










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.43 n.4 Buenos Aires sept./dic. 2006
Revisión de la Biozona de Ogygiocaris araiorhachis (Trilobita, Tremadociano tardío) en la región de Pascha-Incamayo, Cordillera Oriental, Argentina. Parte 1: Bioestratigrafía
Beatriz G. Waisfeld1, N. Emilio Vaccari1, Blanca A. Toro2, Claudia V. Rubinstein2y Ricardo A. Astini3
1Centro de Investigaciones Paleobiológicas (CIPAL), Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Av. Vélez Sarsfield 299, 5000 Córdoba, Argentina. CONICET. bwaisfeld@com.uncor.edu
2Departamento de Geología y Paleontología, IANIGLA-CRICyT, Av. Ruiz Leal s/n, Parque General San Martín, 5500 Mendoza, Argentina. CONICET.
3Cátedra de Estratigrafía y Geología Histórica, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Pabellón Geología, Ciudad Universitaria, 2º Piso, Oficina 7, Av. Vélez Sarsfield 1611, 5016 Córdoba, Argentina. CONICET.
Abstract. Revision of the Ogygiocaris araiorhachis zone (Trilobita, Late Tremadocian) in the Paschaincamayo Region, Cordillera Oriental, Argentina. Part 1: Biostratigraphy. A revision of the Ogygiocaris araiorhachis trilobite Zone recorded in the Saladillo and Parcha formations (Pascha-Incamayo area, quebrada del Toro, Salta Province) is conducted. Stratigraphic distribution of the trilobite fauna, as well as graptolites and palynological associations are presented for the first time for the Pascha-Incamayo area. The O. araiorhachis Zone is late Tremadocian in age, and not Arenigian as previously supposed, extending from the Araneograptus murrayi to the Hunegraptus copiosus graptolite zones, and associated with the messaoudensis-trifidum acritarch assemblage. Its range is extended to the upper part of the Saladillo Formation and to the Parcha Formation. The association of the trilobite Kayseraspis asaphelloides Harrington with the messaoudensis-trifidum acritarch assemblage in the Cerro San Bernardo area (Salta) suggests that the Kayseraspis asaphelloides and the Ogygiocaris araiorhachis zones were broadly coeval. Hence, these late Tremadocian zones do not represent a true temporal succession as proposed by Harrington in the trilobite scheme of 1957. In contrast, they portray geographically restricted ecological expressions in response to lateral environmental shifts. Differences in oxygenation levels appear to be the most influential control. Accordingly, the K. asaphelloides and O. araiorhachis zones are of limited use for biostratigraphical purposes and for regional correlations.
Resumen. Se llevó a cabo una revisión de la Biozona de Ogygiocaris araiorhachis presente en las formaciones Saladillo y Parcha (área de Pascha-Incamayo, quebrada del Toro, Provincia de Salta). Se presenta, por primera vez para el área de Pascha-Incamayo, la distribución estratigráfica de la fauna de trilobites así como de las asociaciones palinológicas y biozonas de graptolitos. La Biozona de O. araiorhachis tiene una edad tremadociana tardía y no arenigiana temprana como se aceptaba tradicionalmente, extendiéndose entre las biozonas de graptolitos de Araneograptus murrayi y Hunegraptus copiosus y asociada con la asociación palinológica de messaoudensis-trifidum. El rango de esta biozona se extiende entre la parte superior de la Formación Saladillo y la Formación Parcha. La relación entre la especie Kayseraspis asaphelloides Harrington y la asociación de acritarcos de essaoudensis-trifidum en el área del Cerro San Bernardo (Salta) sugiere que las biozonas de K. asaphelloides y O. araiorhachis fueron relativamente contemporáneas. Por lo tanto, estas biozonas de edad tremadociana tardía no representarían una verdadera sucesión temporal como fuera propuesto por Harrington en el esquema de 1957. En contraste, estas asociaciones corresponderían a expresiones ecológicas diferenciales, geográficamente restringidas, en respuesta a cambios laterales del ambiente. Posiblemente, las diferencias en los niveles de oxigenación habrían sido el factor con mayor influencia. En consecuencia, las biozonas de K. asaphelloides y O. araiorhachis tienen escaso valor desde el punto de vista bioestrátigráfico y su utilidad para establecer correlaciones a escala regional es muy limitada.
Key words. Trilobites; Biostratigraphy; Late Tremadocian; Pascha; Cordillera Oriental; Argentina
Palabras clave Trilobites; Bioestratigrafía; Tremadociano tardío; Pascha; Cordillera Oriental; Argentina.
Introducción
Los trilobites del Cambro-Ordovícico representan elementos de gran valor para establecer precisiones cronológicas y correlaciones a escala regional. En particular, la abundancia, diversidad y amplia distribución del grupo en las unidades del Grupo Santa Victoria (cuenca del noroeste argentino) convierte a los trilobites en herramientas clave para este fin.
El primer intento en este sentido fue llevado adelante por Harrington (1938) quien, a partir de colecciones provenientes de localidades y horizontes estratigráficos diferentes, distinguió una serie de "Faunas" sucesivas con distinta composición taxonómica, procedentes de la cuenca del noroeste (quebrada del Toro, Azul Pampa, Cajas, Iruya, Purmamarca, Alfarcito, cerro San Bernardo) y del Sistema de Famatina (río Volcancito).
Posteriormente, Harrington (en Harrington y Leanza, 1957) revisó las "Faunas" definidas en 1938 sobre la base del estudio de nuevas y extensas colecciones de trilobites procedentes de numerosas localidades. Este autor sintetizó e integró la información taxonómica y litológica, estableciendo un grupo de biozonas de trilobites para distintas unidades ordovícicas del noroeste argentino, Sistema de Famatina y Precordillera Argentina. El esquema elaborado por Harrington (en Harrington y Leanza, 1957) representó un avance sustancial para comprender las sucesiones ordovícicas de Argentina y fue empleado durante varias décadas para establecer correlaciones regionales y aún intercontinentales.
En años recientes algunas de las biozonas definidas fueron objeto de revisiones críticas, tanto desde el punto de vista de la clasificación sistemática como de la posición estratigráfica de los taxones involucrados. En tal sentido, la revisión de las clásicas biozonas de "Proetiella" tellecheai (Vaccari, 1993), "Heokaspis" megacantha (Vaccari y Waisfeld, 1994), y "Hoekaspis schlagintweiti" (Waisfeld, 1996), han puesto de manifiesto algunos de los inconvenientes generados empleando el esquema propuesto por Harrington (en Harrington y Leanza, 1957). Recientemente, Waisfeld y Vaccari (2003) realizaron una puesta al día de los progresos en la revisión del esquema de biozonas definidas en 1957.
La región de Pascha-Incamayo es una localidad clásica para el estudio de las sucesiones cambro-ordovícicas de la cuenca del noroeste de Argentina. La estratigrafía de la región se conoce a partir de los trabajos pioneros de Keidel (1937, 1943), Vilela (1956) y Harrington (en Harrington y Leanza, 1957). Kobayashi (1935, 1937), Harrington (1937, 1938) y Harrington y Leanza (1957) dan a conocer parte de la abundante fauna de trilobites. El estudio de la fauna y de las unidades expuestas en el área ha cobrado un renovado interés en los últimos años. En tal sentido, recientes contribuciones de Rao y Tortello (1998), Albanesi et al. (1997), Ortega et al. (1997), Ortega y Albanesi (2002, 2003), Rubinstein et al. (1999), Tortello (1999), Tortello y Rao (2000), Astini (2003, 2005), Toro et al. (2003) y Waisfeld et al. (2003) abordan aspectos taxonómicos, estratigráficos y bioestratigráficos de la región.
La región de Pascha- Incamayo es un área clásica para el estudio de las sucesiones cambro-ordovícicas por dos razones fundamentales. Por un lado, representa el área tipo de varias unidades estratigráficas tales como las formaciones Sococha, Lampazar, Cardonal, Saladillo, Parcha y Devendeus (Keidel, 1937; Astini, 2003; Astini, 2005). Por otro lado, las biozonas de trilobites de Parabolina (Neoparabolina) frequens argentina, Jujuyaspis keideli, Kainella meridionalis y Ogygiocaris araiorhachis han sido documentadas en la región (Kobayashi, 1937; Harrington, en Harrington y Leanza, 1957; Tortello y Rao, 2000).
Sin embargo, las potentes sucesiones del Grupo Santa Victoria expuestas en el área exhiben una significativa complejidad estratigráfica y estructural. Por tal motivo, tanto las relaciones entre las unidades mencionadas como las biozonas de trilobites registradas requieren una detallada revisión con criterios modernos mediante un enfoque interdisciplinario.
En relación a las biozonas de trilobites, recientemente, Tortello y Rao (2000) mencionan por primera vez la especie Parabolina (Neoparabolina) frequens argentina (Kayser) en el Angosto de Lampazar. Estos autores documentan la biozona homónima en la Formación Lampazar y tramo inferior de la Formación Cardonal. Harrington y Leanza (1957) indican la presencia de Jujuyaspis keideli Kobayashi en las cabeceras de la quebrada Lampazar y en la divisoria de aguas entre las quebradas Huaico Hondo y Devendeus. Hasta el momento y en las secciones relevadas para este trabajo (figura 1) no se cuenta con registros de esta última especie.

Figura 1. Mapa de ubicación / location map.
Kobayashi (1935) describe la especie Kainella meridionalis Kobayashi con material procedente de la localidad de Pascha-Incamayo (Angosto de Pascha) y acuña el término "Fauna de Kainella" para designar la asociación de trilobites que acompaña esta especie en la región (e.g. Asaphellus catamarcensis Kobayashi 1937, "Kainella" conica Kobayashi, 1935, Leptoplastides argentiniensis (Kobayashi, 1935), etc.). Posteriormente, Harrington (1957) define la Biozona de K. meridionalis sobre la base de los registros de la "Fauna de Kainella" en las áreas de Pascha-Incamayo, Quebrada de Humahuaca y Santa Victoria-Iruya. Recientemente, Waisfeld y Vaccari (2003) documentan la especie K. meridionalis y la Fauna de Kainella en bloques retrabajados de la Formación Cardonal y de la Formación Devendeus. Particularmente, en esta última los bloques contienen asociaciones de trilobites de un rango de edad amplio (Tremadociano- Arenigiano basal?). Por lo tanto, dado el particular registro de K. meridionalis en bloques retrabajados, la zona homónima no se reconoce en las secciones del área de Pascha-Incamayo relevadas para este estudio. Sin embargo, Tortello y Rao (2000) y Ortega y Albanesi (2003) indican la presencia de Kainella meridionalis en los niveles de la Formación Saladillo expuesta en el Angosto de Lampazar, asociada a conodontes de la Biozona de Cordylodus angulatus y graptolitos de la Zona de Bryograptus. Una revisión de la fauna presente en los bloques de las formaciones Cardonal y Devendeus y de la Biozona de K. meridionalis se encuentra actualmente en progreso por los autores de este trabajo.
La Biozona de Ogygiocaris araiorhachis fue definida por Harrington (1957) para el área de Pascha- Incamayo, y extendida a las regiones de Santa Victoria y Potrero de Castillo, fundamentalmente sobre la base del registro de la especie O. araiorhachis Harrington y Leanza en dichas localidades. El objetivo principal de esta contribución es llevar a cabo una revisión de la Biozona de Ogygiocaris araiorhachis. Para tal fin, se reevaluó el material asignado a esta especie, se realizó una revisión taxonómica completa de la fauna de trilobites asociada, y se examinó su distribución espacio-temporal. Estos aspectos fueron analizados sobre la base de nuevas colecciones procedentes de aquellas localidades donde Harrington (en Harrington y Leanza, 1957) documentó esta biozona (Pascha-Incamayo y Santa Victoria). En particular, nuestro estudio se centró en las formaciones Saladillo y Parcha expuestas en el área de Pascha-Incamayo (quebrada del Toro, 40 km al noroeste de Salta, figura 1) donde se presenta por primera vez la distribución estratigráfica de los taxones de trilobites y se establece su calibración cronológica sobre la base de graptolitos y palinomorfos. El estudio sistemático de la fauna de trilobites incluidos en la Biozona de O. araiorhachis se presenta en la segunda parte de este trabajo (Waisfeld y Vaccari, parte 2 -este volumen-).
Estratigrafía de la región Pascha-Incamayo
El espesor estratigráfico registrado por las unidades pertenecientes al ciclo del Grupo Santa Victoria en la región de Pascha-Incamayo es de aproximadamente 1700 m. La totalidad de la estratigrafía ordovícica expuesta en la región fue incluida dentro del Grupo Incamayo (Aceñolaza, 1983), sin embargo, el conjunto de las formaciones Lampazar (~40 m), Cardonal (~150 m) y Saladillo (~800 m) definidas originalmente por Keidel (1937, 1943) fue incluido con anterioridad en el Grupo Guayoc Chico (Ramos, 1973). Recientemente Astini (2003) enmendó y restringió el alcance geográfico de este último grupo a los contrafuertes occidentales de la Cordillera Oriental (figura 2). Para la región de Pascha- Incamayo propuso una estratigrafía conformada por las siguientes unidades de base a techo (figura 2): Formación Sococha (Astini, 2003), Formación Lampazar (Keidel, 1937), Formación Cardonal (Keidel, 1937), Formación Devendeus (Astini, 2003), Formación Saladillo (Keidel, 1937) y Formación Parcha (Keidel, 1937). Astini (2003) señala que la columna expuesta en esta región constituye una excelente exposición de la estratigrafía del Ordovícico Inferior en la Cordillera Oriental a pesar de sus relaciones laterales no muy claras con otras columnas del ámbito de la Cordillera Oriental.
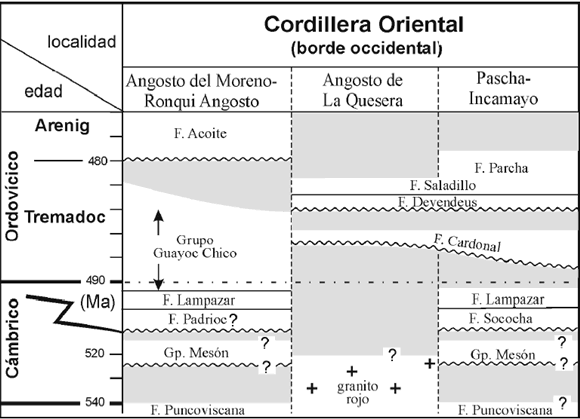
Figura 2. Cuadro estratigráfico general de las principales unidades del Grupo Santa Victoria en el borde occidental de la Cordillera Oriental (regiones de Ronqui Angosto-El Moreno, Angosto de la Quesera y Pascha-Incamayo) (modificado de Astini, 2005) / general stratigraphicchart showing main units of the Santa Vicotoria Group on western foot of the Cordillera Oriental (Ronqui Angosto-El Moreno, Angosto dela Quesera and Pascha-Incamayo) (modified from Astini, 2005)
La columna más completa de la región es la expuesta sobre el curso de la quebrada de Sococha (figuras 1 y 2) donde aflora la sección tipo de la formación homónima compuesta por un paquete de 37.4 m de espesor de areniscas cuarzosas amalgamadas apoyadas en discordancia erosiva sobre el Grupo Mesón. Conglomerados cuarzosos lenticulares separan a esta unidad de las sub y suprayacentes. El contacto con la suprayacente Formación Lampazar es neto y está representado por un salto repentino a una asociación rítmica con depósitos de plataforma externa en un contexto con abundancia de pelitas. Le suprayace la Formación Cardonal que se apoya en marcada discordancia erosiva. La superficie basal de esta última unidad es fuertemente erosiva y constituye una superficie muy irregular con grandes calcos de flujo que reflejan una fuerte etapa erosiva sobre fondos blandos. Los bancos basales contienen abundancia de intraclastos junto a gránulos y clastos de cuarzo y areniscas cuarzosas redondeadas. Internamente la unidad es muy heterogénea y los diferentes bancos carecen de continuidad lateral y están afectados por una intensa deformación sinsedimentaria. Se destacan cuerpos lenticulares de flujos de detritos conteniendo frecuentes clastos calcáreos con abundante y variada macrofauna de invertebrados marinos. Hacia el tope se intercalan paquetes de areniscas subfeldespáticas gruesas de color blanquecino con importante desarrollo de conjuntos de estratificación cruzada con terminación tangencial y frecuentes intercalaciones pelíticas. Estos bancos poseen abundantes asociaciones de baja diversidad de Cruziana localmente atravesadas o alternando con tapices de Skolithos que, localmente, forman verdaderos pipe-rocks. El conjunto de las facies e icnofacies constituyen elementos característicos de ambientes con dominio mareal.
A continuaci ón, en contacto neto erosivo, se dispone la Formación Devendeus compuesta por un conjunto predominantemente conglomerádico con algunas intercalaciones arenosas y más finas. Se cuentan hasta siete bancos de conglomerados (con espesor individual >1m) relativamente tabulares, caracterizados por clastos de composición calcárea y un contenido faunístico muy variado. Acompañan clastos de areniscas y, raramente, alguno de granito alterado; estos últimos bien redondeados. Esta variedad de composiciones litológicas representa una estratigrafía retrabajada, ausente en la región (Astini, 2003), es decir una estratigrafía que fue expuesta, denudada y resedimentada implicando una importante discontinuidad estratigráfica (Astini, 2003, 2005). La Formación Devendeus posee importantes variaciones laterales con depósitos dominantemente verde oliva de flujos de detritos y fangolitas guijosas, ocasionalmente portadores de ostrácodos, alternantes con pelitas y areniscas. Internamente posee superficies erosivas que truncan a esta unidad y lateralmente llegan a acuñarla y fue Keidel (1943) quien sugirió que estas superficies fueron labradas por efectos glaciales. Le suprayace en un contacto neto regional una unidad dominantemente pelítica conocida como Formación Saladillo que en su base posee algunas intercalaciones areniscosas y un color más verde-amarillento. Esta unidad se destaca por su coloración gris oscura y la generación de relieves suaves, al ser las pelitas negras su constituyente principal. Sin embargo, en parte y fundamentalmente hacia el tope, contiene varios intervalos con intercalaciones de areniscas gradadas, algunas de ellas con deformación por carga y evidencias de escape de agua, como así también intercalaciones de limolitas calcáreas y capas con abundantes concreciones. Finalmente la sucesión pasa en forma transicional a la Formación Parcha que se reconoce por la tonalidad más verdosa de sus pelitas y la mayor frecuencia de alternancias areniscosas que conforman ciclos estratocrecientes y decrecientes de entre 5 y 15 m de espesor, aunque el contacto entre formaciones se hace coincidir arbitrariamente con un cuerpo grueso y tabular de areniscas descripto como una megacapa (Astini, 2002).
Desde un punto de vista estratigráfico se interpreta el desarrollo de ciclos transgresivos en la base de las formaciones Lampazar y Saladillo y de importantes ciclos regresivos en la base de las formaciones Sococha, Cardonal y Devendeus. A su vez, la Formación Cardonal representaría el relleno complejo de sistemas estuarinos con fuerte dominio mareal y la Formación Devendeus un pulso conglomerádico tras una prolongada etapa erosiva, incluyendo exposición subaérea. Aunque no es del todo claro el origen del aporte sefítico en la Formación Devendeus, la gran mezcla de componentes, y su ordenamiento estratigráfico indicarían que la unidad precedería al conjunto de las formaciones Saladillo y Parcha. No obstante, es posible que esta unidad esté representando un intervalo erosivo algo más joven y, en consecuencia, su intercalación podría no ser normal con las unidades que le suprayacen. El contacto entre las formaciones Saladillo y Parcha, si bien está localmente representado por una megacapa de areniscas plásticamente deformadas que implican una sedimentación repentina, no representa un marcado cambio paleoambiental sino un fenómeno de sismicidad dentro de una tendencia de progresiva somerización, representada dentro de un ciclo mayor regresivo.
Nuestro estudio se centró en las formaciones Saladillo y Parcha expuestas en las quebradas Aguada del Altillo y del Barranco, ubicadas al este del caserío de Pascha (figura 1). Ambas constituyen afluentes del Río Incamayo por la margen izquierda, situándose la primera aguas arriba y la segunda aguas abajo del mencionado poblado. Entre ambas hay una cierta continuidad lateral aunque localmente la sección se encuentra cubierta por abanicos coluviales modernos y diferentes niveles de agradación cuaternarios. El corrimiento del alto de Salamanca, vergente al oeste, trunca la sección ordovícica en la región, exponiendo las areniscas del Grupo Mesón. En ambas quebradas existe una traza de falla subparalela al corrimiento del alto de Salamanca que aparentemente repite espesores de la Formación Parcha generando un inusual espesor de la misma (figura 3). Entre ambas la mayor diferencia la constituye el acuñamiento lateral de la Formación Devendeus, dado que mientras que en el núcleo del anticlinal de Incamayo ésta se dispone sobre la Formación Cardonal, en la quebrada del Barranco la unidad sobrepuesta a la Formación Cardonal es la Formación Saladillo. La estratigrafía de estas secciones fue comparada con la expuesta en la dorsal de la lámina corrida de Sococha, expuesta hacia el oeste del poblado, donde Astini (2003) describiera la estratigrafía más completa de la región. En particular, se establece una estrecha correlación con hallazgos realizados en los perfiles (e.g., de sur a norte, quebradas de Sococha, Devendeus, Lampazar; véase figura 1) expuestos en esta faja de afloramientos por lo que nuestro estudio puede considerarse como representativo del área Pascha-Incamayo.
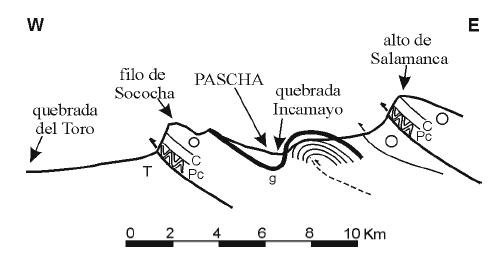
Figura 3. Perfil esquemático E-O en la región de Pascha-Incamayo (corte AA de la figura 1). Pc, Formación Puncoviscana, C, Grupo Mesón, O, Grupo Santa Victoria, T, Terciario, g, capa guía que separa las formaciones Saladillo y Parcha / schematic E-W cross sectionin the Pascha-Incamayo region (section AA from figure 1). Pc,Puncoviscana Formation, C, Mesón Group, O, Santa Victoria Group, T,Tertiary, g, key horizon separating the Saladillo and Parcha formations.
Consideraciones bioestratigráficas
Recientemente, Albanesi et al. (2001) y Ortega y Albanesi (2002, 2003) estudiaron la fauna de graptolitos y conodontes asociados en la región de Pascha- Incamayo. Ortega y Albanesi (2003, p. 81) proponen un esquema bioestratigráfico integrado por las biozonas de Bryograptus, Kiaerograptus, K. supremus, Araneograptus murrayi y Hunnegraptus copiosus, asignadas al Tremadociano tardío.
De acuerdo a nuestros registros (figura 4), en la parte superior de la Formación Saladillo se registran los graptolitos Paradelograptus sp., Adelograptus sp. y Kiaerograptus sp. Esta asociación sería equivalente a la Biozona de Kiaerograptus, propuesta por Ortega y Albanesi (2003) y correlacionable con la zona de Aorograptus victoriae presente en otras localidades de la Cordillera Oriental (Monteros y Moya, 2002, 2003) y recientemente localizada en Bolivia (Maletz y Egenhoff, 2001; Egenhoff et al., 2004). Los acritarcos más antiguos registrados en la Formación Saladillo están asociados a los graptolitos mencionados. Son pobres en abundancia, diversidad y preservación, pudiendo reconocerse los géneros Acanthodiacrodium, Stelliferidium y Stellechinatum?

Figura 4. Columna estratigráfica generalizada de la región de Pascha-Incamayo mostrando el rango vertical de las especies de trilobites y palinomorfos y biozonas de graptolitos / generalized stratigraphic column of the Pascha-Incamayo region showing the vertical ranges of trilobite and palynomorph species, and graptolite zones.
La primera aparición de la especie Araneograptus murrayi (J. Hall) se registra en el tramo cuspidal de la Formación Saladillo, marcando el comienzo de la biozona homónima, y su registro continúa en los niveles inferiores de la Formación Parcha. En esta biozona se han identificado además de la especie guía, ejemplares de Kiaerograptus supremus Lindholm y representantes de los géneros Paradelograptus y Didymograptus.
En el tramo cuspidal de la Formación Saladillo, los acritarcos continúan siendo escasos y pobremente preservados, sumándose los géneros Vavrdovella y Cymatiogalea? En la Formación Parcha se ha identificado una asociación de acritarcos característica de la transición tremadociana-arenigiana, la cual fue descripta por primera vez para la Formación Watch Hill, en el English Lake District, noroeste de Inglaterra (Molyneux y Rushton, 1988), donde fue datada sobre la base de graptolitos. Posteriormente, la misma fue dividida en cinco sub-asociaciones y se efectuó una detallada bioestratigrafía para esta área, basada en graptolitos y acritarcos (Cooper et al., 1995). Esta asociación, posteriormente denominada de messaoudensis- trifidum (Servais y Molyneux, 1997) es considerada de gran importancia bioestratigráfica debido a su amplio desarrollo en áreas peri-gondwánicas (Servais y Mette, 2000). Taxones característicos de la asociación de messaoudensis-trifidum se registran en los niveles inferiores de la Formación Parcha, correspondientes a la Biozona de Araneograptus murrayi. Si bien la asociación está pobremente preservada pueden identificarse especies como Caldariola glabra var. glabra (Martin) Molyneux y Rushton, 1988, Cymatiogalea messaoudensis var. messaoudensis Jardiné et al., 1974, Vavrdovella areniga (Vavrdova) Loeblich y Tappan, 1976, Rhopaliophora palmata (Combaz y Peniguel) Playford y Martin, 1984, Stelliferidium sp., Coryphidium? sp. y Acanthodiacrodium spp.
Cabe destacar, que de acuerdo a nuestras observaciones, la especie K. supremus comienza a registrarse junto con A. murrayi, al igual que lo que ocurre en Bolivia (Egenhoff et al., 2004). Esto permite apoyar el cuestionamiento de la validez de la Biozona de K. supremus para Sudamérica, postulado por Egenhoff et al. (2004), a partir de la distribución de dicho taxón en Bolivia. La presencia de la Biozona de Araneograptus murrayi certifica que este tramo de la secuencia se depositó durante el Tremadociano tardío (La2 de Australia), pudiendo correlacionarse con esta misma biozona reconocida en Bolivia (Egenhoff et al., 2004) y con las biozonas de Kiaerograptus supremus y Araneograptus murrayi de Escandinavia (Lindholm, 1991).
A partir de los niveles medios de la Formación Parcha comienza a registrarse Hunnegraptus copiosus Lidholm, definiendo la biozona del mismo nombre. Ésta continúa desarrollándose por el resto de la unidad y permite asignar a los niveles portadores una edad tremadociana tardía, equivalente a las establecidas en Escandinavia (Lindholm, 1991), en el suroeste de China (Zhang y Chen, 2003) y Bolivia (Egenhoff et al., 2004).
En el tramo cuspidal de la Formación Parcha y dentro de la Biozona de H. copiosus (figura 4) se registra nuevamente la asociación de acritarcos de messaoudensis- trifidum. Los taxones dominantes de esta asociación son los acritarcos denominados "galeate" (Servais y Eiserhardt, 1995) que se caracterizan por poseer vesículas de forma hemisférica, con una amplia abertura (pyloma), cerrada por un opérculo. Éstos son Cymatiogalea messaoudensis var. messaoudensis, Stelliferidium cf. trifidum (Rasul) Fensome et al., 1990, Cymatiogalea deunffi y ?Caldariola glabra var. glabra. La asignación dudosa de esta última especie se debe a que los especímenes estudiados podrían ser formas intermedias entre los géneros Caldariola y Cymatiogalea. Las formas intermedias entre Acanthodiacrodium y acritarcos corynomorfíticos (Coryphidium- Vavrdovella) les continúan en abundancia, aunque representantes típicos de los géneros Coryphidium y Vavrdovella también se encuentran presentes en la asociación. Es importante destacar la presencia de formas intermedias entre Cymatiogalea messaoudensis y Cymatiogalea deunffi, como ha sido observado en la asociación de Rügen, Alemania (Servais y Molyneux, 1997) y en la de la Formación Barriga Shale, en la Sierra Morena, sudoeste de España (Servais y Mette, 2000).
Biozona de Ogygiocaris araiorhachis
Definición original
Harrington (en Harrington y Leanza, 1957, p. 29) advirtió un notable recambio faunístico entre la Fauna de Ceratopyge (Tremadociano) y la asociación de trilobites ubicada por encima de ésta, a la que denominó Fauna de Thysanopyge. Sobre la base de los trilobites procedentes fundamentalmente de las secciones aflorantes en el río Santa Victoria, quebrada del Toro y Cerro San Bernardo, este autor distinguió dos asociaciones dentro de la Fauna de Thysanopyge, una inferior: Biozona de Kayseraspis asaphelloides y otra superior: Biozona de Ogygiocaris araiorhachis. A pesar del fuerte carácter endémico de los trilobites de ambas biozonas, Harrington llevó adelante un análisis cronológico basado en el registro de especies afines presentes en otras regiones de Estados Unidos y oeste de Europa, obteniendo resultados contradictorios. Finalmente, adjudicó una edad arenigiana a ambas biozonas en virtud de la fauna de graptolitos asociada en las localidades mencionadas y en las regiones de Yavi y Tafna.
Harrington (en Harrington y Leanza, 1957) indicó la presencia de O. araiorhachis en tres localidades de la Cordillera Oriental: quebrada del Toro (área de Pascha-Incamayo), río Santa Victoria (Departamento Santa Victoria), y Río Potrero de Castillo (Yacones, noroeste de Salta) y refirió los estratos portadores al Arenigiano.
De acuerdo a Harrington en la región de Pascha Ogygiocaris araiorhachis aparece en la mitad superior de la Formación Parcha asociado a Hypermecaspis inermis Harrington y Leanza, H. armata Harrington y Leanza, Megalaspis saltaensis Harrington y Leanza, Araiopleura reticulata Harrington y Leanza y Raphiphorus? lamasi Harrington y Leanza. Según este autor, en la sección del Río Santa Victoria O. araiorhachis aparece en la parte superior de la Formación Acoite junto a Niobides armatus Harrington y Leanza, y Thysanopyge clavijoi Harrington y Leanza. Estos registros suponen la correlación entre la parte superior de la Formación Parcha (quebrada del Toro) y la parte superior de la Formación Acoite (región de Santa Victoria). Respecto a la mención de O. araiorhachis en la localidad de Potrero de Castillo, Harrington y Leanza (1957) hacen referencia a un único pigidio asociado a Thysanopyge argentina, sin mayores precisiones respecto a su procedencia.
Distribución y edad
De acuerdo a nuestra revisión taxonómica (véase Waisfeld y Vaccari, Parte 2 -este volumen-), sólo el material de la localidad tipo (Pascha-Incamayo) corresponde a la especie O. araiorhachis. En consecuencia, la distribución de este taxón queda restringida a esta área. Los ejemplares de la Formación Acoite (Santa Victoria) corresponden a un taxón diferente de O. araiorhachis. La configuración de los pigidios es claramente diferente de la del material procedente del área de Pascha por ser menos transversos, por poseer menor número de anillos axiales y segmentos pleurales, y por la ausencia de surcos interpleurales. Este material podría ser coespecífico con Thysanopyge? latelimbata Harrington y Leanza documentado también en la región de Santa Victoria. Según nuestros estudios, aún en progreso, el material procedente del río Santa Victoria debería ser acomodado en un nuevo género, diferente de Ogygiocaris y de Thysanopyge . Por su parte, el único pigidio conocido de Potrero de Castillo sugiere una estrecha relación con el material de Santa Victoria, por lo que también es excluido de O. araiorhachis.
Tradicionalmente, y tomando en cuenta la propuesta de Harrington (en Harrigton y Leanza, 1957) la Formación Parcha fue considerada de edad arenigiana. Esta asignación fue mantenida hasta épocas recientes (Benedetto, 1976; Benedetto y Malanca, 1975; Albanesi et al., 1997; Ortega et al., 1997; Moya, 1999; Moya et al. 1994; entre otros) y, corrientemente sustentada por la presencia de O. araiorhachis y Thysanopyge "argentina". Más recientemente, Ortega y Albanesi (2002, 2003) sobre la base de la fauna de graptolitos y conodontes, establecen una edad tremadociana tardía para esta unidad.
En función de nuestras observaciones de campo y nuevas colecciones, el primer registro de O. araiorhachis coincide con el de Araneograptus murrayi, en el tramo superior de la Formación Saladillo, mientras que la última aparición de este trilobite está en la Formación Parcha, dentro de la Biozona de H. copiosus (figura 4). El primer registro cierto del género Coryphidum se ubica en los niveles medio-superiores de la Formación Parcha, correspondientes a la Biozona de H. copiosus. De los acritarcos relacionados a O. araiorhachis y entre los registrados en el tramo inferior de la Formación Parcha se destaca la presencia de formas asignadas con dudas al género Coryphidium debido a su pobre preservación. Éstas se encuentran junto a Araneograptus murrayi. El género Coryphidium aparece por primera vez en la sub-asociación 3 (Cooper et al., 1995), del Tremadociano tardío y por lo tanto su presencia, podría indicar la relación con esta sub-asociación. Los niveles superiores de la Formación Parcha, correspondientes a la Biozona de Hunnegraptus copiosus, presentan mayor diversidad de acritarcos, muchos de los cuales caracterizan la microflora de messaoudensis-trifidum, típica de la transición tremadociana-arenigiana. La relación con H. copiosus confirma una edad tremadociana terminal para los niveles portadores.
Por lo tanto, en contraposición a lo que se supuso originalmente, la edad de O. araiorhachis es tremadociana tardía y no arenigiana. Asimismo, su distribución no está restringida a la parte superior de la Formación Parcha como tradicionalmente se acepta, sino que se extiende desde el tramo cuspidal de la Formación Saladillo hasta la parte superior de la Formación Parcha.
Fauna de trilobites
Las especies de trilobites asociados a O. araiorhachis y su rango vertical se detallan en la figura 4. Los rangos de Thysanopyge taurinus (Harrington) y Bienvillia parchaensis Harrington y Leanza coinciden con el de O. araiorhachis, estando sus primeros registros en los tramos superiores de la Formación Saladillo y los últimos en la parte superior de la Formación Parcha. Hypermecaspi inermis Harrington y Leanza e Hypermecaspis armata Harrington y Leanza aparecen asociados a O. araiorhachis sólo en la parte inferior de la Formación Parcha (Biozona de A. murrayi) mientras que Kayseraspis sp. se asocia a la última aparición de O. araiorhachis en la parte alta de la Biozona de H. copiosus. Bienvillia kichka sp. nov., Megistaspis (Ekeraspis)? sp., e Hypermecaspis sp. están restringidos a la parte media de la Formación Saladillo (Biozona de Kiaerograpus). En la parte superior de la Formación Saladillo y dentro de esta última biozona se registran Bienvillia jana sp. nov. e Hypermecaspis sp. nov. La edad de las especies mencionadas se refiere también al Tremadociano tardío.
Cabe destacar que no contamos, hasta el momento, con registros de Araiopleura reticulata Harrington y Leanza y Raphiphorus? lamasi Harrington y Leanza. De acuerdo a Harrington y Leanza (1957) ambos taxones están asociados a O. araiorhachis en la parte superior de la Formación Parcha. Por otro lado, en la parte superior de la Formación Parcha se registra Psilocara liliae Tortello dentro de la Biozona de H. copiosus (Toro et al., 2003), indicando una edad tremadociana tardía para esta especie. Hasta el presente no contamos con registros de edad arenigiana temprana para P. liliae (cf. Tortello, 1999).
Discusión
Las consideraciones anteriores respecto a la Biozona de O. araiorhachis ponen de manifiesto una serie de aspectos importantes a tener en cuenta al emplear el esquema establecido por Harrington (en Harrington y Leanza, 1957). En primer lugar, es necesaria una revisión taxonómica con criterios de clasificación modernos de los taxones involucrados en el esquema. Tanto en este caso, como en aquéllos derivados de revisiones anteriores (Vaccari, 1993; Vaccari y Waisfeld, 1994; Waisfeld, 1996) existen confusiones taxonómicas, por las cuales formas consideradas como coespecíficas corresponden, en realidad, a especies, o aún a géneros diferentes. Esta situación es recurrente al considerar especies procedentes de distintas localidades. Determinaciones taxonómicas equívocas de especimenes de distintas localidades produjo, como consecuencia directa, correlaciones erróneas entre las unidades portadoras de los taxones.
En algunas de las biozonas propuestas en 1957 la asignación de edades se basó en supuestas relaciones estratigráficas entre las unidades portadoras. Dado que prácticamente ninguna de las secciones expone las unidades en forma completa, y que presentan significativa complejidad estratigráfica y estructural, esta práctica condujo a fuentes de error adicionales. Por su naturaleza esencialmente bentónica, la distribución de los trilobites está fuertemente controlada por las condiciones de fondo. Por ello, la definición de biozonas no sólo debe llevarse a cabo bajo un estricto control ecológico-ambiental, sino también resulta imperativa la calibración de edades a partir de datos cronológicos independientes. En tal sentido, los trilobites con algún valor bioestratigráfico deben evaluarse en conjunto con grupos de amplia distribución como graptolitos, conodontes o palinomorfos.
Como se mencionó precedentemente, el esquema de biozonas de 1957 plantea dos asociaciones sucesivas para el Arenigiano: la Biozona de Kayseraspis asaphelloides y la Biozona de Ogygiocaris araiorhachis. La Biozona de K. asaphelloides está restringida a la Formación San Bernardo que aflora en el cerro San Bernardo. Esta unidad fue tradicionalmente considerada de edad arenigiana sobre la base de la propuesta original de Harrington (en Harrington y Leanza,1957) y de los estudios de graptolitos de Loss (1951). Moya (1998) sugiere una edad tremadociana tardía-arenigiana media para esta unidad y, más recientemente, Monteros y Moya (2003) dan a conocer una fauna de graptolitos de edad tremadociana tardía para la parte inferior de la Formación San Bernardo (Biozona de Kiaerograptus y parte inferior de la Biozona de A. murrayi/ pulchellus). Estos autores mencionan el registro de formas de trilobites de la Fauna de Thysanopyge asociada a los graptolitos estudiados (Thysanopyge argentina Kayser, Kayseraspis asaphelloides Harrington y Leanza, Kayseraspis brackebuschi Harrington y Leanza, Sanbernardaspis pygacantha Aceñolaza).
De acuerdo a nuestros datos, en la Formación San Bernardo K. asaphelloides está asociado a acritarcos y quitinozoos, escasos en número y diversidad y no muy bien preservados, lo que dificulta su asignación sobre todo a nivel específico. La asociación de acritarcos contiene Caldariola glabra var. glabra, formas intermedias entre Cymatiogalea messaoudensis y Cymatiogalea deunffi, Acanthodiacrodium spp., Vavrdovella sp. y formas intermedias entre Acanthodiacrodium y Vavrdovella. Estos taxones evidencian la presencia de la asociación de messaoudensis-trifidum en esta localidad. El conjunto de los mismos indicaría la relación con las sub-asociaciones 1 ó 2 (Cooper et al., 1995). En Inglaterra (English Lake District), estas subasociaciones se relacionan a la Biozona de A. murrayi.
Si se toma en cuenta como primer registro de Coryphidium (Coryphidium? sp.) el correspondiente al tramo inferior de la Formación Parcha en la Biozona de Araneograptus murrayi (figura 4), los acritarcos de la Formación San Bernardo se ubicarían posiblemente por debajo de los acritarcos del tramo inferior de la Formación Parcha. Como alternativa, si se considera como la primera aparición para el género Coryphidium la correspondiente a los niveles medios a superiores de la Formación Parcha (Biozona de H. copiosus), los acritarcos de los niveles inferiores de Parcha y los de la Formación San Bernardo podrían ser relativamente coetáneos.
En consecuencia, también la Biozona de K. asaphelloides debe ser considerada tremadociana superior y no arenigiana inferior.
Las biozonas de K. asaphelloides y de O. araiorhachis integran la Fauna de Thysanopyge. La Fauna de Thysanopyge es una entidad ecológica, representada por asociaciones de trilobites diferentes en distintas localidades y asociadas a distintas especies de Thysanopyge (Waisfeld et al., 1999; Waisfeld y Astini, 2003). Los registros más antiguos de la Fauna de Thysanopyge son los del cerro San Bernardo (Formación San Bernardo), asociados a T. argentina y los de Pascha, asociados a T. taurinus (Formaciones Saladillo y Parcha), mientras que el registro más joven corresponde al Arenigiano inferior alto (Biozona de B. deflexus, Toro, 1999) y se documenta en la localidad de Santa Victoria (Formación Acoite), con T. calvijoi como especie de referencia.
Las biozonas de O. araiorhachis y de K. asaphelloides representan un buen ejemplo de la composición heterogénea de la Fauna de Thysanopyge. La asociación de O. araiorhachis está dominada exclusivamente por trilobites asáfidos y olénidos, siendo la presencia de otros grupos de carácter excepcional y restringida a ostrácodos y filocáridos. El registro de esta asociación en pelitas negras depositadas en un ambiente de plataforma externa por debajo del nivel de base de las olas, indica que la misma se habría desarrollado en un ambiente de baja energía, deficitario en oxígeno y sustratos de naturaleza fangosa. La abundancia de trilobites olénidos y la virtual exclusión de otros elementos bentónicos constituyen evidencias independientes de bajos niveles de oxigenación.
En contraste, de acuerdo a los datos disponibles hasta el momento, la asociación de K. asaphelloides está dominada por trilobites asáfidos (T. argentina Kayser, T. saltaensis (Kayser), K. brackebuschi (Kayser), Megaslaspidella kayseri Kobayashi), otras familias como Shumardiidae (Conophrys sp.) y Calymenidae ("Colpocoryphoides" trapezoidalis Harrington y Leanza) son francamente minoritarias, mientras que los olénidos están ausentes. Es remarcable la presencia de otros grupos fósiles tales como ostrácodos, braquiópodos rhynchonelliformes y linguliformes, bivalvos, gastrópodos, y restos de crinoideos. Esta asociación se registra en los tramos inferiores de la Formación San Bernardo en niveles de pelitas limosas verdes bioturbadas con delgadas intercalaciones de areniscas, depositadas posiblemente en un ambiente de plataforma interna, bien oxigenada y de moderada energía.
Las consideraciones anteriores indican que el desarrollo de las asociaciones que caracterizan las biozonas de K. asaphelloides y O. araiorhachis no sería el resultado de un verdadero control temporal dado que las diferencias en este sentido son sutiles (ambas se ubican dentro de la biozona de A. murrayi y de la asociación de messaoudensis-trifidum). Por lo tanto, estas asociaciones no representarían una secuencia temporal de biozonas, sino serían expresiones ecológicas de variaciones laterales del ambiente. Estas variaciones podrían haber estado relacionadas, por ejemplo, a diferencias en los niveles de oxigenación, características del sustrato, energía del medio, etc. Posiblemente, las condiciones de oxigenación del fondo hayan sido el factor con mayor influencia. Esta alternativa se sustenta en el contraste en la composición taxonómica dentro del grupo de los trilobites, en la distribución de otros grupos fósiles bentónicos, y en las diferentes litofacies en las que se habrían desarrollado las asociaciones de O. araiorhachis y K. asaphelloides.
Asimismo, el patrón de distribución espacial de estas "biozonas", restringidas geográficamente a localidades particulares, y las marcadas diferencias composicionales entre ambas, podría ser explicado por la respuesta diferencial de las especies de trilobites frente a los factores ambientales mencionados. En consecuencia, en contraposición a otros taxones de trilobites de amplia distribución (e.g. Parabolina (Neoparabolina), Jujuyaspis, Kainella) empleados en el esquema de 1957, O. araiorhachis y K. asaphelloides poseen un limitado valor desde el punto de vista bioestratigráfico y las biozonas homónimas resultan de poca utilidad para establecer correlaciones regionales.
Conclusiones
A partir de la presente revisión, la distribución geográfica de la Biozona de Ogygiocaris araiorhachis queda restringida al área de Pascha-Incamayo. Referencias previas de esta especie en la región de Santa Victoria y Potrero de Castillo corresponden a asignaciones taxonómicas erróneas. O. araiorhachis no se registra en la parte superior de la Formación Parcha como se supuso previamente, su primera aparición se documenta en la parte superior de la Formación Saladillo, y su rango se extiende a la parte media-superior de la Formación Parcha.
Los datos aportados por la fauna de graptolitos y asociaciones palinológicas estudiados indican una edad tremadociana tardía para las formaciones Parcha y Saladillo, coincidente con la edad propuesta por Ortega y Albanesi (2003). En estas unidades se han podido documentar las biozonas de Bryograptus, Kiaerograptus, Araneograptus murrayi y Hunnegraptus copiosus. Cabe destacar que la especie Kiaerograptus supremus comienza a registrarse junto a Araneograptus murrayi, en los niveles superiores de la Formación Saladillo. Esto permite cuestionar la validez local de la Biozona Kiaerograptus supremus.
Por su parte, la asociación de acritarcos en el Tremadociano superior confirma la presencia de la microflora de messaoudensis-trifidum para la Cordillera Oriental (Rubinstein y Toro, 2002; Toro et al., 2003). De los numerosos registros de la asociación de messaoudensis-trifidum a través de peri-Gondwana, sólo unos pocos tienen dataciones independientes basadas en otros grupos fósiles. Los estudios bioestratigráficos integrados de graptolitos y acritarcos se encontraban hasta ahora limitados a Inglaterra y España. Los acritarcos de la región de Pascha- Incamayo y su relación con las biozonas de graptolitos proveen, por lo tanto, una de las pocas dataciones independientes para esta asociación.
En relación a la calibración cronológica de la Biozona de O. araiorhachis, su edad no es arenigiana como se la consideró clásicamente, sino tremadociana tardía, extendiéndose su registro entre las biozonas de A. murrayi y H. copiosus. A esta misma edad se refiere aquí la fauna de trilobites asociada a O. araiorhachis: T. taurinus, Kayseraspis sp., Bienvillia parchaensis, Bienvillia jana sp. nov., Hypermecaspis inermis, e Hypermecaspis armata.
La biozona de K. asaphelloides requiere aún una revisión detallada tanto de las especies que la integran como de sus distribuciones estratigráficas. El registro de graptolitos del Tremadociano tardío asociados a especies de trilobites presentes en la Biozona de K. asaphelloides (cf. Monteros y Moya, 2003) y la estrecha asociación de esta especie con la microflora de messaoudensis- trifidum (probablemente con las subasociaciones más antiguas) indican una edad tremadociana tardía para esta biozona.
Las biozonas de K. asaphelloides y O. araiorhachis propuestas en el esquema de Harrington (en Harrington y Leanza,1957) poseen un limitado valor desde el punto de vista bioestratigráfico, dado que no representarían una verdadera secuencia temporal de biozonas. Estas asociaciones corresponderían a expresiones ecológicas de restringida distribución geográfica en respuesta a cambios laterales del ambiente, posiblemente ligadas a diferentes niveles de oxigenación.
Agradecimientos
Los autores agradecen los comentarios de Gladys Ortega y Edsel Brussa con motivo de la revisión de este trabajo. ANPCyTFONCyT (PICT 2000 Nº 8920) y CONICET (PEI Nº 6227) aportaron el financiamiento para la realización de este trabajo.
Bibliografía
Aceñolaza, F.G. 1983. The Tremadocian beds and the Cambrian- Ordovician boundary problems in Latin America. Papers on Cambrian-Ordovician Boundary Symposium, Nanjing, pp. 88-93. [ Links ]
Albanesi, G.L., Ortega G., Tortello, M.F. y Aceñolaza, G.F. 1997. Conodontes, graptolitos y trilobites de la Formación Parcha en la Cordillera Oriental de Salta, Argentina. Ameghiniana 34: 114. [ Links ]
Albanesi, G.L., Ortega, G. y Zeballo, F.J. 2001. Late Tremadocian conodont-graptolite biostratigraphy from NW Argentine basins. The Guide Book, abstracts & Ordovician Silurian correlation Chart for the joint field meeting IGCP 410 and IGCP 421, Mongolia, pp. 121-123. [ Links ]
Astini, R.A. 2002. La megacapa Parcha (Ordovícico temprano de la Cordillera Oriental): ¿Sismita, regresión forzada o evento extraordinario?. 9º Reunión Argentina de Sedimentología, Resúmenes, p. 48. [ Links ]
Astini, R.A. 2003. The Ordovician Proto-Andean basins. En: J.L. Benedetto (ed.), Ordovician fossils of Argentina. Secretaría de Ciencias y Tecnología, Universidad Nacional de Córdoba, pp. 1-74. [ Links ]
Astini, R.A. 2005 Las unidades sedimentarias que reposan en no concordancia sobre el granito rojo en el angosto de la Quesera (Cordillera Oriental, Salta) y su interpretación estratigráfica: una revisión crítica a más de 60 años de los trabajos pioneros de J. Keidel. Revista de la Asociación Geológica Argentina, 60: 513- 523. [ Links ]
Benedetto, J.L. 1976. Una nueva especie del género Hoekaspis Kobayashi de la Provincia de Jujuy, con consideraciones acerca de las faunas llanvirnianas de Argentina y Bolivia. Ameghiniana 13: 300-311. [ Links ]
Benedetto, J.L. y Malanca, S.M. 1975. Los trilobites Ordovícicos de Los Colorados (Departamento Tumbaya, Provincia de Jujuy). 1º Congreso de Paleontología y Bioestratigrafía (San Miguel de Tucumán), Actas 1: 145-173. [ Links ]
Cooper, R.A., Rushton, A.W.A., Molyneux, S.G., Hughes R.A., Moore R.M. y Webby, B.C. 1995. The stratigraphy, correlation, provenance and palaeogeography of the Skiddaw Group (Ordovician) in the English Lake District. Geological Magazine 132: 185- 211. [ Links ]
Egenhoff, S.O., Maletz, J. y Erdtmann, B.-D. 2004. Lower Ordovician graptolite zonation and lithofacies of southern Bolivia: relevance for palaeogeographic interpretations. Geological Magazine 141: 287-299. [ Links ]
Fensome, R.A., Williams, G.L., Barss, M.S., Freeman, J.M., y Hill, J.M. 1990. Acritarchs and fossil prasinophytes: an index to genera, species, and infraspecific taxa. American Association of Stratigraphic Palynologists Foundation (AASP). Contribution Series nº 25, p. 771. [ Links ]
Harrington, H.J. 1937. On some fossils from northern Argentina. Geological Magazine 74: 97-124. [ Links ]
Harrington, H.J. 1938. Sobre las faunas del Ordoviciano inferior del norte Argentino. Revista del Museo de La Plata, Sección Paleontología 4: 109-289. [ Links ]
Harrington, H.J. y Leanza, A. 1957. Ordovician trilobites of Argentina. University of Kansas Press, Lawrence, 276 pp. [ Links ]
Jardiné, S., Combaz, A., Magloire, L., Peniguel, G. y Vachey, G. 1974. Distribution stratigraphique des acritarches dans le Paéozoïque du Sahara algérien. Review of Palaeobotany and Palynology 18: 99- 129. [ Links ]
Keidel, J. 1937. La Prepuna de Salta y Jujuy. Revista Centro de Estudios Doctorales de la Facultad de Ciencias Naturales 1: 125- 154. [ Links ]
Keidel, J. 1943. El Ordovícico inferior en los Andes del norte argentino y sus depósitos marino-glaciales. Boletín de la Academia Nacional de Ciencias 36: 140-229. [ Links ]
Kobayashi, T. 1935. The Kainella fauna of the basal Ordovician age found in Argentina. Japanese Journal Geology and Geography 12: 59-67. [ Links ]
Kobayashi, T. 1937. The Cambro-Ordovician shelly faunas of South America. Journal of the Faculty of Science, Imperial University of Tokyo (section 2) 4: 369-522. [ Links ]
Lindholm, K. 1991. Ordovician graptolites from the early Hunneberg of southern Scandinavia. Palaeontology 34: 283-327. [ Links ]
Loeblich, A.R. y Tappan, H. 1976. Some new and revised organicwalled phytoplankton microfossil genera. Journal of Paleontolology 50: 301-308. [ Links ]
Loss, R. 1951. Contribuciones al conocimiento de las faunas graptolíticas del Norte argentino, I: Graptolites del Cerro San Bernardo (Salta) y zona del dique La Ciénaga (Jujuy). Revista de la Asociación Geológica Argentina 6: 21-61. [ Links ]
Maletz, J. y Egenhoff, S.O. 2001. Late Tremadoc to early Arenig graptolite faunas of southern Bolivia and their implications for a worldwide biozonation. Lethaia 34: 47-62. [ Links ]
Molyneux, S.G. y Rushton A.W.A. 1988. The age of the Watch Hill Grits (Ordovician, English Lake District: structural and palaeogeographical implications. Transactions of the Royal Society of Edinburgh: Earth Sciences 79: 43-69. [ Links ]
Monteros, J.A. y Moya, M.C. 2002. La zona de Aorograptus victoriae (Ordovícico Inferior) en la sierra de Mojotoro, Cordillera Oriental argentina. 8° Congreso Argentino de Paleontología y Bioestratigrafía, Resúmenes p. 88. [ Links ]
Monteros, J.A. y Moya, M.C. 2003. Late Tremadocian graptolites from the Mojotoro Range, Argentine Eastern Cordillera. En: G. Ortega y G.F. Aceñolaza, (eds.), Proceedings of the 7º Internacional Graptolite Conference. Serie Correlación Geológica 18: 73-78. [ Links ]
Moya, M.C. 1998. El Paleozoico inferior en la sierra de Mojotoro, Salta - Jujuy. Revista de la Asociación Geológica Argentina 53: 219- 238. [ Links ]
Moya, M.C. 1999. El Ordovícico en los Andes del norte argentino. En: G. González Bonorino, R. Omarini y J. Viramonte (eds.), 14º Congreso Geológico Argentino, Geología del noroeste argentino, Relatorio 1: 134-152. [ Links ]
Moya, M.C., Malanca, S., Monteros, J.A. y Cuerda, A.J. 1994. Bioestratigrafía del Ordovícico Inferior en la Cordillera Oriental Argentina basada en graptolitos. Revista Española de Paleontología 9: 91-104. [ Links ]
Ortega, G. y Albanesi, G.L. 2002. Bioestratigrafía de graptolitos y conodontes del Tremadociano tardío de la Cordillera Oriental Argentina. 15º Congreso Geológico Argentino (el Calafate), Actas 1: 542-547. [ Links ]
Ortega, G. y Albanesi, G.L. 2003. Late Tremadoc graptolite sequence and conodonts from the Parcha area, Eastern Cordillera, Argentina. En: G. Ortega y G.F. Aceñolaza (eds.), Proceedings of the 7º Internacional Graptolite Conference. Serie Correlación Geológica 18: 79-86. [ Links ]
Ortega, G., Tortello, M.F., Rao, R.I. y Aceñolaza, F.G. 1997. Faunas del Ordovícico Inferior en el área del Angosto de Lampazar, Cordillera Oriental, provincia de Salta, Argentina. Ameghiniana 34: 123. [ Links ]
Playford, G. y Martin, F. 1984. Ordovician acritarchs from the Canning Basin, Western Australia. Alcheringa 8: 187-223. [ Links ]
Ramos, V.A. 1973. Estructura de los primeros contrafuertes de la Puna saltojujeña y sus manifestaciones volcánicas asociadas. Revista de la Asociación Geológica Argentina 27: 84-89. [ Links ]
Rao, R.I. y Tortello, M.F. 1998. Tremadoc conodonts and trilobites from the Cardonal Formation, Incamayo Creek, Salta Province, northwestern Argentina. Palaeontologia Polonica 58: 31-45. [ Links ]
Rubinstein, C.V. y Toro, B.A. 2002. The messaoudensis-trifidum acritarch assemblage (late Tremadoc-early Arenig) from Parcha, Eastern Cordillera, northwestern Argentina. CIMP International Meeting and Workshops, Lille, France. Palaeozoic Palynology in the Third Millenium: new directions in acritarch, chitinozoan and miospore research. Abstract Volume, p. 48. [ Links ]
Rubinstein, C.V., Toro, B.A. y Waisfeld, B.G. 1999. Acritarch biostratigraphy of the upper Tremadoc-Arenig of the Eastern Cordillera, northwestern Argentina: relationships with graptolite and trilobite fauna. Bolletino della Società Paleontologica Italiana 38: 267- 286. [ Links ]
Servais, T. y Eiserhardt, K.H. 1995. A discussion and proposals concerning the Lower Paleozoic "galeatre" acritarch plexus. Palynology 19: 191-210. [ Links ]
Servais, T. y Mette W. 2000. The messaoudensis-trifidum acritarch assemblage (Ordovician: late Tremadoc-early Arenig) of the Barriga Shales Formation, Sierra Morena (SW-Spain). Review of Palaeobotany and Palynology 113: 145-163. [ Links ]
Servais, T. y Molyneux, S. G. 1997. The messaoudensis-trifidum acritarch assemblage (Ordovician: late Tremadoc-early Arenig) from the subsurface of Rügen (Baltic Sea, NE Germany). Palaeontographia Italica 84: 113- 161. [ Links ]
Toro, B.A. 1999. Nuevos datos paleontológicos (Graptolithina), bioestratigráficos y paleogeográficos del área de Santa Victoria, Cordillera Oriental, Argentina. Ameghiniana 36: 477- 487. [ Links ]
Toro, B.A., Rubinstein, C.V., Waisfeld, B.G. y Astini, R.A. 2003. Calibración de las asociaciones de palinomorfos y trilobites, con las zonas de graptolitos del Ordovícico temprano del área de Parcha, Cordillera Oriental, Argentina. Ameghiniana 40 (supl.): 94-95R. [ Links ]
Tortello, M.F. 1999. El género Psilocara Fortey (Trilobita, Olenidae) en la Formación Parcha (Ordovícico temprano), Salta, Argentina. Ameghiniana 36: 339-344. [ Links ]
Tortello, M.F. y Rao, R.I. 2000. Trilobites y conodontes del Ordovícico temprano del Angosto de Lampazar (provincia de Salta, Argentina). Boletín Geológico y Minero 111: 61-84. [ Links ]
Vaccari, N.E. 1993 El género Annamitella Mansuy, 1920 (Trilobita, Leiostegiidae) en el Ordovícico de la Precordillera Argentina. Ameghiniana 30: 395-405. [ Links ]
Vaccari, N.E. y Waisfeld, B.G. 1994. Nuevos trilobites de la Formación Suri (Ordovícico Inferior) en la región de Chaschuil, Provincia de Catamarca. Implicancias bioestratigráficas. Ameghiniana 31: 72-86. [ Links ]
Vilela, C. R. 1956. Descripción Geológica de la hoja 7d, Rosario de Lerma (provincia de Salta). Boletín de la Dirección Nacional de Minería 84, 58pp. [ Links ]
Waisfeld, B.G. 1996. Revisión de la Zona de "Hoekaspis schlagintweiti" Harrington y Leanza, Ordovícico del Noroeste de Argentina. 12º Congreso Geológico de Bolivia (Tarija), Actas 3: 915-921. [ Links ]
Waisfeld, B.G. y Astini, R.A. 2003. Environmental constraints in faunal patterns: an example in Early Ordovician (Arenig) trilobite assemblages from the Argentine Cordillera Oriental. En: G.L. Albanesi, M.S. Beresi y S.H. Peralta (eds.), Ordovician from the Andes. Serie Correlación Geológica 17: 341-346. [ Links ]
Waisfeld, B.G. y Vaccari, N.E. 2003. Trilobites. En: J.L. Benedetto (ed.), Ordovician Fossils of Argentina. Secretaría de Ciencia y Tecnología, Universidad Nacional de Córdoba, pp. 295-410. [ Links ]
Waisfeld, B.G. y Vaccari, N.E. 2006. Revisión de la Biozona de Ogygiocaris araiorhachis (Trilobita, Tremadociano tardío) en la región de Pascha-Incamayo, Cordillera Oriental, Argentina. Parte 2: Sistemática. Ameghiniana 43: 729-745. [ Links ]
Waisfeld, B.G., Sánchez, T.M. y Carrera, M.G. 1999. Biodiversification patterns in the Early Ordovician of Argentina. Palaios 14: 198-214. [ Links ]
Waisfeld, B.G., Vaccari, N.E. y Toro, B.A. 2003. Hacia un nuevo esquema bioestratigráfico basado en trilobites para el Ordovícico del noroeste de Argentina. Revisión de la Zona de Ogygiocaris araiorhachis. Ameghiniana Suplemento Resúmenes 40: 97R. [ Links ]
Zhang, Y. and Chen, X. 2003. The Early-Middle Ordovician graptolite sequence of the Upper Yangtze region, South China. En: G.L. Albanesi, M.S. Beresi y S.H. Peralta (eds.), Ordovician from the Andes. Serie Correlación Geológica 17: 173-180. [ Links ]
Recibido: 9 de mayo de 2005.
Aceptado: 23 de noviembre de 2005.

