










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.1 Buenos Aires ene./mar. 2007
A new araucarian fossil species from the Paleogene of southern Argentina
Carolina Panti1, Silvia N. Césari1, Sergio A. Marenssi2 and Eduardo B. Olivero3
1Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Av. Ángel Gallardo 470, 1405 Buenos Aires, Argentina. caropanti@macn.gov.ar, scesari@macn.gov.ar
2Instituto Antártico Argentino, Cerrito 1248, 1010 Buenos Aires, Argentina. smarenssi@dna.gov.ar
3Centro Austral de Investigaciones Científicas, Bernardo A. Houssay 200, CC. 92, 9410 Ushuaia, Tierra del Fuego, Argentina. emolivero@ciudad.com.ar
Abstract. A new Araucariaceaea from the Paleogene of Tierra del Fuego Province, Argentina is described. Araucaria pararaucana sp. nov. has imbricate, ovate-lanceolate leaves with an acute apex, characterized by the presence of discontinuous and longitudinally oriented stomatal rows separated by epidermal cells, which may be slightly sinuous. Stomatal guard cell surface is granular. These fossil leaves are compared with the extant sections Bunya, Intermedia and Araucaria. Their anatomical characters along with the external morphology suggest a close affinity with the extant southern Patagonian species A. araucana (Molina) K. Koch. The record of this new late Eocene species provides more evidence for a probable unique representation of Section Araucaria in the area during the Tertiary.
Resumen. Una nueva especie fósil de Araucaria del Paleógeno de Tierra del Fuego, Argentina. Se describe una nueva especie de Araucariaceae del Paleógeno de Tierra del Fuego, Argentina. Araucaria pararaucana sp. nov. se caracteriza por sus hojas imbricadas, ovado-lanceoladas con un ápice agudo. Sus caracteres anatómicos permiten una estrecha comparación con las actuales secciones Bunya, Intermedia y Araucaria. La presencia de hileras discontinuas de estomas orientados longitudinalmente, separadas por células epidérmicas que pueden ser algo sinuosas, la superficie granular de las células de cierre, junto con la morfología externa nos permiten compararla con la especie Araucaria araucana (Molina) K. Koch, actualmente restringida a la Patagonia austral. Esta especie del Eoceno tardío es una nueva evidencia de la exclusiva representación de la Sección Araucariadurante el Terciario en el área.
Key words. Gymnosperms; Araucariaceae; Araucaria; Cuticle; Paleogene; Argentina
Palabras clave. Gimnospermas; Araucariaceae; Araucaria; Cutícula; Paleógeno; Argentina
Introduction
Araucariaceae is a coniferous family now confined to the Southern Hemisphere, where the genus Araucaria de Jussieu is a characteristic member of the South American forests from southern Brazil and western Patagonia (represented by A. angustifolia (Bertol.) O. Kuntze and A. araucana (Molina) K. Koch, respectively), New Caledonia, Australia, Norfolk Island and New Guinea. The fossil record of Araucaria leaves in southern South America is scarce and includes Jurassic, Cretaceous and Tertiary species (Berry, 1928, 1938; Calder, 1953; Del Fueyo and Archangelsky, 2002; Menéndez and Caccavari, 1966).
The fossil specimens described here were collected from Tertiary deposits at Sloggett Bay in Tierra del Fuego Province, Argentina (figure 1). The presence of fossil plants in the Sloggett Formation was reported originally by Andersson (1907). Palynological assemblages recovered from this unit suggest a Late Eocene-Early Oligocene age for the sequence (Olivero et al., 1998). Podocarpaceae, Nothofagaceae and Proteaceae dominate the palynoflora with subordinated ferns of the Lophosoriaceae, Dicksoniaceae, Schizaeaceae, Pteridaceae, Athyriaceae, Polypodiaceae and Cyatheaceae (Olivero et al., 1998). Also, pollen grains of Araucariacites australis Cookson were identified, indicating the presence of the Araucariaceae (Olivero et al., 1998).
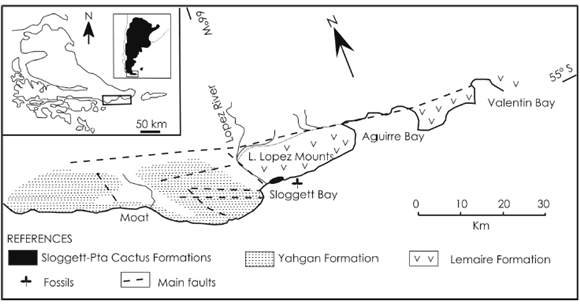
Figure 1. Location map (after Olivero et al., 1998) / mapa de ubicación (tomado de Olivero et al., 1998).
Material and methods
The studied specimens were collected from the Sloggett Formation at Sloggett Bay, the type locality of the stratigraphic unit (Caminos et al., 1981). Olivero et al. (1998) recognized two members in the sequence; the lower Gris Member consists of massive carbonaceous mudstones cut by conglomerate and pebbly sandstone channels. The plant remains are found in thin, black banks with abundant mudstone matrix inside this member, which represents a fluvial system with humid and vegetate areas (Caminos et al., 1981; Zanettini and Zapettini, 1988; Olivero et al., 1998). Besides the araucarian remains, the paleobotanical association includes angiosperm leaves that will be analysed in a forthcoming paper. The upper Bayo Member is a major complex of lenticular conglomerates and sandstones grading laterally and vertically to heterolithic mudstones and coal measures (Olivero et al., 1998).
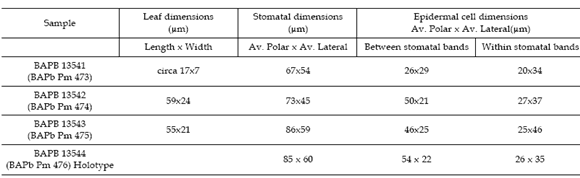
The organically preserved leaf remains were examined under light microscope and with scanning electron microscope (SEM). Cuticles were prepared by soaking leaf fragments in nitric acid; small amounts of 10% nitric acid were successively added until the fragments were cleared, followed by sodium hypochlorite (20%). Material for SEM observation was sputter-coated with gold-palladium (40%- 60%) and examined in a microscope Phillips XL 30 at the Museo Argentino de Ciencias Naturales B. Rivadavia. Twenty stomata and epidermal cells were measured for each sample and the averages were calculated (table 1).
Table 1. Leaf dimensions and internal cuticular features for the samples analyzed / dimensiones de las hojas y caracteres cuticulares internos de las muestras analizadas.
All specimens were photographed using a Coolpix 4500 camera adapted to a Nikon stereomicroscope and an Olympus microscope. The specimens studied are stored in the Paleobotanical Collection of the Museo Argentino de Ciencias Naturales B. Rivadavia (BAPb) together with the thin sections (BAPbPm).
Systematic paleontology
Family ARAUCARIACEAE Henkel and Hochst., 1865
Genus Araucaria de Jussieu, 1789
Section Araucaria Wilde and Eames, 1952
Type species. Araucaria araucana (Molina) K. Koch, 1873.
Araucaria pararaucana sp. nov. Figures 2.A, C-G and 3
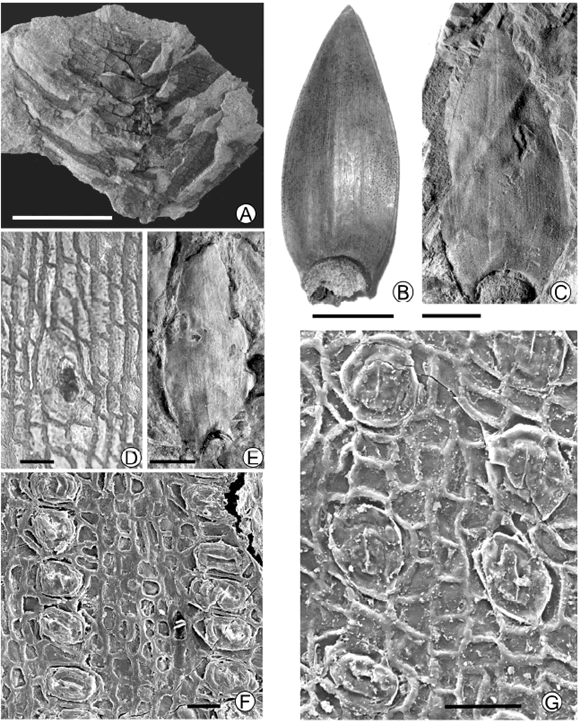
Figure 2. A, C-G, Araucaria pararaucana sp. nov.; A, Imbricate leaves in tight helix / hojas imbricadas en espiral apretada, BAPb 13541. B, Leaf of Araucaria araucana / hoja de Araucaria araucana. C, Complete leaf / hoja completa, BAPb 13542. D, Surface of cuticle with few stomata and slightly sinouous epidermal cells / superficie de la cutícula con pocos estomas y células epidérmicas algo sinuosas, BAPb Pm 473C. E, Holotype / holotipo, BAPb 13544. F, surface with densely packed horizontally oriented stomata / superficie con estomas densamente distribuidos y orientados horizontalmente, BAPb Pm 475. G, One surface of cuticle with longitudinally oriented stomata/ superficie de la cutícula con estomas con orientación longitudinal, BAPb 13541. Scale bar / escala gráfica, A-C, E = 1 cm, D, F-G = 50 µm.
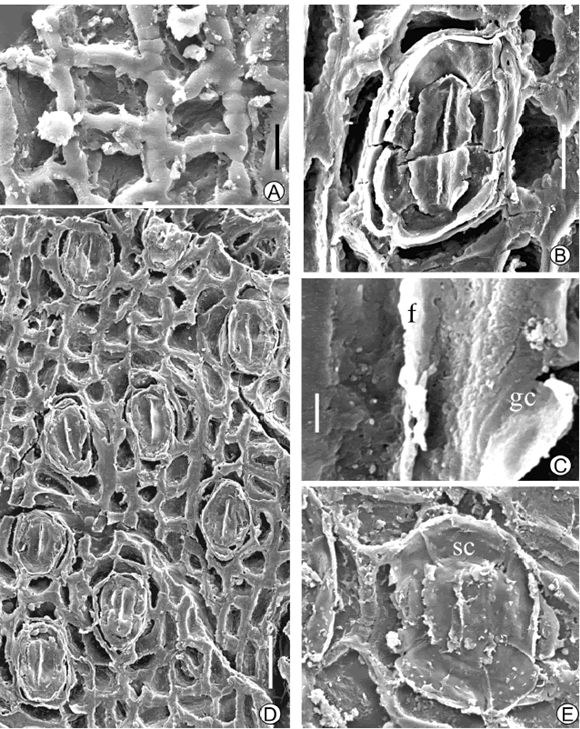
Figure 3. Araucaria pararaucana sp. nov., A, Detail of slightly sinuous wall of epidermal cells / detalle de la pared ligeramente sinuosa de las células epidérmicas, BAPb 13541. B, Oval stomata / estoma oval, BAPb 13544. C, Detail of guard cells of stomata, f: flange, gc: guard cell / detalle de las células de cierre del estoma, f: reborde, gc: célula de cierre, BAPb 13544. D, Subrounded stomata / estoma subredondeado, BAPb 13541. E, General view of discontinuous stomatal rows / vista general de las hileras discontinuas de estomas, BAPb 13544. Scale bar / escala gráfica, A= 4 µm, B, D= 10 µm, C= 1 µm, E= 50 µm.
Holotype. BAPb 13544 (BAPbPm 476).
Other material studied. BAPb 13541 (BAPbPm 473), BAPb 13542 (BAPbPm 474), BAPb 13543 (BAPbPm 475).
Etymology. From Latin par: equal, like.
Type locality and horizon. Sloggett Bay, Tierra del Fuego Province, Argentina. Sloggett Formation, Gris Member, Late Eocene-Early Oligocene.
Diagnosis. Leaves imbricate, ovate to lanceolate, with acute apex and length/width ratio about 2.5. Leaves amphistomatic, more stomata occur on one surface. Stomata in parallel, discontinuous rows; elliptical to sub-rounded, most oriented parallel to leaf margin, oblique and horizontally oriented stomata rare. Subsidiary cells four; five and six occur. Cuticle of the guard cells is granular. Epidermal cells are rectangular to quadrangular between stomata rows, irregular around stomata; outlines slightly sinuous.
Description. The holotype BAPb 13544 corresponds to a complete, ovate-lanceolate leaf with a broad base (figure 2.E). It seems to have been thick and coriaceous. Leaves are amphistomatic, with more stomata in one surface (in the fossil specimens the abaxial and adaxial surfaces could not be differentiated). Stomata are arranged in parallel and discontinuous rows (figure 3.D), they are elliptical to sub-rounded in shape (figure 3.B), sometimes with perpendicular orientation of the stomata to the leaf margin. Cuticle of the guard cells surface is granular (figure 3.C). The common subsidiary cell number is four, but five and six were observed. Epidermal cells are rectangular to quadrangular between stomata rows and irregular around stomata. Their outlines are slightly sinuous.
A second smaller specimen (BAPb 13543) has a length/width ratio of 2.6. This specimen shows stomata densely arranged in one surface. Oblique and horizontally oriented stomata occur less frequently in some leaf fragments (figure 2.F). The complete leaf of sample BAPb 13542 (figure 2.C) is similar to the holotype and preserves the acute apex, with a length/width ratio of 2.4.
All the specimens analyzed are detached leaves except the sample BAPb 13541, which shows imbricate leaves, arranged in a tight helix (figure 2.A). They are preserved without the apex. At high magnification the stomata can be observed arranged in rows parallel to leaf margins. Stomata are sparser on one surface (figure 2.D) and subsidiary cells have a different shape giving them a more sub-rounded shape (figure 3.E). They are mostly oriented parallel to the leaf margin but oblique and horizontally oriented stomata are observed less frequently (figure 2.G). The most common subsidiary cell number is four, but five and six were observed as in the holotype, and result from the division of both lateral and polar subsidiaries (Stockey and Ko, 1986). Epidermal cells are rectangular to quadrangular between stomata rows and irregular around stomata as in the holotype. Their outlines are slightly sinuous (figure 3.A) but usually these margins are masked by the cuticle thickness that partially covers the inner wall surface.
Discussion and comparisons
Wilde and Eames (1952) classified the genus Araucaria into four extant Sections: Araucaria (= Columbea), Bunya, Eutacta and Intermedia. This classification, based on morphological characters, agrees with the rbcL phylogeny proposed by Setoguchi et al. (1998), who suggested that the number of cotyledons, position of the pollen cone and cuticular micromorphologies are phylogenetically informative. Among the cuticular characters, stomatal orientations to long axis of leaf and stomatal distribution have resulted useful to split Section Eutacta from the other sections as a clade (Setoguchi et al., 1998). That section is characterized by stomata mostly oblique or perpendicular, distributed in two bands or groups and the prominent polar extensions of the guard cells. The other three sections constitute another clade (with stomata in discontinuous rows), which divides into two clades, one of them including the two South American species (A. araucana and A. angustifolia). Moreover, an analysis of the evolutionary trend in stomatal orientation showed that parallel orientation arose in the ancestor of sections Araucaria, Bunya and Intermedia (Setoguchi et al., 1998) and is also present in Wollemia adult leaves (Chambers et al., 1998).
Therefore, our species characterized by discontinuous rows of stomata parallel to the leaf margin, can be compared with the sections Bunya, Intermedia and Araucaria (adult leaves of Wollemia differ in having prominent extensions of the guard cells). According to the descriptions of Stockey and Ko (1986), this clade shares the oval to rounded outline of the subsidiary cells and the absence of prominent polar extensions in the guard cells. Moreover, the analysis of the different species seems to suggest that sinuous epidermal cells are present in all these sections.
Although mature leaves of Section Bunya (A. bidwillii) are arranged radially around the branchlet and often imbricate, they are more elongate than our species. Cuticle of this Section shares similarities with Araucaria pararaucana sp. nov. such as the oval to rounded outline of the subsidiary cells but differs in the more sinuous epidermal cells and the more reticulate guard cell surface. Moreover, stomata are predominantly oriented parallel to the long axis of the leaf (Stockey and Ko, 1986).
In the same way, the species included in the Section Intermedia are characterized by prominent sinuous epidermal cell outlines and clearly reticulate guard cells surfaces (Stockey and Ko, 1986). Besides, adult leaves of that Section are lanceolate, narrow and have a similar number of stomata in both surfaces.
Our species is referred to section Araucaria, which includes large, lanceolate and imbricate leaves, represented by the extant species Araucaria araucana and A. angustifolia. A. araucana seems to be the most similar in both external and internal characters to the studied leaves. Silba (1986) characterized A. araucana leaves as scale-like, persistent on the trunk even to the base, ovate-lanceolate, 30-50 mm long by 8-25 mm wide, a surface marked with longitudinal lines, and bearing stomata on both faces (more in one surface). The extant and fossil species share the arrangement of the leaves in a tight helix and the same leaf shape (figures 2.B, C). Both species have discontinuous stomatal rows separated by epidermal cells that may be slightly sinuous (Stockey and Ko, 1986). They coincide in the shape and distribution of the stomata, which are characterized by globular or granular guard cell surface. In contrast, A. angustifolia has smaller leaves, sharply acute, keeled, with a distinct sinuous outline of the epidermal cells and the flange between the guard and subsidiary cells has a serrate edge.
Several fossil leaves have been described in Argentina, among them Araucaria nathorstii Dusén, reported by Menéndez and Caccavari (1966) from the Tertiary of Rio Negro Province, is the most similar to A. pararaucana sp. nov. by having ovate-lanceolate leaves, but their tips are more acute. The cuticular features in which they differ are more regular stomatal rows and the very rectangular epidermal cells in A. nathorstii. However, this species was originally described by Dusén (1899) for leaves without cuticle from Upper Oligocene-Lower Miocene strata of Punta Arenas (Chile). Later, the species was used by Berry (1928) to describe impression of leaves from the Rio Negro Province. Our specimens might be similar to the original material described by Dusén (1899), but the absence of cuticle in the Chilean specimens prevents closer comparisons.
Araucaria pichileufensis Berry, another species described by Berry (1938) from the Eocene of Rio Negro province, is characterized by small pointed, triangular, falcate leaves without cuticle preserved. The size and falcate shape differ from those of A. pararaucana sp. nov.
Recently, Del Fueyo and Archangelsky (2002) emended Araucaria grandifolia Feruglio from the Lower Cretaceous of Patagonia. This species differs from A. pararaucana in having strongly-keeled, lanceolate leaves, with stomata only parallel to the leaf margin and epidermal cells with straight walls. Araucarites santaecrucis Calder, a species from the mid-Jurassic of Patagonia (Calder, 1953) has leaves that are rhomboidal and smaller than A. pararaucana sp. nov. with an incurved apex.
Several Antarctic Araucarian fossil species have been described recently, including Araucaria alexandrensis and Araucaria chambersii (Cantrill and Falcon- Lang, 2000) from the late Albian of Alexander Island. A. alexandrensis seems to be similar in shape and size to our species but differs in having concave and keeled leaves, while A. chambersii has leaves that are much larger with a narrowly acute apex and a base that is slightly contracted and twisted. A. sp. nov. leaves described from the Upper Cretaceous of Vega Island (Césari et al., 2001) seem to be similar externally to those of A. pararaucana sp. nov. but the stomatal rows are much more regular and are separated from each other by a constant number of epidermal cells which have straight walls instead sinuous epidermal cells. Araucaria imponens Dusén, from Seymour Island (Antarctica) differs in being a keeled leaf (Dusén, 1908).
Araucarian foliage from the Lower Cretaceous of Australia was studied by Cantrill (1992). He defined six species, four of them referred to Section Eutacta whereas Araucaria seorsum Cantrill and A. lanceolatus Cantrill have similarities to Section Araucaria. A. seorsum differs from our species in having leaves widely separated with broadly triangular bases and stomata with polar extensions prominent. On the other hand, A. lanceolatus have elongate leaves with rounded leaf apex and straight-walled epidermal cells.
The Tertiary araucarian fossils leaves described from Australia (Hill, 1990) present differences in size and shape from A. pararaucana sp. nov. Araucaria lignitici Cookson and Dungan, A. readiae Hill and Bigwood, A. planus Hill, A. prominens Hill and A. uncinatus Hill have very small leaves and stomata oriented obliquely, while A. balcombensis Selling, A. hastiensis Hill and Bigwood and A. fimbriatus Hill, show a stomata parallel orientation (a characteristic shared with A. pararaucana), but they are smaller and the epidermal cells walls are not sinuous (Hill, 1990).
The genus Araucarioides was defined by Bigwood and Hill (1985) to contain species which lack diagnostic characters of extant taxa of the Araucariaceae. The three species described by Bigwood and Hill (1985) from the Eocene of Tasmania differ from A. pararaucana sp. nov. in the stomatal orientation which is predominantly oblique to transverse distribution (Bigwood and Hill, 1985).
Finally, Araucaria haastii Ettingshausen from New Zealand, shares with A. pararaucana numerous characters like the stomata orientation, the number of subsidiary cells, flange of epidermal cells and the leaf shape, but the leaves are keeled (Bose, 1973).
Conclusions
Although the pollen record including Araucariacites australis in the sequence suspected the presence of the family Araucariaceae in the flora of the Sloggett Formation, the discovery of well preserved leaves confirms the presence of these gymnosperms. The external and internal features of A. pararaucana sp. nov. allow us to include this new species in the Section Araucaria and anatomical characters compare with extant Araucaria araucana. Both taxa share the same leaf shape, arrangement and characteristics of the stomata and similar epidermal cell shapes. As suggested by Del Fueyo and Archangelsky (2002), Patagonia could be an important centre of development for Araucaria, since three of its four sections have been identified in the Mesozoic geological record. This new finding in the Late Eocene provides more evidence that during the Tertiary, Section Araucaria seems to be the unique representative in the region.
Acknowledgements
The authors are indebted to Lic. Fabián Tricárico for his valuable help as technician of the Scanning Electronic Microscope at the Museo Argentino de Ciencias Naturales B. Rivadavia. We thank D. Cantrill and an anonymous reviewer their commments which have improved the manuscript. This study was funded by the grant PICT10747 of the Agencia Nacional de Promoción Científica y Tecnológica.
References
1. Andersson, R.D. 1907 Geological fragments of Tierra del Fuego. University of Uppsala. Geological Institutionen Bulletin 8: 169-183. [ Links ]
2. Berry, E.W. 1928. Tertiary fossil plants from the Argentine Republic. Proceedings United States National Museum73: 1-27. [ Links ]
3. Berry, E.W. 1938. Tertiary Flora from the Río Pichileufu, Argentina. Geological Society of America. Special Paper 12: 1- 149. [ Links ]
4. Bigwood, A.J. and Hill, R.S. 1985. Tertiary araucarian macrofossils from Tasmania. Australian Journal of Botany 33: 645- 656. [ Links ]
5. Bose, M.N. 1973. Araucaria haastii Ettingshausen from Shag Point, New Zealand. The Palaeobotanist 22: 76-80. [ Links ]
6. Calder, M.G. 1953. A coniferous petrified forest in Patagonia. Bulletin of the British Museum (Natural History) Geology 2: 99- 138. [ Links ]
7. Caminos, R., Halle, M.J., Lapido, O., Lizuain, A., Page, R. and Ramos, V. 1981. Reconocimiento geológico de los Andes Fueguinos (San Luis). Territorio Nacional de Tierra del Fuego. Actas 8º Congreso Geológico Argentino (San Luis), Actas 3: 759-786. [ Links ]
8. Cantrill, D.J. 1992. Araucarian foliage from the Lower Cretaceous of southern Victoria, Australia. International Journal of Plant Sciences 153: 622-645. [ Links ]
9. Cantrill, D.J. and Falcon-Lang, H.J. 2000. Cretaceous (late Albian) Coniferales of Alexander Island, Antarctica. 2. Leaves, reproductive structures and roots. Review of Palaeobotany and Palynology 115: 119-145. [ Links ]
10. Césari, S.N., Marenssi, S.A. and Santillana S. N. 2001. Conifers from the Upper Cretaceous of Cape Lamb, Vega Island, Antarctica. Cretaceous Research 22: 309-319. [ Links ]
11. Chambers, T.C., Drinnan, A.N. and Mc Loughlin, S. 1998. Some morphological features of Wollemi Pine (Wollemia nobilis: Araucariaceae) and their comparison to Cretaceous plant fossils. International Journal of Plant Sciences 159: 160-171. [ Links ]
12. Del Fueyo, G.M. and Archangelsky, A. 2002. Araucaria grandifolia Feruglio from the Lower Cretaceous of Patagonia, Argentina. Cretaceous Research 23: 265-277. [ Links ]
13. Dusén, P. 1899. Über die tertiäre Flora der Magallans- Länder. In: O. Nordenskjöld (ed.), Wissenschaftliche Ergebnisse der Schwedischen Expedition nach den Megallans- Länder 1895-97. [ Links ]
14. Geologie, Geographine und Anthropologie, Lithographisches Institut des Generalstabs, Stockholm, 1: 87-108. [ Links ]
15. Dusén, P. 1908. Über die tertiäre Flora der Seymour-Insel. In: O. Nordenskjšld (ed.), Wissenschaftliche Ergebnisse der Schwedischen Südpolar-Expedition 1901-1903. Geologie und Paläontologie. Lithographisches Institut des Generalstabs, Stockholm, 4: 1-27.
16. Henkel, J.B. and Hochstetter, W. 1865. Synopsis der Nadelhölzer, deren charakteristischen Merkmale nebst Andeutungen über ihre Cultur und Ausdauer in Deutschlands Klima. Stuttgart, 446 pp. [ Links ]
17. Hill, R.S. 1990. Araucaria (Araucariaceae) species from Australian Tertiary sediments. A micromorphological study. Australian Systematic Botany 3: 203-220. [ Links ]
18. Jussieu de, A.L. 1789. Genera plantarum ordines naturales disposita. Paris, 498 pp. [ Links ]
19. Koch, K. 1873. Pinus araucana Molina. Dendrochronology 2: 206. [ Links ]
20. Kuntze, O. 1898. Revisio generum plantarum. Part 3 (3). Leipzig, 576 pp. [ Links ]
21. Menéndez, C.A. and Caccavari, M.A. 1966. Estructura epidérmica de Araucaria nathorstii Dus., del Terciario de Pico Quemado. Río Negro. Ameghiniana 4: 195-199. [ Links ]
22. Olivero, E.B., Barreda, V.D., Marenssi, S.A., Santillana, S.N. and Martinón, D.R. 1998 Estratigrafía, sedimentología y palinología de la Formación Sloggett (Paleógeno continental), Tierra del Fuego. Revista de la Asociación Geológica Argentina 53: 504-516. [ Links ]
23. Setoguchi, H., Osawa, T.A., Pintaud, J.C., Jaffré, T. and Veillon, J. M. 1998. Phylogenetic relationships within Araucariaceae based on rbcL genes sequences. American Journal of Botany 85: 1507-1516. [ Links ]
24. Silba, J. 1986. Encyclopaedia coniferae. Phytologia Memoir 8: 1- 127. [ Links ]
25. Stockey, R.A. and Ko, H. 1986. Cuticle micromorphology of Araucaria de Jussieu. Botanical Gazette 147: 508-548. [ Links ]
26. Wilde, M.H. and Eames, A.J. 1952. The ovule and seed of Araucaria bidwilli with discussion of the taxonomy of the genus. II Taxonomy. Annals of Botany (New Series) 16: 27- 47. [ Links ]
27. Zanettini, J.C. and Zapettini, E.D. 1988. Geología de la comarca de Bahía Sloggett, Territorio Nacional de la Tierra del Fuego, Antártida e Islas del Atlántico Sur. Revista de la Asociación Geológica Argentina 43: 188-197. [ Links ]
Recibido: 1 de septiembre de 2005.
Aceptado: 11 de agosto de 2006.

