octubre de 2005.

