










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.1 Buenos Aires ene./mar. 2007
Análisis de biofacies y cambios relativos del nivel del mar en el Miembro Pilmatué de la Formación Agrio, Cretácico Inferior de cuenca Neuquina, Argentina
Darío G. Lazo1
1Consejo Nacional de Investigaciones Científicas y Técnicas, Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Ciudad Universitaria, 1428 Buenos Aires, Argentina dlazo@gl.fcen.uba.ar
Abstract. Biofacies analysis of the Pilmatué Member of the Agrio Formation, Lower Cretaceous, Neuquén basin, Argentina. A biofacies analysis based on benthic macrofossils was carried out. The study unit is the Pilmatué Member of the Agrio Formation in three sections in the Neuquén Province. Six biofacies controlled by relative sea level changes, sedimentation rate, substrate consistency, oxygenation and salinity were recognised. The "Cucullaea-Mimachlamys-Steinmanella biofacies" is highly diverse and recorded in a shoreface environment characterized by a well-oxygenated soft sandy and patchy bioclastic substrate, reduced siliciclastic input, normal marine salinity, and warm-temperate waters. The "coral colonies biofacies" is associated with the former biofacies, but in a more shallow setting and probably temperatures above 16º-18º C. The "Cucullaea and Steinmanella biofacies" occurred in an open shelf environment characterized by low water energy, reduced siliciclastic input, firm muddy substrate, and normal marine conditions. The "Modiolus biofacies" also occurs on the shelf, but under low oxygenation. The "gregarious serpulids" and "gregarious oysters" biofacies correspond to an inner shelf setting characterized by an alternation of high and low water energy, low siliciclastic input, firm muddy substrate, well-oxygenated bottom, and mixo-euhaline conditions. The "Cucullaea-Mimachlamys-Panopea" and "Cucullaea and Steinmanella" biofacies are related to changes of relative sea level in the scale of parasequences, while the "coral colonies", "Modiolus", "gregarious oysters", and "gregarious serpulids" respond to changes in the scale of sequences.
Resumen. Sobre la base de estudios tafonómicos y paleoambientales realizados previamente en el Miembro Pilmatué de la Formación Agrio se analizaron las biofacies macrofaunísticas bentónicas registradas en tres localidades del centro-norte de Neuquén. Se reconocieron seis biofacies influidas por cambios relativos del nivel del mar, tasa de sedimentación, consistencia del sustrato, oxigenación y salinidad. La biofacies de Cucullaea-Mimachlamys-Panopea caracteriza un shoreface de moderada a alta energía, baja tasa de sedimentación, sustrato arenoso blando con parches bioclásticos, buena oxigenación, salinidad marina normal y aguas templado-cálidas. La biofacies de corales coloniales aparece asociada a la anterior, pero en una zona más proximal de la cuenca y a una temperatura superior a 16º-18º C. La biofacies de Cucullaea y Steinmanella se registra en la plataforma interna y media-externa en un ambiente de baja energía, baja tasa de sedimentación, fangos firmes y salinidad marina normal. La biofacies de Modiolus se registra también en la plataforma, pero a diferencia de la anterior llegaba al fondo durante períodos de baja oxigenación. Las biofacies de ostras y serpúlidos gregarios se registran en la plataforma interna. Habitaban un ambiente de energía alternante entre alta y baja, sustrato fangoso firme, baja tasa de sedimentación, oxigenación normal y aguas de salinidad menor a la marina normal. Las biofacies de Cucullaea- Mimachlamys-Panopea y la biofacies de Cucullaea y Steinmanella responden a cambios relativos del orden de parasecuencias, mientras que las biofacies de corales coloniales, Modiolus y ostras y serpúlidos gregarios responden a cambios del orden de secuencias.
Key words. Benthic macrofossils; Paleoecology; Paleoenvironments; Valanginian-Hauterivian; Argentina.
Palabras clave. Macrofauna bentónica; Paleoecología; Paleoambientes; Valanginiano-Hauteriviano; Argentina.
Introducción
En la literatura el término biofacies ha sido utilizado con diferentes significados de los cuales el ambiental o ecológico es el más importante (Ludvigsen et al., 1986). Esta connotación del término cobró fuerza en los estudios de faunas bentónicas actuales los cuales demostraron que distintos sectores de las plataformas están caracterizados por asociaciones o biofacies particulares. Estas biofacies generalmente se van reemplazando con el aumento de la profundidad respondiendo a cambios físicoquímicos e interacciones biológicas y en el registro pueden aportar datos útiles en interpretaciones paleoambientales (Scott, 1970; Brenchley, 1990; Wignall, 1990).
Cabe mencionar que antes de realizar un análisis de biofacies en el registro fósil es aconsejable contar previamente con un estudio litofacial de la unidad bajo estudio y un análisis tafonómico de los fósiles utilizados. Las litofacies permiten interpretar los paleoambientes de sedimentación mientras que las tafofacies en combinación con datos bioestratigráficos permiten estimar los grados de mezcla temporal (time- averaging) y espacial de los fósiles. De esta manera es posible detectar asociaciones fósiles autóctonas o parautóctonas con un cierto time-averaging con posibilidades de ser evaluadas paleoecológicamente.
En nuestro país se han realizado estudios combinados de litofacies y biofacies que demostraron ser muy útiles en el refinamiento de interpretaciones paleoambientales y paleoecológicas (e.g., Scasso et al., 1991; Carrera, 2001; Damborenea y Manceñido, 2005). En particular, la rica fauna de invertebrados del Cretácico Inferior de la cuenca Neuquina representa una oportunidad ideal para realizar estudios de este tipo. Sin embargo, hay una escasez notable de análisis paleoecológicos (véase Damborenea et al., 1979). Esta escasez puede ser explicada en parte por la falta de datos sistemáticos y paleoautoecológicos. Afortunadamente el estudio sistemático de los amonites ha tenido un gran desarrollo en los últimos años y esto ha permitido establecer biozonaciones entre el Berriasiano y el Barremiano de fácil correlación regional en la cuenca (Aguirre-Urreta y Rawson, 1997, 2003). A su vez las biozonaciones permitieron correlacionar y acotar el grado de time-averaging de la fauna bentónica. También se ha avanzado en el conocimiento sistemático y paleoecológico de bivalvos, nautílidos y equinoideos de la Formación Agrio lo cual permitió analizar con más detalle a la fauna en su conjunto (Rodríguez, 2001; Cichowolski, 2003; Lazo, 2003a, 2003b, 2004).
Teniendo en cuenta que ya se han hecho estudios litofaciales, tafofaciales, sistemáticos y paleoautoecológicos de la macrofauna bentónica del Miembro Pilmatué de la Formación Agrio (Lazo, 2004, 2006), es posible avanzar en el conocimiento y plantear para este trabajo los siguientes objetivos: (1) descripción e interpretación de las biofacies macrobentónicas del Miembro Pilmatué; (2) inferencia de las implicancias paleoecológicas y paleoambientales de dichas biofacies; y (3) discusión de la distribución de las biofacies en un esquema estratigráfico secuencial y su posible dependencia con los cambios relativos del nivel del mar.
Marco geológico y metodología
La zona de estudio se ubica en el centro-norte de Neuquén entre las localidades de Zapala y Chos Malal. La unidad analizada es el Miembro Pilmatué de la Formación Agrio el cual fue depositado por detrás de un arco de volcánico en conexión con el océano Pacífico durante el intervalo Valanginiano Tardío- Hauteriviano Temprano. Las aguas oceánicas penetraban desde el oeste e inundaban la depresión del retroarco orientada en sentido noroeste-sudeste, aunque la presencia de un arco volcánico insular habría causado cierto aislamiento temporal de la cuenca.
Las secciones son de norte a sur las siguientes: Agua de la Mula (38°04' S; 70°01' W), Salado Sur (38°12' S; 70°03' W) y Bajada del Agrio (38°27' S; 70°04' W) (figura 1). Agua de la Mula se ubica a 90 km al sur de Chos Malal en el sector noroeste de la cordillera del Salado (cerro Mula). La sección Salado Sur está ubicada a 16,3 km al sur de Agua de la Mula en el sector sudoeste de la misma cordillera (cerro Salado). La sección Bajada del Agrio está ubicada a 27,9 km al sur de Salado Sur en un paraje denominado Bajada Vieja sobre la margen derecha del río Agrio. Las tres secciones están alineadas aproximadamente en dirección norte-sur y por lo tanto se encuentran a distancias similares con respecto al margen este de la cuenca. Sin embargo, el perfil Bajada del Agrio se encuentra más próximo al margen sur de la cuenca, siendo muy probablemente la sección más proximal de las tres analizadas.

Figura 1. Esquema geológico del centro-norte de Neuquén mostrando la ubicación de las tres secciones analizadas: A, Agua de la Mula; B, Salado Sur; C, Bajada del Agrio. Nótese que los perfiles abarcan 44,2 km de distancia en línea recta N-S, Mapa geológico tomado de Leanza et al. (2001) / geological sketch of central-north Neuquén showing the position of the study sections: A, Agua de la Mula; B, Salado Sur; C, Bajada del Agrio. Note that the three sections are in a N-S line of 44.2 km. Geological map from Leanza et al. (2001).
Previamente al análisis de biofacies el autor ha realizado análisis sedimentológicos y tafonómicos completos en el miembro Pilmatué en los mismos perfiles (véase Lazo, 2004, 2006). Dichos análisis indican que: (1) el Miembro Pilmatué fue depositado en un ambiente de shoreface-plataforma influido por tormentas, sin quiebres de pendiente hacia el offshore y ubicado dentro de la zona fótica; (2) el grado de transporte lateral de la macrofauna bentónica fue nulo o bajo en dirección al offshore y (3) el grado de time- averaging de la macrofauna aumenta en dirección al offshore hasta un máximo de 0,5 Ma (Lazo, 2006). De las nueve tafofacies planteadas para el Miembro Pilmatué, sólo siete tienen valor paleoecológico dado que resultan ser autóctonas o parautóctonas (cuadro 1). Las tafofacies P2 y Dc pueden tener un grado muy variable de transporte lateral por lo cual quedan descartadas del análisis paleoecológico. Por otro lado, a excepción de la tafofacies Vi, las tafofacies presentan un cierto time-averaging. Esto implica que las asociaciones preservadas pueden representar una superposición de generaciones, poblaciones e incluso comunidades de hasta 0,5 Ma (Kidwell, 1998).
Cuadro 1. Grado de transporte lateral, time-averaging y potencial paleoecológico de las tafofacies del Miembro Pilmatué de la Formación Agrio y su relación con las biofacies analizadas. Tafofacies tomadas de Lazo (2006) / degree of lateral transport, time-averaging, and paleoecologic value of the taphofacies of the Pilmatué Member of the Agrio Formation and its relation to the study biofacies. Taphofacies from Lazo (2006).

Las biofacies analizadas en este trabajo se basan en corales, serp úlidos, bivalvos y gastrópodos los cuales conforman la mayor parte de la macrofauna bentónica del Miembro Pilmatué. Las biofacies se definieron mediante observaciones cualitativas de campo sobre la base de análisis sedimentológicos y tafonómicos previos que permitieron evaluar procesos depositacionales y bioestratinómicos (figura 2). Estos estudios se complementaron con un análisis detallado de la sistemática y la paleoautoecología de gastrópodos, bivalvos, serpúlidos y corales (Lazo, 2004). De esta forma se pudieron distinguir a grandes rasgos seis biofacies principales, aunque es muy probable que cuantificando la abundancia relativa en cada una de ellas se las pueda subdividir más finamente y en consecuencia refinar la interpretación paleoambiental y paleoecológica. En el caso de la biofacies diversa de shoreface además de su identificación cualitativa se ha realizado una serie de conteos de bivalvos en Agua de la Mula tendientes a cuantificar la abundancia relativa e identificar a las especies más abundantes (véase apéndice; Lazo, 2006).

Figura 2. Diagrama de flujo que muestra la lógica utilizada en este trabajo hasta arribar a la reconstrucción de biofacies con significado paleoambiental y paleoecológico. Las litofacies y tafofacies se basaron en Lazo (2006); los datos taxonómicos y paleoauotecológicos se tomaron de Lazo (2004) / scheme of the strategy followed in this paper. Lithofacies and taphofacies from Lazo (2006); taxonomy and paleoautoecology from Lazo (2004).
Con el objetivo de analizar la relaci ón de las biofacies con los cambios relativos del nivel del mar se analizaron secuencialmente los perfiles sobre la base de un modelo de plataforma sin quiebre de talud que incluye los siguientes cortejos: cuña de margen de plataforma (SMW, shelf margin wedge), cortejo transgresivo (TST, transgressive system tract) y cortejo de mar alto (HST, highstand system tract) (figura 3). El marco bioestratigráfico está basado en la zonación detallada de amonites propuesta por Aguirre-Urreta y Rawson (1997, 2003). La profundidad de los niveles de base de ola de buen tiempo y de tormenta son promedios basados en Howard y Reineck (1981). Estos datos sirven para dar una idea de la profundidad a la cual se desarrollaban cada una de las biofacies.

Figura 3. Perfiles litológicos del Miembro Pilmatué de la Formación Agrio mostrando los límites de secuencia, la zonación de amonites y la distribución de biofacies / lithological columns of the Pilmatué Member of the Agrio Formation showing sequence boundaries, ammonite zonation, and the distribution of biofacies.
Resultados
Biofacies de Cucullaea-Mimachlamys-Panopea (figura 4)

Figura 4. Reconstrucción de la biofacies de Cucullaea-Mimachlamys-Panopea en un ambiente de shoreface en Agua de la Mula, Hauteriviano Temprano / reconstruction of the "Cucullaea-Mimachlamys-Panopea biofacies" in a shoreface environment in Agua de la Mula, Early Hauterivian. A, Holcoptychites agrioensis (Weaver, 1931); B, Myoconcha transatlantica Burckhardt, 1903; C, Mimachlamys robinaldina (d'Orbigny, 1847); D, Trigonia carinata Agassiz, 1840; E, Panopea gurgitis (Brongniart, 1822); F, Ptychomya koeneni Behrendsen 1892; G, Eriphyla argentina Burckhardt, 1903; H, Gervillella aviculoides (J. Sowerby, 1814); I, Pinna (P.) robinaldina d'Orbigny, 1844; J, Cucullaea (Noramya) gabrielis Leymerie, 1842; K, Pterotrigonia sp.
Descripción. Esta biofacies se registra en las litofacies Sm/Sg, Shm y Cg/Cm. La litofacies Sm/Sg corresponde a areniscas muy finas, finas y medianas macizas o con gradación normal seguidas de ondulitas de oleaje y corriente. La litofacies Shm son areniscas finas y medianas macizas o con gradación normal, estructuras de escape de agua, laminación paralela, estratificación entrecruzada hummocky y ondulitas de oleaje. La litofacies Cg/Cm corresponde a depósitos bioclásticos con empaquetamiento denso y pobre, matriz fangosa a arenosa, base erosiva, gradación normal, canalizaciones con laminación paralela y tope con ondulitas de oleaje y corriente. Las tres litofacies poseen abundantes trazas pertenecientes a la icnofacies de Cruziana y fueron interpretadas como tempestitas proximales amalgamadas de shoreface (Lazo, 2006).
La biofacies de Cucullaea-Mimachlamys-Panopea presenta una alta riqueza ya que involucra a la mayor parte de la fauna de bivalvos del Miembro Pilmatué (véase Lazo, 2004). Además, es la más importante dado que se registra abundantemente a lo largo de todo el Miembro Pilmatué en los tres perfiles analizados (figura 3). Los bivalvos están representados por 16 especies que conforman el núcleo principal (apéndice), aunque también aparecen serpúlidos coloniales, gastrópodos epifaunales e infaunales, amonites y nautílidos. La mayoría de los bivalvos corresponde a formas cavadoras (48,4% someras y 12,9% profundas) mientras que el resto corresponde a epibisados (21,3%), semiinfaunales bisados (13,2%) y finalmente reclinantes (4,2%). Todos son suspensívoros. Las tres especies más abundantes y que superan un 50 % en abundancia son Cucullaea (Noramya) gabrielis Leymerie, 1842, Mimachlamys robinaldina (d'Orbigny, 1847) y Panopea gurgitis (Brongniart, 1822).
Interpretación. Los organismos de esta biofacies habitaban sustratos arenosos a bioclásticos de consistencia principalmente blanda dentro del shoreface. Este ambiente abarca desde la línea de baja marea hasta el nivel de base de ola normal, aproximadamente a 5 m de profundidad. Por lo tanto los organismos estaban sometidos al oleaje de buen tiempo y a tormentas cuyos flujos de relajación tendían a transportarlos pendiente abajo hacia la plataforma media-externa. Dado el predominio de formas suspensívoras la tasa de sedimentación debió ser relativamente baja durante el crecimiento de los organismos. Debido a la energía moderada a alta típica de este ambiente los nutrientes se encontraban principalmente en suspensión lo que concuerda con un predominio de formas suspensívoras. Generalmente los bivalvos, incluso los infaunales profundos, aparecen fuera de su posición de vida y por lo tanto puede inferirse que existía erosión profunda por corrientes de fondo. Hay un buen aprovechamiento del espacio ya que hay especies epifaunales, semiinfaunales, infaunales someras y profundas. Las trazas fósiles asociadas también poseen un buen escalonamiento. Estos datos indican que los niveles de oxígeno sobre el fondo eran suficientes como para mantener la parte superior del sedimento oxigenada. La zona de anaerobiosis estaba al menos por debajo de los 30 cm de profundidad ya que hasta allí llegaban los cavadores profundos. La riqueza alta confirma la buena disponibilidad de oxígeno y a su vez apunta a un ambiente de aguas de salinidad marina normal. Esto se confirma con la presencia de grupos euhalinos como los trigonioideos, amonites y nautílidos (cuadro 2).
Cuadro 2. Parámetros ambientales inferidos a partir de las distintas biofacies del Miembro Pilmatué de la Formación Agrio con indicación de la posición de cada biofacies en el esquema secuencial propuesto. Véase abreviaturas en el texto / Inferred environmental parameters based on biofacies and its location within sequences, Pilmatué Member of the Agrio Formation. For abbreviations see text.
Hay especies que aparecen exclusivamente en esta biofacies, por ejemplo Mimachlamys robinaldina (d'Orbigny, 1847) y Gervillella aviculoides (J. Sowerby, 1814), lo que implica que sus larvas lograban fijarse y hacer su metamorfosis sólo sobre fondos arenosos o bioclásticos del shoreface. Sin embargo, en esta biofacies hay también especies generalistas de sustrato, por ejemplo Cucullaea (N.) gabrielis y Pinna (P.) robinaldina d'Orbigny, 1844. Dichas especies aparecen asociadas tanto a fondos arenosos del shoreface como a fangos de plataforma. La presencia de bivalvos de valvas muy espesas o muy ornamentadas, como Gervillaria alatior (Imlay, 1940) o Myoconcha transatlantica Burckhardt, 1903 indicarían aguas templadas a cálidas (cuadro 2).
Biofacies de Cucullaea y Steinmanella (figura 5)
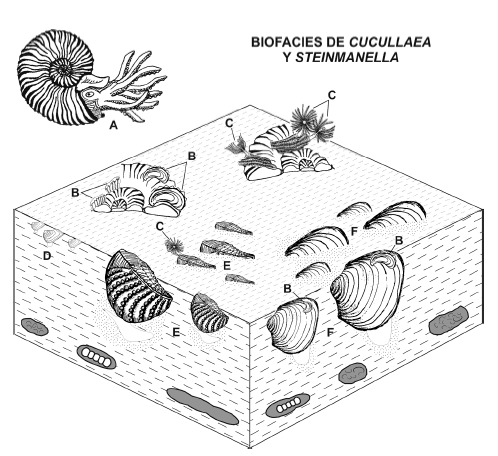
Figura 5. Reconstrucción de la biofacies de Cucullaea y Steinmanella en Bajada del Agrio, Valanginiano Tardío. Nótese la asociación con amonites caídos y concreciones carbonáticas / reconstruction of the "Cucullaea and Steinmanella biofacies" in Bajada del Agrio, Late Valanginian. Note association with sunken ammonites and carbonate concretions. A, Pseudofavrella angulatiformis (Behrendsen, 1892); B, Amphidonte (Ceratostreon) sp.; C, Parsimonia antiquata (J. de C. Sowerby, 1829); D, Disparilia sp.; E, Steinmanella pehuenmapuensis Leanza, 1998; F, Cucullaea (Noramya) gabrielis Leymerie, 1842.
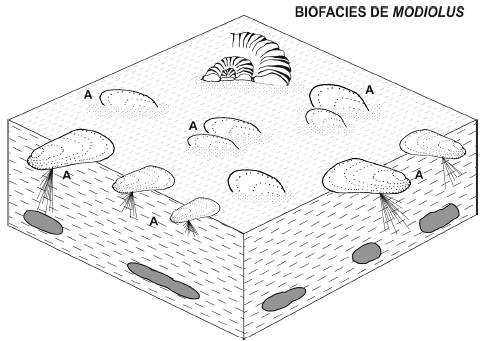
Figura 6. Reconstrucción de la biofacies de Modiolus es sustratos fangosos de plataforma en Bajada del Agrio, Subzona de Holcoptychites neuquensis, Hauteriviano Temprano. Nótese la asociación con amonites "llovidos" / reconstruction of the "Modiolus biofacies" in shelf muddy substrates in Bajada del Agrio, Early Hauterivian. Note association with sunken ammonites. A, Modiolus cf. subsimplex (d'Orbigny, 1850).
Descripción. Esta biofacies se registra en los tres perfiles analizados en las litofacies Fl y Fm las cuales fueron interpretadas como depósitos fangosos de plataforma de consistencia soposa a blanda (figura 3). La biofacies de Cucullaea y Steinmanella aparece junto a excavaciones con límites bien definidos y relleno contrastante con las pelitas y marcas de raspaduras las cuales fueron asignadas a cf. Thalassinoides e interpretadas como trazas de la icnofacies de Glossifungites. Cucullaea (N.) gabrielis, Steinmanella pehuenmapuensis Leanza, 1998 y S. transitoria (Steinmann, 1881) son las especies principales en esta biofacies. Se trata de formas cavadoras y suspensívoras no sifonadas que se ubicaban con el margen posterior en contacto con la interfase agua-sedimento. Presentan valvas relativamente gruesas y en el caso de Steinmanella una ornamentación prominente. En forma minoritaria aparecen Pinna (P.) robinaldina, Sphaera koeneni (Behrendsen, 1892), Disparilia sp., Ptychomya koeneni Behrendsen, 1892, pequeños venéridos indeterminados y amonites. Es muy común encontrar las conchillas de bivalvos y amonites incrustadas por Amphidonte (Ceratostreon) sp. y Parsimonia antiquata (J. de C. Sowerby, 1829) (véase tafofacies P1 en Lazo, 2006).
Interpretación. Esta biofacies se desarrolló en la plataforma. Este ambiente se ubica desde el nivel de base de ola de buen tiempo, aproximadamente a 5 m de profundidad, hacia mar adentro. Estos organismos colonizaban el fondo fangoso de baja energía. El predominio de suspensívoros de conchilla gruesa en un sustrato fangoso indica que la tasa de sedimentación era baja y que el sustrato tenía una consistencia relativamente firme (Rhoads y Young, 1970). La incrustación también apunta a una baja tasa de sedimentación mientras que la asociación de trazas indica la presencia de sustratos firmes (véase Lazo, 2006). El aprovechamiento del espacio es bajo ya que hay un claro predominio de cavadores someros (apéndice). Esto puede deberse a que la zona de anaerobiosis estaba cercana a la interfase agua-sedimento. La presencia ocasional de S. koeneni también indica que la zona anaeróbica estaba cercana a esa interfase (Taylor y Glover, 2000). Los bivalvos que componen esta biofacies son generalistas de sustrato ya que aparecen también en la biofacies de Cucullaea-Mimachlamys- Panopea. Por ello se podría pensar en iguales condiciones de salinidad y temperatura que la biofacies anterior (cuadro 2).
Descripción. Al igual que en el caso anterior esta biofacies se registra las litofacies Fl y Fm interpretadas como depósitos fangosos de plataforma, aunque sólo dentro de la subzona de Holcoptychites neuquensis (figura 3). Es monoespecífica ya que está representada sólo por la especie Modiolus cf. subsimplex (d'Orbigny, 1850). Esta forma es de hábito semiinfaunal bisado y suspensívoro. El registro incluye restos de valvas muy delgadas y moldes internos de individuos pequeños (largo máximo = 63 mm). Los moldes presentan frecuentemente trazas criptococleares (Damborenea y Manceñido, 1996).
Interpretación. Esta biofacies se desarrolló en la plataforma en un ambiente de baja energía y sobre un sustrato fangoso. El sustrato debió tener una consistencia relativamente firme ya que estaba colonizado por semiinfaunales bisados. La presencia de trazas en los moldes internos sugiere una baja tasa de sedimentación. Hay un bajo aprovechamiento del espacio ya que hay sólo semiinfauna (cuadro 2). La monoespecificidad, el pequeño tamaño y la ausencia de incrustantes sugieren un ambiente estresado probablemente por un descenso en el nivel de oxígeno del fondo (véase Wignall, 1993).
Biofacies de corales coloniales (figura 7)

Figura 7. Reconstrucción de la biofacies de corales coloniales en Bajada del Agrio, Hauteriviano Temprano / reconstruction of the "colonial corals biofacies" in Bajada del Agrio, Early Hauterivian. A, Holcoptychites agrioensis (Weaver, 1931); B, Columastrea antiqua (Gerth, 1928); C, Placocoenia neuquensis Gerth, 1928; D, Myoconcha transatlantica Burckhardt, 1903; E, Amphidonte (Ceratostreon) sp.; F, Isognomon (I.) lotenoensis (Weaver, 1931); G, Gervillaria alatior (Imlay, 1940); H, Cucullaea (Noramya) gabrielis Leymerie, 1842; I, Steinmanella transitoria (Steinmann, 1881); J, Pterotrigonia sp.; K, Eriphyla argentina Burckhardt, 1903; L, Ptychomya koeneni Behrendsen 1892; M, Gastrochaenolites.
Descripción. Los rasgos preservacionales de esta biofacies corresponden a la tafofacies R3 de Lazo (2006). Se registra en la litofacies Cg/Cm la cual corresponde a depósitos bioclásticos con empaquetamientos denso y pobre, base erosiva, gradación normal y tope con ondulitas. Estos depósitos fueron interpretados como tempestitas proximales amalgamadas de shoreface. La distribución vertical de esta biofacies es acotada ya que aparece en un solo nivel dentro de la Subzona de Holcoptychites agrioensis (figura 3). Su distribución espacial también es acotada ya que se restringe al perfil Bajada del Agrio en el sector sur de la zona de estudio, aunque algunas colonias pequeñas y aisladas han sido también registradas en Agua de la Mula en niveles equivalentes. Está compuesta por corales coloniales ramosos y globosos pertenecientes a Columastrea antiqua (Gerth, 1928) y Placocoenia neuquensis Gerth, 1928 respectivamente. Al igual que los corales actuales ambas formas poseían probablemente un modo de alimentación microcarnívoro. Las colonias ramosas son las más abundantes. Se las registró cementadas a sustratos duros aislados como gruesas conchillas de bivalvos y fragmentos de ramas previamente retrabajados. En cambio las colonias globosas, menos abundantes, aparentemente eran cementantes en los primeros estadios de crecimiento y luego reclinantes en estadios posteriores ya que presentan un área de cementación menor al área basal total. Todas las colonias se registraron fuera de su posición de vida y dado su modo de preservación se interpretaron como restos parautóctonos que han estado expuestos durante un tiempo significativo en la interfase agua sedimento ya que presentaron incrustaciones y bioerosión (véase Lazo, 2006). Los incrustantes más abundantes corresponden a Amphidonte (C.) sp., Parsimonia antiquata y Demospongea indeterminada. La bioerosión corresponde a Gastrochaenolites sin lineamientos internos (Lazo, 2006). Las trazas fósiles de la roca hospedante fueron asignadas a la icnofacies de Cruziana.
Interpretación. Al igual que la biofacies de Cucullaea- Mimachlamys-Panopea esta biofacies se desarrolló en un ambiente de shoreface, ubicado entre la línea de baja marea y el nivel de base de ola normal aproximadamente a los 5 m de profundidad. El sustrato era arenoso blando, sin embargo poseía valvas concentradas localmente que actuaban como parches de sustrato duro. La ocurrencia de corales junto a una fauna rica de bivalvos indica niveles normales de oxígeno en el fondo y condiciones de salinidad marina normal. El aprovechamiento del espacio es máximo en esta biofacies e incluso mayor que en la biofacies de Cucullaea-Mimachlamys-Panopea, ya que los corales explotan la parte más alta de la columna de agua (cuadro 2). Es difícil estimar una temperatura exacta para estos depósitos, aunque se podría suponer sobre la base de ejemplos actuales, una temperatura no menor a 16°-18° C (Spalding et al., 2001).
Durante el crecimiento de las colonias la tasa de sedimentación debió ser baja (cuadro 2). Dadas las dimensiones centimétricas de las colonias y la falta de sobrecrecimento generalizado de unas colonias sobre otras, es probable que estos corales no hayan constituido un arrecife o una barrera ambiental sino simplemente un jardín de baja altura desarrollado sobre fondos blandos. Si bien no es posible decidir si poseían o no zooxanthelas simbiontes es muy probable que hayan vivido dentro de la zona fótica dada la baja profundidad del ambiente donde se registran. La presencia de macrobioerosión también indica un ambiente marino somero iluminado (Bromley, 1990). Tanto un moderado influjo clástico como la falta de sustratos duros generalizados pueden haber inhibido el desarrollo de un arrecife. Este tipo de control ya ha sido documentado en el Jurásico Medio de la India (Fürsich et al., 1994). El hecho de que esta biofacies esté restringida al perfil Bajada del Agrio probablemente tenga que ver con la posición más proximal de esta localidad donde la acción de fuertes corrientes de fondo relacionadas con olas de buen tiempo o tormentas facilitaron la llegada de nutrientes y larvas planctónicas.
Biofacies de ostras gregarias (figura 8)

Figura 8. Reconstrucción de la Biofacies de ostras gregarias en Bajada del Agrio, Subzona de Olcostephanus (O.) laticosta, Hauteriviano Temprano / reconstruction of the "gregarious oysters biofacies" in Bajada del Agrio, Olcostephanus (O.) laticosta Subzone, Early Hauterivian.
Descripción. Esta biofacies corresponde a la tafofacies Gr debido a que la presencia de ostras con hábito gregario tiene implicancias paleoambientales particulares (Lazo, 2006). Se registra en las litofacies Fl y Fm correspondientes a depósitos fangosos de plataforma interna ubicados en el tope de la Subzona de Holcoptychites agrioensis y en la Subzona de Olcostephanus (O.) laticosta en Salado Sur y Bajada del Agrio (figura 3). Las diferencias paleogeográficas en cuanto al desarrollo de los agregados son significativas. En Agua de la Mula sólo se registran parches centimétricos de ostras agregadas mientras que hacia el sur se ve que las dimensiones de los agregados aumentan significativamente, pasando de 0,02 a 2,1 m de espesor en Bajada del Agrio. La geometría también cambia, pasando de lentiforme a lenticular. Los agregados se componen fundamentalmente de ostras pertenecientes a Amphidonte (Ceratostreon) sp. de pequeño tamaño (largo máximo = 50 mm). Las ostras están acompañadas por una menor proporción del serpúlido Parsimonia antiquata que aparece como incrustante y en forma muy ocasional suelen encontrarse amonites u otros bivalvos. Las trazas fósiles registradas en las pelitas que rodean los agregados son excavaciones con límites bien definidos y relleno contrastante con las pelitas y fueron asignadas a la icnofacies de Glossifungites. Una asociación de ostras similar a esta biofacies se presenta en la Formación Chachao en Mendoza, aunque en éste caso acompañada de pectínidos y serpúlidos coloniales (Damborenea et al., 1979).
Interpretación. Esta biofacies se desarrolló en fangos de la plataforma interna, la cual se ubica entre los niveles de base de ola de buen tiempo y de tormenta y entre los 5 y 11 m de profundidad. Este ambiente presenta una energía alternante entre alta y baja. Durante períodos de buen tiempo ocurre sedimentación lenta de material fino, mientras que durante las tormentas ocurre un fuerte retrabajo del fondo. Dado que una alta tasa de sedimentación inhibe el establecimiento de las larvas y el crecimiento de los adultos, es probable que en los momentos de máxima expansión de los agregados, el aporte clástico y por consiguiente la turbidez del agua hayan sido relativamente bajos. Las capas de ostras están a menudo separadas por niveles delgados de pelitas y esto indica que la decantación frenó su crecimiento. Dado que no se han detectado niveles de hardgrounds verdaderos, las larvas fundadoras se establecieron directamente sobre el fondo fangoso o sobre bioclastos sueltos disponibles sobre la interfase. Las trazas asociadas indican que el sustrato fangoso era de consistencia firme. A partir de la colonización por larvas fundadoras, nuevas larvas se fueron cementando sobre los primeros individuos adultos formando clusters o pequeños parches y luego bajo condiciones favorables prolongadas en el tiempo se habrían formado los agregados de mayor tamaño. El aprovechamiento del espacio es menor que en otras biofacies dado que la mayoría de los organismos explotaban sólo la columna de agua. Sin embargo, existían organismos infaunales que bioturbaban el fondo e indicarían niveles normales de oxigenación. Las ostras actuales toleran un amplio rango de temperaturas y por ello es difícil estimar un intervalo de temperatura para los agregados descriptos. El gregarismo de ostras en la actualidad ocurre en bahías y estuarios que generalmente presentan una salinidad algo menor a la marina normal (Norris, 1953; Stenzel, 1971). Los organismos estenohalinos nadadores o con una cierta movilidad, si bien no tienen su mayor distribución en este tipo de ambientes, pueden hacer incursiones temporarias (Puffer y Emerson, 1953). Por lo tanto se puede asumir que la salinidad en el momento en que los agregados crecían era algo menor a la marina normal. Dado que existe un cierto time-averaging es probable que no se puedan resolver en forma delicada los cambios temporales y/o espaciales en la salinidad. Por ello pueden quedar preservados amonites junto a los agregados. Las diferencias paleogeográficas que presentan los agregados indican que hacia Bajada del Agrio las condiciones fueron más favorables ya que por su posición más proximal poseía variaciones más importantes en la salinidad que los otros dos perfiles.
Biofacies de serpúlidos gregarios (figura 9)

Figura 9. Reconstrucción de la biofacies de serpúlidos gregarios en Bajada del Agrio, Subzona de Neocomites sp., Valanginiano Tardío / reconstruction of the "gregarious serpulids biofacies" in Bajada del Agrio, Neocomites sp.Subzone, Late Valanginian.
Descripción. Esta biofacies corresponde a la tafofacies Gr debido a que la presencia de serpúlidos con hábito gregario tiene implicancias paleoambientales particulares (Lazo, 2006). Al igual que la biofacies de ostras gregarias la biofacies de serpúlidos gregarios se registra en las litofacies Fl y Fm interpretadas como depósitos fangosos de plataforma interna dentro de la Subzona de Neocomites sp. en Agua de la Mula y Bajada del Agrio. Los agregados de serpúlidos registran un aumento de espesor y extensión lateral hacia la sección de Bajada del Agrio, en forma similar a lo que ocurre con los agregados de ostras. En dicha localidad alcanzan 2 m de espesor. La biofacies se compone de serpúlidos de la especie Parsimonia antiquata cuyos tubos se cementan entre sí en forma subparalela en grupos de hasta 30 tubos. Amphidonte (Ceratostreon) sp. y Plicatula sp. aparecen como incrustantes de los tubos. Las pelitas laterales a los agregados son ricas en Aetostreon sp., ostras reclinantes, cuyos individuos adultos pueden alcanzar un gran tamaño (altura máxima = 300 mm). Las trazas asociadas son excavaciones con límites bien definidos y relleno contrastante con las pelitas. Las mismas fueron asignadas a la icnofacies de Glossifungites.
Interpretación. Las implicancias paleoambientales de esta biofacies son muy similares a las de la biofacies de ostras gregarias ya que se registran en facies y ambientes análogos aunque en distinta posición estratigráfica (figura 3, cuadro 2). Sin embargo, a diferencia de los agregados de ostras actuales, los de serpúlidos no se restringen a ambientes salobres sino que aparecen también en ambientes hipersalinos (Ten Hove y van den Hurk, 1993). Por lo tanto, si bien los agregados del Miembro Pilmatué pudieron haberse desarrollado bajo condiciones hipo o hipersalinas, dado que aparecen alrededor del nivel de máxima inundación y en un área proximal de la cuenca lo más probable es que se hayan formado en aguas hiposalinas al igual que los agregados de ostras.
Biofacies y estratigrafía secuencial del Miembro Pilmatué
En esta sección se discute la distribución vertical de las biofacies del miembro Pilmatué en función de un esquema estratigráfico secuencial para relacionar cambios relativos del nivel del mar con la aparición de determinadas biofacies. Los cambios relativos del nivel del mar producen cambios drásticos en los parámetros ambientales causando una alternancia de hábitats y un consiguiente recambio en la fauna bentónica. En particular, los cambios en la tasa de sedimentación pueden variar no sólo la composición y abundancia de una concentración fósil sino que también pueden modifican su impronta tafonómica (Kidwell, 1986).
En el Miembro Pilmatué se identificaron cinco límites de secuencias y cuatro secuencias deposicionales completas correlacionables entre los tres perfiles analizados (figura 3, cuadro 2). La primera secuencia deposicional no está completa ya que le falta la cuña de margen de plataforma que corresponde al tope de la Formación Mulichinco. El límite de secuencia número cinco coincide con la base del Miembro Avilé y es un límite de secuencia de tipo I (véase Rossi, 2001 y referencias citadas allí).
La SMW está dominada por la biofacies de Cucullaea-Mimachlamys-Panopea y la biofacies de Cucullaea y Steinmanella. La biofacies de corales coloniales aparece también en la SMW, pero sólo dentro de la Subzona de Holcoptychites agrioensis en Bajada del Agrio. En el TST se registran todas las biofacies a excepción de la biofacies de corales coloniales. En particular, las biofacies de ostras y serpúlidos gregarios y la de Modiolus son únicas de este cortejo. La ubicación de dichas biofacies coincide aproximadamente con la posición de la superficie de máxima inundación indicando que se desarrollaron en momentos de baja tasa de sedimentación y máxima disponibilidad de espacio (figura 3). En el HST pueden aparecer ocasionalmente las biofacies de Cucullaea-Mimachlamys- Panopea y la biofacies de Cucullaea y Steinmanella, sin embargo en relación a los otros cortejos es muy poco fosilífero (figura 3).
Discusión
Los factores ambientales que más influencia tuvieron sobre las biofacies analizadas fueron los cambios relativos en el nivel del mar, la tasa de aporte sedimentario, la consistencia del sustrato, la oxigenación del fondo y los cambios en la salinidad.
Las biofacies de ostras y serpúlidos gregarios se registran en el TST, especialmente en las etapas de inundación máxima donde aumenta rápidamente el espacio de acomodación y disminuye la tasa de sedimentación. Los agregados, prácticamente monoespecíficos, reflejan un ambiente de salinidad distinta a la marina normal y una baja tasa de aporte sedimentario. Su expresión máxima ocurrió hacia Bajada del Agrio lo cual sugiere que en zonas proximales de la cuenca los cambios de salinidad eran más importantes. Dado que poseen un cierto time-averaging los agregados pueden aparecer mezclados, aunque en baja proporción, con organimos estenohalinos nectónicos como los amonites.
De lo expuesto se desprende que el Miembro Pilmatué de la Formación Agrio, a pesar de ser interpretado tradicionalmente como depositado en un ambiente netamente marino, sufrió variaciones temporales en la salinidad las cuales fueron más pronunciadas hacia sectores proximales posiblemente por la mayor cercanía a la desembocadura de cursos fluviales. Los cambios de salinidad probablemente no fueron bruscos ni de mucha magnitud, ya que no se observan variaciones en las litofacies que así lo indiquen. Sin embargo, las modificaciones fueron lo suficientemente prolongadas en el tiempo como para desencadenar el agregado de organismos de hasta 2 m de espesor. La forma de golfo y la presencia de un arco volcánico insular en el límite este probablemente favorecieron los cambios de salinidad por mantener la cuenca relativamente aislada. Sería muy interesante comprobar la magnitud de los cambios de salinidad analizando isótopos de oxígeno a partir de conchillas no recristalizadas, lo cual está fuera del alcance de este trabajo.
La biofacies de Modiolus en pelitas es también típica del TST, pero dentro de la subzona de Holcoptychites neuquensis. Esta biofacies ocupa una posición intermedia en la columna entre las dos subzonas en las que aparecen los agregados de serpúlidos y ostras. Esto indica que las condiciones ambientales de los cortejos transgresivos no fueron iguales. En este caso un sustrato firme y una baja oxigenación debida probablemente a una combinación de un gran espacio de acomodación y una restricción parcial de la cuenca favorecieron la colonización por parte de esta especie.
La biofacies de Cucullaea-Mimachlamys-Panopea y la biofacies de Cucullaea y Steinmanella responden a cambios relativos en la escala de parasecuencias, de mayor frecuencia que en las biofacies del TST mencionadas anteriormente. Es por esto que dichas biofacies se reiteran a lo largo de todo el perfil sin importar el cortejo sedimentario analizado. La biofacies de corales coloniales aparece asociada a la biofacies de Cucullaea-Mimachlamys-Panopea, pero aparece sólo en un sector proximal del shoreface en Bajada del Agrio. Las formas ramosas probablemente predominaron sobre las globosas ya que pueden acomodar más fácilmente el sedimento entre las colonias. Además en las ramosas los coralitos pueden tener distintas orientaciones a resguardo de la sedimentación. La falta de hardgrounds, la ausencia de sobrecrecimientos y el pequeño tamaño alcanzado por las colonias indican que el aporte sedimentario nunca cesó por completo lo cual limitó el crecimiento de los corales. De esta manera conformaron un jardín de baja altura sobre un fondo mixto arenoso y bioclástico.
En el TST se presenta la mayor diversidad de biofacies lo cual sugiere que en momentos de ascenso relativo se generaban ambientes heterogéneos capaces de alojar a biofacies con distinto significado paleoambiental. En cambio en la SMW aparecen sólo dos biofacies, por lo cual probablemente los ambientes generados durante períodos de mar bajo fueron relativamente homogéneos. A su vez, el HST es muy poco fosilífero probablemente debido a la dilución por el gran aporte clástico en un contexto altamente progradante.
Por último cabe destacar las importantes variaciones paleogeográficas que ocurren entre los tres perfiles analizados, los cuales están separados sólo por 44,2 km de distancia. En Bajada del Agrio, hacia el sur de la zona de estudio, aparece una mayor complejidad tanto espacial como temporal en la fauna bentónica. Allí hay una buena expresión de todas las biofacies descriptas, siendo el perfil con fauna más diversificada. En cambio, Agua de la Mula y Salado Sur son secciones más monótonas en cuanto al reemplazo temporal de biofacies ya que probablemente no estaban tan afectadas por los cambios de salinidad.
Conclusiones
Se distinguen seis biofacies cuya distribución vertical responde a los cambios relativos del nivel del mar, tasa de sedimentación, consistencia del sustrato, oxigenación y la salinidad.
La biofacies de Cucullaea-Mimachlamys-Panopea se registra en un shoreface de moderada a alta energía, baja tasa de sedimentación, sustrato arenoso de consistencia blanda con parches de bioclastos, buena oxigenación, salinidad marina normal y aguas templado- cálidas.
La biofacies de Cucullaea y Steinmanella se registra en la plataforma interna y media-externa y caracteriza un ambiente de baja energía, baja tasa de sedimentación, sustrato fangoso firme con la zona de anaerobiosis relativamente cercana a la interfase agua-sedimento y salinidad marina normal.
La biofacies de Modiolus es también típica de sustratos fangosos de la plataforma, pero a diferencia de la biofacies anterior colonizaba el fondo durante períodos de baja oxigenación.
La biofacies de corales coloniales corresponde a un jardín de formas ramosas y globosas de baja altura que no conformó una barrera ambiental. Se asocia a la biofacies de Cucullaea-Mimachlamys-Panopea apareciendo sólo en un sector proximal sur de la cuenca.
La biofacies de Cucullaea-Mimachlamys-Panopea y la biofacies de Cucullaea y Steinmanella responden a cambios relativos cuya frecuencia es del orden de las parasecuencias, mientras que el resto de las biofacies responden a cambios del orden de secuencias.
Las biofacies de ostras y serpúlidos gregarios responden a cambios en la salinidad marina normal ocurridos durante períodos de ascenso relativo del nivel del mar y se asocian a la superficie de máxima inundación.
El cortejo transgresivo alojó a cinco de las seis biofacies analizadas lo cual probablemente refleja que durante los ascensos relativos del nivel del mar había una mayor heterogeneidad ambiental que en el resto de los cortejos. Por el contrario, en la cuña de margen de plataforma se registraron sólo tres de las biofacies, lo cual indicaría una menor diversidad de ambientes durante los descensos relativos. El cortejo de mar alto es muy poco fosilífero debido probablemente a una gran dilución por llegada de un importante aporte clástico en un contexto altamente progradante.
La sección de Bajada del Agrio presenta la mayor diversidad de fauna bentónica y mayor complejidad en el patrón espacial y reemplazo temporal de las biofacies descriptas. Esto probablemente se deba a su posición más proximal dentro de la cuenca lo cual se vincula con una menor tasa de sedimentación promedio, una mayor abundancia de superficies de amalgamación y una mayor influencia de los cambios de salinidad.
Apéndice. Lista de especies de cada una de las biofacies reconocidas en el Miembro Pilmatué de la Formación Agrio. En la biofacies de Cucullaea-Mimachlamys-Panopea se muestra la abundancia de cada especie que resultó de un conteo realizado en Agua de la Mula en 30 niveles del Miembro Pilmatué. Abreviaturas: MA, modo de alimentación; S, suspensívoro; M, microcarnívoro / list of species of the biofacies of the Pilmatué Member of the Agrio Formation. For the "Cucullaea-Mimachlamys-Panopea biofacies" counts of taxa are presented. Abbreviations are as follows: MA, feeding habit; S, suspension feeder; M, microcarnivorous.
Agradecimientos
Deseo agradecer a M.B. Aguirre-Urreta, A. Concheyro, S. Lanés, M. Cichowolski, D. Rodríguez, P. Rawson, L. Buatois, M.G. Mángano, R. Palma, L. Legarreta, C. Gulisano y E. Olivero por su ayuda y estímulo permanente. G. Iovine, C. Rodriguez y R. Sanci colaboraron en los trabajos de campo en Neuquén. A. Zamorano realizó los dibujos. Agradezco a F. Fürsich y S. Lamés que arbitraron el trabajo. El financiamiento de este trabajo provino de una beca de formación de postgrado del CONICET de cuatro años de duración y un subsidio para estudiantes de postgrado de la International Association of Sedimentologists ambos otorgados al autor, y los subsidios UBACyT X 084 y ANPCYT PICT 03/14143 a cargo de M.B. Aguirre-Urreta.
Bibliografía
1. Agassiz, L. 1840. Etudes critiques sur les mollusques fossiles. Mémoire sur les Trigonies. Neuchâtel, 58 pp., lám. 1-11. [ Links ]
2. Aguirre-Urreta, M.B. y Rawson, P.F. 1997. The ammonite sequence in the Agrio Formation (Lower Cretaceous), Neuquén Basin, Argentina. Geological Magazine 134: 449-458. [ Links ]
3. Aguirre-Urreta, M.B. y Rawson, P.F. 2003. Lower Cretaceous ammonites from the Neuquén Basin, Argentina: the Hauterivian genus Holcoptychites. Cretaceous Research 24: 589-613. [ Links ]
4. Behrendsen, O. 1892. Zur Geologie des Ostabhanges der argentinischen Cordillere. II. Theil. Zeitschrift der Deutschen geologischen Gesellschaft 44: 1-42, lám. 1-4. [ Links ]
5. Brenchley, P.J. 1990. Biofacies. En: D.E.G. Briggs y P.R. Crowther (eds.), Palaeobiology a synthesis, Blackwell Scientific Publications, Cambridge, pp. 395-400. [ Links ]
6. Bromley, R.G. 1990. Trace fossils. Biology and Taphonomy. Special Topics in Palaeontology 3: 280 pp. [ Links ]
7. Brongniart, A. 1822. Lutraria ? gurgitis, lámina 9, figura 15. En: G. Cuvier y A. Bromgniart (eds.), Description géologique des environs de Paris, Paris, 428 p., 17 lám. [ Links ]
8. Burckhardt, C. 1903. Beirträge zur Kenntnis der Jura- und Kreideformation der Cordillere. Palaeontographica 50: 1-144. [ Links ]
9. Carrera, M. 2001. Análisis de la distribución y composición de las biofacies de la Formación San Juan (Ordovícico Temprano), Precordillera Argentina. Ameghiniana 38: 169-184. [ Links ]
10. Cichowolski, M. 2003. The nautiloid genus Cymatoceras from the Cretaceous of the Neuquén and Austral basins, Argentina. Cretaceous Research 24: 375-390. [ Links ]
11. Damborenea, S.E. y Manceñido, M.O. 1996. Icnofósiles (nucleocavia) preservados sobre moldes internos de conchillas del Jurásico Medio del oeste argentino. Asociación Paleontológica Argentina, Publicación Especial 4, 1° Reunión Argentina de Icnología, pp. 111-120. [ Links ]
12. Damborenea, S.E. y Manceñido, M.O. 2005. Biofacies analysis of Hettangian-Sinemurian bivalve/brachiopod associations from the Neuquén Basin (Argentina). Geologica Acta 3: 163-178. [ Links ]
13. Damborenea, S.E., Manceñido, M.O. y Riccardi, A.C. 1979. [ Estudio paleontológico de la Formación Chachao. Informe Interno YPF, 152 pp., Inédito.] . [ Links ]
14. Fürsich, F.T., Pandey D.K., Oschmann W., Jaitly A.K. y Singh, I.B. 1994. Ecology and adaptive strategies of corals in unfavourable environments: Examples from the Middle Jurassic of the Kachchh Basin, Western India. Neues Jarhbuch für Geologie und Paläontologie Abhanlungen 194: 269-303. [ Links ]
15. Gerth, H. 1928. Beiträge zur kenntnis der mesozoischen korallenfaunen von Südamerika. Leidsche Geologische Mededeelingen 3: 1-16. [ Links ]
16. Howard, J.D. y Reineck, H-E. 1981. Depositional facies of highenergy beach-to-offshore sequence: comparison with lowenergy sequence. American Association of Petroleum Geologists Bulletin 65: 807-830. [ Links ]
17. Imlay, R.W. 1940. Neocomian faunas of northern Mexico. Bulletin of the Geological Society of America 51: 117-190. [ Links ]
18. Kidwell, S.M. 1986. Models for fossil concentrations: paleobiologic implications. Paleobiology 12: 6-24. [ Links ]
19. Kidwell, S.M. 1998. Time-averaging in the marine fossil record: overview of strategies and uncertainties. Geobios 30: 977-995. [ Links ]
20. Lazo, D.G. 2003a. The genus Steinmanella (Bivalvia, Trigonioida) in the Agrio Formation (Lower Cretaceous), Neuquén Basin, Argentina. Journal of Paleontology 77: 1069-1085. [ Links ]
21. Lazo, D.G. 2003b. Taxonomy, facies relationships and palaeobiology of bakevelliid bivalves from the Lower Cretaceous of west-central Argentina. Cretaceous Research 24: 765-788. [ Links ]
22. Lazo, D.G. 2004. [ Análisis de concentraciones fósiles del Cretácico Inferior de cuenca Neuquina. Tesis doctoral, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 337 pp., Inédito.] . [ Links ]
23. Lazo, D.G. 2006. Análisis tafonómico e inferencia del grado de mezcla temporal y espacial de la macrofauna del Miembro Pilmatué de la Formación Agrio, Cretácico Inferior de cuenca Neuquina, Argentina. Ameghiniana 43: 311-326. [ Links ]
24. Leanza, H.A. 1998. Una nueva especie de Steinmanella Crickmay (Bivalvia, Trigonioida) del Cretácico Inferior de la Cuenca Neuquina, Argentina. Revista Geológica de Chile 25: 57-67. [ Links ]
25. Leanza, H.A., Hugo, C.A. y Repol, D. 2001. Hoja Geológica 3969-I, Zapala. Provincia del Neuquén, Instituto de Geología y Recursos Minerales, Servicio Geológico Minero Argentino, Boletín 275: 128 pp. [ Links ]
26. Leymerie, M.A. 1842. Suite du mémoire sur le terrain Crétacé du département de l'Aube. Second partie (partie paléontologique). Mémoires de la Société Géologique de France 5: 1-34. [ Links ]
27. Ludvigsen, R., Westrop, S.R., Pratt, B.R., Tuffnell, P.A. y Young, G.A. 1986. Paleoscene # 3. Dual biostratigraphy: Zones and biofacies. Geoscience Canada 13: 139-154. [ Links ]
28. Norris, R. M. 1953. Buried oyster reefs in some Texas bays. Journal of Paleontology 27: 569-576. [ Links ]
29. Orbigny d', A. 1844-1847. Paléontologie Française. Description des Mollusques et Rayonnés Fossiles. Terrains Crétacés. III. Lamellibranches. Mason, Paris, 807 pp. [ Links ]
30. Orbigny d', A. 1850. Prodrome de Paléontologie stratigraphique universelle des animaux mollusqués et rayonnés. Masson, Paris, 2: 1-428. [ Links ]
31. Puffer, E.L. y Emerson, W.K. 1953. The molluscan community of the oyster-reef biotope on the central Texas coast. Journal of Paleontology 27: 537-544. [ Links ]
32. Rhoads, D.C. y Young, D.K. 1970. The influence of deposit-feeding organisms on sediment stability and community trophic structure. Journal of Marine Research 28: 150-178. [ Links ]
33. Rodriguez, D.L. 2001. [ Equinoideos irregulares del Cretácico Inferior de la cuenca Neuquina. Tesis de Licenciatura, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 56 pp. Inédito.] . [ Links ]
34. Rossi, G.C. 2001. [ Arenisca Avilé: Facies, ambientes sedimentarios y estratigrafía de una regresión forzada del Hauteriviano inferior de la cuenca Neuquina. Tesis Doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 231 pp. Inédito.] . [ Links ]
35. Scasso, R.A., Olivero, E.B. y Buatois, L.A. 1991. Lithofacies, biofacies, and ichnoassemblage evolution of a shallow submarine volcaniclastic fan-shelf depositional system (Upper Cretaceous, James Ross Island, Antarctica). Journal of South American Earth Sciences 4: 239-260. [ Links ]
36. Scott, R.W. 1970. Paleoecology and paleontology of the Lower Cretaceous Kiowa Formation, Kansas. The University of Kansas Paleontological Contributions 52 (Cretaceous 1): 1-94. [ Links ]
37. Sowerby, J. 1812-1822. The Mineral Conchology of Great Britain, Meredith. London, 1-383 pp. [ Links ]
38. Sowerby, J. de C. 1823-1846. The Mineral Conchology of Great Britain. London, 384-648 pp. [ Links ]
39. Spalding, M.D., Ravilious, C. y Green, E.P. 2001. World Atlas of Coral Reefs. University of California Press, Berkeley, 424 pp. [ Links ]
40. Steinmann, G. 1881. Zur Kenntniss der Jura- und Kreideformation von Caracoles (Bolivia). Neues Jahrbuch für Mineralogie, Geologie und Palaeontologie, Beilage-Band I, 239-301, lám. 9-14. [ Links ]
41. Stenzel, H. 1971. Oysters. En: R.C. Moore y C. Teichert (eds.), Treatise on Invertebrate Paleontology. Part N. Mollusca 6. Bivalvia Volume 3, University of Kansas and Geological Society of America, pp. N953-N1224. [ Links ]
42. Taylor, E.M. y Glover, E.A. 2000. Functional anatomy, chemosymbiosis and evolution of the Lucinidae. En: E.M. Harper, J.D. Taylor y J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia. Geological Society, London, Special Publication 177: 207-225. [ Links ]
43. Ten Hove, H.A. y van den Hurk, P. 1993. A review of recent and fossil serpulid "reefs"; actuopalaeontology and the "Upper Malm" serpulid limestones in NW Germany. Geologie en Mijnbouw 72: 23-67. [ Links ]
44. Weaver, C.E. 1931 Paleontology of the Jurassic and Cretaceous of West Central Argentina. Memoirs of the University of Washington 1: 1-595. [ Links ]
45. Wignall, P.B. 1990. Benthic palaeoecology of the late Jurassic Kimmeridge Clay of England. Special Papers in Palaeontology 43: 1-74. [ Links ]
46. Wignall, P.B. 1993. Distinguishing between oxygen and substrate control in fossil benthic assemblages. Journal of the Geological Society of London 150: 193-196. [ Links ]
Recibido: 27 de julio de 2005.
Aceptado: 1 de junio de 2006.

