










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.1 Buenos Aires ene./mar. 2007
Los Metatheria sudamericanos de comienzos del Neógeno (Mioceno Temprano, Edad-mamífero Colhuehuapense). Parte I: Introducción, Didelphimorphia y Sparassodonta
Francisco Goin1, Alejandra Abello1, Eduardo Bellosi3, Richard Kay2, Richard Madden2 y Alfredo Carlini1
1División Paleontología Vertebrados, Museo de la Plata, Paseo del Bosque s /n, 1900 La Plata, Argentina. fgoin@museo.fcnym.unlp.edu.ar, mabello@museo.fcnym.unlp.edu.ar, acarlini@museo.fcnym.unlp.edu.ar.
2Department of Biological Anthropology, School of Arts and Sciences, Duke University, Box 3170, DUMC, Durham NC 27710, USA. rich_kay@baa.duhs.duke.edu, rick_madden@baa.duhs.duke.edu.
3Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Avenida Angel Gallardo 470, 1405 Buenos Aires, Argentina. ebellosi@sei.com.ar
Abstract. South american Metatherians from the begining of the Neogene (Early Miocene, Colhuehuapan Land-Mammal Age). Part 1: Introduction, Didelphimorphia and Sparassodonta. This work is the first, of a series of three, taxonomic reviews of the Early Miocene (Colhuehuapian Land-mammal Age) South American metatherians. Colhuehuapian levels from the Gran Barranca south of the Colhue Huapi lake (Sarmiento Department, Chubut Province, Argentina) are the richest ones in specimens of this age. Colhuehuapian landscapes from Central Patagonia developed in a coastal plain located in a peninsular area, which originally suffered an intense erosion and subsequentely was agradded with fluvial and loessic (eolian) sediments. Palinomorphs and phytoliths suggest a period characterized by significant environmental changes, from xeric (middle Early Miocene) to humid-warm conditions (late Early Miocene). In uplands or areas far from the sea, temperate-humid closed forests prevailed, but in lowlands or litoral areas herbaceous (grass) plants and shrubs dominated, with subordinated or patchy arboreal vegetation. Paleoclimate would have been warm-temperate to warm and probably subhumid, more rainy in highlands. The most important results of this analysis are: (1) the oldest record of Didelphoidea didelphimorphians, including, probably, the oldest known caluromyid; (2) the recognition of a great diversity of carnivores belonging to the Order Sparassodonta, including the oldest record of a Thylacosmilidae; (3) the recognition of a new species of the incertae sedis mammal Necrolestes. Among the taxa reviewed here we describe the new species Necrolestes mirabilis.
Resumen. Este trabajo constituye el primero, de una serie de tres, dedicado a revisar la taxonomía de los Metatheria del Mioceno Temprano (Edad-mamífero Colhuehuapense) de América del Sur. Los niveles colhuehuapenses de la Gran Barranca del Lago Colhue Huapi (Departamento de Sarmiento, Provincia del Chubut, Argentina) son los más ricos en especímenes de esta edad. Los paisajes colhuehuapenses de Patagonia central se desarrollaron en la planicie costera de un área peninsular, inicialmente sometida a una intensa erosión y luego agradada por sedimentos fluviales y loéssicos (eólicos). Los registros de la vegetación (palinomorfos y fitolitos) sugieren una etapa de importantes cambios ambientales, desde condiciones xéricas a cálido-húmedas. En zonas elevadas o alejadas del mar prevalecieron bosques templado-húmedos, mientras que en áreas bajas-litorales dominaron hierbas (gramíneas) y arbustos con vegetación arbórea subordinada o en parches. El clima habría sido templado-cálido a cálido y posiblemente subhúmedo, con mayores precipitaciones en zonas elevadas. Del análisis taxonómico se destacan: (1) el registro más antiguo de didelfimorfios Didelphoidea, incluyendo, probablemente, el más antiguo calurómido, (2) la gran diversidad de carnívoros del Orden Sparassodonta, incluyendo el más antiguo registro de un Thylacosmilidae; (3) el reconocimiento de una nueva especie del mamífero incertae sedis Necrolestes. Entre los taxones aquí revisados se describe la nueva especie Necrolestes mirabilis.
Key words: South America; Patagonia; Early Miocene; Colhuehuapian Age; Marsupialia; Didelphimorphia; Sparassodonta.
Palabras clave: América del Sur; Patagonia; Mioceno Temprano; Edad Colhuehuapense; Marsupialia; Didelphimorphia; Sparassodonta.
Introducción
Desde mediados de la década de 1990 un equipo integrado por investigadores de la Duke University, el Museo de La Plata, el Museo Argentino de Ciencias Naturales y la Universidad de Buenos Aires ha venido realizando excavaciones sistemáticas en niveles correspondientes a la transición Paleógeno- Neógeno en la Patagonia central argentina. Los trabajos han cubierto aspectos estratigráficos, sedimentológicos, geocronológicos y paleontológicos en una escala nunca antes abordada, lo que ha permitido, entre otros aspectos, la recuperación de miles de restos fósiles de vertebrados. Es así que los hallazgos de mamíferos colhuehuapenses (Mioceno Temprano; Edad-mamífero Colhuehuapense), llevados a cabo en sucesivas campañas a lo largo de la pasada década, han permitido aumentar significativamente nuestro conocimiento taxonómico y bioestratigráfico sobre este grupo de vertebrados de comienzos del Neógeno. En el caso de los metaterios esto último es particularmente notable, ya que hasta el momento buena parte de los taxones reconocidos para esta edad era de tamaño mediano a grande; tal es el caso de las formas carnívoras, los Sparassodonta Borhyaenidae. Sin embargo, la mayor parte de los metaterios sudamericanos fueron, tal como ocurre en la actualidad, formas de pequeño tamaño, por lo que sus restos son difíciles de observar en el campo a ojo desnudo. En años recientes, la aplicación de técnicas de concentrado sedimentario y posterior análisis bajo lupa binocular han permitido reconocer una diversidad hasta el momento insospechada (Goin y Candela, 2004). Con la excepción parcial de los carnívoros asignados al orden Sparassodonta, resulta mucho más significativa la información ahora disponible sobre el resto de los grupos de metaterios colhuehuapenses, grupos que, no casualmente, están representados por formas de pequeño tamaño.
La disponibilidad de numerosos especímenes como consecuencia de estos trabajos nos ha permitido encarar una exhaustiva revisión de los metaterios representativos de la Edad-mamífero Colhuehuapense. Si bien nuestro análisis se centra en los materiales recolectados en la Patagonia argentina, debe señalarse que, hasta el momento, esta región incluye (al menos en el registro édito) las únicas localidades sudamericanas portadoras de mamíferos incuestionablemente referibles a la Edad-mamífero Colhuehuapense (figura 1.A). Queda clara, entonces, la parcialidad del registro, que sólo nos permite vislumbrar una parte de la radiación neógena de los metaterios sudamericanos: aquella restringida a su porción austral o, en términos de la reciente propuesta biogeográfica de Morrone (2002), a la porción sur de la Región Andina del Reino Austral. Los Didelphimorphia sudamericanos vivientes, por ejemplo, alcanzan su mayor diversidad en la parte tropical extra-andina de América del Sur (la Región Neotropical del Reino Holotropical sensu Morrone, 2002), circunstancia que probablemente ocurrió también a comienzos del Neógeno en este continente. Es así que el capítulo neotropical de los metaterios sudamericanos (como así también de muchos otros grupos de vertebrados) está aún por escribirse.

Figura 1. A, Localidades patagónicas con faunas de edad Colhuehuapense mencionadas en el texto / Patagonian localities including faunas of Colhuehuapian age, mentioned in the text; B, Detalle de la Gran Barranca al sur del Lago Colhue Huapi / detail of the Gran Barranca south of Lake Colhue Huapi.
Este trabajo constituye la primera entrega correspondiente a la revisión en curso de los metaterios sudamericanos de la Edad-mamífero Colhuehuapense y comprende el análisis de los Didelphimorphia y Sparassodonta, más una discusión sobre la situación sistemática de los Necrolestidae -grupo del cual se registra una nueva especie- y que fuera previamente referido a los Sparassodonta por Patterson (1958). Una segunda entrega comprenderá los Microbiotheria y Polydolopimorphia, mientras que la última incluirá a los Paucituberculata, más una discusión general sobre el conjunto de la asociación de metaterios. El concepto de Metatheria aquí utilizado sigue a Rougier et al. (1998). Sobre la extensión y contenidos de los órdenes Didelphimorphia, Polydolopimorphia y Paucituberculata, en este trabajo se sigue el criterio taxonómico sustentado por Goin (1991, 2003) y Goin y Candela (2004).
Abreviaturas. Acrónimos: DU, Duke University, Durham; FMNH, Field Museum of Natural History, Chicago; MACN, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires; MLP, División Paleontología Vertebrados, Museo de La Plata; MNHN, Muséum National d'Histoire Naturelle, Paris; MPEF-PV, Sección Paleontología Vertebrados, Museo Paleontológico "Egidio Feruglio", Trelew; UNSW, University of New South Wales, Sydney. Otras abreviaturas: L, largo; A, ancho; Ma, millones de años antes del presente; I/i, C/c, P/p, M/m, incisivos, caninos, premolares y molares superiores/inferiores, respectivamente; dP3/dp3, premolares deciduos superior e inferior, respectivamente. M/mx, molar de locus incierto. Las homologías de cúspides y crestas adoptadas en este trabajo siguen a Goin et al. (2003). Todas las medidas craneomandibulares y dentarias son en milímetros (mm). Los espesores estratigráficos están expresados en metros (m).
Antecedentes, localidades y contextos geológico y paleoambiental
Sobre la edad Mioceno Temprano de la Edad-mamífero Colhuehuapense
Desde mediados del siglo pasado y hasta comienzos de la década de 1980, la asociación faunística colhuehuapense (el "Colpodenèen" de Ameghino) fue referida al Oligoceno Tardío (e.g., Simpson, 1948; Marshall et al., 1983). Pocos años después Marshall et al. (1986) propusieron su asignación al Mioceno Temprano-Medio en base al hallazgo de fauna deseadense en depósitos datados radiométricamente en el Mioceno más temprano (Scarritt Pocket, provincia del Chubut). Marshall et al. (1986) consideraron que la Edad-mamífero Colhuehuapense resultaba de corta duración, estando comprendida entre los 19 y 18 Ma. Esta asignación temporal fue en parte corroborada por las primeras y más precisas dataciones de depósitos colhuehuapenses en Gran Barranca (Kay et al., 1999a), las cuales indican una edad entre 20 y 19 Ma (Burdigaliano Temprano; véase más abajo).
Estudios previos sobre los metaterios colhuehuapenses
No existía, hasta el momento, una revisión comprehensiva de los metaterios referibles a la Edad-mamífero Colhuehuapense. Sin embargo, su estudio se remonta a los comienzos del siglo XX con trabajos fundacionales de Florentino Ameghino, quien realizó un primer listado de estos metaterios a partir de los restos fósiles colectados por Carlos Ameghino en el año 1898 en las "Capas con Colpodon" o "Piso Colpodonense" (Ameghino, 1900-1902, 1902, 1906). Las nuevas especies reconocidas por F. Ameghino (1902) son actualmente consideradas representantes de los órdenes Paucituberculata, Microbiotheria y Sparassodonta. A este trabajo pionero le siguió el realizado por Simpson (1932) quien estudió los ejemplares colectados por la Scarritt Patagonian Expedition, dando a conocer tres nuevas especies pertenecientes a Microbiotheriidae y Paucituberculata. Además de los ya citados, entre los trabajos puntuales sobre la sistemática de distintos grupos de metaterios de esta edad se destacan los de Sinclair (1930), Ringuelet (1953), Reig (1955) y Marshall (1978, 1980, 1982a). Durante las últimas dos décadas del siglo XX las adiciones más importantes al conocimiento de los metaterios colhuehuapenses fueron el reconocimiento de una nueva superfamilia, Patagonioidea (Pascual y Carlini, 1987), el primer registro de Argyrolagidae para esta edad (Pascual y Carlini, 1985) y una revisión de los Paucituberculata Palaeothentidae (Bown y Fleagle, 1993), en donde se describen nuevos géneros y especies para esta Edad-mamífero.
Los mamíferos sudamericanos terrestres han probado ser una de las herramientas más útiles para el reconocimiento y secuenciación de eventos geológicos y biológicos durante el Cenozoico de América del Sur (Pascual et al., 1996). Por este motivo la diversidad y evolución de los metaterios colhuehuapenses también ha sido considerada en numerosos trabajos concernientes a la sucesión de Edades-mamífero (e.g., Pascual y Odreman Rivas, 1973; Marshall et al., 1983), así como también a los cambios climático-ambientales relacionados (e.g., Pascual y Odreman Rivas, 1971; Pascual et al., 1985; Pascual y Bond, 1986; Ortiz Jaureguizar, 1986; Pascual y Ortiz Jaureguizar, 1990; Pascual et al., 1996; Bellosi, 1995, 1996).
Localidades fosilíferas
La mayor parte de los mamíferos recuperados en niveles de la Edad-mamífero Colhuehuapense procede de una serie de sitios fosilíferos, muy cercanos entre sí, ubicados sobre la Barranca Sur del Lago Colhue Huapi (o Gran Barranca), en la Patagonia central argentina (figuras 1.A y B). Entre los Metatheria, el 95% de todos los especímenes estudiados en este trabajo procede de esta área. Unos pocos restos adicionales proceden de otras localidades, mayormente patagónicas, que incluyen niveles referibles a esta edad. A continuación se mencionan brevemente las localidades portadoras de mamíferos de edad colhuehuapense, la mayor parte de las cuales incluye los metaterios estudiados en este trabajo.
Gran Barranca. Sobre un total de más de 800 especímenes de metaterios aquí analizados, sólo 45 no proceden de la Barranca Sur, o Gran Barranca, del Lago Colhue Huapi (Departamento Sarmiento, provincia del Chubut). Las secciones portadoras de fauna colhuehuapense se exponen en el extremo occidental de la Gran Barranca (i.e., perfil "A" de Simpson, notas inéditas; Spalletti y Mazzoni, 1979: fig. 2 ; Kay et al., 1999a: fig. 1; véase la figura 2 de este trabajo). El nombre de "Puesto Almendra" constituye la clásica referencia geográfica a los afloramientos del Miembro Colhue Huapi en esta área. En rigor de verdad, con este nombre se conocen todos los afloramientos del Miembro Colhue Huapi accesibles desde donde termina la huella que pasa por las ruinas del viejo Puesto Almendra, hoy abandonado (figura 1.B). La zona de afloramientos correspondientes a dicho miembro en esta localidad no excede un kilómetro cuadrado, y durante los trabajos de campo fueron definidos distintos sitios específicos que señalan la procedencia de los materiales exhumados: "Nivel C", "Nivel C Gris", "Nivel B", "Mono Este Superficial", "Mono Oeste Superficial" y "Mono 2". El nombre "Puesto Almendra", sin embargo, sigue siendo la referencia generalizada entre los especímenes recolectados en expediciones previas a 1990, tal como consta en los rótulos de diversas colecciones (MLP, MACN, AMNH, etc.). Sobre la bibliografía paleontológica relativa a los metaterios y otros mamíferos colhuehuapenses en esta localidad, véase el punto anterior (Antecedentes).

Figura 2. Perfil estratigráfico esquemático de la Fm. Sarmiento en su localidad tipo (oeste de la Gran Barranca, véase la indicación del área del perfil en la figura 1.B). Modificado de Spalletti y Mazonni (1979) y Kay et al. (1999a). La sucesión colhuehuapense apoya en discordancia erosiva (valle fluvial inciso) sobre el Miembro Puesto Almendra (Bellosi et al., 2002). La flecha indica los niveles de procedencia de la mayor parte de los especímenes de marsupiales aquí estudiados / schematic stratigraphic profile of the Sarmiento Formation at its type locality (West of Gran Barranca; see the indication of the profile area in figure 1.B). Modified from Spalletti and Mazzoni (1979) and Kay et al. (1999a). The colhuehuapian beds lies in erosive unconformity (incised fluvial valley) over the Puesto Almendra Member (Bellosi et al., 2002). The arrow indicates the levels from which most marsupial specimens here studied come from.
Gaiman. Las principales localidades fosilíferas en esta región se ubican al sur del curso inferior del Río Chubut en las inmediaciones de Gaiman (Departamento de Gaiman, provincia del Chubut). Todas estas localidades corresponden a niveles del Miembro Trelew de la Fornación Sarmiento (véase más abajo y Simpson, 1935; Mendía y Bayarsky, 1981; Fleagle y Bown, 1983; Bown y Fleagle, 1993), constituyendo depósitos eólicos separados de los depósitos marinos costaneros de la suprayacente Formación Gaiman por una discordancia erosiva menor. Un total de 39 especímenes procede de niveles del Miembro Trelew en esta localidad. Bown y Fleagle (1993: 7 y Fig. 3.2) mencionan una rica localidad fosilífera, La Cantera, en niveles del Miembro Trelew en esta área. Más probablemente, esta localidad corresponde a la denominada "Lomas Blancas" o "Bryn Gwyn", tal como figura en los rótulos de los especímenes recolectados por las expediciones MLP-DU.
Sacanana. De la localidad de Sacanana, al oeste de Gan Gan (Departamento de Gastre, provincia del Chubut; véase Rusconi, 1933; Feruglio, 1949; Fleagle y Bown, 1983; Bown y Fleagle, 1993) han sido colectados numerosos restos fósiles sumamente concrecionados, lo que vuelve difícil su preparación e identificación taxonómica. Sobre la edad Colhuehuapense de estos niveles, véase Rusconi (1933). Procedente de un sitio cercano a esta localidad, se estudia aquí un ejemplar del Sparassodonta Acrocyon riggsi Sinclair. La estratigrafía de esta localidad es poco conocida, en tanto que las exposiciones son de poco relieve y desconectadas pero notoriamente fosilíferas (Fleagle y Bown, 1983). Ardolino et al. (1999) denominaron "Tobas colhuehuapenses de la Meseta de Somuncurá" a estas sucesiones piroclásticas, las cuales yacen sobre vulcanitas alcalinas del Oligoceno y están cubiertas por efusiones traquíticas de entre 22 y 18 Ma.
La Curandera. Referencias previas a esta localidad pueden consultarse en Scillato Yané (1977) y Marshall et al. (1983). Durante la campaña MLP-Duke de 1997 se distinguieron tres niveles fosilíferos en la zona de La Curandera (Departamento de Paso de Indios, provincia del Chubut). "La Curandera 1" corresponde a la parte estratigráficamente más alta, en donde fue colectada una fáunula de aspecto colhuehuapense. Los fósiles aparecen generalmente en concreciones silíceas, semejantes a las de Sacanana. Un ejemplar de Borhyaena sp., colectado por Pascual y colaboradores en 1982, procede de esta localidad.
El Pajarito. Una escasa fáunula de probable edad colhuehuapense fue exhumada en los alrededores de El Pajarito, tanto al norte como al sur de la Ruta N° 25 (Departamento de Paso de Indios, provincia del Chubut, Argentina; Scillato Yané, 1977; Marshall et al., 1983). En esta localidad fueron exhumados cuatro metaterios, referibles a los Paucituberculata Palaeothentidae y Abderitidae.
Paso Córdoba. Barrio et al. (1986) dieron a conocer un breve listado de vertebrados exhumados en niveles de la Formación Chichinales, a unos 10 kilómetros al sudoeste de Paso Córdoba, en la provincia de Río Negro. Los niveles fosilíferos corresponden a un estrato carbonático psefítico ubicado en la base de una secuencia formada por tobas medianas a finas y tufitas. La presencia de restos de Colpodon entre los mamíferos exhumados en Paso Córdoba permitió asignar esta fauna a la Edad-mamífero Colhuehuapense. El único metaterio recuperado consiste en un maxilar aislado del hatliacínido Cladosictis cf. C. patagonica (Barrio et al., 1986). Más recientemente Kramarz et al. (2004) comunicaron el hallazgo de nuevos restos de mamíferos exhumados en niveles de esta misma formación, "en el área de Paso Córdoba".
Localidades de la Formación Cerro Bandera. Cuatro localidades fosilíferas cercanas entre sí y ubicadas a pocos kilómetros al oeste, suroeste y sur de Cutral-Có y Plaza Huincul han sido analizadas recientemente (Kramarz et al., en prensa): Cerro Bandera (localidad tipo de la formación), Cerro Bayo Mesa, Sierra del Portezuelo y Barda Negra. Kramarz et al. (2001) dieron a conocer una fauna de mamíferos colhuehuapenses procedente de niveles de la Formación Cerro Bandera, aflorantes al noreste de la denominada Barda Negra y al sur del mismo Cerro Bandera (Departamento de Zapala, provincia del Neuquén, Argentina). La secuencia sedimentaria incluye niveles fosilíferos de bentonitas y tufitas, estas últimas con desarrollo de paleosuelos. Entre la asociación de mamíferos exhumados en estos niveles los autores mencionan un ejemplar de paleoténtido indeterminado. Los autores infieren "... una antigüedad no mayor que Mioceno Temprano (Colhuehuapense) para estos sedimentos" (Kramarz et al., 2001: 11). Finalmente, Kramarz et al. (en prensa) dan a conocer el conjunto de estas localidades, su contexto estratigráfico y contenidos fosilíferos. Para la localidad de Sierra del Portezuelo Norte señalan la presencia de Hathliacynidae y Borhyaenidae indet., sobre la base de restos sumamente fragmentarios.
Otras localidades extra-patagónicas. Son varias las probables localidades fosilíferas extra-patagónicas que han sido señaladas como portadoras de asociaciones de Edad-mamífero Colhuehuapense, aunque su estudio pormenorizado podría revelar una diferente asignación temporal. De una sola de estas localidades se ha registrado la presencia de metaterios. (a) Carlini et al. (1996: 461) mencionan el descubrimiento de fósiles de edad "Colhuehuapense-Mayoense (i.e., Mioceno Inferior-Mioceno Medio)" en la zona de Las Hornillas, sobre las márgenes del río de Los Patos, a unos 48 kilómetros al sud-sudeste de Barreal, provincia de San Juan. Los niveles fosilíferos corresponden a la Formación Chinches, de los cuales procede un paleoténtido indeterminado. (b) Sobre la base de información aportada por Stirton (1953), Marshall et al. (1983) refieren a la edad Colhuehuapense una serie de restos de mamíferos recolectados cerca del pueblo de Coyaima (Departamento de Tolima, Colombia), en niveles de areniscas ferruginosas pertenecientes a una unidad del Grupo Honda (pero véase Guerrero, 1997). (c) Sánchez-Villagra y Clack (2004) señalan la presencia de una fauna referida al Mioceno Temprano en depósitos de costa de la Formación Castillo, Venezuela; su eventual contemporaneidad con la edad mamífero Colhuehuapense no fue discutida. (d) Flynn et al. (1995) describen un primate de edad Mioceno Temprano de los Andes chilenos correspondiente a la Formación Abanico, con una edad 40Ar-39Ar de 20,09 ± 0,27 Ma, probablemente asociado a una fauna colhuehuapense.
Contexto geológico
Se detallan aquí aspectos estratigráficos, sedimentológicos y geocronológicos de los niveles aflorantes en las dos localidades fosilíferas que han aportado restos más abundantes de metaterios (y de mamíferos en general) de edad Colhuehuapense: los miembros Colhue Huapi (Puesto Almendra, en la Gran Barranca; figura 2) y Trelew (en los alrededores de Gaiman) de la Formación Sarmiento.
Miembro Colhue Huapi. El Miembro Colhue Huapi es la unidad superior de la Formación Sarmiento, aflorante en el extremo occidental de Gran Barranca (figura 1.B). Son varios los estudios geológicos que han permitido clarificar la pertenencia estratigráfica de las faunas colhuehuapenses. Con posterioridad a los trabajos de Ameghino (1900-1902, 1902, 1906), ya en la década de 1930 George G. Simpson registró numerosas observaciones de campo sobre la geología y paleontología de la Barranca Sur del Lago Colhue Huapi. Si bien el autor realizó varias notas y perfiles estratigráficos de las rocas conteniendo faunas de edad Deseadense y Colhuehuapense, sus observaciones permanecieron inéditas, constando sólo en sus libretas de campo -su trabajo édito se concentró en los términos inferiores de la Formación Sarmiento, fundamentalmente en lo que respecta a rocas y faunas de las edades Casamayorense y Mustersense (Simpson, 1948, 1967; Cifelli, 1984). La estratigrafía de los niveles superiores de la Formación Sarmiento fue descripta en detalle en sucesivos trabajos de Spalletti y Mazzoni (1977, 1979) y Mazzoni (1985). Más recientemente Kay et al. (1999a) figuraron los niveles superiores de esta sección de la Gran Barranca y dieron a conocer una serie de dataciones practicadas sobre las rocas que componen los distintos niveles de la misma (figura 2).
Nuestro uso del término es algo distinto al de la descripción original del miembro por Spalletti y Mazzoni (1979). En nuestra concepción, el Miembro Colhue Huapi abarca todo el paquete de sedimentos tobáceos blanquecinos y friables de la Formación Sarmiento que descansan encima de una discordancia erosiva significativa (Bellosi et al., 2002, Fig. 2). La discordancia muestra un relieve inciso de 27 m de profundidad, correspondiente a un paleovalle labrado sobre los paleosuelos anaranjados y endurecidos de la sección cuspidal del Miembro Puesto Almendra. Por encima de esta discordancia el Miembro Colhue Huapi mide aproximadamente 65 m de espesor total. Dentro del Miembro Colhue Huapi fueron reconocidas dos biozonas portadoras de mamíferos. Las secciones basal y media incluyen los niveles fosilíferos más ricos (Biozona Inferior) y miden 40 m de espesor. Ambas están constituidas por ciclos granodecrecientes, con conglomerados finos a medianos de base plana y poco erosiva, seguidos por tobas finas. Estas litofacies registran, por un lado, la acumulación fluvial en canales de poca envergadura y, por otro, la sedimentación loéssica (eólica) sobre las planicies (caídas distales de cenizas). Ambos tipos de depósitos y especialmente los más finos, fueron edafizados dando lugar a paleosuelos con rasgos ándicos (Bellosi et al., 2002). Por encima descansan 22 m de tobas finas macizas, pobremente estratificadas y estériles. Los últimos metros de la unidad corresponden a la Biozona Superior.
En la Gran Barranca, el techo del Miembro Colhue Huapi está cubierto discordantemente por los Rodados Patagónicos (Plio-Pleistoceno), mientras que 20 kilómetros al sudeste (Valle Hermoso) exhibe un contacto concordante con la Formación Chenque ("Superpatagoniano"), unidad marina del Mioceno Inferior (Bellosi y Barreda, 1993). En sectores localizados, los depósitos marinos presentan base erosiva e incluyen vertebrados colhuehuapenses (Ameghino, 1906; Feruglio, 1949).
La Biozona Inferior del Miembro Colhue Huapi es extraordinariamente fosilífera y ha proporcionado casi todos los materiales que ahora incluimos en la fauna Colhuehuapense. Todos los metaterios descriptos en este trabajo y que provienen de los siete sitios fosilíferos ya mencionados, fueron recolectados en esta biozona. Los sitios "Nivel C", "Nivel C Gris", "Nivel B", "Mono Este Superficial", "Mono Oeste Superficial", "Mono 2" y, en un sentido general, "Puesto Almendra", se ubican dentro de un intervalo estratigráfico de 4 m de espesor en la parte media de la Biozona Inferior. Estos niveles basales presentan, dentro de la zona de afloramientos, poca variación lateral.
Kay et al. (1999a, 1999b) dieron a conocer una serie de dataciones 40Ar/39Ar practicadas en muestras de vidrio volcánico y plagioclasa recogidas en la Gran Barranca en niveles asociados con faunas colhuehuapenses. En el contexto de un perfil de polaridad magnética asociado, los autores sugieren que el grueso de las faunas colhuehuapenses de la Gran Barranca cae dentro de los crones 6r y 6n de la escala de Berggren et al. (1995) (19,1 a 20,5 Ma). Flynn et al. (1995) mencionaron dataciones 40Ar/39Ar de 20,09 ±0,27 Ma para una fauna aparentemente colhuehuapense de Chile, lo que concuerda con las dataciones de Kay et al. (1999a, 1999b). Los sedimentos del intervalo estratigráfico en el que fueron colectados los fósiles aquí estudiados poseen una polaridad magnética normal y están próximos a una toba volcánica (la "Toba Mono") fechada en 19,8 Ma por el método 40Ar/39Ar (Kay et al., 1999b), por lo que los sedimentos portadores se corresponden con el Cron C6n de la escala de Berggren et al. (1995).
Miembro Trelew. El Miembro Trelew de la Formación Sarmiento ha proporcionado abundantes mamíferos fósiles en varias localidades correspondientes a la barranca Sur del curso inferior del río Chubut en el área de Gaiman y Trelew. El primer registro de fósiles en esta área corresponde a Roth (1899). Simpson (1935) revisó la geología y fauna asociada en esta área, y concluyó que sus fósiles debían ser referidos a la Edad-mamífero Colhuehuapense (véase también Bordas, 1939). Más recientemente, la estratigrafía de este miembro fue revisada por Fleagle y Bown (1983) y por Scasso y Bellosi (2004). Varios restos de primates y otros mamíferos fueron recolectados por Fleagle y colaboradores justo por encima del Miembro Rojo (Bown y Fleagle, 1993), el cual forma un distintivo resalto hacia el sector medio de la barranca. Se desconoce la ubicación estratigráfica precisa del primate Dolichocebus, colectado en los alrededores de Gaiman, si bien casi con seguridad debería proceder de niveles del Miembro Trelew (no se conocen restos de mamíferos terrestres procedentes de otros niveles de la barranca en esta área; Bordas, 1939; véase Bown y Fleagle, 1993: 7).
El Miembro Trelew yace en discordancia sobre el Miembro Rojo y es cubierto en aparente concordancia por la marina Formación Gaiman, del Mioceno Temprano (Scasso y Bellosi, 2004). Al igual que en la región del Colhue Huapi, este contacto se encuentra intensamente bioturbado por invertebrados marinos correspondientes a la transgresión Superpatagoniana. Los niveles más ricos en fósiles provienen de la sección inferior del Miembro Trelew, en particular del paleosuelo cálcico (calcrete nodular) que ocurre en la base de la unidad y de las tobas finas macizas que le suprayacen (loessitas), las cuales exhiben rasgos pedogénicos de leve desarrollo (entisoles-andisoles; Fleagle y Bown, 1983; Scasso y Bellosi, 2004), posiblemente por ser paleosuelos acumulativos (Bown y Fleagle, 1993).
Bown et al. (1988) mencionaron una edad por trazas de fisión de 15,8 ± 2,5 Ma en el Miembro Trelew de la Formación Sarmiento a la altura de Gaiman, unos 20 m por encima de los niveles con mamíferos colhuehuapenses. Una segunda e independiente fuente de información procede del hecho de que los niveles marinos de la Formación Gaiman, incluyendo restos de tiburones, ballenas, pingüinos y numerosos invertebrados, suprayacen a los niveles colhuehuapenses de Gaiman. Estos depósitos marinos, posiblemente correspondientes al "Superpatagoniano" (Bellosi, 1995), serían de edad burdigaliana, por lo que el Miembro Trelew y su fauna colhuehuapense no serían más jóvenes que el Mioceno Temprano.
Contexto paleogeográfico y paleoambiental
Bellosi (1995) sintetizó la información paleogeográfica y paleoambiental procedente de los registros sedimentarios, volcánicos y paleontológicos de la Patagonia central hacia la transición Paleógeno- Neógeno. A principios del Mioceno la cuenca San Jorge fue inundada por las transgresiones Leonense y Superpatagoniense, entre las cuales habría tenido lugar la mayor parte de la sedimentación colhuehuapense (figura 3). Los principales elementos morfoestructurales positivos que habrían de enmarcar la acomodación de sedimentos en esta cuenca fueron el Macizo del Deseado al sur, la Faja Plegada de San Bernardo (Bernárdides o Patagónides) inmediatamente al oeste del Lago Colhue Huapi, el Alto de Rawson al norte y, en el flanco occidental, la Cordillera Patagónica (Bellosi, 1995). Además de la cuenca San Jorge, el centro de Patagonia incluía las cuencas Mazzaredo al sudeste y la subcuenca Posadas al sudoeste (extremo norte de la cuenca Magallanes), aunque en ellas no se han reconocido fehacientemente registros colhuehuapenses. Estos elementos morfoestructurales definieron dos paleopenínsulas: la península Tehuelche (o Protopatagonia) al norte de la cuenca San Jorge, y la península Deseado al sur (figura 4.A-B). Debido a la directa vinculación de esta cuenca con el Atlántico, los cambios en la línea de costa estuvieron condicionados por oscilaciones eustáticas globales (Bellosi y Barreda, 1993; Legarreta y Uliana, 1994). Se ha inferido, para esta etapa transgresiva, un desplazamiento hacia el oeste y noroeste de la línea de costa en la cuenca San Jorge, siendo ocupada dicha área por un mar somero dominado por corrientes de marea (Bellosi, 1988). En este contexto paleogeográfico, los depósitos mamalíferos más importantes de edad colhuehuapense, los de la Gran Barranca, aparecen en el extremo austral de la península Tehuelche (figura 4.B).
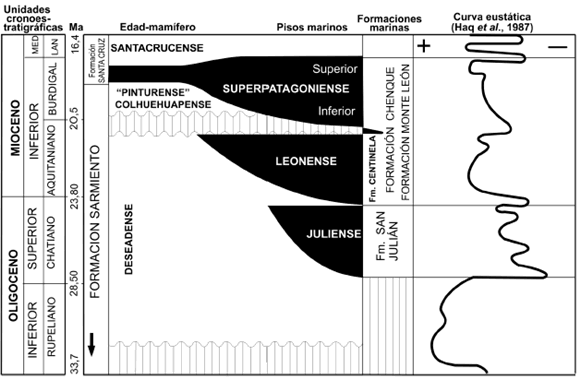
Figura 3. Estratigrafía de secuencias continentales y marinas del Oligoceno-Mioceno en diferentes cuencas de la Patagonia Central (modificado de Bellosi, 1995). La curva de Haq et al. (1987) está adaptada a la cronoestratigrafía de Berggren et al. (1995). De izquierda a derecha se indican las unidades cronoestratigráficas (series y pisos), los millones de años antes del presente (Ma), las formaciones continentales (F), la secuencia de Edades-mamífero, los pisos marinos, las formaciones marinas y la curva eustática global / stratigraphy of terrestrial and marine sequences from the Oligocene-Miocene in different Central Patagonian basins (modified from Bellosi, 1995). Haq et al.'s (1987) curve was adapted to comform Bergreen et al.'s (1995) chronostratigraphy. From left to right are indicated the chronostratigraphic units (series and stages), million years before present (Ma), terrestrial formations (F), the land-mammal ages succession, marine stages, marine formations and the global eustatic curve.

Figura 4. Paleogeografía generalizada de la Patagonia central (cuenca San Jorge) en el Burdigaliano Temprano (A, aproximadamente 20 Ma) y tardío (B, hace 17 Ma). En A se reconstruyen los valles fluviales incisos rellenos con sedimentos del Miembro Colhue Huapi y el posterior desarrollo de estuarinos mareales transgresivos en areas distales. En B se muestra la expansión del mar "Superpatagoniano" que alcanzó su máximo algo después de los tiempos colhuehuapenses (Fm. Chenque, Secuencia 4). Modificado de Bellosi (1995) / generalized paleogeography of east-central Patagonia (San Jorge Basin) during the early Burdigalian (A, approximately 20 Ma) and late Burdigalian (B, 17 Ma). In A, it can be seen the incised fluvial valleys filled with Colhuehuapian sediments and the subsequent development of tidal-controlled estuarine systems in distal areas. In B, it is shown the maximum expansion of the "Superpatagonian" sea, little after of Colhuehuapian times (Chenque Formation, Sequence 4). Modified from Bellosi (1995).
Uno de los rasgos más notables de la composición de las sedimentitas mesoterciarias de la Patagonia es el importante aporte piroclástico cuyo origen habría sido la suspensión eólica directa o caída de cenizas. La Formación Sarmiento consiste en una secuencia de sedimentitas piroclásticas de primero y segundo ciclo (Spalletti y Mazzoni, 1977, 1979; Mazzoni, 1979). Éstas constituirían la manifestación distal de un vulcanismo de tipo pliniano que habría tenido lugar en áreas situadas en el noroeste patagónico (Mazzoni, 1985) y que incluirían el sector extraandino occidental del Macizo Nordpatagónico (Franchi y Nullo, 1986). En sus consideraciones acerca de la génesis de estos depósitos para el conjunto de la formación, Spalletti y Mazzoni (1979) estimaron que el agente principal de transporte y depositación fueron las lluvias distales de cenizas volcánicas, ocurriendo en regiones semiáridas a húmedas de bajo gradiente. Si bien el suministro de material es eólico, la sedimentación colhuehuapense fue en parte subaérea -como lo demuestran los numerosos caracteres loéssicos y la abundancia de paleosuelos-, y en parte también fluvial, especialmente en la mitad inferior del Miembro Colhue Huapi, donde prevalecen cuerpos canalizados de conglomerados intraformacionales. Tal como tempranamente advirtiera Ameghino (1906), buena parte de la Formación Sarmiento está formada por acumulaciones comparables al loess plio-pleistoceno de la región pampeana.
La composición de la fauna atlántica durante los tiempos colhuehuapenses y levemente posteriores a la transgresión superpatagoniana muestra un cambio importante en las temperaturas marinas, cambios que son evidenciados por el registro de invertebrados tropicales y peces de aguas cálidas (Feruglio, 1949; Cione, 1979). La fauna de vertebrados terrestres ha sugerido a varios autores (véase Pascual et al., 1996 y bibliografía citada) cierto grado de fragmentación ambiental, con alternancia de sabanas arboladas, bosques húmedos, escasas y circunscriptas estepas herbáceas y, en áreas determinadas, ambientes abiertos de tipo semiárido a árido (véase más abajo). Un detalle más pormenorizado sobre la información aportada por distintos grupos de mamíferos se ofrece en el siguente párrafo.
Sin el beneficio de la amplia muestra de metaterios que aquí se describe, Pascual y Bond (1986) sugirieron que los mismos muestran una marcada diversificación en relación con aquéllos de edad Deseadense. Los autores destacaron la abundancia de microbiotéridos, el primer registro de boriénidos protilacínidos, la notable radiación de paucituberculados y el primer registro de patagónidos y argirolágidos. Los microbiotéridos vivientes (con una sola especie, Dromiciops gliroides Thomas, 1894) se restringen a los ambientes boscosos y húmedos de la Patagonia andina de Chile y Argentina. Son de hábitos frugívoro-insectívoros y notablemente arborícolas. Los protilacínidos se caracterizan por una estructura molar en la que los talónidos muestran un grado de reducción relativamente escaso, lo que indica cierto componente omnívoro en la dieta, tal vez vinculado con la oferta estacional de alimentos. El primer molar inferior modificado (de tipo "plagiaulacoideo", muy comprimido lateralmente y de aspecto cortante) de los paucituberculados Palaeothentidae y Abderitidae ha sido objeto de variadas especulaciones paleobiológicas. Burramys parvus Broom, 1896, un diprotodonte australiano viviente con morfología dentaria similar y tamaño equivalente, ha sido observado utilizando este diente especializado para romper el caparazón duro de ciertos escarabajos (en contra de interpretaciones sobre su supuesta utilización en dietas estrictamente frugívoras; Turner y McKay, 1989; véase también Wall y Krause, 1992). Para Pascual y Bond (1986) los argirolágidos y patagónidos sugieren, al contrario que los microbiotéridos, ambientes abiertos y más áridos (véase más abajo). Finalmente, el registro de borieninos relativamente grandes (Arctodictis sinclairi Marshall) prefigura a los grandes metaterios carnívoros del Santacrucense. Un análisis reciente de Forasiepi (1999) sobre la morfología carpal y tarsal de A. sinclairi argumenta a favor de su carácter plantígrado (véase también Argot, 2004). Los edentados colhuehuapenses también difieren significativamente de aquéllos de la edad precedente (Scillato-Yané, 1986). Desaparecen las formas gigantescas, aparecen los primeros megaloníquidos, y entre los dasipódidos abundan las formas entomófagas de la Tribu Stegotherinii. En relación con los edentados de la edad subsiguiente, la diversidad de la asociación colhuehuapense es baja. Vucetich (1986) analizó la composición cenozoica de roedores y primates para la Patagonia central. La autora destacó, en comparación con las formas santacrucenses, los siguientes aspectos distintivos de la edad Colhuehuapense: (1) la diferente composición taxonómica entre los roedores equímidos, incluyendo un solo género cuya especie muestra una tendencia definida hacia la hipsodoncia; (2) una mayor abundancia relativa de cefalómidos; (3) la ausencia de chinchíllidos; (4) la menor diversidad de dasipróctidos y eocárdidos; (5) una gran diversidad relativa de eretizóntidos; (6) una mayor diversidad de primates con respecto a la edad Santacrucense. La información provista por el conjunto de los roedores y primates sugiere la existencia de climas relativamente cálidos y húmedos, con amplio desarrollo de superficies boscosas. Los bajos índices de hipsodoncia entre los equímidos, por ejemplo, contrastan con su abundancia en tiempos santacrucenses. Los eretizóntidos vivientes son todos arborícolas; Candela et al. (2005) sugirieron que algunos tipos extinguidos habrían tenido hábitos locomotores, al menos, semiarborícolas. De todos modos, la abundancia de eretizóntidos colhuehuapenses constituye un dato significativo, como también lo es la relativa abundancia de primates platirrinos. Bond (1986) analizó las asociaciones cenozoicas de los ungulados nativos del territorio argentino. En relación con los ungulados deseadenses, el autor señaló la menor diversidad de las formas colhuehuapenses. Abundan los últimos registros taxonómicos de las formas arcaicas de macrauquénidos y cramauquénidos, así como también de leontínidos (Colpodon). Son abundantes los proterotéridos, notoungulados homalodotéridos y notohípidos, y especialmente interatéridos y hegetotéridos. En conjunto, el autor sugirió, concordantemente con otras fuentes de evidencia, la mayor persistencia de tipos ramoneadores entre las especies colhuehuapenses que entre las santacrucenses. Esto habla en favor de ambientes menos abiertos, más húmedos y boscosos que los que caracterizarían la edad subsiguiente en el extremo sur de la Patagonia.
El análisis palinológico de la Formación Chenque permitió a Barreda (1996) y Barreda y Palamarczuk (2000a, 2000b, 2000c) arribar a conclusiones detalladas sobre la flora del Oligoceno Tardío-Mioceno Temprano de Patagonia, con la definición de cuatro asociaciones de polen de angiospermas y dinoflagelados, cuyo ajuste temporal posee limitaciones para intervalos de tiempo tan reducidos como el colhuehuapense.
El registro palinológico de la secuencia marina que mejor correlaciona con esta edad es, por otra parte, relativamente escaso (Barreda y Bellosi, 2003), aunque las Asociaciones G/C y T-B/H serían, desde el punto de vista cronológico, las más próximas. El fuerte contraste composicional entre estas asociaciones indicaría la existencia de un hiato estratigráfico en la Formación Chenque, o bien un rápido cambio en la vegetación (¿y en el clima?) de la Patagonia central. Con todo, es posible inferir que los rasgos paleoflorísticos comunes a ambas asociaciones podrían haber permanecido en tiempos colhuehuapenses. En una reciente reconstrucción de los ambientes continentales del Mioceno Temprano de la Patagonia central, Barreda y Bellosi (2003) destacaron las significativas variaciones climáticas y paleogeográficas ocurridas en dicho lapso a partir de variadas evidencias. Estos autores interpretaron para el Aquitaniano Tardío (Asociación G/C) una flora dominada por arbustos xerofíticos (efedráceas y quenopodiáceas) que ocuparon la planicie costera. En posiciones alejadas de la costa o, alternativamente, altos topográficos se habrían desarrollado bosques húmedos de podocarpáceas, fagáceas y araucariáceas, junto con representantes de sotobosque dominado por helechos, algunos de tipo arborescente. Las podocarpáceas podrían también haber crecido en las partes bajas del relieve o en bosques en galería en los márgenes de los ríos. Este conjunto evidencia condiciones climáticas subhúmedas. Poco después de este lapso (Asociación TB/ H), la vegetación se modificó sustancialmente en consonancia con el óptimo climático de fines del Mioceno Temprano. El cambio florístico resultó más significativo en la planicie costera, donde prevalecieron esparganiáceas acuáticas, colonias de Pediastrum, másulas de salviniáceas, restionáceas, cyperáceas y fagáceas (Barreda, 1996; Barreda y Bellosi, 2003), acompañado por la retracción de representantes arbustivos xerofíticos. Tal asociación indica un clima húmedo y cálido.
La Formación Sarmiento en la Gran Barranca es, asimismo, extraorinarimente prolífica en fitolitos (Mazzoni, 1979), por lo que resulta el registro más antiguo de ecosistemas de pastizales bien desarrollados (Zucol et al., en prensa). El Miembro Colhue Huapi muestra claras fluctuaciones entre ejemplares de pastos meso- y macrotérmicos. Zucol et al. (en prensa) reconocieron a lo largo del Miembro Colhue Huapi una heterogénea y variable asociación de morfotipos entre los que preponderan los componentes panicoides, pooides y chloridoides, con abundantes prismatolitos, 3-flabelolitos, aculeolitos, espículas y diatomeas en los niveles inferiores; menor proporción de globulolitos de arecáceas, 5-nuxolitos, 6-proteolitos y 7-halteriolitos; y escasos 8-estrobilolitos, 9-braquiolitos, 10-doliolitos y 11-pileolitos.
A manera de síntesis, se interpreta que el paisaje colhuehuapense de la Patagonia central se desarrolló en una planicie costera bordeada por un mar somero, cuya temperatura es difícil de definir ya que este lapso corresponde a la transición entre mares posiblemente fríos por efecto del avance de corrientes antárticas de la transgresión Leonense (Nánez, 1988; Bellosi, 1994) y cálidos de la transgresión Superpatagoniana (Bellosi, 1995). De acuerdo al registro de fitolitos y a las características persistentes en las palinofloras temporalmente próximas, en las zonas más elevadas o alejadas del mar prevalecieron bosques de podocarpáceas, araucariáceas y fagáceas mientras que en áreas bajas y litorales dominaron los representantes herbáceoarbustivos (frecuentes gramíneas) y vegetación arbórea subordinada o en parches. El clima debió haber sido templado-cálido a cálido y posiblemente subhúmedo, con mayores precipitaciones en zonas elevadas. Los conjuntos paleoflorísticos evidencian fluctuaciones ambientales propias de un lapso de transición hacia condiciones más húmedas y cálidas, las cuales preevalecieron a fines del Mioceno Temprano (Edad Pinturense; Kramarz y Bellosi, 2005). Las evidencias mencionadas sugieren, al igual que en otras secuencias cenozoicas de la Patagonia (Bellosi et al., 2002; Kramarz y Bellosi, 2005), que las fluctuaciones climático- ambientales habrían sido de una frecuencia tal que, por el momento, la caracterización de las mismas constituye por ahora un problema irresoluble, y/o que los registros paleobiológicos son resultados promediados o condensados de las mismas.
Paleontología sistemática
Infraclase METATHERIA Huxley,1880
Supercohorte MARSUPIALA (Illiger, 1811)
Orden DIDELPHIMORPHIA (Gill, 1872) Aplin y Archer, 1987
Superfamilia DIDELPHOIDEA (Gray, 1821) Osborn, 1910
Familia DIDELPHIDAE Gray, 1821Gen. et sp. indet.
Figuras 5.A-F

Figura 5. A-F, Didelphidae gen. et sp. indet. A-C, MPEF-PV 4774, un molar inferior izquierdo (m2 ó m3) en vistas lingual (A), posterior (B) y oclusal (C). D, MPEF-PV 4707, un molar superior derecho (Mx) en vista oclusal. E-F, MPEF-PV 4773, un fragmento mandibular derecho con un molar (m?1) en vistas anterolabial (E) y anterolingual (F). G-H ?Caluromyidae, gen. et sp. indet., MPEF-PV 4708, un molar inferior izquierdo en vistas oclusal (G) y lingual (H). I, Didelphimorphia indet., MLP 77-VI-13-25, un fragmento mandibular izquierdo con parte de la dentición yugal preservada; vista lingual (detalle). J-M, Didelphimorphia o Sparassodonta, gen. et sp. indet. J-K, MLP 77-VI-13-8, un trigónido derecho aislado en vistas lingual (J) y oclusal (K). L-M, MLP 92-X-10-11, un M4 izquierdo aislado, completo; vistas anterior (L) y oclusal (M). Referencias: En, entocónido; Hy, hipocónido; Hyl, hipoconúlido; Me, metacónido; Pa, paracónido; Pr, protocónido / A-F, Didelphidae gen et sp. indet. A-C, MPEF-PV 4774, a lower left molar (m2 or m3) in lingual (A), posterior (B), and occlusal (C) views. D, MPEF-PV 4707, an upper right molar (Mx) in occlusal view. E-F, MPEF-PV 4773, a fragment of a right dentary with one molar (m?1) in antero-labial (E) and antero-lingual (F) views. G-H, ?Caluromyidae, gen. et sp. indet., MPEF-PV 4708, a lower left molar in occlusal (G), and lingual (H) views. I, Didelphimorphia indet., MLP 77-VI-13-25, a left dentary fragment with part of the cheek-teeth in lingual view (detail). J-M, Didelphimorphia or Sparassodonta gen. et sp. indet. J-K, MLP 77-VI-13-8, an isolated right trigonid in lingual (J) and occlusal (K) views. L-M, MLP 92-X-10-11, a complete, isolated left M4 in anterior (L) and occlusal (M) views. References: En, entoconid; Hy, hypoconid; Hyl, hypoconulid; Me, metaconid; Pa, paraconid; Pr, protoconid.
Material. MPEF-PV 4774, un molar inferior izquierdo, aislado (m2 o m3; figuras 5.A-C), MPEF-PV 4709, un molar inferior izquierdo sin el metacónido (m?1), MPEF-PV 4707, un molar superior derecho, muy gastado (Mx; figura 5.D). Material tentativamente referido. MPEF-PV 4773, un fragmento mandibular derecho con un molariforme (m?1; figuras 5.E-F).
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, Provincia del Chubut, Argentina). Fm. Sarmiento, Miembro Colhue Huapi, Nivel C. Mioceno Inferior (Edad-mamífero Colhuehuapense). Los ejemplares fueron coleccionados por personal del MLP y DU en 1999 (MPEF-PV 4774) y 2000 (MPEFPV 4773).
Medidas. MPEF-PV 4774: largo total, 1,52 mm; ancho del trigónido, 0,96 mm; ancho del talónido, 0,84 mm. MPEF-PV 4709: largo total,1,40 mm; ancho del trigónido, 0,84 mm; ancho del talónido, 0,80 mm. MPEFPV 4707: largo total, 1,28 mm; ancho total, 1,76 mm. MPEF-PV 4773: largo total, 1,12 mm; ancho del trigónido, 0,56 mm; ancho del talónido, 0,48 mm.
Descripción. El m2 o m3 MPEF-PV 4774 (figuras 5.AC) está completo, incluyendo ambas raíces. Su tamaño es similar al de los marmosinos actuales relativamente pequeños. Las cúspides del trigónido son notablemente más altas y robustas que aquellas del talónido; la paracrístida y la metacrístida son altas y están bien desarrolladas; el protocónido es muy alto y robusto, con su base ocupando buena parte de la superficie del trigónido; el paracónido es más pequeño y bajo que el metacónido y está algo comprimido an- teroposteriormente; el cíngulo anterobasal es corto pero amplio. El talónido es apenas más corto que el trigónido; el hipocónido es relativamente saliente y mucho más alto que el hipoconúlido y el entocónido; el extremo distal de la crístida oblicua -la cual es relativamente muy alta- contacta con el protocónido en un punto labial y muy por debajo de la muesca de la metacrístida; el hipoconúlido es más alto que el entocónido y se ubica casi por detrás de esta cúspide; la postcrístida es casi transversal al eje dentario. Si bien está roto en el ápice, puede apreciarse que el entocónido está claramente reducido; por su parte, la preentocrístida es muy baja. La morfología del diente sugiere hábitos insectívoros o animalívoros.
El m?1 MPEF-PV 4709 es esencialmente similar al anterior, aunque apenas más pequeño y con el entocónido más alto. El cíngulo anterobasal está un poco menos desarrollado que el del MPEF-PV 4774 y la paracrístida es más corta y baja que la de aquel ejemplar. Por todas estas diferencias, el ejemplar MPEFPV 4709 podría corresponder a un m1; de hecho, su hipoconúlido calza ajustadamente en la muesca hipoconúlida del ejemplar MPEF-PV 4774.
El ejemplar MPEF-PV 4707, un molar superior derecho (figura 5.D), está muy desgastado y roto a la altura de la postmetacrístida. Su morfología general es coherente con la de un antagonista oclusal para los dientes inferiores ya descriptos. El molar es más ancho que largo, el protocono está reducido y la postmetacrista es comparativamente grande y larga; a pesar del desgaste, el tamaño relativo de sus respectivas bases sugiere que el metacono duplicaba o triplicaba al paracono. No se aprecia evidencia alguna de para- o metacónulo. El cíngulo anterobasal es corto y las cúspides estilares están poco desarrolladas, siendo la StD más grande que la StB.
El resto mandibular MPEF-PV 4773 (figuras 5.E-F) es sumamente fragmentario. Como ocurre en otros didelfimorfios, la cara lingual es plana y la labial levemente convexa. Por debajo del alvéolo correspondiente a la raíz posterior del molariforme preservado, la pared labial del dentario está rota, por lo que no se advierten eventuales forámenes mentonianos. Sólo se han preservado dos alvéolos dentarios: el correspondiente a la raíz posterior del único molar preservado, y el alvéolo inmediatamente subsiguiente. El primero es algo más pequeño que el segundo. El diente está bien preservado aunque fuertemente desgastado; el paracónido es bajo y tiene una ubicación claramente mesial; la paracrístida es relativamente larga. El metacónido y el protocónido, fuertemente desgastados, están muy próximos entre sí y parcialmente fusionados en la base. Sobre la pared labial del trigónido no se advierte evidencia alguna de cíngulo anterobasal. El trigónido es algo más largo que el talónido y apenas más ancho. Los rasgos más notables del talónido son la gran altura relativa del hipocónido y de la crístida oblicua, así como también la reducción del entocónido. De hecho, no se advierte evidencia de esta cúspide, siendo imposible de determinar si esto último es debido al fuerte desgaste de todo el diente o al carácter vestigial o ausente de esta cúspide. Una leve saliencia posterior, por detrás de la esquina posterolingual del talónido, sugiere la existencia de un hipoconúlido bajo y poco desarrollado. Sólo se ha preservado la raíz posterior de este diente. La misma no es recta sino que su extremo distal se curva hacia atrás. En vista posterior, la raíz muestra su cara labial moderadamente convexa.
Comentarios. A diferencia de los m1 de la mayor parte de los Didelphimorphia, el paracónido del MPEF-PV 4774 está ubicado en una posición bien lingual y no mesial; a diferencia de los m4 de estos marsupiales, su talónido no está reducido o comprimido lateralmente. Por estos motivos, el ejemplar en cuestión constituye más probablemente un m2 o m3. El ejemplar MPEF-PV 4709 es algo menor que el anterior, con la paracrístida menos desarrollada y el cíngulo anterobasal más angosto, lo que sugiere que este diente es un m1. Por su parte, el ejemplar MPEFPV 4707 podría corresponder a un M2 o M3: el metacono y la postmetacrista están bien desarrollados (i.e., no se trata de un M4) y el molar es muy ancho en relación al largo, con un metacono claramente más grande que el paracono; estos rasgos vuelven improbable su pertenencia a un M1.
El ejemplar MPEF-PV 4773 es claramente más pequeño que el MPEF-PV 4774, tanto en el largo como en el ancho de su trigónido y talónido. Sin embargo, algunos rasgos morfológicos sugieren la pertenencia de ambos a un mismo taxón: crístidas oblicuas altas y entocónidos reducidos. Otros rasgos no son tan concordantes: el cíngulo anterobasal del MPEF-PV 4774 está bien desarrollado, mientras que el del MPEF-PV 4773 es irreconocible (i.e., es vestigial o ausente); en segundo lugar, la posición del paracónido en el MPEF-PV 4773 es claramente más mesial. Estas diferencias, especialmente la segunda, así como también el tamaño, podrían ser indicativas de la ubicación de estos dientes en distintos loci de la serie molar: el MPEF-PV 4773 podría constituir un dp3, mientras que, como ya se señaló, el MPEF-PV 4774 podría representar un m2 o m3. Luego de examinar el cuerpo mandibular y los alvéolos preservados del MPEF-PV 4773 es claro que el alvéolo posterior del diente es completo, no observándose en el mismo oquedad o espacio alveolar alguno que sugiera la existencia de un molar permanente en desarrollo por debajo de aquél. Sin embargo, este espécimen podría corresponder a un individuo juvenil, en el que dicha cripta o espacio alveolar no se ha desarrollado aún; en este último caso, el ejemplar MPEFPV 4773 podría interpretarse como un dp3. Cuatro rasgos derivados pueden apreciarse en los molares aquí descriptos: en los inferiores, la amplitud del cíngulo anterobasal, la altura y desarrollo de la paracrístida y metacrístida y la reducción del entocónido; en el superior, la gran anchura del diente en relación a su largo. Los dos primeros rasgos pueden ser observados en didelfimorfios tanto paleógenos como neógenos; sin embargo, la reducción del entocónido es un rasgo casi exclusivo de las formas neógenas (i.e., los Didelphoidea sensu Goin, 1991,1995). La mayor parte de los didelfimorfios paleógenos, como los Derorhynchidae, Protodidelphidae y Sternbergiidae, así como también los Herpetotheriidae del Paleógeno y Neógeno europeo y norteamericano, se caracterizan precisamente por poseer entocónidos bien desarrollados a muy grandes; la única excepción es Minusculodelphis minimus Paula Couto, 1962 (Paleoceno de Itaboraí) cuyo entocónido es vestigial. En este último caso, sin embargo, la reducción del entocónido es concomitante con una reducción igualmente notable de las otras cúspides del talónido, así como también de la totalidad del talónido, reducción que podría relacionarse con muy peculiares hábitos alimentarios (¿nectarívoros?, tal como lo sugieren su tamaño, la elongación mandibular y la simplificación de la serie dentaria) por parte de estos marsupiales excepcionalmente pequeños. En conclusión, entendemos que el taxón innominado al que pertenecen los ejemplares aquí descriptos constituye más probablemente un didelfoideo; su tamaño y proporciones generales coinciden con los de los didélfidos marmosinos de pequeño tamaño de la fauna sudamericana viviente (véase, e.g., Voss et al., 2004). La reducción del talónido ya presente en este ejemplar sugiere su mayor afinidad con Monodelphis Burnett, 1830 y formas afines (Monodelphini sensu Goin y Rey, 1997). De confirmarse la pertenencia de este taxón aquí innominado a los Didelphidae, constituiría el registro más antiguo para esta familia.
Familia ?CALUROMYIDAE Kirsch, 1977
?Caluromyidae Gen. et sp. indet.
Material. MPEF-PV 4708, un molar inferior izquierdo aislado.
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Fm. Sarmiento, Miembro Colhue Huapi, Nivel C. Mioceno Inferior (Edad- mamífero Colhuehuapense); coleccionados por personal del MLP y DU.
Medidas. Largo total, 1,68 mm; ancho del trigónido, 1,16 mm; ancho del talónido, 1,24 mm.
Descripción. El ejemplar MPEF-PV 4708 está roto a la altura de las cúspides del protocónido y metacónido (figuras 5.G-H), a lo largo de la metacrístida y en la mitad labial del hipoconúlido. El trigónido es más corto y subigual o apenas más angosto que el talónido. Si bien están rotos en la parte apical de sus cúspides, se aprecia que el protocónido y el metacónido eran de baja altura; por el contrario, el paracónido está bien desarrollado, se ubica en una posición menos lingual que el metacónido y es relativamente alto. Las cúspides del paracónido y metacónido están más próximas entre sí que cualquiera de ellas con el protocónido, por lo que el aspecto del trigónido en vista oclusal es el de una estructura comprimida en sentido anteroposterior. El talónido muestra una amplia cuenca central más ancha que larga, bien limitada lingualmente por el muy desarrollado entocónido, la cúspide más alta del diente; el hipocónido es muy saliente labialmente; el hipoconúlido, si bien es de posición relativamente lingual, no está apareado al entocónido. La estructura del hipoconúlido es curiosa: está bien comprimida en sentido dorsoventral. La postcrístida, que forma el borde posterior del talónido, no es transversal al eje dentario sino oblicua, por lo que el hipocónido ocupa una posición más anterior que en la mayor parte de los didelfimorfios. El cíngulo anterobasal está extremadamente reducido y limitado a un pequeño reborde por debajo de la muesca de la paracrístida; no existe postcíngulo. En vista lateral se aprecia que la corona del diente es muy baja; las raíces anterior y posterior están comprimidas. Se observa una incipiente faceta de desgaste a lo largo de todo el borde labial de la paracrístida, faceta que se continúa en la cara anterior del cíngulo anterobasal.
Comentarios. El ejemplar MPEF-PV 4708 constituye un marsupial enigmático. Podría postularse, por ejemplo, la pertenencia de este ejemplar a los Microbiotheria: la posición del hipoconúlido y el carácter saliente del hipocónido así lo sugieren. Sin embargo, la crístida oblicua y la postcrístida forman entre sí un ángulo muy agudo y el entocónido y el paracónido están bien desarrollados. Ninguno de estos rasgos es observable en los Microbiotheria conocidos. Las afinidades con los Didelphimorphia paleógenos o neógenos resultan más probables, si bien tampoco ofrecen resultados concluyentes. La proximidad entre el paracónido y el metacónido, por ejemplo, es infrecuente en la mayor parte, si no en todos, los didelfimorfios neógenos. Un rasgo de estas características se aprecia en algunas formas paleocenas, como Gaylordia macrocynodonta Paula Couto, 1952 del Paleoceno de Itaboraí; sin embargo, el paracónido en Gaylordia está reducido, el cíngulo anterobasal está bien desarrollado y el talónido es sumamente corto y sobre su borde posterior persiste un postcíngulo bien desarrollado. La morfología general del trigónido, pero no así la del talónido, recuerda también a la de Bobbschaefferia fluminensis Paula Couto, 1952, también del Paleoceno de Itaboraí, especialmente en lo que respecta al tamaño y posición del paracónido y a la baja altura de las cúspides en general. Sin embargo, los talónidos de ambos difieren ampliamente. Finalmente, una comparación con los paleógenos Derorhynchidae sugiere ciertas semejanzas en el gran tamaño y estructura columnar del entocónido, aunque no así en el resto de las cúspides y crestas del molar inferior.
Entre los Didelphimorphia neógenos, la estructura del trigónido sugiere, por el tamaño y posición del paracónido, varias semejanzas con la especie viviente Caluromysiops irrupta Sanborn, 1951. Otro rasgo coincidente con los molares inferiores de esta última especie es la posición del hipocónido, bien adelantado con respecto al borde posterior del talónido (véase el m2 de C. irrupta en Reig et al., 1987, Fig. 10.F). Este último rasgo es interesante, ya que es altamente infrecuente, si no inexistente, en el resto de los Didelphimorphia. Finalmente, el escaso apareamiento del hipoconúlido con el entocónido sólo ocurre, entre los Didelphimorphia vivientes, en Caluromysiops (Voss y Jansa, 2003). La silueta labial que describe el diente en vista oclusal sugiere que los molares superiores antagonistas del mismo debieron haber tenido una profunda centrocrista en "v"; la amplitud del talónido, un protocono ancho y, finalmente, el ángulo que forman la pre- y postprotocrístida sugieren un espacio interdental reducido. Asimismo, a juzgar por el desarrollo de la paracrístida, la postmetacrista de los molares superiores debería haber sido de tamaño moderado. Todos estos rasgos inferidos coinciden con la morfología de los molares superiores de los Caluromyidae, especialmente con Caluromysiops irrupta. No hemos podido comparar el ejemplar MPEF-PV 4708 con materiales referibles a las especies de Caluromyidae, por lo que no se proseguirá aquí con esta hipótesis sistemática.
DIDELPHIMORPHIA indet.
Figura 5.I
Material. MLP 77-VI-13-25, un fragmento mandibular izquierdo con el canino, alvéolos del p1, p2 completo, mitad posterior del p3, alvéolos del m1, raíces anteriores y fragmentos de los talónidos de los m2-3 y raíz anterior del m4, y un fragmento mandibular derecho con la dentición ausente; colectado por Hebe Herrera en 1977.
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, Provincia del Chubut, Argentina). Niveles inciertos, probablemente (aunque no con seguridad) referibles al Miembro Colhue Huapi de la Fm. Sarmiento (Edad mamífero Colhuehuapense) (H. Herrera, com. pers. 2005).
Medidas. Largo del canino, 1,24 mm; ancho del canino, 0,8 mm; largo del p2, 1,32 mm, ancho del p2, 0,56 mm; largo aproximado del p3, 1,48 mm; largo aproximado del m1, 1,68 mm; largo aproximado del m2, 1,84 mm; largo aproximado del m3, 1,84 mm; largo aproximado de la serie c-m4, 2,82 mm. Fragmento mandibular derecho: largo aproximado de la serie p2-m4, 20,9 mm.
Descripción y comentarios. El fragmento mandibular izquierdo está mejor preservado que el derecho e incluye unas pocas piezas dentarias pobremente preservadas. La longitud de la serie dentaria es mayor que la del microbiotérido Pachybiotherium acclinum Ameghino, 1902 y, a su vez, el cuerpo mandibular es más bajo y delgado. El canino es menos procumbente que el de los didélfidos; asimismo, y a diferencia de los Didelphoidea neógenos, los p2-3 son subiguales en tamaño a juzgar por las dimensiones inferidas para el p3. Todos los premolares son estrechos y alargados, y al menos los p2-3 terminaban posteriormente en una pequeña cuenca. De los molares sólo se han preservado parcialmete los talónidos de los m2-3; estos tienen el hipocónido poco saliente y la crístida oblicua relativamente paralela al eje dentario.
A pesar de la similitud general en tamaño entre el ejemplar MLP 77-VI-13-25 y los del microbiotérido Pachybiotherium acclinum, las distintas proporciones en las dimensiones de su cuerpo mandibular y serie dentaria no concuerdan con las de aquél ni con las de ningún otro microbiotérido conocido. El borde alveolar mandibular es recto, los premolares son relativamente comprimidos y el canino, si bien poco procumbente, tampoco es subvertical como el de los microbiotéridos. En suma, el conjunto de rasgos mandibulares y dentarios presentes en el ejemplar MLP 77- VI-13-25 permite su asignación a los Didelphimorphia, si bien su carácter fragmentario impide mayores precisiones taxonómicas. El tamaño de los restos preservados es mucho mayor que el de los ejemplares de Didelphidae analizados más arriba.
DIDELPHIMORPHIA o SPARASSODONTA gen. et sp. indet.
Figuras 5.J-M
Material. MLP 77-VI-13-8, un trigónido derecho aislado (figuras 5.J-K); MLP 92-X-10-11, un M4 izquierdo aislado, completo (figuras 5.L-M).
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). El ejemplar MLP 77-VI-13-8 fue coleccionado por Hebe Herrera en 1977. El ejemplar MLP 92-X-10-11 fue coleccionado en Puesto Almendra por personal del MLP y UNSW en noviembre de 1992.
Medidas. MLP 77-VI-13-8: largo, 1,8 mm; ancho aproximado, 1,6 mm; MLP 92-X-10-11: largo, 1,28 mm; ancho, 2,96 mm.
Descripción y comentarios. El ejemplar MLP 77-VI- 13-8 (figuras 5.J-K) muestra un protocónido robusto al igual que el paracónido, el cual, si bien es más pequeño, está bien desarrollado. La muesca hipoconúlida está bien desarrollada y probablemente también el cíngulo anterobasal; a juzgar por sus partes preservadas, el metacónido es muy pequeño y se fusiona basalmente al protocónido; ambas cúspides están algo comprimidas en sentido anteroposterior.
El ejemplar MLP 92-X-10-11 (figuras 5.L-M) muestra una única cúspide central, el paracono, y una amplia cuenca en su mitad lingual correspondiente al cuerpo del protocono. El paracono se conecta con la esquina parastilar a través de una cresta baja; no hay evidencia de cúspides estilares (salvo un reborde de esmalte hacia la esquina posterolabial del diente) ni de metacono.
Ambos ejemplares muestran una combinación de caracteres coincidentes con las adaptaciones esperables en metaterios carnívoros de tamaño relativamente pequeño. Precisamente por estas especializaciones, es sumamente difícil asignarlos a alguno de los grupos reconocibles de Didelphimorphia o Sparassodonta. El trigónido del ejemplar MLP 77-VI-13-8 muestra una paracrístida bien desarrollada y el metacónido reducido, rasgos que constituyen típicas adaptaciones hacia la carnivoría. La reducción y posterior pérdida del metacónido en los Sparassodonta constituye un proceso verificable en varios linajes distintos (e.g., Hathliacynidae y Borhyaenidae; véase Marshall, 1978). Por su parte, el M4 (MLP 92-X-10-11) carece de metacono. Sin embargo, la longitud de la preparacrista (relativamente corta) y la amplitud del talón (relativamente grande) son más generalizados que en la mayor parte de los esparasodontes hasta ahora conocidos, incluyendo las formas generalizadas del Paleoceno y Eoceno. Una excepción a esto último es el caso de los Hondadelphidae, probables Sparassodonta (Marshall et al., 1990) procedentes de niveles laventenses (Mioceno Medio) de La Venta, Colombia. Hondadelphys fieldsi Marshall, 1976 muestra en sus molares inferiores una notable reducción del metacónido y, en el M4, una amplia cuenca del trígono. Sin embargo, la morfología del M4 de Hondadelphys es aun más generalizada que la del ejemplar MLP 92-X-10-11: el metacono, si bien reducido, está claramente presente y el paracónulo está bien desarrollado. La pertenencia de uno o ambos ejemplares a un taxón asignable o afin a los Hondadelphydae constituye una hipótesis atractiva; sin embargo, el carácter fragmentario de los ejemplares colhuehuapenses impide contrastar adecuadamente esta hipótesis. Por esta misma razón tampoco se puede determinar si ambos pertenecen a un mismo taxón. En suma, los mismos podrían corresponder a avanzados Didelphimorhia carnívoros o, por el contario, a muy generalizados Sparassodonta.
Orden SPARASSODONTA Ameghino, 1894
Familia HATHLIACYNIDAE Ameghino, 1894Género Sipalocyon Ameghino, 1887
Sipalocyon externus (Ameghino, 1902) Figuras 6.A-D
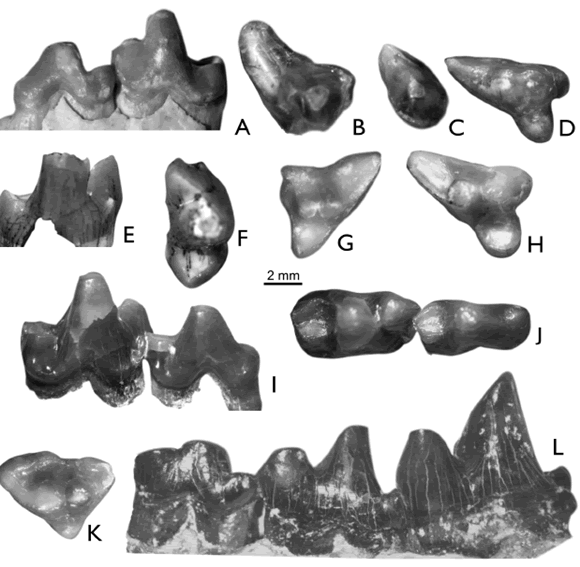
Figura 6. A-D, Sipalocyon externus (Ameghino, 1902). A, MPEF-PV 5840, m1-2 izquierdos en vista labial. B-C, MPEF-PV 5832, M3 derecho sin el protocono (B) y M4 izquierdo completo (C) en vistas oclusales. D, MLP 82-V-2-85, M1 derecho completo en vista oclusal. EG, Sipalocyon sp., E-F, MPEF-PV 4769, un molar inferior derecho aislado en vista labial (E) y oclusal (F), G, MPEF-PV 4545, un M3 izquierdo aislado en vista oclusal. H, Cladosictis centralis Ameghino, 1902, MPEF-PV 5854, un M1 derecho completo en vista oclusal. IJ, Cladosictis sp., MLP 82-V-2-109, m1-2 derechos en vistas labial (I) y oclusal (J). K-L, Hathliacynidae indet. K, MPEF-PV 4766, un M?1 derecho aislado, L, MPEF-PV 4770, m1-3 en vista lingual / A-D, Sipalocyon externus (Ameghino, 1902). A, MPEF-PV 5840, left m1-2 in labial view. B-C, MPEF-PV 5832, a right M3 lacking the protocone (B) and complete left M4 (C) in occlusal views. D, MLP 82-V-2-85, complete right M1 in occlusal view. E-G, Sipalocyon sp., E-F, MPEF-PV 4769, an isolated, right lower molar in labial (E) and occlusal (F) views, G, MPEFPV 4545, an isolated left M3 in occlusal view. H, Cladosictis centralis Ameghino, 1902, MPEF-PV 5854, a complete right M1 in occlusal view. IJ, Cladosictis sp., MLP 82-V-2-109, right m1-2 in labial (I) and occlusal (J) views. K-L, Hathliacynidae indet., K, MPEF-PV 4766, an isolated right M?1, L, MPEF-PV 4770, m1-3 in lingual view.
Cladosictis externa Ameghino, 1902: 129
Sipalocyon externa Marshall, 1978: 42
Especie tipo. MACN 52-383, un fragmento de maxilar izquierdo con los P3-M3 completos.
Material referido. MPEF-PV 5840, un fragmento mandibular izquierdo con los m1-2 completos (figura 6.A); MPEF-PV 5832, un fragmento de maxilar izquierdo con el P2, un M3 sin el protocono y el M4 completo (figuras 6.B-C), todos probablemente asociados; MLP 77-VI-13-7, un fragmento de M2 aislado; MLP 82-V-2-85, fragmento de maxilar derecho con el M1 completo (figura 6.D)
Procedencia geográfica y estratigráfica. Barranca Sur del Lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Fm. Sarmiento, Miembro Colhue Huapi, Mioceno Inferior (Edad-mamífero Colhuehuapense). El ejemplar MLP 77-VI-13- 7 fue colectado por H. Herrera; los materiales pertenecientes a las colecciones del MPEF-PV fueron colectados por personal del MLP y DU en 1999. Medidas. Véanse las tablas 1 y 2.
Tabla 1. Medidas (en milímetros) de la dentición inferior de los Sparassodonta aquí estudiados. El asterisco indica que la medida en cuestión es aproximada / measurements (in mm) of the lower dentition belonging to the Sparassodonta here studied. An asterisk indicates that the corresponding measurement is approximate.
Tabla 2. Medidas (en milímetros) de la dentición superior de los Sparassodonta aquí estudiados. El asterisco indica que la medida en cuestión es aproximada / Measurements (in mm) of the upper dentition belonging to the Sparassodonta here studied. An asterisk indicates that the corresponding measurement is approximate.
Descripción. El ejemplar MPEF-PV 5840 (figura 6.A) es asignable a S. externus en base a las siguientes consideraciones: (1) la morfología dentaria de los m1-2 muestra todas las características de los representantes del género, de cuya especie tipo (S. gracilis Ameghino, 1887) sí se conocen series dentarias inferiores: a diferencia de las especies de Cladosictis Ameghino, 1887, las especies de Sipalocyon son de menor tamaño, los molares inferiores son de aspecto más grácil, presentan el cíngulo posterolabial menos desarrollado y los talónidos son proporcionalmente más amplios. (2) Si bien no se conocen restos dentarios inferiores de S. externus, el tamaño del ejemplar aquí referido a esta especie es menor que el de S. gracilis y, a su vez, se corresponde con aquél de los dientes de la serie superior. Una posible diferencia entre los m1- 2 de S. gracilis y S. externus es la mayor compresión labiolingual en esta última, si bien el avanzado desgaste del ejemplar MPEF-PV 5840 no permite ser concluyentes al respecto.
El ejemplar MPEF-PV 5832 (figuras 6.B-C) consiste en un fragmento maxilar derecho que ha preservado el P2 completo y los alv éolos del canino y primer premolar así como, parcialmente, los de los P3-M3; las piezas dentarias asociadas son un M3 fragmentario y un M4 completo. Tanto la longitud de la serie superior como las dimensiones del M3 coinciden con aquellas del ejemplar tipo; en la porción anterior del maxilar se observan pequeños diastemas entre el alvéolo del canino y el P1, así como también entre este último y el P2. No se ha preservado el segundo premolar superior en el holotipo de Sipalocyon externus. Confrontado el ejemplar MPEF-PV 5832 con el P2 de la especie tipo del género se aprecia que ambos comparten la presencia de cúspulas anterior y posterior desarrolladas y una cresta posterior menos subvertical que la anterior. Sin embargo, y a diferencia de aquella especie, la región posterior es proporcionalmente más ancha y la cúspula más desarrollada. El M3, aunque fragmentario (carece de protocono), es extremadamente similar al del ejemplar tipo de S. externus, si bien la plataforma estilar labial al paracono es más angosta y el ectoflexo es menos marcado. Asociado a los restos antes mencionados existe un M4 completo; el mismo difiere de aquel de S. gracilis principalmente en la ausencia de protocono y en su mayor reducción en relación con M3. Tanto el M3 ya mencionado como este M4, muy probablemente correspondan al mismo ejemplar que el representado por el maxilar asociado. De ser este el caso, es destacable que el M4 está más simplificado (el protocono está ausente) que el diente homólogo de S. gracilis.
El ejemplar MLP 82-V-2-85 (figura 6.D) es un fragmento de maxilar derecho que conserva el M1 completo pero fuertemente gastado. En sus pocos rasgos confrontables es indiferenciable del diente hom ólogo presente en el tipo de S. externus. El ejemplar MLP 77- VI-13-7 constituye un M2 izquierdo; salvo por su preparacrista, aparentemente más desarrollada que la del diente homólogo del ejemplar tipo (aunque este rasgo podría estar exagerado por el mayor desgaste presente en este último), ambos molares son indiferenciables en sus partes confrontables.
Comentarios. Sipalocyon externus era conocido, hasta la revisión de Marshall (1981), sólo por su ejemplar tipo, un maxilar izquierdo aislado con los P3-M3. Aquí se refieren nuevos materiales a esta especie, incluyendo los primeros elementos de la dentición inferior. En líneas generales esta especie es muy similar, aunque algo más pequeña, que S. gracilis de edad Santacrucense. Sin embargo, de confirmarse la pertenencia de todos los elementos del ejemplar MPEF-PV 5832 a S. externus, las mayores diferencias corresponderían a la gran simplificación coronaria (ausencia del protocono) del M4 de esta especie con respecto a S. gracilis.
Sipalocyon sp. Figuras 6.E-G
Material. MPEF-PV 4769 (figuras 6.E-F), un molar inferior derecho aislado (m4).
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi; Localidad y nivel: "Mono Este Superficial". Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por personal del MLP y DU.
Medidas. Véase la tabla 1.
Descripción y comentarios. El ejemplar tiene la cúspide del protocónido y la raíz anterior rotas (figura 6.E). En el extremo anterior del trigónido se aprecia un cíngulo anterobasal corto, verticalmente orientado y de escaso desarrollo, el cual bordea labialmente una fosa hipoconúlida poco profunda. La paracrístida está bien desarrollada y se orienta oblicuamente en relación al eje dentario. El talónido es corto y angosto, con sólo el hipocónido y el hipoconúlido diferenciados.
El ejemplar es similar al m4 de Sipalocyon gracilis, difiriendo de este último fundamentalmente en la menor profundidad y desarrollo anteroposterior del talónido. No se conocen dientes homólogos asignables a S. externus, por lo que es imposible asignar con seguridad el ejemplar MPEF-PV 4769 a esta última especie.
Material. MPEF-PV 4545 (figura 6.G), un M3 izquierdo aislado.
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por personal del MLP y DU.
Medidas. Véase la tabla 1.
Descripción y comentarios. El material es muy similar a los M3 de Sipalocyon gracilis, siendo sus principales diferencias las siguientes: menor tamaño, protocono más reducido y comprimido anteroposteriormente, StB más desarrollada; por último, la muesca parastilar, para el alojamiento de la esquina metastilar del M2, es más profunda.
Género Cladosictis Ameghino, 1887
Cladosictis centralis Ameghino, 1902 Figura 6.H
Especie tipo. MACN 11639, un fragmento de rostro con el paladar casi completo, aunque sin dientes.
Material referido. MPEF-PV 4772, un fragmento de M2 o M3 izquierdo aislado; MPEF-PV 5854, M1 derecho completo (figura 6.H).
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). El ejemplar MPEFPV 4772 proviene del "Nivel C". Ambos materiales fueron colectados por personal del MLP y DU en 1999.
Medidas. Véase la tabla 2.
Descripción y comentarios. Si bien no existían ejemplares conocidos para la dentición superior de Cladosictis centralis, el ejemplar MPEF-PV 5854 (figura 6.H) es referible a esta especie por varias razones: su tamaño es menor que el del diente homólogo de la especie santacrucense C. patagonica Ameghino, 1887 y el protocono es comparativamente más desarrollado. El ectoflexo, muy reducido en C. patagonica, en este ejemplar es casi inexistente. Se trata entonces de una forma más pequeña y generalizada que la correspondiente a la especie C. patagonica; su menor tamaño concuerda con el menor tamaño de C. centralis, del cual sólo se conoce un paladar edéntulo. Confrontado con los alvéolos correspondientes al M1 de dicho paladar, el ejemplar MPEF-PV 5854 coincide en sus proporciones y dimensiones generales, siendo apenas más pequeño.
El ejemplar MPEF-PV 4772 consiste en un M2 o M3 izquierdo parcialmente preservado. Comparte las características generales del ejemplar anterior: tamaño menor y protocono más desarrollado que el de la especie tipo del género.
Los nuevos materiales aquí asignados a C. centralis constituyen el primer registro de dientes superiores para esta especie. Su menor tamaño y mayor desarrollo de los protoconos constituyen rasgos generalizados en relación a la especie santacrucense C. patagonica. Su mayor antigüedad coincide con esta apreciación.
Cladosictis sp. Figuras 6.I-J
Material. MLP 82-V-2-109, un fragmento mandibular derecho con los m1-2 completos.
Procedencia geográfica y estratigráfica. Puesto Almendra, Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por R. Pascual y colaboradores del MLP en 1982.
Medidas. Véase la tabla 1.
Comentarios. El ejemplar (figuras 6.I-J) es un veinte por ciento más pequeño que los m1-2 del MNHN col. 4 de Cladosictis centralis; sin embargo, comparte mayores similitudes con este último que con cualquier otro Hathliacynidae estudiado por nosotros. Tanto la posición del entocónido en los m1-2 como el tamaño relativamente grande del hipoconúlido en ambos es muy similar. Entre las diferencias, se destaca la mayor extensión posterior del cíngulo labial al hipocónido en el m2, el tamaño comparativamente mayor de la fosa hipoconúlida y el aspecto apenas más comprimido de los m1-2 (fundamentalmente el protocónido) en el ejemplar MNHN col.4.
Material. MPEF-PV 4768, un molar inferior derecho aislado (m?3).
Procedencia geográfica y estratigráfica. Puesto Almendra, Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por R. Pascual y colaboradores del MLP.
Medidas. Largo, 7,32 mm; ancho del trigónido, 3,76 mm.
Descripción y comentarios. El molar muestra las cúspides del trigónido muy desgastadas. Anteriormente, el cíngulo anterobasal es corto y subvertical. El protocónido está muy desarrollado, el talónido es muy corto y angosto y está roto a la altura del hipoconúlido. Labialmente muestra un cíngulo moderadamente desarrollado, sobre todo a la altura de la pared posterolabial del hipocónido.
El resto concuerda en tamaño y morfología con aquellos del m3 de Cladosictis centralis (la única especie de Cladosictis de Edad-mamífero Colhuehuapense), si bien el hipocónido del MPEF-PV 4768 está algo menos desarrollado. Por esta razón referimos este ejemplar a Cladosictis sp.
HATHLIACYNIDAE indet. Figuras 6.K-L
Material. MPEF-PV 4771, un premolar inferior aislado.
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por personal del MLP y DU en 1999.
Medidas. Véase la tabla 1.
Comentarios. El ejemplar carece de la base de la corona y de las raíces y muestra un moderado desgaste de su cúspide y cresta posterior; su diámetro anteroposterior coincide con el del p3 de Cladosictis patagonica si bien es proporcionalmente más comprimido labolingualmente, la cresta anterior está poco desarrollada tal como ocurre en otros hatliacínidos.
Material. MPEF-PV 4767, un talónido derecho aislado.
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. "Mono Este Superficial". Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por personal del MLP y DU.
Medidas. Ancho del talónido, 2,41 mm.
Comentarios. El material (un talónido aislado) es demasiado fragmentario como para intentar siquiera una aproximación genérica. Su tamaño y aspecto general recuerdan al de los m1 de Sipalocyon externus .
Material. MPEF-PV 4766 (figura 6.K), un molar superior derecho (M?1).
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. "Mono Este Superficial". Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por personal del MLP y DU.
Medidas. Largo, 6,27 mm; ancho, 4,78 mm.
Descripción y comentarios. El ejemplar muestra una morfología coronaria completa, aunque desgastada a la altura de la postmetacrista; las tres raíces están ausentes. En vista oclusal este molar es bastante simétrico, con poca diferencia de tamaño entre el paracono (menor) y el metacono (mayor). Si bien la postmetacrista es más desarrollada que la preparacrista, ésta no alcanza el desarrollo relativo de otros Hathliacynidae. El protocono es bajo y reducido. La plataforma estilar muestra un reborde labial más desarrollado que en otros Hathliacynidae.
La relativa simetría anterior-posterior observable en el ejemplar MPEF-PV 4766 obedece a la escasa diferencia de tamaño entre el paracono y metacono, a la ubicación poco excéntrica del protocono (i.e., el mismo se ubica más centralmente sobre la cara lingual del diente) y al relativamente escaso desarrollo de la postmetacrista. Este último rasgo sugiere la pertenencia del molar a un M1, si bien es de destacar que la postmetacrista del resto de los Hatliacynidae está más desarrollada. El tamaño del MPEF-PV 4766 coincide aproximadamente con el de los M1 de las especies de Cladosictis, si bien ninguna de ellas muestra reborde alguno sobre la cara labial de la plataforma estilar.
Podría argumentarse que este ejemplar corresponde en realidad a un dP3 de un Borhyaenidae de gran tamaño. Las ilustraciones provistas por Marshall (1978: 38-39, Figs. 8-9) sobre un ejemplar deseadense (Oligoceno Medio-Superior) de cf. Pharsophorus sp. (AMNH 29591) recuerdan a la morfología del ejemplar MPEF-PV 4766, si bien debe señalarse que: (1) el dP3 AMNH 29591 carece casi por completo de protocono y (2) posee dos raíces. Por el contrario, el ejemplar MPEF-PV 4766 muestra un (reducido) protocono y presenta tres raíces.
Material. MPEF-PV 4770, una hemimandíbula casi completa con los i3-m4 (figura 6.L).
Procedencia geográfica y estratigráfica. Colectado por Mario Cozzuol y Teresa Dozo en Gaiman (provincia del Chubut, Argentina). Mioceno Temprano (Edad-mamífero Colhuehuapense).
Medidas. Véase la tabla 1.
Comentarios. A pesar de ser inusualmente completo, resulta muy compleja la asignación de este ejemplar a alguna de las especies de Hathliacynidae reconocidas hasta la fecha, así como también su asignación siquiera genérica. En primer lugar, no es posible asignarlo a ninguna de las especies del género Cladosictis. El único material hasta ahora conocido de Cladosictis centralis es, además del tipo, el ejemplar MNHN col.4, un par de hemimandíbulas izquierda y derecha del mismo ejemplar, con la dentición casi completa. Confrontado con este material, el ejemplar MPEF-PV 4770 muestra diferencias notorias a pesar de su tamaño similar: el canino del ejemplar MPEF-PV 4770 es más ancho y robusto (i.e., menos comprimido lateralmente) que el del ejemplar MNHN col.4; los premolares, sobre todo los p1-2 son más cortos, anchos (i.e., más subovales en sección) y su cúspide media está volcada más anteriormente, por lo que las crestas anterior y posterior de los p1-3 son más asimétricas; los p1-2 carecen de las cúspulas anterobasales que se aprecian en el ejemplar MNHN col. 4 (vestigial en el p1 y más desarrollada en el p2); finalmente, el p1 se implanta algo oblicuamente, con su extremo anterior más labial que el posterior, tal como ocurre con sus raíces. La serie molar también presenta diferencias, a pesar de que la longitud m1-4 es apenas menor que la del ejemplar MNHN col.4. En primer lugar, los m1-3 carecen del hipoconúlido, relativamente bien desarrollado en C. centralis, lo cual seguramente incide en el hecho de que la longitud molar sea más corta; en segundo lugar, los molares no están tan comprimidos labiolingualmente como los materiales conocidos de C. centralis. Tal como puede observarse en el poco desgastado talónido del m3, éste era más corto que en el diente homólogo de C. centralis, con el hipocónido mucho más bajo que el entocónido. Labial y basalmente al hipocónido no se aprecia en los m1-4 del MPEF-PV 4770 el pequeño cíngulo que se observa en el MNHN col.4. Finalmente, el talónido del m4 es más corto y estrecho, con la cuenca claramente más reducida. Los cuerpos mandibulares muestran también algunas diferencias menores: en ambos se aprecian varios forámenes mentonianos, el mayor de los cuales se ubica más anteriormente (aunque en la mandíbula izquierda del ejemplar MNHN col. 4 éste se aprecia como un foramen doble). En el ejemplar MPEF-PV 4770, el foramen mentoniano mayor (anterior) abre en un punto por debajo de la raíz posterior del p2; un segundo foramen mucho más pequeño y algo más dorsal que el anterior abre por debajo de la raíz posterior del p3; el último foramen, aparentemente comprimido en sentido dorsoventral, abre por debajo de la raíz anterior del m2. Por el contrario, los tres forámenes mentonianos visibles en la hemimandíbula derecha del ejemplar MNHN col.4 de C. centralis abren a la misma altura en el cuerpo mandibular, siendo el mediano el más comprimido dorsoventralmente. Los puntos de apertura de dichos forámenes también difieren. El anterior lo hace en un punto por debajo de la raíz posterior del p1 y de la raíz anterior del p2, el mediano por debajo y algo delante de la raíz poste- rior del p3, mientras que el tercero lo hace por debajo de la raíz posterior del m1. En la cara lingual del cuerpo mandibular se aprecia que la sínfisis mandibular en el ejemplar MPEF-PV 4770 es proporcionalmente más baja, más extendida posteriormente (llegando hasta un punto por debajo de la raíz posterior del p3) y con un mayor desarrollo de rugosidades en la superficie sinfisaria que en el ejemplar MNHN col. 4 de C. centralis. Un pequeño foramen sinfisario se localiza algo por delante y por debajo de la raíz anterior del p1, a diferencia del existente en C. centralis, en el cual el mismo es de mayor tamaño abriendo por debajo del canino.
Las diferencias morfológicas entre ambos ejemplares no permiten asignar al ejemplar MPEF-PV 4770 a Cladosictis centralis Ameghino, 1902 a pesar de su contemporaneidad. Tampoco es posible asignarlo a C. patagonica, prácticamente por las mismas diferencias morfológicas que las ya señaladas para C. centralis, con el agregado de que C. patagonica es claramente mayor.
En segundo lugar, el ejemplar MPEF-PV 4770 tampoco puede ser asignado al género Sipalocyon, cuya especie más generalizada, S. externus, está registrada en niveles colhuehuapenses. Esta especie es de tamaño claramente menor, con los premolares y molares más comprimidos labiolingualmente y los talónidos de los molares más desarrollados especialmente el del m4 que contrasta notablemente con el del ejemplar MPEF-PV 4770.
En tercer lugar, es dudosa, aunque menos improbable, la asignación de este material al género Anatherium Ameghino, 1887 (si bien el género Anatherium es cuestionable; Forasiepi et al., en prensa). El mismo incluye dos especies, una de ellas de Edad-mamífero Colhuehuapense (A. herrerae Marshall, 1981) y la otra de Edad-mamífero Santacrucense (A. defossus Ameghino, 1887). Los talónidos de las especies de este género son más reducidos que aquellos de las especies de Cladosictis y Sipalocyon. Sin embargo, los premolares son bien comprimidos labiolingualmente, con sus crestas anterior y posterior más simétricas, y el talónido del m1 es francamente unicuspidado, a diferencia de este ejemplar en el que, a pesar del desgaste, parece haberse desarrollado una cuenca. La confrontación del ejemplar MPEF-PV 4770 con el MACN-9, asignado por Marshall (1981) a A. defossus, sugiere similitudes en al menos dos rasgos derivados: el hipocónido del m3 es muy bajo, por lo que la cuenca del talónido semeja un plano inclinado hacia la cara labial (rasgo más acentuado y exagerado por el desgaste en A. defossus); en segundo lugar, la cuenca del talónido del m4 está notablemente reducida y bien circunscripta (aunque menos en la cara lingual del MPEF-PV 4770). Otros rasgos, como el escaso desarrollo y orientación de los cíngulos anterobasales y la ausencia de cíngulos labiales a los hipocónidos en los m1-4, también aproximan al ejemplar MPEF-PV 4770 a las especies de Anatherium.
Las especies santacrucenses Pseudonotictis pusillus Marshall, 1981 y Perathereutes pungens Ameghino, 1891 tienen dimensiones notablemente más pequeñas y sus mandíbulas son de aspecto más grácil que la del ejemplar MPEF-PV 4770. Ninguno de estos taxones se encuentra representado en niveles de edad Colhuehuapense.
Finalmente, posibles comparaciones con otros Hathliacynidae podrían incluir a la especie deseadense Notogale mitis Ameghino, 1897, similar en tamaño aunque descartable por el mayor tamaño de los talónidos de sus molares, y Procladosictis anomala Ameghino, 1902, de tamaño mucho menor y sin materiales conocidos representativos de la dentición inferior.
En síntesis, la confrontación del ejemplar MPEFPV 4770 con otros hatliacínidos arroja numerosas dudas sobre la asignación de este ejemplar a alguno de los taxones reconocidos. Sin embargo, un análisis de la totalidad de los materiales asignables a ambas especies del género Anatherium podría confirmar la pertenencia de dicho ejemplar a este género.
Familia BORHYAENIDAE Ameghino, 1894
Subfamilia PROTHYLACYNINAE (Ameghino, 1894) Trouessart, 1898Género Pseudothylacynus Ameghino, 1902
Pseudothylacynus rectus Ameghino, 1902 Figuras 7.A-C

Figura 7. A-C, Pseudothylacynus rectus Ameghino, 1902. A-B, MLP 92-X-10-7, M1-2 derechos en vista lingual (A) y oclusal (B), C, MLP 92-X-10-5, p2-3 derechos en vista oclusal. D-G, Borhyaena macrodonta (Ameghino, 1902), D-E, MLP 82-V-4-3, P2-3 derechos en vista lingual (D) y oclusal (E), F, MLP 92-X-10-8, un m2 izquierdo aislado en vista labial, G, MPEF-PV 1467, detalle del M1 izquierdo en vista oclusal. H-J, Arctodictis sinclairi Marshall, 1978, H, MPEF-PV 5846c, un M1 izquierdo en vista oclusal, I-J, MPEF-PV 4752, detalle de los los m3-4 izquierdos en vistas oclusal (I) y labial (J) / A-C, Pseudothylacynus rectus Ameghino, 1902. A-B, MLP 92-X-10-7, right M1-2 in lingual (A) and labial (B) views, C, MLP 92-X-10-5, right p2-3 in occlusal view. D-G, Borhyaena macrodonta (Ameghino, 1902), D-E, MLP 82- V-4-3, right P2-3 in lingual (D) and occlusal (E) views, F, MLP 92-X-10-8, an isolated left m2 in labial view, G, MPEF-PV 1467, detail of left M1 in occlusal view. H-J, Arctodictis sinclairi Marshall, 1978, H, MPEF-PV 5846c, a left M1 in occlusal view, I-J, MPEF-PV 4752, detail of left m3- 4 in occlusal (I) and labial (J) views.
Especie tipo. MACN 52-369, un fragmento mandibular izquierdo con los p1-m4 completos.
Material referido. MNHN col 5, un fragmento mandibular izquierdo con los p3-m4; MPEF-PV 5838, un incisivo aislado, un p1 derecho aislado, un p?3 derecho aislado, un premolar superior o inferior aislado; MLP 92-X-10-7, un fragmento de maxilar derecho con los M1-2 completos (figuras 7.A-B); MPEF-PV 4765, un fragmento mandibular derecho con el m3 y el m4 en proceso de erupción; MLP 92-X-10-5, un fragmento mandibular derecho con los p2-3 completos (figura 7.C).
Procedencia geográfica y estratigráfica. Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). El ejemplar MPEF-PV 5838 fue coleccionado por personal del MLP y DU; el MPEF-PV 4765 lo fue por personal del MPEF, mientras que los ejemplares pertenecientes al MLP fueron colectados por personal del MLP y UNSW en noviembre de 1992.
Medidas. Véanse las tablas 1 y 2 para los ejemplares MLP 92-X-10-7, MLP 92-X-10-5 y MPEF-PV 4765. Para el ejemplar MPEF-PV 5838, las medidas son: p1.largo, 6,11 mm; ancho, 3,38 mm; p?3. largo, 8,75 mm aprox.; ancho, 5,12 mm; ?Px. largo, 8,75 mm aprox.; ancho, 5,36mm.
Descripción y comentarios. El ejemplar MPEF-PV 5838 consiste en cuatro dientes aislados: un incisivo y tres premolares. Hasta ahora no se habían preservado incisivos de Psudothylacynus rectus; sin embargo, su tamaño y el hecho de que el material está asociado a los premolares confirman su pertenencia a esta especie. El p1 es similar al diente homólogo de Pseudothylacynus rectus. Un segundo premolar aquí asignado a un p?3 es algo más grande y ancho que el p3 preservado en el tipo de esta especie. Sin embargo, el ejemplar MNHN col. 5 muestra su p3 algo más grande y ancho que en el tipo, si bien no alcanza las dimensiones del MPEF-PV 5838. El tercer premolar aislado es claramente más ancho en su mitad poste- rior que en la anterior; siendo más ancho y convexo que los premolares inferiores hasta ahora conocidos de P. rectus.
El ejemplar MLP 92-X-10-5 (figura 7.C) es apenas más grande que el del tipo de P. rectus, si bien ambos premolares muestran una morfología casi idéntica. Estos dientes son asimétricos en vista lateral, con la ladera anterior más corta y abrupta que la posterior; el p2 es comprimido lateralmente y su (muy poco desarrollada) cresta anterior termina en una cúspula basal; la cresta posterior es también de escaso desarrollo y culmina en un talón posterior unicuspidado. El p3 es muy similar al anterior con las siguientes diferencias: su tamaño es algo mayor, la cúspula anterobasal se ubica en una posición algo más lingual, la cresta anterior está proporcionalmente más desarrollada y, en el holotipo de P. rectus, el talón posterior de este molar es tricuspidado y de muy escaso desarrollo, mientras que en el MLP 92-X-10-5 está algo más desarrollado y es unicuspidado. Sin embargo, las diferencias observadas podrían obedecer, en vista de las notables similitudes morfológicas entre ambos ejemplares, más bien a la variabilidad intraespecífica.
El ejemplar MPEF-PV 4765 corresponde a un individuo juvenil, apreciándose sólo parte del trigónido del m4, ya que éste estaba aún en proceso de erupción. El m3, completamente erupcionado, muestra una morfología general esencialmente idéntica a la de los ejemplares MACN 52-369 (tipo de Pseudothylacynus rectus) y MNHN Col. 5. Debido a su menor desgaste, el ejemplar MPEF-PV 4765 permite comprender mejor la morfología del talónido de P. rectus: se aprecia un minúsculo entocónido en el extremo anterolingual del talónido, que está conectado a través de una cresta dentada al bien desarrollado hipoconúlido; la cara interna de esta cresta constituye la mayor parte de la cuenca del talónido. Por delante y labialmente al hipoconúlido se aprecia un hipocónido moderadamente desarrollado; el hipocónido emite hacia adelante dos crestas, una anterior dirigida hacia la base posterior del protocónido y otra anterolabial que termina antes de contactar con la pared posterolabial del protocónido. Más adelante (véase la Discusión) se desarrolla una hipótesis sobre las homologías existentes en las estructuras del talónido de los Prothylacyninae versus las de los Borhyaeninae, con los cuales parecen mantener importantes diferencias estructurales.
El ejemplar 92-X-10-7 (figuras 7.A-B) constituye un interesante aporte al conocimiento de Pseudothylacynus rectus, ya que hasta el momento no se conocían dentaduras superiores asignables a esta especie. La confrontación entre el ejemplar MPEF-PV 4765 (véase más arriba) con el maxilar MLP 92-X-10-7 permite apreciar un ajuste oclusal perfecto entre el M2 y el m3 de ambos ejemplares. Por este motivo, asignamos el MLP 92-X-10-7 a Pseudothylacynus rectus. El ejemplar consiste en un fragmento de maxilar derecho con los M1-2 casi completos. Ambos molares muestran un desgaste moderado a nivel de la postmetacrista y en los ápices de las cúspides; a juzgar por el tamaño de los alvéolos del M3 (de los que se han preservado la mitad anterior del álveolo anterolabial y anterolingual) el M3 era notablemene más grande que el M2, lo cual coincide con el rápido aumento de tamaño de los molares superiores que caracterizan a esta subfamilia. La raíz posterior de los M1-2, que soporta el área metastilar del diente, es notablemente mas grande (más del doble) que las raíces anteriores. El área del paladar colindante con estos molares está saltuariamente perforado y, entre los M2-3 (y presumiblemente entre los M3-4 también) muestra una fosa profunda para alojar el protocónido de los molares antagonistas inferiores. Los M1-2 son de contorno subtriangular, con el área metastilar bien desarrollada y el protocono presente aunque reducido. El área protoconal difiere significativamene de aquella de los Borhyaeninae tanto por su desarrollo relativo como por la morfología y sus contactos oclusales. En el ejemplar MLP 92-X-10-7 no existe en esta área una verdadera cuenca del trígono, sino más bien una única cúspide robusta y de sección subcircular (el protocono). Labialmente a esta cúspide se observan las laderas linguales del paracono y metacono fusionadas entre sí, formando una cara profunda y casi plana desde la muesca de la centrocrista hasta la base labial del protocono. Por el efecto del desgaste se forma una muy pequeña cuenca en la ladera labial del protocono, rasgo observable tanto en el M1 como en el M2. El protocono muestra distintas facetas de desgaste en sus tres caras libres: lingual (sobre todo en el M1), anterior (sobre todo en el M2) y posterior (en ambos molares).
El paracono, si bien reducido y fusionado parcialmente al metacono, persiste como una cúspide claramente individualizable y, a diferencia de aquel de los Borhyaeninae no está comprimido anteroposteriormente. Labialmente al paracono se aprecia una pequeña cúspide estilar (StB) conectada a la preparacrista a través de una muesca. En el M2 la StB es mucho más grande y algo comprimida labiolingualmente. Si bien en ambos molares la esquina parastilar está algo deteriorada, no parece haberse desarrollado un cíngulo anterobasal en la misma (a diferencia de los Borhyaeninae, en los que este cíngulo parece estar presente en forma más constante). Inmediatamente por detrás de la StB se desarrolla un pequeño ectoflexo, el cual es apenas menos profundo en el M1 que en el M2. A diferencia de los Borhyaeninae, en los molares de este ejemplar y de otros Prothylacyninae existe una verdadera cara labial independiente de la postmetacrista. Tanto en este ejemplar como en los de Prothylacynus patagonicus se aprecia, en el borde oclusal de dicha cara, una serie de cúspulas de homologías inciertas, alineadas anteroposteriormente en toda su longitud. La postmetacrista está bien desarrollada en ambos molares.
Comparados con los M1-2 de Prothylacynus patagonicus Ameghino, 1891, las mayores diferencias con los de este ejemplar son las siguientes: tamaño menor en P. rectus, StB aparentemente más comprimida en el M2 de esta especie y ectoflexo algo más desarrollado en el M1. Estas diferencias, al igual que las existentes entre las denticiones inferiores de ambas especies, permiten apreciar las estrechas afinidades existentes entre los representantes de ambos géneros. Comparadas las denticiones inferiores de los ejemplares tipo de ambas especies, se observan diferencias que tampoco contradicen lo señalado más arriba. Además de su menor tamaño, las mayores diferencias entre ambas son las siguientes: molares inferiores más gráciles y comprimidos labiolingualmente, p1 más oblicuamente orientado en P. patagonicus (aunque en P. rectus ya se orienta levemente en forma oblicua) y p3 oblicuamente orientado hacia adentro y con su corona de aspecto "torsionado" en P. patagonicus. Las tres primeras diferencias podrían obedecer al carácter más pequeño, grácil y generalizado de P. rectus; sin embargo, el aspecto "torsionado" o "retorcido" de la corona del p3 en P. patagonicus es de más difícil interpretación. Marshall (1979) había notado las mayores afinidades existentes entre ambas especies, más que con cualquier otro Prothylacyninae conocido.
Finalmente, tal como se destaca en la discusión, resultan notables las similitudes morfológicas generales y homologías del talónido entre el "Borhyaeninae" Plesiofelis schlosseri Roth, 1903 y los Prothylacyninae; tanto que en nuestra opinión dicho taxón debería ser referido en lo sucesivo a esta última subfamilia.
Subfamilia BORHYAENINAE (Ameghino, 1894) Cabrera, 1927
Género Borhyaena Ameghino, 1887
Borhyaena macrodonta (Ameghino, 1902) Figuras 7.D-G
Especie tipo. MACN 52-390 (tipo de Pseudoborhyaena macrodonta Ameghino 1902), un resto craneano con los P2-M4 izquierdos, asociado a un fragmento mandibular derecho con la raíz del p1, los p2-m3 completos y un fragmento del m4.
Material referido. MACN 52-366 (tipo de Pseudoborhyaena longeva Ameghino, 1902), un fragmento mandibular derecho con los c-p1 completos, parte del p2, raíces del p3, m1 completo, raíces del m2 y los m3-4 parcialmente rotos; MLP 82-V-4-3, fragmento de maxilar derecho con los P2-3 completos (figuras 7.D-E); MLP 92-X-10-8, un m2 izquierdo aislado (figura 7.F); MPEF-PV 1467, fragmento de maxilar izquierdo con los P3-M3 (figura 7.G, detalle del M1).
Procedencia geográfica y estratigráfica. El ejemplar MLP 82-V-4-3 fue coleccionado en La Curandera (provincia del Chubut, Argentina) por Pascual, Bond, Carlini y Reguero en 1982; Mioceno Inferior (Edad-mamífero Colhuehuapense). El MPEF-PV 1467 fue recolectado por Raúl Vacca en Puesto Almendra. Los especímenes MACN 52-390, MACN 52-366 y MLP 92-X-10-8 proceden de la Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense).
Medidas. Véanse las tablas 1 y 2.
Descripción y comentarios. El ejemplar MLP 82-V-4- 3 (figuras 7.D-E) ha preservado solamente la raíz posterior del P1 y los P2-3 completos. Las dimensiones de estos últimos coinciden ajustadamente con las del tipo del Borhyaena macrodonta. Las diferencias entre las escasas partes confrontables son de índole menor: el cíngulo posterolingual y la cúspide posterior del P2 parecen estar más desarrollados en este ejemplar que en el tipo, así como también el desarrollo apenas mayor del talón posterior del P3, aunque esta última diferencia es difícil de establecer debido al mayor grado de desgaste del material tipo en esta área. El ejemplar MLP 92-X-10-8 (figura 7.F), un m2 casi completo, mantiene muy pocas diferencias con el diente homólogo del ejemplar tipo de esta especie: los cíngulos anterobasal y posterolingual son algo menos desarrollados.
El ejemplar MPEF-PV 1467 (figura 7.G) también muestra diferencias de carácter menor con el maxilar del holotipo: el P3 tiene un talón posterior aparentemente más desarrollado en la cara labial del diente y, en general, el "capuchón" de esmalte que lo recubre está menos desarrollado que en el P3 del ejemplar tipo. Los molares preservados están muy desgastados, por lo que es difícil precisar diferencias significativas con los M1-3 del holotipo; algunas pocas diferencias observadas son las siguientes: el M1 es de menor tamaño (no así los M2-3) y el protocono está menos desarrollado que en el M1 del holotipo; en el M3 del holotipo de Borhyaena macrodonta se aprecian un par de cúspulas (¿supernumerarias?) por detrás de la StB; finalmente, sobre la cara lingual de los M1-3 se aprecia que las coronas de estos molares son más bajas en el ejemplar MPEF-PV 1467 que en el holotipo de B. macrodonta.
Género Arctodictis Mercerat, 1891
Arctodictis sinclairi Marshall, 1978 Figuras 7.H-J
Especie tipo. FMNH P13526, un fragmento mandibular izquierdo con el canino, los p1-2 fragmentarios, el p3 completo, las raíces del m1 y los m2-3 completos aunque desgastados.
Material referido. MLP 85-VII-3-1, un cráneo y mandíbulas casi completos con buena parte del esqueleto postcraneano asociado; MLP 93-XI-18-9, un canino superior derecho aislado; MPEF-PV 5846c, un M1 izquierdo aislado, fragmentario (figura 7.H); MPEFPV 4752, un fragmento mandibular izquierdo con los m3-4 completos (figuras 7.I, J); MLP 82-V-2-4c, un canino superior izquierdo aislado; MLP 77-VI-13-27, un m2 izquierdo aislado; MLP 82-V-2- 4a, un m2 derecho aislado; MPEF-PV 5846d, un P2 derecho aislado; MLP 82-V-2-116, un P3 derecho aislado.
Procedencia geográfica y estratigráfica. Excepto el ejemplar MLP 77-VI-13-27 (cuyo único dato de procedencia es "Gran Barranca"), el resto de los materiales procede de la Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). Todos los ejemplares pertenecientes al MPEF fueron colectados por personal del MLP y DU; el MLP 77-VI-13-27 fue colectado por Hebe Herrera en 1977; el MLP 82-V-2-4a y 4c fueron colectados por Mariano Bond en 1982; el MLP 82-V-2-116 por "Pascual y otros" (según reza la etiqueta acompañante) en 1982; el MLP 85-VII-3-1 fue colectado por Juan Quiroga en 1985.
Medidas. Véanse las tablas 1 y 2.
Descripción y comentarios. El ejemplar MLP 85-VII- 3-1 de A. sinclairi es notablemente completo, por lo que su análisis permitirá ampliar considerablemente nuestro conocimiento sobre esta especie, uno de los mayores carnívoros colhuehuapenses. Este ejemplar está siendo estudiado actualmente por Analía Forasiepi, por lo que no se abundará aquí en mayores detalles sobre el mismo.
Los ejemplares MLP 77-VI-13-27, MLP 82-V-2-4a y MPEF-PV 4752 muestran su morfología molar mucho mejor preservada que la del ejemplar tipo, FMNH P 13526. El MLP 77-VI-13-27 (un m2 izquierdo) es prácticamente idéntico, aunque mejor preservado y apenas más grande, que el diente homólogo del tipo. El gran desarrollo de la paracrístida es el rasgo más destacado de este molar, que muestra un cíngulo anterobasal prácticamente vestigial, un talónido muy corto y unicuspidado y un cíngulo labial al talónido muy desarrollado. El MLP 82-V-2-4a, un m2 derecho, es esencialmente idéntico auque peor preservado que el ejemplar anterior; la cúspula del talónido está algo más comprimida labiolingualmente que la del MLP 77-VI-13-27.
El ejemplar MPEF-PV 4752 (figura 7.I-J) ha preservado los m3-4. Si bien el m3 del tipo de Arctodictis sinclairi está muy desgastado y es algo más grande que el del MPEF-PV 4752, las características de este molar, así como también del que le sigue posteriormente, permiten asignarlos con pocas dudas a los m3-4. El m3 muestra su talón también unicuspidado, aunque en este caso la cúspide está levemente comprimida en sentido labiolingual y el cíngulo labial al talónido está menos expandido labialmente que en el m2 de los ejemplares MLP 77-VI-13-27 y FMNH P 13526. El m4 del ejemplar MPEF-PV 4752 es un diente muy robusto, también con el talónido unicuspidado, aunque ambos (talónido y cúspide) muestran un desarrollo vestigial. La paracrístida es comparativamente enorme, así como también la raíz anterior del diente, la cual soporta al trigónido
El MLP 93-XI-18-9 parece corresponder a un canino superior derecho. Es un diente muy robusto, con el capuchón de esmalte ausente y numerosos surcos y crenulaciones en el área radicular. El ápice preservado muestra en su cara anterolingual una faceta de desgaste. El diente es relativamente plano en la cara lingual, sobre todo en la porción radicular; en la porción media de esta cara se aprecia un surco que la recorre desde la raíz.
El ejemplar MLP 82-V-2-4c corresponde a la porción distal de un canino superior izquierdo. Es muy similar, aunque más fragmentario y apenas más grande, que el ejemplar anterior. La faceta de desgaste presente en su cara anterolingual está mucho más desarrollada que en el ejemplar anterior.
EL MPEF-PV 5846d constituye un P2 derecho aislado. Si bien la raíz anterior está ausente prácticamente desde la base de la corona, a juzgar por su sección es claro que era mucho más pequeña y estrecha que la raíz posterior. El premolar es mucho más ancho en su mitad posterior que en la anterior. Anteriormente, muestra una cresta de aspecto poco cortante. La cúspide mayor de este diente es robusta y se ubica en la porción central. Toda la mitad posterior posterior del diente duplica a la anterior, hecho debido al gran desarrollo de un talón, sobre todo en el extremo posterolingual del diente. Una pequeña cúspide baja se ubica por detrás de la gran cúspide central del diente.
El MLP 82-V-2-116 correponde a un P3 derecho; el diente es grande y unicuspidado, con la raíz anterior más grande y ancha que la posterior. El talón posterior es ancho y se continúa hasta la mitad de la cara lingual del diente. Es difícil precisar si sobre el borde posterior de este talón se desarrolló algún tipo de cúspide.
El MPEF-PV 5846c (figura 7.H) corresponde a un M1 izquierdo roto a la altura de la metacrista, por lo que su tercio posterior está ausente. Si bien la plataforma estilar está reducida, se aprecia como una estrecha repisa sobre el borde labial del diente, culminando anteriormente en un cíngulo o cúspide moderadamente desarrollado. El paracono es más pequeño que el metacono mientras que el protocono, ubicado muy por debajo, está extremadamente reducido.
cf. Arctodictis sinclairi Marshall, 1978
Material referido. MLP 77-VI-13-1, una hemimandíbula izquierda casi completa con los i1-m4 completos, una hemimandíbula derecha con los i1 e i3, y los c-m1 completos, y un canino superior aislado, probablemente asociado; MLP 84-III-15-1, fragmento de rama mandibular derecha con los m2-4 completos y un M1 derecho aislado asociado; MPEF-PV 5846a, un p2 izquierdo aislado.
Procedencia geográfica y estratigráfica. El ejemplar MLP 84-III-15-1 procede de Puesto Almendra, mientras que los ejemplares MLP 77-VI-13-1 y MPEF-PV 5846a lo hacen de "Gran Barranca". Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). El ejemplar MLP 77-VI-13-1 fue colectado en 1977 por Hebe Herrera; el MLP 84-III-15-1 en 1984 por Alfredo Carlini, mientras que el ejemplar MPEF-PV 5846a lo fue por personal del MLP y DU.
Medidas. Véanse las tablas 1 y 2.
Nota. Los ejemplares MLP 77-VI-13-1 y MLP 84-III- 15-1 fueron coleccionados en la Barranca Sur del lago Colhue Huapi con una década de diferencia. Curiosamente, sin embargo, ambos parecen corresponder no solo al mismo taxón sino incluso al mismo individuo. En efecto, los planos de rotura de la hemimandíbula derecha del primero y del borde anterior mandibular del segundo son complementarios de tal modo, que al juntarlos encastran perfectamente entre sí. No sería esta la primera vez que distintos ejemplares colectados en diferentes momentos en una misma localidad resultan pertenecer a un mismo individuo. En consecuencia, en este trabajo se refieren ambos especímenes a un mismo individuo, al cual optamos por denominar como MLP 77-IV-13-1+MLP 84-III-15- 1. Ambos fragmentos mandibulares fueron pegados y así están alojados en las colecciones del MLP.
Descripción y comentarios. El ejemplar MPEF-PV 5846a es casi idéntico en su tamaño y morfología al p2 del MLP 77-IV-13-1+MLP 84-III-15-1. Este último muestra mayores similitudes con los materiales conocidos de Arctodictis sinclairi que con cualquier otro borienino conocido. En efecto, la longitud p1-m4 es casi idéntica y tanto los premolares como los molares muestran una morfología sumamente similar. Entre las diferencias, se destaca el aspecto más grácil de los premolares y la persistencia, en el m4, de una cúspula posterior al protocónido. Sin embargo, las mayores diferencias se registran en el tamaño relativo de los caninos inferiores, mucho más grandes en el tipo de A. sinclairi y en el MLP 85-VII-3-1 que en este ejemplar. Asimismo, el cuerpo mandibular en los primeros es notoriamente más alto y robusto que en este último. Finalmente, los incisivos inferiores del ejemplar MLP 85-VII-3-1 son de mayor tamaño que los de éste. Un mejor conocimiento de la variabilidad intraspecífica en Arctodictis sinclairi podría demostrar la pertenencia del ejemplar MLP 77-IV-13-1+MLP 84-III-15-1 a esta especie. Por el momento, sin embargo, y hasta tanto dicha revisión no se haga efectiva, preferimos referir el ejemplar como cf. Arctodictis sinclairi.
Género Acrocyon Ameghino, 1887
Acrocyon riggsi (Sinclair, 1930) Figuras 8.A-B
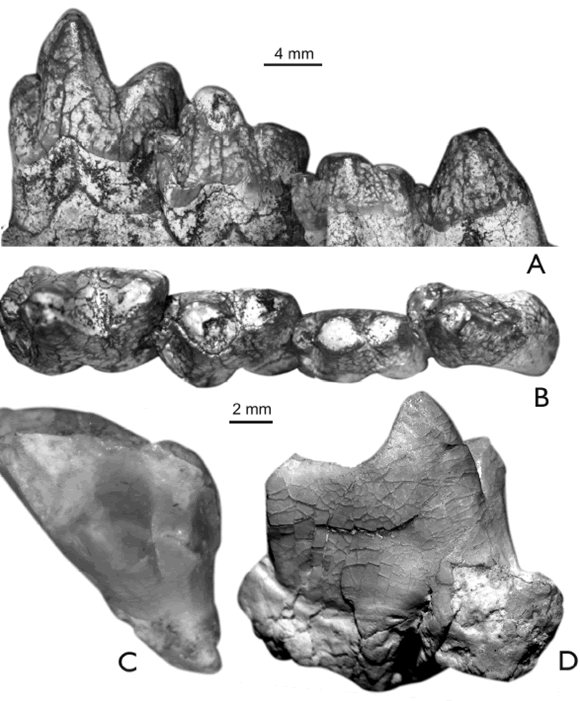
Figura 8. A-B, Acrocyon riggsi (Sinclair, 1930); MLP 85-II-1-1; detalle de los p3-m3 derechos en vistas labial (A) y oclusal (B). C-D, Thylacosmilidae gen. et sp. indet.; MLP 92-X-10-6, un molar superior derecho (M?3) en vistas oclusal (C) y posterior (D) / A-B, Acrocyon riggsi (Sinclair, 1930); MLP 85-II-1-1; detail of right p3-m3 in labial (A) and occlusal (B) views. C-D, Thylacosmilidae gen. et sp. indet.; MLP 92- X-10-6, an upper right molar (M ?3) in occlusal (C) and posterior (D) views.
Especie tipo. FMNH P 13433 (tipo de Borhyaena riggsi Sinclair 1930), un fragmento craneano parcialmente preservado incluyendo la dentición.
Material referido. MACN 52-385, un canino inferior derecho, fragmento de un incisivo y una falange, todos supuestamente asociados. MLP 85-II-1-1, un fragmento mandibular derecho con los p3- m3 completos (figuras 8.A-B).
Material tentativamente referido. MPEF-PV 1620, un fragmento mandibular derecho con el m3 o m4 en proceso de erupción.
Procedencia geográfica y estratigráfica. El ejemplar MLP 85-II-1-1 fue exhumado junto a la Ruta Provincial N° 59, entre Gan Gan y Escorial, a unos 35 km de este último, provincia del Chubut, Argentina. Mioceno Inferior (Edad-mamífero Colhuehuapense).
Medidas. Véase la tabla 1.
Descripción y comentarios. El ejemplar MLP 85-II-1- 1 (figuras 8.A-B), constituye la primera evidencia de dientes postcaninos inferiores para esta especie. Su m1 difiere del diente homólogo de la especie tipo del género, Acrocyon sectorius Ameghino, 1887 (MLP 11- 70) en su carácter más corto, en el menor alineamiento anteroposterior del paracónido, protocónido y cúspide del talónido, como así también en el mayor desarrollo de cíngulos labial y lingual a esta cúspide. Comparados los m3 del nuevo ejemplar de A. riggsi y del MACN 9364 de A. sectorius, se aprecia que ambos mantienen similitudes esenciales. Un rasgo, en particular, parece ser diagnóstico de Acrocyon: la presencia de una cresta corta pero claramente definida en la ladera posterior del protocónido. Dicha cresta podría constituir un vestigio del metacónido (ya muy reducido en otros Borhyaeninae) desplazado más posteriormente; sin embargo, la evidencia disponible no permite ser concluyentes al respecto. Podría argumentarse también su correspondencia con una postprotocrístida vestigial. De todos modos, este rasgo distingue a las especies de Acrocyon del resto de las especies confrontadas de Borhyaeninae. Un segundo rasgo, el mayor alineamiento anteroposerior de las cúspides de los molares inferiores (no sólo del m1) también resulta diagnóstico del género. El resto mandibular preservado en el ejemplar MLP 85-II-1-1, revela que la rama mandibular era robusta y la sínfisis probablemente fusionada en los adultos, llegando por detrás hasta un punto por debajo de la raíz anterior del p3. La cara lingual de la rama horizontal es plana a levemente cóncava, mientras que la labial es muy poco convexa. Se distinguen en esta última no menos de seis forámenes mentonianos, un rasgo inusual en los Borhyaeninae con excepción de las especies de Arctodictis. Finalmente, al igual que lo que ocurre con el P3, el p3 muestra un fuerte talón posterobasal, extendido hacia atrás en una plataforma relativamente bien desarrollada.
Hemos tenido la oportunidad de estudiar un calco parcial del ejemplar MPEF-PV 1620, en el que se aprecia un m3 o m4 en proceso de erupción. El molar muestra una cresta muy notoria por detrás del protocónido, rasgo que, como ya se ha señalado, parece ser diagnóstico de Acrocyon. Su tamaño es algo mayor que el del m3 del ejemplar MLP 85-II-1-1, si bien su aspecto general y, aparentemente, el importante desarrollo del cíngulo anterobasal, coinciden con el m3 de este último.
BORHYAENINAE Indet.
Material. MLP 85-VII-3-46, un fragmento de maxilar izquierdo con parte del alvéolo del canino, el P1 completo y las raíces de los P2-3; MLP 82-V-2-4b, un incisivo inferior aislado; MPEF-PV 5637a, un incisivo inferior aislado; MPEF-PV 5637c, un fragmento mandibular derecho con los alvéolos del p2; MPEF-PV 5637b, un fragmento de m2 derecho; MPEF-PV 5846b, un P3 izquierdo aislado.
Procedencia geográfica y estratigráfica. El único dato de colección de los materiales pertenecientes al MLP es: "Gran Barranca". Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Formación Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edadmamífero Colhuehuapense). Todos los ejemplares del MPEF fueron colectados por personal del MLP y DU.
Medidas. Véase la tabla 2.
Descripción y comentarios. Dado que los incisivos no presentan caracteres diagnósticos a nivel genérico, los ejemplares MPEF-PV 5637a y MLP 82-V-2-4b son asignados aquí a Borhyaeninae indet. Del mismo modo, y teniendo en cuenta su estado de preservación, los ejemplares MPEF-PV 5637b, MPEF-PV 5637c y MLP 85-VII-3-46 carecen de elementos que permitan asignarlos a alguno de los géneros o especies de esta subfamilia. El ejemplar MLP 85-VII-3-46 ha preservado el P1 completo y las raíces de los P2-3; todas las raíces de estos premolares están fuertemente infladas; la orientación de al menos el P1 es casi transversal al eje dentario. Estos rasgos coinciden con aquellos presentes en los grandes Borhyaeninae con inferidos hábitos osífragos, como los de algunas especies de Borhyaena y ambas especies de Arctodictis. La porción de paladar preservada en este ejemplar muestra una serie de forámenes relativamente grandes, rasgo que caracteriza también a los representantes de Sparassodonta. El ejemplar MPEF-PV 5846b, un P3 izquierdo, comparte varios rasgos en común con los dientes homólogos de A. sinclairi; sin embargo, su menor tamaño y el desigual desarrollo del cíngulo lingual nos inclinan a mantenerlo como Borhyaeninae indet.
Familia THYLACOSMILIDAE Riggs, 1933
THYLACOSMILIDAE gen. et sp. indet. Figuras 8.C-D
Material. MLP 92-X-10-6 un molar superior derecho (M?3).
Procedencia geográfica y estratigráfica. Puesto Almendra, Barranca Sur del lago Colhue Huapi (Departamento de Sarmiento, provincia del Chubut, Argentina). Fm. Sarmiento, Miembro Colhue Huapi. Mioceno Inferior (Edad-mamífero Colhuehuapense). Colectado por personal del MLP-UNSW en noviembre de 1992
Medidas. Largo aproximado, 7,15 mm; ancho aproximado, 12,25 mm.
Descripción. El molar (figuras 8.C-D), está casi completo aunque resquebrajado en varios puntos y con toda la cara labial parcialmente arrasada. A pesar de su tamaño relativamente grande es de aspecto grácil, rasgo que lo distingue visiblemente de los molares de los Borhyaenidae. El carácter más conspicuo de este molar es el gran desarrollo relativo de la postmetacrista, la cual forma un plano vertical casi continuo con la cara posterolingual del protocono. La muesca de la postmetacrista es marcadamente en "V". El protocono está reducido y comprimido en sentido anteroposterior, desarrollando una quilla lingual, tal vez exagerada por la rotura del diente en este punto. El paracono es pequeño y más bajo que el metacono; ambas cúspides son coalescentes en la base. La ladera anterior del metacono es relativamente plana excepto en el área donde se desarrolla la centrocrista, esta última es tenue pero claramente visible; la preparacrista se orienta de forma relativamente transversal al eje dentario, no pudiendo precisarse la estructura de su extremo distal debido al grado de preservación en este punto. Las raíces que sostienen al protocono y a la postmetacrista están notablemente comprimidas en sentido anterolabial-posterlingual. La plataforma estilar está rota labialmente, por lo que es imposible precisar su extensión; no obstante, parece evidente la gran reducción e incluso ausencia de cúspides estilares.
Comentarios. El carácter fragmentario de este molar aislado impide asignarlo con precisión a alguno de los grupos de Sparassodonta hasta ahora conocidos. Sin embargo, son varios los rasgos que lo alejan de los Sparassodonta hasta ahora revisados en este trabajo. Fundamentalmente se destacan el aspecto grácil del molar a pesar de su tamaño relativamente grande, la gran compresión de al menos dos de las raíces (la tercera está rota) en sentido anterolabial-posterolingual; el aspecto delgado del área metastilar (i.e., el ancho de la postmetacrista), el desarrollo de una quilla lingual al protocono y el aspecto marcadamente en "v" de la muesca de la postmetacrista (mientras la porción proximal de la postmetacrista desciende abruptamente hacia la muesca, la porción distal asciende claramente después de la misma; en los Hathliacynidae y Borhyaenidae, por el contrario, la porción distal de la postmetacrista se orienta en forma más subhorizontal al plano oclusal del diente). Todos estos rasgos excluyen a este ejemplar de los Hathliacynidae y Borhyaenidae (Prothylacininae + Borhyaeninae) y, en cambio, lo aproximan a los fuertemente especializados Thylacosmilidae, especialmente con un primitivo ejemplar de edad Friasense s. l. ("Colloncurense"; Mioceno Medio), aún no catalogado, presente en las colecciones del MLP.
MAMMALIA incertae sedis
Familia NECROLESTIDAE Ameghino, 1894
Género Necrolestes Ameghino, 1891
Especie tipo. Necrolestes patagonensis Ameghino 1891.
Especies incluidas. N. patagonensis y N. mirabilis sp. nov.
Distribución. Mioceno Inferior de Patagonia.
Necrolestes mirabilis sp. nov. Figuras 9.A-D

Figura 9. A-D, Necrolestes mirabilis sp. nov. A, MPEF-PV 4706, un molariforme superior (P?4) en vista oclusal. B-C, MLP 92-X-10-14 (Tipo), un fragmento mandibular izquierdo mostrando un diente en proceso de erupción y, por encima, el diente deciduo al que reemplaza; B, vista labial, C, vista lingual. La flecha señala la presencia de estructuras óseas por detrás del último alvéolo edéntulo de este ejemplar. D, MPEF-PV 6030, un molariforme inferior en vista oclusal / A-D, Necrolestes mirabilis sp. nov. A, MPEF-PV 4706, an upper molariform (P?4) in occlusal view. B-C, MLP 92-X-10-14 (type), a left dentary fragment showing an erupting tooth and, above it, the deciduous tooth it replaces; B, labial view; C, lingual view. The arrow indicates bone behind the last edentulous alveolus. D, MPEF-PV 6030, a lower molariform in occlusal view.
Etimología. Del latín mirabilia, admirable, en referencia a la notable circunstancia de que el tipo ha preservado un diente deciduo a punto de desprenderse del diente permanente (p?4) al que reemplaza.
Holotipo. MLP 92-X-10-14, un fragmento mandibular izquierdo con un diente en erupción (p?4), el diente deciduo al que reemplaza (dp?4) y los alvéolos edéntulos de la serie molar (figura 9.B, C).
Hipodigma. El tipo y MPEF-PV 4706, un molariforme superior (P?4; figura 9.A); MPEF-PV 6030, un molariforme inferior (m?1; figura 9.D); MPEF-PV 4548, un molar superior izquierdo; MPEF-PV 4549, un premolariforme superior; MPEF-PV 4546, un premolariforme superior.
Procedencia geográfica y estratigráfica. Puesto Almendra, Gran Barranca (Barranca sur del lago Colhue Huapi) Departamento de Sarmiento, provincia del Chubut, Argentina. Miembro Colhue Huapi, Formación Sarmiento; Mioceno inferior (Edad-mamífero Colhuehuapense). El tipo fue coleccionado por personal del MLP-UNSW en 1992. Los ejemplares del MPEF fueron colectados por personal del MLP.DU en 1995.
Medidas. MLP 92-X-10-14 (diente deciduo): largo total, 0,84 mm; ancho total, 1,16 mm. MPEF-PV 6030: largo total, 1,2 mm; ancho total, 1,48 mm. MPEF-PV 4706: largo total, 0,96 mm; ancho total, 1,6 mm.
Diagnosis. Tamaño apenas mayor que la especie tipo del género; alvéolos de los molares inferiores más redondeados en su cara labial; borde ventral mandibular más subhorizontal; mayor diferencia de tamaño entre el p?4 y el m1; cúspides linguales de los postcaninos molariformes más separadas entre sí que en N. patagonensis.
Descripción. El holotipo consiste en un fragmento mandibular izquierdo que ha preservado el dp?4, el p?4 en erupción, y los alvéolos de los tres molares (sobre las homologías de los dientes postcaninos (véase más abajo). El cuerpo mandibular es relativamente alto, más alto a la altura del p?4 que del m3 y claramente más elevado en su cara lingual que en la labial. Los bordes alveolar y ventral de la rama horizontal son subparalelos y relativamente rectos. En vista oclusal se aprecia claramente el carácter oblicuo del plano alveolar, el cual es más alto lingual que labialmente. Los alvéolos de la serie molar (constituida por piezas unirradiculadas) son de aspecto suboval, con el borde labial más redondeado y el lingual más plano. El diámetro transverso del alvéolo del m3 es más corto que el de los m1-2; por lo tanto, su sección es más circular que suboval. Por detrás del último alvéolo, el cuerpo mandibular está roto; no obstante, la estructura del hueso preservado sugiere que no existían otros alvéolos por detrás del aquí identificado como m3. El diente deciduo (dp?4) ha preservado la totalidad de su corona. A diferencia de los molariformes permanentes, éste es un diente birradiculado. La raíz posterior preservada en este ejemplar está, casi en su totalidad, "anclada" todavía en su alvéolo original, mucho más pequeño que el alvéolo del premolar que lo reemplaza, el cual abre inmediatamente por delante de aquel. La raíz posterior del dp?4 está muy comprimida en sentido anteroposterior, probablemente más que la raiz anterior del mismo diente -si bien la rotura de esta última impide mayores precisiones-. La corona de este diente deciduo incluye, al igual que la de los molariformes permanentes, tres cúspides dispuestas en forma triangular, siendo dos de ellas de ubicación lingual mientras que la cúspide mayor lo hace labialmente. Esta última es mucho más alta que las cúspides linguales. La cara anterolabial del diente se proyecta en forma casi plana hasta el nacimiento de la raíz. En la base de la corona por debajo de la cúspide anterolingual se aprecia una cúspula que sobresale del vértice anterolingual del diente.
El p?4 estaba, al momento del deceso del ejemplar, en proceso de erupción, desplazando así al diente anteriormente descripto. Sus cúspides labial y posterolingual están encajadas a ambos lados de la raíz anterior del dp?4. El diámetro transverso de este diente y de su alvéolo es claramente menor que el diámetro del alvéolo correspondiente al m1, lo cual coincide con lo observado por nosotros en la especie tipo del género. La raíz de este diente es abierta, lo cual confirma su carácter protohipsodonte. Debido a la rotura del cuerpo mandibular se ha preservado toda la cara anterolabial del diente, tanto la que estaba erupcionada como la que permanecía en el espacio intralveolar; toda esta cara es plana y está surcada por estriaciones paralelas entre sí y perpendiculares al eje vertical del diente, tanto en el área intralveolar como extralveolar. Es interesante destacar que el mismo tipo de estriaciones aparece en la raíz posterior del diente deciduo, aunque no en la corona del mismo.
Al igual que los otros molariformes de Necrolestes, la corona del diente es prismática e incluye tres caras una más estrecha, la cara lingual, y dos más grandes y planas, anterolabial y posterolingual. En vista oclusal se aprecian tres cúspides, siendo la principal la cúspide labial y las dos más pequeñas orientadas lingualmente. La cresta correspondiente al borde de la cara anterolabial es de aspecto algo cortante y, en vista oclusal, levemente convexa hacia adelante.
El ejemplar MPEF-PV 6030 tiene el mismo patrón morfológico que los molariformes de N. patagonensis; por sus dimensiones parece corresponder a un m1 izquierdo. Como se expresó en la diagnosis, las cúspides linguales de este diente, al igual que las del MLP 92-X-10-14, están algo más separadas que las de los molares de la especie tipo del género; asimismo, la cúspide anterolingual es algo más comprimida que la posterolingual.
El ejemplar MPEF-PV 4706 (figura 9.A) es un diente postcanino superior aislado que ha preservado la corona y parte de la raíz; se trata de un molariforme muy comprimido en sentido anteroposterior. Presenta una cúspide principal, la lingual, y tres cúspides labiales de desarrollo desigual: dos de ellas están muy próximas entre sí y se ubican sobre el vértice anterolabial del diente; la tercera, de ubicación posterolabial, está separada de las anteriores por un surco y ocupa una posición mucho más baja. Las dos cúspides anterolabiales son pequeñas y están fusionadas en la base. La mayor de ellas es la posterior, la cual está comprimida en sentido labiolingual. La cara oclusal del diente se inclina levemente hacia la cara posterior. Comparado con los molariformes superiores de N. patagonensis, este ejemplar semeja más al P?4 que a cualquiera de los molares. En el P?4 de N. patagonensis la esquina posterolabial del diente es más saliente que la anterolabial; en el ejemplar MPEF-PV 4706 esta condición se aprecia con claridad. En líneas generales, este diente es más comprimido y su borde posterolabial más saliente que el de los ejemplares de N. patagonensis aquí estudiados.
El ejemplar MPEF-PV 4548, probablemente un molar superior izquierdo, es aún más comprimido que el MPEF-PV 4706 aquí asignado a un probable P?4. Se trata de un diente apenas más angosto que aquél, en el que la cúspide labial principal se ubica en la esquina posterolabial. Por delante de esta se proyecta una muy corta cresta que termina en una cúspula ubicada hacia el centro de la cara labial del diente. En la esquina anterolabial se alcanza a apreciar un muy pequeño cíngulo de orientación subvertical.
Los ejemplares MPEF-PV 4549 y MPEF-PV 4546 son dientes muy comprimidos en sentido labiolingual; son tricuspidados, con la cúspide central más alta y grande que las restantes. El MPEF-PV 4549 está sólo parcialmente preservado en una de sus caras (¿lingual?), la que carece por completo de la capa de esmalte. La morfología general de ambos ejemplares se corresponde con aquella del segundo premolar superior descripta para la especie tipo del género.
Comentarios. Si bien N. mirabilis sp. nov. constituye una especie algo más antigua que N. patagonensis, es imposible precisar su carácter ancestral a esta última: por un lado, su tamaño es algo mayor a la de la especie tipo del género; por el otro, N. mirabilis sp. nov. muestra un grado de especialización tan avanzado como el de aquella. Los restos aquí descriptos de N. mirabilis sp. nov. ofrecen evidencia contraria a la asignación de los Necrolestidae a los Metatheria (véase la Discusión).
Discusión
Una discusión general sobre los metaterios de la Edad-mamífero Colhuehuapense será ofrecida en la última entrega de esta serie. En consecuencia, en esta sección nos referiremos únicamente a los linajes estudiados en el presente trabajo: Didelphimorphia y Sparassodonta. En el primer caso, se comentan los hallazgos y se sugiere que los didelfimorfios aquí presentados constituyen la más antigua evidencia de "verdaderos" Didelphoidea (Goin, 1991, 1995), referibles tanto a los Didelphidae como, probablemente, a los Caluromyidae. En el segundo caso, se hace hincapié en la notable diversidad de esparasodontes colhuehuapenses, tanto en los aspectos taxonómicos como en la variedad de tamaños y zonas adaptativas representadas. Se discuten puntualmente aspectos vinculados con (a) la diversidad de Hathliacynidae y Borhyaenidae, (b) las más antiguas evidencias de Thylacosmilidae y (c) las homologías cuspidales del talónido de los Borhyaenoidea. Finalmente, se argumenta en contra de las afinidades metaterias de los Necrolestidae, considerados por Patterson (1958) como parte de la radiación Borhyaenoidea.
Didelphimorphia
Los más antiguos Didelphidae neógenos
Muy escasos restos confirman la presencia de didélfidos en la fauna colhuehuapense, los que constituyen la más antigua evidencia de representantes de este grupo para el Neógeno sudamericano. Un primer taxón, aquí innominado (figuras 5.A-C, G), incluye un par de molares inferiores y uno superior, muy desgastado. Representan probablemente al marsupial más pequeño hasta ahora exhumado en niveles colhuehuapenses, por lo que no debe extrañar la escasez de restos referibles a este taxón. Su tamaño es notablemente reducido, aún en el contexto de los más pequeños didelfoideos vivientes. Entre los molares inferiores sobresale el escaso desarrollo del entocónido, un rasgo que caracteriza a todos los Didelphoidea s.s. (Goin 1991, 1995). Recientemente Voss y Jansa (2003: 40) señalaron que la mayor parte de los didélfidos tienen "... a well-developed entoconid that is about as tall as the hypoconid and much exceeds the adjacent hypoconulid in height and breath on m1-3". Sin embargo, esto es correcto sólo en términos relativos; la reducción del hipoconúlido es una sinapomorfía de los Didelphimorphia en relación a los Peradectia, por lo que el tamaño relativo del entocónido debería medirse con respecto a otras cúspides del molar inferior. Con la sola excepción de los Caluromyidae (véase más abajo), los Didelphoidea sudamericanos vivientes tienen entocónidos de apenas moderado a muy escaso desarrollo (particularmente entre los Didelphini y Monodelphini sensu Goin y Rey, 1997). Entre los Didelphimorphia paleógenos el entocónido es una cúspide notoria y prominente en los molares inferiores, particularmente entre los Derorhynchidae y Protodidelphidae. En ambos casos el entocónido es de aspecto cónico, muy alto y comparativamente ancho en la base. Los Protodidelphidae Protodidelphis vanzolinii Paula Couto, 1952 y Guggenheimia brasiliensis Paula Couto, 1952 muestran la base del entocónido tan ensanchada que ésta ocupa buena parte de la mitad lingual de todo el talónido.
Muy posiblemente, el didélfido indeterminado estudiado en este trabajo represente una forma basal de marmosino (Marmosini sensu Reig et al., 1987), aunque la escasez de restos impide mayores precisiones. Ya se señaló que en los didélfidos modernos el entocónido es de desarrollo apenas moderado a muy escaso. Entre los didélfidos de pequeño tamaño, el clado que presenta el menor desarrollo del entocónido es el de Monodelphis y formas afines (como el extinto Thylatheridium cristatum Reig, 1952), esto es, los Monodelphini sensu Goin y Rey (1997). El bien preservado molar inferior correspondiente al ejemplar colhuehuapense MPEF-PV 4774 muestra, precisamente, un entocónido notablemente reducido a vestigial, por lo que en una primera aproximación podría especularse sobre la pertenencia de este taxón innominado a un grupo natural conjuntamente con Monodelphis y Thylatheridium.
La morfología molar del didélfido indeterminado que aquí se describe es compatible con hábitos alimentarios más probablemente insectívoros; de ello dan cuenta su pequeño tamaño y la gran altura y aspecto puntiagudo de las cúspides del trigónido, así como también el desarrollo moderado del talónido.
¿El más antiguo Caluromyidae?
Un pequeño molar inferior aislado (figuras 5.GH) podría constituir el más antiguo registro de un calurómido. La evidencia es sumamente fragmentaria, si bien debe señalarse que son varios los rasgos de este molar que así lo sugieren. En primer lugar, el trigónido no es comprimido lateralmente (i.e., en sentido labiolingual) sino anteroposterior. El protocónido es comparativamente bajo y el paracónido está bien desarrollado; el hipoconúlido no está apareado con el entocónido y este último es muy grande y alto, superando en altura al hipocónido. Finalmente, el hipocónido está algo "adelantado", por lo que la postcrístida no es perpendicular al eje dentario sino oblicua al mismo. El conjunto de estos rasgos es único entre los Didelphimorphia vivientes, si bien varios de ellos son compartidos con un taxón en particular: Caluromysiops irrupta Sanborn. Tal es el caso del hipoconúlido separado del entocónido, el gran desarrollo del entocónido y la posición adelantada del hipocónido. En esta última especie, aunque menos notoriamente que en el ejemplar MPEF-PV 4708, también se verifica un acortamiento del talónido, el cual es progresivo desde el m1 al m3 (véase Reig et al., 1987: Fig. 10.F). Las similitudes entre el MPEF-PV 4708 y otros calurómidos es más discutible. Al menos en Caluromys lanatus (Olfers, 1818) (no hemos podido estudiar especímenes o calcos de otros calurómidos) existe un cierto grado de reducción del cíngulo anterobasal en los molares inferiores. C. lanatus también muestra un entocónido agrandado, si bien este no es de forma cónica sino que está comprimido labiolingualmente. Un último rasgo también es compartido entre el ejemplar MPEF-PV 4708 y los Caluromyidae: la relativa reducción en altura del protocónido. En definitiva, si bien la evidencia es insuficiente, los escasos elementos disponibles sugieren que el ejemplar MPEF-PV 4708 sería referible a los Caluromyidae. De confirmarse esta asignación, el taxón innominado representado por este ejemplar podría resultar clave en la interpretación de las relaciones filogenéticas de esta familia. En efecto, algunos de los rasgos ya mencionados también están presentes en grupos de Didephimorphia paleógenos; tal es el caso de los Protodidelphidae: escaso desarrollo del cíngulo anterobasal, baja altura del protocónido y gran desarrollo del entocónido. Una hipótesis sistemática que relacione a protodidélfidos y calurómidos argumentaría a favor de la muy antigua diferenciación de este clado hipotético (contra Goin, 1994) y de su gran distancia filogenética de los didélfidos sudamericanos modernos. Al respecto es destacable que los Caluromyidae aparecen siempre como grupos basales en distintas hipótesis filogenéticas de didelphimorfios vivientes (e.g. Reig et al., 1987; Kirsch y Palma, 1995; Patton et al., 1996; Kirsch et al., 1997; Jansa y Voss, 2000; Voss y Jansa, 2003). Aún más interesante resulta el hecho de que en varios de estos estudios los Caluromyidae no aparecen como un grupo natural sino que Glironia venusta Thomas, 1912 ocupa una posición más basal que las especies de Caluromys y Caluromysiops (Kirsch et al., 1997; Jansa y Voss, 2000; Voss y Jansa, 2003)
Sparassodonta
La radiación neógena de los Borhyaenoidea
La tabla 3 muestra la clasificación de los metaterios estudiados en este trabajo. De ella se desprende que no menos de siete, y tal vez hasta diez especies de Sparassodonta fueron coetáneas en la Edad-mamífero Colhuehuapense, así como también distribuidas en un mismo espacio geográfico (sur y nordeste de la actual provincia del Chubut). Esta notable diversidad de metaterios carnívoros contrasta significativamente con aquélla de las edades precedentes y anticipa el pico de mayor diversidad alcanzado por los Sparassodonta en la Era Cenozoica, ocurrido en la subsiguiente Edad-mamífero Santacrucense. En síntesis, el registro disponible sugiere que a comienzos del Neógeno ocurrió una importante radiación en casi todos los grupos representativos de este orden, con dos excepciones: (1) los Proborhyaenidae, cuyo último registro corresponde a la inmediatamente precedente Edad-mamífero Deseadense; (2) los Hondadelphyidae (si es que este grupo es efectivamente referible a los Sparassodonta), restringidos al Mioceno Tardío de los trópicos sudamericanos. Todos los demás linajes de Sparassodonta están abundantemente representados en el Colhuehuapense de Patagonia.

Tabla 3. Clasificación de los taxones estudiados en este trabajo / classification of the studied taxa.
Origen de los Thylacosmilidae
Uno de los resultados más notables de este estudio es el registro más antiguo de un Sparassodonta Thylacosmilidae. El material aquí descripto está representado por un molar superior aislado que muestra las características básicas de los representantes de esta familia: corona dentaria muy alta y labiolingulamente comprimida, protocono vestigial, posmetacrista muy bien desarrollada y comprimida labiolingualmente. Los Thylacosmilidae eran hasta el momento conocidos para el Mioceno Medio-Tardío de Colombia (Edad-mamífero Laventense), el Mioceno Tardío del noroeste y centro de la Argentina (Edadmamífero Huayqueriense) y el Plioceno Tardío de la Región Pampeana (Edad-mamífero Chapadma_lalense). Un ejemplar aún no descripto, referible a un nuevo género de tilacosmílido, procede de niveles "colloncurenses" (Mioceno Medio) del oeste de la provincia de Río Negro. No se conocen Thylacosmilidae de edad Santacrucense. El notable grado de especialización de los Thylacosmilidae sugiere una mayor antigüedad de este linaje que la revelada por su biocrón hasta ahora conocido (Mioceno Medio-Plioceno Superior). En consecuencia, era esperable la presencia de representantes de esta familia en niveles de edad colhuehuapense. Tal como se señaló en la descripción de este material, resultan notables algunas de las especializaciones presentes en el molar superior de este tilacosmílido colhuehuapense, por lo que puede presumirse un origen más antiguo para los miembros de este grupo.
Muizon (1999) fue el último autor en realizar una revisión sobre las relaciones filogenéticas de los Sparassodonta y concluyó que tilacosmílidos y proboriénidos constituyen grupos hermanos, claramente derivados entre los representantes de este orden. El material colhuehuapense aquí descripto no permite contrastar esta hipótesis sistemática (las sinapomorfías que agrupan a los tilacosmílidos y proboriénidos en el análisis de Muizon no corresponden a rasgos de los molares superiores).
Homologías cuspidales del talónido de los Borhyaenidae
Para Marshall et al. (1990) los Sparassodonta incluyen: Stagodontidae (del Cretácico Superior de América del Norte) y las familias (del Cenozoico sudamericano) Hathliacynidae, Borhyaenidae (Prothylacyninae + Borhyaeninae), Proborhyaenidae, Thylacosmilidae y Hondadelphidae. Con posterioridad Muizon (1999) incluyó a los Mayulestidae, con un solo género y especie (Mayulestes ferox Muizon, 1994), del Paleoceno Inferior de Bolivia, como taxón basal de sus Borhyaenoidea. En la actualidad se excluye a los Stagodontidae de los Sparassodonta, así como también se han reconsiderado hipótesis sobre el origen y filogenia de los representantes de este grupo (véase, e.g., Rougier et al., 1998; Muizon, 1999). Dos de los grupos mejor representados de Sparassodonta sudamericanos son los protilacininos y los borieninos, los que tradicionalmente se agrupan en la familia Borhyaenidae. Tanto los protilacininos como los borieninos están bien representados en la fauna colhuehuapense, especialmente a partir de sus estructuras dentarias (véase más arriba).
Resulta instructivo contrastar la morfología del talónido de los más antiguos protilacininos conocidos (Pseudothylacynus, de edad colhuehuapense) con la de borhiénidos precolhuehuapenses (figura 10). El holotipo de Pseudothylacynus rectus (MACN 52-369) muestra un claro alineamiento de tres cúspides principales en los m1-3. En estos tres primeros molares de la serie es evidente que: (1) el paracónido, el protocónido y el hipocónido están alineados anteroposteriormente; (2) hacia el centro de la cara posterior del protocónido se aprecia el desarrollo de una pequeña cresta que acompaña la ladera del mismo con una orientación casi vertical, que en su extremo inferior se dirige a, pero no contacta con, el hipocónido. Esta cresta está más desarrollada en los m1-2 que en los m3-4. (3) Si bien no existe metacónido, en la ladera posterolingual del m4 y, en menor medida, la del m3, se aprecia una muy reducida cresta que se interrumpe bruscamente sin contactar con ninguna cúspide del talónido. Posiblemente esta cresta constituye un remanente del metacónido; (4) lingualmente al hipocónido se desarrolla en estos molares una pequeña cuenca o cíngulo, la que posiblemente constituye el remanente del talónido. Dicha cuenca o cíngulo se desarrolla en Pseudothylacynus progresivamente del m1 al m3; en el m1 está apenas esbozada y culmina anteriormente en un pequeño entocónido, cuya cresta anterior no alcanza a cerrar lingualmente la cuenca; en el m2 se aprecia una cresta (preentocrístida) que limita lingualmente al talónido; en el m3, si bien muy desgastado en el holotipo, se aprecia un mayor desarrollo relativo de toda la cuenca del talónido. El m4, por su parte, muestra un talónido visible aunque menos desarrollado que en el m3, por lo cual es imposible establecer con certeza las homologías implicadas en sus cúspides.

Figura 10. A, Pseudothylacynus rectus Ameghino, 1902, MACN 52-369, m1-4 izquierdos en vista oclusal. B, Plesiofelis schlosseri Roth, 1903; MLP 11-114, m1-4 izquierdos en vista oclusal. C, Pharsophorus lacerans Ameghino, 1897; MACN 52-391, m1-4 izquierdos en vista oclusal. D, Borhyaena macrodonta (Ameghino, 1902), MLP 77-VI-13-1, m1-4 izquierdos en vista oclusal / A, Pseudothylacynus rectus Ameghino, 1902, MACN 52-369, left m1-4 in occlusal view. B, Plesiofelis schlosseri Roth, 1903; MLP 11-114, left m1-4 in occlusal view. C, Pharsophorus lacerans Ameghino, 1897; MACN 52-391, left m1-4 in occlusal view. D, Borhyaena macrodonta (Ameghino, 1902), MLP 77-VI- 13-1, left m1-4 in occlusal view.
Resultan notables las similitudes existentes entre el patrón hasta aquí mencionado y aquel presente en el "Borhyaeninae" mustersense Plesiofelis schlosseri Roth, cuyo holotipo, MLP 11-114, muestra la dentición yugal casi completa. En este ejemplar, cuyos m2- 4 aun presentan el metacónido en la esquina posterolingual del protocónido, se aprecia incluso un mayor desarrollo relativo del "talónido" en los m1-2, además de un pequeño cíngulo basal y labial al hipocónido. En P. schlosseri se aprecia que todavía persiste el metacónido en la ladera posterolingual del protocónido de los m2 y m4 (el m3 está roto en este punto). El m1, por su parte, desarrolla una corta cresta casi central a la ladera posterior del protocónido; presumiblemente esta cresta es el remanente del metacónido más desplazado en este molar hacia el centro de la cara posterior del diente.
Notablemente distinta es la morfología del talónido en los m1-4 del Borhyaeninae Pharsophorus lacerans Ameghino, del Deseadense de Patagonia. En sus molares también se ha preservado el metacónido en los m2-4 (y vestigialmente en el m1, a juzgar por la pequeña cresta preservada en la ladera posterolingual del protocónido de este molar); asimismo, también se observa un alineamiento entre el paracónido, protocónido e hipocónido, pero a diferencia de Plesiofelis y Pseudothylacynus la pequeña cresta posterior al trigónido en los m1-3 no se conecta con el hipocónido sino con una minúscula cúspide lingual a aquél, i.e., el muy reducido entocónido. De hecho, en los m2-3 de Pharsophorus la cresta que relaciona al entocónido con el trigónido nace directamente del metacónido, mientras que en el m1 la pequeña cresta posterior al trigónido apunta claramente (aunque sin conectarse) al entocónido. Paralelamente, el hipocónido en los m2-4 no sólo disminuye de tamaño sino que todo el borde labial del talónido desciende notablemente en altura hasta confundirse con el cíngulo posterolabial del diente.
Tratándose Pharsophorus de un borienino cuya morfología molar es relativamente plesiomorfa, saltan a la vista las radicales diferencias en la conformación del talónido entre estos boriénidos (i.e., Borhyaeninae) y su grupo hemano, los Prothylacyninae: (1) en los Borhyaeninae persiste el hipocónido bien desarrollado sólo en el m1, mientras que en los m2-4 se reduce progresivamente mostrando una tendencia a fusionarse con el cíngulo posterolabial del diente; la cúspide posterior al trigónido en estos molares sería o bien el entocónido o bien el entocónido fusionado al metacónido; (2) En los m2-3 de los Borhyaeninae se produce un mayor desarrollo del cíngulo labial al talónido que de la propia cuenca del talónido (ya de por sí vestigial), formando así un pseudotalónido labial al hipocónido; por el contrario, en los Protylacyninae no se desarrolla ningún cíngulo labial a esta cúspide sino que, en cambio, se aprecia un mayor desarrollo relativo del talónido o pseudotalónido (véase más abajo); (3) mientras que en los Borhyaeninae primitivos (Pharsophorus) el metacónido se conecta con el entocónido, en los Prothylacininae primitivos (en nuestra interpretación, Plesiofelis) esta cúspide, ya de aspecto crestiforme, apunta directamente al hipocónido. Las homologías de la cúspide principal posterior al trigónido en cada uno de estos linajes ya fueron señaladas: hipocónido en los Prothylacyninae y entocónido (o entocónido más metacónido) en los Borhyaeninae.
De confirmarse las radicales diferencias en las homologías implicadas en la principal cúspide del talónido en ambos linajes de Borhyaenidae, las mismas podrían conducir a una nueva aproximación a las relaciones filogenéticas tanto dentro como entre cada uno de estos grupos:
1. Según nuestra interpretación Plesiofelis no sería un Borhyaeninae sino un verdadero Prothylacyninae. Una serie de rasgos adicionales apoyan la pertenencia de este taxón a esta última subfamilia: gran foramen mentoniano en el cuerpo mandibular por debajo del p2, los molares inferiores incrementan marcadamente su tamaño del m1 al m4, p1 no orientado transversalmente (ni siquiera muy oblicuamente) con respecto al eje dentario; finalmente, y a juzgar por la porción anterior mandibular preservada en el ejemplar MLP 11-114, el canino estaba sólo moderadamente desarrollado (véase Marshall, 1978, 1979).
2. La interpretación de las homologías aquí implicadas explica varios rasgos curiosos presentes en el talónido de boriénidos más avanzados, de edad post- Deseadense. Por ejemplo, el ejemplar MLP 77-VI-13- 1, tentativamente asignable a Borhyaena macrodonta del Colhuehuapense de Patagonia, muestra una conformación asimétrica en la estructura del talónido del m1 con respecto a la de los m2-4. En el primero existe una cúspide notoria por detrás del trigónido y un cíngulo relativamente bien desarrollado lingualmente a ésta; por el contrario, en los m2-4 de este mismo ejemplar no existe tal cíngulo lingual sino un mayor desarrollo del cíngulo posterolabial. En nuestra interpretación, la cúspide del talónido del m1 es homologable al hipocónido, y el "cíngulo lingual" a esta cúspide es un resto vestigial de talónido, mientras que la cúspide principal del talónido de los m2-4 es en realidad el entocónido, siendo el cíngulo labial el resultado de la desaparición del hipocónido o de su fusión con el cíngulo posterolabial al mismo, formando así un pseudotalónido ya presente en borieninos más primitivos como Pharsophorus.
3. Como las homologías implicadas en la porción posterior de los molares inferiores de borieninos y protilacininos son parcialmente inciertas (a excepción del hipocónido de los m1-4 de los protilacininos y del m1 de los borieninos, y por el otro lado del entocónido de los m2-4 de los borieninos), en rigor de verdad debería hablarse de un pseudotalónido para dichas estructuras en cada uno de estos grupos. En el caso de los Prothylacyninae esto se confirma al confrontar los dientes antagónicos de un mismo ejemplar y observar los contactos oclusales entre las respectivas cúspides y crestas. Así, se aprecia en el holotipo de Prothylacynus patagonicus que la cara anterior del protocónido ocluye con el área de la faceta de desgaste número 5 sensu Crompton (e.g. 1971), esto es, con la base de la postmetacrístida (en el caso de los Borhyaeninae está constituida por la cara posterior del protocónido). Sin embargo, la cara posterior del protocono, que debería ocluir con la cara anterolingual del entocónido según el modelo de relaciones oclusales propuesto por Crompton (i.e., con la faceta de desgaste número 6) ocluye aparentemente con el extremo anterior del paracónido (y cíngulo anterobasal?). Esto significa que no existe un verdadero entocónido y estructuras colindantes en los molares inferiores de los Prothylacininae. En consecuencia, es razonable deducir la existencia en este grupo de un pseudotalónido desarrollado a partir de los vestigios del talónido original. En contrapartida, dada la fusión parcial del hipocónido con el cíngulo posterolabial en los Borhyaeninae, resulta también razonable hablar de un pseudotalónido labial al hipocónido en los representantes de este grupo.
4. Como corolario de esto último puede deducirse que el origen de los Prothylacyninae debió haberse producido a partir de un ancestro cuya estructura del talónido ya mostraba una reducción importante. Si bien no es éste el único caso de estructuras homoplásicamente desarrolladas en grupos afines, resulta interesante constatar dicha evolución, con diferente significado funcional, en ambos linajes de Borhyaenidae: en un caso (Borhyaeninae) mediante un pseudotalónido labial vinculado con las estructuras de corte, en el otro (Prothylacyninae) a través de un pseudotalónido lingual al hipocónido relacionado con una nueva función masticatoria vinculada con la trituración del alimento.
Necrolestes mirabilis sp. nov. y las afinidades de los Necrolestidae
Necrolestes patagonensis, de edad Santacrucense, fue reconocido por Ameghino (1891) sobre la base de un resto mandibular izquierdo parcialmente preservado. Scott (1905) asignó tentativamente a esta especie una serie de restos mucho más completos también procedentes de niveles santacrucenses. Ameghino (1891), Scott (1905) y Gregory (1910) consideraron a Necrolestes como perteneciente a los "Insectivora". Tanto Ameghino como Scott sugirieron afinidades entre Necrolestes y los insectívoros africanos del género Chrysochloris Lacépède. Leche (1907) señaló varias similitudes tanto con los crisoclóridos como con el topo marsupial Notoryctes typhlops Stirling. Discusiones posteriores sobre la situación taxonómica de Necrolestes incluyen las de Romer (1945), Simpson (1945), Saban (1954), Winge (1941), Patterson (1958), Archer (1984), Aplin y Archer (1987) y Szalay (1994). Patterson (1958) discutió extensamente las afinidades de Necrolestes a partir del re-estudio de los mejores materiales disponibles hasta entonces. Este autor afirmó las afinidades marsupiales de Necrolestes y sostuvo que las mismas estaban más allá de cualquier duda. De la lectura del trabajo de Patterson (1958; véase las páginas 3 y 9) se deduce que el autor dió un peso importante al hecho de que Necrolestes posee, efectivamente, una fórmula incisiva marsupial (cinco incisivos superiores y cuatro incisivos inferiores). Curiosamente, Patterson culminó su ensayo refiriendo a Necrolestes tentativamente a los Borhyaenoidea, ninguno de los cuales (excepto Mayulestes, si es que este taxón es efectivamente referible a los Borhyaenoidea; véase Goin, 2003: 31-32) incluye una fórmula incisiva como la de Necrolestes. Más recientemente Szalay (1994) consideró a Necrolestes un Ameridelphia incertae sedis, si bien manifestó sus dudas sobre buena parte de los rasgos supuestamente metaterios argumentados previamente por Patterson (fórmula incisiva y molar, hueso yugal extendido hacia la cavidad glenoidea).
Tres rasgos principales fueron utilizados por Patterson (1958) para afirmar la condición marsupial de Necrolestes: proceso angular inflexionado (al respecto, véase Sánchez-Villagra y Smith, 1997), contribución del yugal a la cavidad glenoidea y fórmula incisiva 5/4. Szalay (1994) señaló que los últimos dos rasgos no son diagnósticos de Metatheria. Nuestras propias observaciones realizadas sobre los mismos materiales estudiados por Scott y Patterson nos indican que el proceso angular del dentario no está inflexionado sino, por el contrario, claramente dirigido hacia atrás; la mínima orientación lingual del extremo distal de dicho proceso no constituye, en nuestra opinión, un argumento definitivo a favor a su pertenencia a los Metatheria.
Las denticiones superior e inferior de la especie tipo del género, N. patagonensis, incluyen seis dientes postcaninos cada una. En ambas, el primero de ellos es birradiculado, de aspecto premolariforme y bien comprimido lateralmente. Por el contrario, los cinco restantes son unirradiculados, de sección subtriangular, molariformes y bien comprimidos en sentido anteroposterior, excepto el primero (algo menos comprimido). Patterson (1958) sostuvo que los dos primeros postcaninos corresponden a premolares, mientras que los cuatro restantes son referibles a la serie molar de los marsupiales. El análisis del tipo de Necrolestes mirabilis sp. nov. sugiere, sin embargo, una interpretación diferente; a saber, que la fórmula dentaria postcanina es de tres premolares y tres molares:
1. Por detrás del diente deciduo sólo existen en el cuerpo mandibular tres alvéolos, y por lo tanto tres dientes unirradiculados (aquí identificados como pertenecientes a la serie molar; véase más abajo). Ya se mencionó que por detrás del tercero de estos alvéolos no parecen haber existido otros. En consecuencia, y teniendo en cuenta que son los premolares (y no los molares) los que están precedidos en la ontogenia por piezas deciduas, es razonable concluir que la fórmula dentaria adulta de los necroléstidos incluía no más de tres molares. Una observación adicional fortalece este último argumento: el alvéolo más posterior preservado en N. mirabilis sp. nov. es más corto en su diámetro transverso que el inmediatamente precedente, lo que coincide con el menor tamaño del último molar de Necrolestes patagonensis con respecto al molar precedente.
2. El diente que reemplaza a la pieza decidua en el tipo de N. mirabilis sp. nov. es homólogo al tercer postcanino (segundo molariforme). En efecto, su morfología no se corresponde con aquélla del segundo postcanino (primer molariforme), el cual es bien distinguible porque la cúspide anterior de este último se proyecta hacia delante. A su vez, el diente es mucho más angosto labiolingualmente que el cuarto postcanino (tercer molariforme). Este rasgo es observable tanto en el tipo de N. mirabilis sp. nov. como en los dos restos mandibulares de N. patagonensis examinados por nosotros.
3. La diferencia en el ancho del diámetro transverso entre el tercer postcanino inferior y los tres posteriores a éste parece estar marcando el límite entre las series premolar y molar. La posición del tercer postcanino en la serie, y el hecho de que el diente es claramente un premolar (al estar reemplazando a una pieza decidua), sugiere que este diente sólo puede ser un tercer premolar, mientras que los dientes posteriores corresponderían al m1, m2 y m3. La presencia de diastemas entre el canino y el primer postcanino y entre el primer y segundo postcanino sugieren la posible pérdida de uno de los premolares anteriores, suponiendo una fórmula premolar ancestral superior a tres. Sin embargo, la existencia de diastemas no es, por sí misma, indicativa de una eventual pérdida de piezas dentarias.
Si bien es cierto que entre los marsupiales sudamericanos existen casos de pérdida del m4, estos son muy contados, ocurriendo sólo entre los Polydolopimorphia: los polidolópidos Polidolopinae (Marshall, 1982b), el prepidolópido Punadolops (Goin et al., 1998) y los argirolagoideos Klohnia (Flynn y Wyss, 1999) y Patagonia (Pascual y Carlini, 1987). Por el contrario, la pérdida de al menos un premolar en la fórmula postcanina de los Eutheria es común a muchos grupos. Si bien no apoyamos explícitamente aquí la pertenencia de Necrolestes a los Eutheria, entendemos que la suma de rasgos descriptos para el mismo no arroja evidencia inequívoca de su pertenencia a los Metatheria. No descartamos, incluso, eventuales afinidades con linajes ya extintos de Theria no referibles ni a Metatheria ni a Eutheria.
Conclusiones preliminares
En comparación con aquellos del Paleógeno, los Metatheria patagónicos de comienzos del Neógeno parecen indicar un recambio dramático en la composición taxonómica y tipos adaptativos de sus distintos linajes. Decimos "parecen" en función de dos aspectos que no deben ser soslayados: en primer lugar, existe información creciente sobre la diversidad de metaterios durante el Paleoceno y el Eoceno, no así durante el Oligoceno; esta última época es tal vez la más oscura de todo el Período Terciario en lo que respecta al conocimiento de los mamíferos de pequeño a mediano tamaño. Una mejor comprensión de las asociaciones oligocenas de metaterios podría revelar la existencia de linajes y tipos adaptativos transicionales entre las faunas paleógenas y neógenas, los cuales no son hoy discernibles a partir de la evidencia disponible. En segundo lugar, los metaterios vivientes alcanzan su mayor diversidad en las áreas tropicales de América del Sur, y es altamente probable que así haya sido también en el pasado. Su mayor diversidad se produjo entre el Máximo Térmico del Paleoceno Tardío y el Optimo Climático del Eoceno Temprano; la historia posterior de este grupo de mamíferos es la de una lenta pero sostenida declinación de linajes y tipos adaptativos. La crisis climático-ambiental del Eoceno/Oligoceno seguramente produjo eventos de extinción regional en las altas latitudes sudamericanas. En consecuencia, el registro patagónico muestra sólo una parte (la parte más austral) de su evolución, muy probablemente poco representativa de lo que debió haber ocurrido en latitudes bajas y regiones intertropicales de este continente. Como señalamos al comienzo de este trabajo, el capítulo tropical de los metaterios sudamericanos está aún por escribirse. Dicho esto, conviene repasar los datos más salientes que nos proveen las fuentes de evidencia, positiva y negativa, hasta ahora disponibles. Si bien somos cautos con respecto a la evidencia negativa, más de 800 especímenes de Metatheria colhuehuapenses sugieren que no hay, en la muestra analizada, sesgo alguno relacionado con el tamaño de los restos exhumados.
Evidencia positiva
Los Didelphimorphia y Sparassodonta de la Edad-mamífero Colhuehuapense muestran, en su composición taxonómica, una franca "modernización". Entre los primeros aparecen los Didelphoidea s.s. (Goin, 1991, 1995) a expensas de todos los linajes paleógenos. Entre los segundos sobresale la notable diversificación de Hathliacynidae y Borhyaenidae (Prothylacyninae + Borhyaeninae), así como el más antiguo registro de Thylacosmilidae. En los siguientes trabajos de esta serie se describe el clímax que alcanzan los Microbiotheria Microbiotheriidae, la consolidación de tipos adaptativos rodentiformes entre los Polydolopimorphia (Argyrolagidae, Patagoniidae) y la gran radiación de los Paucituberculata (Caenolestidae, Palaeothentidae y Abderitidae).
Evidencia negativa
Se repiten en esta edad ausencias significativas, tal como lo sugiere el aún muy incompleto registro del Oligoceno Medio-Tardío (Edad-mamífero Deseadense): desaparecen los Peradectia (Peradectidae, Caroloameghiniidae), numerosos linajes de Didelphimorphia paleógenos (Derorhynchidae, Protodidelphidae, Sternbergiidae, Pucadelphyidae, Jaskhadelphyidae, entre otros, véase Oliveira y Goin, en prensa), así como también numerosos linajes de Polydolopimorphia paleógenos (Glasbiidae, Prepidolopidae, Bonapartheriidae, Gashterniidae y Polydolopidae s. s.; sobre estos últimos, véase Goin et al., 2003). La mayor parte de estos linajes ausentes corresponde a tipos adaptativos omnívoro-frugívoros o frugívoro-insectívoros, muchos de los cuales debieron haber sido arborícolas. En el tercer trabajo de esta serie se debatirá si la aparición de roedores caviomorfos y primates platirrinos fue decisiva o no en la extinción de estos grupos.
La existencia de ciclos faunísticos definidos entre las asociaciones de mamíferos sudamericanos cenozoicos ha sido largamente argumentada (véase Pascual et al., 1996 y bibliografía citada). La evidencia fáctica determinante para estos análisis procede mayoritariamente de la Patagonia y del centro del territorio argentino, por lo que su potencial explicativo para otras regiones de América del Sur podría ser puesta en duda. Sin embargo, su valor descriptivo y predictivo para el entendimiento de la sucesión de asociaciones mamalíferas del Cono Sur sudamericano es indudable; al respecto sobresalen los aportes de Pascual et al. (1985, 1996). Los metaterios de la Edad-mamífero Colhuehuapense se enmarcan en el denominado Ciclo Faunístico Patagoniano; a su vez, este ciclo marca las etapas tempranas del Superciclo Mesocenozoico y del Megaciclo Neocenozoico (Pascual et al., 1996 y bibliografía citada). En síntesis, el Ciclo Patagoniano incluye el mayor evento de recambio mamalífero de toda la Era Cenozoica. La crisis climático-ambiental del Eoceno-Oligoceno, con la consiguiente formación de hielos permanentes en el casquete antártico, la consolidación de la Corriente Circumpolar Antártica y su consecuencia: la zonación latitudinal de los climas sudamericanos y, en tercer lugar, la fase Pehuenche del diastrofismo andino, han sido citados como factores causales de estos cambios (Pascual et al. 1996). La evolución de los metaterios patagónicos se ajusta notablemente a este contexto tectónico, climático-ambiental y faunístico. Adicionalmente, la diversificación de los roedores caviomorfos y los primates platirrinos, arribados probablemente desde Africa hacia el límite Eoceno- Oligoceno, habría de impactar significativamente en las comunidades de pequeños y medianos mamíferos sudamericanos nativos. El conjunto de metaterios colhuehuapenses sugiere el fin de su etapa "experimental", estrictamente paleógena, caracterizada por una explosiva diversidad taxonómica y una gran disparidad de tipos morfológicos, al tiempo que señala el inicio de una nueva etapa evolutiva: la estandarización neógena de relativamente pocos linajes y tipos adaptativos, etapa que habría de continuar hasta fines del Terciario.
Agradecimientos
Agradecemos a M.G. Vucetich y a C. Deschamps por toda la ayuda recibida en torno a los especímenes recolectados en el campo. Al personal del MLP, MACN, UBA y Duke (técnicos, estudiantes, investigadores y responsables del proyecto) que colectó y/o revisó los concentrados fosilíferos de los cuales proceden muchos de los especímenes estudiados. A A. Kramarz por su ayuda en la identificación de las localidades fosilíferas de la Formación Cerro Bandera y por su colaboración en los trabajos de campo. A A. Forasiepi por su colaboración con los Sparassodonta analizados. A N. Czaplewski por sus sugerencias en torno a la identificación taxonómica de los Didelphimorphia. A M. Reguero por la información brindada sobre las localidades fosilíferas colhuehuapenses. A T. Dozo, quien colectara uno de los Hathliacynidae aquí estudiados. A V. Barreda, cuya colaboración fue crucial en la interpretación paleoambiental que se ofrece en la Introducción de este trabajo. A M. Tomeo por la confección de las figuras que ilustran este trabajo. A A. Forasiepi y a un evaluador anónimo por sus valiosos comentarios sobre el manuscrito original. A los editores de la revista Ameghiniana. F. Goin agradece a la Fundación Alexander von Humboldt por el equipamiento de lupa binocular, microfotografía y software para el tratamiento de las imágenes de los especímenes aquí figurados. Parte de los trabajos de campo y laboratorio fueron financiados por la Agencia Nacional de Investigaciones Científicas y Técnicas (PICT R0074, Grupo 3), el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET; PIP 2585, 5621), y la National Science Foundation (Grants SBR 93-18942 y DEB 99-07985).
Bibliografía
1. Ameghino, F. 1887. Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de la Patagonia austral y depositados en el Museo de La Plata. Boletín del Museo de La Plata 1: 1-26. [ Links ]
2. Ameghino, F. 1891. Nuevos restos de mamíferos fósiles descubiertos por Carlos Ameghino en el Eoceno inferior de la Patagonia austral. Especies nuevas, adiciones y correcciones. Revista Argentina de Historia Natural 1: 289-328. [ Links ]
3. Ameghino, F. 1894. Enumération synoptique des espécies de mammiféres fossiles des formations éocènes de Patagonia. Boletín de la Academia de Ciencias (Córdoba) 13: 259-455. [ Links ]
4. Ameghino, F. 1897. Mammiféres crétacés de l´Argentine (Deuxième contribution à la conniasance de la faune mammalogique des couches à Pyrotherium). Boletín del Instituto Geográfico Argentino 18: 406-521. [ Links ]
5. Ameghino, F. 1900-1902. L' âge des formations sédimentaires de Patagonia. Anales de la Sociedad Científica Argentina L:109-130, 145-165, 209-229 (1900); LI: 20-39, 65-91 (1901); LII: 189-197, 244-250 (1901); LIV: 161-180, 220-249, 283-342 (1902). [ Links ]
6. Ameghino, F. 1902. Première contribution à la connaissance de la faune mammalogique des couches à Colpodon. Boletín de la Academia Nacional de Ciencias de Córdoba XVII: 71-141 (1902- 1904). [ Links ]
7. Ameghino, F. 1906. Les formations sédimentaires du Crétacé Supérieur et du Tertiaire de Patagonie avec un parallèle entre leurs faunes mammalogiques et celles de l`ancien continent. Anales del Museo Nacional de Historia Natural de Buenos Aires 8: 1-568. [ Links ]
8. Aplin, K. P. y Archer, M. 1987. Recent advances in marsupial systematics with a new syncretic classification. En: M. Archer (ed). Posums and Opossums: studies in evolution, Vol. 1, Surrey Beatty and Sons and the Royal Zoological Society of New South Wales, Sydney, pp. XV-LXXII. [ Links ]
9. Archer, M. 1984. Origins and early radiations of marsupials. En: M. Archer and G. Clayton (eds.), Vertebrate zoogeography and evolution in Australasia, Hesperian Press, Marrickville, pp. 585-631. [ Links ]
10. Ardolino, A., Franchi, M., Remesal, N. y Salani, F. 1999. El volcanismo en la Patagonia extraandina. En: R. Camino (ed.), Geología Regional Argentina, SEGEMAR, Buenos Aires: 579-612. [ Links ]
11. Argot, C. 2004. Evolution of South American mammalian predators (Borhyaenoidea): anatomical and paleobiological implications. Zoological Journal of the Linnean Society 140: 487-521. [ Links ]
12. Barreda, V.D. 1996. Bioestratigrafía de polen y esporas de la Formación Chenque, Oligoceno Tardío?-Mioceno de las provincias de Chubut y Santa Cruz, Patagonia, Argentina. Ameghiniana 33: 35-56. [ Links ]
13. Barreda, V.D. y Palamarczuk, S. 2000a. Palinoestratigrafía de depósitos del Oligoceno tardío-Mioceno, en el área sur del Golfo San Jorge, Provincia de Santa Cruz, Argentina. Ameghiniana 37: 103-117. [ Links ]
14. Barreda, V.D. y Palamarczuk, S. 2000b. Palinomorfos continentales y marinos de la Formación Monte León en su área tipo, provincia de Santa Cruz, Argentina. Ameghiniana 37: 3-12. [ Links ]
15. Barreda, V.D. y Palamarczuk, S. 2000c. Estudio palinoestratigráfico integrado del entorno Oligoceno Tardío-Mioceno en secciones de la costa patagónica y plataforma continental argentina. En: F.G. Aceñolaza y R. Herbst (eds.), El Neógeno de Argentina, Instituto Superior de Correlación Geológica , Serie Correlación Geológica 14: 103-138. [ Links ]
16. Barreda, V.D. y Bellosi, E.S. 2003. Ecosistemas terrestres eomiocenos de la Patagonia central. 12º Simposio Argentino de Paleobotánica y Palinología (Buenos Aires), Ameghiniana Suplemento Resúmenes 40: 5R. [ Links ]
17. Barrio, C., Carlini, A.A. y Goin, F.J. 1986. Litogénesis y antigüedad de la Formación Chichinales de Paso Córdoba (Río Negro, Argentina). 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 4: 149-156. [ Links ]
18. Bellosi, E.S. 1988. Régimen y velocidades tidales del mar patagoniano (Terciario medio) de la Cuenca San Jorge: estudio de periodicidades en secuencias laterales. 2° Reunión Argentina de Sedimentología (Buenos Aires), Actas 1: 16-20. [ Links ]
19. Bellosi, E.S. 1994. Hardgrounds miocenos de la Cuenca San Jorge (Argentina), un caso atípico en secuencias no carbonáticas de mares fríos. 5° Reunión Argentina de Sedimentología (Tucumán): 139-144. [ Links ]
20. Bellosi, E.S. 1995. Paleogeografía y cambios ambientales de la Patagonia Central durante el Terciario Medio. Boletín de Informaciones Petroleras (BIP) 44: 50-83. [ Links ]
21. Bellosi, E.S. 1996. La transición Paleógeno-Neógeno en cuencas centropatagónicas. Reunión de Comunicaciones Científicas Asociación Paleontológica Argentina, Ameghiniana Suplemento Resúmenes 33: 228R. [ Links ]
22. Bellosi, E.S. y Barreda, V. 1993. Secuencias y Palinología del Terciario medio en la cuenca de San Jorge, registro de oscilaciones eustáticas en Patagonia. 12° Congreso Geológico Argentino y 2º Congreso de Exploraciones de Hidrocarburos (Mendoza), Actas 1: 78-86. [ Links ]
23. Bellosi, E.S., González, R., Kay, R. y Madden, R. 2002. El valle inciso colhuehuapense de Patagonia central (Mioceno Inferior). 9° Reunión Argentina de Sedimentología (Córdoba), Resúmenes: 49. [ Links ]
24. Berggren, W.A., Kent, D.V., Swisher III, C.C. y Aubry, M.P. 1995. A revised Cenozoic geochronology and chronostratigraphy. En: W.A. Berggren, D.V. Kent, M.P. Aubry y J. Hardenbol (eds.), Geochronology, time scales and global stratigraphic correlation: a unified temporal framework for an historical geology. Society for Economic Paleontology and Mineralogy Special Publication 54: 129-212. [ Links ]
25. Bond, M. 1986. Los ungulados fósiles de Argentina: Evolución y paleoambientes. 4º Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 2: 173-185. [ Links ]
26. Bordas, A.F. 1939. Diagnosis sobre algunos mamíferos de las capas con Colpodon del Valle del Río Chubut (República Argentina). Physis 14: 413-433. [ Links ]
27. Bown, T.M. y Fleagle, G. F. 1993. Systematics, Bioestratigraphy, and dental evolution of the Palaeothentidae, Later Oligocene to Early-Middle Miocene (Deseadan-Santacrucian) Caenolestoid marsupials of South America. Journal of Paleontology 67: 1-76. [ Links ]
28. Bown, T.M., Larriestra, C.N., Powers, D.W., Naesser y Tabutt, K. 1988. New information on age, correlation, and paleoenvironments of fossil platyrrhine sites in Argentina. Journal of Vertebrate Paleontology 8: 9A. [ Links ]
29. Broom, R. 1896. On a small fossil marsupial with large grooved premolars. Proceedings of the Linnean Society of New South Wales 10: 563-567. [ Links ]
30. Burnett, G.T. 1830. Illustrations of the Quadrupeda, or quadrupeds, being the arrangement of the true footed beasts indicated in the outline. Quarterly Journal of Science and Literature 28: 351. [ Links ]
31. Cabrera, A. 1927. Datos para el conocimiento de los dasyuroideos fósiles argentinos. Revista del Museo de la Plata 30: 271-315. [ Links ]
32. Candela, A.M., Picasso, M. y Abello, M.A. 2005. Inferring locomotor behavior in the Miocene porcupine Steiromys duplicatus Ameghino (Rodentia, Erethizontidae). Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina (Puerto Madryn), Ameghiniana Suplemento Resúmenes 42: 19R. [ Links ]
33. Carlini, A.A., Madden, R., López, G., Kay, R., Vucetich, M.G., Bond, M. y Jordan, T. 1996. Nuevos mamíferos de la Formación Chinches (Mioceno) de la Cordillera Frontal de San Juan (Argentina). 12º Jornadas Argentinas de Paleontología de Vertebrados (Santa Rosa) Ameghiniana 33: 461. [ Links ]
34. Cifelli, R.L. 1984. The Casamayoran Land mammal Age in Patagonia. Ameghiniana 21: 9-10. [ Links ]
35. Cione, A.L. 1979. Aportes paleoictiológicos al conocimiento de la evolución de las paleotemperaturas en el área austral de América del Sur durante el Cenozoico. Ameghiniana 15: 183- 208. [ Links ]
36. Crompton, A.W. 1971. The origin of the tribosphenic molar. En: D. M. Kermack y K.A. Kermack (eds.), Early Mammals, Zoological Journal of the Linnean Society (supplement 1) 50: 65-70. [ Links ]
37. Feruglio, E. 1949. Descripción geológica de la Patagonia. Ministerio de Industria y Comercio de la Nación, División General Y.P.F. II: 1-349. [ Links ]
38. Fleagle, J.G. y Bown, T.M. 1983. New primate fossils from Late Oligocene (Colhuehuapian) localities of Chubut province, Argentina. Folia Primatologica 41: 240-266. [ Links ]
39. Flynn, J.J. y Wyss, A.R. 1999. New marsupials from the Eocene- Oligocene transition of the Andean Main Range, Chile. Journal of Vertebrate Paleontology 19: 533-549. [ Links ]
40. Flynn, J.J., Wyss, A.R., Charrier, R. y Swisher, C.C. 1995. An Early Miocene anthropoid skull from the Chilean Andes. Nature 373: 603-607. [ Links ]
41. Forasiepi, A.M. 1999. Plantigradismo en un Borhyaenidae (Metatheria) del Mioceno Temprano de Patagonia. 15º Jornadas de Paleontología de Vertebrados (La Plata y Luján), Ameghiniana Suplemento Resúmenes 36: 11R. [ Links ]
42. Forasiepi, A., Sánchez-Villagra, M., Goin, F.J., Kay, R., Madden, R. y Takai, M. En prensa. A new Hathliacynidae (Metatheria, Sparassodonta) from the Middle Miocene of Quebrada Honda, Bolivia. Journal of Vertebrate Paleontology. [ Links ]
43. Franchi, M. y Nullo, F. 1986. Las tobas de Sarmiento en el Macizo de Somuncurá. Revista de la Asociación Geológica Argentina 41: 219-222. [ Links ]
44. Goin, F.J. 1991. [Los Didelphoidea (Mammalia, Marsupialia, Didelphimorphia) del Cenozoico tardío de la Región Pampeana. Tesis doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. 327 pp. Inédito.] . [ Links ]
45. Goin, F.J. 1994. Living South American opossums are not living fossils. 6° International Theriological Congress (Sydney), Abstracts: 112-113. [ Links ]
46. Goin, F.J. 1995. Los Marsupiales. En: M.A. Alberdi, G. Leone y E. P. Tonni (eds.), Evolución biológica y climática de la Región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental. Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas, Madrid, pp. 165-179. [ Links ]
47. Goin, F. 2003. Early Marsupial Radiations in South America. En: M. Jones, C. Dickman y M. Archer (eds.), Predators with Pouches: The Biology of Carnivorous Marsupials, CSIRO Publishing, Colingwood, pp. 30-42. [ Links ]
48. Goin, F.J. y Candela, A.M. 2004. New Paleogene marsupials from the Amazonian basin, Southeastern Perú. En: K.E. Campbell, Jr. (ed.). The Paleogene Mammalian Fauna of Santa Rosa, Amazonian Perú. Contributions in Science, Natural History Museum of Los Angeles County, Science Series, Los Angeles, 40: 15-59. [ Links ]
49. Goin, F.J. y Rey, P. 1997. Sobre las afinidades de Monodelphis Burnett, 1830 (Mammalia, Marsupialia, Didelphidae, Marmosinae). Neotropica 43: 93-98. [ Links ]
50. Goin, F.J., Candela, A.M. y López, G. 1998. Middle Eocene Marsupials from Antofagasta de la Sierra, Northwestern Argentina. Geobios 31: 75-85. [ Links ]
51. Goin, F.J., Candela, A.M. y Muizon, C. de. 2003. The affinities of Roberthoffstetteria nationalgeographica (Marsupialia) and the origin of the Polydolopine molar pattern. Journal of Vertebrate Paleontology 23: 869-876. [ Links ]
52. Guerrero, J. 1997. Stratigraphy, sedimentary environments, and the Miocene uplift of the Colombian Andes. En: R.F Kay, R. H. Madden, R.L. Cifelli y J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene fauna of La Venta, Colombia, Smithsonian Institution Press, Washington y Londres, pp. 15-43. [ Links ]
53. Gray, J.E. 1821. On the natural arrengement of vertebrose animals. London Medical Repository, Monthley Journal and Review 15: 296-311. [ Links ]
54. Gregory, W.K. 1910. The orders of mammals. Bulletin of the American Museum of Natural History 27: 1-524. [ Links ]
55. Haq, B.U., Hardenbol, J. y Vail P.R. 1987. Chronology of fluctuating sea levels since the Triassic. Science 237: 1156-1167. [ Links ]
56. Jansa, S.A. y Voss, R.S. 2000. Phylogenetic studies on didelphid marsupials I. introduction and preliminary results from nuclear IRBP gene sequences. Journal of Mammalian Evolution 7: 43-77. [ Links ]
57. Kay, R.F., Madden, R.H., Vucetich, M.G., Carlini, A.A., Mazzoni, M.M., Re, G.H., Heizler, M. y Sandeman, H. 1999a. Revised geochronology of the Casamayoran South American Land Mammal Age: Climatic and biotic implications. Proceedings of the National Academy of Science 96: 13.235-13.240. [ Links ]
58. Kay, R.F., Madden, R.H., Mazzoni, M.M., Vucetich, M.G., Re, G. H., Heizler, M. y Sandeman, H. 1999b. Sixty-Eighth Annual Meeting of the Association of Physical Anthropologists, Abstracts, American Journal of Physical Anthropology, Suppl. 28, pp. 166. [ Links ]
59. Kirsh, J.A.W. 1977. The comparative serology of Marsupialia. Australian Journal of Zoology, Supplementary Series 52: 1-152. [ Links ]
60. Kirsch, J.A.W. y Palma, R.E. 1995. DNA/DNA hybridization studies of carnivorous marsupials. V. A further estimate of relationships among opossums (Marsupialia, Didelphidae). Mammalia 59: 403-425. [ Links ]
61. Kirsch, J.A.W., Lapointe F.J. y Springer, M. 1997. DNA-hybridisation studies of Marsupials and their Implications for Metatherian Classification. Australian Journal of Zoology 45: 211-280. [ Links ]
62. Kramarz, A.G. y Bellosi, E.S. 2005. Hystricognath rodents from the Pinturas Formation, Early-Middle Miocene of Patagonia, biostratigraphic and paleoenvironmental implications. Journal of South American Earth Sciences 18: 199-212. [ Links ]
63. Kramarz, A., Garrido, A.C., Ribeiro, A.M. y Ortiz, R. 2004. Nuevos registros de vertebrados fósiles de la Formación Chichinales, Mioceno Temprano de la Provincia de Río Negro, Argentina. 20° Jornadas Argentinas de Paleontología de Vertebrados (La Plata), Ameghiniana Suplemento Resúmenes 41: 53R. [ Links ]
64. Kramarz, A., Forasiepi, A., Tauber, A., Garrido, A., Tambussi, C. y Saldivia, S. 2001. Mamíferos fósiles de la Formación Cerro Bandera, Mioceno Temprano de la Provincia del Neuquén. 17º Jornadas Argentinas de Paleontología de Vertebrados (Esquel), Ameghiniana Suplemento Resúmenes 38: 10R. [ Links ]
65. Kramarz, A., Garrido, A. Forasiepi, A. Bond, M. y Tambussi, C. En prensa. Estratigrafía y vertebrados (Aves - Mammalia) de la Formación Cerro Bandera, Mioceno Temprano de la provincia del Neuquén, Argentina. Revista Chilena de Historia Natural. [ Links ]
66. Leche, W. 1907. Zur Entwicklungsgeschichte des Zahn-Systems der Saügetiere, zugleich ein Beitrag zur Stammesgeschichte dieser Tiergruppe. Zweite Teil: Phylogenie. Zweites Heft: Die Familien der Centetidae, Solenodontidae und Chrysochloridae. Zoologica 20: 1-158. [ Links ]
67. Legarreta, L. y Uliana, M.A. 1994. Asociaciones de fósiles y hiatos en el Supracretácico-Neógeno de Patagonia: una perspectiva estratigráfico-secuencial. Ameghiniana 31: 257-281. [ Links ]
68. Marshall, L. G.1976. New didelphine marsupials from the La Venta fauna (Miocene) of Colombia, South America. Journal of Paleontology 50: 402-418. [ Links ]
69. Marshall, L.G.1978. Evolution of the Borhyaenidae, extinct South American predaceous marsupials. University of California Publications in Geological Sciences 117: 1-89. [ Links ]
70. Marshall L.G. 1979. Review of the Prothylacyninae, an extinct subfamily of South American 'dog-like' marsupials. Fieldiana Geology, new series 3: 1-50. [ Links ]
71. Marshall, L.G. 1980. Systematics of the South American marsupial family Caenolestidae. Fieldiana Geology, new series 5: 1-145. [ Links ]
72. Marshall, L.G. 1981. Review of the Hatlyacyninae, an extinct subfamily of South American "dog-Like" marsupials. Fieldiana Geology, new series 7: 1-120. [ Links ]
73. Marshall, L.G. 1982a. Systematics of the South American marsupial family Microbiotheriidae. Fieldiana Geology, new series 10: 1-75. [ Links ]
74. Marshall, L.G. 1982b. Systematics of the extinct South American marsupial family Polydolopidae. Fieldiana Geology, new series 12: 1-109. [ Links ]
75. Marshall, L.G., Case, J. y Woodburne, M.O. 1990. Phylogenetic relationships of the families of marsupials. En: H.H. Genoways (ed.), Current Mammalogy, Plenum Press, New York, 2: 433- 505. [ Links ]
76. Marshall L.G., Hoffstetter, R. y Pascual, R. 1983. Mammals and stratigraphy: geochronology of the continental mammal-bearing Tertiary of South America. Palaeovertebrata. Mémorie Extraordinaire, Montpellier, pp. 1-93. [ Links ]
77. Marshall, L.G., Cifelli, R., Drake, R. y Curtis, G. 1986. Vertebrate paleontology, geology and geochronology of the Tapera Lopez and Scarrit Pocket, Chubut province, Argentina. Journal of Geology 60: 920-951. [ Links ]
78. Mazzoni, M.M. 1979. Contribución al conocimiento petrográfico de la Formación Sarmiento. Barranca sur del lago Colhue Huapi, provincia de Chubut. Revista de la Asociación Argentina de Mineralogía, Petrología y Sedimentología 10: 31-54. [ Links ]
79. Mazzoni, M.M. 1985. La formación Sarmiento y el vulcanismo paleógeno. Revista de la Asociación Geológica Argentina 40: 60-68. [ Links ]
80. Mendía, J. y Bayarsky, R. 1981. Estratigrafía del Terciario del valle inferior del rio Chubut. 8º Congreso Geológico Argentino (Buenos Aires), Actas 3: 593-606. [ Links ]
81. Mercerat, A. 1891. Caracteres diagnósticos de algunas especies de Creodonta conservadas en el Museo de La Plata. Revista del Museo de La Plata 2: 51-56. [ Links ]
82. Morrone, J.J. 2002. Biogeographical regions under track and cladistic scrutiny. Journal of Biogeography 29: 149-152. [ Links ]
83. Muizon, C. de. 1994. A new carnivorous marsupial from the Paleocene of Bolivia and the problem of the marsupial monophyly. Nature 370: 208-211. [ Links ]
84. Muizon, C. de. 1999. Marsupial skulls from the Deseadan (late Oligocene) of Bolivia and phylogenetic analysis of the Borhyaenoidea (Marsupialia, Mammalia). Geobios 32: 483-509. [ Links ]
85. Nánez, C. 1988. Foraminíferos y bioestratigrafìa del Terciario medio de Santa Cruz oriental. Revista de la Asociación Geológica Argentina 43: 493-517. [ Links ]
86. Olfers, I. von. 1918. Bemerkungen Zu fuger's Ueber-blid der Saugthiere, nach ihrer Bertheilung über die Welttheile rücksichtlich der Südamericanischen Ärten (Species). En: W.L. Eschwege (ed.), Neue Bibliothek der wicktigsten Reisenbeschreibungen zur Ereweiterung der Erd-und Volkerkunde. Journal von Brasilien 15: 192-237. [ Links ]
87. Oliveira, E.V. y Goin, F.J. En prensa. Marsupiais fósseis do Paleoceno de Itaboraí: Origem, Irradiação e História Biogeografica. En: N.C. Caceres y E.L.A. Monteiro Filho (eds.), Os marsupiais do Brasil: biologia, ecologia e evoluçao. Ed. Campo Grande, UNIDERP/UFMS, Campo Grande. [ Links ]
88. Ortiz Jaureguizar, E. 1986. Evolución de las comunidades de mamíferos cenozoicos sudamericanos: un estudio basado en técnicas de análisis multivariados. 4º Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 2: 191-207. [ Links ]
89. Pascual, R. y Bond, M. 1986. Evolución de los marsupiales cenozoicos de Argentina. 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 2: 143-150. [ Links ]
90. Pascual, R. y Carlini, A.A. 1985. Nuevos testimonios de la filogenia de los Argyrolagoidea (Marsupialia). 1° Jornadas Argentinas de Mastozoología (Mendoza) Resúmenes, p. 25. [ Links ]
91. Pascual, R. y Carlini, A.A. 1987. A new Superfamily in the extensive radiation of South American Paleogene marsupials. Fieldiana Zoology, new series 39: 99-110. [ Links ]
92. Pascual, R. y Odreman Rivas, O. 1971. Evolución de las comunidades de los vertebrados del Terciario argentino. Los aspectos paleozoogeográficos y paleoclimáticos relacionados. Ameghiniana 8: 372-412. [ Links ]
93. Pascual, R. y Odreman Rivas, O. 1973. Las unidades estratigráficas del Terciario portadoras de mamíferos. Su distribución y sus relaciones con los acontecimientos diastróficos. 5° Congreso Geológico Argentino (Córdoba) Actas 3: 293-338. [ Links ]
94. Pascual, R. y Ortiz Jaureguizar, E. 1990. Evolving climates and mammal faunas in Cenozoic South America. Journal of Human Evolution 19: 23-60. [ Links ]
95. Pascual, R., Ortiz Jaureguizar, E. y Prado, J.L. 1996. Land mammals: Paradigm for Cenozoic South American geobiotic evolution. Münchner Geowissenschaften Abhadlungen 30: 265-319. [ Links ]
96. Pascual, R., Vucetich, M.G., Scillato Yané, G.J. y Bond, M. 1985. Main pathways of mammalian diversification in South America. En: F. Stehli y S.D. Webb (eds.), The Great American Biotic Interchange, Plenum Press, New York and London, pp. 219-247. [ Links ]
97. Patterson, B. 1958. Affinities of the Patagonian fossil mammal Necrolestes. Breviora 94: 1-14. [ Links ]
98. Patton, J.L., dos Reis, S.F. y da Silva, M.N.F. 1996. Relationships among didelphid marsupials based on sequence variation in the mitochondrial cytocrome b gene. Journal of Mammalian Evolution 3: 3-29. [ Links ]
99. Paula Couto, C. de. 1952. Fossils Mammals from the Beginning of the Cenozoic in Brazil. Marsupialia: Didelphidae. American Museum Novitates 1567: 1-26. [ Links ]
100. Paula Couto, C. de. 1962. Didelfídeos fósiles del Paleoceno de Brazil. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 112: 135-166. [ Links ]
101. Reig, O.A. 1952. Descripción previa de nuevos ungulados y marsupiales fósiles del Plioceno y del Eocuartario argentinos. Revista del Museo Municipal de Ciencias Naturales y Tradicional de Mar del Plata 1: 119-129. [ Links ]
102. Reig, O.A. 1955. Un nuevo género y especie de cenolestinos del Plioceno de la Provincia de Buenos Aires (República Argentina). Revista de la Asociación Geológica Argentina 10: 60- 71. [ Links ]
103. Reig, O.A., Kirsh, J.A.W. y Marshall, L.G. 1987. Systematic and relationships of the living and neocenozoic american "opossumlike" marsupials (Suborder Didelphimorphia), with comments on the classification of these and of the Cretaceous and Paleogene new world and european metatherians. En: M. Archer (ed.), Possums and Opossums: studies in evolution, Vol. 1, Surrey Beatty and Sons and the Royal Zoological Society of New South Wales, Sydney, pp. 1-89. [ Links ]
104. Riggs, E.S. 1933. Preliminary description of a new marsupial saber- tooth from the Pliocene of Argentina. Field Museum of Natural History, Geological Series 6: 61-66. [ Links ]
105. Ringuelet, A.B. de. 1953. Revisión de los didélfidos fósiles argentinos. Revista del Museo y Universidad Eva Perón (nueva serie), Paleontología 3: 265-308. [ Links ]
106. Romer, A.S. 1945. Vertebrate Paleontology, University of Chicago Press, Chicago, 687 pp. [ Links ]
107. Roth, S. 1899. Aviso preliminar sobre mamíferos mesozoicos encontrados en Patagonia. Revista del Museo de La Plata 9: 381- 388. [ Links ]
108. Roth, S. 1903. Noticias sobre nuevos mamíferos fósiles del Cretáceo Superior y Terciario de la Patagonia. Revista del Museo de La Plata 11: 133-158. [ Links ]
109. Rougier, G.W., Wible, J.R. y Novacek, M.J. 1998. Implications of Deltatheridium specimens for early marsupial history. Nature 396: 459-463. [ Links ]
110. Rusconi, C. 1933. Nuevos restos de monos fósiles del Terciario an- tiguo de Patagonia. Anales de la Sociedad Científica Argentina 116: 286-289. [ Links ]
111. Saban, R. 1954. Phylogénie des insectivores. Bulletin du Muséum National d`Histoire Naturelle 26: 419-432. [ Links ]
112. Sánchez-Villagra, M. R. y Clack J. A. 2004. Fossils of the Castillo Formation, Venezuela: Contributions on Neotropical Paleontology. Special Papers in Paleontology 71: 1-112. [ Links ]
113. Sánchez-Villagra, M.R. y Smith K. 1997. Diversity and Evolution of the Marsupial Mandibular Angular Process. Journal of Mammalian Evolution 4: 119-144. [ Links ]
114. Sanborn, C.C. 1951. Two new mammals from southern Peru. Fieldiana, zoological series 31: 473-477. [ Links ]
115. Scasso, R. y Bellosi, E. 2004. Cenozoic continental and marine trace fossils at the Bryn Gwyn Paleontological Park, Chubut. Bryn Gwyn Guidebook, 1º International Congress on Ichnology (Trelew), Resúmenes, pp. 18. [ Links ]
116. Scillato Yané, G.J. 1977. Sur quelques Gliptodontidae nouveaux (Mammalia, Edentata) du Déséadien (Oligocène inférieur) de Patagonie (Argentine). Bulletin du Muséum National d'Histoire Naturelle, 3° séries, 487: 249-262. [ Links ]
117. Scillato Yané, G.J. 1986. Los Xenarthra fósiles de Argentina (Mammalia, Edentata) 4º Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 2: 151-155. [ Links ]
118. Scott, W.B. 1905. Palaeontology. Part II. Insectivora. Reports of the Princeton University Expeditions to Patagonia 5: 365-383. [ Links ]
119. Simpson, G.G. 1932. Some new or little-known mammals from the Colpodon beds of Patagonia. American Museum Novitates 575: 1- 12. [ Links ]
120. Simpson, G.G. 1935. Early and Middle Tertiary geology of the Gaiman region, Chubut, Argentina. American Museum Novitates 775: 1-29. [ Links ]
121. Simpson, G.G. 1945. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85: 1-350. [ Links ]
122. Simpson, G.G. 1948. The beginning of the age of mammals in South America. Part. 1. Bulletin of the American Museum of Natural History 91: 1-232. [ Links ]
123. Simpson, G.G. 1967. The beginning of the age of mammals in South America. Part. 2. Bulletin of the American Museum of Natural History 137: 1-259. [ Links ]
124. Sinclair, W.J. 1930. New carnivorous Marsupialia from the Deseado Formation of Patagonia. Field Museum of Natural History, Geological Memories 1: 35-39. [ Links ]
125. Spalletti, L.A. y Mazzoni, M.M. 1977. Sedimentología del Grupo Sarmiento en un perfil ubicado al sudeste del Lago Colhue Huapi, provincia de Chubut. Revista del Museo de La Plata, Obras del Centenario del Museo, Geología 4: 261-283. [ Links ]
126. Spalletti, L.A. y Mazzoni, M.M. 1979. Estratigrafía de la Formación Sarmiento en la barranca sur del lago Colhue Huapi, provincia de Chubut. Revista de la Asociación Geológica Argentina 34: 271-281. [ Links ]
127. Stirton, R.A. 1953. Vertebrate paleontology and continental stratigraphy in Colombia. Geological Society of America Bulletin 64: 603-622. [ Links ]
128. Szalay, F.S. 1994. Evolutionary history of the marsupials and an analysis of osteological characters. Cambridge University Press, Cambridge, 480 pp. [ Links ]
129. Thomas, O. 1894. On Micoureus griseus, Desm. With the description of a new genus and species of Didelphyidae. Annals and Magazine of Natural History 14: 184-188. [ Links ]
130. Thomas, O. 1912. A new genus of opossum and a new tuco-tuco. Annals and Magazine of Natural History 9: 239-241. [ Links ]
131. Trouessart, E.L. 1898. Catalogus Mammalian tam viventium quam fossilium. Ed. New Berlin, pp. 665-1264. [ Links ]
132. Turner, V. y McKay, G. M. 1989. Burramyidae. En: D.W. Walton y B.J. Richardson (eds.), Fauna of Australia, Volume 1B: Mammalia. Fauna of Australia Series, Australian Government Publishing Service, Canberra, pp. 1-27. [ Links ]
133. Voss, R.S. y Jansa, S.A. 2003. Phylogenetic studies on didelphid marsupials II. Nonmolecular data and new IRBP sequences: separate and combined analyses of didelphine relationships with denser taxon sampling. Bulletin of the American Museum of Natural History 276: 1-82. [ Links ]
134. Voss, R.S., Tarifa T. y Yensen E. 2004. An introduction to Marmosops (Marsupialia: Didelphidae), with the description of a new species from Bolivia and notes on the taxonomy and distribution of other Bolivian Forms. American Museum Novitates 3466: 1-40. [ Links ]
135. Vucetich, M.G. 1986. Historia de los roedores y primates en Argentina: su aporte al conocimiento de los cambios ambientales durante el Cenozoico. 4° Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza), Actas 2: 157-165. [ Links ]
136. Wall, C.E. y Krause D.W. 1992. A biomechanical analysis of the masticatory apparatus of Ptilodus (Multituberculata). Journal of Vertebrate Paleontology 12: 172-187. [ Links ]
137. Winge, H. 1941. The interrelationships of the mammalian genera. Volume 1. Monotremata, Marsupialia, Insectivora, Chiroptera, Edentata. København; C.A. Reitzel, 1-12, pp 1-412. [ Links ]
138. Zucol, A., Brea M., Madden R., Bellosi E., Carlini A. A. y Vucetich M.G. En prensa. Preliminary phytolith analysis of Sarmiento Formation in the Gran Barranca (Central Patagonia, Argentina). Fourth International Meeting of Phytolith Research. The McDonald Institute for Archaeological Research, University of Cambridge. Balkema. [ Links ]
Recibido: 9 de diciembre de 2004.
Aceptado: 1 de junio de 2006.

