










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.1 Buenos Aires ene./mar. 2007
Fabaceae de la Formación El Morterito (Mioceno Superior) del valle del Cajón, provincia de Catamarca, Argentina
Luisa M. Anzótegui1, Silvina S. Garralla1 y Rafael Herbst2
1Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste y Centro de Ecologia. Casilla de Correo 291, 3400 Corrientes, Argentina. luisaanzotegui@arnet.com.ar
2Instituto Superior de Geología- Consejo Nacional de Investigaciones Científicas y Técnicas, Las Piedras 201 7º/B, 4000 Tucumán, Argentina. rafaherbst@uolsinectis.com.ar
Abstract. Fabaceae from the El Morterito Formation (Upper Miocene), valle del Cajón, Catamarca province, Argentina. Fabaceae leaves from the floral assemblage of the El Morterito Formation (Upper Miocene) from the valle del Cajón in Catamarca Province, Argentina, are described. The taxa are assigned to Caesalpinoideae: Senna cf. bicapsularis (L.) Rouxbourgh, Senna cf. obtusifolia (L.) Irwin and Barneby, Chamaecrista cf. flexuosa (L) Greene; to Mimosoideae: Mimosa cf. flagellaris, Bentham, Mimosa sp., Prosopis sp. 1; and to Papiliodoideae: Desmodium cf. ascendens (Swart) De Candolle, cf. Desmodium sp., Eriosema sp. and Tipuana tipu (Bentham) Kuntze. These species are compared with those treated in older literature and, also, some taxonomic considerations are made. Based on the characters of the most similar living taxa and additional sedimentological, invertebrate and vertebrate fossil data, an open dry environment with xerophytic vegetation is inferred, associated with more humid habitats with riparian forests, characteristic of a warm climate.
Resumen. Se describen hojas de Fabaceae que constituyen parte de la asociación florística de la Formación El Morterito (Mioceno Superior) del valle del Cajón, provincia de Catamarca. Los taxones son incluidos en Caesalpinoideae: Senna cf. bicapsularis (L.) Rouxbourgh, Senna cf. obtusifolia (L.) Irwin y Barneby, Chamaecrista cf. flexuosa (L) Greene; en Mimosoideae: Mimosa cf. flagellaris, Bentham, Mimosa sp., Prosopis sp. 1 y en Papiliodoideae: Desmodium cf. ascendens (Swart) De Candolle, cf. Desmodium sp. Eriosema sp.y Tipuana tipu (Bentham) Kuntze. Estas formas son comparadas con otras mencionadas en la literatura regional más antigua y se hacen algunas consideraciones taxonómicas. En base a las características de los taxones actuales, más afines a los datos sedimentológicos, de invertebrados y vertebrados fósiles, se infiere el predominio de ambientes abiertos y secos con vegetación xerófila y otros algo más húmedos con bosques ribereños, característicos de clima cálido.
Key words. Fabaceae; Leaf impressions; Upper Miocene; Catamarca; Argentina.
Palabras clave. Fabaceae; Impresiones de hojas; Mioceno Superior; Catamarca; Argentina.
Introducción
Los estudios paleoflorísticos (hojas, frutos, polen y maderas) de Formaciones neógenas del noroeste argentino: formaciones San José, Chiquimil y Palo Pintado (Herbst et al., 1987; Anzótegui y Cuadrado, 1996; Anzótegui, 1998; Acevedo et al., 1997, 1998; Anzótegui y Cristalli, 2000; Herbst et al., 2000; Starck y Anzótegui, 2001; Anzótegui y Ferraro, 2000; Anzótegui, 2002a, 2002b y 2002c; Mautino et al. 1997; Mautino y Anzótegui, 1998, 2000, 2002, 2004; Mautino et al. 2004; Anzótegui y Herbst, 2004; Anzótegui, 2004; Anzótegui et al. 2004; Martínez y Lutz, en prensa) reflejan que la paleovegetación es de extracción Neotropical. Esto está dado, pues su composición se caracteriza por la ausencia de Nothofagus, presencia de comunidades xerofíticas, de ribera, palustres y acuáticas, afines por sus integrantes a las distintas provincias fitogeográficas del actual Dominio Chaqueño (Cabrera, 1976). Por otra parte, esta paleoflora, que se habría desarrollado bajo condiciones climáticas cálidas, por su ubicación geográfica y edad se encuentra comprendida en la "Paleoflora Subtropical Neógena" del esquema Paleofitogeográfico de Hinojosa y Villagran (1997) e Hinojosa (2005).
La paleoflora de la Formación El Morterito, también del noroeste argentino, de la localidad río Totoral, provincia de Catamarca (figura 1), está compuesta por numerosas impresiones de láminas foliares, entre las que predominan especies de Fabaceae. Dada esta particularidad, en la presente contribución se dan a conocer 10 especies de folíolos dispersos. Esto, junto con las características de los análogos modernos de las especies fósiles, permite inferir algunas condiciones paleoclimáticas y paleoambientales predominantes en el Mioceno Superior de la región de estudio. La paleoflora estudiada y otros hallazgos de la región y de la Argentina en general, demuestran que las Fabaceae tuvieron una representación importante en la vegetación del Neógeno en el país.
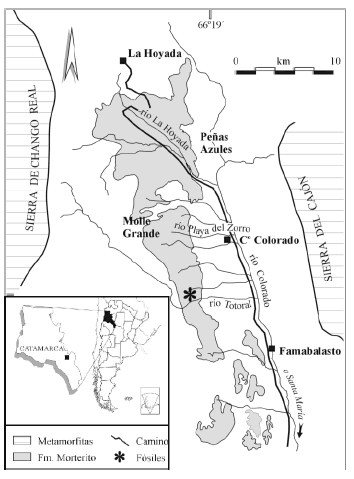
Figura 1. Mapa de ubicación de la localidad fosilífera / map of the fossiliferous locality.
La familia Fabaceae (Leguminosae, nomen conservandum) está dividida en tres subfamilias, Caesalpinoideae, Mimosoideae y Papilionoideae (Polhill y Raven, 1981), e incluye unos 686 géneros y más de 18.000 especies distribuidas en todo el mundo, principalmente en regiones tropicales y subtropicales, bajo clima árido o semiárido (Kurkbride et al., 2003). En Argentina se conocen 113 géneros y 732 especies, ubicadas preferentemente en el centro y norte de país (Zuloaga y Morrone, 1999). El registro fósil en el Paleógeno es relativamente escaso ya que cuenta hasta el momento con 11 especies descriptas, llegando en el Neógeno a unas 40 especies (cuadro 1).
Cuadro 1. Fabaceae fósiles de Argentina / fossil Fabaceae from Argentina.

Geología
Turner (1973) estableció un esquema estratigráfico del Neógeno, constituido por el Grupo El Bolsón, con las Formaciones El Morterito, El Áspero y El Cajón. Malizia et al. (1990) publicaron un perfil detallado del río Totoral, (localidad de donde provienen las plantas aquí descriptas) en el que individualizan sedimentitas finas, de colores amarillento y verdoso en su facies B. Posteriormente Bossi et al. (1993) estudiaron la sedimentología del Neógeno de este valle y establecieron un nuevo esquema estratigráfico, basado en Aloformaciones: Peñas Azules, Playa del Zorro y Totoral. Ambos esquemas (Turner, 1973 y Bossi et al., 1993) no son estrictamente equivalentes entre sí. La Aloformación Playa del Zorro tiene un espesor de 1288 m a la altura del puesto Molle Grande (figura 1). Por sus características está dividida en tres secciones: inferior, media y superior; la inferior está constituida por areniscas macizas y conglomerados volcánicos, de color verde oliva oscuro a castaño. La sección media (lacustre, subsecuencia II) por limolitas amarillas y verde oscuras con estratificación delgada y laminación paralela y la superior por areniscas finas a medianas con algunos niveles edáficos. En los mismos niveles que las plantas (sección media) se hallan abundantes pelecípodos, gasterópodos, ostrácodos y escamas de peces. En general esta unidad representa un sistema fluvial de ríos sinuosos con etapas de inundación permanente y formación de cuerpos de agua transitorios, en los que se encuentran los fósiles. Más tarde Bossi et al. (2000) complementan varios aspectos de esta secuencia, definen las paleocorrientes así como la procedencia de los materiales y establecen además algunas consideraciones paleoambientales. A los fines de este trabajo, se utiliza la nomenclatura tradicional establecida por Turner (1973) que permite realizar correlaciones con otras formaciones de la región.
Los sedimentos portadores de esta flora son equivalentes a los de la Formación Chiquimil, del Grupo Santa María, que aflora en el valle homónimo, los que pueden servir como columna tipo para el Neógeno del noroeste argentino.
Edad
Turner (1973) ubicó la Formación El Morterito en el "Calchanquense" y la asignó al Mioceno sensu lato. Malizia et al. (1990) correlacionaron su "facies B" con la Formación San José (Mioceno Medio) del valle de Santa María, pero datos más recientes provistos por Bossi et al. (2000) señalan que dos dataciones (Ar/Ar) de una toba ubicada por encima de los niveles lacustres establecen valores de 5.38 ± 0.07 y 6.04 ± 0.07 lo que confiere a estos niveles una antigüedad dentro del Mioceno Superior. Estas cifras concuerdan con las obtenidas para los depósitos lacustres de la Formación Chiquimil (Bossi et al., 2001) y los del Miembro El Jarillal de la misma Formación en el valle de Villavil (Muruaga, 2001) con los que, además, comparte algunas especies de la fauna.
Materiales y métodos
El sedimento contiene abundantes láminas foliares, de ellas muchas se encuentran carbonizadas conservando sólo la forma y unas pocas permiten observar la venación en detalle; del conjunto, Senna cf. bicapsularis, Senna cf. obtusifolia y Chamaecrista flexuosa son las dominantes. Para su estudio simplemente fueron limpiadas y en algunos casos destapadas con pequeños cinceles. La terminología empleada en las descripciones es básicamente la propuesta por Hickey (1974, 1979) y Ash et al. (1999). Las láminas foliares contienen algunos o todos los elementos que Herendeen (en Herendeen y Dilcher, 1992) considera típicos de esta familia: pulvínulos abultados sobre el peciólulo, margen entero y lámina o base asimétricas. Las especies fósiles han sido determinadas por comparación con ejemplares actuales del Herbario CTES y con la bibliografía especializada. Las características de cada forma fósil se encuentran comprendidas en la variabilidad que presentan las hojas de las especies actuales afines. Se las incluye en el sistema natural de clasificación (Lindley,1830; Cronquist et al., 1966) porque tradicionalmente, para este tipo de fósiles, en la literatura existen pocos morfogéneros. Con el fin de indicar mayor semejanza con la especie actual afín, en aquellos géneros que contienen numerosas especies y morfología foliar variada, se emplea la abreviatura cf. para una asignación específica insegura pero probable (Meléndez, 1998). Las fotografías han sido tomadas con una cámara digital Cannon Power Shot A70.
Repositorio. Los ejemplares estudiados se encuentran depositados en la colección Paleobotánica de la Facultad de Ciencias Exactas y Naturales de Universidad Nacional del Nordeste (Corrientes), bajo el acrónimo CTES-PB.
Paleontología sistemática
División MAGNOLIOPHYTA Cronquist, Taktajan y Zimmermann 1966
Clase MAGNOLIOPSIDA Cronquist, Taktajan y Zimmermann 1966
Familia FABACEAE Lindley 1830
Subfamilia CAESALPINOIDEAE De Candolle 1825Género Senna Miller 1754
Especie tipo. Senna alexandrina Miller 1754.
Senna cf. bicapsularis (L.) Rouxbourgh 1832 Figuras 2.A, 3.A, B

Figura 2. A-J, Esquema de folíolos con detalle de venación / sketches of leaflets with detail of venation. A, Senna cf. bicapsularis, reconstrucción / reconstruction CTES-PB 10857 y/and CTES-PB 10925 (Escala / scale 0,25 cm). B, Senna cf. obtusifolia CTES-PB 10877 (Escala / scale 0,8 cm). C, Chamaecrista flexuosa CTES-PB 10985 (Escala / scale 5,2 cm). D, Mimosa cf. flagellaris CTES-PB 10965 (Escala / scale 0,1 cm). E, Mimosa sp. Fragmento de hoja / leaf fragment CTES-PB 10888a (Escala / scale 0,4 cm). F, Prosopis sp 1, reconstrucción / reconstruction CTES-PB 10988 a y b (Escala / Scale 0,3 cm). G, Cf. Desmodium sp. CTES-PB 10973 (Escala / Scale 0,5 cm). H, Eriosema sp. CTES-PB 10916 (Escala / scale 0,4 cm). I, Tipuana tipu CTES-PB 10923 (Escala / scale 0,5 cm). J, Desmodium cf. ascendens CTES-PB 10915 (Escala / scale 0,5 cm).

Figura 3. A, B, Senna cf. bicapsularis. A, aspecto de la hoja / leaf general aspect CTES-PB 10857 (x 2). B, detalle de la venación / detail of venation CTES-PB 10925 (x 8). C, Cf. Desmodium sp. Detalle de un fragmento de la lámina / detail of the leves fragment CTES-PB 10973 (x 1,8). D, Mimosa sp. Fragmento de hojas / leaves fragment CTES-PB (x 5,5). E, F Senna cf. obtusifolia CTES-PB 10877. E, hoja /leaf CTES-PB 10877 (x 2). F, detalle de la venación / detail of venation CTES-PB 10877(x 4). G,H, Tipuana tipu detalle de venación en un fragmento de hoja / detail of venation in a leaf fragment CTES-PB 10935. G, detalle de la venación / detail of venation CTES-PB 10935 (x 2). H, aspecto de la hoja / leaf general aspect CTES-PB 10935 (x 2,5). I, Desmodium cf. ascendens CTES-PB 10915 (x 4,5). J, K, Prosopis sp. 1 CTES-PB 10918a y 10988b. J, aspecto de la hoja / leaf general aspect CTES-PB 10918a (x 3,5). K, detalle de la venación / detail of venation CTES-PB 10988b (x 2,5). L, Chamaecrista flexuosa CTES-PB 10985 (x 10). M, O, Eriosema sp. CTES-PB 10916. O, detalle de la venación / detail of venation CTES-PB 10916 (x 8). M, aspecto de la hoja / leaf general aspect CTES-PB 10916 (x5). N, Mimosa cf. flagellaris CTES-PB 10965 (x 10).
Descripción. Folíolos dispersos, aparentemente coriáceos, ovado - laceolados de 1-1,5 cm de largo por 0,3- 0,5 cm de ancho, asimétricos, de margen entero y liso; ápice agudo mucronado, base asimétrica levemente cuneada en una hemilámina y con una leve lobulación redondeada en la otra hemilámina, peciólulo con pulvínulo muy pequeño y excéntrico. Venación broquidódroma. Vena primaria excéntrica, recta, llega hasta el ápice del folíolo, menor de 0,1 cm, de grosor moderado (1,25-2%). Venas secundarias débiles, opuestas a subopuestas, en número de 6-8 de cada lado, las venas secundarias que se encuentran en la hemilámina más desarrollada emergen con ángulo agudo ancho (80-90º) mayor que los de la otra hemilámina que son agudo-moderados (40-60º). Venación terciaria aparentemente reticulada ortogonal.
Material estudiado. CTES-PB 10802, 10857, 10874, 10884, 10903, 10925, 10926, 10932, 10947, 10955, 10968, 10969, 10970.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Medio (Malizzia et al. 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Esta especie se compara con Cassia patagonica Berry, de la localidad río Pichileufú (Berry, 1938), del Paleoceno-Eoceno de Argentina y con C. singewaldi Berry, de la localidad Corocoro (Mioceno Medio) de Bolivia (Berry, 1919; Gregory-Wodzicki et al., 1998), porque tienen base asimétrica, ápice mucronado y son mayores de 1 cm de longitud; no obstante, ambas se diferencian de la especie aquí descrita porque la de Argentina (C. patagonica) tiene la base más redondeada en ambas hemiláminas y la de Bolivia (C. singewaldi) tiene mayor cantidad de venas secundarias; Senna sp. del Mioceno Medio (Anzótegui y Herbst, 2004), difiere de Senna cf. bicapsularis por la forma, tamaño y tipo de venación.
Los ejemplares fósiles aquí descritos se parecen por la forma, tipo de ápice, base y venación, a los folíolos más pequeños de la especie actual de Senna bicapsularis; también son parecidos por los mismos caracteres a los de S. birostris (Dombey Ex Vogel) H.S. Irwin y Barneby, pero los folíolos de esta especie son pubescentes y menos coriáceos, en consecuencia la venación es poco visible.
Hábitat y hábito de la especie actual. S. bicapsularis actualmente es una planta arbórea que vive a orillas de ríos y arroyos en clima cálido húmedo. Es una especie sudamericana, aunque no ha sido citada para Argentina (Irwin y Barneby, 1982).
Senna cf. obtusifolia (L.) Irwin y Barneby, 1982 Figuras 2.B, 3.E, F
Descripción. Lámina foliar entera elíptica a obovada, de 2,4-2,6 cm de largo por 1-1,5 cm de ancho, ligeramente asimétrica, de ápice agudo y base cuneada, peciólulo con pulvínulo, de 0,03 cm de largo. Venación broquidódroma, vena primaria recta, menor de 0,01 cm de grosor moderado (2,5%) uniforme en todo su trayecto. Venas secundarias en número de 5 - 8, subopuestas, ángulo de divergencia agudo-moderado (40º- 60º), presentan un recorrido levemente sinuoso. Venas intersecundarias compuestas, visibles en algunos espacios intercostales. Venas terciarias describen un modelo reticulado ortogonal. Venación de orden mayor no se observa.
Material estudiado. CTES-PB , 10856,10868a, 10869, 10870 a y b, 10871, 10872, 10877 a, 10889, 10934, 10941, 10951e, 10960.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Entre las especies bolivianas de la flora fósil del Mioceno Medio -Superior de Corocoro (Berry, 1919) y Pislepampa (Berry, 1922a; Gregory- Wodzicki et al., 1998), Pithecolobium palcanum Berry y Cassia rigidulifolia Engelhardt, son semejantes por la venación broquidódroma, tamaño y base cuneada pero difieren porque la primera tiene ápice redondeado y la base más recta que la especie aquí descrita, y la segunda el ápice retuso. Cassia singewaldi Berry y C. cristoides Engelhardt, tienen el tamaño y algunos caracteres de la forma (obovados, base cuneada) semejantes a los de Senna cf. obtusifolia, pero se diferencian porque la venación es camptódroma. Entre las especies actuales de la flora argentina, es semejante por el tamaño, forma y tipo de venación a Senna obtusifolia; no obstante algunos ejemplares fósiles tienen forma elíptica por lo que también se parecen a los folíolos de Senna corymbosa (Lam.) Irwin y Barneby, 1982.
Hábitat y hábito de la especie actual. Senna obtusifolia es una hierba integrante de la vegetación de ribera y de pastizales del planalto brasileño y del noroeste argentino (Irwin y Barneby, 1982).
Género Chamaecrista (L.) Moench, 1794
Especie tipo. Chamaecrista nictitans Moench 1794.
Chamaecrista flexuosa (L.) Greene 1899 Figuras 2.C, 3.L
Descripción. Folíolos dispersos aparentemente coriáceos, asimétricos, oblongo-loriformes, de márgenes lisos de 0,5-0,7 cm de largo por 0,2 cm de ancho. Ápice agudo y base asimétrica. Venación actinódroma, tres venas primarias rectas menores de 0,1 cm (<1,25%) emergen del pulvínulo; dos de ellas son largas y llegan hasta la zona apical sin unirse, la restante termina en la región media del folíolo. Venas secundarias cortas y rectas, apenas perceptibles, emergen de las primarias con ángulos agudo-angostos (30º-40º) y terminan libres sin anastomosarse entre sí. Venación de orden mayor no se observa.
Material estudiado. CTES-PB 10905, 10934 a, 10941 a, b, 10946, 10948, 10951 a, b, c, d, 10959, 10958, 10957, 10958, 10964, 10967 a, b.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Entre las especies fósiles de la Patagonia argentina, Cassia sp. (Mioceno Inferior) en Fiori (1938), posee la misma forma, pero se diferencia porque contiene una sola vena media. Berry (1919) y Singewald y Berry (1922) en la paleoflora de Corocoro (Mioceno Medio) de Bolivia citan Calliandra obliqua Engelhardt, que tiene folíolos pequeños (0,7- 2,8 cm x 0,2-0,8 cm, largo por ancho) oblongos, con ápices y bases agudos y tres venas primarias divergiendo de la base, pero se diferencia de la especie aquí descrita porque las venas se unen en el ápice. Mimosa arcuatifolia Engelhardt, se diferencia por los folíolos que son linear- laceolados, más pequeños (0,3-0,4 por 0,1 cm de largo por ancho) y sólo presentan una vena media. La especie actual, Chamaecrista flexuosa es idéntica en sus caracteres morfológicos.
Hábitat y hábito de la especie actual. Chamaecrista flexuosa es una planta herbácea o subarbustiva integrante de la vegetación abierta (campos y sabanas), de regiones cálidas de América del Sur; en Argentina se la encuentra en Misiones, Corrientes y Entre Ríos (Irwin y Barneby, 1982).
Subfamilia MIMOSOIDEAE De Candolle 1825
Género Mimosa Linnaeus 1753
Especie tipo. Mimosa sensitiva Linnaeus 1753.
Mimosa cf. flagellaris Bentham 1842 Figuras 2.D, 3.N
Descripción. Folíolos dispersos, escasos, asimétricos, ovados de margen entero de 0,4 cm de largo por 0,2 cm de ancho. Ápice redondeado y base asimétrica cordada en la hemilámina más ancha y aguda en la hemilámina más angosta, peciólulo con pulvínulo de 0,01 cm desplazado hacia la hemilámina más angosta. Venación broquidódroma; vena primaria menor de 0,1 cm de grosor moderado (1,25%) y recorrido sinuoso, difusa en el ápice; venas secundarias subopuestas o alternas, 4 emergen en la hemilámina mayor y 3 en la opuesta, en ambos casos emergen con ángulos agudo - angostos (entre 30º- 40º), el espacio intercostal basal en la hemilámina más desarrollada es el más amplio. Solamente están preservadas las venas terciarias que se encuentran en el espacio marginal por fuera de las secundarias, en la hemilámina más desarrollada, nacen de las secundarias o de las terciarias y determinan ojales amplios uniéndose a las mismas o entre ellas en los tramos superiores. Venación de orden mayor no se observa.
Material estudiado. CTES-PB: 10888, 10961, 10962, 10963, 10965 a y b, 10982.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. La especie Leguminosites calliandraformis Berry 1928, del río Ñirihuau (Mioceno inferior) de la Patagonia argentina, es muy semejante en forma y tipo de venación a la especie aquí descripta, no obstante se diferencia porque en la hemilámina más desarrollada muestra un solo espacio intercostal grande que abarca casi todo el folíolo, una vena intersecundaria dicotomizada y una terciaria con terminación libre, además es levemente más grande (1 x 0,6 cm). Otras especies fósiles con folíolos pequeños menores de 1 cm asimétricos, de márgenes lisos y enteros y ápices redondeados, fueron hallados por Berry (1919) en la paleoflora de Corocoro (Mioceno Medio) de Bolivia, entre ellos se encuentran Caesalpinia sessilifoliodes Berry que se diferencia porque tiene mayor cantidad de venas secundarias (5-7) y Aeschynomene bolivianum (Engelhardt) Berry, que tiene forma obovada y base cuneada. Por la forma, número y disposición de las venas secundarias y terciarias, la especie aquí descrita, se encuentra comprendida en la variabilidad que presentan los folíolos de las hojas de la especie actual Mimosa flagellaris Bentham, de la flora argentina.
Hábitat y hábito de la especie actual. Mimosa flagellaris es una hierba rastrera o subarbusto subfructicoso de Corrientes y Entre Ríos (Argentina). Forma parte de la vegetación de sabanas y de las orillas de los montes (Burkart, 1948).
Mimosa sp. Figuras 2.E, 3.D
Descripción. Fragmentos de dos hojas compuestas pinnadas; pinnas paralelas de 1,5 cm de largo por aproximadamente 0,4 cm de ancho, muy próximas entre sí, mediando entre ellas 0,25 cm de separación. El raquis es de 0,05 cm de ancho y tiene costillas más o menos paralelas. Folíolos imbricados, aparentemente plegados sobre una hemilámina, como en "posición de sueño" (Burkart, 1948); los folíolos se insertan sobre el raquis con ángulos de 25-30º, sólo en algún sector, se puede observar que son lanceolados, coriáceos, de 0,3 cm de largo y 0,05 cm de ancho, con márgenes enteros, ápice agudo y la base, irregular, es más desarrollada y cordada del lado acroscópico con superficie glabra. Solamente se observa una vena media.
Material estudiado. CTES-PB 10888 a y b.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Calliandra obliqua Engelhardt, en Berry, 1919 (lám. 15, fig. 29), de la paleoflora de Corocoro (Mioceno Medio) de Bolivia es la única especie fósil en la que se preservó un fragmento de pinna con folíolos pequeños (0,7- 2,8 cm, longitud x 0,2- 0,8 cm, anchura) pero difiere de Mimosa sp. porque el ángulo de inserción de los folíolos sobre el raquis es mayor y por la forma y venación de los folíolos dispersos relacionados a la especie (oblongos, con ápices y bases agudos y tres venas primarias divergiendo de la base). Son varias las especies actuales de Mimosa de la flora argentina que poseen la particularidad de ser sensibles al tacto y reaccionar ante los estímulos externos plegando sus folíolos ("posición de sueño") (Burkart, 1948) y que tienen folíolos pequeños, lanceolados, coriáceos, glabros, ápices agudos, bases irregulares y 1 vena principal más destacada. Entre ellas se pueden mencionar a Mimosa strigillosa Torr. y Gray, M. polycarpa Kunth, M. farinosa Grise., M. cruenta Benth., M. paupera Benth., M. velloziana Herter y M. pigra L.
Hábitat y hábito de las especies actuales. Las especies de Mimosa aquí citadas son hierbas o arbustos, crecen desde el centro al norte de Argentina y están relacionadas a ambientes húmedos (orilla de ríos o praderas húmedas); sólo M. farinosa es xerófila.
Prosopis L. emend. Burkart 1940
Especie tipo. Prosopis spicigera Linnaeus 1767.
Prosopis sp. 1 Figuras 2.F, 3.J, K
Descripción. Folíolos dispersos, escasos, oblongo-lineales, de 1,5 cm de largo por 0,3-0,4 cm de ancho, margen liso, levemente falcados, de ápice agudo; base asimétrica, en una hemilámima es más desarrollada y semicordada y en la opuesta cuneada. Peciólulo con pulvínulo de 0,2 cm de largo. Lámina pubescente. Venación broquidódroma, vena media de grosor moderado. Venas secundarias en número de 10-12, emergen con ángulos variables agudo-moderados las basales (40º-60º) y agudo-anchos (80º) las restantes; la unión de las venas secundarias entre sí se produce muy próxima al margen, por lo que se confunde con él, dando el efecto de una vena marginal. Venación terciaria difusa, aparentemente reticulada.
Material estudiado. CTES-PB 10918 a y b.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Prosopis sp. de río Seco (Catamarca) en Anzótegui y Herbst (2004), se diferencia porque es más pequeño (0,3 - 0,6 cm x 0,1 - 0,2 cm por ancho) y la venación secundaria presenta diferente comportamiento a ambos lados de la vena media.
Las especies actuales similares a la fósil aquí descrita se encuentran en el modelo V (Sección Algarobia, serie Chilenses) establecido por Martínez (1984). Por la forma y tamaño es comparable con P. chilensis (Molina) Stuntz y P. alpataco Phil., también lo es por el comportamiento de las venas secundarias en las que sólo el primer par basal emerge con ángulos más cerrados que las restantes. Además, es semejante a P. nigra (Grises.) Hieron., P. pugionata Burkart, P. alba Grises., P. caldenia Burkart, P. juliflora (Sw.) DC., P. velutina Wooton y P. laevigata (Humb. y Bonpl. ex Willd.) Johnston.
Hábitat y hábito de las especies actuales. Las especies afines de la sección Algarobia serie Chilenses son árboles, excepto P. alpataco que es un arbusto, la mayoría pertenece a la flora argentina (excepto P. juliflora, P. velutina y P. laevigata) y crecen en las provincias del centro y norte, generalmente en regiones áridas o semiáridas (Burkart, 1976).
Subfamilia PAPILIONOIDEAE De Candolle 1825
Género Desmodium Desvaux 1813
Especie tipo. Desmodium scorpiurus (Swart) Desvaux 1813.
Desmodium cf. ascendens (Swart) De Candolle 1825 Figuras 2.J, 3.I
Descripción. Láminas foliares oblongas a elípticas de 0,6 - 1,4 cm de largo por 0,4 - 1 cm de ancho, ligeramente asimétricas; ápice retuso y base aguda a obtusa, asimétrica; margen liso y entero; peciólulo con pulvínulo muy pequeño. Venación broquidódroma. Vena primaria recta, menor de 0,1 cm de grosor moderado (1,5 - 2%) y uniforme a lo largo de toda la lámina. Venas secundarias débiles, en número de 5, opuestas a subopuestas, emergen de la primaria con ángulo agudo moderado (40 - 50º), presentan un recorrido recto a levemente sinuoso. Venas intersecundarias compuestas, presentes en la mayoría de las áreas intercostales. Las venas terciarias emergen de las venas secundarias y primarias siguiendo el modelo reticulado ortogonal. Venas cuartarias reticuladas. Venación de orden mayor no se observa.
Material estudiado. CTES-PB: 19856, 10915, 10929, 10934, 10960.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Cassia pisllypampensis Berry, de Pislepampa, (Berry,1922a) y C. rigidulifolia Engelhardt, de Corocoro (Berry, 1919) ambas del Mioceno Medio - Superior de Bolivia, tienen venación broquidódroma, pero son de mayor longitud (2 - 2,5 por 1 cm), obovadas y sésiles y la primera especie posee además ápice mucronado. Otros ejemplares fósiles de Corocoro, tales como Pithecolobium brittonianum Berry y Caesalpinia gmehlingi Engelhardt, comparten algunos caracteres (forma, tipo de base y/o ápice) pero difieren porque poseen venación camptódroma. Las folíolos en ejemplares actuales de Desmodium ascendens son similares a la especie fósil descrita, en cuanto al ápice, venación y ángulo de divergencia de las venas secundarias y sólo difieren levemente porque tienen forma obovada y son algo más anchos.
Hábitat y hábito de la especie actual. Desmodium ascendens es una planta rastrera ascendente o postrada, de bosques abiertos y sabanas. Se la encuentra desde la región central y ecuatorial de Brasil hasta el noroeste de Argentina (Burkart, 1952).
Cf. Desmodium sp. Figuras 2.G, 3.C
Descripción. Porción de lámina foliar de la parte media inferior, simétrica, de forma oblongo-linear, de márgenes lisos, de 3,5 cm de largo por 0,5 cm de ancho. Base cuneada. Venación broquidódroma. Vena primaria recta de 0,1 cm de espesor, de grosor moderado (1,25 - 2%); 5 pares de venas secundarias subopuestas, emergen con ángulos agudo-angostos de 15º - 20º, uniéndose a las superadyacentes en arcos largos y estrechos. Venas terciarias tenuemente marcadas forman un reticulado ortogonal. Lámina pubescente.
Material estudiado. CTES-PB 10973.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. No se halló en la paleoflora argentina ni boliviana ejemplares semejantes.
Desmodium cuneatum Hooker y Arnott, posee caracteres que se asemejan a los del ejemplar fósil descrito (tamaño, tipo de venación, ángulo de emisión de las venas secundarias), no obstante por la ausencia de la porción apical y por ser único, este ejemplar permanece con nomenclatura abierta.
Género Eriosema (DC.) Reichenbach 1828
Especie tipo. Eriosema rufum (H.B.K.) G. Don 1832.
Eriosema sp. Figuras 2.H, 3.M, O
Descripción. Folíolos dispersos, lanceolados, de 2 cm de largo por 0,4 cm de ancho, levemente asimétricos, de márgenes enteros y lisos. Ápice y base agudos, esta última asimétrica. Peciólulo con pulvínulo excéntrico. Venación broquidódroma. Vena primaria de grosor moderado, menor de 0,1 cm, recta. Venas secundarias tenues, alternas, emergen de la primaria con ángulos agudo - moderados (40 - 60º), se encuentran 7 - 8 venas en una hemilámina y 5 - 6 en la otra; en la hemilámina con menor cantidad de venas el espacio intercostal basal mide 0,5 cm de largo y es mayor que en el resto de los espacios intercostales, en este lugar se observa también una vena intersecundaria compuesta. Las venas terciarias describen un modelo reticulado al azar con aréolas pentagonales.
Material estudiado. CTES-PB: 10936, 10916, 10983.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Astragalus sp. en Spegazzini (1924), del Paleoceno-Eoceno y Dalbergia patagonica Berry (Berry, 1938) del Mioceno Inferior, ambas de la Patagonia argentina, tienen forma semejante a la especie aquí descrita, pero no se encuentran detalles de la venación en las descripciones ni en las ilustraciones. Calliandra jancocatana Berry, de la flora miocénica boliviana de Jankokkota (Berry, 1922b) muestra el mismo tipo de venación, incluso en la hemilámina más desarrollada, el espacio intercostal basal es de mayor tamaño, no obstante se diferencia de Eriosema sp. porque es más pequeña (1 cm de largo), las venas secundarias emergen con ángulos más abiertos y el peciólulo es excéntrico. Entre las especies actuales, es semejante a los folíolos más angostos y pequeños de las hojas de Eriosema crinitum Benth., especie citada en la flora brasileña, y de Eriosema glabrum Mart. ex Benth., en la flora argentina, porque comparte en general la forma de los folíolos, el tipo de venación y el espacio intercostal basal más desarrollado en una hemilámina. No obstante se diferencia de ambas especies porque el tamaño de la lámina es menor y de Eriosema glabrum, porque tiene mayor cantidad de venas y el espacio intercostal basal es de menor tamaño.
Hábitat y hábito del género actual. El género Eriosema es esencialmente pantropical, sus especies son hierbas o subarbustos erectos, postrados, raramente volubles, son plantas heliófilas, típicas de las sabanas (Grear, 1970).
Género Tipuana (Benth.) Bentham 1860
Especie tipo. Tipuana speciosa (Benth.) Kuntze 1898.
Tipuana tipu (Benth.) Kuntze, 1898 Figuras 2.I, 3.G, H
Descripción. Lámina foliar oblonga de 1,7 - 3 cm de largo por 0,6 - 1 cm de ancho, levemente asimétrica, de márgenes enteros y lisos. Ápice redondeado a emarginado y base aguda, peciólulo pequeño de 0,01 cm. Venación broquidódroma; vena primaria recta de grosor moderado (1,25 - 2%); venas secundarias (aproximadamente 12) de recorrido recto, subopuestas a opuestas, emergen de la vena primaria con ángulos agudos (40º - 50º), próximo al márgen se curvan y se unen a la supra-adyacente. Venas intersecundarias simples, se desprenden con el mismo ángulo que las venas secundarias, están presentes en casi todos los espacios intercostales. Venación terciaria reticulada ortogonal. Venación de orden mayor no se observa.
Material estudiado. CTES-PB: 10920, 10923 a y b, 10935, 10953, 10891.
Localidad. Río Totoral, valle del Cajón (próximo a río Cerro Colorado).
Horizonte. Formación El Morterito, Sección Media (Malizzia et al., 1990). Sección II, Aloformación Playa del Zorro (Bossi et al., 1993).
Edad. Mioceno Superior (Bossi et al., 2000).
Comparaciones. Dalbergia patagonica Berry, es semejante por el tamaño y forma, pero no se brindan detalles de la venación. Sweetia tertiraria Engelhardt, de Corocoro (Mioceno Medio) Bolivia, (Berry, 1919) es semejante en forma y tamaño, pero el tipo de venación es camptódroma y no se establecen mayores detalles acerca de las venas secundarias. Por otra parte, las características señaladas en la especie fósil aquí descrita, se encuentran en la variabilidad que presentan los folíolos de las hojas de la especie actual Tipuana tipu de la flora argentina.
Hábitat y hábito de la especie actual. Tipuana tipu es un árbol alto, inerme, corpulento, característico de la selva tucumano - boliviana (Burkart, 1952).
Discusión y conclusiones
El hábito y hábitat de las especies actuales más cercanas a los fósiles descriptos (cuadro 2), nos permiten inferir las condiciones paleoclimáticas y paleoambientales predominantes en el Mioceno Superior de la región de estudio. De ello se desprende que la vegetación fósil hallada se habría desarrollado bajo un clima en general cálido. Algunas especies, tales como Tipuana tipu, Senna cf. bicapsulares y Senna cf. obtusifolia, habitaron ambientes húmedos e integraron bosques ribereños. Otras (Prosopis sp. 1, Desmodium cf. ascendens, Eriosema sp., Chamaecrista flexuosa y Mimosa cf. flagellaris) ambientes abiertos y secos constituyendo sabanas xerófilas, siendo en su mayoría elementos herbáceo/arbustivos. Estas inferencias se ven además confirmadas por las características foliares de los taxones descriptos, pues son láminas pequeñas (nanófilas, leptófilas a micrófilas) y de márgenes enteros. En los ambientes húmedos terrestres lagunares, perilagunares y en riberas de cursos de agua de la Formación El Morterito, se encuentra, además, una abundante y rica fauna de vertebrados (flamencos, tortugas y mamíferos pequeños) acompañada por pelecípodos, gasterópodos y ostrácodos (Bossi et al, 1993). En estos ambientes habrían prosperado las plantas mencionadas (cuadro 2) que requieren elevada humedad, pero hasta el momento no se han encontrado fósiles propios de condiciones xerofíticas en la Formación El Morterito. No obstante, en unidades de edad similar, tales como las formaciones San José del Mioceno Medio y Chiquimil del Mioceno Superior (ambas del valle de Santa María) se identificaron, por medio del polen (Mautino y Anzótegui, 2002) y hojas (Anzótegui y Herbst, 2004), asociaciones indicadoras de sabanas xéricas o vegetación abierta. También en la Formación Anta, del Grupo Orán (Mioceno Temprano/ Medio) en la provincia de Salta, a partir de palinomorfos, Rivero de Dibi et al. (2001) y Quattrocchio et al. (2003), infirieron este tipo de vegetación. La Formación Chiquimil posee un registro de una abundante y diversa fauna de mamíferos (Herbst et al., 2000) que permiten inferir las condiciones paleoambientales durante su depositación. Entre los roedores, los Chinchillidae, Abrocomidae y Octodontidae están bien representados y han sido tradicionalmente asociados a ambientes cálidos, secos y hasta desérticos (Vucetich, 1986). Entre los dasipódidos, las formas de gran tamaño como Vetelia y Macrochorobates están asociados a regiones cálidas, por otra parte Paleophractus habría habitado en áreas abiertas de condiciones climáticas cálidas y semiáridas (Scillato Yané, 1982; Carlini y Scilliato Yané, 1995).
Cuadro 2. Taxones fósiles descriptos y taxones actuales más cercanos con su hábito, paleoclima y paleoasociación sugerida para los taxones fósiles de la Formación El Morterito / described fossil taxa and extant taxa with their habit, habitat, paleoclimate and paleoassociation inferred for the fossil taxa of the El Morterito Formation.
Desde el punto de vista taxonómico, se debe señalar que en los trabajos de Berry (1919, 1922 a, 1922 b, 1928, 1938) y Fiori (1938 y 1940), que contienen descripciones foliares de Fabaceae fósiles, se encuentran pocos elementos descriptivos y en la mayoría de los casos, sus ilustraciones no aportan mayores datos para realizar una adecuada comparación con los fósiles aquí estudiados. Cuando describen el modelo de venación principal, dichos autores utilizan el término camptódroma; sólo en pocas especies refieren que las venas secundarias describen arcos y se unen entre sí; de ello se infiere que en realidad se trata de venación broquidódroma y el análisis de las ilustraciones apoya también esta apreciación. Si se considera que el modelo de la venación principal de las Fabaceae, generalmente, es broquidódromo, se torna difícil su comparación. Además, es necesario resaltar que muchas de las especies descritas por los mencionados autores, han sido incluidas en el género Cassia; actualmente las especies de este género han pasado a sinonimia de Chamaecrista, Senna y Cascaronia (Zuloaga y Morrone, 1999). Coincidentemente, Senna cf. bicapsularis y Senna cf. obtusifolia descritas en este trabajo han sido comparadas con varias especies fósiles de Cassia. Por otra parte, Mimosa cf. flagellaris posee la misma forma, tamaño y tipo de venación que Leguminosites calliandraiformis, pero no fue posible su incorporación al morfogénero, porque Leguminosites Bowerbank 1840, ha sido erigido originalmente para frutos, lo mismo que Mimosites Bowerbank 1840.
Las Fabaceae f ósiles halladas en Argentina (impresiones de hojas, leños y polen) se enumeran en el cuadro 1, en éste se consignan los nombres originales de las especies en orden cronológico y la cita bibliográfica. Los granos de polen fósil de Fabaceae citados en dicho cuadro se caracterizan por ser políades, que ofrecen un diagnóstico seguro para algunos géneros de las tribus Acacieae, Mimosae e Ingae de la subfamilia Mimosoideae. Los restantes géneros de la familia contienen granos de polen del tipo tricolporado, carácter muy común en varias familias de las Magnoliofitas, excepto Margocolporites vanwijhei Germeraad, Hopping y Müller 1968 que siendo tricolporado ha sido asignado a una Caesalpinoideae (Ottone et al., 1998) por sus colpos con amplios margos.
Del an álisis del cuadro 1 se desprende que las Fabaceae se registran a partir del Paleoceno Superior, y desde ese momento hasta fines del Eoceno, sólo se han descripto 11 especies (no ocurriendo hasta el momento registros en el Oligoceno). En el Mioceno su representación llegó a unas 54 especies, incluyendo las aquí estudiadas. Si se considera que actualmente en Argentina la familia Fabaceae cuenta con aproximadamente 730 especies (Zuloaga y Morrone, 1999), el registro fósil hallado hasta el momento implicaría que durante el Neógeno se produjo una notoria diversificación de esta familia. Un antecedente similar en América del Sur que apoya esta conclusión fue registrado por Berry (1919, 1922a y 1922b), quien en Bolivia (Corocoro, Pislepampa y Jakokkota), reconoció en el Mioceno Medio (Gregory-Wodzicki et al., 1998) más de 50 taxones asignados a esta familia, aunque algunos autores actualmente no aceptan dichas determinaciones (Taylor, 1991). Coincidentemente, Caccavari y Barreda (2000) sugieren, a partir del análisis de políadas fósiles, que durante el Mioceno las Mimosoideae han tenido un alto grado de diversificación en América del Sur.
Finalmente, las especies determinadas en la Formaci ón El Morterito señalan que durante el Mioceno Superior, imperaba un clima cálido tropical a subtropical en la región, predominando ambientes secos y algunos más húmedos asociados a cursos de agua. Esta asociación florística, al igual que las de las citadas Formaciones Neógenas de los valles Calchaquíes (San José, Chiquimil y Palo Pintado), estaría también integrando la "Paleoflora Subtropical Neógena" del esquema paleofitogeográfico de Hinojosa y Villagran (1997) e Hinojosa (2005).
La elevada representaci ón de integrantes de Fabaceae durante el Mio-Plioceno de Argentina y Bolivia indica que los mismos formaron una parte importante de la Paleoflora Subtropical Neógena. Los resultados obtenidos de los estudios paleoflorísticos en el Neógeno del noroeste argentino permiten incorporar por primera vez esta región en el esquema paleofitogeográfico del sur de América del Sur y ampliar el área geográfica de la Paleoflora Subtropical Neógena al noroeste de Argentina.
Agradecimientos
A la Secretaría de Ciencia y Técnica de la Universidad Nacional del Nordeste y al Consejo Nacional de Investigaciones de Ciencia y Técnica, por el apoyo financiero. A L. Simón que realizó las ilustraciones. A P.P. Medina por su inestimable ayuda en los viajes de campaña. Al Comité Editor de la Revista y a los árbitros, por las sugerencias que realizaron aportes significativos a este trabajo.
Bibliografía
1. Acevedo, T.L., Mautino, R.L., Anzótegui, L.M. y Cuadrado G.A. 1997. Estudio palinológico de la Formación Palo Pintado (Mioceno Superior), Provincia de Salta, Argentina. Parte II: Esporas. Geociencias 2: 112-120. [ Links ]
2. Acevedo, L.T., Anzótegui, L.M. y Mautino, L.R. 1998. Análisis palinológico preliminar de la Formación Palo Pintado (Mioceno Superior), en Quebrada del Estanque. Salta, Argentina. Comunicaciones Científicas y Técnicas de la Universidad Nacional del Nordeste. Resistencia: 47-49 pp. [ Links ]
3. Anzótegui, L.M. 1998. Hojas de angiospermas de la Formación Palo Pintado, Mioceno Superior, Salta, Argentina. Parte 1: Anacardiaceae, Lauraceae y Moraceae. Ameghiniana 35: 25-32. [ Links ]
4. Anzótegui, L.M. 2002a. Fabaceae en la Formación San José, Mioceno Medio, Tucumán, Argentina. 8º Congreso Argentino de Paleontología y Bioestratigrafía (Corrientes), Resúmenes: p. 23. [ Links ]
5. Anzótegui, L.M. 2002b. Paleocomunidades vegetales de tres Formaciones del Mioceno del noroeste argentino. 8º Congreso Argentino de Paleontología y Bioestratigrafía (Corrientes), Resúmenes: p. 24. [ Links ]
6. Anzótegui, L. 2002c. Hojas de Myrtacae en la Formación San José (Mioceno Medio) Provincia de Tucumán, Argentina. Ameghiniana 39: 321-330. [ Links ]
7. Anzótegui, L.M. 2004. Megaflora de la Formación Chiquimil, (Mioceno Superior), en los valles de Santa Maria y Villavil, provincias de Catamarca y Tucumán, Argentina. Ameghiniana 41: 303-314. [ Links ]
8. Anzótegui, L.M. y Cuadrado, G.A. 1996. Palinologia de la Formación Palo Pintado, Mioceno Superior, Provincia de Salta, Repúblia Argentina. Parte I: Taxones nuevos. Revista Española de Micropaleontología 28: 77-92. [ Links ]
9. Anzótegui, L.M. y Cristalli, P. 2000 Hojas de Malvaceae en el Neógeno de Argentina y Brasil. Ameghiniana 37: 169-180. [ Links ]
10. Anzótegui, L.M. y Ferraro, L.I. 2000. Líquenes folícolas en la Formación San José (Mioceno Medio) Tucumán, Argentina. Geociencias 5: 256. [ Links ]
11. Anzótegui, L.M. y Garralla, S.S. 1985. Estudio palinológico de la Formación Paraná (Mioceno Superior), (Pozo Josefina), Provincia de Santa Fe, Argentina. 1º parte: Descripciones sistemáticas. Facena 6: 101-177. [ Links ]
12. Anzótegui, L.M. y Herbst, R. 2004. Megaflora (hojas y frutos) de la Formación San José (Mioceno Medio) en río Seco, departamento Santa María, provincia de Catamarca, Argentina. Ameghiniana 41: 423-436. [ Links ]
13. Anzótegui, L.M., Horn, Y. y Herbst, R. 2004. Contribución a la paleoflora de la Formación Andalhuala (Plioceno inferior?), Tucumán, Argentina. 11 Reuniao de Paleobotánicos e Palinólogos. (Gramado), Resúmenes: 8. [ Links ]
14. Ash, A., Ellis, B., Hickey, L.J., Johnson, K., Wilf, P. y Wing, S. 1999. Manual of Leaf Architecture- morphological description and categorization of dicotyledonous and net-veined monocotyledonous angiosperms by Leaf Architecture Working Group. Smithsonian Institution, Washington, 65 pp. [ Links ]
15. Barreda, V.D. y Caccavari, M. 1992. Mimosoideae (Leguminosae) ocurrences in the Early Miocene of Patagonia (Argentina). Paleogeography, Paleoclimatology and Paleoecology 94: 243-252. [ Links ]
16. Bentham, G. 1842. Mimosa flagellaris. En: Hooker's Journal of Botany 4, 372 pp. [ Links ]
17. Bentham, G. 1860. Tipuana. En: Journal of the Linnean Society Botany 4. Suppl., 72 pp. [ Links ]
18. Berry, E. 1919. Fossil plants from Bolivia and their bearing upon the age of uplift of the eastern Andes. Proceedings of the United States National Museum 54: 103-164. [ Links ]
19. Berry, E. 1922a. Pliocene fossil plants from eastern Bolivia. En: E. B. Mathews (ed.), Contributions to the Paleobotany of Perú, Bolivia and Chile. The Johns Hopkins University, Studies in Geology 4: 145-185. [ Links ]
20. Berry, E. 1922b. Late tertiary plants from Jancocata, Bolivia. En: E. B. Mathews (ed.), Contributions to the Paleobotany of Perú, Bolivia and Chile. The Johns Hopkins University, Studies in Geology 4: 205-220. [ Links ]
21. Berry, E.W. 1925. Miocene flora from Patagonia. Johns Hopkins University Studies in Geology 6: 183-223. [ Links ]
22. Berry, E.W. 1928. Tertiary fossil plants from the Argentine Republic. Proceedings of the United States National Museum 73: 1-27. [ Links ]
23. Berry, E.W. 1938. Tertiary flora from the Río Pichileufú, Argentina. Geology Society American. Special Paper 12: 1-149. [ Links ]
24. Bossi, G.E., Muruaga, C.M., Sanuaga, J.G., Hernando, A. y Ahumada A.L. 1993. Geología y estratigrafía de la Cuenca Neógena Santa María-Hualfin (Deptos. Santa María y Belén, provincia de Catamarca). 12º Congreso Geológico Argentino de Exploración de Hidrocarburos (Mendoza), Actas: 2: 156-165. [ Links ]
25. Bossi, G.E., Vides, M.E., Georgieff, S.M., Muruaga, C. M. e Ibáñez, L.M. 2000. Análisis de las paleocorrientes y de la varianza de los componentes a tres niveles, Neógeno del valle del Cajón, Catamarca, Argentina. Revista de la Asociación Argentina de Sedimentología 7: 23-47. [ Links ]
26. Bossi, G.E., Georgieff, S.M., Gavriloff, I.J., Ibáñez, L.M. y Muruaga, C.M. 2001. Cenozoic evolution of the intramontane Santa Maria basin, Pampean Ranges, Northwestern Argentina. Journal of South American Earth Sciences 14: 725-734. [ Links ]
27. Bowerbank, J.S., 1840. A Hystory Fossil Fruit and Seeds of the London Clay. John Van Voorst, London, 144 pp. [ Links ]
28. Brea, M., Aceñolaza, P.G. y Zucol, A.F. 2001. Estudios paleoxilológicos en la Formación Paraná, Entre Rios, Argentina. Asociación Paleontológica Argentina, Publicación Especial 8: 7-17. [ Links ]
29. Burkart, A. 1940. Materiales para una monografía del género Prosopis (Leguminosae). Darwiniana 4: 57-128. [ Links ]
30. Burkart, A. 1948. Las especies de Mimosa de la Flora Argentina. Darwiniana 8: 9-231. [ Links ]
31. Burkart, A. 1952. Las Leguminosas argentinas, silvestres y cultivadas. Ed. Acme, 2º ed., Buenos Aires, 104 pp. [ Links ]
32. Burkart, A. 1976. A monograph of the genus Prosopis. Journal of the Arnold Arboretum 57: 219-249. [ Links ]
33. Cabrera, A.L. 1976. Regiones fitogeográficas argentinas. Enciclopedia argentina de agricultura y ganadería, Ed. Acme, 2º ed., Buenos Aires, 85 pp. [ Links ]
34. Caccavari, M. y Anzótegui, L.M. 1987. Polen de Mimosoideae (Leguminosae) de la Formación Ituzaingó, Plioceno Superior de Corrientes, Argentina. 4º Congreso Latinoamericano de Paleontología Bolivia (Bolivia), Actas: 443-458. [ Links ]
35. Caccavari, M.A y Barreda, V. 2000. A new calymate mimosoid polyad from the Miocene of Argentina. Review of Palaeobonaty and Palynology 109: 197-203. [ Links ]
36. Caccavari, M.A. y Guler, V. 2002. Una nueva especie de Acaciapollenites del Neógeno de la Argentina. 8º Congreso Argentino de Paleontología y Bioestratigrafía (Corrientes), Resúmenes: p. 77. [ Links ]
37. Carlini, A. y Scillato Yané, G. 1995. Chorobates recens (Xenarthra, Dasypodidae) y un análisis de la filogenia de los Euphractini. Revista Museo La Plata (Nueva Serie) Paleontología 9: 225-338. [ Links ]
38. Cronquist, A., Takhtajan, A. y Zimmermann, W. 1966. On the higher taxa of Embryobionta. Taxon 15: 129-143. [ Links ]
39. De Candolle, A.P. 1825. Prodromus Systematics Naturalis. Regni vegetabilis, Pars 2. Trewttei y Würtz (ed.), 644 pp. [ Links ]
40. Desvaux, N.A., 1813. Desmodium scorpiurus. Journal de Botanique 1, 122 pp. [ Links ]
41. Don, G. 1832. A general History of the Dichlamydeous Plants 2: 347. [ Links ]
42. Fiori, A. 1938 Filliti terziarie della Patagonia II. Filliti del Río Ñirihuau. Giornale di Geologia 13: 1-27. [ Links ]
43. Fiori, A. 1940. Filliti terziarie della Patagonia III. Filliti di Chenque-Niyeu. Giornale di Geologia S2, 14: 93-133. [ Links ]
44. Germeraad, J.J., Hopping, G.A. y Muller, J. 1968. Palynology of Tertiary sediments from tropical areas. Review of Palaeobotany and Palynology 6: 189-348. [ Links ]
45. Georgieff, S.M., Herbst, R., Esteban, G.I.y Nassif, N. 2004. Análisis paleoambiental y registro paleontológico de la Formación Desencuentro (Mioceno Superior), Alto de San Nicolás, La Rioja, Argentina. Ameghiniana 41: 45-56. [ Links ]
46. Grear, J.W. 1970. A revision of the American species of Eriosema (Leguminosae -Lotoideae). Memories the New York Botanical Garden 20: 1-98. [ Links ]
47. Greene, S.W. 1899. Chamaecrista flexuosa. Pittonia 4, 27 pp. [ Links ]
48. Gregory-Wodzicki, K.M., Mcintosh, W.C. y Velasquez, K. 1998. Climatic and tectonic implications of the Miocene Jakokkota flora, Bolivian Altiplano. Journal of South American Earth Sciences, 6: 533-560. [ Links ]
49. Guler, M.V., Guerstein, M.E. y Quattrocchio, M.E. 2001. Palinología del Neógeno de la Perforación Cx-1, Cuenca del Colorado, Argentina. Revista Española de Micropaleontología 33: 183-204. [ Links ]
50. Herbst, R., Anzótegui, L.M. y Jalfin, G. 1987. Estratigrafía, paleoambientes y dos especies de Salvinia Adanson (Filicopsida) del Mioceno Superior de Salta, Argentina. Facena 7: 15-42. [ Links ]
51. Herbst, R., Anzótegui, L.M., Esteban, G., Mautino, L. R., Morton, L.S., y Nassif, N. 2000. Síntesis paleontológica del Mioceno de los valles Calchaquíes, noroeste argentino. En: F. Aceñolaza y R. Herbst R. (eds.), El Neógeno de Argentina. Instituto Superior de Correlación Geológica, Serie Correlación Geológica (Tucumán) 14: 263-288. [ Links ]
52. Herendeen, P.S y Dilcher, D.L. 1992. Advances in Legume Systematics. The fossil Record. Part 4. Royal Botanic Gardens, Kew, 326 pp. [ Links ]
53. Hickey, LJ. 1974. Clasificación de la arquitectura de las hojas de Dicotiledóneas. Boletín de la Sociedad Argentina de Botánica 16: 1-26. [ Links ]
54. Hickey, L.J. 1979. A revised classification of the architecture of Dicotyledonous leaves. En: C.R. Melcalfe y L. Chalk (eds.), Anatomy of the Dicotyledons, Vol. 1. 2da. ed., Clarendon Press. Oxford: 25-39 pp. [ Links ]
55. Hinojosa, F. L. 2005. Cambios climáticos y vegetacionales a partir de paleofloras cenozoicas del sur de Sudamérica. Revista Geológica de Chile 32: 95 -115. [ Links ]
56. Hinojosa, F.L. y Villagran, C. 1997. Historia de los bosques del sur de Sudamérica, I: antecedentes paleobotánicos, geológicos y climáticos del Terciario del cono sur de América. Revista Chilena de Historia Natural 70: 225 -239. [ Links ]
57. Irwing, H. S. y Barneby, R.C. 1982. The American Cassinae. A synoptical revision of Leguminosae tribe Cassieae, subtribe Cassiinae. Memorial New York Botanical Garden, 35: 1-918. [ Links ]
58. Kuntze, J.E. 1898. Tipuana tipu. En: Reveue Gènèrale de Botanique, 3. 2, 72 pp. [ Links ]
59. Kurkbride, J.H., Gum, Ch.R. y Wetzman, A.L. 2003. Fruit and seed of genera in the subfamily Faboideae (Fabaceae). United States Department of Agriculture, 1208 pp. [ Links ]
60. Lindley, J. 1830. An introduction to the natural system of botany. Logman, Res, Orme, Brow and Green, 148 pp. [ Links ]
61. Linnaeus, C. 1753. Species plantarum. Holmiae, Stockholm 2 vol., 1200 pp. [ Links ]
62. Linnaeus, C. 1767. Mantissa plantarum. Specierum editions 2. Holmiae, Stockholm, 327 pp. [ Links ]
63. Lutz, A.I. 1979. Maderas de angiospermas (Anacardiaceae y Leguminosae) del Plioceno de la provincia de Entre Ríos, Argentina. Facena 3: 39-63. [ Links ]
64. Lutz, A.I. 1980. Entrerrioxylon victoriensis nov. gen. et sp. (Leguminosae) del Plioceno de la provincia de Entre Ríos, Argentina. Facena 4: 21-29. [ Links ]
65. Lutz, A.I., 1987. Estudio anatómico de maderas terciarias del valle de Santa María (Catamarca-Tucumán), Argentina, Facena 7: 125-144. [ Links ]
66. Lutz, A.I. 1991. Descripción anatómica de Mimosoxylon sp. del Plioceno (Formación Ituzaingó) de la provincia de Corrientes, Argentina. Revista de la Asociación de Ciencias Naturales del Litoral 22: 3-10. [ Links ]
67. Malizia, D.C., Strecker, M., Herbst, R. y Carrión, M. 1990. La Formación El Morterito (Neógeno de Sierras Pampeanas); características sedimentológicas y paleogeográficas. 3º Reunión Argentina de Sedimentología, Actas: 175-180 pp. [ Links ]
68. Martinez, L.C. A y Lutz, A.I. 2004. Xilotaflora de la Formación Chiquimil (Mioceno Superior) en el Valle de Santa María, Catamarca, Argentina. Ameghiniana, Suplemento Resúmenes 41. [ Links ]
69. Martinez, L.C.A y Lutz, A.I. Nuevo género y especie de liana del Mioceno Superior (Formación Palo Pintado), provincia de Salta, Argentina. Ameghiniana (en prensa). [ Links ]
70. Martinez, S. 1984. Arquitectura foliar de las especies del género Prosopis. Darwiniana 25: 279-297. [ Links ]
71. Mautino, L.R. 2002. Cactaceae en la Formación San José (Mioceno Medio), Tucumán, Argentina. 8º Congreso Argentino de Paleontologia y Bioestratigrafía, Resúmenes: p. 26. [ Links ]
72. Mautino, R.L., Anzótegui, L.M. y Herbst, R. 1997. Análisis palinológico de la localidad Nacimientos de Abajo, Neógeno, en la Sierra de Hualfin, Departamento Belén, Catamarca, Argentina. Geociencias 2: 121-127. [ Links ]
73. Mautino, L.R. y Anzótegui, L.M. 1998. Palinología de la Formación Chiquimil, en la localidad Vallecito (Mioceno Superior). Parte 1: Esporas, especies nuevas. Ameghiniana 35: 227-233. [ Links ]
74. Mautino, L.R. y Anzótegui, L.M. 2000. Esporas del Mioceno y retrabajadas del Mesozoico en la Formación Chiquimil, Vallecito, Provincia de Catamarca. Ameghiniana 37: 13 - 22. [ Links ]
75. Mautino, LR. y Anzótegui, L.M. 2002. Palinología de la Formación Chiquimil (Mioceno Superior) Vallecito, provincia de Catamarca. Parte 2. Polen. Ameghiniana 39: 257-270. [ Links ]
76. Mautino, L.R. y Anzótegui, L.M., 2004. Clorophyta from the Middle and Late Miocene Calchaquí valleys, northwestern argentina. Taxonomy, paleoecology and biostratigraphic considerations. 9º International Palynological Congress Polen (Granada, España), Abstracts: p. 27. [ Links ]
77. Mautino, L.R., Cuadrado, G.A. y Anzótegui, L.M. 2004. Diversidad y significado evolutivo del polen de Malvaceae (Mioceno Medio) en la Formación San José, Tucumán, Argentina. Revista Española de Micropaleontología 36: 467-483. [ Links ]
78. Meléndez, B. 1998. Tratado de Paleontología, Tomo 1. 3º ed., Consejo Superior de Investigaciones Científicas, Madrid, 457 pp. [ Links ]
79. Miller, P. 1754. The gardeners dictionary (Abridged). John and James Rimington 3: 999-1582. [ Links ]
80. Moench, C. 1794. Methodii Plantarum Horticole. Botanica Marburguensis: 272 pp. [ Links ]
81. Muruaga, C.M. 2001. Estratigrafía del Miembro El Jarillal (Formación Chiquimil, Mioceno Superior) Sierra de Hualfín, provincia de Catamarca. Acta Geológica Lilloana 18: 265-280. [ Links ]
82. Ottone, E.G, Barreda, V.D y Perez, D.E. 1998. Basin evolution as reflected by Miocene palynomorphs from the Chinches Formation, Frontal Cordillera (32ºS), San Juan Province, Argentina. Revista Española de Micropalentología 30: 35-47. [ Links ]
83. Polhill, R.M. y Raven, P.H. 1981. Advances in Legume Systematics. Parts 1 and 2. Royal Botanic Gardens. Ministry of Agriculture, Fisheries and Food, Kew, 425 pp. [ Links ]
84. Quattrochio, M., Durango de Cabrera, J. y Galli, C. 2003. Formación Anta (Mioceno temprano/Medio), subgrupo Metán (Grupo Orán), en el río Piedras, Provincia de Salta. Datos palinológicos. Revista de la Asociación Geológica Argentina 58: 117-127. [ Links ]
85. Reichenbach, H.G.L. 1828. Conspectus Regni Vegetabilis 150 pp. [ Links ]
86. Rivero de Dibi, L., Rodríguez de Sarmiento, M y Durango de Cabrera, J. 2001. Esporas de hongos de la Formación Anta (Grupo Orán) Mioceno Medio, provincia de Salta, Argentina. Acta Geológica Lilloana 18: 257-264. [ Links ]
87. Rouxbourgh, W. 1832. Senna bicapsularis en Flora Indica 2. 2º ed. Willam Carey, 342 pp. [ Links ]
88. Scillato Yané, G. 1982. [ Los Dasypodidae (Mammalia, Edentata) del Plioceno y Pleistoceno de Argentina. Tesis doctoral . Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 159 pp. Inédita.]. [ Links ]
89. Singewald, J.T y E.W. Berry, 1922. The Geology of the Corocoro Copper district of Bolivia. The Johns Hopkins University. Studies in Geology 1: 85-117. [ Links ]
90. Spegazzini, C. 1924. Sobre algunas impresiones vegetales Eocénicas de Patagonia. Comunicaciones del Museo Nacional de Historia Natural de Buenos Aires 2: 95-107. [ Links ]
91. Starck, D. y Anzótegui, L.M. 2001 The late climatic change persistence of a climatic signal through the orogenic stratigraphic record in northwestern of Argentina. Journal of South American Earth Sciences 14: 763-774. [ Links ]
92. Taylor, D.W. 1991. Paleobiogeographic relationship of Andean Angiosperms of Cretaceous to Pliocene age. Palaeogeography, Palaeoclimatology, Palaeoecology 88: 69-84. [ Links ]
93. Turner, J.C.M. 1973. Descripción geológica de la Hoja 11d, Laguna Blanca, provincia de Catamarca. (Carta Geológico-Económica de la República Argentina, escala 1:200.000). Servicio Nacional Minero Geológico, Boletín 142, 72 pp. [ Links ]
94. Vucetich, M.G. 1986. Historia de los roedores y primates en Argentina: su aporte al conocimiento de los cambios ambientales durante el cenozoico. 4º Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza) Actas 2: 157-165. [ Links ]
95. Zuloaga, F.O. y Morrone, O. 1999. Catálogo de la Plantas Vasculares de la República Argentina II. Monographs in Systematic Botany from the Missouri Botanical Garden, Vol. 74. Missouri Botanical Garden Press. Saint Louis, 1269 pp. [ Links ]
Recibido: 3 de junio de 2005.
Aceptado: 19 de julio de 2006.

