










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.2 Buenos Aires abr./jun. 2007
Foraminíferos bentónicos de un testigo del Holoceno de la cuenca del Colorado, Argentina
Emiliana Bernasconi1 y Gabriela Cusminsky1
1Centro Regional Universitario Bariloche, Departamento de Ecología, Universidad Nacional del Comahue, Quintral 1250, Bari-loche, 8400 Río Negro, Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina. bernasconie@crub.uncoma.edu.ar gcusmins@crub.uncoma.edu.ar
Resumen. Se reconocen 50 especies de foraminíferos bentónicos recuperados de un testigo proveniente de la cuenca del Colorado. Predominaron Elphidium discoidale (d'Orbigny), Buccella peruviana campsi (Boltovskoy), Quinqueloculina patagonica d'Orbigny y Quinqueloculina seminula (Linné). La asociación de especies encontrada indica un ambiente marino de plataforma interna. A partir de análisis cualitativos y cuantitativos se pudieron distinguir dos tramos. En el tramo inferior se infiere un ambiente restringido, mientras que en el tramo superior se observa una tendencia al establecimiento de las condiciones marinas normales. Los sedimentos del testigo se habrían depositado durante la transgresión del último máximo glacial, hace aproximadamente 10.000 años B.P.
Abstract. Benthic foraminifers of a Holocene core from the Colorado Basin, Argentina. Fifty species of benthic foraminifera from a core of the Argentine Continental Shelf were identified. Elphidium discoidale (d'Orbigny), Buccella peruviana campsi (Boltovskoy), Quinqueloculina patagonica d'Orbigny and Quinqueloculina seminula (Linné) were the dominant species. The association is characteristic of inner shelf environments. Qualitative and quantitative analyses allow the recognition of a lower zone with an assemblage typical of restricted environments, and an upper zone which, according to foraminifers, reflects a change towards normal marine conditions. The core was deposited during the post last glacial maximum transgression, about 10.000 yr B.P.
Palabras clave. Foraminíferos bentónicos; Holoceno; Plataforma Continental Argentina; Variaciones relativas del nivel del mar.
Key words. Benthic foraminifers; Holocene; Argentine Continental Shelf; Relative sea level variations.
Introducción
Los foraminíferos bentónicos son ampliamente estudiados y constituyen una herramienta útil en la determinación de condiciones paleoambientales (Basov y Krasheninikov, 1983), paleoecológicas (Laprida, 1998; Laprida y Bertels-Psotka, 2003), paleoclimáticas (Alperin y Cusminsky, 1992) y paleoceanográficas (Bertels, 1984; Cusminsky, 1994; Alperín et al., 1995). Además, los foraminíferos fueron usados como indicadores de masas de agua (Kahn y Watanabe, 1980) y de cambios en la circulación oceánica de las aguas profundas (Van Leeuwen, 1989).
En este trabajo se presentan datos sobre los foraminíferos bentónicos y la edad de la depositación de sedimentos de un testigo proveniente de la plataforma continental argentina con el objetivo de obtener datos sobre la dinámica de la transgresión holocénica en la cuenca del Colorado. Los resultados obtenidos permiten realizar inferencias paleoecológicas respecto de las variaciones relativas del nivel del mar acontecidas durante los últimos 10.000 años.
Área de estudio
El área de estudio corresponde a la cuenca del Colorado. Se trata de un sector de la plataforma con suave pendiente en dirección noroeste-sudeste. Su relieve submarino actual no presenta marcadas variaciones. El mismo es escaso o nulo hasta profundidades de algo más de 70 m, a partir de allí el mismo posee ondulaciones suaves y desde los 140 m presenta irregularidades más marcadas (Gelós et al., 1988; Cortelezzi y Mouzo, 1979). Los sedimentos son texturalmente maduros presentando una buena selección y una composición relativamente homogénea integrada por arenas finas a muy finas (Gelós et al., 1988).
La circulación oceánica de este tramo de la plataforma está en relación con el sistema del Agua Costera Argentina. Esta masa de agua está influenciada por la acción de los vientos y las mareas que resultan muy importantes en su movimiento, siendo su dirección preponderantemente hacia el norte. Este agua es de origen subantártico, su temperatura superficial oscila entre 9ºC y 20ºC pero exhibe considerable variaciones latitudinales y estacionales. La salinidad presenta valores entre 33‰ y 33,5‰ (Boltovskoy, 1981).
Materiales y métodos
El testigo T1 fue extraído por el buque Oceanográfico ARA Puerto Deseado con un sacatestigo a pistón tipo Ewing durante la campaña realizada en 1984 por el Instituto Argentino de Oceanografía. El testigo, de 560 cm de largo, se extrajo a una profundidad de 45 m a los 40º 30' 00" S y los 60º 59´ 05" O (figura 1).

Figura 1. Mapa de ubicación del testigo estudiado (modificado de Gelós et al. (1988) / location map of the studied core (modified from Gelós et al. (1988).
En el mismo se reconoce, desde el techo hasta los 185 cm, un sedimento arenoso fino, cuyo color en húmedo varía del grisáceo claro al oscuro, con restos de conchillas y finas intercalaciones de material arcilloso de hasta 1 cm de espesor. Desde los 185 cm hasta los 230 cm hay una gradación decreciente desde un limo arenoso a una arcilla limosa; el límite inferior es neto. A partir de los 230 cm hasta los 425 cm hay una secuencia de arcilla limosa y arcillas de tono oscuro a gris verdoso con restos de conchillas. Entre los 425 cm y los 570 cm hay material muy lavado y suelto, formado principalmente por restos de conchillas, arena y algo de material arcilloso. El último tramo entre 570 cm y los 585 cm (base del testigo) lo constituye un tapón arcilloso con restos de conchillas trituradas (Gelós y Chaar, 1988).
Para este estudio se dispuso del tramo comprendido entre el techo del testigo y los 433 cm con un intervalo de muestreo de aproximadamente 10 cm. Se tomaron 31 muestras de 15 gramos, restringido a veces a la cantidad de material disponible. Las mismas se lavaron con agua a través de un tamiz de 63 µm (230 Tyler Screen System). Se extrajo la totalidad de los foraminíferos presentes bajo lupa binocular. En las muestras donde el residuo fue inferior al gramo se ponderó la cantidad de ejemplares presentes en ese peso. La clasificación sistemática de los organismos a nivel genérico se basó en la propuesta de Loeblich y Tappan (1988), mientras que a nivel específico se usaron Boltovskoy et al. (1980), Cusminsky (1992), Laprida (1998), y Laprida y Bertels-Psotka (2003). Las especies más abundantes se fotografiaron mediante microscopio electrónico de barrido (Philips, Modelo SEM 515 del Centro Atómico Bariloche). Estos ejemplares se encuentran depositados en el laboratorio de Micropaleontología del Museo de La Plata bajo el número MLP-Mi 1535 al 1545.
Se realizaron estudios cuantitativos determinando la frecuencia relativa de las especies en cada nivel. Se calcularon la riqueza específica S y los índices de diversidad de Shannon-Wiener (H') y α de Fisher. La riqueza específica S es el número de especies presente en una muestra y no toma en cuenta el número de individuos ni la proporción de éstos en la muestra. El índice de Shannon-Wiener (H') involucra tanto el tamaño de la asociación como la proporción de especies presentes en cada muestra. La función de información H' está determinada por la ecuación (Buzas y Gibson, 1969):
s
H'(S) = - Σ pi . ln pi
i=1
donde S es el número de especies y pi la proporción de cada especie en la muestra. Para calcular el índice α de Fisher (Fisher et al. 1943) se utiliza un gráfico en el cual se coloca en las abcisas el logaritmo del número de individuos y en las ordenadas el número de especies en muestras con más de 100 individuos (Murray, 1991).
Adicionalmente se realizó el análisis de agrupamiento respetando el orden estratigráfico usando el programa Coniss incluido en el paquete estadístico Tilia 2.0 (Grimm, 1991). El análisis considera sólo aquellas especies cuya abundancia relativa alcanza o supera el 2% en al menos 2 niveles del testigo (Gómez et al., 2000). Como coeficiente de distancia se utilizó la distancia euclidiana estandarizada. Se empleó la transformación de datos por estandarización a media 0 y desviación típica 1. Los grupos se formaron según el método de agrupamiento jerárquico de la suma de los cuadrados del error (Grimm, 1991).
Resultados
Los foraminíferos recuperados son exclusivamente bentónicos (cuadro 1). La abundancia fue considerablemente mayor en los niveles inferiores del testigo para decrecer drásticamente desde el nivel 290 cm hasta el techo. Con respecto a la riqueza específica S y al índice de diversidad de Shannon-Wiener (H´), los mismos tuvieron un comportamiento coincidente obteniéndose valores bajos hasta el nivel 290 cm, a partir del cual se presenta un incremento de los mismos hasta el nivel 200 cm, volviendo a decaer en los niveles superiores (figura 2).
Cuadro 1. Distribución de las especies de foraminíferos en el testigo T1 (número de individuos por gramo) / distribution of foraminiferal species in core T1 (individuals per gram).
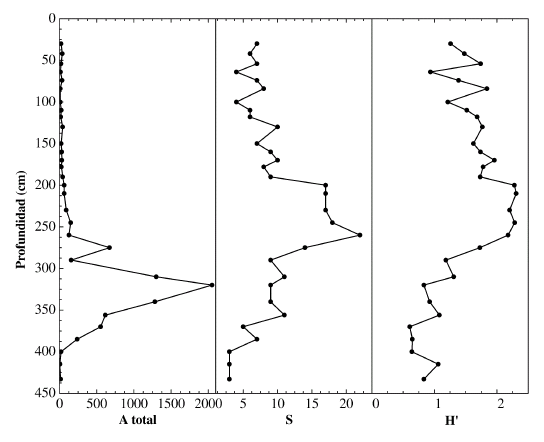
Figura 2. Abundancia total (A total), diversidad S y función de información (H') / plot of total abundance (A total), diversity S and information function (H').
La abundancia total varió entre 2.051 ejemplares en el nivel 320 cm y 5 ejemplares en el nivel 415 cm. La mayor riqueza específica se encontró en el nivel 260 cm con 22 especies mientras que el valor más bajo, de 3 especies se obtuvo en los niveles 433 cm, 415 cm y 400 cm. La diversidad H' más alta fue 2,3 y se la determinó en los niveles 245 cm, 210 cm y 200 cm, mientras que el valor más bajo de 0,6 se calculó para los niveles 400 cm, 385 cm y 370 cm. El índice α de Fisher osciló entre 0,8 en el nivel 370 cm y 7,7 en el nivel 260 cm.
Tramos y/o asociaciones determinados
El análisis de agrupamiento permitió establecer 2 tramos (figura 3). El primer tramo (T1 I) abarca desde la base hasta el nivel 275 cm inclusive y el segundo (T1 II) desde el nivel 260 cm hasta el nivel 30 cm. Los taxones más representativos se ilustran en la figura 4. Tramo T1 I (433 - 275 cm). Este tramo está caracterizado fundamentalmente por la dominancia de Elphidium discoidale (d´Orbigny) (33,3-84,4%) y la presencia en menores porcentajes de Quinqueloculina patagonica d´Orbigny (7,1-44,4%), Buccella peruviana campsi (Boltovskoy) (1 al 64,3%) y Quinqueloculina seminula (Linné) (0,5-10%). El resto de las especies se registraron en pocos niveles y con una proporción menor a 3,7%. Debe resaltarse que si bien los niveles inferiores (433 cm y 415 cm) poseen un mayor porcentaje de B. peruviana campsi y Q. patagonica, se trata de muestras donde la abundancia total es muy baja.
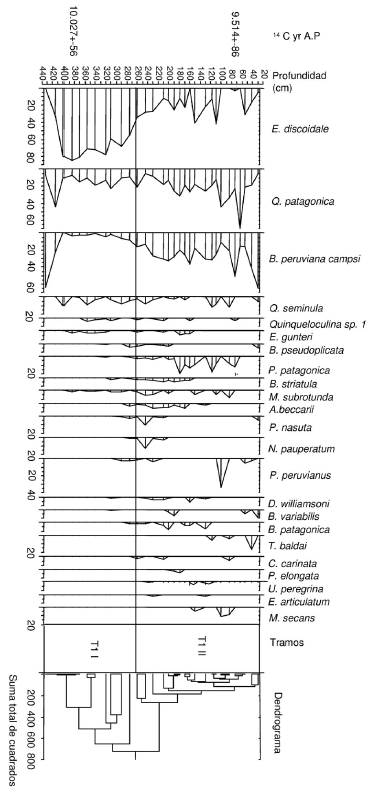
Figura 3. Distribución de las especies de foraminíferos presentes en el testigo T1 con 2% de abundancia relativa en al menos dos muestras / distribution of foraminiferal species in core T1, with 2% of relative abundance in at least two samples.
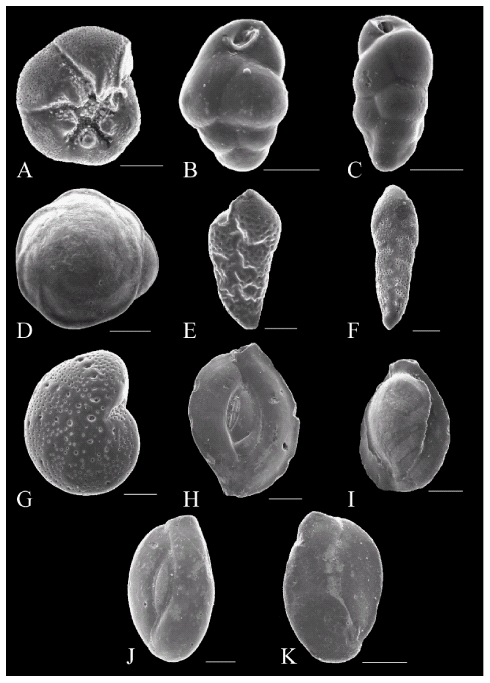
Figura 4. A, Ammonia beccarii (Linné), vista umbilical /umbilical view, MLP-Mi 1535. B, Globobilimina affinis (d´Orbigny), vista apertural / apertural view, MLP-Mi 1536. C, Bulimina patagonica d´Orbigny, vista lateral / lateral view, MLP-Mi 1537. D, Buccella peruviana campsi (Boltovskoy), vista dorsal / dorsal view, MLP-Mi 1538. E, Bolivina pseudoplicata Heron-Allen y Earland, vista lateral / lateral view, MLP-Mi 1539. F, Bolivina striatula Cushman, vista lateral / lateral view, MLP-Mi 1540. G, Elphidium discoidale (d´Orbigny), vista lateral / lateral view, MLP-Mi 1541. H, Massilina secans (d´Orbigny), vista lateral / lateral view, MLP-Mi 1542. I, Pyrgo nasuta Cushman, vista lateral / lateral view, MLP-Mi 1543, J, Quinqueloculina seminula (Linné), vista lateral / lateral view, MLP-Mi 1544. K, Triloculina baldai Bermúdez y Seiglie, vista lateral / lateral view, MLP-Mi 1545. Escala = 50 μm (figuras A, B, E, F, G); 100 μm (figuras C, D, I, J, K); 200 μm (figura H) / scale = 50 μm (figures A, B, E, F, G); 100 μm (figures C, D, I, J, K); 200 μm (figure H).
El tramo T1 I incluye las muestras con los valores de abundancia más altos de todo el testigo, alcanzando los 2.051 organismos. Presenta un número variable de especies (entre 3 y 14) y valores bajos de H' (entre 0,6 y 1,7).
Tramo T1 II (260 - 30 cm). Q. patagonica (5-69,2%), B. peruviana campsi (11,1-65%) y E. discoidale (5-41,7%) siguen predominando en este tramo. Sin embargo se observa un incremento en el porcentaje de B. peruviana campsi y Q. patagonica en desmedro de E. discoidale. El resto de las especies se encuentran en proporciones menores. Varias especies fueron registradas sólo en este tramo, entre las cuales pueden citarse Nonion pauperatum Blakwill y Wright, Triloculina baldai Bermúdez y Sieglie, Cassidulina carinata Silvestri, Pyrgo elongata (d´Orbigny), Elphidium articulatum (d´Orbigny) y Massilina secans (d´Orbigny).
Se registró una abundancia entre 9 y 150 individuos. La riqueza específica alcanza valores de hasta 22 especies. Se calcularon valores de H' entre 0,9 y 2,3. El índice se calculó sólo para los dos niveles que superaron los 100 ejemplares, resultando 7,7 (nivel 260 cm) y 5,4 (nivel 245 cm) .
Edad del testigo
Los fechados radimétricos de 14C fueron realizados por la NSF Arizona AMS Facility, Universidad de Arizona, sobre caparazones de foraminíferos bentónicos sin retrabajo. La edad de depositación de los sedimentos determinada por las dataciones para el nivel 385 cm (AA61785) es de 10.027 ± 56 años 14C B.P. y para el nivel 74 cm (AA61786) de 9.514 ± 86 años 14C B.P. Es posible hacer una corrección por las variaciones de la concentración de 14C en la atmósfera (Guilderson et al., 2000), obteniéndose 9.427 ± 56 años cal B.P. y 8.914 ± 86 años cal B.P. para los niveles 385 cm y 74 cm respectivamente. Por estos datos se observa que la depositación de los sedimentos habría ocurrido en un lapso breve durante la primera etapa del Holoceno.
Interpretación de los resultados
La ausencia de especies planctónicas en el testigo T1, cuenca del Colorado, sugiere un ambiente de plataforma interna (Boltovskoy, 1976; 1979; Caramés y Malumián, 2000). Se observó gran variabilidad de la riqueza de especies a lo largo del testigo, lo que según Sen Gupta y Kilbourne (1974) y Laprida y Bertels-Psotka (2003) podría estar indicando paleoambientes inestables.
Las especies más representadas a lo largo del testigo analizado, E. discoidale y B. peruviana campsi, junto con A. beccarii, fueron registradas por Laprida (1998) para ambientes marino marginales tales como lagunas y estuarios de las costas bonaerenses. La composición y la pobreza de la fauna hallada en algunos de los niveles analizados son comparables con las encontradas en la Formación Río Salado por Laprida (1998), quien sugiere que la depositación de los sedimentos habría ocurrido en un ambiente marino marginal.
Tramo T1 I (433 - 275 cm). Los bajos valores de H´ indicarían un ambiente de alta inestabilidad (Buzas y Gibson, 1969), mientras que los valores bajos del índice α de Fisher (entre 0,8 y 2,6) estarían indicando un ambiente hipohalino.
En este tramo la abundancia es muy alta y está dada por el predominio de E. discoidale. El género Elphidium es referido por Murray (1991) como típico de plataforma interna, encontrándose en profundidades de 0-50 m. Desde el punto de vista biogeográfico Elphidium es sumamente abundante en toda la costa argentina. En la subprovincia Norpatagónica domina E. discoidale, distinguiendo a esta subprovincia de la subprovincia Surpatagónica donde esta especie no se presenta; el límite es el paralelo 42°- 43º S (península Valdés) (Boltovskoy, 1976, 1979; Boltovskoy et al., 1980). E. discoidale vive en aguas hiposalinas de lagunas y estuarios (Boltovskoy et al., 1980). Los géneros Elphidium y Quinqueloculina son característicos de aguas de muy poca profundidad (Boltovskoy, 1966).
Estas observaciones concuerdan con las condiciones inferidas, desde el punto de vista sedimentológico, por Gelós y Chaar (1988) quienes mencionan que los sedimentos fangosos de este tramo estarían indicando un ambiente de depositación en aguas someras.
Tramo T1 II (260 - 30 cm). Si bien la abundancia es marcadamente más baja que en el tramo anterior, se observa en la base un aumento en los valores de H' como así también de la riqueza específica respecto al tramo T1 I. Hacia los niveles superiores se observa que los valores de S descienden y son similares a los del tramo T1 I. El aumento de H´ indicaría una mayor influencia marina, con condiciones próximas a la marina normal.
En este tramo es claro el predominio de Buccella peruviana campsi. El género Buccella es referido por Murray (1991) como un género de plataforma interna (0-50 m) a batial, de temperatura templada-fría. B. peruviana está ampliamente distribuida a lo largo de la plataforma continental argentina (Boltovskoy, 1976, 1979); además es típica en la zona de la corriente de Malvinas (Boltovskoy et al., 1980). El predominio de B. peruviana campsi en relación con E. discoidale sugirie una mayor influencia marina coincidente con lo observado por Gómez et al. (2005). Las inferencias ambientales realizadas para este tramo están en congruencia con lo propuesto por Gelós y Chaar (1988), que mencionan que este tramo caracterizado por sedimentos arenosos podría corresponder a un ambiente de plataforma interna.
En este tramo también se detectó la presencia de otras especies de aguas frías como Cassidulina carinata Silvestri y Uvigerina peregrina Cushman (Caramés y Malumián, 2000; Kafescioglu, 1975; Kahn y Watanabe, 1980; Murray, 1991). Se observó además la presencia de elementos de aguas cálidas como Ammonia beccarii (Linné) (Murray, 1991) y de otros elementos de la subprovincia Sudbrasileña evidenciada por la especie Massilina secans (Boltovskoy, 1979). El hallazgo de especies típicas de distintas masas de agua confirma que es un área de transición oceanográfica (Páez y Zuñiga, 2001).
Cortelezzi y Mouzo (1979) analizaron testigos provenientes del área bajo estudio señalando que los niveles de mayor contenido limo arcilloso fueron depositados durante un período en el que la profundidad era menor que la actual. Esto es avalado por la existencia de conchillas de bivalvos que corresponden a especies litorales de aguas poco profundas. Al producirse la transgresión, esos sedimentos han sido cubiertos por otros semejantes a los que se depositan en la actualidad (Cortelezzi y Mouzo, 1979).
Según Gelós y Chaar (1988) los niveles inferiores del testigo T1 están compuestos por sedimentos fangosos correspondientes a una depositación en aguas someras que representan un ambiente del tipo litoral albúferico o marismal, en el que prevalece un régimen de corrientes de muy baja energía. Los niveles superiores del mismo están caracterizados por sedimentos arenosos que corresponderían a un ambiente litoral costero de playas y cordones litorales en el que se presenta un régimen de mareas y corrientes costeras de alta energía, señalando un ambiente de plataforma interna. El estudio de los foraminíferos del testigo T1 confirma una mayor influencia de las condiciones marinas hacia el tope del testigo, lo que implica desde el punto de vista dinámico un ambiente inestable relacionado con el ascenso del nivel del mar.
En la Argentina, las fluctuaciones del nivel del mar no se expresaron de manera uniforme sino por el contrario, poseyeron fuertes componentes locales y regionales (Schnack et al., 2005). En el área bajo estudio, las dataciones radimétricas indican que la depositación de los sedimentos analizados habría ocurrido durante el Holoceno Temprano (Clapperton, 1993). Según Guilderson et al. (2000), Violante et al. (2001), Violante y Parker (2000; 2004) y Cavallotto et al. (2004) en el momento de la depositación de los sedimentos del testigo analizado el nivel del mar se encontraba en franco ascenso, lo que es reflejado por las asociaciones faunísticas halladas.
Conclusiones
El análisis del testigo T1 proveniente de la plataforma continental Argentina, ubicado a los 40º 30' 00" S y los 60º 59´ 05" O indica que el mismo se depositó a principios del Holoceno durante un episodio transgresivo ocurrido aproximadamente a los 10027 años BP. Los taxa presentes en el testigo T1 sugieren un ambiente marino somero con profundización hacia el techo del testigo, verificando de esa manera una elevación relativa del nivel del mar. Este ascenso del nivel del mar inferido concuerda con lo concluido por varios autores para el intervalo 10.000-8.200 años A P.
Agradecimientos
Este trabajo constituye una contribución a proyectos financiados por la Agencia de Promoción Científica y Tecnológica (BID 1201/OC-AR PICT 07-09659; ANCYT FONCYT 03 Nº 07-14653; CONICET PIP 2000 / 02040 y al proyecto de la Universidad Nacional del Comahue (UNC B940). Las autoras desean agradecer al Instituto Argentino de Oceanografía (IADO) por las muestras facilitadas.
Bibliografía
1. Alperín M.I. y Cusminsky, G.C. 1992. Variaciones paleoclimáticas sobre la base de foraminíferos y radiolarios de un testigo del talud continental argentino. 3° Jornadas Geológicas Bonaerenses (La Plata), Actas: 109-111. [ Links ]
2. Alperín M.I. Caballé, M.F. y Cusminsky, G.C. 1995. Bioestratigrafía y paleooceanografía de sedimentos del océano Atlántico Sur comprendidos entre los 52º y 56º Latitud S y los 56º y 62º Longitud O. 1° Jornadas de Comunicaciones Científicas de la Facultad de Ciencias Naturales y Museo Universidad Nacional de La Plata, p. 1. [ Links ]
3. Basov, I.A. y Krasheninnikov, V.A. 1983. Benthic foraminifers in Mesozoic and Cenozoic sediments of the southwestern Atlantic as an indicator of paleoenvironment. Deep Sea Drilling Project, Leg- 71. Initial Reports Deep Sea Drilling Project 71: 739-787. [ Links ]
4. Bertels, A. 1984. Foraminíferos bentónicos del Neógeno del océano Atlántico sudoccidental austral: inferencias paleoclimáticas y paleooceanográficas. 2° Congreso Argentino de Geología (Bariloche), Actas: 455-474. [ Links ]
5. Boltovskoy, E. 1966. Los Foraminíferos recientes. Editorial Universitaria de Buenos Aires, 510 pp. [ Links ]
6. Boltovskoy, E. 1976. Distribution of Recent Foraminifera of the South American Region. Foraminifera 2: 171-236. [ Links ]
7. Boltovskoy, E. 1979. Paleoceanografia del Atlántico Sudoccidental desde el Mioceno, según estudios foraminiferológicos. Ameghiniana 16: 357-389. [ Links ]
8. Boltovskoy, E. 1981. Masas de agua en el Atlántico Sudoccidental. En: D. Boltovskoy (ed.), Atlas de Zooplancton del Atlántico Sudoccidental (Mar del Plata), pp. 227-236. [ Links ]
9. Boltovskoy, E., Giussani, G., Watanabe, S. y Wright, R. 1980. Atlas of benthic shelf foraminifera of the southwest Atlantic. W. Junk Publishers, The Hague, 147 pp. [ Links ]
10. Buzas, M.A. y Gibson, T.G. 1969. Species diversity: Benthonic foraminifera in Western North Atlantic. Science 163: 72-75. [ Links ]
11. Caramés, A. y Malumián, N. 2000. Foraminíferos bentónicos del Cenozoico del pozo Estrella x -1, Cuenca del Colorado, plataforma continental argentina. Ameghiniana 37: 387-419. [ Links ]
12. Cavallotto, J.L., Violante, R.A. y Parker, G. 2004. Sea-level fluctuations during the last 8600 yrs. in the de la Plata River, Argentina. Quaternary International 114: 155-165. [ Links ]
13. Clapertton, C. 1993. Quaternary Geology and Geomorphology of South America. Elsevier, Amsterdam, 775 pp. [ Links ]
14. Cortelezzi, C.R. y Mouzo, F.H. 1979. Estudio sedimentológico de testigos del Margen Continental Argentino sobre un perfil a los 40º Lat. Sur. 6° Congreso Geológico Argentino (Buenos Aires), Actas 2: 249-264. [ Links ]
15. Cusminsky, G. 1992. Foraminíferos bentónicos provenientes de testigos del océano Atlántico Sudoccidental Austral. Revista Española de Micropaleontología 24: 5-32. [ Links ]
16. Cusminsky, G.C. 1994. Estudio micropaleontológico (Foraminiferida) de dos testigos extraídos del océano Atlántico sudoccidental austral. Revista Española de Micropaleontología 26: 109-123. [ Links ]
17. Fisher, R.A., Corbet, A.S. y Williams, C.B. 1943. The relation between the number of species and the number of individuals in a random sample of an animal population. Journal Animal Ecology 12: 42-58. [ Links ]
18. Gelós, M.E. y Chaar, E. 1988. Estudio sedimentológico - mineralógico de testigos de la plataforma continental argentina entre los paralelos de 39° y 42° de latitud Sur. 2° Jornadas Geológicas Bonaerenses (Bahía Blanca), Actas: 441- 440. [ Links ]
19. Gelós, M.E., Spagnuolo, O. y Lizasoain, O. 1988. Mineralogía y caracterización granulométrica de sedimentos actuales de la plataforma argentina entre los paralelos 39º y 43º de Latitud Sur y del golfo San Matías. Revista de la Asociación Geológica Argentina 18: 63-79. [ Links ]
20. Gómez, E.A., Martínez, D., Cusminsky, G.C., Borel, C.M. y Guerstein, R. 2000. Submerged tidal flats at the Bahía Blanca Estuary, Argentina. Stuttgart. 17 Latein Amerika- Kolloquium, Profil 18, 6 pp. [ Links ]
21. Gómez, E., Martínez, D., Borel, M., Guerstein, G.R. y Cusminsky, G.C. 2005. Submarine evidences of Holocene sea-level fluctuations in the Bahía Blanca Estuary, Argentina. Journal of South America Earth Sciences 20: 139-155. [ Links ]
22. Grimm, E.C. 1991. Tilia version 2.0. Illinois State Museum. Research and Collections Center. Springfield. United States. [ Links ]
23. Guilderson, T.P., Burckle, L., Hemming, S. y Peltier, W. 2000. Late Pleistocene sea level variations derived from the Argentine Shelf. Geochemistry. Geolphysics. Geosystems 1, (Paper number 2000GC000098). Published December 15, 2000. [ Links ]
24. Kafescioglu, I. 1975. Quantitative distribution of foraminifera on the continental shelf and uppermost slope off Massachusetts. Micropaleontology 21: 261-305. [ Links ]
25. Kahn, G. y Watanabe, S. 1980. Foraminíferos bentónicos como indicadores de la corriente de Malvinas. Revista Española de Micropaleontología 12: 169-177. [ Links ]
26. Laprida, C. 1998. Micropaleontological assemblages (Foraminiferida and Ostracoda) from Late Quaternary marginal marine environments (Destacamento Río Salado Formation), Salado Basin, Argentina. Revue de Paleobiologie 17: 461-478. [ Links ]
27. Laprida, C. y Bertels-Psotka, A. 2003. Benthic foraminifers and paleoecology of a Holocene shelly concentration, Salado Basin, Argentina. Geobios 36: 559-572. [ Links ]
28. Loeblich, A, Jr. y Tappan, H. 1988. Foraminiferal Genera and their Classification. Van Nostrand Reinhold Company. New York, 1-2, 1182 pp. [ Links ]
29. Murray, W. 1991. Ecology and Paleoecology of Benthic Foraminifera. Longman, Wiley, Harlow/Essex, New York, 397 pp. [ Links ]
30. Páez, M. y Zúñiga, Ó. 2001. Foraminiferos recientes en el área del Río Loa (21º 25'S; 70º 04'W), Chile. Estudio Oceanológico 20: 3-12. [ Links ]
31. Schnack, E., Isla, F., De Francesc, F. y Fucks, E. 2005. Estratigrafía del Cuaternario marino tardío en la provincia de Buenos Aires. En: R.E. de Barrio, R.O. Etcheverry, M.F. Caballé y E. Llambías (eds.), Geología y Recursos Minerales de la Provincia de Buenos Aires. Relatorio del 16º Congreso Geológico Argentino (La Plata): 159-182. [ Links ]
32. Sen Gupta, B. y Kilbourne, E. 1974. Diversity of Benthic Foraminifera on the Georgia Continental Shelf. Geological Society of American Bulletin 85: 969-972. [ Links ]
33. Van Leeuwen, R.J.W. 1989. Sea-floor distribution and Late Quaternary faunal patterns of planktonic and benthic foraminifers in the Angola Basin. Utrecht Micropaleontological Bulletins 38: 287 pp. [ Links ]
34. Violante, R. y Parker, G. 2000. El Holoceno en las regiones marinas y costeras del nordeste de la provincia de Buenos Aires. Revista de la Asociación Geológica Argentina 55: 337-351. [ Links ]
35. Violante, R. y Parker, G. 2004. The post-last glacial maximum transgression in the de la Plata River an adjacent inner continental shelf, Argentina. Quaternary International 114: 167-181. [ Links ]
36. Violante, R., Parker, G. y Cavallotto, J.L. 2001. Evolución de las llanuras costeras del este bonaerense entre la Bahía Samborombón y la laguna Mar Chiquita durante el Holoceno. Revista de la Asociación Geológica Argentina 56: 51-66. [ Links ]
Recibido: 31 de marzo de 2005.
Aceptado: 11 de agosto de 2006.

