










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAmeghiniana
versión On-line ISSN 1851-8044
Ameghiniana v.44 n.2 Buenos Aires abr./jun. 2007
La paleoflora triásica de Potrerillos, provincia de Mendoza, Argentina
Analía E. Artabe1,2, Eduardo M. Morel1,3 Daniel G. Ganuza1, Ana M. Zavattieri4 y Luis A. Spalletti5
1División Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. aartabe@museo.fcnym.unlp.edu.ar
2Consejo Nacional de Investigaciones Científicas y Técnicas.
3Comisión de Investigaciones Científicas de la Provincia de Buenos Aires.
4Laboratorio de Paleopalinología, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Centro Regional de Investigaciones Científicas y Tecnológicas - Consejo Nacional de Investigaciones Científicas y Técnicas.
5Centro de Investigaciones Geológicas, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata y Consejo Nacional de Investigaciones Científicas y Técnicas.
Resumen. En los cerros Bayo y Cocodrilo (Potrerillos, noroeste de la provincia de Mendoza, Argentina) se levantaron dos perfiles de detalle y durante el relevamiento de datos se determinó la existencia de 16 niveles fosilíferos con plantas fósiles. En esta contribución se presenta el estudio sistemático de la paleoflora. Los 23 taxones reconocidos corresponden a Sphenopsida (Equisetales), Filicopsida (Filicales) y Gymnospermopsida (Corystospermales, Peltaspermales, Cycadales, Ginkgoales, Petriellales y Gnetales). En este trabajo se describen y comparan los taxones identificados: Equisetites fertilis (Frenguelli) Frenguelli, Neocalamites carrerei (Zeiller) Halle, Cladophlebis mesozoica Kurtz ex Frenguelli, Cladophlebis mendozaensis (Geinitz) Frenguelli, Dicroidium argenteum (Retallack) Gnaedinger y Herbst, Dicroidium dubium (Feistmantel) Gothan, Dicroidium odontopteroides (Morris) Gothan, Johnstonia stelzneriana (Geinitz) Frenguelli, Johnstonia coriacea (Johnston) Walkom, Xylopteris elongata (Carruthers) Frenguelli, Zuberia feistmanteli (Johnston) Frenguelli, Feruglioa samaroides Frenguelli, Pachydermophyllum praecordillerae (Frenguelli) Retallack, Kurtziana cacheutensis (Kurtz) Frenguelli emend. Petriella y Arrondo, Sphenobaiera argentinae (Kurtz) Frenguelli, Baiera cuyana Frenguelli, Baiera africana Baldoni, Rochipteris sp., Yabeiella mareyesiaca (Geinitz) Ôishi, Yabeiella spathulata Ôishi, Yabeiella brackebuschiana (Kurtz) Ôishi, Fraxinopsis andium (Frenguelli) Anderson y Anderson y Gontriglossa sp.
Abstract. The Triassic paleoflora of Potrerillos, Mendoza Province, Argentina. At the Bayo and Cocodrilo Hills (Potrerillos, northwestern Mendoza Province, Argentina) two stratigraphic logs were measured in detail and 16 plantiferous levels were determined. In this contribution the systematic study of the paleoflora is presented. The 23 taxa recognized belong to Sphenopsida (Equisetales), Filicopsida (Filicales) and Gymnospermopsida (Corystospermales, Peltaspermales, Cycadales, Ginkgoales, Petriellales and Gnetales). The following taxa are described and compared: Equisetites fertilis (Frenguelli) Frenguelli, Neocalamites carrerei (Zeiller) Halle, Cladophlebis mesozoica Kurtz ex Frenguelli, Cladophlebis mendozaensis (Geinitz) Frenguelli, Dicroidium argenteum (Retallack) Gnae-dinger and Herbst, Dicroidium dubium (Feistmantel) Gothan, Dicroidium odontopteroides (Morris) Gothan, Johnstonia stelzneriana (Geinitz) Frenguelli, Johnstonia coriacea (Johnston) Walkom, Xylopteris elongata (Carruthers) Frenguelli, Zuberia feistmanteli (Johnston) Frenguelli, Feruglioa samaroides Frenguelli, Pachydermophyllum praecordillerae (Frenguelli) Retallack, Kurtziana cacheutensis (Kurtz) Frenguelli emend. Petriella and Arrondo, Sphenobaiera argentinae (Kurtz) Frenguelli, Baiera cuyana Frenguelli, Baiera africana Baldoni, Rochipteris sp., Yabeiella mareyesiaca (Geinitz) Ôishi, Yabeiella spathulata Ôishi, Yabeiella brackebuschiana (Kurtz) Ôishi, Fraxinopsis andium (Frenguelli) Anderson and Anderson and Gontriglossa sp.
Palabras clave. Paleobotánica; Sistemática; Formación Potrerillos; Triásico; Mendoza; Argentina.
Key words. Paleobotany; Systematics; Potrerillos Formation; Triassic; Mendoza; Argentina.
Introducción
En esta contribución se da a conocer el estudio sistemático de la flora fósil exhumada de las sedimentitas triásicas de la Formación Potrerillos, aflorantes en los cerros Bayo y Cocodrilo, sector meridional de la Precordillera mendocina (figura 1). La zona de trabajo corresponde a un área clásica de la literatura geológica del Triásico de la Argentina, por tratarse de la región donde se definieron las secciones tipo de las formaciones Río Mendoza (Borrello, 1962), Cerro de las Cabras (Frenguelli, 1948), Potrerillos (Truempy y Lhez, 1937), Cacheuta (Truempy y Lhez, 1937) y Río Blanco (Fossa Mancini, 1937). En un trabajo anterior se dieron a conocer los perfiles del cerro Bayo y cerro Cocodrilo, relevados a escala de detalle (1:100) de las unidades que componen la sucesión triásica del área de Potrerillos (cf. Spalletti et al., 2005). Asimismo se describieron las facies sedimentarias, se determinaron los sistemas de depositación, se caracterizaron los distintos niveles paleoflorísticos (se realizó la determinación taxonómica y la valoración semicuantitativa de los componentes de cada tafocenosis) y esta información se utilizó con fines bioestratigráficos.
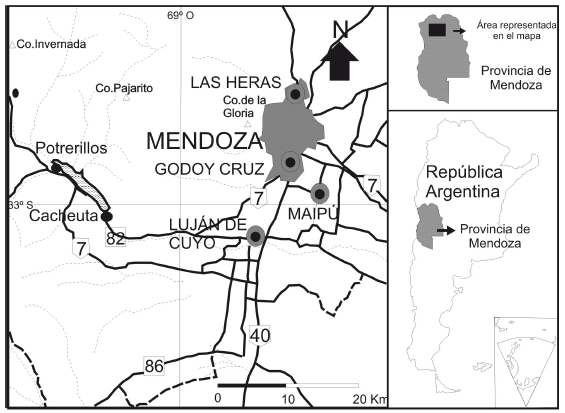
Figura 1. Mapa de ubicación de la región de Potrerillos, provincia de Mendoza, en el oeste de la República Argentina / location map of Potrerillos area in Mendoza Province, in the western of República Argentina.
Desde el punto de vista litoestratigráfico existe bastante acuerdo en lo que concierne a las denominaciones formales de las unidades que constituyen el Grupo Uspallata (véase Spalletti et al., 2005). La Formación Río Mendoza (314 m) está caracterizada por el desarrollo de bancos conglomerádicos en el que se superponen estratos tabulares y lenticulares potentes e integran una sucesión de tonalidad castaño pálida a castaño rojiza. La Formación Cerro de Las Cabras (190 m) muestra dos secciones. La basal es predominantemente pelítica con intercalaciones de cuerpos lenticulares de conglomerados y areniscas conglomerádicas y presenta una tonalidad general rojo ladrillo. La sección superior está constituida por una sucesión de pelitas multicolores con periódicas intercalaciones de tobas y chonitas (o piropelitas) generadas como depósitos piroclásticos de caída. La Formación Potrerillos (735 m) corresponde a la unidad de mayor desarrollo y variabilidad litológica. En ella se reconocen tres secciones con dominio de depósitos conglomerádicos. Así, en la parte inferior de la unidad se identifican los gruesos conglomerados ocres de la base del cerro Bayo. La segunda, comprende a los depósitos gris amarillentos que coronan al cerro Bayo. Por último, la tercera es un tramo psefítico ubicado en la parte superior de la unidad y que se destaca por su coloración rojo ladrillo. Esta última corona el cerro Cocodrilo, baja a la quebrada de las Minas y se puede seguir por el faldeo sur del cerro Bayo. El resto de la sucesión de la Formación Potrerillos se caracteriza por la alternancia de dos conjuntos, uno en el que prevalecen las fangolitas grises y gris verdosas, y otro compuesto por areniscas y areniscas conglomerádicas de tonos grises, ocres, rosado-rojizos y castaños. Finalmente en la Formación Cacheuta (44 m) se distingue un marcado predominio de fangolitas y lutitas gris oscuras a negras con ocasionales intercalaciones de areniscas y areniscas conglomerádicas, particularmente en la parte inferior de la Formación.
En el perfil del cerro Bayo, se identificaron dos secciones estratigráficas con niveles plantíferos. Una inferior desarrollada entre los conglomerados ocráceos de la base del cerro Bayo y el conglomerado rojo (Spalletti et al., 2005), que involucra a los estratos plantíferos 1 a 10, y otra superior, que se reconoce en los cerros Bayo y Cocodrilo, ubicada entre el conglomerado rojo y las lutitas negras atribuidas a la Formación Cacheuta. La sección estratigráfica (apéndice 1) comprendida entre el conglomerado de la base del cerro Bayo y el conglomerado rojo del cerro Bayo (con los niveles fosilíferos 1 al 10) fue asignada por Spalletti et al. (2005) a la Biozona de Asociación Yabeiella mereyesiaca - Scytophyllum bonettiae - Protophyllocladoxylon cortaderitaensis (MBC). La sección estratigráfica desarrollada por encima del conglomerado rojo y la base de las pelitas negras de fondo lacustre del cerro Bayo (niveles fosilíferos 11 al 13) y del cerro Cocodrilo (niveles fosilíferos del 1 al 3) fue referida por Spalletti et al. (2005) a la Biozona de Asociación Yabeiella brackebuschiana - Scytophyllum neuburgianum - Rhexoxylon piatnitzkyi (BNP). En los perfiles estudiados, las biozonas reconocidas se presentan en concordancia y continuidad estratigráfica y el límite entre ambas está representado por el techo del conglomerado rojo. Recientemente Spalletti et al. (en prensa) dataron zircones en bancos tobáceos en el perfil del Cerro Bayo (apéndice 1). Una muestra ubicada por debajo de los niveles fosilíferos hallados en la Formación Potrerillos presenta una edad de 239.7 +/- 2.2 Ma. Otra muestra analizada, que se ubica por encima del nivel fosilífero 10 (NF 10) tiene una edad de 230.3 +/- 2.2 Ma. Estas edades radimétricas confirman la antigüedad asignada a las biozonas respectivas por Spalletti et al. (1999) y Morel et al. (2003). Por lo tanto se concluye que la Formación Potrerillos -en este sector de la cuenca- abarca el lapso Triásico Medio Tardío a Triásico Tardío Temprano.
Cabe aclarar que la tarea de colección de material fosilífero en la Formación Potrerillos se vio favorecida por los trabajos llevados a cabo en la nueva traza de la ruta nacional nº 7, que en la región de Potrerillos atraviesa el flanco este y norte del cerro Bayo y la quebrada de la Mina (figura 1).
Los estudios paleobotánicos realizados en los cerros Bayo y Cocodrilo son escasos. La primera referencia corresponde a Stappenbeck (1910) quien presentó un perfil de unos 50 metros de espesor del área de Potrerillos al sur del río Mendoza, e incluyó una lista de plantas fósiles "Réticas" (determinadas por Kurtz). Con posterioridad se dieron a conocer los aportes de Borrello (1942) y Frenguelli (1948) en los que además de aspectos geológicos y estratigráficos, se incluyen listas de plantas fósiles, que aunque discriminadas por niveles estratigráficos involucran registros de varias localidades del norte de Mendoza.
Por el contrario, los antecedentes referidos a otras localidades del norte de la provincia de Mendoza - como Paramillo de Uspallata, El Challao, Divisadero Largo y cerro Cacheuta- son más numerosos y su paleoflora, en muchos casos, ha sido bien estudiada (Kurtz, 1921; Harrington, 1941; Frenguelli, 1948; Jain y Delevoryas, 1967; Morel, 1994; Spalletti et al., 1995, 1999; Stipanicic et al., 1996; Brea, 1995, 1997, 2000; Artabe et al., 1999a; Brea y Artabe, 1999; Artabe y Brea, 2003).
Las impresiones que componen el material analizado en el presente trabajo fueron depositadas en la colección de la División Paleobotánica del Museo de La Plata, bajo la sigla LPPB y en el Museo Municipal de Ciencias Naturales y Antropológicas "Juan Cornelio Moyano" de la ciudad de Mendoza (MCNAM-PB).
Sistemática paleontológica
Durante los últimos años se ha producido un considerable avance en el conocimiento de las floras triásicas, de las paleocomunidades que se desarrollaron durante este período y de las plantas que las integraron (Anderson y Anderson, 1983, 1989, 2003; Anderson et al., 1998; Artabe y Brea, 2003; Artabe et al., 1999a, 1999b, 2001, 2003; Axsmith et al., 1997, 1998, 2000; Cúneo et al., 2003; Herbst y Lutz, 1988, 1995; Klavins et al., 2002; Lutz y Herbst, 1992; Meyer-Berthaud y Taylor, 1991; Meyer-Berthaud et al., 1992, 1993; Morel et al., 2001, 2003; Perovich y Taylor, 1989; Smoot et al., 1985; Taylor, 1992, 1996; Taylor et al., 1994; Yao et al., 1995, 1997; Zamuner et al., 1999, 2001). El hallazgo de hojas y estructuras reproductivas permineralizadas y con cutícula, y de tallos en asociación estrecha y repetida, así como el descubrimiento reciente de conexiones orgánicas entre hojas -tratadas tradicionalmente como Incertae Sedis- y estructuras reproductivas permitió avanzar en la reconstrucción de plantas enteras y en la definición de grupos naturales. El valor de estos aportes trasciende al conocimiento particular de las floras triásicas gondwánicas, ya que la elaboración de reconstrucciones consistentes de plantas enteras conduce a un mejor entendimiento de los grupos, de sus relaciones filogenéticas y genera un salto cualitativo en el esquema de clasificación. Teniendo en cuenta que la información aún no ha sido integrada en un esquema único que respete las relaciones filogenéticas de los grupos, se utiliza en forma provisoria la clasificación del Reino Plantae propuesta por Stewart y Rothwell (1993) y utilizado por Zamuner et al. (2001).
La asignación sistemática de la paleoflora de la Formación Potrerillos hallada en los cerros Bayo y Cocodrilo se muestra en la tabla 1. En la tabla 2 se presenta la distribución de los 23 taxones en los estratos plantíferos reconocidos en la mencionada unidad estratigráfica.
Tabla 1. Asignación sistemática de los taxones registrados en la Formación Potrerillos (cerros Bayo y Cocodrilo) / systematic assignation of the recorded taxa in Potrerillos Formation (Bayo and Cocodrilo Hills).
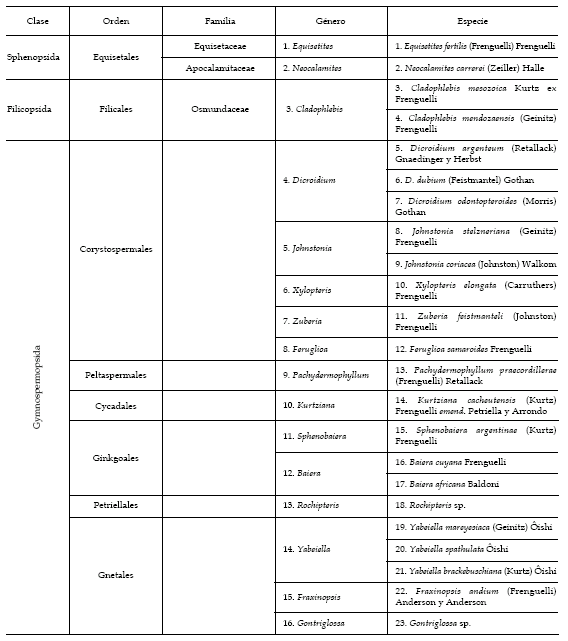
Tabla 2. Contenido paleoflorístico por nivel fosilífero de la Formación Potrerillos en los cerros Bayo y Cocodrilo / palaeofloristic record per fossiliferous level of the Potrerillos Formation at Bayo and Cocodrilo Hills.

División TRACHEOPHYTA sensu Stewart y Rothwell 1993
Clase SPHENOPSIDA sensu Stewart y Rothwell 1993
Orden EQUISETALES sensu Boureau 1964
Familia EQUISETACEAE Richard ex DeCandolle 1805
Género Equisetites Sternberg 1833
Especie tipo. Equisetites münsteri Sternberg 1833.
Equisetites fertilis (Frenguelli) Frenguelli 1944a Figuras 2.A, B

Figura 2. A-B, Equisetites fertilis (Frenguelli) Frenguelli. A. LPPB 11886a; B, LPPB 11887; C-F, Neocalamites carrerei (Zeiller) Halle. C, LPPB 11889; D, LPPB 11894; E, Diafragma nodal / nadal diagram LPPB 11888b; F, LPPB 11895.
Descripción. Tallos articulados de hasta 1,4 cm de ancho, con entrenudos que oscilan entre 0,6 -1,5 cm y hojas reducidas y fusionadas a nivel del nudo.
Material estudiado. Perfil cerro Bayo: NF6: LPPB 11883; NF10: LPPB 11884, 11885; MCNAM-PB 1401, 1402; NF12: LPPB 11886a, 11886b, 11887.
Comparaciones. Se asignaron a Equisetites aquellas formas articuladas con caracteres megascópicos semejantes a los del género Equisetum. En este sentido los tallos encontrados presentan hojas fusionadas en la base -conformando vainas adpresas a nivel del nudo- y libres en la zona distal.
Familia APOCALAMITACEAE Radczenko 1957
Género Neocalamites Halle 1908
Especie tipo. Neocalamites hoerensis (Schimper) Halle 1908.
Neocalamites carrerei (Zeiller) Halle, 1908 Figuras 2.C-F
Descripción. Impresiones externas de tallos articulados, con longitud superior a 30 cm y un ancho que oscila entre 2 y 9 cm. Los entrenudos alcanzan hasta 6 cm de largo. Presentan costillas principales marcadas, continuas a través de los nudos, contándose aproximadamente 2 o 3 por cm. Presentan verticilos de ramas de hasta 25 elementos que se observan como rastros circulares por encima del nudo.
Material estudiado. Perfil cerro Bayo: NF6: LPPB 11933, MCNAM- PB 1403, 1404a, 1404b; NF7: MCNAM-PB 1405a, 1405b; NF8: MCNAM-PB 1406a, 1406b, 1409; NF10: LPPB 11888a, 11888b, 11889, 11890, MCNAM-PB 1407a, 1407b, 1408, 1410, 1411, 1412; NF12: LPPB 11891a, 11891b, 11892, 11893a, 11893b, 11894, MCNAM- PB 1413, 1414, 1415; NF13: LPPB 11895; Perfil cerro Cocodrilo: NF1: LPPB 11896, MCNAM-PB 1416, 1417; NF2: LPPB 11897, 11898a, 11898b, MCNAM-PB 1418, 1419a, 1419b, 1420, 1421; NF3: MCNAM-PB 1422.
Comparaciones. N. carrerei se distingue de otras especies triásicas del género citadas para Argentina (N. ramaccionii Frenguelli 1944b y N. ischigualasti Frenguelli 1944c) por su hábito arborescente, con tallos de grandes dimensiones y con entrenudos que oscilan entre 3 a 12 cm de largo. Los tallos poseen ramas dispuestas en verticilos sobre ciertos nudos, y dejan al caer rastros rameales ovales-circulares. El hallazgo de impresiones de tallos en asociación estrecha con estructuras reproductivas no bracteadas (Brea y Artabe, 1999) y diafragmas nodales con crecimiento secundario en espesor (Artabe y Zamuner, 1991) permitió realizar la reconstrucción de la planta y asignarla a la Familia Apocalamitaceae. Holmes (2000) no reconoce la reconstrucción de la planta entera (Neocalamites carrerei-Nododendron suberosum - Neocalamostachys arrondoi) realizada con material de la Argentina y sugiere que todos los tallos con hojas del Gondwana -previamente incluidos en Neocalamites o Phyllotheca- deberían ser reasignados a dos géneros Incertae Sedis de Equisetales: Zonulamites Holmes,
2000 o Townroviamites Holmes, 2000.
Clase FILICOPSIDA sensu Stewart y Rothwell, 1993
Orden FILICALES sensu Stewart y Rothwell, 1993
Familia OSMUNDACEAE Berchtold y Presl, 1820
Género Cladophlebis Brongniart emend. Frenguelli 1947
Especie tipo. Cladophlebis albertsii (Dunker) Brongniart 1849.
Cladophlebis mesozoica Kurtz ex Frenguelli 1947 Figuras 3.A, B

Figura 3. A-B, Cladophlebis mesozoica Kurtz ex Frenguelli. A, LPPB 11902; B, LPPB 11899; C-D, Cladophlebis mendozaensis (Geinitz) Frenguelli. C, LPPB 11913b; D, LPPB 11922a; E-G, Dicroidium argenteum (Retallack) Gnaedinger y Herbst. E, LPPB 11956a; F, LPPB 11957a; G, 11957a; H, Dicroidium dubium (Feistmantel) Gothan, LPPB 11966.
Descripción. Fronde bipinnada de hasta 12 cm de largo por 16 cm de ancho. Disponemos de sectores medios y distales. Las pinnas son lanceoladas y se insertan con un ángulo de 70º; las de la zona media de la fronde alcanzan 9 cm de largo por 3,4 cm de ancho. Las pínnulas son de forma falcada, con ápice agudo; las de la zona media de la pinna miden 1,5 cm de largo por 1 cm de ancho; se insertan por su ancho máximo, con un ángulo de 70º, en forma opuesta, subopuesta o alternas; el margen lateral es entero o levemente ondulado y en la zona de inserción, el basiscópico es decurrente y confluente con el acroscópico que es recto o levemente expandido. La venación es cladofleboide; tienen una vena media que alcanza el tercio distal pinnular y se resuelve en varias dicotomías; las venas secundarias se bifurcan hasta dos veces, primero muy cerca de la vena media y luego en la mitad de su recorrido al margen; cada vena puede resolverse en cuatro, o en tres si una vena de la horquilla permanece libre.
Material estudiado. Perfil cerro Bayo: NF3: LPPB 11899, 11900, 11901, MCNAM-PB 1423; NF4: LPPB 11902, 11903, MCNAM-PB 1424, 1425, 1426; NF5: LPPB 11904a, 11904b, 11905; NF8: LPPB 11906; NF12: LPPB 11907. Perfil cerro Cocodrilo: NF2: LPPB 11897, 11908, 11909, 11910, MCNAM-PB 1418, 1419a, 1419b, 1421, 1429, 1430.
Comparaciones. El polimorfismo observado en esta especie dificulta la determinación específica cuando la preservación es fragmentaria (Herbst, 1971). Una especie nueva recientemente creada para Australia (C. retallackii) por Holmes (2003) es bastante similar a C. mesozoica, y en el caso de que fuesen sinónimos esta última tendría claramente prioridad.
Cladophlebis mendozaensis (Geinitz) Frenguelli 1947 Figuras 3.C, D
Descripción. Fragmentos de frondes bipinnadas; disponemos de sectores de pinnas de hasta 5 cm de largo por 2,5 cm de ancho; las pínnulas miden 1,4 cm de largo por 0,6 cm de ancho; son de forma falcada, con márgenes lobulados o crenulados y ápice agudo o redondeado; se insertan por su ancho máximo, con un ángulo de 75º, en forma alterna o subopuesta. La vena media es decurrente respecto del raquis secundario y alcanza el tercio distal pinnular, resolviéndose luego en sucesivas dicotomías; las venas laterales se bifurcan dos veces, primero muy cerca de la vena media y luego en el tercio distal; por lo tanto cada vena se resuelve en cuatro, que corresponden a un lóbulo o crena.
Material estudiado. Perfil cerro Bayo: NF4: LPPB 11911, 11912, 11913a, 11913b, 11914 al 11920, MCNAM-PB 1431 al 1438; NF5: LPPB 11921, MCNAM-PB 1427, 1428; NF8: LPPB 11922a, 11922b, 11922c, 11923a, 11923b, 11924, MCNAM-PB 1439A, 1439b, 1440 al 1442; NF12: LPPB 11925, 11926, 11927a, 11927b, 11928a, 11928b, 11929, MCNAM-PB 1443.
Comparaciones. La presencia de pínnulas alternas, margen suavemente ondulado y la doble bifurcación de las venas laterales, con la particularidad de que un juego de cuatro venillas se corresponde con cada lóbulo, distingue a esta especie de las formas más afines, C. mesozoica y C. kurtzi. Cladophlebis sinuata Holmes es bastante similar a C. mendozaensis y se distingue según Holmes (2003) por presentar la vena media de las pínnulas más delgada y sinuosa, mayor superposición en la distribución pinnular y diferente relación largo/ancho en la pínnula.
Clase GYMNOSPERMOPSIDA sensu Stewart y Rothwell, 1993
Orden CORYSTOSPERMALES Petriella 1981
Familia CORYSTOSPERMACEAE Thomas 1933
Género Dicroidium Gothan 1912 emend. Townrow 1957
Especie tipo. Dicroidium odontopteroides (Morris) Gothan 1912.
Dicroidium argenteum (Retallack) Gnaedinger y Herbst 2001 Figuras 3.E-G
Descripción. Fragmentos de frondes monopinnadas con raquis principal bifurcado de largo superior a los 9 cm. Pinnas subcirculares, de ápice obtuso-redondeado y base constricta, insertas en forma opuesta-subopuesta, con ángulo que oscila entre 70º y 90º. Largo entre 0,8 a 1,5 cm y ancho entre 0,4-0,8 cm. Venación odontopteroide; en las pinnas se observa una insipiente vena media.
Material estudiado. Perfil cerro Bayo: NF1: LPPB 11956a, 11956b, 11957a, 11957b, 11957c, 11958 al 11965, MCNAM-PB 1455 al 1459.
Comentarios. Esta especie fue considerada como una variedad de dos especies distintas. Así Retallack (1977) la incluye en D. odontopteroides y Petriella (1979) en D. crassum. Según Gnaedinger y Herbst (2001) el tratamiento de este taxón como una especie diferente se justifica por la forma pinnular (pinnas subcirculares, de ápice obtuso-redondeado y base constricta) claramente distinta de la presente en los taxones antes mencionados. Esta especie fue hallada en la Formación Potrerillos en cerro Cacheuta (Jain y Delevoryas, 1967 como Thinnfeldia dutoiti), Formación Cañadón Largo (Gnaedinger y Herbst, 1998a); Formación Los Rastros (Frenguelli, 1942a); Formación Las Breas, Chile (Gnaedinger y Herbst, 2001).
Dicroidium dubium (Feistmantel) Gothan 1912 Figura 3.H
Descripción. Fragmento de fronde pinnatífida. Pinnas de 1,2 cm de largo por 0,5 cm de ancho; distribuidas en forma alterna, presentan márgenes lobados a profundamente lobatisectos; ápice redondeado y venación aletopteriforme.
Material estudiado. Perfil cerro Bayo: NF2: LPPB 11966.
Comentarios. Se trata de un solo fragmento de fronde en donde los extremos distales de las pinnas están enteramente fusionadas mientras que la parte mediaproximal es profundamente lobada.
Dicroidium odontopteroides (Morris) Gothan 1912 Figura 4.A
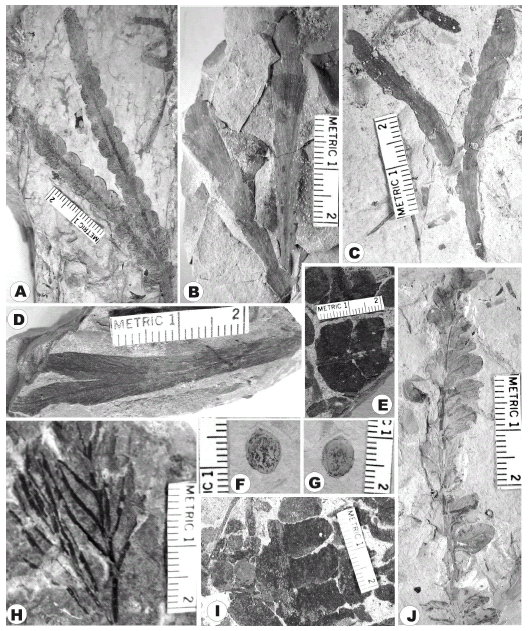
Figura 4. A, Dicroidium odontopteroides (Morris) Gothan, LPPB 12788b; B, Johnstonia stelzneriana (Geinitz) Frenguelli, LPPB 11969; C-D, Johnstonia coriacea (Johnston) Walkom, LPPB 11984; D, LPPB 11981a; E, Zuberia feistmanteli (Johnston) Frenguelli, LPPB 11990; F-G, Feruglioa samaroides Frenguelli, LPPB 12761b; G, LPPB12761a; H, Xylopteris elongata (Carruthers) Frenguelli, LPPB 11988; I, Zuberia feistmanteli, LPPB 11990; J, Pachydermophyllum praecordillerae (Frenguelli) Retallack, LPPB 11984.
Descripción. Fragmentos de frondes monopinnadas con raquis principal bifurcado, de largo superior a los 7 cm. Pinnas insertas por su ancho máximo, opuestas, subopuestas o alternas, con ápice redondeado. El largo oscila entre 0,3 a 1,8 cm, mientras que el ancho entre 0,4 - 0,7 cm, en la zona media de la hoja, observándose una relación largo/ancho menor a 3/1. La venación es odontopteroide, a veces con una vena media diferenciada en la zona proximal de la pinna.
Material estudiado. Perfil cerro Bayo: NF5: LPPB 11930a, 11930b; NF6: LPPB 11931 al 11933, MCNAM-PB: 1444, 1445; NF7: LPPB 11934, 11935, MCNAM-PB: 1446a, 1446b, 1447; NF8: LPPB 11936a, 11936b; NF9: LPPB 11937 al 11947, 11948a, 11948b, 11949 al 11951, MCNAM-PB 1448 al 1454; NF12: LPPB 11952 al 11955, 12788a, 12788b; Perfil cerro Cocodrilo: NF 2: LPPB 11898a, 11898b.
Comentarios. El material estudiado del cerro Bayo y cerro Cocodrilo presenta pinnas con una relación largo/ ancho inferior a 2/1 y se correspondería con D. odontopteroides var. odontopteroides (Morris) Gothan 1912 y con D. odontopteroides var. moltenense Retallack, 1977.
Género Johnstonia Walkom 1925
Especie tipo. Johnstonia coriaceae (Johnston) Walkom 1925.
Johnstonia stelzneriana (Geinitz) Frenguelli 1943 Figura 4.B
Descripción. Frondes bifurcadas de hasta 14 cm de largo, con lámina pinnatiserrada, margen dentado y lóbulos obtusos. Venación odontopteroide.
Material estudiado. Perfil cerro Bayo: NF3: LPPB 11901, 11967 al 11971; NF12: LPPB 11972; Perfil cerro Cocodrilo: NF3: LPPB 11973a, 11973b.
Comentarios. El material estudiado del cerro Bayo se corresponde con Johnstonia stelzneriana var. serrata Retallack 1977, mientras que en el cerro Cocodrilo se encontró un solo ejemplar fragmentario que se ajusta a J. stelzneriana var. stelzneriana (Geinitz) Frenguelli 1943.
Johnstonia coriacea (Johnston) Walkom 1925 Figuras 4.C, D
Descripción. Frondes bifurcadas de hasta 7 cm de largo, con lámina de márgenes enteros o levemente lobados de 0,4 cm de ancho. Venación rara vez visible, taeniopteroide.
Material estudiado. Perfil cerro Bayo: NF2: LPPB 11974, 11975; NF3: LPPB 11901, 11967, 11968, 11970, 11971, 11976 al 11980, 11981a, 11981b, MCNAM-PB 1460a, 1460b; NF 4: LPPB 11982, MCNAM-PB 1461; NF5: MCNAM-PB 1462a, 1462b, 1463; NF9: LPPB 11951; NF 11: LPPB 12797, MCNAM-PB 1494; NF12: LPPB 11907, 11952, 11983 al 11985, 12762, MCNAM-PB 1464; NF13: LPPB 11993.
Comentarios. El material estudiado se corresponde con Johnstonia coriacea var. coriacea (Johnston) Walkom, 1925 .
Género Xylopteris Frenguelli 1943
Especie tipo. Xylopteris elongata (Carruthers) Frenguelli 1943.
Xylopteris elongata (Carruthers) Frenguelli 1943 Figura 4.H
Descripción. Fragmentos de frondes monopinnadas de hasta 6 cm de largo; pecíolo de 1,5 cm de largo por 0,2 cm de ancho; ráquises primarios con pinnas de hasta 2 cm de largo por 0,1 cm de ancho; la distribución pinnular es espaciada, con una separación lateral de 0,4 cm. Venación no visible.
Material estudiado. Perfil cerro Bayo: NF9: LPPB 11986, 11948a, 11948b, MCNAM-PB 1465; NF12: LPPB 11987a, 11987b, 11988, 11989a, 11989b.
Comentarios. Las frondes presentan pinnas numerosas sobre el raquis primario. Este carácter permite asignar nuestro material a Xylopteris elongata y diferenciarlo de Xylopteris argentina (Kurtz) Frenguelli 1943, caracterizada por presentar un máximo de cuatro pinnas por raquis primario.
Género Zuberia Frenguelli 1943 emend. Artabe 1990
Especie tipo. Zuberia zuberi (Szajnocha) Frenguelli 1943.
Zuberia feistmanteli (Johnston) Frenguelli emend. Artabe 1990 Figuras 4.E, I
Descripción. Fragmentos de pinnas, con pínnulas subcuadrangulares, subromboidales o subtriangulares, de margen entero. Sus dimensiones oscilan entre 0,8-0,9 cm de ancho por 1-1,3 cm de largo.
Material estudiado. Perfil cerro Bayo: NF13: LPPB 11993, MCNAM-PB 1466; Perfil cerro Cocodrilo: NF3: LPPB 11994 al 11997, 11998a, 11998b, 11999; MCNAM-PB 1467 al 1470.
Comentarios. Se trata de material fragmentario correspondiente a porciones de pinnas, con pínnulas de gran tamaño de forma subcuadrangular o subromboidal y margen entero, y es en estos caracteres que se diferencia de las otras especies descriptas (Frenguelli, 1944d; Artabe, 1990).
Feruglioa Frenguelli 1944e
Especie tipo. Feruglioa samaroides Frenguelli 1944e
Feruglioa samaroides Frenguell, 1944e Figuras 4.F, G
Descripción. Semillas aladas, platispérmicas, con simetría bilateral y micrópila bífida. Miden entre 0,3-0,5 cm de ancho por 0,4-0,5 cm de largo.
Material estudiado. Perfil cerro Bayo: NF12: LPPB 11984, 12761a, 12761b.
Comparaciones. Las semillas aladas del género Feruglioa fueron encontradas en conexión orgánica con estróbilos laxos asignados a Fanerotheca Frenguelli 1944f (Anderson y Anderson, 2003). Este género revalidado por Anderson y Anderson (2003), fue anteriormente considerado como sinónimo de Antevsia por Townrow (1960) quien interpretó a los lóbulos de las cúpulas como sacos polínicos. Feruglioa samaroides fue descripta anteriormente en el cerro Cacheuta, Formación Potrerillos, NF1 y en la Formación Cacheuta NF4 (Morel, 1991)
Orden PELTASPERMALES Meyen 1987
Familia PELTASPERMACEAE Thomas 1933
Género Pachydermophyllum Thomas y Bose 1955
Especie tipo. Pachydermophyllum papillosum Thomas y Bose 1955.
Pachydermophyllum praecordillerae (Frenguelli) Retallack 1981 Figura 4.J
Descripción. Fragmentos de frondes monopinnadas de largo superior a los 8,5 cm y ancho aproximado de 3,2 cm; pinnas opuestas, subopuestas; las de la parte media son linear-lanceoladas, con ápice redondeado o agudo y base asimétrica; el margen es entero, siendo el acroscópico contraído, formando un lóbulo, y el basiscópico decurrente; miden hasta 2 cm de largo por 0,5 cm de ancho; se insertan con un ángulo de 60º-70º en la parte basal-media de la fronde. La venación es aletopteroide; la vena media llega casi hasta el extremo distal de la pinna donde se resuelve en varias dicotomías; las venas secundarias salen con ángulo agudo y se bifurcan más de una vez. El raquis mide 0,3 cm.
Material estudiado. Perfil cerro Bayo: NF12: LPPB 11983, 11984, 12762; NF13: LPPB 11993, 12763a, 12763b.
Comparaciones. De las cinco especies reconocidas por Retallack (1981) sólo tres están presentes en Argentina. P. dubium (Burgues) Rettallack 1981 fue descripta para la Formación Potrerillos (Morel, 1994), mientras que Pachydermophyllum pinnatum Retallack 1981 fue hallada en la Formación Llantenes (Artabe et al., 1999b). P. praecordillerae se diferencia de ambas, porque sus pinnas son lineares, con frecuente auriculación en la base, posición oblicua y notable reducción de las pinnas basales.
Orden CYCADALES Dumortier, 1829
Género Kurtziana Frenguelli emend. Petriella y Arrondo 1982
Especie tipo. Kurtziana cacheutensis (Kurtz) Frenguelli 1942b.
Kurtziana cacheutensis (Kurtz) Frenguelli, 1942b Figuras 5.A, B

Figura 5. A-B, Kurtziana cacheutensis (Kurtz) Frenguelli emend. Petriella y Arrondo, LPPB, 12765; B, LPPB 12764; C, Baiera africana Baldoni, LPPB 12776; D, Yabeiella brackebuschiana (Kurtz) Ôishi, LPPB 12790a; E, Yabeiella mareyesiaca (Geinitz) Ôishi, LPPB 12781; F, Yabeiella spathulata Ôishi, LPPB 12789; G, Sphenobaiera argentinae (Kurtz) Frenguelli, LPPB 12771; H, Rochipteris sp., LPPB 12777a; I, Fraxinopsis andium (Frenguelli) Anderson y Anderson, LPPB 12792a; J, Baiera cuyana Frenguelli, LPPB 12772a; K, Gontriglossa sp., LPPB 12793.
Descripción. Hojas monopinnadas, imparipinnadas de hasta 9 cm de largo por 4,5 cm de ancho. Las pinnas son linear-oblongas, se insertan a los lados del raquis, con un ángulo que oscila entre 55º-60º, en forma alterna. Las de la zona distal miden 1,5 cm de largo por 0,7 cm de ancho y las de la media 2,5 cm de largo por 1,1 cm de ancho. El sector basal de la pinna es asimétrico, el margen acroscópico se contrae hasta alcanzar la vena media, el margen basiscópico es recto o levemente contraído formando un lóbulo basal. Los márgenes laterales son enteros, subparalelos y confluyen en un ápice redondeado u obtuso. La venación es aletopterídea. La vena media alcanza el tercio distal de la pinna y se resuelve en varias dicotomías. Las venas laterales se bifurcan en dos dicotomías sucesivas. El raquis primario es estriado y mide hasta 0,2 cm.
Material estudiado. Perfil cerro Bayo: NF2: LPPB 11966, 12764; NF3: LPPB 12765 al 12768; MCNAM-PB 1471; NF5: LPPB 11921, 12769, 12770, MCNAM-PB 1472 al 1476; NF9: LPPB 12796.
Comparaciones. En Argentina se reconocen dos especies de Kurtziana: K. brandmayri y K. cacheutensis que se diferencian porque la primera posee frondes de mayor tamaño, raquis más robusto, pinnas sésiles, densas, a veces imbricadas, distribuidas sobre la superficie del raquis. K. cacheutensis presenta frondes más pequeñas, con pinnas subpecioladas o pecioladas, insertas a los lados del raquis, sin superposición de las mismas. El género Kurtziana - creado por Frenguelli (1942b)- fue enmendado por Petriella y Arrondo (1982), quienes adicionan caracteres megascópicos y epidérmicos. Artabe et al. (1991) sugieren la filiación sistemática con Cycadales sobre la base del estudio cuticular de una de sus especies (K. brandmayri). Más recientemente, Herbst y Gnaedinger (2002) retienen al género Kurtziana para frondes sin cutícula, crean una nueva especie K. paipotensis para el Triásico de Chile y segregan a las formas con cutícula en un nuevo género Alicurana. Por último, Anderson y Anderson (2003) incluyen en Kurtziana formas asignables a Pachydermophyllum y reconocen para Sudáfrica 16 especies aún no formalmente descriptas. A su vez, estos últimos autores asignan a Kurtziana (= Pachydermophyllum) al Orden Matatiellales por haber encontrado hojas en asociación estrecha con estructuras reproductivas femeninas (Matatiella). Holmes y Anderson (2005) reconocen en Australia dos "complejos" basados en las dos especies originalmente establecidas por Frenguelli (1942b). Teniendo en cuenta que existen diferencias morfológicas entre Kurtziana y Pachydermophyllum, se sigue en este trabajo una posición conservadora y se los considera como entidades genéricas diferentes.
Orden GINKGOALES sensu Stewart y Rothwell 1993
Género Sphenobaiera (Florin) Harris y Millington 1974
Especie tipo. Sphenobaiera spectabilis (Nathorst) Florin 1936.
Sphenobaiera argentinae (Kurtz) Frenguelli 1946 Figura 5.G
Descripción. Hojas de forma triangular que miden 0,9 cm de ancho por 5,4 cm de largo. Ángulo basal de 10º. La lámina se divide por una incisión leve en dos segmentos divergentes. Venación dicotomizada.
Material estudiado. Perfil cerro Bayo: NF12: LPPB 11984, 12771.
Comparaciones. Por tratarse de hojas pequeñas, triangulares, divididas en dos segmentos similares por una única incisión se asignaron a S. argentinae.
Género Baiera Braun emend. Florin 1936
Especie tipo. Baiera muensteriana (Presl) Florin 1936.
Baiera africana Baldoni 1980 Figura 5.C
Descripción. Hojas pecioladas de contorno triangular, de hasta 10 cm de largo por 2 cm de ancho, con un ángulo basal de 20° aproximadamente. Hendidas por una incisión que las divide en dos hemiláminas. Estas últimas vuelven a bifurcarse hasta dos veces formando segmentos distales con márgenes paralelos, de hasta 0,1 cm de ancho. Venación no visible.
Material estudiado. Perfil cerro Bayo: NF12: LPPB 11907, 11954, 12775, 12776, MCNAM-PB 1479.
Comparaciones. De las especies mencionadas para Argentina, las más afines serían B. furcata y B. cuyana. De la primera se diferencia por poseer hojas de menor tamaño, menos divididas. De la segunda por tener los pecíolos más largos y hojas con dicotomías que determinan segmentos filiformes de ancho menor.
Baiera cuyana Frenguelli 1942c Figura 5.J
Descripción. Hojas cortamente pecioladas (0,2 cm), de contorno triangular, de hasta 2,6 cm de largo por 3,5 cm de ancho, con un ángulo basal de 60° aproximadamente. La lámina se divide dicotómicamente cinco veces. La primera división es profunda y alcanza la base de la hoja dividiéndola en dos hemiláminas aparentemente asimétricas. Estas últimas vuelven a dividirse hasta cuatro veces formando segmentos distales con márgenes paralelos, de hasta 0,1 cm de ancho. La última división es muy cercana al extremo distal de la hoja formando lacinias de 0,2 cm de largo de ápice redondeado. Venación no visible.
Material estudiado. Perfil cerro Bayo: NF2: LPPB 11966, 11974, 12772a, 12772b, 12773a, 12773b, MCNAM-PB 1477, 1478; NF3: LPPB 12774
Comparaciones. De las especies mencionadas para Argentina, las más afines serían B. furcata (Lindley y Hutton) Braun 1843 y B. africana Baldoni 1980. De la primera se diferencia por poseer hojas de menor tamaño, menos divididas. De la segunda por presentar pecíolos más cortos y hojas con dicotomías que determinan segmentos filiformes de ancho mayor.
Orden PETRIELLALES Taylor et al. 1994
Género Rochipteris Herbst, Troncoso y Gnaedinger 2001
Especie tipo. Rochipteris lacerata (Arber) Herbst et al. 2001.
Rochipteris sp. Figura 5.H
Descripción. Hojas apecioladas, triangulares, con ápice levemente curvado y márgenes laterales rectos. Ángulo basal de 20º-30º. Mide 3,5 cm de largo por 1,6 cm de ancho total en el ápice. Venas dicotomizadas, frecuentemente anastomosadas en toda la lámina, formando aréolas angostas, romboidales y fusiformes, alargadas en la mitad de la hoja y más cortas en la zona apical. La densidad de venación es 33 venas por cm en la zona apical.
Material estudiado. Perfil cerro Bayo: NF12: LPPB 1277a, 12777b, 12778, MCNAM-PB 1480a, 1480b.
Comparaciones. El género Rochipteris fue creado por Herbst et al. (2001) para incluir algunas hojas gondwánicas previamente asignadas a Chiropteris Kurr ex Bronn (1858) o a Ginkgophytopsis Høeg, 1967 (Seward, 1903; Arber, 1917; du Toit, 1927; Frenguelli, 1942d; Retallack, 1980). Este género incluye hojas apecioladas, cuneadas, elongadas a flabeliformes, con márgenes desde enteros hasta profundamente divididos y con venación anastomosada. Se reconocen siete especies: R. cuneata, R. copiapensis, R. chilensis, R. lacerata, R. alexandriana, R. etheridgei, R. ginkgoides en Argentina, Chile y Australia (Herbst et al., 2001; Barone-Nugent et al., 2003). Las hojas encontradas se asemejan a R. cuneata (Carruthers) Herbst et al. 2001 aunque presentan menores dimensiones y margen distal recto. El la Argentina R. cuneata fue hallada en la Formación Barreal (Bonetti, 1963) y en Cañadón Largo (Herbst et al., 2001). Formas comparables a Rochipteris fueron encontradas en conexión orgánica con estructuras reproductivas femeninas (Kannaskoppia / Kannaskoppifolia) y masculinas (Kannaskoppianthus / Kannaskoppifolia) y asignadas al Orden Petriellales y a la Familia Kannaskoppiaceae por Anderson y Anderson (2003).
Orden GNETALES sensu Stewart y Rothwell 1993
Género Yabeiella Ôishi 1931
Especie tipo. Yabeiella mareyesiaca (Geinitz) Ôishi 1931.
Yabeiella mareyesiaca (Geinitz) Ôishi 1931 Figura 5.E
Descripción. Hojas simples, coriaceas, lanceoladas de largo superior a los 15 cm. El ancho oscila entre 2,5-5,5 cm, en la parte media, más frecuentemente miden entre 3 y 3,5 cm. El ápice se observa en pocos ejemplares y es agudo o redondeado y la base se angosta gradualmente. La vena media es ancha, estriada y mide hasta 0,3 cm. Las venas secundarias están bifurcadas a distintos niveles y algunas sufren anastomosis; respecto de la vena principal salen con un ángulo que oscila entre 70º y 80º; en la zona media de la lámina se cuentan alrededor de 20 venas por cm. Al llegar al margen las venas secundarias se anastomosan con las venas vecinas formando arcos, lo que sugiere la presencia de una vena colectora marginal.
Material estudiado. Perfil cerro Bayo: NF2: LPPB 11974, 12779, MCNAM-PB 1481 al 1483; NF3: LPPB 11900, 11901, 11967, 11977, 11978a, 11978b, 11979, 11980, 12780 al 12786, MCNAM-PB 1484 al 1490; NF4: LPPB 12787; NF5: MCNAM-PB 1491.
Comparaciones. No existe acuerdo en el número de especies que se reconocen dentro del género. Así, Anderson y Anderson (1989) asignan dos especies al género (Y. mareyesiaca, Y. brackebuschiana), mientras que Stipanicic et al. (1996) determinan seis (Y. mareyesiaca, Y. brackebuschiana, Y. crassa, Y. du-toiti, Y. spathulata y Y. wielandii) proponiéndose incluso variedades de ellas. Aunque el género está ampliamente distribuido en todo el Triásico de Gondwana, las formas más abundantes corresponden a Y. mareyesiaca y a Y. brackebuschiana (Spalletti et al., 1999). Y. mareyesiaca se diferencia por presentar hojas de grandes dimensiones, de forma lanceolada; las venas secundarias se insertan con ángulo agudo a recto y la densidad aproximada es de 18 venas por cm.
La filiación sistemática del género ha sido objeto de controversia y se lo ha asignado tanto a las Cycadales (Ganuza et al., 1998) como a las Bennettitales (Jain y Delevoryas, 1967; Taylor y Hickey, 1992). Basándose en la asociación repetida de Yabeiella con Fraxinopsis Wieland en Australia, Sudáfrica y Argentina, en los caracteres de arquitectura foliar -presentan patrones semejantes a los de las angiospermas basales-, y en análisis filogenéticos recientes (Nixon et al., 1994), Axsmith et al. (1997) vinculan al género con el clado de las antophytas. Anderson y Anderson (2003) consideran que la venación y los caracteres cuticulares vinculan al género Yabeiella con las Gnetopsidas y asignan la planta Fraxinopsis/ Yabeiella a un nuevo orden: Fraxinopsiales y una nueva familia: Fraxinopsiaceae.
Yabeiella brackebuschiana (Kurtz) Ôishi 1931 Figura 5.D
Descripción. Fragmentos de hojas enteras, lineal-lanceolada de 6,5 cm de largo por 0,8 cm de ancho. La vena media es fuerte de 0, 1 cm de ancho con estrías longitudinales. Las venas laterales salen de la vena media en ángulos de aproximadamente 65º; las venas son simples o dicotomizadas. Se observan pocas anastomosis. La densidad en la parte media de la hoja es de 12 venas por cm. Todas las venas laterales confluyen en una vena colectora marginal.
Material estudiado. Perfil cerro Bayo: NF12: LPPB 11928a, 11928b, 12790a, 12790b, 12791, MCNAM-PB 1492.
Comentarios. Aunque el ejemplar estudiado no conserva el ápice se asigna a Y. brackebuschiana por presentar una hoja linear con márgenes paralelos diferente del presente en Yabeiella spathulata donde la lámina es lineal-espatulada.
Yabeiella spathulata Ôishi 1931 Figura 5.F
Descripción. Hoja entera, lineal-espatulada, de 5,5 cm de largo con un ancho máximo en la zona distal de 0,9 cm, con ápice redondeado. La vena media es fuerte, de 0, 1 cm de ancho con estrías longitudinales. Las venas laterales salen de la vena media con ángulos que oscilan entre 55º-60º; las venas son simples o dicotomizadas, principalmente las del sector medio y distal de la hoja. Se observan pocas anastomosis. La densidad en la parte media de la hoja es de 12 venas por cm. Todas las venas laterales confluyen en una vena colectora marginal.
Material estudiado. Perfil cerro Bayo: NF5: LPPB 12789.
Comentarios. Yabeiella spathulata se diferencia de Y. brackebuschina por la forma de la lámina (lineal-espatulada con el ápice redondeado) y por la venación (presenta una fuerte vena media y venas laterales simples o bifurcadas con escasas anastomosis).
Género Fraxinopsis Wieland 1929
Especie tipo. Fraxinopsis minor Wieland 1929.
Fraxinopsis andium (Frenguelli) Anderson y Anderson 2003 Figura 5.I
Descripción. Semilla alada de 0,4 cm de ancho por 1,5 cm de largo. Semilla pequeña de 0,2 cm de ancho por 0,5 cm de largo con una quilla longitudinal; ala elíptica con venación paralela; se observan 9 venas; 7 centrales largas (dos están bifurcadas en la zona distal) y dos laterales cortas.
Material estudiado. Perfil cerro Bayo: NF12: LPPB 12792a, 12792b.
Comparaciones. Fraxinopsis andium presenta semillas aladas que oscilan entre 1,2-2,1 cm de largo, con alas elípticas con 9 venas -7 centrales y dos marginalesocasionalmente bifurcadas y anastomosadas y semillas con quilla central y pequeñas aurículas. Entre las restantes especies reconocidas: F. minor, F. major Wieland, F. auriculata Anderson y Anderson y F. cornicoides Anderson y Anderson (Frenguelli, 1941, 1944g; Anderson y Anderson, 2003) la especie más afin es F. minor. Esta última se diferencia porque presenta un ala con márgenes paralelos y venación simple (generalmente 7 venas). F. major tiene mayores dimensiones, un ala elíptica con venas sucesivamente dicotomizadas y con repetidas anastomosis. Ganuza et al. (1995) describen e ilustran para la Formación Paso Flores a F. andium y F. major. Como fuera indicado por Anderson y Anderson (2003) el material LPPB 12538b (citado como F. andium) por sus mayores dimensiones podría corresponder a F. cornicordes Anderson y Anderson 2003.
Género Gontriglossa Anderson y Anderson 1989
Especie tipo. Gontriglossa verticillata (Thomas) Anderson y Anderson 1989.
Gontriglossa sp. Figura 5.K
Descripción. Fragmentos de hojas elípticas, con ápice y base desconocida, margen entero a suavemente ondulado, de largo superior a los 12 cm y 2,5 cm de ancho. Vena media marcada, estriada, de 0,25 cm en la zona media y adelgazándose hacia el ápice. Venación lateral reticulada. En el tercio proximal las venas laterales se bifurcan y mantienen un ángulo agudo respecto de la vena media. En los tercios medio y distal la venación es reticulada y perpendicular a la vena media con mallas que disminuyen de tamaño hacia el margen.
Material estudiado. Perfil cerro Bayo: NF4: LPPB 12793; NF8: LPPB 11906, 12794, 12795, MCNAM-PB 1493.
Comentarios. Los ejemplares estudiados concuerdan con los descriptos por Jain y Delevoryas (1967) y Morel (1991) para el cerro Cacheuta. Aunque se desconoce la forma de inserción de estas hojas, se han encontrado segmentos de hojas (LP-PB 11821) que tienden a converger en sus bases, lo que apoyaría la idea de una distribución verticilada. La especie más afín, Gontriglossa verticilata fue asignada a las gnetopsidas, sobre la base de los caracteres cuticulares de las hojas y el hallazgo de estructuras reproductivas asociadas. G. verticillata se diferencia del taxón de la Formación Potrerillos (cerro Bayo) en que la venación lateral reticulada presenta un ángulo de aproximadamente 50º respecto de la vena media. Otros géneros posiblemente vinculados, con hojas elípticas y venación lateral reticulada, Santaecruzia Gnaedinger y Herbst, 1998b y Cetiglossa Anderson y Anderson 2003, se diferencian de Gontriglossa por presentar hojas de mayor tamaño.
Discusión y conclusiones
Aun cuando el área de Potrerillos es clave para el estudio del Triásico del oeste argentino, no se cuenta hasta el momento con estudios sistemáticos de la paleoflora en el perfil tipo de la Formación Potrerillos, por lo que los taxones descriptos en este trabajo son nuevos para la localidad.
En lo que hace al conocimiento paleobotánico integral de la Formación Potrerillos en la Cuenca Cuyana, en este trabajo se identifican y describen por vez primera ejemplares de Zuberia feistmanteli, Baiera africana y Rochipteris sp.
Desde el punto de vista bioestratigráfico y sobre la base del análisis de las tafocenosis en relación con los paleoambientes sedimentarios (cf. Spalletti et al., 1999, 2005), en la Formación Potrerillos se reconocen dos biozonas de asociación en concordancia y continuidad secuencial. La Biozona MBC se atribuye al Triásico Medio Tardío, mientras que la Biozona BNP se asigna al Triásico Tardío Temprano.
En las tablas 3 y 4 se puede apreciar la distribución estratigráfica y regional (Argentina y Chile) de los taxones identificados en la Formación Potrerillos. Nótese que por el criterio de presencia las mayores afinidades en la Argentina se dan con las formaciones Potrerillos (otras áreas de afloramiento), El Alcázar, Cortaderita y Los Rastros. Con respecto a las paleofloras de Chile los elementos comunes son escasos (tabla 4) muy posiblemente debido a que la Formación Potrerillos corresponde al Triásico Medio Tardío-Triásico Tardío Temprano, en tanto que las unidades chilenas se ubican con preferencia en el Triásico Tardío.
Tabla 3. Lista paleoflorística de la Formación Potrerillos en los cerros Bayo y Cocodrilo. Comparaciones con otras unidades estratigráficas. A, Formación Cerro de las Cabras; B, Formación Potrerillos en Cerro Cacheuta; C, Formación Cacheuta; D, Formación Río Blanco; E, Formación Chihuido; F, Formación Llantenes; G, Formación Cañadón Largo; H, Formación Laguna Colorada; I, Formación Vera; J, Formación Paso Flores; K, Formación Ischichuca; L, Formación Los Rastros; M, Formación Ischigualasto; N, Formación Agua de los Pajaritos; O, Formación El Alcázar; P, Formación Hilario; Q, Formación Barreal; R, Formación Cortaderita; S, Formación Quebrada de la Mina; T, Formación Carrizal / palaeofloristic list of the Potrerillos Formation at Bayo and Cocodrilo Hills. Comparisons with other stratigraphic units. A, Cerro de las Cabras Formation; B, Potrerillos Formation at Cacheuta Hill; C, Cacheuta Formation; D, Río Blanco Formation; E, Chihuido Formation; F, Llantenes Formation; G, Cañadón Largo Formation; H, Laguna Colorada Formation; I, Vera Formation; J, Paso Flores Formation; K, Ischichuca Formation; L, Los Rastros Formation; M, Ischigualasto Formation; N, Agua de los Pajaritos Formation; O, El Alcázar Formation; P, Hilario Formation; Q, Barreal Formation; R, Cortaderita Formation; S, Quebrada de la Mina Formation; T, Carrizal Formation.
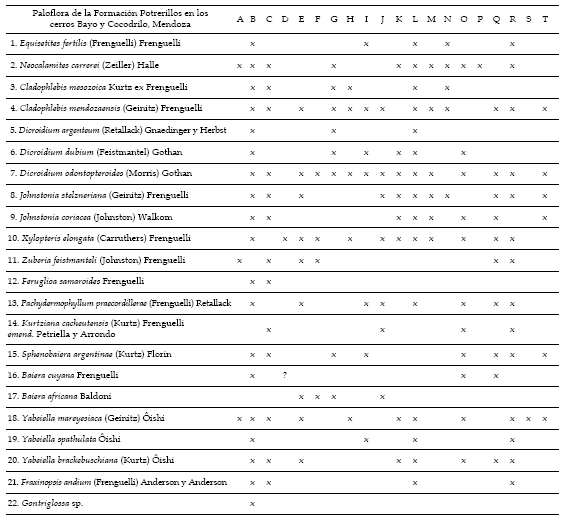
Tabla 4. Lista florística para las unidades triásicas chilenas. 1. Formación La Ternera, 2. Formación San Felix (Alto del Carmen), 3. Formación Las Breas, 4. Formación El Puquén, 5. Formación Panguipulli, 6. Formación Tralcán, 7. Formación Cajón Troncoso, 8. Formación Santa Juana. Las determinaciones fueron realizadas por Stipanicic (1983), Mohr y Schöner (1985), Arrondo et al. (1988), Lutz et al. (1999, 2001, 2003), Troncoso y Herbst (1999, 2000), Herbst (2000), Herbst y Troncoso (2000), Herbst et al. (1998, 2001, 2005), Gnaedinger y Herbst (2001, 2004a, 2004b), Leppe y Moisan (2003) y Leppe et al. (2006). Sólo los taxones resaltados están presentes en la Formación Potrerillos aflorante en los cerros Bayo y Cocodrilo, Mendoza, Argentina / floristic list of Chilean Triassic units. 1. La Ternera Formation, 2. San Felix (Alto del Carmen) Formation, 3. Las Breas Formation, 4. El Puquén Formation, 5. Panguipulli Formation, 6. Tralcán Formation, 7. Cajón Troncoso Formation, 8. Santa Juana Formation. The systematic assignments were made by Stipanicic (1983), Mohr and Schöner (1985), Arrondo et al. (1988), Lutz et al. (1999, 2001, 2003), Troncoso and Herbst (1999, 2000), Herbst (2000), Herbst and Troncoso (2000), Herbst et al. (1998, 2001, 2005), Gnaedinger and Herbst (2001, 2004a, 2004b) and Leppe and Moisan (2003). Only the taxa in bold are present in the Potrerillos Formation at Bayo and Cocodrilo Hills, Mendoza, Argentina.
Desde el punto de vista paleoclimático y paleogeográfico, las floras analizadas se ubican en la zona subtropical seca del Área Extratropical del Sudoeste de Gondwana (Artabe et al., 2003). Se infiere un régimen climático megamonzónico a partir de la presencia de grupos con adaptaciones estacionales y xeromórficas (Artabe et al., 2003; Spalletti et al., 2003). En tal contexto, y de acuerdo a lo indicado por Spalletti et al. (2005), las comunidades estudiadas se desarrollaron en ambientes sedimentarios dominantemente fluviales, en especial en las áreas bien drenadas de las planicies de inundación, en las vecindades de los canales principales y en los lóbulos de desbordamiento de cursos de moderada a alta sinuosidad.
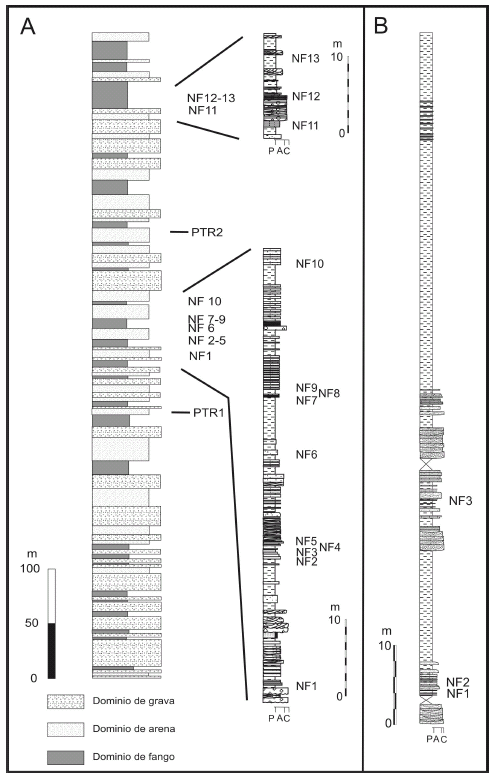
Apéndice 1. A, Perfil general de la Formación Potrerillos en el Cerro Bayo, acompañado por secciones de detalle en las que se muestran los niveles con plantas fósiles. PTR1 y PTR 2 son muestras de tobas que han sido datas por Spalletti et al. (en prensa) en 239.7 ± 2.2 Ma y 230.3 ± 2.3 Ma (U-Pb SHRIMP sobre circones) / A, Simplified log of the Potrerillos Formation at Cerro Bayo showing the location of fossiliferous plant levels. PTR1 and PTR 2 are tuff samples dated by Spalletti et al. (in press) using U-Pb SHRIMP on zircon grains in 239.7 ± 2.2 Ma and 230.3 ± 2.3 Ma, respectively. B, Perfil estratigráfico de la parte superior de la Formación Potrerillos (por encima del conglomerado rojo) y de la Formación Cacheuta en el faldeo sur del Cerro Cocodrilo / B, Log of the upper section of the Potrerillos Formation (above the red conglomerate) and the Cacheuta Formation in the southern side of Cerro Cocodrilo.
Agradecimientos
El presente trabajo se realizó en el marco de proyectos financiados por la Agencia Nacional de Promoción Científica y Tecnológica, Argentina (PICT 07-08467), Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 5430) y Universidad Nacional de La Plata, Argentina (Proyecto N 412 y N 458). Al Sr. Sergio Londero (Profesional de Apoyo del Consejo Nacional de Investigaciones Científicas y Técnicas, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, Centro Regional de Investigaciones Científicas y Tecnológicas - Consejo Nacional de Investigaciones Científicas y Técnicas.) por la asistencia en las primeras tareas de campo para recolección de muestras y a E. Cerdeño, curadora del Museo de Ciencias Naturales y Antropológicas "J.C. Moyano", por el préstamo de materiales para su estudio.
Referencias
1. Anderson, J.M. y Anderson, H.M. 1983. Palaeoflora of Southern Africa Molteno Formation (Triassic). Volume 1, Parts 1-2. Introduction: Dicroidium. A.A. Balkema, Rotterdam, 227 pp. [ Links ]
2. Anderson, J.M. y Anderson, H.M. 1989. Paleoflora of Southern Africa. Molteno Formation (Triassic). Volume 2 Gymnosperms (excluding Dicroidium). A.A. Balkema, Rotterdam, 567 pp. [ Links ]
3. Anderson, J.M. y Anderson, H.M. 2003. Heyday of Gymnosperms: systematics and biodiversity of the Late Triassic Molteno fructifications. Strelitzia 15: 1-398. [ Links ]
4. Anderson, J.M., Anderson, H.M. y Cruickshank, A.R.I. 1998. Late Triassic ecosystems of the Molteno/Lower Elliot Biome of Southern Africa. Palaeontology 41: 387-421. [ Links ]
5. Arber, E.A.N. 1917. Earlier Mesozoic flora of New Zealand. New Zealand Geological Survey, Paleontological Bulletin 6: 1-80. [ Links ]
6. Arrondo, O.G., Morel, E.M., Ganuza, D.G., Hervé, F., Parada, M. y Muehlebach, W. 1988. La Formación Panguipulli y su contenido paleoflorístico, Chile. 5º Congreso Geológico Chileno (Santiago), Actas 1: 137-145. [ Links ]
7. Artabe, A.E. 1990. Revalidación del género triásico Zuberia Frenguelli 1943, Familia Corystospermaceae. Revista del Museo de La Plata, nueva serie, Paleontología 9: 145-157. [ Links ]
8. Artabe, A.E. y Brea, M. 2003. A new approach to Corystospermales based on petrified stems from the Triassic of Argentina. Alcheringa 27: 209-229. [ Links ]
9. Artabe, A.E. y Zamuner A.B. 1991. Una nueva Equisetal del Triásico de Cacheuta, Argentina, con estructura interna preservada. Ameghiniana 28: 287-294. [ Links ]
10. Artabe, A. E., Zamuner, A. B. y Archangelsky, S. 1991. Estudios cuticulares en Cycadópsidas fósiles. El género Kurtziana Frenguelli 1942. Ameghiniana 28: 365-374. [ Links ]
11. Artabe, A.E., Brea, M. y Zamuner, A.B. 1999a. Rhexoxylon brunoi Artabe, Brea et Zamuner, nov. sp., a new Triassic Corystosperm from the Paramillo de Uspallata, Mendoza, Argentina. Review of Palaeobotany and Palynology 105: 63-74. [ Links ]
12. Artabe, A.E., Morel, E.M., Spalletti, L.A. y Brea, M. 1999b. Paleoambientes sedimentarios y paleoflora asociada en el Triásico superior de Malargüe (Sur de Mendoza, Argentina). Revista de la Asociación Geológica Argentina 53: 526-548. [ Links ]
13. Artabe, A.E., Morel, E.M. y Spalletti, L.A. 2001. Paleoecología de las floras triásicas argentinas. En: A.E. Artabe, E.M. Morel, y A.B. Zamuner (eds.), El Sistema Triásico de Argentina. Fundación Museo de La Plata "Francisco Pascasio Moreno", pp. 199-225. [ Links ]
14. Artabe, A.E., Morel, E.M. y Spalletti, L.A. 2003. Caracterización de las Provincias Fitogeográficas Triásicas del Gondwana Extratropical. Ameghiniana 40: 387-405. [ Links ]
15. Axsmith, B.J., Taylor, T.N., Fraser, N.C. y Olse, P.E. 1997. An occurrence of the gondwanan plant Fraxinopsis in the Upper Triassic of Eastern North America. Modern Geology 21: 299-308. [ Links ]
16. Axsmith, B.J., Taylor, T.N. y Taylor, E.L. 1998. Anatomically preserved leaves of the conifer Notophytum krauselii (Podocarpaceae) from the Triassic of Antartica. American Journal of Botany 85: 704-713. [ Links ]
17. Axsmith, B.J., Taylor, E.L., Taylor, T.N. y Cúneo, R.N. 2000. New perspectives on the Mesozoic seed fern Order Corystospermales based on attached organs from the Triassic of Antarctica. American Journal of Botany 87: 757-768. [ Links ]
18. Baldoni, A. 1980. Baiera africana, una nueva especie de Ginkgoal del Triásico de Sudáfrica. Ameghiniana 17: 156-162. [ Links ]
19. Barone-Nugent, E. D, McLoughlin, S. y Drinnan, A. N. 2003. New species of Rochipteris from the Upper Triassic of Australia. Review of Palaeobotany and Palynology 123: 273-287. [ Links ]
20. Bertchold, B. W. von y Presl, J. S. 1820. Pfirozenosti rostlin 1: pp. 272. [ Links ]
21. Bonetti, M.I.R. 1963. [Contribución al conocimiento de la Flora fósil de Barreal, Departamento de Calingasta (provincia de San Juan. Tesis Doctoral. Facultad de Ciencias Exactas, Físicas y Naturales. Universidad Nacional de Buenos Aires, 246 pp. Inédita]. [ Links ]
22. Borrello, A.V. 1942. [Estratigrafía y tectónica del Triásico Retiense en los alrededores de Potrerillos, provincia de Mendoza. Tesis Doctoral Nº 32, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 181 pp. Inédita]. [ Links ]
23. Borrello, A.V. 1962. Fanglomerado Río Mendoza (Triásico-Provincia de Mendoza). Notas Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, 1: 3-9. [ Links ]
24. Boureau, E., 1964. Traité de Paléobotanique. Sphenophyta Noeggerathiophyta, 3 : 1-544. Masson et Cie. Paris. [ Links ]
25. Braun, C.F.W. 1843. Beiträge zur Urgeschichte der Pflanzen. En: Münster Graf. G. zu: Beiträge (ed.) Petrefactenkunde, Bayreuth, 6 : 5-25. [ Links ]
26. Brea, M. 1995. [Estudio de la paleoflora de la secuencia triásica de Agua de la Zorra, provincia de Mendoza. Tesis Doctoral N° 642, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 202 pp. Inédita]. [ Links ]
27. Brea, M. 1997. Una nueva especie fósil del género Araucarioxylon Kraus 1870 emend. Maheshwari 1972 del Triásico de Agua de la Zorra, Uspallata, Mendoza, Argentina. Ameghiniana 34: 485- 496. [ Links ]
28. Brea, M. 2000. Paleoflora triásica de Agua de la Zorra, Uspallata, provincia de Mendoza, Argentina: Lycophyta y Filicophyta. Ameghiniana 37: 199-204. [ Links ]
29. Brea, M. y Artabe, A.E. 1999. Apocalamitaceae (Sphenophyta) triásicas de la Formación Paramillo, Agua de la Zorra, provincia de Mendoza, Argentina. Ameghiniana 36: 389-400. [ Links ]
30. Brongniart, A. 1849. Tableau des genres de végétaux fossiles considerés sous le point de vue de leur classification botanique et de leur distribution geólogique. Dictionnaire universel de Histoire Naturelle 13 : 1-127. Paris. [ Links ]
31. Bronn, H.G. 1858. Beiträge zur Triasischen Fauna und Flora der bituminosen Schiefer von Raibl nebst anhang uber die KURR´sche Sippe Chiropteris aus dem Lettenkohlen-sandsteinen. Neues Jahrbuch für Mineralogie, Geologie und Paläeontologie 1: 129-144. [ Links ]
32. Cúneo, R.N., Taylor, E.L., Taylor, T.N. y Krings, M. 2003. In situ fossil forest from the upper Fremouw Formation (Triassic) of Antarctica: paleoenvironmental setting and paleoclimate analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 197: 239-261. [ Links ]
33. Dumortier, B.Ch.J. 1829. Analyse des Familles de Plantes: Avec l' indication des principaux genres qui s'y ratachent. Imprimerie de J. Casterman, Tournay. 104 pp. [ Links ]
34. du Toit, A. L. 1927. The fossil flora of the Upper Karroo beds. Part. 2. Annals of the South African Museum 22: 289-420. [ Links ]
35. Fossa Mancini, E. 1937. Las investigaciones geológicas de YPF en la Provincia de Mendoza y algunos problemas de estratigrafía regional. Boletín de Informaciones Petroleras, vol. 14, N° 154, p. 51-118. [ Links ]
36. Florin, R. 1936. Die fossilen Ginkgophyten von Franz-Joseph-Lang nebst Erörterungen über vermeintliche Cordaitales mesozoischen Alters, 2. Allgemeiner Teil. Palaeontographica 82 B: 1-71. [ Links ]
37. Frenguelli, J. 1941. Sobre Cycadocarpidium andium n. sp., del Rético de Cacheuta, Mendoza. Notas del Museo de La Plata, Paleontología 6: 485-498. [ Links ]
38. Frenguelli, J. 1942a. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. 1. Harringtonia argentinica (Arber) n. gen. Notas del Museo de La Plata, Paleontología 7: 265-273. [ Links ]
39. Frenguelli, J. 1942b. Contribución al conocimiento de la flora del gondwana superior en la Argentina. 9. Kurtziana cacheutensis Kurtz sp., n. gen et n. comb. Notas del Museo de La Plata, Paleontología 7: 332-338. [ Links ]
40. Frenguelli, J. 1942c. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. 4. Baiera cuyana n. sp. Notas del Museo de La Plata, Paleontología 7: 298-302. [ Links ]
41. Frenguelli, J. 1942d. Contribución al conocimiento de la flora del gondwana superior en la Argentina. 10. Chiropteris barrealensis n. sp. Notas del Museo de La Plata, Paleontología 7: 341-353. [ Links ]
42. Frenguelli, J. 1943. Reseña crítica de los géneros atribuidos a la "Serie de Thinnfeldia". Revista del Museo de La Plata nueva serie, Paleontología 2: 225-342. [ Links ]
43. Frenguelli, J. 1944a. Contribuciones al conocimiento de la flora del gondwana superior en la Argentina. 24. Equisetites fertilis n. comb. Notas Museo La Plata, Paleontología 9: 501-509. [ Links ]
44. Frenguelli, J. 1944b. Contribución al conocimiento de la flora del gondwana superior en la Argentina. 21. Neocalamites ramaccionii n. sp. Notas del Museo de La Plata, Paleontología 9: 479-485. [ Links ]
45. Frenguelli, J. 1944c. Contribución al conocimiento de la flora del gondwana superior en la Argentina. 22. Neocalamites ischigualasti n. sp. Notas del Museo de La Plata Paleontología 9: 487-492. [ Links ]
46. Frenguelli, J. 1944d. Las especie del género Zuberia en la Argentina. Anales del Museo de La Plata, Paleontología, sección B; Paleobotánica 1: 1-30. [ Links ]
47. Frenguelli, J. 1944e. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. 19. Feruglioa samaroides n. gen. et n. sp. Notas del Museo de La Plata, Paleontología 9: 403-412. [ Links ]
48. Frenguelli, J. 1944f. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. 18. Fanerotheca exstans n. g., n. sp. Notas Museo La Plata, Paleontología 9: 393-402. [ Links ]
49. Frenguelli, J. 1944g. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. 31. Cycadocarpidium majus y C. minus (Wiel). Notas Museo La Plata, Paleontología 9: 557-568. [ Links ]
50. Frenguelli, J. 1946. La flora del Gondwana superior en la Argentina. 33. Ginkgoales de los estratos de Potrerillos en la Precordillera de Mendoza. Notas del Museo de La Plata, Paleontología 11: 102-127. [ Links ]
51. Frenguelli, J. 1947. El género Cladophlebis y sus representantes en la Argentina. Anales del Museo de La Plata, Paleontología nueva serie 2: 1-74. [ Links ]
52. Frenguelli, J. 1948. Estratigrafía y edad del llamado "Rético" en la Argentina. Anales de la Sociedad Argentina de Estudios Geográficos 8: 159-309. [ Links ]
53. Gothan, W. 1912. Übber die Gattung Thinnfeldia Ettingshausen. Abhandlung der Naturhistorische Gesellschaft zu Nürnberg 19: 67-80. [ Links ]
54. Ganuza, D.G., Spalletti, L.A., Morel, E. y Arrondo, O.G. 1995. Paleofloras y sedimentología de una asociación lacustre-fluvial del Triásico tardío; la Formación Paso Flores en Cañadón de Pancho, Neuquén. Argentina. Ameghiniana 32: 3-18. [ Links ]
55. Ganuza, D.G., Zamuner, A.B., Artabe, A.E. y Spalletti, L.A. 1998. Sistemática y Paleoecología de la flora triásica de Hilario-Agua de Los Pajaritos (Formación El Alcázar), Provincia de Mendoza, Argentina. Ameghiniana 35: 271-283. [ Links ]
56. Gnaedinger, S. y Herbst, R. 1998a. La Flora Triásica del Grupo El Tranquilo, Provincia de Santa Cruz (Patagonia), parte 4: Pteridospermae. Ameghiniana 35: 33-52. [ Links ]
57. Gnaedinger, S. y Herbst, R. 1998b. La Flora Triásica del Grupo El Tranquilo, Provincia de Santa Cruz (Patagonia), parte 5: Pteridophylla. Ameghiniana 35: 53-65. [ Links ]
58. Gnaedinger, S. y Herbst, R. 2001. Pteridospermas triásicas del Norte Chico de Chile. Ameghiniana 38: 281-298. [ Links ]
59. Gnaedinger, S. y Herbst, R. 2004a. Pteridophylla triásicas del Norte de Chile. 1. El género Taeniopteris Brongniart. Ameghiniana 41: 91-110. [ Links ]
60. Gnaedinger, S. y Herbst, R. 2004b. Pteridophylla triásicas del Norte de Chile. 2. Géneros Dejerseya Herbst, Linguifolium (Arber) Retallack y Yabeiella Ôishi. Revista del Museo Argentino de Ciencias Naturales nueva serie 6: 49-59. [ Links ]
61. Halle, T.G. 1908. Zur Kenntnis der Mesozoischen equisetales Schwedens. Kungliska Svenska Vetenskapsademiens Handlingar, 43: 3-42. [ Links ]
62. Harrington, H. J. 1941. Investigaciones geológicas en las sierras de Villavicencio y Mal País (provincia de Mendoza). Dirección Nacional de Geología y Mineria, Boletín 49: 1-54. [ Links ]
63. Harris, T.M. y Millington, W. 1974. The Yorkshire Jurasic Flora. 4. Ginkgoales and Czekanowskiales. Trustees of the Brithish Museum (Natural History), London, 150 pp. [ Links ]
64. Herbst, R. 1971. Paleophytología Kurtziana 3. 7. Revisión de las especies argentinas del género Cladophlebis. Ameghiniana 8: 265-281. [ Links ]
65. Herbst, R. 2000. Dipteridaceae (Filicales) del Triásico Superior de Chile. Revista Geológica de Chile 27: 65-81. [ Links ]
66. Herbst, R. y Gnaedinger, S., 2002. Kurtziana (Pteridospermae? incertae sedis) y Alicurana nov. gen. (Cycadopsida) del Triásico y Jurásico temprano de Argentina. Ameghiniana 39: 331-341. [ Links ]
67. Herbst, R. y Lutz, A.I. 1988. Rhexoxylon brasiliensis n. sp. (Corystospermaceae, Pteridospermales) from the Upper Triassic Caturrita Formation, Brazil, with comments on biology and environment. Mededelingen rriks Geologische Dienst 42: 21-28. [ Links ]
68. Herbst, R. y Lutz, A.I. 1995. Tranquiloxylon petriellai nov. gen. et sp. (Pteridospermales) from the Upper Triassic Laguna Colorada Formation, Santa Cruz province, Argentina. Ameghiniana 32: 231-236. [ Links ]
69. Herbst, R. y Troncoso, A. 2000. Las Cycadophyta del Triásico de las Formaciones La Ternera y El Puquén (Chile). Ameghiniana 37: 283-292. [ Links ]
70. Herbst, R., Melchor, R. y Troncoso, A. 1998. Las Pteridophyta y el paleoambiente de la parte media de la Formación La Ternera (Triásico Superior) en quebrada La Cachivarita, III Región, Chile. Revista Geológica de Chile 25: 85-107. [ Links ]
71. Herbst, R., Troncoso, A. y Gnaedinger, S. 2001. Rochipteris nov. gen., hojas incertae sedis (= Chiropteris pro parte) del Triásico Superior de Argentina y Chile. Ameghiniana 38: 257-269. [ Links ]
72. Herbst, R., Troncoso, A. y Muñoz, J. 2005. Las tafofloras triásicas de la región de los Lagos, X ma Región, Chile. Ameghiniana 42: 377-394. [ Links ]
73. Høeg, O.A., 1967. Ordre Incertae Sedis des Palaeophyllales. En: E. Boureau (ed.), Traité de Paleobotanique 2. Briophyta, Psilophyta, Lycophyta. Masson et Cie., Paris, pp. 362-399. [ Links ]
74. Holmes, K.B.K. 2000. The Middle Triassic megafossil flora of Basin Creek Formation, Nymboida Coal Measures, NSW, Australia. Part 1: Bryophyta, Sphenophyta. Proceedings of the Linnean Society of New Soutw Wales 122: 43-68. [ Links ]
75. Holmes, K.B.K. 2003. The Middle Triassic megafossil flora of Basin Creek Formation, Nymboida Coal Measures, NSW, Australia. Part 3: Fernlike plant. Proceedings of the Linnean Society of New South Wales 123: 53-109. [ Links ]
76. Holmes, K.B.K. y Anderson, H.M. 2005. The Middle Triassic Megafossil Flora of the Basin Creek Formation, Nymboida Coal Measures, NSW, Australia. Part. 5: The Genera Lepidopteris, Kurtziana, Rochipteris and Walkomiopteris. Proceedings of the Linnean Society of New South Wales 126: 39-79. [ Links ]
77. Jain, R. y Delevoryas, T. 1967. A Middle Triassic Flora from the Cacheuta Formation, Minas de Petróleo, Argentina. Palaeontology 10: 564-589. [ Links ]
78. Klavins, S.D, Taylor, T.N. y Taylor, E.L. 2002. Anatomy of Umkomasia (Corystospermales) from the Triassic of Antarctica. American Journal of Botany 89: 664-676. [ Links ]
79. Kurtz, F. 1921. Atlas de las plantas fósiles de la República Argentina. Actas Academia Nacional de Ciencias, Córdoba, 7: 129-153. [ Links ]
80. Leppe, M. y Moisan, P. 2003. Nuevos registros de Cycadales y Cycadeoideales del Triásico superior del Río Bío Bío, Chile. Revista Chilena de Historia Natural 76: 475-484. [ Links ]
81. Leppe, M., Moisan, P., Abad, E. y Palma-Heldt, S. 2006. Paleobotánica del Triásico Superior del valle del río Biobío, Chile: Clase Filicopsida. Revista Geológica de Chile 33: 81-107. [ Links ]
82. Lutz, A.I. y Herbst, R. 1992. Una nueva especie de Rhexoxylon del Triásico de Barreal, San Juan, Argentina. 7° Simposio Argentino de Paleobotánica y Palinología. Asociación Paleontológica Argentina, Publicación Especial 2: 73-76. [ Links ]
83. Lutz, A.I., Crisafulli, A. y Herbst, R. 1999. Gymnospermous woods from the Upper Triassic of northern Chile. The Palaeobotanist 48: 31-38. [ Links ]
84. Lutz, A.I., Crisafulli, A. y Herbst, R. 2001. Contribución al estudio xiloflorístico de la Formación La Ternera, Triásico Superior, Chile. Ameghiniana 38: 119-127. [ Links ]
85. Lutz, A.I., Crisafulli, A. y Herbst, R. 2003. Vladiloxylon troncosoi nov. gen. et sp. (Cycadales) de la Formación La ternera (Triásico Superior), 3º Región, Chile. Revista del Museo Argentino de Ciencias Naturales, nueva serie, 5: 31-38. [ Links ]
86. Meyen, S. V. 1987. Fundamentals of Palaeobotany. Chapman and Hall, London, 432 pp. [ Links ]
87. Meyer-Berthaud, B. y Taylor, T.N. 1991. A probable conifer with podocarpacean affinities from the Triassic of Antarctica. Review of Palaeobotany and Palynology 67: 79-198. [ Links ]
88. Meyer-Berthaud, B., Taylor, E.L. y Taylor, T.N. 1992. Reconstructing the Gondwana seed fern Dicroidium: evidence from the Triassic of Antarctica. Geobios 25: 341-344. [ Links ]
89. Meyer-Berthaud, B., Taylor, T.N. y Taylor, E.L. 1993. Petrified stems bearing Dicroidium leaves from the Triassic of Antarctica. Palaeontology 36: 337-356. [ Links ]
90. Mohr, B. y Schöner, F. 1985. Eine obertriassische Dicroidium-Flora südöstlich Alto del Carmen, Región de Atacama (Chile). Newes Jahrbuch für Geologie und Palaontologie H 6: 368-379. [ Links ]
91. Morel, E.M. 1991. [Estudio Paleoflorístico y Paleoambiental de la secuencia triásica en el área de Cacheuta, provincia de Mendoza. Tesis Doctoral, Facultad de Ciencias Naturales y Museo de La Plata, Universidad Nacional de La Plata, 368 pp. Inédita]. [ Links ]
92. Morel, E.M. 1994. El Triásico del Cerro Cacheuta, Mendoza (Argentina). Parte I. Geología, contenido paleoflorístico y cronoestratigrafía. Ameghiniana 31: 161-176. [ Links ]
93. Morel, E.M. Artabe, A.E., Zavattieri, A.M. y Bonaparte, J. 2001. Cronología del Triásico de Argentina. En: A.E. Artabe, E.M. Morel y A.B. Zamuner (eds.), El Sistema Triásico de Argentina. Fundación Museo de La Plata "Francisco Pascasio Moreno", pp. 227-253. [ Links ]
94. Morel, E.M., Artabe, A.M. y Spalletti, L.A. 2003. The Triassic floras of Argentina: Biostratigraphy, Floristic events and comparison with other areas of Gondwana and Laurasia. Alcheringa 27: 231-243. [ Links ]
95. Nixon, K.C., Crepet, W.L., Stevenson, D. y Friis, E.M. 1994. A reevaluation of seed plant phylogeny. Annals of the Missouri Botanical Garden 81: 484-533. [ Links ]
96. Ôishi, S. 1931. On Fraxinopsis Wieland and Yabeiella Ôishi gen. nov. Japan Journal of Geology and Geography 8: 259-267. [ Links ]
97. Petriella, B. 1979. Sinopsis de las Corystospermaceae (Corystospermales, Pteridospermophyta) de Argentina. 1. Hojas. Ameghiniana 16: 81-102. [ Links ]
98. Petriella, B. 1981. Sistemática y vinculaciones de las Corystospermaceae H. Thomas. Ameghiniana 18: 221-234. [ Links ]
99. Petriella, B. y Arrondo, O.G. 1982. El género Kurtziana Frenguelli: su morfología y vinculaciones. Ameghiniana 19: 209-215. [ Links ]
100. Perovich, N. y Taylor, E.L. 1989. Structurally preserve fossil plants from Antarctica. 4. Triassic ovules. American Journal of Botany 76: 992-999. [ Links ]
101. Radczenko, G. P. 1957. Particularités morphologiques et anatomiques de certains types de végétaux du Carbonifère ancien de la Kuznetsk. Sbornik Pamjati A.N. Kysofovich Institut Bot. Komorov. Akademii Nauk. S.S.S.R.: 33-54. [ Links ]
102. Retallack, G.J. 1977. Reconstructing Triassic vegetation of eastern Australasia: a new approach for the biostratigraphy of Gondwanaland. Alcheringa 1: 253-283. [ Links ]
103. Retallack, G.J. 1980. Middle Triassic Megafossil Plants and Trace Fossils from Tank Gully, Canterbury, New Zealand. Journal of the Royal Society of New Zealand 10: 31-63. [ Links ]
104. Retallack, G.J. 1981. Middle Triassic megafossil plants from Long Gully, near Ottematata, nord Otago, New Zealand. Journal of the Royal Society of New Zealand 11: 167-200. [ Links ]
105. Seward, A.C. 1903. Fossil flora of Cape Colony. Annals of the South African Museum, 4: 1-122. [ Links ]
106. Smoot, E.L., Taylor, T.N. y Delevoryas, T. 1985. Structurally preserved fossil plants from Antarctica. I. Antarcticycas, gen. nov., a Triassic cycad stem from the Beardmore Glacier area. American Journal of Botany 72: 1410-1423. [ Links ]
107. Spalletti, L.A., Artabe, A.E., Brea, M. y Ganuza, D. 1995. Ambientes de acumulación y paleoflora en capas rojas triásicas de la Cuenca Cuyana, Mendoza. Revista de la Asociación Geológica Argentina 50: 175-188. [ Links ]
108. Spalletti, L.A., Artabe, A.E., Morel, E.M. y Brea, M., 1999. Biozonación paleoflorística y cronoestratigrafía del Triásico Argentino. Ameghiniana 36: 419 -451. [ Links ]
109. Spalletti, L.A., Artabe, A.E. y Morel, E.M. 2003. Geological factors and evolution of Southwestern Gondwana Triassic Plants. Gondwana Research 6: 119-134. [ Links ]
110. Spalletti, L.A., Morel, E.M., Artabe, A.E., Zavattieri, A.M. y Ganuza, D.G. 2005. Estratigrafía, facies y paleoflora de la sucesión triásica de Potrerillos, Mendoza, República Argentina. Revista Geológica de Chile 32: 249-272. [ Links ]
111. Spalletti, L.A., Fanning, C.M. y Rapela, C.W., en prensa. Age of the Triassic Potrerillos Formation, Cuyo Basin, Mendoza Province, Argentina. Gondwana Research. [ Links ]
112. Stappenbeck, R. 1910. La Precordillera de San Juan y Mendoza. Ministerio de Agricultura de la Nación, Sección Geología, Mineralogía y Minería. Anales 4: 3-187. [ Links ]
113. Sternberg, W.P. 1833. Versuch einer geognostischen botanischen. Darstellung der flora der Vorwelt 2: 1-80. [ Links ]
114. Stewart, W.R. y Rothwell, G. W. 1993. Paleobotany and the evolution of plants. Cambridge University Press, 521 pp. [ Links ]
115. Stipanicic, P. N. 1983. The Triassic of Argentina and Chile. En: M. Moullade y A. E. M. Nairn (eds.): The Phanerozoic geology of the World. 2, The Mesozoic, pp. 181-199. Amsterdam. [ Links ]
116. Stipanicic, P.N., Herbst, R. y Bonetti, M.I.R. 1996. Floras Triásicas. En: P.N. Stipanicic y M. A. Hünicken (eds.): Revisión y actualización de la obra paleobotánica de Kurtz en la República Argentina. Actas de la Academia Nacional de Ciencias 11: 127-184. [ Links ]
117. Taylor, D.W. y Hickey, L.J. 1992. Phylogenetic evidence for the herbaceous origin of angiosperms. Plant Sytematics and Evolution 180: 137-156. [ Links ]
118. Taylor, E.L. 1992. The occurrence of a Rhexoxylon-like stem in Antarctica. Courrier Forschungsinstitut Senckenberg 147: 183-189. [ Links ]
119. Taylor, E.L. 1996. Enigmatic gymnospermous? structurally preserved Permian and Triassic seed ferns from Antarctica. Review of Palaeobotany and Palynology 90: 303-318. [ Links ]
120. Taylor, T.N., Del Fueyo, G. M. y Taylor, E.L. 1994. Permineralized seed fern cupules from the Triassic of Antarctica: implications for cupule and carpel evolution. American Journal of Botany 81: 666-677. [ Links ]
121. Thomas, H. H. 1933. On some Pteridospermous plants from the Mesozoic rocks of South Africa. Philosophical Transactions of the Royal Society London B 222: 193-254. [ Links ]
122. Thomas, H.H. y Bose, M.N. 1955. Pachydermophyllum papillosum gen. et sp. nov. from the Yorkshire Jurassic. Annals and Magazine of Natural History 12: 335-543. [ Links ]
123. Townrow, J.A. 1957. On Dicroidium probably a Pteridospermous leaf, and other leaves now removed from this genus. Transactions Geological Society South Africa 60: 21-56. [ Links ]
124. Townrow, J. A. 1960. The Peltaspermaceae, a Peteridosperm Family of Permian and Triassic age. Palaeontology 3: 333-361. [ Links ]
125. Troncoso, A. y Herbst, R. 1999. Ginkgoales del Triásico del norte de Chile. Revista Geológica de Chile 26: 255-271. [ Links ]
126. Troncoso, A. y Herbst, R. 2000. La Tafoflora Triásica del Cajón Troncoso, Alta Cordillera del Maule, 7ª Región, Chile. Revista Museo Argentino Ciencias Naturales, nueva serie 2: 137-144. [ Links ]
127. Truempy, E. y Lhez, R. 1937. División estratigráfica de los terrenos aflorantes en la región comprendida entre Luján de Cuyo, Potrerillos y Tupungato. Boletín de Informaciones Petroleras 223: 39-42. [ Links ]
128. Walkom, A.B. 1925. Notes on some Tasmanian Mesozoic Plants. Part 1. Papers and proceedings of the Royal Society of Tasmania, 1924: 73-89. [ Links ]
129. Wieland, G. R. 1929. Antiquity of the Angiosperms. International Congress of Plant Sciences, Ithaca, New York, Proceedings: 429-456. [ Links ]
130. Yao, X., Taylor, T.N. y Taylor, E.L. 1995. The corystosperm pollen organ Pteruchus from the Triassic of Antactica. American Journal of Botany 82: 535-546. [ Links ]
131. Yao, X., Taylor, T.N. y Taylor, E. 1997. A taxodiaceous seed cone from the Triassic of Antarctica. American Journal of Botany 84: 343-354. [ Links ]
132. Zamuner, A.B., Artabe, A.E. y Ganuza, D.G. 1999. A new peltasperm (Gymnospermopsida) from the Middle Triassic of Argentina. Alcheringa 23: 185-191. [ Links ]
133. Zamuner, A.B., Zavattieri, A.M., Artabe, A.M. y Morel, E.M. 2001. Paleobotánica. En: A.E. Artabe, E.M. Morel y A.B. Zamuner (eds.) El Sistema Triásico de Argentina. Fundación Museo de La Plata "Francisco Pascasio Moreno", pp. 143-184. [ Links ]
Recibido: 4 de mayo de 2006.
Aceptado: 3 de mayo de 2007.

